大豆转录因子GmAREB3在植物油脂代谢调控中的应用的制作方法
本发明涉及生物技术领域,尤其涉及一种大豆转录因子gmareb3在植物油脂代谢调控中的应用。
背景技术:
人类饮食中71%的油脂来自于植物。在世界上的几种主要产油作物中,大豆总产油量约占30%,居世界植物油产量的第一位(表1)。
表1为世界上主要的产油作物
脂肪酸的合成是植物体中最重要的代谢途径之一,它存在于植物体的任何一个细胞中,是生长发育所必须的。对它的阻断会导致细胞的死亡,因而至今为止还没有发现一个阻断脂肪酸合成的植物突变体。
植物同其它真核生物在参与脂肪酸合成途径的酶上有很大差异。从乙酰coa和丙二酰coa合成16或18个碳原子的脂肪酸至少需要30个不同的酶催化的反应来完成这一过程,而在动物、真菌及一些细菌中,以上反应是由一个存在于胞质中的多酶复合体来完成的。植物中,参与脂肪酸合成的酶以可溶的形式存在于质体的胞质中。
大多数植物中,油脂都以三酰甘油(triacylglycerols,tag)的形式储藏,它的含量是一个非常重要的农艺性状,tag的生物合成称之为kennedy途径,如同真核生物中合成膜甘油酯的途径,脂肪酸去除coa后被转移到3-磷酸甘油的1和2位,形成中间产物pa。pa去磷酸化产生dag。在tag合成的最后一步,第三个脂肪酸分子被转移到空的dag3’-oh位置,这一步反应是由二酰甘油乙酰转移酶(diacylglycerolacyltransferase,dgat)催化的,此反应被认为是tag生物合成中唯一的限速步骤。人们已对脂类合成途径有了认知,并且已经克隆了很多参与脂类合成的酶基因。然而,植物中,对脂类合成的调控机理及其相关基因仍然知之甚少。
植物中,bzip类转录因子是一个大家族,在植物中其成员高达100多个,又可分为几个亚族,其功能很不相同,在植物的生长、发育中起重要作用。申请人实验室早期聚类了大豆基因组中的131个bzip类转录因子,进而发现,gmbzip44、gmbzip62和gmbzip78三个基因应答非生物胁迫,并参与耐盐、耐低温调控。它们的过表达提高了转基因植株的耐盐和耐低温。之后,申请人在大豆中克隆了bzip类转录因子基因gmbzip123的编码基因gmbzip123,同源聚类分析发现,gmbzip123基因与拟南芥s1亚族bzip类转录因子的同源关系最近,因此属于s族。已经报道,bzip家族中的许多a亚族基因参与植物对于aba的响应,如abi5、areb1/2等,参与种子成熟干燥、萌发、休眠以及环境胁迫反应。拟南芥中areb3编码aba响应元件结合蛋白,是aba不敏感类蛋白,定位在细胞核内,尚未报道其与油脂代谢相关。鉴于大豆基因组序列已经完成,gmareb3基因序列与glyma06g47220.2一致,启动子分析显示其为aba响应元件结合因子,但是没有功能的描述。大豆gmareb3与拟南芥areb3的蛋白质序列相似性小于20%,因此大豆gmareb3的功能尚属未知。
技术实现要素:
本发明的一个目的是提供如下1)-3)中任一种物质的新用途。
本发明提供的如下1)-3)中任一种物质在调控植物组织总油脂含量和/或调控植物组织中脂肪酸含量中的应用或在培育油脂和/或脂肪酸含量提高的植物中的应用:
1)蛋白gmareb3;
2)编码蛋白gmareb3的dna分子;
3)含有编码蛋白gmareb3的dna分子的重组载体、表达盒、转基因细胞系或重组菌;
所述蛋白gmareb3为如下(1)或(2):
(1)由序列表中序列2所示的氨基酸序列组成的蛋白质;
(2)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的由(1)衍生的蛋白质。
上述应用中,所述编码蛋白gmareb3的dna分子是如下1)至4)中任一所述的dna分子:
1)序列表中序列1所示的dna分子;
2)序列表中序列1第1-948位所示的dna分子;
3)在严格条件下与1)或2)所限定的dna分子杂交且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子;
4)与1)或2)所限定的dna分子至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子。
