一种人源化单克隆抗体及应用的制作方法
本发明涉及一种人源化单克隆抗体及应用,属于医药
技术领域:
。
背景技术:
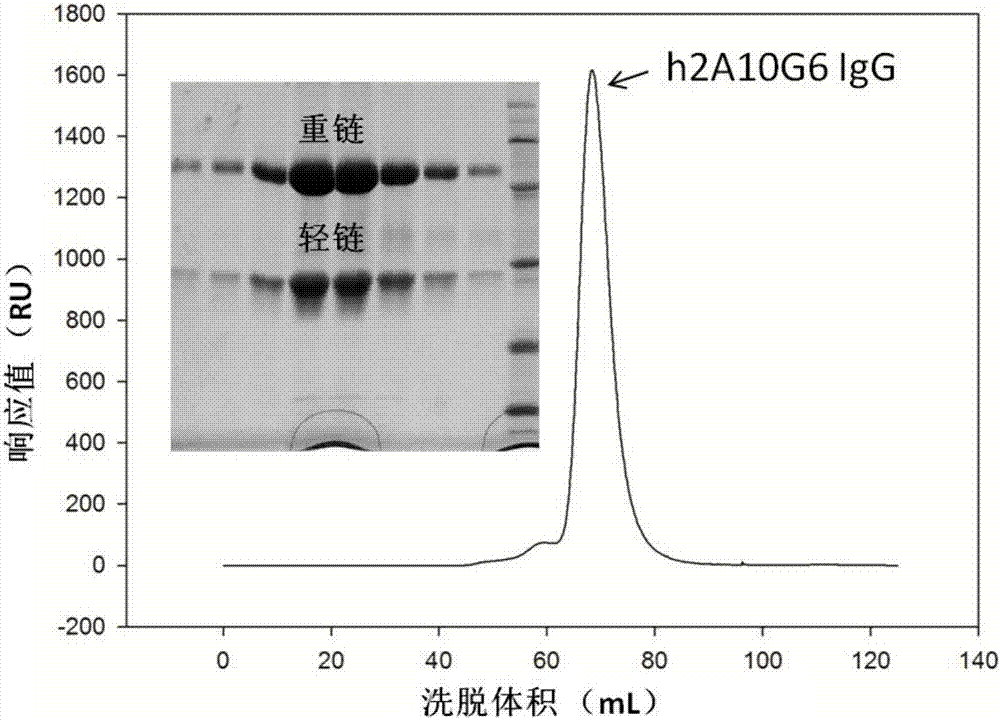
:黄病毒是一类囊膜单股正链rna病毒,包括寨卡(zikv)、登革热(denv)、西尼罗热(wnv)、黄热(yfv)等对人有致病力的病毒。这些病毒主要由蚊虫传播,感染之后对人产生严重的后果。黄热病毒感染主要引起发热、肌肉疼痛(腰痛和下肢疼痛等)、头痛、寒战、食欲不振、恶心和呕吐,严重的产生高热、黄疸腹痛并伴有呕吐,甚至死亡;登革热感染则引起高热、头疼、肌肉骨关节疼痛等临床表现,是东南亚地区儿童死亡的主要原因之一;西尼罗热病毒感染严重时将引起无菌性脑膜炎,甚至死亡。这几种病毒都尚缺乏有效的疫苗和治疗手段。中和抗体特异性高、能阻断病毒的入侵,是治疗病毒感染的有效手段。黄病毒囊膜蛋白e蛋白介导病毒与宿主受体的结合和入侵,是病毒最主要的保护性抗原。大部分的中和抗体都识别黄病毒的e蛋白。x-射线晶体学结构显示黄病毒e蛋白有三个结构域:di、dii和diii。一般认为diii负责介导病毒与受体的结合,大多数有效的中和抗体都是识别diii的中和表位。dii头部(98-110位氨基酸)包含一个高度保守的融合区(fusionloop),在病毒入侵的膜融合过程中起关键作用。diii在黄病毒中保守程度不高,而dii的融合区保守度很高,因此有报道黄病毒的广谱的中和抗体通过识别dii的融合区表位,从而中和不同的黄病毒。鼠单克隆抗体2a10g6是一个黄病毒广谱的中和抗体,能够识别e蛋白的融合区表位,结合并中和登革热病毒1-4型、黄热病毒、西尼罗热病毒。由于2a10g6为鼠单克隆抗体,尚不能应用于临床上,所以抗体亟需进行改进以使其能够获准临床使用。因此,亟需找到一种有效的能够获准临床使用的具有防治黄热病毒,登革病毒各血清型以及西尼罗热病毒的中和抗体。目前的鼠单克隆抗体的人源化改造,一般是保留cdr区,而其他部分都由鼠的替换成人的。然而,这样替换常常会导致抗体的构想发生变化,进而导致亲和力下降甚至消失。技术实现要素:为了解决上述问题,本发明对鼠单克隆抗体2a10g6进行人源化改造,通过杆状病毒表达,获得人源化抗体h2a10g6。本发明的h2a10g6抗体,对黄热病毒,登革热和西尼罗热病毒的高亲和力和中和活性,可应用与临床治疗和预防黄热病毒、登革热病毒以及西尼罗热等病毒。本发明的第一个目的是提供一种人源化单克隆抗体,所述抗体含有氨基酸序列为seqidno:1的重链可变区和氨基酸序列为seqidno:2的轻链可变区。在本发明的一种实施方式中,所述抗体的重链的氨基酸序列为seqidno:3、轻链的氨基酸序列为seqidno:4。在本发明的一种实施方式中,所述抗体的重链的核苷酸序列为seqidno:5、轻链的核苷酸序列为seqidno:6。本发明的第二个目的是提供所述人源化单克隆抗体在制备治疗或预防黄病毒的药物方面的应用。在本发明的一种实施方式中,所述应用,是用于制备试剂盒;所述试剂盒中含有所述抗原,或者编码所述抗原的dna分子,或者表达所述抗原的重组载体/表达盒/转基因细胞系/重组菌。在本发明的一种实施方式中,所述黄病毒,包括黄热病毒、登革热病毒1-4型以及西尼罗热等病毒。本发明的第三个目的是提供一种药物组合物,所述药物组合物含有所述的人源化单克隆抗体。