多功能等位基因的制作方法
本申请为申请号为201080056458.9的中国申请的分案申请。
发明领域
本发明涉及用于修饰基因组的核酸构建体,包括敲除构建体和用于将COIN置于基因组中的构建体。包括经遗传修饰的非人动物,例如具有以所选重组酶识别位点排列的基因或核酸元件的经遗传修饰的小鼠,所述重组酶识别位点允许基因或核酸元件的缺失或倒转,以在非人动物例如小鼠和大鼠中形成无效(null)等位基因、可选择的等位基因、报道等位基因和/或条件等位基因。
背景
一般地,通过用选择的另一种序列通常为报道分子和选择盒同源替换靶基因进行敲除,其中所述另一种序列优选邻侧为位点特异性重组酶位点,以准许经由同源位点特异性重组酶的作用去除选择盒。随后可以通过用相应同源重组酶处理细胞或通过使小鼠后代与“缺失者(deletor)”品系繁殖去除选择盒。例如,在floxed等位基因的情况下(其中目的序列邻侧为loxP位点),同源重组酶是Cre,并且在基因组中剩余的是单个loxP位点和报道分子。
相关策略已照常规用于生成条件-无效等位基因。这涉及以这样的方式使目的基因与位点特异性重组酶识别位点(例如用于Cre的lox,和用于Flp的FRT)部分侧接,从而使得在同源重组酶作用后,邻侧为位点特异性重组酶识别位点的区域被缺失,并且所得等位基因是无效等位基因。
尽管已做出将无效和条件功能性两者都并入一种靶向载体中且在单个靶向步骤中实现构建相应经修饰的等位基因的尝试,但由此类尝试产生的方法具有几个缺点且不是完全成功的。这些缺点包括例如真正功能性的缺乏(即无效形式不是真实无效的,条件等位基因不是真正条件的,缺乏报道分子功能等)或不能实现具有所需特征的实际工作等位基因。
因此,本领域需要当经改造的基因座是多功能基因座时,经由靶向生成经遗传修饰的生物体,例如真正的KO-first等位基因和随后条件-无效或其他条件-突变体等位基因。
附图简述
图1举例说明重组酶识别位点的使用,以同时缺失一个元件(U)并倒转另一个(D)。
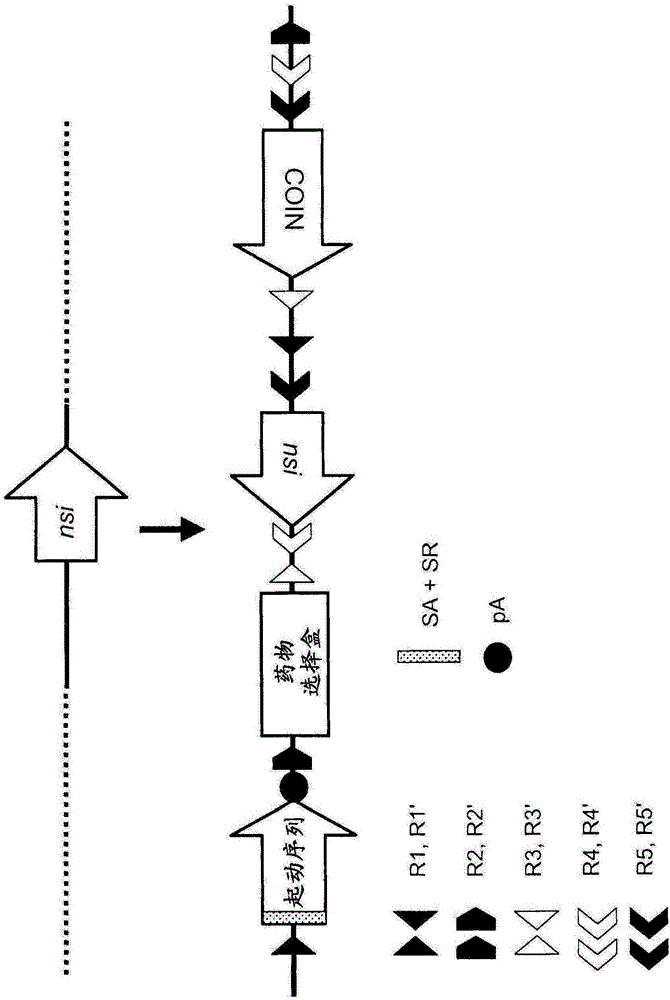
图2举例说明用于目的核苷酸序列(NSI)显示的,多功能等位基因(MFA)等位基因的一个实施方案,采用具有剪接区的剪接受体和起动序列随后为多聚腺苷酸化(pA)信号、药物选择盒(DSC;以选择的合适方向)、COIN和5对重组酶识别位点。R1/R1’、R2/R2’、R3/R3’、R4/R4’和R5/R5’代表重组酶识别位点的同源对。
图3举例说明实施MFA等位基因的一个实施方案的可重组单位(由R1/R1’、R2/R2’、R3/R3’、R4/R4’和R5/R5’定义)的概念图,所述MFA等位基因采用起动序列(为简单起见,未显示在序列前的剪接受体和剪接区、以及在序列后的polyA信号)、DSC(以选择的合适方向)、NSI和COIN。
图4举例说明具有特异性重组酶识别位点的MFA的特定实施方案用于举例说明目的(上),且生成包括包含LacZ序列的起动序列的“净化”无效等位基因,且使用单个重组酶步骤去除来自最初MFA实施方案的DSC以及NSI和COIN元件。为简单起见,未显示在LacZ序列前的剪接受体和剪接区以及在LacZ序列后的多聚腺苷酸信号。
图5举例说明生成条件等位基因的MFA的特定实施方案,所述条件等位基因使用单一重组酶(此处,Flp重组酶首先作用于等位基因实施方案的FRT3位点)含有/并入来自MFA的COIN。为简单起见,未显示在LacZ序列前的剪接受体和剪接区以及在LacZ序列后的多聚腺苷酸信号。
图6举例说明生成条件无效等位基因的MFA的实施方案,所述条件等位基因使用单一重组酶(此处,Flp重组酶首先作用于等位基因实施方案的FRT位点)含有/并入来自MFA的COIN。为简单起见,未显示在LacZ序列前的剪接受体和剪接区以及在LacZ序列后的多聚腺苷酸信号。
图7举例说明这样的实施方案,其中在重组酶处理后(如图5或图6中所示的Flp暴露),使等位基因暴露于第二种重组酶(Cre),导致NSI的缺失和COIN以用于转录的有义方向的放置。
图8举例说明这样的实施方案,其中在重组酶处理后(Flp暴露),使等位基因暴露于第二种重组酶,导致COIN和NSI的倒转,由于在NSI的5'位置关于第二种重组酶的重组酶识别位点的放置(备选)。
图9举例说明由Ensembl小鼠基因组服务器(上图–www.ensembl.org)修改的,在外显子2-4的区域中小鼠Hprt1基因的外显子-内含子结构,并且来自外显子2-外显子4的区域在ECR浏览器(http://ecrbrowser.dcode.org)中扩展,以突出显示保守区域。外显子3通过虚线椭圆形突出显示。黑色垂直箭头指出起动序列和DSC的插入点,而灰色箭头指出COIN元件的插入点,所有都用于改造Hprt1MFA等位基因。应当指出侧接外显子3的进化保守的内含子序列无一在所得等位基因中被破坏。虚线平行四边形指示在Hprt1MFA等位基因中将变成NSI的区域。
图10举例说明MFA特别是关于Hprt1基因的MFA的例子。Hprt1的外显子3加上侧接外显子3的进化保守的内含子序列(如图9中举例说明的)变成NSI。在靶向后,就Hprt1基因的转录方向而言将NSI置于反义链内。将起动序列-SA-lacZ-polyA-和DSC置于NSI的上游。就Hprt1基因的转录方向而言以有义方向放置起动序列,有效充当基因捕获元件,且取消起动序列下游的转录。就Hprt1基因的转录方向而言以反义方向将COIN元件置于NSI下游。COIN元件和NSI都不并入生产性Hprt1mRNA内,并且因此所得等位基因Hprt1MFA是具有报道分子(LacZ)的无效等位基因。包含Hprt1MFA等位基因的元件邻侧为如下排列的位点特异性重组酶识别位点:FRT-起动序列-Rox-DSC-FRT3-(LoxP)-(NSI)-(Lox2372)-(FRT)-(FRT3)-(COIN)-(Lox2372)-(LoxP)-Rox,其中括号指示放置是就Hprt1基因的转录方向而言以反义方向,或在位点特异性重组酶位点的情况下,就互相识别的对而言的相反方向。
图11举例说明MFA的实施方案,显示通过第一种重组酶(B)或第二种(C)和第三种(D)重组酶的作用生成的特定重叠的可重组单位(A)和所得等位基因。
图12举例说明MFA的实施方案,显示通过第一种重组酶(B)或第二种(C)和第三种(D)重组酶的作用生成的特定重叠的可重组单位(A)和所得等位基因。
图13举例说明MFA(A)的实施方案,显示起因于第一种重组酶和第二种重组酶的作用的所得等位基因,所述第一种重组酶以有义方向放置NSI(B),所述第二种重组酶以反义方向放置NSI同时以有义方向放置COIN(C)。
图14举例说明MFA(A)的实施方案,显示起因于第一种重组酶和第二种重组酶的作用的所得等位基因,所述第一种重组酶以有义方向放置NSI(B),所述第二种重组酶以反义方向放置NSI同时以有义方向放置COIN(C)。
图15举例说明MFA(A)的另一个实施方案,显示起因于第一种重组酶和第二种重组酶的作用的所得等位基因,所述第一种重组酶以有义方向放置NSI(B),所述第二种重组酶以反义方向放置NSI同时以有义方向放置COIN(C)。
图16举例说明MFA实施方案的例子,其中报道分子是SA(adml)-gtx-LacZ-pA,DSC是Neo,NSI是关键外显子(ec),并且COIN是Gtx-SA-HA-myc3-TM-T2A-GFP-pA(A),通过重组酶的作用以有义方向放置NSI,同时维持以反义方向的COIN(B),和NSI的进一步切除与COIN以有义方向的伴随放置(C);箭头指出用于证实在MFA中(A)、和在重组酶处理后(B和C)的重组酶位点的鉴定和方向的引物。
图17显示分别地关于Hprt1+/Y、Hprt1MFA/Y、Hprt1COIN/Y和Hprt1COIN-INV/YES细胞的细胞生存力和增殖测定的结果,所有细胞在不含6-TG(无6-TG;上图)或补充有10μM6-TG(6-TG;下图)的标准ES细胞培养基中培养。
图18显示衍生自Hprt1+/Y细胞(WT)、Hprt1MFA/Y(MFA)-即由图16A的MFA靶向的细胞、Hprt1COIN/Y细胞(MFA+FLPo)-即由图16A的MFA靶向且随后用FLPo处理的细胞、和Hprt1COIN-INV/Y细胞(MFA+FLPo+Cre)-即用Cre处理的Hprt1COIN/Y细胞的总蛋白质制备物的Western印迹;上图显示Hprt1蛋白质的检测,中图显示LacZ(报道分子)蛋白质的检测,并且下图显示作为上样对照的GAPDH蛋白质的检测。
概述
提供了用于制备无效等位基因和条件等位基因、和组合无效和COIN特征的等位基因的方法和组合物。在多种实施方案中,提供了用于在单个靶向步骤中将多功能等位基因改造到基因组内的方法和组合物。提供了用于在经遗传修饰的非人动物中的敲除补充分析的方法和组合物,包括包含单个靶向步骤的方法。
在一个方面,提供了经修饰的等位基因,其包含3'剪接区和剪接受体,和就剪接受体而言3'的起动序列,和就起动序列而言3'的目的核苷酸序列(NSI),其中NSI以就靶基因(或待修饰的基因座,或就起动序列而言)而言的反义方向。
在一个实施方案中,起动序列选自微小RNA、转录停止信号(例如多聚腺苷酸化区)、编码cDNA的核苷酸序列或其任何组合,并且可以包括调节元件例如操纵子、增强子和隔离子。在特定实施方案中,cDNA编码报道分子(例如编码LacZ)。在一个实施方案中,起动序列包含外显子。在特定实施方案中,外显子是基因座的最5'外显子。
在一个实施方案中,NSI包含外显子。在一个实施方案中,NSI包含外显子和邻近内含子序列。在特定实施方案中,侧接外显子与内含子序列5'和3'侧接。在一个实施方案中,核苷酸序列包含2个或更多个外显子,并且在特定实施方案中,包含一种或多种内含子序列。在另一个实施方案中,NSI缺乏外显子,或缺乏外显子的片段。
在一个实施方案中,经修饰的等位基因包含COIN。在一个实施方案中,COIN是就NSI而言3'的;在另一个实施方案中,COIN是就NSI而言5'的。
在一个实施方案中,COIN选自报道分子、基因捕获样元件(GT样元件)和基因捕获样报道分子(GT样报道分子)。在特定实施方案中,GT样元件选自SA-药物抗性cDNA-polyA。在特定实施方案中,GT样元件选自SA-报道分子-polyA。
在一个实施方案中,COIN包含3'剪接区。在特定实施方案中,3'剪接区随后为选自cDNA、外显子-内含子序列、微小RNA、微小RNA簇、小RNA、密码子跳过元件、IRES、多聚腺苷酸化序列或其任何组合的序列。在特定实施方案中,小RNA是mirtron。在特定实施方案中,密码子跳过元件是T2A、E2A或F2A。
在一个实施方案中,经修饰的等位基因包含药物选择盒(DSC)。
在一个实施方案中,经修饰的等位基因在靶向构建体上,所述靶向构建体包含上游和下游同源臂。在一个实施方案中,至少一个同源臂是小鼠同源臂。在特定实施方案中,2个同源臂都是小鼠同源臂。
在一个实施方案中,经修饰的等位基因从5'到3'包含剪接受体、起动序列、DSC、NSI和COIN,其中目的核苷酸序列和COIN都处于就起动序列而言的反义方向,和5对位点特异性重组酶识别位点。在一个实施方案中,在暴露于独立地识别且倒转在第一对位点特异性重组酶识别位点之间的序列,且缺失在第二对位点特异性重组酶识别位点之间的序列的第一种位点特异性重组酶后,经修饰的等位基因产生如下等位基因,其包含就用于转录的有义方向的NSI,缺乏DSC,并且包含以反义方向的COIN。在一个实施方案中,经修饰的等位基因包含这样排列的第三个和第四个位点特异性重组酶识别位点,从而使得等位基因对于第二种重组酶的进一步暴露导致缺失NSI且将COIN以用于转录的有义方向放置,所述第二种重组酶独立地识别第三个和第四个位点特异性重组酶识别位点。
在一个方面,提供了核酸构建体,其包含(a)以有义方向的报道分子和以选择的合适方向的DSC,以及以反义方向的NSI和COIN;(b)5对位点特异性重组酶识别位点,其中5对重组酶识别位点由不超过3种重组酶识别;其中在用第一种重组酶处理核酸构建体后,形成经修饰的等位基因,其中(i)NSI以有义方向放置,(ii)COIN保持反义方向,(iii)报道分子和DSC缺失,和(iv)在用第二种重组酶处理后,经修饰的等位基因倒转和/或缺失NSI,且将COIN以有义方向放置。
