用于提高的冻存后细胞活力和滞留的胞外基质组分和/或基质细胞蛋白的选择的制作方法
技术背景
本发明的主题涉及附着于基质的贴壁细胞的冻存及后续解冻的方法。还揭示了鉴定一种或多种胞外基质(ecm)组分和/或基质细胞蛋白的方法,所述胞外基质组分和/或基质细胞蛋白提高细胞在从冻存状态解冻期间及之后的活力和滞留。
减少在研究中使用动物数量的持续需求驱动着基于细胞和基于组织的体外分析的发展,并提供关于各种化学制品、化合物和制剂的准确毒性数据。在2009年,欧盟禁止了使用动物来测试化妆品成分。这之后可能会禁止使用动物来测试其他类型化合物(包括药物和家用化学制品)的毒性。
对于该需求,已采用通过将细胞和组织的温度降至水的冰点以下进行的细胞和组织的冻存来保护和保存生物系统。细胞的冻存通常在悬浮形式中进行,并且对于细胞在固定基质上的冻存的研究极少。因此,开发用于细胞悬液的常用的冻存方案也通常应用于固定基质上的贴壁细胞,这常常导致冻存后细胞脱离和膜的破坏。只能通过使用合适的冷冻保护剂来使细胞在冻存处理中的冻融的严苛条件下存活。因此,冷冻生物学中的大多数研究着眼于寻找并测试新型冷冻保护剂。
然而,在冻融的严苛条件之后,对于ecm(天然或合成)的细胞贴附的滞留对于保存天然和工程改造的组织以及其它应用的如体外毒理学测试而言是至关重要的。贴壁机制的破坏不可避免地具有严重的后果;甚至对于贴壁机制的可逆作用也可能是灾难性的,因为与基础性的ecm的空间分离足以阻止再附着。
因此,需要用于贴壁细胞的改善的冻存方案。具体地说,希望在将附着于基质的细胞从冻存状态升温后,获得改善的细胞贴附和活力。
技术实现要素:
本文公开了一种用于附着于基质的贴壁细胞的冻存方法,所述方法提高冻存细胞的细胞活力和滞留(retention)。所述方法包括:用至少一种ecm组分处理基质和/或向所述基质或细胞培养基添加至少一种基质细胞蛋白,将细胞铺板于经处理的基质上;和通过使细胞降温至冻存温度来在经处理的基质上冻存所述细胞。
还公开了鉴定一种或多种ecm组分和/或基质细胞蛋白质的方法,所述ecm组分和/或基质细胞蛋白质提高基质上的细胞在从冻存状态解冻后的活力和滞留。所述方法包括:选择特定细胞类型的细胞;用一种或多种不同的ecm组分处理基质和/或向所述基质或细胞培养基添加一种或多种基质细胞蛋白;将所述特定细胞类型的细胞铺板于经处理的基质上;通过使细胞降温至冻存温度,在经各种处理的基质上冻存所述细胞;通过如下方式解冻所述细胞:将含有所述细胞的经处理的基质暴露于第一环境,所述第一环境具有高于所述冻存温度的第一升温温度,使细胞从冻存温度经历第一次升温,然后,通过使所述细胞暴露于第二环境,所述第二环境具有高于第一升温温度的第二升温温度,使所述细胞从第一升温温度进一步升温;评估解冻的细胞以确定细胞活力和滞留;和,鉴定一种或多种ecm组分和/或至少一种基质细胞组分,所述ecm组分和/或基质细胞组分能提高特定细胞类型的细胞在从冻存状态解冻后的活力和滞留。
附图说明
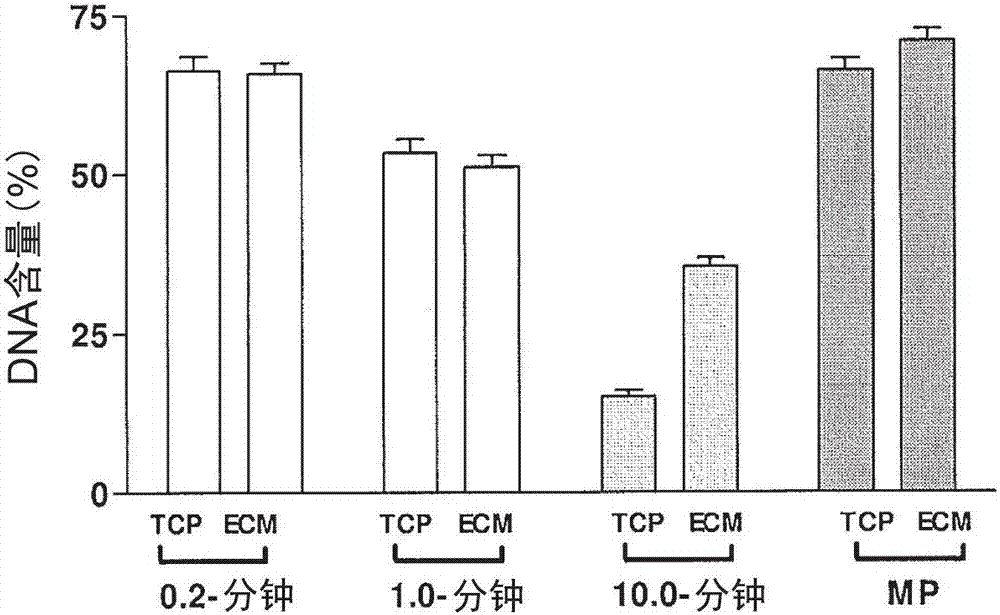
附图1a和1b的柱状图说明在以各种降温速率进行冻存后,ecm或组织培养物处理的塑料(tcp)上的贴壁牛角膜内皮(bce)细胞的(a)细胞活力和(b)dna含量(%)(即细胞滞留)。
附图2a-2d的柱状图说明在采用(a)单一和成对的ecm组分、(b)三种ecm组分的组、(c)四种ecm组分的组、和(d)五种ecm组分的组进行冻存后,贴壁bce细胞的细胞活力。
附图3a-3d的柱状图说明在采用(a)单一和成对的ecm组分、(b)三种ecm组分的组、(c)四种ecm组分的组,和(d)五种ecm组分的组进行冻存后,贴壁bce细胞的dna含量(%)(即细胞滞留)。
附图4a-4d的柱状图说明在采用(a)单一和成对的ecm组分、(b)三种ecm组分的组、(c)四种ecm组分的组、和(d)五种ecm组分的组进行冻存后,贴壁人间充质干细胞(hmsc)的细胞活力。
附图5a和5b的柱状图说明在通过单一快速解冻步骤或两步法解冻方案进行冻存和解冻后,各种基质上的贴壁hmsc的(a)细胞活力和(b)dna含量(%)(即细胞滞留)。
附图6的柱状图说明在明胶上或ecm与层粘连蛋白、胶原i、胶原iii和胶原v的组合上,采用或不采用基质细胞蛋白生腱蛋白x,进行冻存之后,贴壁hmsc的细胞活力。
具体实施方式
本文所使用的与数量相关使用的修饰语“约”包括所述的值,并且具有上下文描述的含意。例如,它至少包括与该具体数值的测量相关联的误差度。在使用范围的情况下,修饰语“约”也应该被认为是公开了由两个端点的绝对值定义的范围。例如,“约2-约4”的范围也公开了“2-4”的范围。
除非在本文中另有明确说明,述及温度(℃)时,修饰语“约”是指所述值或值的范围以及所述值或值范围+/-1-4%。
除非在本文另有明确说明,述及细胞活力和细胞滞留或附着力(%)时,修饰语“约”是指所述值或值的范围以及所述值或值范围+/-1-3%。
除非在本文另有明确说明,述及浓度(μg/ml)时,修饰语“约”是指所述值或值的范围以及所述值或值范围+/-1-4%。
除非在本文另有明确说明,述及摩尔浓度(m)时,修饰语“约”是指所述值或值的范围以及所述值或值范围+/-1-2%。
除非在本文另有明确说明,述及降温速率(℃/分钟)时,修饰语“约”是指所述值或值的范围以及所述值或值范围+/-1-3%。
如本文所用,术语“室温”是指标准压强下约为18℃到25℃的温度。在不同的实施例中,室温可能约为18℃、约为19℃、约为20℃、约为21℃、约为22℃、约为23℃、约为24℃,或约为25℃。
本方法涉及固定在基质上的贴壁细胞的冻存(即通过冷冻保存)以及所述细胞的后续解冻,即从冻存温度升温,以用于各种应用,如体外毒理学测试。例如,术语“冷冻”是指低于水的冰点的温度,即低于0℃。冻存通常包括将细胞冷冻至远低于冰点的温度,例如至-80℃或更低,更通常是至-130℃或更低。可使用任何本领域从业人员已知的任何冻存方法,没有限制。冻存温度可低于-20℃,例如-80℃或更低,或者-130℃或更低。冻存温度可约为-20℃至-200℃,约为-30℃至-175℃,约为-50℃至-160℃,约为-65℃至-150℃,约为-75℃至-135℃,约为-80℃至-130℃,约为-90℃至-125℃,或约为-100℃至-115℃。参见例如,armitage等,“降温速率对冷冻细胞存活的影响在单层与悬液中不同”("theinfluenceofcoolingrateonsurvivaloffrozencellsdiffersinmonolayersandsuspensions"),cryo-letters17:213-218(1996)。