在本发明的一种实施方式中,所述药物组合物还含有医学生可接受的载体。本发明的第四个目的是提供一种编码人源化单克隆抗体的基因,所述基因编码的人源化单克隆抗体的重链可变区的氨基酸序列为seqidno:1、轻链可变区的氨基酸序列为seqidno:2。在本发明的一种实施方式中,所述基因编码的人源化单克隆抗体的重链的氨基酸序列为seqidno:3、轻链的氨基酸序列为seqidno:4。本发明还要求保护含有所述基因的表达载体、表达所述基因的细胞。本发明也要求保护对含有氨基酸序列为seqidno:1的重链可变区或氨基酸序列为seqidno:2的轻链可变区的基因进行改进得到的人源化单克隆抗体,所述改造包括亲和性改造。本发明的有益效果:通过杆状病毒表达h2a10g6,纯化后可获得每升20mg以上的纯化抗体。抗体人源化后,它与黄热、登革热、西尼罗热病毒的结合能力仍然维持在40-70nm之间。通过与活病毒进行中和试验显示,人源化抗体h2a10g6有着很强的活病毒中和活性(表2)。这提示,人源化抗体h2a10g6是成功改造的黄病毒广谱中和抗体,有着临床治疗和预防黄热、登革热、西尼罗热以及其他黄病毒的应用价值。附图说明图1:2a10g6fab的分子筛分离纯化图以及收集峰sds-page凝胶电泳图;图2:h2a10g6igg的分子筛分离纯化图以及收集峰sds-page凝胶电泳图;图3:h2a10g6对黄热e蛋白的结合动力学分析;图4:h2a10g6对2型登革病毒e蛋白的结合动力学分析;图5:h2a10g6对西尼罗河病毒e蛋白的结合动力学分析。具体实施方式实施例1:鼠单克隆抗体2a10g6的人源化改造2a10g6鼠源抗体fab设计策略如下:phpromoter-bamh1-gp67-ecor1-lchain-hind3p10promoter-sma1-hbmsp-nhe1-hchain-his-kpn1鼠单克隆抗体2a10g6的fab重链氨基酸序列(vh-ch1)如seqidno:7所示、核苷酸序列如seqidno:8所示,鼠单克隆抗体2a10g6mfab轻链氨基酸序列(vl-cl)如seqidno:9所示、核苷酸序列如seqidno:10所示。将2a10g6重链和轻链v区与已报道的人源抗体氨基酸序列分别比对,按照氨基酸同源性,分别排序,选择轻重链排序都靠前的已知抗体10个,再依据cdr长度相差最小、轻重链相互作用氨基酸是否一致,结构的模拟,选择最优抗体。依据antibodyengineeringlabmanual(ed.:duebel,s.andkontermann,r.,springer-verlag,heidelberg),通过氨基酸序列分别判断2a10g6抗体与3hc4抗体的v区cdr区域,将2a抗体的cdr区域氨基酸整体替换到3hc4抗体cdr上。抗体的亚型为igg1,轻链为kappa。得到的人源化后2a10g6的重链可变区的氨基酸序列如下(seqidno:1),其中cdr区下滑线标出、2a替换3hc4氨基酸标为斜体:得到的人源化后2a10g6的轻链可变区的氨基酸序列如下(seqidno:2),其中cdr区下滑线标出:改造后的人源化抗体iggh2a10g6的重链的氨基酸序列(ch)如seqidno:3所示,核苷酸序列如seqidno:5所示。改造后的人源化抗体iggh2a10g6的轻链的氨基酸序列(vl-cl)如seqidno:4所示,核苷酸序列如seqidno:6所示。