一种优化的pIFN‑γ肽链及在提高毕赤酵母分泌表达猪IFN‑γ产量和活性中的应用的制作方法
本发明涉及基因工程
技术领域:
,具体地,涉及一种优化的pIFN-γ肽链及在提高毕赤酵母分泌表达猪IFN-γ产量和活性中的应用。
背景技术:
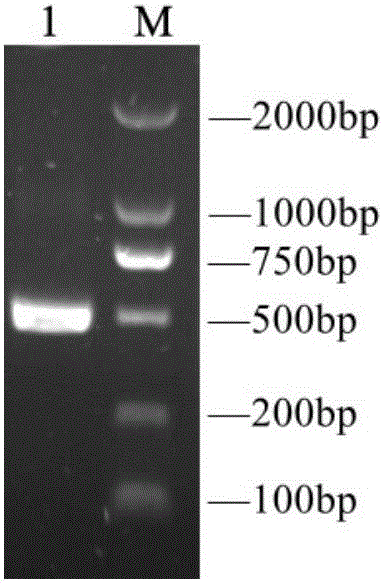
:猪伽马干扰素(porcineInterferonGamma,pIFN-γ)是一种具有广谱抗病毒、抗肿瘤活性和免疫调节功能的细胞因子。猪伽马干扰素属于Ⅱ型干扰素中的唯一成员,主要是由活化的T细胞和NK细胞产生的一种具有广谱抗病毒活性和免疫调节功能的细胞因子。猪伽马干扰素在生猪养殖中具有广阔的运用前景,在生猪养殖中可作为抗生素类替代药品、猪肿瘤性疾病的治疗及流行性疾病的防治等用途。目前国内研究者构建的各类pIFN-γ表达菌株存在目的蛋白表达效率低下,采用分泌表达方式表达量一般为100~300mg/L,采用胞内表达方式目的蛋白纯化成本高昂,产量一般占细胞总蛋白1/5~1/3;此外,表达的pIFN-γ还存在活性较低的问题,阻碍了pIFN-γ在生猪养殖中的进一步运用。为此,构建一种能够高效分泌表达具有高活性的pIFN-γ工程菌对于推动pIFN-γ在生猪养殖业中的应用是十分必要的。目前,未见有关能够提高pIFN-γ分泌表达水平及其活性的改良氨基酸序列的报道。技术实现要素:本发明为了克服现有技术的上述不足,提供了一种优化的pIFN-γ肽链,优化的pIFN-γ肽链能提高毕赤酵母分泌表达猪IFN-γ产量和活性。本发明的另一个目的是提供一种优化的pIFN-γ肽链提高毕赤酵母分泌表达猪IFN-γ产量和活性中的应用。本发明的另一个目的是提供一种提高毕赤酵母分泌表达猪IFN-γ产量和活性的方法。为了实现上述目的,本发明是通过以下方案予以实现的:一种提高毕赤酵母分泌表达猪IFN-γ产量和活性的pIFN-γ肽链,pIFN-γ肽链的氨基酸序列如SEQIDNO:1所示。优化的pIFN-γ肽链命名为pIFN-γ-C。天然未经过改造的pIFN-γ肽链序列如SEQIDNO:2所示。本发明提供的优化的pIFN-γ肽链是在天然pIFN-γ肽链序列第129和131位氨基酸残基由精氨酸(R)定向突变为组氨酸(H)得到的。SEQIDNO:1所示氨基酸序列的pIFN-γ肽链在提高毕赤酵母分泌表达猪IFN-γ产量和活性中的应用。一种提高毕赤酵母分泌表达猪IFN-γ产量和活性的方法,具体为将SEQIDNO:1所示序列的pIFN-γ肽链整合到毕赤酵母中,得到的重组毕赤酵母分泌表达的猪IFN-γ的产量和活性大大提高。一种提高毕赤酵母分泌表达猪IFN-γ产量和活性的方法,包括如下步骤:将SEQIDNO:1所示序列的pIFN-γ肽链对应的核苷酸片段插入质粒pPICZαA中,重组质粒转入毕赤酵母X33野生型感受态细胞,利用质粒上的抗性基因,通过高抗筛选得到具有目的基因高拷贝的毕赤酵母重组子,获得能高效分泌表达具有高活性pIFN-γ的毕赤酵母工程菌。与现有技术相比,本发明具有如下有益效果:将本发明提供的优化后的pIFN-γ肽链整合到毕赤酵母中,可以大大提高毕赤酵母分泌表达猪IFN-γ产量和活性,因此,本发明提供的优化后的pIFN-γ肽链对于推动pIFN-γ在生猪养殖业中的应用是十分必要的。