本方法经设计以使细胞能在再升温后立即使用,这消除了对细胞铺板、扩增及再铺板的需要。为此,已经开发了用于将冻存的贴壁细胞从冻存温度升温的两阶段升温方案。参见例如,campbell等,美国专利号6,596,531("campbell'531"),其通过引用全文纳入本文,并显示了贴壁细胞可在多孔板中以贴壁分化细胞单层的形式冻存。
本方法针对附着于基质的贴壁细胞(例如bce细胞和hmsc)的冻存。参见例如,ji等,“贴壁人胚胎干细胞的冻存”("cryopreservationofadherenthumanembryonicstemcells"),biotechnologyandbioengineering88(3):299-312(2004);katkov等,“完全解离的贴壁人诱导型多能干细胞的无dmso编程的的冻存”("dmso-freeprogrammedcryopreservationoffullydissociatedandadherenthumaninducedpluripotentstemcells"),stemcellsinternational2011,2011:981606.doi:10.4061/2011/981606,于2011年1月1日电子公开;以及xu等,“在冻存期间,渗透和冷激击对贴壁人间充质干细胞的影响”("effectsofosmoticandcoldshockonadherenthumanmesenchymalstemcellsduringcryopreservation"),j.biotech.162(2-3):224-231(2012)。
可使用任何合适的基质,没有限制。例如,可使细胞附着于组成具有多个孔的微滴定板(即多孔板)的表面的组织培养塑料(tcp)、胶原凝胶、天然基质,或合成材料。对于基质的细胞附着是本领域已知的。参见例如,campbell等,“用于冻存细胞的无血清溶液”("serumfreesolutionsforthecryopreservationofcells"),invitrocelldev.biol.,43:269-275(2007);campbell等,“猪主动脉瓣膜小叶衍生肌成纤维细胞的冻存”("cryopreservationofporcineaorticheartvalveleaflet-derivedmyofibroblasts"),biopreservationandbiobanking,8(4):211-217(2010);campbell等,“冻存后用海藻糖培养产生活性内皮细胞”("culturingwithtrehaloseproducesviableendothelialcellsaftercryopreservation"),cryobiology,64(3):240-244(2012);hornung等,“固定于结构化玻璃和硅基质上的锚定依赖性哺乳动物细胞的冻存”("cryopreservationofanchorage-dependentmammaliancellsfixedtostructuredglassandsiliconsubstrates"),cryobiology,33:260-70(1996);mcgann等,“冻存期间细胞与细胞间和细胞与表面间的相互作用反应”("cell-to-cellandcell-to-surfaceinteractionsaffectresponsesduringcryopreservation"),transfusion,33(7):611(1993);ohno,“一种原位冷冻锚定依赖性细胞的简单方法”("asimplemethodforinsitufreezingofanchorage-dependentcells"),刊于:adoyle,jbgriffiths,dgnewell(编),《细胞和组织培养:实验步骤》(cellandtissueculture:laboratoryprocedures),奇切斯特:约翰威力父子公司(johnwileyandsons)(1994);pasch等,“悬液和单层角质形成细胞的冻存中hes浓度的变化”("variationofthehesconcentrationforthecryopreservationofkeratinocytesinsuspensionsandinmonolayers"),cryobiology,41(2):89-96(2000);以及pasch等,“单层角质形成细胞的冻存”("cryopreservationofkeratinocytesinamonolayer"),cryobiology,39(2):158-168(1999),上述每篇文献均通过引用全文纳入本文,并显示贴壁细胞的成功活力和对于tcp的细胞附着。
本发明的方法涉及通过采用ecm或一种或多种ecm组分处理基质来促进和提高细胞在冻存期间的活力和附着力。ecm是为组织提供结构框架的动态结构的小生境,也密切涉及细胞加工,例如信号转导、分化、增殖、附着、极性及存活。参见例如,frantz等,“胞外基质概况”("theextracellularmatrixataglance"),j.cellsci.,123:4196-4200(2010),该文献揭示了ecm在细胞的最终细胞健康中起到重要作用。
例如,短语“改善的细胞活力”或“改善的活力”是指细胞活力(%)至少为60%,例如80%或更高。改善的细胞活力(%)可以是65%或更高、67%或更高、70%或更高、73%或更高、75%或更高、77%或更高、80%或更高、83%或更高、85%或更高、87%或更高、90%或更高、93%或更高、95%或更高、97%或更高、98%或更高,或99%或更高。同样地,术语“细胞滞留”,“滞留”,“细胞附着”或“附着”是指,例如,dna含量的测量,其可用作细胞数量的指标。短语“改善的细胞滞留”、“改善的滞留”、“改善的细胞附着”或“改善的附着”是指,例如,至少为80%、81%或更多、82%或更多、83%或更多、84%或更多、85%或更多、87%或更多、89%或更多、90%或更多、92%或更多、95%或更多、96%或更多、97%或更多、98%或更多,或99%或更多的dna的含量(%)。参见例如,malpique等,“贴壁细胞的冻存:提高解冻后细胞活力和功能的策略”("cryopreservationofadherentcells:strategiestoimprovecellviabilityandfunctionafterthawing"),tissueengineeringpartcmethods,15(3):373-386(2009)。