实施例2:单克隆抗体的制备与纯化鼠源抗体2a10g6和人源化抗体iggh2a10g6的制备方法如下:i)质粒构建:将鼠源抗体2a10g6的fab基因序列插入pfastbac-dual载体中,在重链的基因后面加入命名为pfastbac-dual-2amfab。将鼠源抗体2a10g6的人源化抗体igg基因插入到pfastbac-dual载体中,在基因后面加入6组氨酸标签,命名为pfastbac-dual-2ahigg。重组质粒通过pcr、酶切鉴定和测序证实插入的外源片段完全正确。ii)鼠源单克隆抗体2a10g6的fab片段2amfab蛋白和人源化抗体h2a10g6igg蛋白制备和纯化:首先,将重组质粒转化dh10bac感受态细胞,37℃过夜培养,通过蓝白斑筛选和pcr鉴定出阳性克隆,并提取重组bacmid,即已含有2amfab或2ahigg基因序列的质粒。将重组bacmid转染sf9细胞,转染3d后收取培养上清,得到p1代杆状病毒。连续扩增3代病毒即可得到大量高滴度病毒。然后感染hi5细胞,48h离心收取细胞上清,将2amfab细胞培养上清液与ni-ntaresin(ge)于4℃结合过夜。以缓冲液a(20mmtris,150mmnacl,ph8.0)洗涤resin,以去除非特异结合的蛋白。最后将目的蛋白以缓冲液b(20mmtris,150mmnacl,ph8.0,300mmimidazole)从resin上洗脱下来,并以10k截留(10kcutoff)的浓缩管将洗脱液浓缩至5ml。将2ahigg细胞培养上清液与proteina于4℃结合过夜。以缓冲液a(20mmna3po4,ph7.0)洗涤resin,以去除非特异结合的蛋白。最后将目的蛋白以缓冲液b(100mmglycine,ph3.0)从resin上洗脱下来,并以10k截留(10kcutoff)的浓缩管将洗脱液浓缩至5ml。将浓缩后的蛋白溶液进一步以分子排阻层析纯化,使用akta-purifier(ge)和superdex200hiload16/60柱子(ge),使用缓冲液a(20mmtris,150mmnacl,ph8.0,同时监测280nm的紫外吸收值,收取目的蛋白,并通过sds-page鉴定蛋白纯度。典型的2a10g6fab和h2a10g6蛋白的分子筛图谱和sds-page分析分别如图1和图2。由图1、图2可知,通过分子筛分离纯化,发明人获得了预期大小的2a10g6fab和人源化的h2a10g6igg的洗脱峰收集液。通过sds变性的聚丙烯酰胺凝胶电泳我们知道2a10g6fab我们看到fab区的重链和轻链,抗体纯度在95%以上;h2a10g6igg我们看到了全长的重链和轻链,纯度在95%以上。用于做黄病毒体外中和试验的2a10g6的制备如下:表达2a10g6的杂交瘤细胞腹腔注射balb/c小鼠,每106细胞注射一支小鼠。2周后,抽取小鼠腹水。腹水用20mmna3po4ph7.0的缓冲液对半稀释,过滤,通过proteing亲和层析柱吸附于柱子上。随后通过0.1mglycine,ph3.0洗脱收集到含有0.2mtris-hclph9.0的收集液中。及获得2a10g6全抗。用于做黄病毒结合力实验的人源化抗体h2a10g6的fab的制备如下:将全抗2a10g6igg浓缩至10mg/ml,浓缩换液为20mmna3po4,ph7.0,10mmedta。