所述含有编码蛋白gmareb3的dna分子的重组载体为将所述编码蛋白gmareb3的dna分子插入表达载体中,得到表达蛋白gmareb3的重组载体。在本发明的实施例中,表达载体为pgwb412,重组载体为将序列表中序列1所示的核苷酸插入载体pgwb412,得到重组载体;采用同源重组的方式插入,具体方法见实施例。
上述应用中,所述调控植物组织总油脂含量为提高植物组织总油脂含量;
所述调控植物组织中脂肪酸含量为提高植物组织中脂肪酸含量;
所述脂肪酸为棕榈酸、油酸、亚油酸和/或贡多酸。
上述总油脂含量通过种子中脂类重量与种子总重量的百分比体现。
上述应用中,所述组织为种子;所述植物为单子叶植物或双子叶植物。所述双子叶植物具体为豆科植物或十字花科植物。在本发明的实施例中,双子叶植物为拟南芥。
本发明的另一个目的是提供一种培育油脂和/或脂肪酸含量提高的转基因植物的方法。
本发明提供的方法,为将编码蛋白gmareb3的dna分子导入目的植物中,得到转基因植物;所述转基因植物组织中总油脂含量和/或脂肪酸含量高于所述目的植物;
所述蛋白gmareb3为如下(1)或(2):
(1)由序列表中序列2所示的氨基酸序列组成的蛋白质;
(2)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的由(1)衍生的蛋白质。
上述总油脂含量通过种子中脂类重量与种子总重量的百分比体现。
上述方法中,所述脂肪酸为棕榈酸、油酸、亚油酸、亚麻酸和/或贡多酸;
所述编码蛋白gmareb3的dna分子通过重组载体导入所述目的植物中;
所述编码蛋白gmareb3的dna分子是如下1)至4)中任一所述的dna分子:
1)序列表中序列1所示的dna分子;
2)序列表中序列1第1-948位所示的dna分子;
3)在严格条件下与1)或2)所限定的dna分子杂交且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子;
4)与1)或2)所限定的dna分子至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子。
所述重组载体为将所述编码蛋白gmareb3的dna分子插入表达载体中,得到表达蛋白gmareb3的重组载体。在本发明的实施例中,表达载体为pgwb412,重组载体为将序列表中序列1所示的核苷酸插入载体pgwb412,得到重组载体;采用同源重组的方式插入,具体方法见实施例。在本发明的实施例中,应用invitrogen公司生产的gateway系统的
上述方法中,所述组织为种子;所述植物为单子叶植物或双子叶植物。在本发明的实施例中,双子叶植物具体为豆科植物或十字花科植物,双子叶植物为拟南芥。
本发明的第三个目的是提供一种重组载体。
本发明提供的重组载体,为将编码蛋白gmareb3的dna分子插入表达载体中,得到表达蛋白gmareb3的重组载体;
所述蛋白gmareb3为如下(1)或(2):
(1)由序列表中序列2所示的氨基酸序列组成的蛋白质;
(2)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的由(1)衍生的蛋白质。
上述重组载体中,所述编码蛋白gmareb3的dna分子的核苷酸序列为序列表中的序列1。
上述重组载体是将上述基因插入表达载体中,得到表达上述蛋白的重组载体。在本发明的实施例中,表达载体为pgwb412,重组载体为将序列表中序列1所示的核苷酸插入载体pgwb412,得到重组载体;采用同源重组的方式插入,具体方法见实施例。
上述用于构建所述植物表达载体的出发载体可为任意一种双元农杆菌载体或可用于植物微弹轰击的载体等,如pbin438、pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa或pcambia1391-xb(cambia公司)。使用gmareb3构建植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,如花椰菜花叶病毒(camv)35s启动子、泛生素基因ubiquitin启动子(pubi)等,它们可单独使用或与其它的植物启动子结合使用;此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、具有抗性的抗生素标记物(庆大霉素标记物、卡那霉素标记物等)或是抗化学试剂标记基因(如抗除莠剂基因)等。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