如附图1说明,对于ecm而不是tcp的细胞的附着,提高了冻存期间和之后的细胞活力和滞留。完全形成和组织的ecm的存在提供了更近似于细胞的天然环境的表面。因此,细胞附着可能较少地受冻存过程影响。此外,ecm组合物可能影响细胞在冻存期间维持附着的能力。而且,通过采用ecm来冻存细胞,细胞所处的构造可能更有利于其整体健康并且可能改善其应对冷冻温度的耐受力和恢复力。
一些实施方式涉及贴壁细胞在完全形成的ecm上的冻存。其他实施方式涉及采用某些ecm组分的贴壁细胞的冻存,以促进冻存期间细胞的附着,并且在细胞从冻存状态重新升温后在使用前为其提供更天然的环境。用ecm组分处理基质可涉及将ecm组分被覆于细胞基质上。
所述ecm组分可包括本领域已知的任何ecm组分。例如,本领域已知,细胞对ecm基质的附着是通过接合复合物(粘着斑)介导的。这些接合复合物包括特异性粘附受体,其中许多属于称为整联蛋白的同源基质受体的大型超家族,其大部分识别它们结合的胞外蛋白中的arg-gly-asp(rgd)三肽序列。一些细胞在与胶原的结合中利用其他明显无关的跨膜糖蛋白,并且许多细胞具有将细胞直接连接到ecm的完整的膜蛋白聚糖。参见例如,rixen等,“在体外i型和iv型胶原上,角膜内皮细胞的附着与扩散:研究内皮修复机制的模型”("adhesionandspreadingofcornealendothelialcellsoncollagentypeiandivinvitro:amodeltostudymechanismsofendothelialrepair"),res.exp.med.,190:203-211(1990)。
在培养条件下,各种细胞类型,例如牛角膜内皮细胞,对于组织培养聚苯乙烯的最初附着,可能取决于细胞-粘附糖蛋白,例如纤连蛋白和/或玻连蛋白,对培养表面的吸附。参见例如,underwood等,“细胞-粘附蛋白玻连蛋白和纤连蛋白的生物活性的比较”("acomparisonofthebiologicalactivitiesofthecell-adhesiveproteinsvitronectinandfibronectin"),j.cell.sci.,93(pt.4):641-649(1989)。在细胞接种之前,所述细胞-粘附蛋白可以纯化形式预被覆在聚合物上;它们也可以从用于补充培养基的血清吸附到培养表面上;或者,它们可由细胞合成并沉积到塑料表面上。
已经报道内源性蛋白质至少部分地有助于在没有血清来源的蛋白质(例如纤连蛋白和玻连蛋白)的情况下将细胞附着于合成的基质上。可以使用任何新鉴定的或众所周知的内源性蛋白质和细胞粘附蛋白。参见例如,gordon等,“损伤期间细胞骨架的作用-诱导的角膜内皮的细胞迁移”("roleofthecytoskeletonduringinjury-inducedcellmigrationincornealendothelium"),cellmotil.&cytoskeleton,6:47-57(1990)。蛋白聚糖介导的相互作用可促进附着细胞中肌动蛋白丝的组织。肌动蛋白的动态作用涉及肌动蛋白结合蛋白质的作用,其调节肌动蛋白丝的长度和缔合,并且取决于球状和丝状肌动蛋白之间的平衡。这受到温度、ph、离子强度和有机溶剂的存在的影响,所有这些都是冻存中的因素。因此,在本方法中,肌动蛋白结合蛋白质可用作ecm蛋白。
bce细胞生产许多ecm蛋白,以建立成熟的基质,包括纤连蛋白、层粘连蛋白和i型、iii型、iv型和v型胶原。纤连蛋白和层粘连蛋白由分离的细胞和单层中的细胞差异表达。类似地,不同的胶原类型也不以与优势胶原iii等同的方式表达。参见例如,gospodarowicz等,“在培养的角膜内皮细胞中鉴定和定位纤连蛋白:细胞表面极性和生理意义”("theidentificationandlocalizationoffibronectininculturedcornealendothelialcells:cellsurfacepolarityandphysiologicalimplications"),exp.eyeres.,29:485-509(1979);gospodarowicz等,“在培养的血管和角膜内皮细胞中生产和定位层粘连蛋白”("theproductionandlocalizationoflaminininculturedvascularandcornealendothelialcells",j.cellphysiol.,107:171-183(1981);及scheffer等,“由培养的牛角膜内皮细胞合成的胶原的特征”("characterizationofcollagenssynthesizedbyculturedbovinecornealendothelialcells"),j.biol.chem.,256(7):3361-3365(1981)。例如,层粘连蛋白,在单层形成前大量产生,随后蛋白质产量下降,并且可能不利于冻存期间的粘附,如纤连蛋白,其生成不随细胞密度而变化。此外,可证明多种组分的组合是维持附着和活力的更好的基质。例如,已显示纤连蛋白有利于牛角膜内皮细胞附着于基质中的胶原。参见例如,scott等,“牛角膜内皮细胞对胶原和其他亚内皮组分的附着的研究”("investigationoftheattachmentofbovinecornealendothelialcellstocollagensandothercomponentsofthesubendothelium"),exp.cellres.,144:472-478(1983)。
hmsc也生产ecm蛋白,包括纤连蛋白、层粘连蛋白以及i型、iii型、iv型和v型胶原,并且可受到它们所在的ecm小生境的大幅影响。ecm组合物影响间充质干细胞如何并且向什么谱系分化,当开发用于将在板上冻存的干细胞的ecm覆层时,需要考虑该方面。参见例如,singh等,“纤连蛋白和干细胞分化——来自软骨形成的研究结果”("fibronectinandstemcelldifferentiation-lessonsfromchondrogenesis"),j.cellsci.,125:3703-3712(2012)。