在一个实施方案中,5对位点特异性重组酶识别位点是FRT3、Rox、FRT、loxP和lox2372对。
在一个实施方案中,第一种重组酶是Flp重组酶,并且第二种重组酶是Cre重组酶。
在一个实施方案中,在用第二种重组酶处理后,经修饰的等位基因产生不能通过第一种或第二种重组酶缺失或倒转的等位基因。
在一个实施方案中,目的核苷酸序列是基因的野生型外显子。在另一个实施方案中,NSI是具有一个或多个核酸置换、缺失或添加的基因的外显子。
在一个实施方案中,NSI是基因的野生型外显子加上邻侧的内含子。在另一个实施方案中,NSI是具有一个或多个核酸置换、缺失或添加的基因的外显子加上邻近内含子序列。
在一个实施方案中,NSI是基因的野生型内含子。在另一个实施方案中,NSI是具有一个或多个核酸置换、缺失或添加的基因的内含子。
在一个实施方案中,COIN包含基因的一个或多个外显子,其包含一个或多个核酸置换、缺失或添加。在特定实施方案中,COIN包含哺乳动物的外显子。在特定实施方案中,哺乳动物是人、小鼠、猴或大鼠。
在一个实施方案中,COIN包含3'剪接区。在特定实施方案中,3'剪接区随后为选自cDNA、外显子-内含子序列、微小RNA、微小RNA簇、小RNA、密码子跳过元件、IRES、多聚腺苷酸化序列及其组合的序列。在特定实施方案中,小RNA是mirtron。在特定实施方案中,密码子跳过元件是T2A。
在一个实施方案中,COIN选自报道分子、基因捕获样元件(GT样元件)和基因捕获样报道分子(GT样报道分子)。在特定实施方案中,GT样元件选自SA-药物抗性cDNA-polyA。在特定实施方案中,GT样元件选自SA-报道分子-polyA。
在一个实施方案中,构建体进一步包含上游和下游同源臂。在一个实施方案中,上游和下游同源臂是小鼠或大鼠同源臂。在特定实施方案中,同源臂是小鼠同源臂,并且NSI包含人序列。在特定实施方案中,人序列包含其为小鼠外显子的人同系物的人外显子。
在一个实施方案中,报道分子选自:荧光蛋白质、发光蛋白质或酶。在特定实施方案中,报道分子选自GFP、eGFP、CFP、YFP、eYFP、BFP、eBFP、DsRed、MmGFP、萤光素酶、LacZ和碱性磷酸酶。
在一个实施方案中,DSC包含编码选自下述活性的序列:新霉素磷酸转移酶(neor)、潮霉素B磷酸转移酶(hygr)、嘌呤霉素-N-乙酰基转移酶(puror)、杀稻瘟菌素S脱氨酶(bsrr)、黄嘌呤/鸟嘌呤磷酸核糖基转移酶(gpt)、诺尔丝菌素乙酰基转移酶(nat1)、和单纯疱疹病毒胸苷激酶(HSV-tk)。
在一个方面,提供了核酸构建体,其包括包含3'剪接受体随后为以有义方向的报道分子的起动序列,以选择的合适方向的DSC,以反义方向的NSI和以反义方向的COIN,其中起动序列和报道分子上游邻侧为重组酶识别位点R1,重组酶识别位点R2置于报道分子和DSC之间,重组酶识别位点R3置于DSC和目的核苷酸序列之间,重组酶识别位点R4置于位点R3和NSI之间,重组酶识别位点R5置于NSI和COIN之间,重组酶识别位点R1’置于位点R5和COIN之间,重组酶识别位点R3’置于R1’和COIN之间,重组酶识别位点R5’置于COIN的下游,重组酶识别位点R4’置于位点R5’的下游,并且重组酶识别位点R2’置于位点R4’的下游;其中R1和R1’处于相反方向,R2和R2’处于相同方向,R3和R3’处于相反方向,R4和R4’处于相同方向,并且R5和R5’处于相同方向。
在一个实施方案中,报道分子随后为多聚腺苷酸化区域。
在一个实施方案中,R1和R1’由识别R3和R3’的重组酶识别。在一个实施方案中,R4和R4’由识别R5和R5’的重组酶识别。在一个实施方案中,R2和R2’不由识别R1/R1’、R3/R3’、R4/R4’或R5/R5’的任何重组酶识别。在一个实施方案中,R1和R1’、R3和R3’以及R2和R2’不由识别R4和R4’以及R5和R5’的任何重组酶识别。在一个实施方案中,R4和R4’、R5和R5’以及R2和R2’不由识别R1和R1’以及R3和R3’的任何重组酶识别。
在一个实施方案中,用单一重组酶的处理产生缺乏DSC、NSI和COIN的核酸构建体。在特定实施方案中,所得到的核酸构建体基本上由起动序列、R1和R2或R2’组成。在特定实施方案中,R1是FRT3位点,并且R2(或R2’)是Rox位点。
在一个实施方案中,用单一重组酶的处理产生这样的核酸构建体,其包含以有义方向的起动序列,但缺乏DSC,缺乏NSI且缺乏COIN。在特定实施方案中,R2和R2’是Rox位点,并且单一重组酶是Dre重组酶。
在一个实施方案中,用单一重组酶的处理产生这样的核酸构建体,其包含以反义方向的NSI和以反义方向的COIN。在一个实施方案中,单一重组酶是Flp重组酶,R1和R1’是不与R3和R3’(其也是FRT或FRT变体)交叉反应的FRT变体序列,R2和R2’是Rox序列,并且R4和R4’是不与R5和R5’交叉反应的loxP或lox变体序列,其中R5和R5’是lox变体序列。
在一个实施方案中,用单一重组酶的处理产生这样的核酸构建体,其包含以有义方向的NSI和以反义方向的COIN。在一个实施方案中,单一重组酶是Flp重组酶,R1和R1’是FRT3序列,R2和R2’是Rox序列,R3和R3’是FRT序列,R4和R4’是loxP序列,R5和R5’是lox2372序列。
在一个实施方案中,NSI是基因的野生型外显子。在另一个实施方案中,NSI是具有一个或多个核酸置换、缺失或添加的基因的外显子。
在一个实施方案中,COIN包含基因的一个或多个外显子,其包含一个或多个核酸置换、缺失或添加。在特定实施方案中,COIN包含哺乳动物的外显子。在一个实施方案中,哺乳动物是人、小鼠、猴或大鼠。
在一个实施方案中,构建体进一步包含构建体上游的同源臂(上游同源臂)和构建体下游的同源臂(下游同源臂)。在一个实施方案中,上游和下游同源臂是小鼠或大鼠同源臂。在特定实施方案中,同源臂是小鼠同源臂,并且NSI包含人序列。在特定实施方案中,人序列包含与小鼠外显子同源的人外显子。
在一个实施方案中,报道分子选自:荧光蛋白质、发光蛋白质或酶。在特定实施方案中,报道分子选自GFP、eGFP、CFP、YFP、eYFP、BFP、eBFP、DsRed、MmGFP、萤光素酶、LacZ和碱性磷酸酶。在一个实施方案中,DSC包含编码选自下述活性的序列:新霉素磷酸转移酶(neor)、潮霉素B磷酸转移酶(hygr)、嘌呤霉素-N-乙酰基转移酶(puror)、杀稻瘟菌素S脱氨酶(bsrr)、黄嘌呤/鸟嘌呤磷酸核糖基转移酶(gpt)、诺尔丝菌素乙酰基转移酶(nat1)、和单纯疱疹病毒胸苷激酶(HSV-tk)。
在一个方面,提供了用于修饰基因座的核酸构建体,其包含第一个、第二个、第三个、第四个和第五个重叠的可重组单位,其中可重组单位包括一对同源位点特异性重组酶识别位点,并且其中(a)外框为以相反方向的重组酶位点R1和R1’(允许经由R1/R1’的倒转)的第一个可重组单位,其中在R1和R1’之间放置就靶基因的转录方向而言以有义方向的起动序列,随后为重组酶位点R2,随后为以选择的合适方向的DSC,随后为重组酶位点R3,随后为重组酶位点R4,随后为以反义方向的NSI,随后为重组酶位点R5;(b)外框为以相同方向的重组酶位点R2和R2’(允许经由R2/R2’的缺失)的第二个可重组单位,其中在R2和R2’之间放置以选择的合适方向的DSC,随后为R3,随后为R4,随后为以反义方向的NSI,随后为R5,随后为R1’,随后为重组酶位点R3’,其中R3’处于就R3而言的相反方向(使得能够经由R3/R3’倒转),随后为以反义方向的COIN,随后为R5’,其中R5’处于就R5而言的相同方向,随后为R4’,其中R4’处于就R4而言的相同方向,随后为R2’,其中R2’处于就R2而言的相同方向(允许经由R2/R2’的缺失);(c)外框为以相反方向的重组酶位点R3和R3’(允许经由R3/R3’的倒转)的第三个可重组单位,其中在R3和R3’之间放置R4,以反义方向的NSI,随后为R5,随后为R1’;(d)外框为以相同方向的重组酶位点R4和R4’的第四个可重组单位,其中在R4和R4’之间放置以反义方向的NSI,随后为R5,随后为R1’,随后为R3’,随后为以反义方向的COIN,随后为R5’,随后为R4’;和(e)外框为相同方向的重组酶位点R5和R5’的第五个可重组单位,其中在R5和R5’之间放置R1’,随后为R3’,随后为以反义方向的COIN。
在一个实施方案中,R1/R1’和R3/R3’就相同位点特异性重组酶而言是功能性的,并且所述相同的位点特异性重组酶就R4/R4’和R5/R5’、和R2/R2’而言不是功能性的。
在一个实施方案中,R4/R4’和R5/R5’就相同位点特异性重组酶而言是功能性的,并且所述相同的位点特异性重组酶就R1/R1’和R3/R3’、和R2/R2’而言不是功能性的。
在一个实施方案中,R2/R2’对于重组酶是功能性的,其中所述重组酶就R1/R1’、R3/R3’、R4/R4’和R5/R5’中的任何而言不是功能性的。
在一个实施方案中,R1/R1’是FRT、FRT3、loxP或lox2372位点。在一个实施方案中,R3/R3’是FRT、FRT3、loxP或lox2372位点。在一个实施方案中,R4/R4’是FRT、FRT3、loxP或lox2372位点。在一个实施方案中,R5/R5’是FRT、FRT3、loxP或lox2372位点。在一个实施方案中,R2/R2’是Rox位点。在一个实施方案中,R2/R2’是attP/attB位点。
在特定实施方案中,R1/R1’和R3/R3’对于Flp重组酶是功能性的。在另一个特定实施方案中,R1/R1’和R3/R3’对于Cre重组酶是功能性的。
在特定实施方案中,R4/R4’和R5/R5’对于Cre重组酶是功能性的。在另一个特定实施方案中,R4/R4’和R5/R5’对于Flp重组酶是功能性的。
在一个实施方案中,R2/R2’是对于Dre重组酶是功能性的Rox位点。在另一个实施方案中,R2/R2’是对于PhiC31整合酶(phiC31\int)是功能性的attP/attB位点。
在一个实施方案中,报道分子选自:荧光蛋白质、发光蛋白质或酶。在特定实施方案中,报道分子选自GFP、eGFP、CFP、YFP、eYFP、BFP、eBFP、DsRed、MmGFP、萤光素酶、LacZ和碱性磷酸酶。
在一个实施方案中,DSC包含编码选自下述活性的序列:新霉素磷酸转移酶(neor)、潮霉素B磷酸转移酶(hygr)、嘌呤霉素-N-乙酰基转移酶(puror)、杀稻瘟菌素S脱氨酶(bsrr)、黄嘌呤/鸟嘌呤磷酸核糖基转移酶(gpt)、诺尔丝菌素乙酰基转移酶(nat1)、和单纯疱疹病毒胸苷激酶(HSV-tk)。
在一个实施方案中,NSI是基因的野生型外显子。在另一个实施方案中,NSI是具有一个或多个核酸置换、缺失或添加的基因的外显子。
在一个实施方案中,COIN包含基因的外显子,其包含一个或多个核酸置换、缺失或添加。在特定实施方案中,COIN包含人、小鼠、猴或大鼠的外显子。
在一个实施方案中,COIN包含3'剪接区。在特定实施方案中,3'剪接区随后为选自cDNA、外显子-内含子序列、微小RNA、微小RNA簇、小RNA、密码子跳过元件、IRES、多聚腺苷酸化序列及其组合的序列。在特定实施方案中,小RNA是mirtron。在特定实施方案中,密码子跳过元件是T2A、E2A或F2A。
在一个实施方案中,COIN选自报道分子、基因捕获样元件(GT样元件)和基因捕获样报道分子(GT样报道分子)。在特定实施方案中,GT样元件选自SA-药物抗性cDNA-polyA。在特定实施方案中,GT样元件选自SA-报道分子-polyA。
在一个实施方案中,构建体进一步包含构建体上游的同源臂(上游同源臂)和构建体下游的同源臂(下游同源臂)。在一个实施方案中,上游和下游同源臂是小鼠或大鼠同源臂。在特定实施方案中,同源臂是小鼠同源臂,并且NSI包含人序列。在特定实施方案中,人序列包含与小鼠外显子同源的人外显子。