携带有本发明gmareb3的植物表达载体可通过使用ti质粒、ri质粒、植物病毒载体、直接dna转化、微注射、电导、农杆菌介导等常规生物学方法转化植物细胞或组织,并将转化的植物细胞或组织培育成植株。被转化的植物宿主既可以是单子叶植物,如水稻、小麦、玉米等,也可以是双子叶植物,如大豆、烟草、拟南芥或棉花等。
本发明的实验证明,本发明提供了将转录因子gmareb3转入野生型拟南芥中,得到转基因拟南芥,该转基因拟南芥与野生型拟南芥相比,其种子中的油脂含量和部分脂肪酸含量(如棕榈酸、油酸、亚油酸和/或贡多酸)均提高。说明转录因子gmareb3及其编码基因可以调控植物种子中油脂和/或脂肪酸含量,过表达后提高植物种子中油脂和/或脂肪酸含量。该基因对提高和改良作物油脂成份、特别是对于提高大豆等油料植物种子中油脂成份,培育高油脂品种具有重要的理论和现实意义。
下面结合具体实施例对本发明做进一步详细说明。
附图说明
图1为克隆载体和植物表达载体示意图。
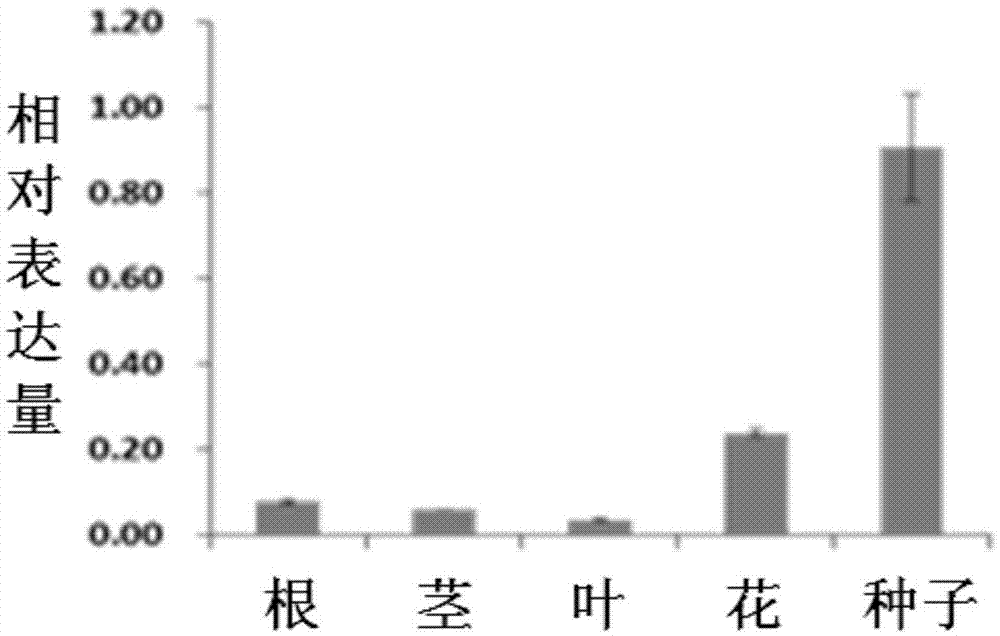
图2为gmareb3在大豆不同器官的表达分析。
图3为转gmareb3植株纯系的分子鉴定。
图4为转gmareb3植株种子中油脂含量测定。
图5为转gmareb3植株种子中脂肪酸含量测定。
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中所用引物均由三博生物公司合成。
大豆黑农44(hn44)记载在如下文献中:满为群等,大豆新品种黑农44的选育及不同种植方式对其产量和品种的影响,黑龙江农业科学2004年5期,1-5;公众可从中国科学院遗传与发育生物学研究所获得;该大豆2006年获自黑龙江农业科学院大豆研究所;由黑龙江农业科学院大豆研究所2002年经黑龙江省农作物品种审定委员会审定的大豆品种,第一育成人为杜维广研究员,专利号为:cna20020216.2,审定号为:黑审豆2002003。
表达载体pgwb412记载在如下文献中:departmentofmolecularandfunctionalgenomics,shimaneuniversity,aatsue,shimane690-8504,japan,e.mail:tnakagaw@life.shimane-u.ac.jpisuyoshinakagawa,etal.,gatwayvectorsforplanttransformation,plantbiotechnology,2009,26,275-284。由tsuyoshinakagawa博士提供,公众得到tsuyoshinakagawa博士同意后可从中科院遗传与发育生物学研究所获得。
农杆菌gv3101,公众可从中国科学院遗传与发育生物学研究所获得,记载在如下文献中:leecw等,agrobacteriumtumefacienspromotestumorinductionbymodulatingpathogendefenseinarabidopsisthaliana,plantcell,2009,21(9),2948-62。
实施例1、大豆转录因子gmareb3的cdna克隆和植物表达载体的构建
1、转录因子gmareb3的获得
在进行大豆种子发育过程中脂肪酸积累代谢的分析时获得一个bzip转录因子家族中gmareb3基因有较高的表达。因此对该基因与油脂合成调控的相关性进行了研究。
提取大豆黑农44幼苗的总rna,将rna用逆转录酶反转录合成cdna。
根据在plantgdb的大豆基因组序列中gmareb3全长cdna序列的信息,设计引物,引物序列如下:
gmareb3-up:5’-atggggtctcaatgtggtggtg(序列3);
gmareb3-dp:5’-gaatgaagccgatgatgttcggc(序列4)。