基质可用ecm组分处理,所述ecm组分包括:纤连蛋白、层粘连蛋白、胶原i、胶原iii、胶原iv和胶原v中一种或多种。基质可用浓度约为0.5-25μg/ml,例如约1-15μg/ml,或约1-10μg/ml的ecm组分处理。基质可用ecm组分处理,所述ecm组分的浓度为约0.75-20μg/ml、约2-20μg/ml、约2-15μg/ml、约5-10μg/ml、约5-12μg/ml、约5-15μg/ml、约5-18μg/ml、约5-20μg/ml、约5-22μg/ml、约7-20μg/ml、约7-15μg/ml、约7-10μg/ml、约10-25μg/ml、约10-20μg/ml、约10-15μg/ml、约15-25μg/ml,或约15-20μg/ml。
当冻存的细胞是牛角膜内皮细胞时,基质可用ecm组分处理,所述ecm组分可包括:纤连蛋白、层粘连蛋白、胶原i、胶原iii、胶原iv和胶原v中的一种、两种、三种、四种、五种或六种。ecm组分的示例性组合包括:纤连蛋白、胶原i和胶原v;纤连蛋白、层粘连蛋白、胶原i和胶原iv;纤连蛋白、胶原iii、胶原iv和胶原v;或者胶原i、胶原iii、胶原iv和胶原v。
当冻存的细胞是人间充质干细胞时,ecm组分可以是包括纤连蛋白、层粘连蛋白、胶原i、胶原iii、胶原iv和胶原v的一种或多种蛋白质。ecm组分的示例性组合可包括:纤连蛋白、胶原iv和/或胶原v;纤连蛋白、层粘连蛋白、胶原i和胶原v;胶原iv和胶原v;纤连蛋白、层粘连蛋白和胶原v;层粘连蛋白、胶原iv和胶原v;胶原iii、胶原iv和胶原v;以及层粘连蛋白、胶原iii和胶原v。
除了ecm组分外,可将基质细胞蛋白添加至基质,或可将它们添加至细胞培养基。近年来,已揭示基质细胞蛋白的功能和重要性。虽然这些蛋白质存在于ecm中,它们在维持基质的结构方面不起重要作用。相反,它们参与细胞功能的调节。基质细胞蛋白通过与细胞表面受体、激素和其它效应分子(包括ecm)相互作用来调节细胞功能。它们分泌并存在于胞外环境中,但不像常规的ecm蛋白一样起到结构性作用。虽然在发育过程中更为显著,但它们仍然存在于成人中,特别是在受伤部位。它们发挥各种各样的功能,这由它们所在的环境决定。在本方法中,可以使用任何已知的基质细胞蛋白或未来发现的基质细胞蛋白。参见例如,bornstein等,“基质细胞蛋白:细胞功能的胞外调节剂”("matricellularproteins:extracellularmodulatorsofcellfunction"),curr.opin.cellbiol.,14:608-616(2002);bornstein等,“基质细胞蛋白:概述”("matricellularproteins:anoverview"),j.cellcommun.signal.,3:163-165(2009);frangogiannis,“心脏适应和疾病中的基质细胞蛋白”("matricellularproteinsincardiacadaptationanddisease"),physiol.rev.,92:635-688(2012);morris等,“基质细胞蛋白和生物材料”("matricellularproteinsandbiomaterials"),matrixbiol.mar.,pii:s0945-053x(14)00051-1.doi:10.1016/j.matbio.2014.03.002(2014);roberts,“基质细胞蛋白的新兴功能”("emergingfunctionsofmatricellularproteins"),cellmol.lifesci.,68(19):3133-3136(2011);wong等,“基质细胞蛋白:启动癌症发展和转移的肿瘤微环境”("matricellularproteins:primingthetumourmicroenvironmentforcancerdevelopmentandmetastasis"),brit.j.cancer,108:755-761(2013)。迄今为止的大多数研究已经用敲除小鼠研究了基质细胞蛋白的各种功能,而几乎没有体外评估这些蛋白的研究。
基质细胞蛋白影响细胞行为的能力可被利用以促进细胞在体外系统的活力和附着力。同样,基质可用一种或多种基质细胞蛋白,单独或与ecm组分组合,来处理。也即,所述方法也可包括向细胞基质或培养基添加基质细胞蛋白。合适的基质细胞蛋白包括血小板反应蛋白-1、生腱蛋白-c,生腱蛋白-x,sparc(分泌型蛋白,酸性且富含半胱氨酸)、骨膜素(periostin)、ccn-1,以及骨桥蛋白。
可使用浓度约为0-5μg/ml的基质细胞蛋白。基质细胞蛋白可以如下浓度使用:约0.25-5μg/ml、约0.5-5μg/ml、约0.75-5μg/ml、约1-5μg/ml、约2-5μg/ml、约3-5μg/ml、约4-5μg/ml、约0-4μg/ml、约0-3μg/ml、约0-2μg/ml、约0-1μg/ml、约0.5-4μg/ml、约0.5-3μg/ml、约1-4μg/ml、约1-3μg/ml,或约1-2μg/ml。
通过将细胞与二糖,如海藻糖,孵育,可在冻存前进一步保护细胞(campbell等,在保存前用糖处理细胞材料的方法(methodfortreatmentofcellularmaterialswithsugarspriortopreservation),见于2007年9月18日出版的美国专利7,270,946)
通过在将细胞在冷冻至冻存温度前使其与冻存组合物接触,可在冻存期间进一步保护细胞与冻存组合物接触是指使细胞以某种方式与冻存组合物接触,从而在降温至冻存温度期间,细胞被冻存组合物保护。例如,可通过填充待保护的细胞的附着的板中合适的孔来使细胞与冻存组合物接触。