在一个方面,提供了多功能等位基因,其包含由2种或更多种不同重组酶识别的2个或更多个可重组单位,每个可重组单位由限定可重组单位边界的一对相容的重组酶识别位点限定。每个可重组单位包含一个或多个内部重组酶识别位点。一个或多个内部重组酶识别位点这样进行选择,使得在通过多功能等位基因的可重组单位的第一种重组酶重组后,在一个可重组单位内的一个或多个内部重组酶识别位点随后与在另一个可重组单位内的一个或多个内部重组酶单位配对,以允许通过序列的第一种重组酶的倒转和/或缺失,所述序列跨越多功能等位基因的2个或更多个可重组单位,其中倒转和/或缺失仅在一个或多个内部重组酶识别位点倒转后才是可能的。
在一个实施方案中,倒转和/或缺失伴随多功能等位基因的其他重组酶识别位点的倒转,其中其他重组酶识别位点的倒转允许通过第二种重组酶倒转或缺失多功能等位基因的元件。
在一个方面,提供了多功能等位基因,其包含:(a)第一个、第二个、第三个、第四个和第五个可重组单位,其中每个可重组单位被相容的重组酶识别位点限制,并且其中第一个可重组单位重叠第二个可重组单位,并且其中第三个、第四个和第五个可重组单位包含在第二个可重组单位内;(b)第一个可重组单位包含与起动序列可操作地连接的3'剪接受体和剪接区、DSC和NSI;(c)第二个可重组单位包含DSC、NSI和COIN;(d)第三个可重组单位包含NSI;(e)第四个可重组单位包含NSI和COIN;(f)第五个可重组单位包含COIN;其中多功能等位基因包含侧接第一个可重组单位上游和下游的第一对重组酶识别位点,其允许第一个可重组单位的第一次倒转,其中第一次倒转导致在第二个可重组单位内重组酶位点的第二次倒转,其中第二次倒转定向在第二个可重组单位内的重组酶位点,以便缺失起动序列且缺失DSC。
在一个实施方案中,单一重组酶识别第一对重组酶识别位点并且还缺失起动序列和药物选择盒。
在一个实施方案中,第二次倒转定向重组位点,从而使得在倒转后,形成允许NSI的缺失和/或COIN的倒转的第二组重组酶识别位点。
在一个方面,提供了包含MFA的核酸构建体,所述MFA就转录方向而言从5'到3'包含以反义方向的COIN、以反义方向的NSI、DSC和以有义方向的报道分子,其中在用所选重组酶处理MFA后,COIN、NSI和DSC被切除,并且报道分子保持有义方向;并且其中在用不同的所选重组酶备选处理后,报道分子和DSC被切除,并且COIN保持反义方向,并且NSI以有义方向放置,从而使得在用另外一种不同的所选重组酶进一步处理后,NSI被切除,并且COIN以有义方向放置。
在一个实施方案中,MFA包含第一个可重组单位、第二个可重组单位和第三个可重组单位,其中第一个可重组单位重叠第二个和第三个可重组单位,并且其中第二个可重组单位重叠第一个和第三个可重组单位。
在一个实施方案中,第一个可重组单位包含以倒转(反义)方向的COIN和以倒转方向的NSI,其中可重组单位在COIN的上游和NSI的下游邻侧为定向以指导缺失的相容的重组酶位点R2和R2’;第二个可重组单位重叠第一个可重组单位,并且第二个可重组单位是通过重组酶对DSC上游的重组位点和报道分子下游的重组位点的作用而可重组的,其中重组位点定向以指导倒转,并且其中DSC上游的重组位点随后为包含NSI的序列。在特定实施方案中,MFA就在有义链上的方向而言从5'到3'包含第一个重组酶位点R1、第二个重组酶位点R2、第三个重组酶位点R3、以反义方向的COIN、第四个重组酶位点R4、第五个重组酶位点R5、与R3相容且定向以指导在R3和R3’之间的序列缺失的第六个重组酶位点R3’、以反义方向的NSI、与R2相容且定向以指导在R2和R2’之间的序列缺失的第七个重组酶位点R2’、第八个重组酶位点R4’、DSC、与R1相容且定向以指导在R1和R1’之间的序列缺失的第九个重组酶位点R1’、以有义方向的报道分子、和与R5相容且定向以指导在R5和R5’之间的序列倒转的第十个重组酶位点R5’。
在特定实施方案中,R1/R1’是Rox位点,R2/R2’是loxP位点,R3/R3’是lox2372位点,R4/R4’是FRT位点,并且R5/R5’是FRT3位点。在特定实施方案中,MFA包含如图11,小图A中所示的重组酶位点以及COIN、NSI、DSC和报道分子的放置。在特定实施方案中,在暴露于识别R1/R1’的单一重组酶后,形成如图11,小图B中所示的等位基因。在特定实施方案中,在暴露于识别R4/R4’和R5/R5’的单一重组酶后,形成如图11,小图C中所示的等位基因。在特定实施方案中,在图11,小图C的等位基因暴露于识别R2/R2’和R3/R3’的其他重组酶后,形成如图11,小图D中所示的等位基因。
在一个方面,提供了包含MFA的核酸构建体,所述MFA就转录方向而言从5'到3'包含以反义方向的NSI、DSC、以有义方向的报道分子和以反义方向的COIN;其中在用所选重组酶处理MFA后,NSI和DSC被切除,报道分子保持有义方向,并且COIN保持反义方向;并且其中在用不同的所选重组酶备选处理后,DSC和报道分子被切除,并且NSI以有义方向放置,并且COIN处于反义方向,并且其中在用不同的所选重组酶备选处理后,用另外一种不同的所选重组酶处理等位基因,导致NSI切除和COIN以有义方向的放置。
在一个实施方案中,MFA包含第一个可重组单位、第二个可重组单位和第三个可重组单位,其中第一个可重组单位重叠第二个和第三个可重组单位,并且其中第二个可重组单位重叠第一个和第三个可重组单位。在一个实施方案中,第一个可重组单位包含DSC和以有义方向的报道分子,并且其中可重组单位在DSC的上游邻侧为重组酶位点R2,随后为R3,并且在报道分子的下游邻侧为重组酶位点R3’,其中R2/R3’定向以指导倒转,并且其中就R2而言定向以指导倒转的R2’在DSC前;第二个可重组单位在反义NSI的上游邻侧为R4,并且在反义COIN的下游邻侧为R4’,其中R4/R4’定向以指导切除,并且其中第二个可重组单位包括DSC和报道分子;并且第三个可重组单位上游邻侧为R1并且下游邻侧为R1’,其中R1/R1’定向以指导切除,并且其中在R1’的上游且与R1’邻近的是DSC,并且其中在R1的下游且与R1邻近的是R2。在特定实施方案中,MFA就转录方向而言从5'到3'包含R1、R2、R3、R4、以反义方向的NSI、R5、R2’(其中R2/R2’定向以指导倒转)、DSC、R1’(其中R1/R1’定向以指导倒转)、报道基因、R3’(其中R3/R3’定向以指导倒转)、以反义方向的COIN、R5’(其中R5/R5’定向以指导切除)、和R4’(其中R4/R4’定向以指导切除)。
在特定实施方案中,R1/R1’是Rox位点,R2/R2’是FRT或FRT3位点,R3/R3’是与R2/R2’不同的FRT或FRT3位点,R4/R4’是lox2372位点或loxP位点,并且R5/R5’是与R4/R4’不同的lox2372位点或loxP位点。
在特定实施方案中,MFA包含如图12,小图A中所示的重组酶位点以及COIN、NSI、DSC和报道分子的放置。用所选重组酶处理产生图12,小图B中所示的等位基因。用不同的所选重组酶备选处理产生图12,小图C中所示的等位基因。用另外一种不同的重组酶处理图12,小图C的等位基因产生图12,小图D中所示的等位基因。
在一个方面,提供了包含MFA的核酸构建体,所述MFA就转录方向而言从5'到3'包含以有义方向的报道分子、DSC、以反义方向的NSI和以反义方向的COIN;其中在用第一种所选重组酶处理MFA后,报道分子被切除,NSI置于有义方向,并且COIN保持反义方向,并且其中等位基因包含允许序列倒转的重组酶位点,在用第二种所选重组酶处理后将COIN置于有义方向,并且NSI置于反义方向。在一个实施方案中,在用第一种所选重组酶处理后,用第二种所选重组酶处理等位基因。在一个实施方案中,COIN发信号NSI在用第二种重组酶处理后已置于反义方向。
在一个实施方案中,MFA就转录方向而言从5'到3'包含重组酶位点R1、报道分子、第二个重组酶位点R2、DSC、第三个重组酶位点R3、以反义方向的NSI、第四个重组酶位点R4、第五个重组酶位点R5、与R1相容且就R1而言定向以指导倒转的第六个重组酶位点R1’、与R3相容且就R3而言定向以指导倒转的第七个重组酶位点R3’、以反义方向的COIN、与R5相容且就R5而言定向以指导切除的第八个重组酶位点R5’、与R4相容且就R4而言定向以指导切除的第九个重组酶位点R4’、和与R2相容且就R2而言定向以指导切除的第十个重组酶位点R2’。在特定实施方案中,R1/R1’是FRT3或FRT位点,R2/R2’是Rox位点,R3/R3’是与R1/R1’不同的FRT3或FRT位点,R4/R4’是loxP或lox2372位点,并且R5/R5’是与R4/R4’不同的loxP或lox2372位点。
在特定实施方案中,MFA包含如图13,小图A中所示的重组酶位点以及COIN、NSI、DSC和报道分子的放置。用所选重组酶处理产生图13,小图B中所示的等位基因。用不同的重组酶处理图13,小图B的等位基因产生图13,小图C中所示的等位基因。
在一个方面,提供了包含MFA的核酸构建体,所述MFA就转录方向而言从5'到3'包含以反义方向的COIN、以反义方向的NSI、DSC和以有义方向的报道分子;其中在用第一种所选重组酶处理MFA后,报道分子被切除,NSI置于有义方向,并且COIN保持反义方向,并且其中等位基因包含允许序列倒转的重组酶位点,在用第二种所选重组酶处理后将COIN置于有义方向,并且NSI置于反义方向。在一个实施方案中,在用第一种所选重组酶处理后,用第二种所选重组酶处理等位基因。在一个实施方案中,COIN发信号NSI在用第二种重组酶处理后已置于反义方向。
在一个实施方案中,MFA就转录方向而言从5'到3'包含重组酶位点R1、第二个重组酶位点R2、第三个重组酶位点R3、以反义方向的COIN、第四个重组酶位点R4、第五个重组酶位点R5、与R3相容且就R3而言定向以指导切除的第六个重组酶位点R3’、与R2相容且就R2而言定向以指导切除的第七个重组酶位点R2’、以反义方向的NSI、与R4相容且就R4而言定向以指导倒转的第八个重组酶位点R4’、DSC、与R1相容且就R1而言定向以指导切除的第九个重组酶位点R1’、以有义方向的报道分子、和与R5相容且就R5而言定向以指导倒转的第十个重组酶位点R5’。在特定实施方案中,R1/R1’是Rox位点,R2/R2’是loxP或lox2372位点,R3/R3’是与R2/R2’不同的loxP或lox2372位点,R4/R4’是FRT或FRT3位点,并且R5/R5’是与R4/R4’不同的FRT或FRT3位点。
在特定实施方案中,MFA包含如图14,小图A中所示的重组酶位点以及COIN、NSI、DSC和报道分子的放置。用所选重组酶处理产生图14,小图B中所示的等位基因。用不同的重组酶处理图14,小图B的等位基因产生图14,小图C中所示的等位基因。
在一个方面,提供了包含MFA的核酸构建体,所述MFA就转录方向而言从5'到3'包含以反义方向的NSI、DSC、以有义方向的报道分子和以反义方向的COIN;其中在用第一种所选重组酶处理MFA后,报道分子被切除,DSC被切除,NSI置于有义方向,并且COIN保持反义方向,并且其中在用第一种所选重组酶处理后,等位基因包含允许序列倒转的重组酶位点,在用第二种所选重组酶处理后将COIN置于有义方向,并且NSI置于反义方向。在一个实施方案中,在用第一种所选重组酶处理后,用第二种所选重组酶处理等位基因。在一个实施方案中,COIN发信号NSI在用第二种重组酶处理后已置于反义方向。
在一个实施方案中,MFA就转录方向而言从5'到3'包含重组酶位点R1、第二个重组酶位点R2、第三个重组酶位点R3、以反义方向的NSI、第四个重组酶位点R4、第五个重组酶位点R5、与R2相容且就R2而言定向以指导倒转的第六个重组酶位点R2’、DSC、与R1相容且就R1而言定向以指导切除的第七个重组酶位点R1’、以有义方向的报道分子、与R3相容且就R3而言定向以指导倒转的第八个重组酶位点R3’、以倒转方向的COIN、与R5相容且就R5而言定向以指导切除的第九个重组酶位点R5’、和与R4相容且就R4而言定向以指导切除的第十个重组酶位点R4’。在特定实施方案中,R1/R1’是Rox位点,R2/R2’是FRT或FRT3位点,R3/R3’是与R2/R2’不同的FRT或FRT3位点,R4/R4’是loxP或lox2372位点,并且R5/R5’是与R4/R4’不同的loxP或lox2372位点。