吸取0.5ml充分混合的偶联酶beans到反应容器中,1000g/min离心2min去除保存液。用3ml20mmcysteine,ph7.0清洗beans三次,弃去洗脱。然后加入0.5ml20mmcysteine,ph7.0温和颠倒重悬。给反应管加上堵头,加入浓缩好的抗体,拧紧盖子,用封口膜封住空隙。将反应管固定到旋转混合仪,37℃孵育4h,1000g/min离心5分钟,保存下液。用1ml20mmna3po4,ph7.0,10mmedta清洗beans2次,回收下液,测浓度。浓缩后,通过proteina柱,收集流穿液,即为h2a10g6的fab。本发明通过人源化改造鼠单克隆抗体2a10g6,获得h2a10g6。通过杆状病毒表达h2a10g6,纯化后可获得每升20mg以上的纯化抗体。实施例3:黄病毒蛋白的表达黄热e蛋白、登革热病毒e蛋白、西尼罗热e蛋白的表达:yfv_e胞外区dna片段(氨基酸序列如seqidno:11所示、核苷酸序列如seqidno:12所示)通过ndei和xhoi酶切后,连接到pet21a载体上。其中yfv_e蛋白编码区的3’端连上6个组氨酸标签(hexa-his-tag)的编码序列及翻译终止密码子。再将连接产物转化到bl21大肠杆菌感受态细胞。单克隆接种到40mllb培养基中,培养6-8小时。接种到4l的lb培养基中,37℃培养至od600=0.4-0.6,加入iptg至终浓度1mm,37℃继续培养4-6小时。收获包涵体,通过在稀释法复性包涵体。复性液(100mmtrisph8.0,400mml-arghcl,2mmedta,5mmgsh,0.5mmgssg,5%glycerol)浓缩后换成20mmtris,150mmnacl,ph8.0缓冲液。将浓缩后的蛋白溶液进一步以凝胶过滤层析纯化,使用akta-purifier(ge)和superdex200hiload16/60柱子(ge),使用缓冲液a(20mmtris,150mmnacl,ph8.0),同时监测280nm的紫外吸收值,收取目的蛋白,并通过sds-page鉴定蛋白纯度。将编码dv2e蛋白的dna片段(氨基酸序列如seqidno:13所示、核苷酸序列如seqidno:14所示)用pstⅰ和xhoⅰ酶切位点连入pfastbac1载体中,wnve蛋白的dna片段(氨基酸序列如seqidno:15所示、核苷酸序列如seqidno:16所示)用ecorⅰ和xhoⅰ酶切位点连入pfastbac1载体中,并在蛋白的编码片段前加入信号肽gp67有助于蛋白分泌表达,在蛋白的编码片段后插入6组氨酸标签(hexa-his-tag)的编码序列及翻译终止密码子,这样会得到一个c-端带有组氨酸标签的重组蛋白,利于后续的蛋白纯化,分别命名为pfastbac1-dv2e和pfastbac1-wnve。首先,将重组质粒转化dh10bac感受态细胞,37℃过夜培养,通过蓝白斑筛选和pcr鉴定出阳性克隆,并提取重组bacmid,即已含有dv2e或wnve基因序列的质粒。将重组bacmid转染sf9细胞,转染3d后收取培养上清,得到p1代杆状病毒。连续扩增3代病毒即可得到大量高滴度病毒。然后感染hi5细胞,48h离心收取细胞上清,将细胞培养上清液与ni-ntaresin(ge)于4℃结合过夜。以缓冲液a(20mmtris,150mmnacl,ph8.0)洗涤resin,以去除非特异结合的蛋白。