也可使待冻存的细胞与冷冻相容的ph缓冲液接触,所述ph缓冲液通常至少主要包含:碱性盐溶液、能量源(例如葡萄糖),以及能够在降温温度下保持中性ph的缓冲剂。众所周知的这样的材料包括,例如杜尔贝科改良的伊格培养基(dulbecco'smodifiedeaglemedium)(dmem)。该材料也可作为冻存组合物的一部分。参见例如,campbell等,“贴壁平滑肌和内皮细胞与二糖一起冻存”("cryopreservationofadherentsmoothmuscleandendothelialcellswithdisaccharides"),刊于:katkovi.(编),《冻存前沿》(currentfrontiersincryopreservation),克罗地亚:科技公司(intech)(2012);以及campbell等,“胰腺储存解决方案的发展:供于低温储存后β细胞存活和代谢状态的细胞保护补充剂的初步筛选”("developmentofpancreasstoragesolutions:initialscreeningofcytoprotectivesupplementsforβ-cellsurvivalandmetabolicstatusafterhypothermicstorage"),biopreservationandbiobanking,11(1):12-18(2013)。
冻存组合物可能包含本领域已知的任何冷冻保护性物质。已知的冷冻保护剂化合物包括乙酰胺、琼脂糖、藻酸盐、l-苯胺、白蛋白、醋酸铵、丁二醇、硫酸软骨素、氯仿、胆碱、右旋糖、二甘醇、二甲基乙酰胺、二甲基甲酰胺、二甲基亚砜(dmso)、赤藻糖醇、乙醇、乙二醇、甲酰胺、葡萄糖、甘油、α-甘油磷酸酯、甘油单乙酸酯、甘氨酸、羟乙基淀粉、肌醇、乳糖、氯化镁、硫酸镁、麦芽糖、甘露糖醇、甘露糖、甲醇、甲基乙酰胺、甲基甲酰胺、甲基脲、苯酚、普朗尼克(pluronic)多元醇、聚乙二醇、聚乙烯吡咯烷酮、脯氨酸、丙二醇、吡啶n-氧化物、核糖、丝氨酸、溴化钠、氯化钠、碘化钠、硝酸钠、硫酸钠、山梨醇、蔗糖、海藻糖、三甘醇、三甲胺醋酸盐、尿素、缬氨酸、木糖等。
冷冻保护剂化合物可存在于冻存组合物中,例如其含量是约0.05m-6m、约0.1-3m、约0.25-6m、约1-6m、约2-6m、约3-6m、约4-6m、约5-6m、约0.25-1m、约0.25-2m、约0.25-3m、约0.25-4m、约0.25-5m、约1-4m、约1-3m、约1-2m、约3-5m、约2-4m、约0.5-6m、约0.5-5m、约0.5-4m、约0.5-3m、约0.5-2m,或约0.5-1m。
冷冻保护剂组合物可能包含至少一种环己烷二醇(chd)化合物,例如顺式或反式的1,3-环己烷二醇(1,3chd)或1,4-环己烷二醇(1,4chd),或其外消旋混合物,作为冷冻保护剂化合物。
chd化合物可在冻存组合物中存在,例如其含量是约0.05-2m、约0.1-1m、约0.1-2m、约0.1-1m、约0.1-1.5m、约0.1-0.5m、约0.1-0.25m、约1-2m、约1.5-2m、约0.75-2m、约0.75-1.5m、约0.75-1m、约0.05-1m、约0.05-0.75m、约0.05-0.5m,或约0.05-0.1m。冻存组合物也可包含良好适用于细胞、组织和器官的储存的溶液。溶液可包括上文讨论过的缓冲液。例如,溶液可以是eurocollins溶液,所述溶液由右旋糖、磷酸二氢钾和磷酸氢二钾、碳酸氢钠以及氯化钾组成。参见例如,taylor等,“作为冷冻保护剂的运载体溶液的unisol和euro-collins溶液的比较”("comparisonofunisolwitheuro-collinssolutionasavehiclesolutionforcryoprotectants"),transplantationproceedings,33:677-679(2001)。
冻存组合物可同时包含至少一种chd化合物和至少一种额外的冷冻保护剂化合物。
另外,冻存组合物也可包含抗冷冻蛋白质/多肽(afp)或抗冷冻糖脂(afgl)。afp也包括抗冷冻糖蛋白(afgp)和昆虫抗冷冻或“热滞(thermalhysteresis)”蛋白(thp)。最近发现的afgl已在昆虫和植物中观察到。据信,天然产生的afp能够结合于正在发展的冰晶体的棱柱面,从而改变它的形成。由于鱼和昆虫产生这些蛋白,这意味着它们冰点的下降,所以它们能够在一般应会导致它们的体液被冻住的条件下存活。就此而言,本方法可使用任何新发现的或众所周知的afp。参见例如,sicheri和yang,nature,375:427-431,(1995),描述八种这样的蛋白;devries,“抗冷冻糖肽和多肽:与冰和水的相互作用”("antifreezeandicenucleatorproteinsinterrestrialarthropods"),meth.enzymol.,127:293-303(1986);duman,“陆地节肢动物的抗冰冻和冰核蛋白”("antifreezeandicenucleatorproteinsinterrestrialarthropods"),annualrev.physiol.,63:327-3257(2001);holmstrup等,“在北极跳虫onychiurusarcticus中的脱水和抗旱”("dehydrationandcoldhardinessinthearcticcollembolanonychiurusarcticus"),j.comp.physiol.b,168:197-203(1998);kuiper等,“通过吸附冰来纯化抗冷冻蛋白”("purificationofantifreezeproteinsbyadsorptiontoice"),biochem.biophys.res.commun.,300(3):64-68(2003);miller等,“一些成年的和未成熟的昆虫在内陆阿拉斯加越冬的抗寒策略”("cold-hardinessstrategiesofsomeadultandimmatureinsectsoverwinteringininterioralaska"),comp.biochem.physiol.,73a:595-604(1982);neven等,“昆虫血淋巴脂蛋白冰核剂的纯化和特征:磷脂酰肌醇和载脂蛋白在冰核活动中的重要性的证据”("purificationandcharacterizationofaninsecthemolymphlipoproteinicenucleator:evidencefortheimportanceofphosphatidylinositolandapolipoproteinintheicenucleatoractivity"),j.comp.physiol.b,159:71-82(1989);sformo等,“阿拉斯加树皮甲虫的幼虫中的深度过冷,玻璃化和-100℃的极限生存”("deepsupercooling,vitrificationandlimitedsurvivalto-100℃inthealaskanbeetlecucujusclavipespuniceuslarvae"),j.exp.biol.,213(3):502-509(2010);storey等,“动物的耐寒性”("freezetoleranceinanimals"),physiol.rev.,68:27-84(1988);storey等,“昆虫中抗寒的生化适应”("biochemicaladaptationforcoldhardinessininsects"),phil.trans.r.soc.lond.,b326:635-54(1990);walters等,“北极阿拉斯加石蝇(nemouraarctica)的耐寒性”("freezetoleranceinthearcticalaskastonefly,nemouraarctica"),j.exp.biol.,212:305-12(2009a);walters等,“在耐寒的阿拉斯加甲虫(upisceramboides)中冷冻保护剂的生物合成以及苏糖醇的选择性积累”("cryoprotectantbiosynthesisandtheselectiveaccumulationofthreitolinthefreezetolerantalaskanbeetle,upisceramboides"),j.biol.chem.,284:16822-16831(2009b);walters等,“在耐寒的阿拉斯加甲虫(upisceramboides)中的非蛋白的产生热滞后的木甘聚糖防冻剂”("anon-proteinthermalhysteresis-producingxylomannanantifreezeinthefreeze-tolerantalaskanbeetle,upisceramboides"),proc.natl.acad.sci.,106,20210-20215(2009c);walters等,“在不同的分类中发现与耐寒相关的产生热滞后的木甘聚糖糖脂抗冻剂”("athermalhysteresis-producingxylomannanglycolipidantifreezeassociatedwithcoldtoleranceisfoundindiversetaxa"),j.comp.physiol.b.,181(5):631-40(2011);wang等,“加拿大拟步甲虫的抗冻蛋白增强彼此的活性”("antifreezeproteinsofthebeetledendroidescanadensisenhanceoneanother'sactivities"),biochemistry,44:10305-10312(2005);worland等,“极地陆生节肢动物在零度温度下的脱水应激”("desiccationstressatsubzerotemperaturesinpolarterrestrialarthropods"),j.insect.physiol.,49:193-203(2003);zachariassen等,“昆虫抗血清中的成核剂耐寒”("nucleatingagentsinthehaemolymphofinsectstoleranttofreezing"),nature,262:285-87(1976);以及zachariassen,“昆虫中耐寒性生理学”("physiologyofcoldtoleranceininsects"),physiol.rev.,65:799-832(1985)。
示例性afp包括afpi(i型afp)、afpiii(iii型afp)以及/或afgp。冻存组合物中存在的afp含量,例如,对每一种存在的afp而言,约为组合物的0.001-1mg/ml、约0.05-0.5mg/ml,或约0.1-0.75mg/ml。
一旦细胞已经接触了冻存组合物,细胞即可随后被冷冻用于冻存。用于冻存的降温可用任何方式进行,并且可以使用任何上述以外的物质。
例如,用于冻存的降温(冷冻)方案可以是任何合适的类型。本领域技术人员熟知多种类型的降温方案。降温方案可包括以连续速率降温的方式从冰成核点降至-80℃或任何上述的降温温度,降温速率取决于冷冻的细胞/组织的特性。例如,降温速率可以是每分钟约-0.1℃至约-10℃,或每分钟约-1℃至约-2℃。降温速率可以是每分钟约-0.1℃至-9℃、每分钟约-0.1℃至-8℃、每分钟约-0.1℃至-7℃、每分钟约-0.1℃至-6℃、每分钟约-0.1℃至-5℃、每分钟约-0.1℃至-4℃、每分钟约-0.1℃至-3℃、每分钟约-0.1℃至-2℃、每分钟约-0.1℃至-1℃、每分钟约-0.1℃至-0.5℃、每分钟约-1℃至-2℃、每分钟约-1℃至-3℃、每分钟约-1℃至-4℃、每分钟约-1℃至-5℃、每分钟约-1℃至-6℃、每分钟约-1℃至-7℃、每分钟约-1℃至-8℃、每分钟约-1℃至-9℃、每分钟约-1℃至-10℃、每分钟约-2℃至-3℃、每分钟约-2℃至-5℃、每分钟约-2℃至-7℃、每分钟约-2℃至-8℃、每分钟约-2℃至-20℃、每分钟约-4℃至-10℃、每分钟约-4℃至-8℃、每分钟约-4℃至-6℃、每分钟约-6℃至-10℃、每分钟约-6℃至-9℃、每分钟约-6℃至-8℃、每分钟约-6℃至-7℃、每分钟约-7℃至-10℃、每分钟约-7℃至-9℃、每分钟约-7℃至-8℃、每分钟约-8℃至-9℃,或每分钟约-9℃至-10℃。一旦细胞以该连续降温速率方式降温至约-40℃至-80℃或更低,即可将它们转移到液氮或液氮的汽相中以进一步降温至冻存温度,冻存温度通常低于冷冻溶液的玻璃化转变温度。在进一步降温至冻存温度前,可使细胞降温至约-40℃至-75℃、约-45℃至-70℃、约-50℃至-60℃、约-55℃至-60℃、约-70℃至-80℃、约-75℃至-80℃、约-40℃至-45℃、约-40℃至-50℃、约-40℃至-60℃、约-50℃至-70℃,或约-50℃至-80℃。
升温方案可包括两步升温法。在两步升温方案中,可将冻存的细胞(冻存在冻存温度)从冻存冰箱中移出。两步方案的第一步中,使冻存细胞在第一环境中经历第一次缓慢的升温。该环境不需要经受任何特殊的处理或具有任何具体的修饰,并且可以使用任何环境。该环境可以是气体气氛,例如空气。为了实现第一阶段的缓慢升温,可使该环境处于高于冻存温度的第一升温温度。第一升温温度可以接近室温。例如,可以使用30℃或更低,例如约15℃至30℃、约20℃至25℃,或者约为20℃至30℃的温度。