在特定实施方案中,MFA包含如图15,小图A中所示的重组酶位点以及COIN、NSI、DSC和报道分子的放置。用所选重组酶处理产生图15,小图B中所示的等位基因。用不同的重组酶处理图15,小图B的等位基因产生图15,小图C中所示的等位基因。
在一个方面,提供了多功能等位基因,其包含DSC、报道分子、COIN、NSI和沿着报道分子排列的5对重组酶位点、DSC、COIN和NSI,其中没有重组酶位点对与任何其他对相同,并且其中第一个2对重组酶位点由相同的第一种重组酶识别,第二个2对重组酶位点由相同的第二种重组酶识别,并且第五对重组酶位点由第三种重组酶识别,其中第一种、第二种和第三种重组酶不相同,并且其中就转录方向而言,MFA包含(从5'到3'):(a)以有义方向的起动序列(例如具有报道分子)、以有义或反义方向的DSC、以反义方向的NSI、以反义方向的COIN;(b)以反义方向的COIN、以反义方向的NSI、以有义或反义方向的DSC、以有义方向的报道分子;(c)以反义方向的NSI、以有义或反义方向的DSC、以有义方向的报道分子、以反义方向的COIN;(d)以有义方向的报道分子、以有义或反义方向的DSC、以反义方向的NSI、以反义方向的COIN;(e)以反义方向的COIN、以反义方向的NSI、以有义或反义方向的DSC、以有义方向的报道分子;或(f)以反义方向的NSI、以有义或反义方向的DSC、以有义方向的报道分子、以反义方向的COIN。
在一个实施方案中,排列如(a)中所示,并且重组酶位点对这样排列,从而使得在暴露于第三种重组酶后,第五对重组酶位点指导DSC、NSI和COIN的切除,其中报道分子维持有义方向。
在一个实施方案中,排列如(a)中所示,并且重组酶位点对这样排列,从而使得在暴露于第一种重组酶后,形成经修饰的MFA,其中第一个2对重组酶位点指导报道分子的切除和DSC的切除和NSI至有义方向的倒转,其中COIN维持反义方向。在进一步的实施方案中,经修饰的MFA包含第二个2对重组酶位点,在暴露于第二种重组酶后,产生其中NSI被切除并且COIN置于有义方向的等位基因。
在一个实施方案中,排列如(b)中所示,并且重组酶位点对这样排列,从而使得在暴露于第三种重组酶后,第五对重组酶位点指导COIN、NSI和DSC的切除,其中报道分子维持有义方向。
在一个实施方案中,排列如(b)中所示,并且重组酶位点对这样排列,从而使得在暴露于第一种重组酶后,形成经修饰的MFA,其中第一个2对重组酶位点指导DSC和报道分子的直接切除以及NSI至有义方向的直接倒转,其中COIN维持反义方向。在进一步的实施方案中,经修饰的MFA包含第二个2对重组酶位点,在暴露于第二种重组酶后,产生其中NSI被切除并且COIN置于有义方向的等位基因。
在一个实施方案中,排列如(c)中所示,并且重组酶位点对这样排列,从而使得在暴露于第五种重组酶后,NSI和DSC被切除,并且报道分子和COIN维持反义方向。
在一个实施方案中,排列如(c)中所示,并且重组酶位点对这样排列,从而使得在暴露于第一种重组酶后,形成经修饰的MFA,其中DSC和报道分子被切除,并且NSI置于有义方向,其中COIN维持反义方向。在进一步的实施方案中,经修饰的MFA包含第二个2对重组酶位点,在暴露于第二种重组酶后,产生其中NSI被切除并且COIN置于有义方向的等位基因。
在一个实施方案中,排列如(d)中所示,并且重组酶位点对这样排列,从而使得在暴露于第五种重组酶后,DSC、NSI和COIN被切除,并且报道分子维持有义方向。
在一个实施方案中,排列如(d)中所示,并且重组酶位点对这样排列,从而使得在暴露于第一种重组酶后,形成经修饰的MFA,其中报道分子和DSC被切除,并且NSI置于有义方向,其中COIN维持反义方向。在进一步的实施方案中,经修饰的MFA包含第二个2对重组酶位点,在暴露于第二种重组酶后,产生其中COIN置于有义方向并且NSI置于反义方向的等位基因。
在一个实施方案中,排列如(e)中所示,并且重组酶位点对这样排列,从而使得在暴露于第五种重组酶后,COIN、NSI和DSC被切除,并且报道分子维持有义方向。
在一个实施方案中,排列如(e)中所示,并且重组酶位点对这样排列,从而使得在暴露于第一种重组酶后,形成经修饰的MFA,其中DSC和报道分子被切除,NSI置于有义方向,并且COIN维持反义方向。在进一步的实施方案中,经修饰的MFA包含这样排列的第二个2对重组酶位点,从而使得在暴露于第二种重组酶后,NSI置于反义方向并且COIN置于有义方向。
在一个实施方案中,排列如(f)中所示,并且重组酶位点对这样排列,从而使得在暴露于第五种重组酶后,NSI和DSC被切除,报道分子维持有义方向,并且COIN维持反义方向。
在一个实施方案中,排列如(f)中所示,并且重组酶位点对这样排列,从而使得在暴露于第五种重组酶后,NSI和DSC被切除,并且报道分子维持有义方向,并且COIN维持反义方向。
在一个实施方案中,排列如(f)中所示,并且重组酶位点对这样排列,从而使得在暴露于第一种重组酶后,形成经修饰的MFA,其中DSC和报道分子被切除,并且NSI置于有义方向,并且COIN维持反义方向。在进一步的实施方案中,经修饰的MFA包含这样排列的第二个2对重组酶位点,从而使得在暴露于第二种重组酶后,NSI置于反义方向并且COIN置于有义方向。
在一个方面,提供了用于制备细胞的方法,所述细胞包含具有以反义方向的目的核苷酸序列和以反义方向的COIN的构建体,所述方法包括将如本文描述的MFA引入细胞的基因组内、鉴定包含MFA的细胞的步骤、随后为使基因组暴露于第一种重组酶的步骤,其中第一种重组酶对基因组中的构建体的作用导致目的核苷酸序列置于有义方向。
在一个实施方案中,细胞是多能细胞、经诱导的多能细胞、全能细胞或ES细胞。在特定实施方案中,ES细胞是小鼠或大鼠ES细胞。
在一个实施方案中,通过同源重组将构建体引入细胞内。在另一个实施方案中,构建体随机整合到细胞的核酸内。在一个实施方案中,细胞的核酸是细胞的基因组。
在一个实施方案中,NSI包含外显子。在一个实施方案中,NSI包含外显子和侧接内含子序列。在特定实施方案中,侧接外显子与内含子序列在5'和3'侧接。在一个实施方案中,核苷酸序列包含2个或更多个外显子,并且在特定实施方案中,包含一种或多种内含子序列。在另一个实施方案中,NSI缺乏外显子,或缺乏外显子的片段。
在一个实施方案中,NSI是基因的一个或多个野生型外显子。在另一个实施方案中,NSI是具有一个或多个核酸置换、缺失或添加的基因的一个或多个外显子。
在一个实施方案中,COIN包含基因的外显子,其包含一个或多个核酸置换、缺失或添加。在特定实施方案中,COIN包含人、小鼠、猴或大鼠基因的外显子。
在一个实施方案中,COIN包含3'剪接区。在特定实施方案中,3'剪接区随后为选自cDNA、外显子-内含子序列、微小RNA、微小RNA簇、小RNA、密码子跳过元件、IRES、多聚腺苷酸化序列及其组合的序列。在特定实施方案中,小RNA是mirtron。在特定实施方案中,密码子跳过元件是T2A、E2A或F2A。
在一个实施方案中,COIN选自报道分子、基因捕获样元件(GT样元件)和基因捕获样报道分子(GT样报道分子)。在特定实施方案中,GT样元件选自SA-药物抗性cDNA-polyA。在特定实施方案中,GT样元件选自SA-报道分子-polyA。
在一个方面,提供了用于将多功能等位基因置于小鼠细胞基因组内的方法,其包括将包含第一个可重组单位的靶向构建体引入小鼠细胞中的基因座内的步骤,所述第一个可重组单位包含(a)起动序列(例如核苷酸序列和/或报道分子);(b)DSC;(c)就基因座而言以反义方向的NSI;(d)就基因座而言以反义方向的COIN;和(e)在可重组单位中排列的位点特异性重组酶识别位点,用于缺失报道分子和DSC、用于将NSI倒转回有义方向、以及用于倒转COIN且缺失或再倒转NSI。
在一个实施方案中,位点特异性重组酶识别位点这样排列在可重组单位中,从而使得NSI将再倒转成反义链,并且COIN将倒转成有义链。在另一个实施方案中,位点特异性重组酶识别位点这样排列在可重组单位中,从而使得NSI将缺失,并且COIN将倒转成有义链。
在一个实施方案中,可重组单位这样排列,从而使得在MFA修饰的靶基因座暴露于第一种重组酶后,包含报道分子和DSC的第一个可重组单位被缺失,并且NSI置于就基因组而言的有义方向,并且COIN维持反义方向,形成第二个可重组单位。
在一个实施方案中,以反义方向的目的核苷酸序列是以反义方向的外显子,或邻侧为内含子序列的外显子,其中外显子和内含子序列各自以反义方向。在特定实施方案中,置于反义方向的外显子与被靶向构建体替换的外显子相同。在特定实施方案中,以反义方向的NSI是外显子和外显子周围的序列。在特定实施方案中,NSI是2个或更多个外显子。在特定实施方案中,NSI是非外显子序列。
在一个实施方案中,通过第一种重组酶的作用生成的第二个可重组单位暴露于第二种重组酶,其中第二种重组酶缺失NSI且将COIN置于有义方向。
在一个实施方案中,通过第一种重组酶的作用生成的第二个可重组单位暴露于第二种重组酶,其中第二种重组酶将NSI置于反义方向且将COIN置于有义方向。
在一个方面,提供了用于补充敲除的方法,其包括将如本文描述的MFA引入非人动物内,其中核酸构建体包含以反义方向的野生型核酸序列和以反义方向的COIN,其中在核酸构建体暴露于第一种重组酶后,野生型核酸序列倒转至有义方向且转录,但COIN保持反义方向;并且其中在暴露于第二种重组酶后,野生型核酸序列被切除或倒转回反义链,并且COIN倒转至有义方向。
在一个实施方案中,非人动物是小鼠。
在一个实施方案中,COIN是报道分子元件。在一个实施方案中,报道分子元件选自荧光蛋白质、发光蛋白质或酶。在特定实施方案中,报道分子选自GFP、eGFP、CFP、YFP、eYFP、BFP、eBFP、DsRed、MmGFP、萤光素酶、LacZ和碱性磷酸酶。
在一个方面,提供了包含依照本发明的多功能等位基因的哺乳动物细胞。
在一个实施方案中,哺乳动物细胞选自小鼠细胞和大鼠细胞。在一个实施方案中,细胞选自干细胞、胚胎干(ES)细胞、经诱导的多能细胞、多能细胞和全能细胞。
在一个方面,提供了包含依照本发明的多功能等位基因的非人胚胎或非人动物。
在一个实施方案中,非人胚胎或非人动物包含已暴露于一种或多种位点特异性重组酶的多功能等位基因。在特定实施方案中,由于育种步骤,多功能等位基因已暴露于一种或多种位点特异性重组酶,其中包含多功能等位基因的非人动物已与包含一种或多种位点特异性重组酶的非人动物交配,并且非人胚胎或非人动物是育种步骤的后代。
在一个方面,提供了包含如本文描述的MFA的细胞,其中细胞是哺乳动物细胞,例如ES细胞或多能或经诱导的多能细胞。在特定实施方案中,细胞是小鼠或大鼠细胞。
在一个方面,提供了包含如本文描述的MFA、或如本文描述的已暴露于一种或多种重组酶的MFA的非人动物。
在一个方面,提供了包含如本文描述的MFA、或如本文描述的已暴露于一种或多种重组酶的MFA的非人胚胎。
在一个方面,提供了使用如本文描述的MFA制备的细胞、非人胚胎或非人动物。
在一个方面,提供了使用如本文描述的MFA制备的细胞、非人胚胎或非人动物。
任何方面或实施方案可以与适当的任何其他方面或实施方案组合使用,例如与任何特定MFA实施方案结合叙述的任何报道分子或DSC可以与本文描述的任何MFA实施方案一起使用,并且与任何特定MFA实施方案结合提及的任何特定重组酶或重组酶位点可以与本文描述的任何MFA实施方案一起使用。
根据随后详述的综述,其他实施方案得到描述且对于本领域技术人员将是显而易见的。
详述
本发明并不限于所述的特定方法和实验条件,因为此类方法和条件可以改变。本文使用的术语仅用于描述特定实施方案的目的,并且不预期是限制性的,因为本发明的范围仅受权利要求限制。
除非另有说明,本文使用的所有技术和科学术语具有与本发明所属领域普通技术人员通常理解相同的含义。尽管目前描述了特定方法和材料,但类似于或等价于本文描述那些的任何方法和材料可以用于本发明的实践或测试中。本文提及的所有出版物通过引用整体合并入本文。