以黑农44的cdna为模板,用gmareb3-up和gmareb3-dp为引物,进行pcr扩增,得到约1kb的pcr产物。经过测序,该pcr产物为951bp,其具有序列表中序列1所示的核苷酸,该核苷酸所示的基因为gmareb3,编码区为序列1第1-951位;该基因编码的蛋白命名为gmareb3,该蛋白含316个氨基酸,其氨基酸序列为序列表中的序列2。
2、植物表达载体的构建
基因克隆使用invitrogen公司提供的gateway系统,载体3′-t突出端,用于直接连接taq酶扩增的pcr产物。
将上述1得到的951bp的pcr产物运用ta克隆的原理克隆与载体
由于
具体方法如下:1ul
重组载体经过测序,该载体为将序列表中序列1所示的dna分子同源重组到载体pgwb412中得到的载体,命名为pgwb412-gmareb3(部分结构示意图如图1b)。
3、gmareb3在大豆不同器官的表达分析
取大豆黑农44的根、茎、叶、花和种子的总rna,用逆转录酶反转录合成cdna。引物为:5’-ggaatctgctgcccgttcta和5’-tgatgttcggcgaatctggt,进行realtime-pcr鉴定。大豆tublin基因为内标,所用引物为primer-tf:5’-aacctcctcctcatcgtact,和primer-tr:5’-gacagcatcagccatgttca-3’。
以野生型拟南芥(col-0)(种子购自arabidopsisbiologicalresourcecenter(abrc))为对照。
结果如图2所示,在根、茎、叶和花中几乎检测不到gmareb3基因的转录,而在种子中其表达量很高,因此gmareb3是种子特异表达的基因。
实施例2、gmareb3在调控植物组织总油脂或脂肪酸含量中的应用
一、重组农杆菌的获得
将实施例1的2得到重组载体pgwb412-gmareb3用电击法导入农杆菌gv3101,得到重组菌。
提取重组菌的质粒,经过测序,该质粒为pgwb412-gmareb3,将含有该质粒的重组菌命名为gv3101/gmareb3,即为重组农杆菌。
二、转gmareb3拟南芥的获得及鉴定
将重组农杆菌gv3101/gmareb3培养至对数期,然后用抽真空法将其转化哥伦比亚生态型拟南芥(col-0)(种子购自arabidopsisbiologicalresourcecenter(abrc))花中,经培育后收获种子,将种子播于含卡那霉素(50mg/l)的ms筛选培养基上,待筛选得到的t1代植株长至6叶时移到蛭石上生长,收获t1代单株,各单株种子分别播种,用相同的ms筛选培养基继续筛选以观察t2代的分离情况,如此重复数代直至获得遗传稳定的转基因纯合株系,获得14个t5代转gmareb3拟南芥纯系。
提取上述获得的14个t5代转gmareb3拟南芥株纯系苗的rna,反转录得到cdna作为模板,引物为:5’-ggaatctgctgcccgttcta和5’-tgatgttcggcgaatctggt,进进行realtime-pcr鉴定。拟南芥atactin2基因为内标,所用引物为primer-tf:5’-atgcccagaagtcttgttcc,和primer-tr:5’-tgctcatacggtcagcgata-3’。以野生型拟南芥(col0)为对照。实验重复三次,结果取平均值±标准差。
gmareb3表达量的结果如图3所示,野生型拟南芥(col0)中未能检测出gmareb3的表达量;t5代转gmareb3拟南芥株系中均能检测出gmareb3的转录,oe-2、oe-3、oe-7、oe-17和oe-18中gmareb3的相对表达量约为0.095、0,051、0.117、0,083和0.086。上述结果证明,gmareb3转入拟南芥中且得到表达,证明编号为oe-2、oe-3、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥为阳性转基因植株。
选取编号为oe-2、oe-3、oe-7、oe-17和oe-18五个t5代转gmareb3拟南芥纯系做进一步功能检测。
采用同样的方法将空载体pgwb412转入野生型拟南芥中,得到t0代转空载体拟南芥,播种、收种,直到得到t5代转空载体拟南芥。
三、转gmareb3基因拟南芥的表型分析
测定野生型拟南芥(col0)、t5代转空载体拟南芥、编号为oe-2、oe-3、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥的种子总油脂含量。