两步升温步骤中的第二个步骤包括在第二升温温度的第二环境中快速解冻细胞,第二升温温度高于第一升温步骤中所使用的升温温度。第二升温温度可以是32℃或更高、约32℃至50℃、约35℃至45℃、约40℃至50℃、约45℃至50℃、约32℃至40℃、约35℃至40℃,或约37℃。同样,任何合适的环境,例如气体(空气)、液体或流化床可用作第二环境。例如,可以使用升温温度的水浴来实现该快速解冻。
附图5说明两步升温策略改善了细胞活力和附着。ecm蛋白的添加促进了细胞活力和附着,并且有效的ecm组合可取决于细胞类型。两步升温策略可应用于像贴壁细胞一样在固体基质上冻存的所有细胞类型。
某些ecm组分和/或ecm组分的某些组合可增强一种类型细胞的活力和滞留,但不能增强另一种类型的细胞。因此,本发明主题还涉及鉴定一种或多种ecm组分的方法,所述ecm组分能够改善细胞在从冻存状态解冻后在基质上细胞活力和滞留。该方法涉及,首先选择一种特定细胞类型的细胞,例如bce细胞或人间充质干细胞。方法还包括鉴定已知与所选细胞类型相关联或由所选细胞类型生成的ecm组分。或者,在缺乏与所选细胞类型相关联的特定ecm组分的现有知识的情况下,可筛选已知的ecm组分。然后,可用ecm组分和ecm组分的各种组合处理基质,并且将所选细胞类型的细胞铺板于各种经处理的基质上,然后如上所述,将细胞在经处理的基质上冻存并随后解冻。一旦细胞已经解冻,即可评估细胞的活力和滞留,以鉴定何种ecm组分或ecm组分的组合导致为60%或更高,例如80%或更高的细胞活力(%),以及所选细胞类型的至少80%的dna含量(%)。可以根据任何已知的方法来评估细胞活力和滞留。已知的用于测量细胞活力(%)的方法包括活力实验,例如基于荧光的实验。已知的测量dna含量(%)的方法包括用于提供细胞数量准确测量结果的细胞增殖实验。
也可以鉴定与特定细胞类型的ecm组分协作最好的冷冻保护剂,因为冷冻保护剂化合物的选择可影响冻存后的细胞活力和滞留。例如,甘油和dmso促进体外微管的组装,然而其他冷冻保护剂,例如丙二醇,而不是dmso,在卵母细胞中解聚肌动蛋白。参见例如,vincent等,“在兔胚胎单细胞冻存期间冷冻保护剂对肌动蛋白丝的影响”("effectsofcryoprotectantsonactinfilamentsduringthecryopreservationofone-cellrabbitembryos"),cryobiology,27:9-23(1990)。在一些细胞中,dmso导致应力纤维的解体以及胞内肌动蛋白束的形成。所以,冻存附着于基质的细胞的重要方面是在冷冻和解冻的严苛条件后细胞保留在基质上的能力。
实施例在下文中列出,并且说明可用于实施具体实施方式中的不同方法和条件。对本领域技术人员来说多种替代方式、改进和变化方式是显而易见的。因此,实施例仅仅是说明性的而不是限制性的。
实施例
实施例1-在ecm对比tcp上的细胞冻存的细胞活力与滞留
分化的细胞系,bce细胞(bce细胞系,atcc#crl-2048)维持在37℃、5%二氧化碳下的杜尔贝科改良的伊格培养基(dmem)中,dmem具有10%胎牛血清(fcs)、1.0mm丙酮酸钠,以及4mm谷氨酰胺,以及青霉素(100u)/链霉素(100μg/ml)。bce细胞铺板于组织培养塑料上或天然bce细胞衍生的ecm上。为了制得ecm,将bce细胞以接近融合密度(50,000个细胞/孔)铺板于96孔板中,并在具有10%fcs和82μg/ml抗坏血酸磷酸镁(购自日本和光纯药工业株式会社(wakochemical))的dmem中置留6天。细胞培养基中的抗坏血酸促进ecm的沉积和形成。6天以后,通过加入水中的25mmnh4oh溶液,之后水洗3次,移出细胞。参见例如,roemer等,“炎性细胞介导的间质胞外基质的损伤的体外实验系统”("invitroassaysystemsforinflammatorycell-mediateddamagetointerstitialextracellularmatrix"),invitrotoxicol.,7(2):75-81(1994)。
随后使铺板的bce细胞随后暴露于2mdmso,并通过如下方式冻存:将板置于冰上并用0.5m甘露醇预处理细胞以使细胞准备好暴露至预期在添加冷冻保护剂(例如2mdmso)时使用的高渗环境。在添加终浓度的冷冻保护剂后,以可控速率使板降温至-80℃,然后置于-130℃。使用以下可控降温速率:-0.2℃/分钟,-1.0℃/分钟,-10.0℃/分钟,和修改的-1.0℃/分钟,该概况(mp)包括在成核步骤中。在至少储存24小时之后,根据campbell'531(同上)中的两步升温方案,将板升温。首先,将板置于-20℃,30分钟,然后将板置于37℃以快速解冻。一旦板解冻(无可见冰),就将其置于冰上,并用加有10%fcs的dmem中的0.5m甘露醇反复洗涤孔。在用加有10%fcs的dmem另外洗涤后,细胞在37℃于dmem(10%fcs)中放置1小时,回收,然后评估细胞活力和滞留。解冻后,分别使用alamar
用alamar
通过
如附图1中说明的,无关于所用的降温速率,将细胞铺板于其本身天然的ecm上而不是tcp上时,细胞活力显著提高(p<0.001)。除了-10.0℃/分钟,对于所有降温速率的细胞滞留,铺板于ecm上均等同或优于铺板于tcp上的细胞。然而,在-10.0℃/分钟的降温速率下,铺板于其ecm上的细胞比铺板于tcp上的细胞就细胞滞留而言有显著改善(p<0.0001)。附图1说明相较于较高的降温速率,较低的降温速率普遍提供更好的活力和附着(p<0.001)。尽管在所有降温速率进行冻存的过程中,ecm的存在均改善细胞活力,但是在以较高降温速率进行冻存的过程中,ecm的存在增强了细胞滞留。在较低的降温速率下,铺板于ecm和tcp上细胞的细胞滞留是等同的。一种可能的解释是存在于细胞培养基中所用的血清中的ecm蛋白(例如玻连蛋白),产生基础的基质,由此提供在冻存期间供于细胞锚着的框架。由于这种可能性,后续的实施例(2-4)是在无血清的条件下实施的。
完全形成和组织的ecm的存在提供更近似于细胞的天然环境的表面。因此,细胞附着可能较少地受冻存步骤影响。此外,ecm组合物可能影响细胞在冻存期间维持附着的能力。而且,通过ecm来冻存细胞,细胞所处的构造可能更有利于其整体健康并且可能改善其应对冷冻温度的耐受力和恢复力。