短语“有义方向”或“有义”指在基因组的局部背景中可转录核酸序列的编码方向或有义链,例如当序列以“有义方向”置于基因组中的可转录序列中或附近时,序列的方向与转录相容,并且对于蛋白质编码基因,还与序列置于其中的区域或基因座或基因中的序列翻译相容。短语“反义方向”或“反义”指序列在区域或基因座或基因中的放置,其中序列处于与转录相容那种相反(或反义)的链中。因此,在特定例子中,如果序列置于基因中的“有义”方向,那么它一般可以被转录。如果序列置于“反义”方向,那么它一般不被转录。对于其中在有义和反义链之间的转移可能导致来自任一链的转录的基因座,序列可以这样进行选择或改造,从而使得从有义到反义或从反义到有义的转移将导致来自一条链而不是任一的转录。
术语“COIN”包括提及条件元件。条件元件包含其表达(或未能表达)视独立事件的发生而定的核苷酸序列。例如,以有义方向的编码区将编码蛋白质或其片段,或非编码RNA(ncRNA)置于反义方向并且在两侧上邻侧为以相反方向的位点特异性重组位点。在不存在识别侧接位点的位点特异性重组酶的情况下,编码区不转录,因为它置于就靶基因而言的反义链中。在用同源位点特异性重组酶处理后,COIN序列倒转,并且因此,它变得并入转录的信使内,导致蛋白质或其片段或ncRNA的表达。
当用于描述2个或更多个重组酶识别位点时,术语“不相容的”指2个或更多个重组酶识别位点不能彼此重组(但2个或更多个重组酶识别位点可以与其他同源(例如等同)重组酶识别位点重组)的特性。
敲除和条件等位基因
通过遗传方法的基因功能研究已依赖于发现天然存在的变体或突变体等位基因,或此类变体或突变体等位基因的有意生成。后者已通过下述进行:随机诱变,随后为基于表型的筛选和气候原因突变的阐明—已称为“正向遗传学”的过程,或基因工程方法,由此在特异性“靶”基因或基因座中致使突变—已称为“反向遗传学”的方法。
在小鼠中—最广泛使用的哺乳动物模型生物体—经由基因靶向改造特异性、分子上非常充分限定的突变的能力已在反向遗传学领域的支配地位。然而,迄今为止制备的大多数变体是相对简单的无效等位基因,通常称为“敲除等位基因”或简单地“敲除体”,并且通常涵盖基因或其部分的外显子-内含子区的缺失,并且近年来伴随那个区域被报道分子cDNA例如LacZ的同时替换。衍生自噬菌体或酵母且修饰用于在哺乳动物细胞中使用的位点特异性重组酶及其同源识别位点(例如Cre/lox、Dre/Rox、PhiC31\int/attP-attB、Flp/FRT)的修改是更近期的开发,其不仅使DSC的靶向后切除变成可能(只要它邻侧为位点特异性重组酶识别位点),还已使得条件-无效等位基因的改造成为可能。已开发出条件-无效等位基因,其中靶基因的外显子-内含子区—或更经常地其部分—邻侧为重组酶识别位点,致使经修饰的等位基因顺应通过同源重组酶的作用而转换为无效状态。这种方法相比于常规敲除的优点是通过控制位置(器官、组织或细胞类型)、时间以及有时同源重组酶将有活性的持续时间,可以在空间时间上控制经修饰的基因至敲除的转换。
常规地,作为后续,条件-无效等位基因已改造为相应简单敲除等位基因,主要是在其中后者是胚胎致死和/或展示出多重表型的情况下,因此致使靶基因功能的研究在成年设置背景中是不可能的(在胚胎致死率的情况下),或在特异性细胞类型或生物学过程中难以解释(在其中基因展示出多重表型的情况下)。考虑到经由基因靶向生成经遗传修饰的小鼠花费的努力、时间和费用的量,首先生成敲除体,随后译码其表型,且随后改造条件-无效(conditional-null)的这个逐步方式已由越来越多的研究者视为繁重的。此外,对于小数目的基因,常规敲除等位基因无法经历种系,因为它们即使在杂合无效(heterozygousnull)状态下也导致胚胎致死率。因此,在单个基因靶向步骤中能够改造双重(无效和条件)或甚至多模式等位基因的需要已是本领域涉及那些以及最终用户的共同体的持续目标。2种方法已事实上设法解决这个需要:FlEx和首先敲除(KO-first)。
FlEx方法已用于靶向和作为基因捕获(GT),但基本设计原则是相同的,与最终应用无关。FlEx的基本设计显示于图1中,其中U代表例如DSC并且D代表报道分子。对FlEx构建体的重组酶作用结果是U元件(例如DSC)的永久缺失和D元件(例如报道分子)的倒转(和表达)。
在其最初实施方案和应用中,FlEx开发作为改造条件等位基因的方法。通过将FlEx盒插入这个基因内,FlEx首先用于生成用于Rarg的“条件-无效”等位基因,从而使得loxP/lox511偶联体插入Rarg的外显子8的上游,并且FlEx盒的其余部分—包括5'至3'地就Rarg而言以反义方向的SA-lacZ-SV40polyA(GT样元件),和随后就第一个loxP/lox511偶联体而言以反义方向的另一个loxP/lox511偶联体,并且含有以有义方向插入Rarg的内含子8内的新霉素磷酸转移酶小基因(neo)。这种设计准许Cre介导的GT样元件SA-lacZ-SV40polyA的倒转,从而使得它进入有义链,并且充当基因捕获;同时,Rarg的外显子8带进反义方向内(有效确保即使在其中转录不在GT样元件SA-lacZ-SV40polyA末端时终止的情况下,外显子8也不并入通读信息内),虽然neo被同时缺失,并且从而产生Rarg的无效等位基因,其中Rarg的表达被lacZ的表达替换。
然而,尽管这种方法在生成无效等位基因(通过暴露于Cre)中成功,但Rarg的未重排(前Cre)FlEx等位基因不是如最初设计的真实的条件-无效的,而是严重减效的等位基因,其中RargFLEx的表达与Rarg比较是显著减少的。因此,RargFLEx/FLEx小鼠展示出类似较不严重形式的Rarg敲除小鼠的表型,并且揭示FlEx的这个起始实施方案不能生成真实的条件-无效等位基因。
FlEx方法也已适合于在基因捕获中使用。在该方法的那种变化中,GT元件(SA-βgeo-polyA,其中βgeo是lacZ与neo开放读码框的框内融合物,由此组合经由LacZ报道和经由Neo选择的能力)邻侧为2个FlEx样阵列,由FRT/FRT3偶联体组成的外部阵列,和由loxP/lox511偶联体组成的内部阵列,两者在就彼此而言的镜像配置中。以这种方式,所得到的GT载体成功并入活性转录的基因内将导致βgeo的表达和由此允许通过就G418选择来选择这些事件,并且取决于GT元件的并入位点理论上还导致生成关于相应基因的功能无效等位基因。一旦基因已捕获以生成相应FlEx等位基因,所得等位基因就可以是敲除等位基因或减效等位基因,即其中被捕获的基因的表达被下调的等位基因。用Flp重组酶处理这些FlEx等位基因原则上应将GT元件倒转至反义链,从而减轻在捕获元件内的转录终止,并且由此将经修饰的基因转换为条件GT。目前“隐藏”在反义链中的这种条件GT可以通过暴露于Cre再活化,这将通过作用于FlEx阵列的loxP/lox511偶联体再将其倒转。
FlEx技术的这种应用依赖于GT元件以生成无效等位基因。它因此受到基因捕获技术的局限性,这不保证将生成真实敲除且具有灭活调节元件(通过随机插入灭活)的另外风险。GT元件的替换以及它在终止转录中的有效程度都可以影响任何给定等位基因是否将是无效等位基因。另外的问题是对于不具有已确定功能的大多数基因,并且通过确定无效等位基因研究测定的基因与表型的相连尤其如此,非常难以决定性证明GT等位基因是真实地无效等位基因。事实上,对于这些以及其他,主要是技术原因,在通过大规模小鼠诱变共同体—EUCOMM的4年适应和使用后,基于FlEX的基因捕获方法已被抛弃,支持使用KO-first的基因靶向。
类似于FlEx方法,一般的目前KO-first等位基因至少部分依赖于GT样元件(SA-LacZ-polyA或SA-βgeo-polyA),以生成敲除样等位基因。然而,在已与那种方法相关的局限性的认识中(由用GT获得的经历有效学习的,以及理论考虑),KO-first还要求GT样元件下游的floxed(即,在序列两侧加上loxp位点的)关键外显子必须被缺失(使用Cre),以便生成真实的无效等位基因。因此,在实践中,这种方法首先要求FRT侧接的报道分子/GT样盒加上药物小基因置于靶基因的内含子内,有时在待缺失的外显子上游,同时对提名为待缺失的外显子进行序列两侧加上loxp位点(floxing)。这种外显子已称为“关键外显子”,并且与用于限定“关键外显子”的标准无关,KO-first方法明确要求其去除,以便致使所得等位基因成为真实无效的。因此,在靶向后,所得等位基因不是真实无效的也不是条件-无效等位基因。所得等位基因不是真实无效的原因已归于下述事实:如果没有通过Cre去除关键外显子(其是经floxed的),存在GT样盒周围的通读转录和剪接,以及由于药物小基因的存在,GT样盒下游的基因信使的转录的可能性。所得等位基因不是条件-无效等位基因的原因在于下述事实:如果没有报道分子/GT样盒和药物小基因的去除,正常信息(正常组成、以及表达水平和位点)的生成不能发生。
因此,取决于所需用途—无效或条件-无效—必须对KO-first等位基因实施第二个靶向后步骤。在其中需要真实无效的情况下,KO-first等位基因必须用Cre重组酶处理,以缺失关键外显子(其是floxed)。
相反,在Flp介导的报道分子/GT样盒和药物小基因的去除后,可以生成条件等位基因,其一起邻侧为FRT位点。以这种方式,保留的唯一修饰是FRT位点和floxed“关键”外显子。这种等位基因依次可以通过Cre介导的floxed外显子去除转换为无效的。
尽管KO-first方法解决了FlEx的某些局限性,但它仍受限制其效用的3个主要缺点所阻碍:首先,尽管它校正GT样元件生成真实的KO-first的可靠性的缺乏,但它未能提供真实的KO-first而无需另外的靶向后步骤;其次,由于用于限定“关键外显子”的标准,KO-first局限于蛋白质编码基因,有效放置所有非蛋白质编码基因(即编码‘非编码’RNA的那些,非常重要的生物分子类别)是力所不及的。此外,在蛋白质编码基因中,仅对于其可以限定“关键外显子”的那些顺应KO-first设计。用于限定关键外显子的标准是其缺失导致在其前的开放读码框(ORF)部分和在其后的ORF部分之间的移码。这是因为这个移码的诱导对于KO-first方法是强制性的,以提供限定的敲除。因此,即使特定类别的蛋白质编码基因也不顺应KO-first设计。这些包括其中ORF包含在一个外显子内的基因,以及其中所有或大多数串联外显子脱离相同框的基因。第三,一旦KO-first等位基因已转换成条件-无效等位基因(通过Flp的作用),在条件等位基因转换为无效状态后,所得等位基因就不提供用于无效的肯定报道的任何机制。一般地,首先敲除等位基因去除报道分子(例如lacZ)连同DSC,使用(使用Flp重组酶完成的步骤,因为在KO-first中使用的SA-lacZ-polyA和DSC是FRTed),仅留下floxed外显子(加上其上游的FRT位点)。因此,在常规首先敲除方法中不存在关于报道无效等位基因达到的报道分子功能的选项。
多功能等位基因
提供了多功能等位基因(MFA)方法,其通过引入包含起动序列(其证实去除或灭活)的一组功能元件,允许去除或灭活在基因组中的核苷酸序列,导致真实的敲除,并且还含有一种或多种条件元件,其在等位基因中的表达是条件的(即取决于特定分子事件或线索)和可报道的。
MFA方法提供了生成真实首先敲除等位基因的靶向选项,其不需要第二个靶向后步骤以将靶向等位基因转换为无效状态,从而提供超过一般的目前首先敲除等位基因(其需要靶向后步骤以将靶向等位基因转换为无效等位基因)的优点和概念突破。MFA方法也不限制于与“关键外显子”的使用并且不限于通过移码的敲除,但一般可应用于修饰任何目的核苷酸序列。MFA提供了在单个靶向步骤后增强的多样性和多重等位基因选项。