最后将目的蛋白以缓冲液b(20mmtris,150mmnacl,ph8.0,300mmimidazole)从resin上洗脱下来,并以10k截留(10kcutoff)的浓缩管将洗脱液浓缩至5ml。将浓缩后的蛋白溶液进一步以凝胶过滤层析纯化,使用akta-purifier(ge)和superdex200hiload16/60柱子(ge),使用缓冲液a(20mmtris,150mmnacl,ph8.0,同时监测280nm的紫外吸收值,收取目的蛋白,并通过sds-page鉴定蛋白纯度。实施例4:黄病毒蛋白与抗体亲和力分析采用表面等离子共振技术检测yfv_e、wnv_e、dv2_e蛋白与抗体亲和力。表面等离子共振分析是用biacore3000(biacoreinc.)进行的。具体步骤如下:首先,将yfv、dv2和wnv的e蛋白溶于10mmnaac(ph5.5)固定在cm5芯片(gehealthcare)上,然后将浓度梯度为1.9nm-250nm浓度梯度的抗体2a10g6mfab和h2a10g6fab分别流过芯片,分析在恒温25℃进行,使用的缓冲液是pbst缓冲液(含0.005%tween-20),流速为30μl/min。芯片表面的再生使用的是10mmnaoh。结合动力学曲线见图3、图4、图5。通过图3-5可以计算结合动力学常数,结合动力学常数的计算是利用biaevaluationsoftwareversion4.0(biacore,inc.)软件进行的,计算结果见表1。由表1可知,抗体人源化后,抗体对e蛋白的结合能力有所下降。但它与黄热、登革热、西尼罗热病毒的结合能力仍然维持在40-70nm之间,表明其结合能力依然很高。表1.2a10g6和h2a10g6与黄病毒蛋白亲和力的检测实施例5:中和试验将实施例2纯化的抗体5倍倍比稀释,与200pfu黄病毒(yfv、dv2、wnv)混合,37℃共孵育60分钟。然后将混合液加入到已铺满bhk21细胞的6孔板中。37℃孵育1小时后,弃掉病毒液,pbs洗涤细胞2次,换为含3ml1%(w/v)低熔点琼脂糖(promega)、4%胎牛血清的dmem培养基。感染后4至7天,6孔板用含有1%(w/v)结晶紫、4%(v/v)福尔马林试剂来观察孔板,通过空斑减少量来计算中和活力,公式如下:空斑减少比例=100-[(孵育抗体的空斑数/不孵育抗体的空斑数)*100]nt80表示空斑数下降80%的时候的抗体浓度。计算结果如表2,可知人源化抗体h2a10g6有着很强的活病毒中和活性。表2.2a10g6和h2a10g6对黄病毒的中和作用ic50(μg/ml)鼠单克隆抗体2a10g6人源化单克隆抗体h2a10g6yfv1.42.6dv24.0120wnv4698综上,由实施例2-5可知,本发明通过杆状病毒表达h2a10g6,纯化后可获得每升20mg以上的纯化抗体。抗体人源化后,它与黄热、登革热、西尼罗热病毒的结合能力仍然维持在40-70nm之间。通过与活病毒进行中和试验显示,人源化抗体h2a10g6有着很强的活病毒中和活性(表2)。这提示,人源化抗体h2a10g6是成功改造的黄病毒广谱中和抗体,有着临床治疗和预防黄热、登革热、西尼罗热以及其他黄病毒的应用价值。虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1