实施例2-用ecm组分的不同组合冻存的bce细胞的活力和滞留
bce细胞(bce细胞系,atcc#crl-2048)保存在37℃、5%二氧化碳下的杜尔贝科改良的伊格培养基(dmem)中,dmem具有10%胎牛血清(fcs)、1.0mm丙酮酸钠,以及4mm谷氨酰胺,和青霉素(100u)/链霉素(100μg/ml)。
确定ecm组分——纤连蛋白、层粘连蛋白、胶原i、胶原iii、胶原iv和胶原v——的最佳浓度,然后以单一组分或与一种或多种其它组分的组合形式进行评估。ecm蛋白从商业来源获得,并且在铺板细胞之前,使用浓度约为1-10μg/ml的ecm蛋白处理微量滴定板。在磷酸盐缓冲盐水(pbs)中,用各种浓度的ecm蛋白处理各孔,并在37℃放置2小时,然后根据本领域已知的方法,在37℃用1%牛血清白蛋白(bsa)孵育1小时,以封闭未被占据的结合位点。参见例如,underwood,“胞外基质分子对牛内皮细胞体外性能的影响”("theeffectofextracellularmatrixmoleculesontheinvitrobehaviorofbovineendothelialcells"),exp.cell.res.,205:311-319(1993)。孔用无血清培养基漂洗,然后置于培养基中,直到接种细胞。
如实施例1中所述,冻存细胞,随后解冻。如实施例1中,使用alamar
如附图2和3所示,ecm组分的几种组合表现出极好的活力和附着。细胞活力和百分比指示细胞数量的dna含量是基于与处理的细胞同时铺板的分开的未处理的对照细胞计算的。数据的统计分析没有鉴定出任何一种ecm组分组合是最显著的。相反,认为几组ecm组合显著优于其余组,并且取决于所使用的冷冻保护剂(p<0.01)。注意到的趋势包括,更好的活力和附着随着更复杂的ecm组分混合物(4种或更多)出现。附图2说明了在冻存和后续的解冻后,ecm蛋白对显著提高细胞活力。对于活力而言,最佳组合包括纤连蛋白和胶原i,或对于附着而言,包括纤连蛋白和胶原v,其中层粘连蛋白至少有助于改善bce细胞的活力和附着。4种ecm组分的几种组合(f+l+ci+civ、f+ciii+civ+cv,以及ci+ciii+civ+cv)导致近乎100%细胞在冻存后存活并附着。
附图3说明,甚至是单一ecm蛋白的存在都能改善细胞在单独的tcp上的附着。对于所有测试的ecm组合,包括由bce细胞产生的基质,细胞附着>80%。通过用至少一种ecm组分冻存细胞,观察到细胞附着的总体改善。
实施例3-用ecm组分的不同组合冻存的hmsc的细胞活力和滞留
将源于人骨髓的间充质干细胞维持在37℃、5%二氧化碳下的dmem/f12培养基中,所述培养基具有10%fcs、非必需氨基酸,2mmglutamax,以及青霉素(50u)/链霉素(50μg/ml)。
初始实验评估ecm组分——纤连蛋白、层粘连蛋白、胶原i、胶原iii、胶原iv和胶原v——以分别确定最佳浓度。然后,使用就个体蛋白质建立的浓度研究蛋白质对。基于由个体蛋白质和不同的蛋白质对获得的结果来选择并研究三种或四种蛋白质的进一步组合。ecm蛋白从商业来源获得,并且在铺板细胞之前,使用浓度约为1-10μg/ml的ecm蛋白处理微量滴定板。在磷酸盐缓冲盐水(pbs)中,用不同浓度的ecm蛋白处理单个孔,并在37℃放置2小时,然后在37℃用1%牛血清白蛋白(bsa)孵育1小时,以封闭未被占据的结合位点。孔用无血清培养基漂洗,然后留置于培养基中,直到铺板细胞。
如实施例1中所述,冻存细胞,随后解冻。解冻后,检测并比较附着于tcp或明胶(其常规用于hmsc的常规生长和维持)上的细胞的活力,以确定哪些ecm组分在冻存期间为细胞的持续活力和附着提供最多支持。在这些实验中,细胞活力百分比是基于冻存前每个孔中的细胞的活力来计算的。如实施例1中所述,使用alamar
虽然显现的是多于一种的ecm蛋白的组合能改善bce细胞的细胞活力,但对于hmsc而言并不一定是这种情况。单个ecm组分将hmsc的活力改善至与2种或更多ecm组分的组合相似的程度。4种或更多种ecm蛋白的组合对bce细胞而言效果最好,但hmsc对于产生最佳活力的组合有明确的偏好。ecm组分的两种组合产生大于70%的活力,两种组合为f+l+ci+cv和civ+cv。另一组组合产生大于60%的活力,并且包括:f+l+cv、l+civ+cv、ciii+civ+cv、f+l+civ+cv、f+ci+civ+cv、f+ciii+civ+cv和l+ci+ciii+cv。有趣的是,通常用于hmsc细胞培养的明胶没有促进冻存和解冻后的细胞活力。这说明为特定细胞类型建立的基质处理方法不一定适用于冻存。这也在包括基质细胞蛋白时观察到(参见实施例4)。一般来说,至少一种ecm蛋白的存在能改善冻存后hmsc的活力。那些表现出最佳活力的组合趋于在其混合物中包括纤连蛋白、胶原iv和/或胶原v。
实施例4-用基质细胞蛋白,生腱蛋白x冻存后hmsc的活力
源于人骨髓的间充质干细胞铺板于明胶或层粘连蛋白、胶原i、胶原iii和胶原v的ecm组合上。附图6所示样品,在细胞铺板时,包含浓度为500ng/ml的细胞间质蛋白,生腱蛋白x。细胞冻存在含有2.5%硫酸软骨素的1mdmso中,随后解冻,如实施例1所述。解冻后,测量细胞活力,并相对于未处理的对照进行标准化。相较于ecm单独存在的情况(57%),生腱蛋白x的添加促进细胞活力的改善(100%),并且在ecm±生腱蛋白x上的hmsc的整体活力优于细胞在明胶上铺板时的情况(p<0.05,附图1)。有趣的是,与ecm组合相比,生腱蛋白x与明胶一起使用时没有显示出任何改善,可能是因为明胶是胶原的加工形式,而组合中使用的ecm组分都作为天然蛋白质产生。这进一步强调,为特定细胞类型培养建立的基质处理方法不一定在贴壁细胞冻存期间起作用。
本文所示分化细胞系和间充质干细胞的结果说明,ecm组分的添加能改善活力,并维持冻存后细胞贴附于多孔板的附着。观察到两种类型细胞间的差异,这强调了ecm可在体外和体内细胞的健康和维持中起关键作用。这对干细胞尤为重要,因为ecm可影响其分化能力以及它们将分化为何种细胞谱系。这些结果也说明ecm的组合物影响所述细胞及其作为贴壁群体在冻存中存活的能力。
应理解以上内容的各种变形以及其他特征和功能或其变形,可与许多其他不同的系统或应用结合。本领域技术人员也可进行各种目前未预见或未预料到的替代方案、变形、变化或改进,它们也包括在所附权利要求书中。
- 还没有人留言评论。精彩留言会获得点赞!