MFA方法提供了真实的KO-first等位基因,其提供了用于在与起动序列连接的任何所选序列倒转后,在受体基因组中产生第二种状态的机会,并且其中在倒转后,可以报道所选序列的转录。所选序列可以是例如COIN,无限制地,其自身可以包含起动序列,其例如包含阻遏子以控制另一种基因或调节序列的转录。在一个例子中,将MFA置于基因座上的单个靶向步骤可以允许野生型基因座修饰为特定状态,状态A(例如内源基因的敲除)。MFA设计为通过第一种重组酶的作用,使得状态改变至另一个特定状态,状态B(例如野生型表型的修复)的变化成为可能。状态B可以通过第二种重组酶转换为状态C(例如敲除的重新建立和COIN的表达),等等。因此,当采用MFA时,改变在所选基因座上的状态可以由单一起始等位基因达到。
MFA方法可以用于放置作为基因捕获的MFA,从而使得采用内源转录活性的启动子,包含MFA的转基因获得MFA元件的表达。
MFA方法可以用于常规转基因生成(transgenesis)应用中,其中起动序列包含启动子、外显子、或外显子和内含子、和任选地转录控制元件以及随后为DSC(任选的,因为这对于常规、基于原核注射的转基因生成是不需要的)、就第一种起动序列而言以反义方向的NSI(其可以是第二种起动序列)、和也置于就第一种起动序列而言的反义方向的COIN(其可以是第三种起动序列)。
MFA方法提供了用于在单个靶向步骤中产生基因座的多重选项,所述基因座含有无效等位基因、条件无效等位基因、COIN、起动序列(其可以包括报道分子)和DSC、及其他元件。基因座的靶向后操作提供了在单个靶向步骤中通过使用引入含MFA基因座内的可重组单位与MFA构建体一起的选项。通过采用不相容的不同对的同源位点特异性重组酶识别位点(例如一对FRT位点和一对FRT3位点、一对loxP位点和一对lox2372位点等),减少在基因座靶向后上实行多个可操作选项所需的不同重组酶的数目。因此,含MFA基因座暴露于单一重组酶可以独立地作用于至少2个不同的可重组单位,以这样重组单位,从而使得单位将目的元件(例如报道分子、DSC、外显子、COIN等)置于所需方向,以及在可重组单位内重新定向位点特异性重组酶识别位点,以形成新的可重组单位。
MFA方法提供了关于COIN的选项,所述COIN可以含有任何所需序列,包括但不限于报道分子、或编码突变体或变体形式的靶基因或靶基因部分的cDNA、或靶基因的相关物和同系物、或甚至非蛋白质编码序列例如微小RNAs或微小RNA的簇、或这些元件的任何组合(因为它们可以通过内部核糖体进入位点或“自切割”肽在不同元件之间的放置进行调节—取决于元件的选择)。
MFA方法提供了其中通过靶向达到敲除的选项,第一种重组酶用于使敲除元件重新建立回到活性(野生型)状态,并且第二种重组酶用于重新建立敲除,其中COIN置于有义方向伴随敲除元件的重新建立或敲入,因此报道在其中第二种重组酶已活化的细胞中敲除的重新建立。
尽管目前的首先敲除方法缺乏关于在条件等位基因转换为无效后报道无效的机制,但MFA方法提供了关于报道元件(例如COIN,参见图5和图6,在底部;通过基因分型和/或通过显现或其他定性或定量测定可检测的,任选在细胞水平)的选项,在第二种重组酶作用后,其倒转或倒转且切除目的核苷酸序列(例如外显子和周围序列,在图5和图6中的NSI),且将报道元件置于有义方向,有效报道NSI的倒转和/或切除。在这个实施方案中,在单个靶向步骤后无效等位基因通过第一种重组酶转换为恢复的等位基因(在这个实施方案中,图5和图6的NSI是由靶向载体替换的基因或其部分的外显子和周围序列),且通过第二种重组酶缀合物转换为无效等位基因,这通过将COIN置于有义方向报道其存在。
因此,MFA方法提供了关于下述的选项:评估敲除的表型效应(在靶向步骤后),随后暴露于第一种重组酶以重新建立敲除的外显子或基因或其区域,评估重新建立的表型效应——即,转换回野生型(等价于补充测定的步骤但缺乏生成新转基因小鼠系的需要,已常规地伴随补充分析的需要),随后任选暴露于第二种重组酶以重新建立敲除,且评估重新建立无效等位基因的表型效应。因此,MFA等位基因组合真实的首先敲除方法与另外(条件)元件的多样性,和在包含单个靶向步骤的方案中在经遗传修饰的动物中进行真实补充型分析的能力。
在一种MFA应用中,提供了用于补充测定的方法,其包括用依照本发明的MFA靶向细胞的内源等位基因,随后在靶向后步骤中,由MFA生成条件-无效等位基因(通过暴露于第一种重组酶),其中在MFA中的目的核苷酸序列包含以有义方向的外显子或外显子加上周围序列或另一个目的区域(与例如表型相关),并且评估条件-无效等位基因(其应是野生型的)的表型效应。在进一步的实施方案中,MFA进一步暴露于重新建立无效的第二种重组酶,并且任选地再次测量表型效应。在特定实施方案中,第二种重组酶还将条件报道分子(例如COIN)置于有义方向,其中条件报道分子报道条件-无效等位基因至无效等位基因的转换,或如情况可以是报道无效的重新建立。
在一个实施方案中,NSI包含外显子和邻近内含子序列、或靶基因的外显子-内含子区。在另一个实施方案中,NSI包含编码ncRNA、微小RNA、微小RNA簇或其他一种或多种小ncRNA的区域。
MFA是可以在基因组中随机放置或靶向选择的基因座的等位基因。通过序列在同源位点特异性重组酶识别位点对的阵列中的慎重放置,将MFA进行改造以产生无效、条件、或组合条件/无效等位基因。通过构建体置于基因组中产生的所得等位基因可使用所选重组酶进行操作,所述重组酶可以瞬时或通过在其基因组中包含构建体的动物与包含关于所选重组酶的基因的动物(例如Cre-、Flp-或PhiC31\int-表达品系)的育种引入至基因组中的构建体。
在多种实施方案中,提供了用于生成真实的首先敲除等位基因的方法和组合物,其中无效不取决于执行第二个步骤,例如通过重组酶的作用去除“关键外显子”或“关键区”。相应地,提供了用于在单个靶向步骤中生成其为多功能的等位基因的实施方案,因为它是在单个靶向重组步骤中达到的具有报道分子的真实的首先敲除等位基因。
通过MFA方法用于生成敲除等位基因的方法和组合物并不限于经由关键外显子倒转的缺失生成移码的需要,以生成无效等位基因,如由某些类型的敲除等位基因需要的(例如KO-first或FlEx的某些实施方案)。相反,MFA方法依赖于其在靶向时去除来自靶基因的转录单位的NSI的能力,同时NSI的表达替换为起动序列的那种。起动序列可以包含GT样元件(例如报道分子例如SA-lacZ-polyA)、cDNA、一个或多个外显子、调节元件(例如增强子、绝缘子、操纵子)。因为起动序列是实验者限定的,所以可以同样良好地致使除无效外的等位基因成为可能。例如,起动序列可以编码显性失活或者组成活性或活化形式的基因。
在多种实施方案中,MFA包含各自在所得等位基因内以反义方向的目的核苷酸序列和COIN,且进一步包含在等位基因中都以有义方向(或以有义和反义方向,或各自独立地以有义或反义方向)的起动序列和/或DSC,因此在暴露于第一种重组酶后,起动序列和/或DSC缺失,目的核苷酸序列倒转至有义方向,并且COIN维持在反义方向。等位基因进一步包含这样放置的重组位点,以便允许通过第二种重组酶后续同时倒转目的核苷酸序列和COIN,从而使得在第二种重组酶作用后,目的核苷酸序列置于反义方向,并且COIN置于有义方向。在进一步的实施方案中,目的核苷酸在用第二种重组酶处理后缺失,留下以有义方向的COIN。在特定实施方案中,COIN是报道分子或DSC。在另一个特定实施方案中,目的核苷酸是一个物种的基因的外显子或目的区域(例如小鼠、大鼠、非人灵长类动物或人外显子),并且COIN是另一个物种的基因的外显子(例如小鼠、大鼠、非人灵长类动物或人外显子)。
MFA方法还允许基因捕获方法。在MFA方法的这个实施方案中,MFA插入转录活性的基因座上。这可以通过随机重组或通过“靶向捕获”(参见例如,通过引用合并入本文的美国专利号7,473,557)来实现。在剪接受体和剪接区后且随后为polyA信号的起动序列提供了任何现有转录的基因组序列的敲除或“敲减(knockdown)”。包括无启动子DSC即其表达取决于在转录活性的基因座的有义链内的插入的DSC,确保含有MFA的细胞的阳性选择。包括以反义方向的目的核苷酸序列(NSI)连同以反义方向的COIN,与位点特异性重组酶识别位点的推荐排列结合,提供了在暴露于第一种重组酶后条件表达NSI(来自捕获基因座的启动子)的能力。随后,在暴露于第二种重组酶后,NSI的表达可以关闭且同时被COIN的那种所替换。无启动子DSC确保所选的任何细胞将具有表达无启动子NSI和无启动子COIN的能力,并且表达将依照来自转录活性的基因座的内源表达模式。
为方便起见且不作为限制,使用MFA的特定有利方法与特定实施方案(即,就包含如图中所示的特异性命名的重组酶位点和核苷酸序列的等位基因而言)结合方便地描述,即合适的重组酶和重组酶识别位点、起动序列、报道分子、DSC和目的核苷酸序列可以基于本文公开内容常规地选择。“目的核苷酸序列”或“NSI”可以是任何目的核苷酸序列,例如外显子、外显子加上一种或多种侧接序列、2个或更多个外显子、编码序列的片段、完整编码序列、调节元件或序列、非蛋白质编码序列、内含子或其任何组合等。COIN可以包含cDNA以及非蛋白质编码序列,并且可以并入元件例如多聚腺苷酸化信号和位点、微小RNAs或其他非蛋白质编码RNA、IRES、密码子跳过肽及其任何组合。特定COIN和关于使用其的某些系统可以例如在美国专利号7,205,148中找到。
提供了用于在任何细胞包括非人动物细胞和非人动物中制备且使用MFA的方法和组合物。可以采用使用同源重组(或随机整合)的方法和组合物,以将有用的等位基因置于细胞的基因组中的任何所选位点(或随机位点)上。该方法和组合物可以用于多能、经诱导的多能和全能细胞中。用于与该方法和组合物一起使用的合适细胞包括ES细胞,例如小鼠或大鼠ES细胞。在多种实施方案中,提供了真实的KO-first等位基因,其提供了关于具有嵌入报道分子功能的条件功能性的选项。
在图2中举例说明了元件和重组酶识别位点的排列可以如何设计以产生构建体的例子,所述构建体将取消靶基因的功能(即,产生无效等位基因),或改变靶基因的功能(例如,将其变成显性失活、组成活性或减效等位基因),同时包埋所有下游元件,其将允许(a)条件等位基因的生成,和(b)其使用报道分子恢复至无效。
图2显示可以置于基因组内的MFA的实施方案(例如使用关于所示MFA的左和右侧的同源臂)。在靶向后,所得等位基因可以转换为条件等位基因,其伴随缺失第一种所选序列且倒转第二种所选序列。缺失和倒转可以通过相同重组酶或不同重组酶而实现。例如,可以使用2对不相容的Flp识别位点—一种指导缺失而另一种指导倒转。2个此类Flp位点的一个例子是FRT位点和FRT3位点。在另一个例子中,可以使用2对不相容的Cre位点,例如loxP和lox2372—一种指导缺失而另一种指导倒转。进一步地,可以使用2种不同的重组酶(例如关于Cre的一对loxP位点和关于Flp的一对FRT位点)。对于这个实施方案可以选择任何合适位点,只要位点可以指导图2中所示MFA的重组酶位点对的缺失和倒转(其特异性实施方案显示于图5和图6中)。
尽管图2的构建体设计可以与任何目的序列(即NSI是任何目的序列)一起使用,但构建体设计可以特别用于将目的外显子替换为经修饰的外显子。
在一个实施方案中,NSI是天然存在的外显子(或多个外显子),并且COIN是经修饰的外显子(或包含突变的外显子)。MFA通过例如同源重组(使用合适的小鼠同源臂)置于例如小鼠ES细胞的基因组内,并且ES细胞用于制备在小鼠种系中包含构建体的经遗传修饰的小鼠。在一个实施方案中,通过重组酶对MFA的作用实现从天然存在的外显子到经修饰的外显子的状态变化。
在一个实施方案中,构建体置于例如小鼠的基因组内,并且小鼠进一步包含其活性可以调节的重组酶(例如Cre)。重组酶可以通过例如采用融合蛋白进行调节,所述融合蛋白将重组酶置于效应物或代谢产物(例如CreERT2,其活性受它莫西芬正面控制)的控制下,将重组酶置于组织特异性启动子的控制下,或将重组酶置于在特定发育阶段时有活性的启动子(例如Nanog启动子)(或其他调节元件)的控制下,或诱导型启动子(例如其活性受多西环素控制的启动子和TetR或TetR变体)的控制下,或这些技术的组合。
图2中所示的MFA实施方案具有元件,其包含编码起动序列的序列、DSC、目的核苷酸序列(NSI)和COIN,其中元件置于这样选择的重组酶识别位点的阵列中,以便对MFA提供所需功能性。