具体方法如下:
种子总油脂含量测定:将干燥的种子研磨成粉,称取100mg到离心管中,平行称取四份。加入500μl的正己烷,充分混匀,37℃过夜。慢速离心3分钟,将正己烷吸入称量过新管中。剩下的粉末继续加正己烷重复浸泡、然后离心、然后收集正己烷到同一的离心管中。将离心管放入真空泵中,抽真空,使正己烷完全挥发。然后再次称取离心管的重量。离心管前后重量的变化即是提取的脂类重量;总油脂量(%)的计算公式为总油脂量(%)=(提取的脂类重量/种子总重量)x100%。
每个株系取30株的种子,实验重复三次,结果取平均值±标准差。
结果如图4所示,
野生型拟南芥种子总油脂量为34.2±0.1%(即为种子总重量的百分比),野生型拟南芥和转空载体拟南芥的结果无显著差异。
t5代转gmareb3拟南芥株系oe-2、oe-3、oe-7、oe-17和oe-18种子总油脂量约为36.0±0.4%;34.5±0.4%;35.7±0.4%;34.8±0.4%和34.6±0.3%。其中oe-2和oe-7的种子油脂含量极显著高于对照,oe-17和oe-18种子总油脂量显著高于对照,而oe-3种子总油脂量仅略高于对照。
结果表明,4个转基因株系种子中的总油脂含量明显高于对照。
上述实验表明,大豆bzip类转录因子gmareb3对种子中总油脂的合成呈正调控作用,其编码基因gmareb3的过量,可提高转基因植株种子中总油脂的含量。
各植株的种子中脂肪酸含量的检测:彻底干燥待测种子,研磨成粉,取10mg加入螺口的2ml离心管中,每份样品平行称取四份。加入10μl的17:0脂肪酸(10mg/ml)做内标。加含2.5%浓硫酸的甲醇溶液1ml,85℃水浴中保温1小时,期间晃动数次。自然冷却后,取上清500μl到新管中,加入600μl的0.9%nacl溶液、300正己烷,震荡混匀几分钟,4000转离心10分钟,取上清至新管中。通风橱中过夜使正己烷挥发完全,然后加入50μl乙酸乙酯溶解甲酯化的脂肪酸。将甲酯化的脂肪酸样品用气相色谱质谱联用仪测(perkin-elmerturbomass)各个组分的相对含量,然后各个成分的脂肪酸与加入的十七烷酸17:0内标(sigma,51610)比较得出相对含量(方法可以参见:shen,b.,etal.,thehomeoboxgeneglabra2affectsseedoilcontentinarabidopsis,plantmol.biol.,60,377-387,2006)。每个株系取30株的种子,实验重复三次,结果取平均值±标准差。
结果如图5所示,其中编号为oe-2、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥种子中芥子酸(22:1)的含量与野生型拟南芥相比均无明显差异;亚麻酸(18:3)和硬脂酸(18:0)的含量比对照低,但是总油脂量是显著上升的。
野生型拟南芥(col0)、编号为oe-2、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥种子中棕榈酸(16:0)占种子总重量的百分比分别约为2.8%、3.2%、3.3%、3.1%和3.2%;
野生型拟南芥、编号为oe-2、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥种子中油酸(18:1)约为4.3%、5.3%、5.3%、5.4%和5.5%;
野生型拟南芥、编号为oe-2、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥种子中亚油酸(18:2)约为10.0%、12.1%、11.4%、11.1%和11.3%;
野生型拟南芥、编号为oe-2、oe-7、oe-17和oe-18的t5代转gmareb3拟南芥种子中贡多酸(20:1)约为5.1%、6.6%、6.2%、6.1%和6.3%。
上述实验表明,大豆bzip类转录因子gmareb3对种子中油脂的合成呈正调控作用,其编码基因gmareb3的过量表达,可提高转基因植株种子中总油脂的含量和一些脂肪酸如棕榈酸(16:0)、油酸(18:1)、亚油酸(18:2)和贡多酸(20:1)的含量。
- 还没有人留言评论。精彩留言会获得点赞!