图2的顶部举例说明在选择的基因组中的目的核苷酸序列(NSI)(例如在小鼠基因组中的NSI)。如所示的MFA通过例如同源重组引入基因组内以替换NSI。NSI替换为所示的MFA,其中MFA的NSI如所示的倒转且因此不再并入靶基因的转录物内。如果起动序列含有报道分子(例如lacZ),那么MFA的存在可以方便地加以证实。DSC同样存在,以帮助选择经修饰的细胞(例如用MFA修饰的小鼠ES细胞)。
图2的MFA实施方案包含通过5组重组酶识别位点限定的5个不同的序列单位。图3含有邻侧为相容的重组酶识别位点的5个不同的序列单位。
第一个不同的可重组单位包含以相反方向(即指导倒转)的R1/R1’位点(例如FRT3位点),其中在R1/R1’位点之间排列下述:起动序列(为简单起见,在图3中未显示就起动序列而言5'的3'剪接区和受体、和就起动序列而言3’的polyA信号)、R2位点(例如Rox位点)、DSC、以与R1位点相同方向的R3位点(例如FRT位点)、R4位点(例如loxP位点)、和就靶基因的转录方向(即由反义链编码)而言以反义方向的目的核苷酸序列(NSI)、和以与R4位点相同方向的R5位点(例如lox2372位点)。当R3’位点(以图3A中所示的R3位点的相反方向)进一步包括在3’R1’位点的下游时,在识别R1/R1’的重组酶的存在下,单位将NSI倒转成用于转录的位置且缺失起动序列和DSC。在一个实施方案中,进一步序列包括置于反义链上的COIN,且随后为就单位的R4位点而言以相反方向的R4’位点(例如loxP位点),从而使得在暴露于识别R4/R4’的重组酶(例如Cre)后,COIN这样倒转,从而使得COIN的编码序列目前处于NSI的转录下游的位置中。
第二个不同的可重组单位(图3B)包含以相同方向(即指导缺失)的R2/R2’位点(例如Rox位点),其包含在R2/R2’位点之间排列的下述序列:DSC、以与第一个不同的可重组单位的R1位点相同方向的第一个R3位点(例如第一个FRT位点)、第一个R4位点(例如第一个loxP位点)、就靶基因而言以倒转(即反义)方向的NSI、就R4位点而言以相同方向的第一个R5位点(例如第一个lox2372位点)、以与R3位点相反方向的R1’位点(例如第二个FRT3位点)和R3’位点(例如第二个FRT位点)、COIN(就靶基因的转录而言以反义方向)、以与R5位点相同方向的R5’位点(例如第二个lox2372位点)、和以与R4相同方向的R4’位点(例如第二个loxP位点)。这种第二个不同的可重组单位可通过识别R2/R2’的重组酶切除。当包括在MFA中时,这个单位可以切除,以留下邻侧为R1和R2或R2’位点的起动序列(例如在某些实施方案中,报道分子,例如编码lacZ的序列)。
第三个不同的可重组单位(图3C)包含以相反方向(即指导倒转)的R3/R3’位点(例如FRT位点),其包含在R3/R3’位点之间排列的下述序列:R4位点(例如loxP位点)、就靶基因的转录而言以倒转(即反义)方向的NSI、以与单位(即图3C的)的R4位点相同方向的R5位点(例如lox2372位点)、和以与R3位点相反方向的R1’位点(例如FRT位点)。这个单位可以通过识别R3/R3’的重组酶(例如当R3/R3’位点是FRT位点时,Flp重组酶)的作用倒转,导致NSI置于用于转录和翻译的合适方向。
第四个不同的可重组单位(图3D)包含以相同方向(即指导缺失)的R4/R4’位点(例如2个loxP位点),其包含在R4/R4’位点之间排列的下述序列:以倒转(即反义)方向的NSI、各自就R4位点以相同方向的R5位点(例如第一个lox2372位点)和R1’位点(例如FRT3位点)和R3’位点(例如FRT位点)、COIN(以反义方向)、和以与R5位点相同方向的R5’位点(例如第二个lox2372位点)。在识别R4/R4’的重组酶(例如如果R4/R4’是loxP位点,那么例如Cre)的存在下,这个单位是可切除的。如果置于MFA内且暴露于R4/R4’重组酶(在不存在暴露于识别R1/R1’、R3/R3’的重组酶的情况下),这个单位将缺失且留下起动序列(例如在某些实施方案中,报道分子,例如编码lacZ的序列)和DSC。因此,这个单位允许这样的实施方案,其中当替换基因组中的序列(例如替换外显子)时,在识别R4/R4’的重组酶的存在下,MFA可以充当包含起动序列和DSC的无效等位基因。需要时,在识别R2/R2’的重组酶(例如当R2/R2’是Rox位点时,Dre重组酶)的作用后,MFA的DSC可以被去除,因为DSC将在上游和下游侧接以相同方向的R2/R2’位点。
第五个不同的可重组单位(图3E)包含以相同方向(即指导缺失)以及与第四个不同的可重组单位的R4/R4’位点相同方向的R5/R5’位点(例如2个lox2372位点),其包含在R5和R5’位点之间排列的下述序列:就彼此而言以相同方向但以对于第一个不同的可重组单位的R1位点以相反方向的R1’位点(例如FRT3位点)和R3’位点(例如FRT位点)、和就靶基因的转录而言的COIN(以反义方向)。
如本领域技术人员应认识到的,重叠的可重组单位这样描述为传达MFA的结构,而不是限制在MFA中的可能可重组元件。例如,本领域技术人员应认识到每个可重组单位包含在可重组单位内的位点特异性重组位点,并且重组酶对位点(跨越可重组单位)的作用实现MFA的所需和所述操作,其达到MFA的预期功能。例如,就图3而言,识别R1/R1’和R3/R3’的重组酶的作用起作用,以操纵所有5个可重组单位的部分,如它们在图3中概念上展示的。
一旦MFA置于基因组中的所需位置上,它就可以这样进行改造,从而使得它提供了具有报道功能的无效等位基因,其中无效等位基因可以这样再修饰(在靶向后重组酶介导的步骤中),从而使得它缺乏被以相同方向定向的由重组酶识别位点所侧接的所有序列。这个实施方案的例子显示于图4中,显示合适的重组酶识别位点的例子,其中除起动序列(此处,编码lacZ)外的所有元件在上游和下游邻侧为Rox位点。在暴露于Dre重组酶后,仅存在起动序列。Roxed序列的切除可以通过DSC(此处,含有neor)的丧失、和/或COIN的丧失、和/或NSI的丧失加以证实。结果是缺乏DSC、NSI和COIN的真实的无效等位基因。
如图2中举例说明和如图3顶部例示的MFA可以用于产生条件等位基因。通过选择在第一种情况下等位基因暴露于其的合适重组酶可以生成条件等位基因。在这个实施方案中的合适重组酶是将NSI倒转回有义链且留下以反义方向的COIN的重组酶。这可以例如通过使MFA暴露于识别R1/R1’以及R3/R3’的重组酶(例如当R1/R1’和R3/R3’选自FRT和FRT3位点时,Flp重组酶;关于特定实施方案,参见图5)来实现。简言之,一旦MFA置于基因组中的所需位置上,它就可以用于生成条件等位基因,其中图2和图3顶部的倒转NSI置于用于靶基因转录的方向,同时留下以反义方向的COIN且缺失起动序列和DSC。这个实施方案的例子显示于图5中,其中通过首先使等位基因暴露于Flp重组酶,引起通过FRT3位点指导的元件倒转,随后为Flp介导的通过FRT位点指导的缺失,去除含有lacZ的起动序列和含有neor的DSC。所得等位基因呈现以用于转录的方向的NSI,但留下以反义方向的COIN。
如图6中所示,无论Flp介导的倒转首先经由FRT位点(如图5中)还是FRT3位点(如图6中)发生,可以实现相同的条件等位基因。
在生成条件等位基因的实施方案中,在等位基因中保留的重组酶位点这样加以选择,从而使得用一种或多种合适的重组酶处理导致NSI的后续缺失(或NSI的再倒转)和COIN的倒转,从而使得等位基因导致就NSI而言的无效等位基因,还将COIN置于用于转录的方向。这个的例子使用loxP和lox2372位点进行显示,其各自独立地指导Cre介导的重组。尽管使用Cre反应位点,但可以使用任何合适的位点代替Cre位点。
如图7中所示,以有义方向(即处于用于转录和翻译的位置)的NSI置于就第一个lox2372位点而言的3’。在NSI后是以与第一个lox2372位点相同方向的第一个loxP位点,并且倒转(即反义)COIN置于第一个loxP位点的下游,并且倒转的COIN置于就第一个lox2372位点而言相反方向的第二个lox2372位点的上游。在第二个lox2372位点下游放置的是就第一个loxP位点而言以相反方向放置的第二个loxP位点。在用Cre处理后,这个排列允许经由lox位点的倒转,随后经由lox位点的缺失(参见图7)。所得等位基因含有以有义方向的COIN,即处于用于转录和翻译的位置。
在备选排列中(参见图8),loxP位点置于就NSI而言的5'(代替置于NSI和COIN之间),从而使得暴露于Cre导致NSI至反义方向和COIN至有义方向的倒转。
MFA方法提供了用于许多实施方案的选项。在特定实施方案中,在暴露于第一种重组酶后,元件和重组酶位点的排列如图5或图6的底部构建体中所示,其中如所示的FRT3位点是在所得等位基因中不具有同源位点的位点R1,如所示的FRT位点是在所得等位基因中不具有同源位点的重组酶位点R3,最左侧lox2372位点是与占据如所示的最右侧lox2372位点的同源重组酶位点R5’配对的位点R5,如所示的最左侧loxP位点是与由如所示的最右侧loxP位点提供的同源重组酶位点R4’配对的位点R4,并且如所示的Rox位点是在所得等位基因中不具有同源重组酶位点的位点R2。
在特定实施方案中,在暴露于第二种重组酶后,所得等位基因的元件和重组酶位点的排列如图7的底部构建体中所示,其中所示的FRT3位点是在所得等位基因中不具有同源位点的位点R1,lox2372位点是不与所得等位基因中的同源重组酶位点配对的位点R5,所示的FRT位点是不与所得等位基因中的同源重组酶位点配对的位点R3,所示的loxP位点是不与所得等位基因中的同源重组酶位点配对的位点R4,并且所示的Rox位点是不与所得等位基因中的同源重组酶位点配对的位点R2’。
在特定实施方案中,所得等位基因允许在暴露于第二种重组酶后COIN的表达。在特定实施方案中,COIN是报道分子或DSC。
在一个方面,提供了包含COIN、NSI、DSC、报道分子和重组酶位点的MFA,其这样排列,从而使得通过一种重组酶的作用将切除COIN、NSI和DSC而不切除报道分子(图11B),而用不同重组酶的作用将生成这样的等位基因,其缺乏DSC但将NSI置于有义方向,同时维持以反义方向的COIN(图11C)。这个所得等位基因具有这样排列的重组酶位点,从而使得通过进一步重组酶的作用将切除NSI且将COIN置于有义方向(图11D)。因此,在讨论的实施方案中,这个MFA将允许选择具有报道分子功能的真实敲除和DSC的去除、或NSI的放置,其中NSI的后续去除通过COIN以有义方向的伴随放置加以证实。关于此类等位基因的某些重叠的重组酶单位的示意图显示于图11A中,其中相似重组酶单位由相似虚线形状代表。
在一个方面,提供了包含COIN、NSI、DSC、报道分子和重组酶位点的MFA,其这样排列,从而使得通过一种重组酶的作用将切除NSI和DSC,但维持报道分子和COIN的方向(图12B),而用不同重组酶的作用将生成这样的等位基因,其缺乏DSC和报道分子但将NSI置于有义方向,同时维持以反义方向的COIN(图12C)。这个所得等位基因具有这样排列的重组酶位点,从而使得通过进一步重组酶的作用将切除NSI且将COIN置于有义方向(图12D)。因此,在讨论的实施方案中,这个MFA将允许选择具有报道分子功能的真实敲除和DSC的去除;或NSI的放置,其中NSI的后续去除通过COIN以有义方向的伴随放置加以证实。关于此类等位基因的某些重叠的重组酶单位的示意图显示于图12A中,其中相似重组酶单位由相似虚线形状代表。
在一个方面,提供了包含COIN、NSI、DSC、报道分子和重组酶位点的MFA,其这样排列,从而使得通过一种重组酶的作用将切除报道分子和DSC,且将NSI置于有义方向(图13B)。这个所得等位基因具有这样排列的重组酶位点,从而使得通过进一步重组酶的作用将NSI置于反义方向,同时将COIN置于有义方向(图13C)。因此,在讨论的实施方案中,这个MFA将允许由MFA产生条件等位基因。
在一个方面,提供了包含COIN和NSI、DSC、报道分子和不同排列的重组酶位点的MFA,其这样排列,从而使得通过所选重组酶的作用将切除报道分子和DSC,且将NSI置于有义方向(图14B)。这个所得等位基因具有这样排列的重组酶位点,从而使得通过进一步重组酶的作用将NSI置于反义方向,同时将COIN置于有义方向(图14C)。因此,这个MFA将允许由MFA产生条件等位基因。
在一个方面,提供了包含NSI、DSC、报道分子、COIN和重组酶位点的MFA,其这样排列,从而使得通过所选重组酶的作用将切除报道分子和DSC,且将NSI置于有义方向,同时维持以反义方向的COIN(图15B)。这个所得等位基因具有这样排列的重组酶位点,从而使得通过进一步重组酶的作用将NSI置于反义方向,同时将COIN置于有义方向(图15C)。因此,这个MFA也将允许由MFA产生条件等位基因。
实施例
实施例1:Hprt1MFA
Hprt1是在小鼠中X连锁的基因,并且Hprt1-无效ES细胞【203】对于核碱基类似物6-硫鸟嘌呤是抗性的(6-TG)。这个特性提供了简单和强壮的表型测试,因为对于Hprt1为野生型的细胞在6-TG的存在下死亡,而对于Hprt1无效的细胞存活。此外,如果靶向衍生自雄性胚泡的ES细胞(如一般地情况,以及对于目前在靶向中使用的大多数ES细胞系的情况),那么仅需要一轮靶向以生成Hprt1MFA/Y ES细胞。为了生成Hprt1MFA/Y ES细胞,根据图5(上)中所示的等位基因,根据美国专利号6,586,251和Valenzuela等人(2003)High-throughput engineering of the mouse genome coupled with high-resolution expression analysis,Nature Biotech.21(6):652-659中所述的方法(该专利和论文在此通过引用合并),通过标准基因工程方法和细菌同源重组,制备靶向载体中的MFA。Hprt1MFA等位基因设计在外显子3周围,限定外显子3及其直接5’和3’的保守内含子序列作为NSI(图9)。关于这个选择的原因在于外显子3以框2(f2)开始且以框0(f0)结束;通过延伸,在前的外显子(即外显子2)以框2(f2)结束,并且随后的外显子(即外显子4)以框0(f0)开始。这意味着如果这个NSI倒转成反义方向,那么外显子2致使离开就外显子4而言的框,因为外显子2以f2结束,并且外显子4以f0起始。以这种方式,如果在Hprt1MFA等位基因中存在经过起动序列—SA-lacZ-polyA(图10)的任何转录,并且还存在从最终mRNA中去除起动序列的剪接,那个mRNA不包含外显子3且编码无义序列,那么有效产生Hprt1-无效mRNA和表型。相反,对于Hprt1COIN-INV等位基因(通过用FLP或FLP的变体处理Hprt1MFA生成,以首先生成Hprt1COIN等位基因,随后通过用Cre处理,以生成Hprt1COIN-INV),如果存在经过COIN元件的SA-eGFP-polyA的转录(图10),并且还存在从最终mRNA中去除SA-eGFP-polyA序列的剪接,那个mRNA不包含外显子3且因此编码无义序列,那么有效产生Hprt1-无效mRNA和表型。
反义定向的NSI是Hprt1的外显子3和周围进化保守的内含子序列(图9),并且反义定向的COIN是SA-eGFP-polyA。靶向载体具有在第一个位点FRT3上游和第二个Rox位点下游的小鼠同源臂,其指导靶向进入Hprt1基因座内,从而使得它由其MFA形式替换,由此(a)就Hprt1的转录方向而言以有义方向的SA-LacZ-polyA元件,随后为就Hprt1的转录方向而言以反义方向的DSC,两者都在Hprt1的外显子3前,(b)外显子3置于就Hprt1的转录方向而言的反义方向,和(c)COIN元件置于外显子3下游就Hprt1的转录方向而言的反义方向,并且其中这些不同元件邻侧为位点特异性重组酶识别位点,一起置于如图3和图10中详述的可重组单位中,其中SA-LacZ-polyA是起动序列,并且Hprt1的外显子3加上侧接内含子序列是NSI。
根据美国专利号6,586,251和Valenzuela等人(2003)High-throughput engineering of the mouse genome coupled with high-resolution expression analysis,Nature Biotech.21(6):652-659中所述的方法(该专利和论文在此通过引用合并),制备靶向载体且电穿孔到ES细胞内。所得到的ES细胞具有Hprt1的MFA等位基因代替Hprt1的野生型形式。在任何进一步修饰前,Hprt1MFA/Y ES细胞对于用6-TG处理是抗性的(因为它们对于Hprt1是有效地无效的),证实MFA方法生成真实的首先敲除等位基因的有用性。在用Dre处理后,这个特性保存,同时细胞的基因型转换为Hprt1SA-LacZ-polyA/Y。尽管对于Hprt1基因座,这种修饰可以不改变报道分子(LacZ)的表达水平,也不具有任何表型后果(改变对于6-TG的抗性),这可能不是对于其他基因座的情况。在用FLP或FLP变体处理后,在有效等价于补充测试的步骤中,Hprt1MFA/Y ES细胞转换为Hprt1COIN/Y ES细胞,其是有效野生型的且因此对于6-TG敏感的。此外,这个操作恢复Hprt1信息的表达回到其野生型同一性。在用Cre处理后,Hprt1COIN/YES细胞转换为Hprt1COIN-INV/YES细胞,其对于Hprt1是有效地无效的并且因此对于6-TG抗性的。此外,这个操作导致Hprt1信使的野生型信使表达的取消,并且其伴随替换为由Hprt1的第一个外显子和eGFP(由COIN元件编码)组成的杂交信息,从而生成表达eGFP代替Hprt1的等位基因。表达eGFP的这个新特性可以任选用于评分COIN元件至有义链的倒转,并且具有在使得细胞分离成为可能中的进一步效用,其中这个事件已由其中存在2类细胞(Hprt1COIN/Y ES细胞和Hprt1COIN-INV/Y ES细胞)的细胞群体发生。因此,不仅COIN等位基因转换为无效的,而且该事件还通过新的可容易测量和有用的事件标记。
实施例2:Hprt1 MFA结果
用如图16A中所示的重组酶位点排列构建MFA,其具有以有义方向的LacZ报道分子(SA(adml)-gtx-LacZ-pA)、新霉素DSC(Neo)、涵盖关于Hprt1(外显子3)的关键外显子(ec)且侧接进化保守的内含子序列的以反义方向的NSI、和COIN(Gtx-SA-HA-myc3-TM-T2A-GFP-pA)。将MFA电穿孔到F1H4ES细胞内,并且就对于G418的抗性进行选择。随后,将G418抗性集落进行基因分型,以测定靶向。从筛选的总共96个集落中获得5个靶向的克隆(Hprt1MFA/Y)。当在补充有10μM6-TG的标准ES细胞培养基中培养时(其是利用的标准6-TG存活测定),发现所有5个这些克隆存活且繁殖,如对于其为Hprt1-无效的细胞所预期的(Doetschman,T.等人(1987)Targeted correction of mutant HPRT gene in mouse embryonic stem cells,Nature 330:576-578)。相比之下,亲本细胞系F1H4以及测试的任何非靶向克隆未能在6-TG的存在下生长。这些结果与先前已报道的那些一致(Doetschman等人(1987))。
在用重组酶FLPo处理后(Raymond,C.S.和Soriano,P.(2007)High-efficiency FLP and PhiC31 site-specific recombination in mammalian cells,PLoS ONE2:e162),Hprt1MFA等位基因转换为Hprt1COIN等位基因(图16B),产生Hprt1MFA/Y ES细胞。这个操作导致LacZ报道分子、DSC的去除、以及NSI再倒转成有义链。因此,所得等位基因(Hprt1COIN)是功能上野生型的,因为编码且表达野生型Hprt1mRNA。
在用重组酶Cre进一步处理后(Sauer,B.和Henderson,N.(1988)Site-specific DNA recombination in mammalian cells by the Cre recombinase of bacteriophage P1,Proc.Natl.Acad.Sci.USA 85:5166-5170),Hprt1COIN等位基因转换为Hprt1COIN-INV等位基因(图16C),产生Hprt1COIN-INV/Y ES细胞。这个等位基因(Hprt1COIN-INV)是功能上无效的,因为Hprt1mRNA由编码eGFP的mRNA替换(并且还缺乏如设计阶段时限定的NSI—即Hprt1的外显子3和侧接内含子序列)。
就对于核苷酸类似物6-TG的抗性来测试具有MFA(Hprt1MFA/Y)的细胞,并且与野生型细胞比较(图17)。Hprt1MFA/Y ES细胞存活,而Hprt1+/Y ES细胞死亡,指出Hprt1MFA/Y是功能上Hprt1-无效的。Hprt1MFA/YES细胞随后用FLPo处理,以测试Hprt1MFA等位基因是否转换为Hprt1COIN等位基因。所得到的Hprt1COIN/YES细胞预期为表型上野生型的,因为Hprt1表达恢复。这显示为事实上的情况,因为当在6-TG的存在下培养时,Hprt1COIN/YES细胞死亡,正如同其野生型(Hprt1+/Y)配对物。最后,Hprt1COIN/YES细胞用Cre处理,以生成Hprt1COIN-INV/YES细胞,其预测对于Hprt1是无效的,因为COIN模块被活化,同时缺失Hprt1的外显子3(图16,小图C)。当在6-TG的存在下培养时,Hprt1COIN-INV/YES细胞存活且增殖,证实它们对于Hprt1是功能上无效的,如由MFA设计和应用所预期的。
通过对属于每个基因型类别的ES细胞的蛋白质制备物执行Western印迹,在蛋白质水平进一步证实上文获得的表型结果:野生型(Hprt1+/Y)、Hprt1MFA/Y(MFA)、Hprt1COIN/Y(MFA+FLPo)和Hprt1COIN-INV/Y(MFA+FLPo+Cre)。就报道分子和NSI(即Hprt1)表达检查这些蛋白质制备物。Hprt1MFA/YES细胞缺乏Hprt1蛋白质,但表达报道分子(LacZ)。在Hprt1COIN/YES中,Hprt1的表达恢复为野生型水平,反映NSI的放置(Hprt1的外显子3)回到有义方向,并且未显示报道分子(LacZ)蛋白质,证实通过FLPo的报道分子切除。这确定Hprt1MFA等位基因是事实上无效的,并且在去除报道分子和DSC、和NSI伴随再倒转成有义链(实验上通过FLPo完成的操作)后,可以转换为功能上野生型的等位基因。在Hprt1蛋白质表达水平上Hprt1COIN等位基因等同于野生型(Hprt1+)的事实,进一步证实这种方法在一个重组酶介导的靶向后步骤中生成真实的条件-无效且执行等价补充测定的稳固性。最后,Hprt1COIN-INV/YES细胞缺乏Hprt1蛋白质,有效证实使用6-TG抗性测定做出的表型观察。这进一步证实基于COIN的条件-无效等位基因(Hprt1COIN)如预期的起作用。
- 还没有人留言评论。精彩留言会获得点赞!
- 油菜BnaSPT1基因在促进双子叶植物角果生长中的应用的制造方法与工艺
- 油菜BnaSPT3基因在促进双子叶植物角果生长中的应用的制造方法与工艺
- 一种木质素合成途径中上位性效应检测方法与制造工艺
- 用于检测人类HLA‑B*5701等位基因的核酸、试剂盒及方法与制造工艺
- 玉米低镁条件下特异性转运镁离子基因ZmMGT10及其应用的制造方法与工艺
- 与癌症相关的基因融合体和基因变异体的制造方法与工艺
- CRISPR‑Cas9特异性敲除猪SLA‑3基因的方法及用于特异性靶向SLA‑3基因的sgRNA与制造工艺
- 一种高通量测序检测无创亲子鉴定的方法与制造工艺
- 构建在海马体区域特异性敲除IKKα基因的小鼠模型的方法及打靶载体和试剂盒与制造工艺
- 通过分析预示性基因集诊断亚临床和临床的急性排异的方法与制造工艺