附图说明图1为引入酶切位点的pIFN-γ-C基因高保真PCR扩增结果,图中M为DL2000标准分子量Marker,泳道1为引入酶切位点的pIFN-γ(484bp)。图2为菌落PCR筛选DH5α阳性转化子(阳性克隆约1000bp),图中M为DL2000标准分子量Marker,阳性克隆长度约1000bp。图3为质粒pPICZαA-pIFN-γ-C反向测序结果。图4毕赤酵母转化子的高抗筛选结果,图中1为0.5mg/mLZeocin平板筛选结果,2为1mg/mLZeocin平板筛选结果,1和2中菌落为原位一一对应关系。图5为诱导发酵上清SDS-PAGE电泳结果;1-4号泳道为抗1.0mg/mLZeocin的酵母转化菌株诱导表达发酵上清,第5号泳道为X33野生型诱导表达发酵上清,表达的pIFN-γ-C理论分子量为17.5Kda。图6为pIFN-γ-C蛋白的Western-blot鉴定结果;泳道1-8为抗1.0mg/mLZeocin的酵母转化菌株诱导表达发酵上清,M为Bio-RadDualColor标准分子量预染Marker,表达的pIFN-γ-C理论分子量为17.5Kda。图7为Ni-亲和层析纯化pIFN-γ-C结果;泳道1-8为Eltionbuffer洗脱样品,表达的pIFN-γ理论分子量为17.5Kda。具体实施方式下面结合具体实施例对本发明作出进一步地详细阐述,所述实施例只用于解释本发明,并非用于限定本发明的范围。下述实施例中所使用的试验方法如无特殊说明,均为常规方法;所使用的材料、试剂等,如无特殊说明,为可从商业途径得到的试剂和材料。实施例1天然未经过改造的pIFN-γ肽链序列如SEQIDNO:2所示,本发明利用本领域常用的基因突变方法将天然pIFN-γ肽链序列第129和131位氨基酸残基由精氨酸(R)定向突变为组氨酸(H),得到SEQIDNO:1所示的优化的pIFN-γ肽链。(1)根据本发明提供的优化的pIFN-γ基因的核苷酸序列SEQIDNO:1为表达基因,分泌表达pIFN-γ突变体。优化的pIFN-γ-C基因通过体外全基因合成得到。全基因合成得到的pIFN-γ序列高保真PCR结果如图1所示(目的产物上下游分别引入XhoⅠ与XbaⅠ酶切位点)。(2)构建pIFN-γ毕赤酵母重组分泌表达载体:将毕赤酵母分泌表达载体pPICZαA与上述步骤(1)中得到的pIFN-γ基因高保真PCR产物用XhoⅠ与XbaⅠ进行双酶切,用PCR纯化回收试剂盒和DNA琼脂糖凝胶回收试剂盒分别对基因和质粒双酶切产物进行回收,将回收的基因与质粒用T4连接酶连接,转化大肠杆菌DH5α感受态细胞,构建得到pIFN-γ重组毕赤酵母分泌表达载体pPICZαA-pIFN-γ-C,保存载体的菌株命名为DH5α-pPICZαA-pIFN-γ-C。用质粒通用引物5’AOXⅠ、3’AOXⅠ通过菌落PCR筛选DH5α阳性转化子结果如图2所示,将所有阳性克隆菌落送样测序,将测序结果完全一致的菌落保种,测序结果如图3所示。(3)pIFN-γ-C毕赤酵母分泌表达菌株的构建与高拷贝菌株的筛选:将上述步骤(3)中构建的重组表达载体用SacⅠ单酶切线性化后,电击转化毕赤酵母野生型X33感受态细胞,涂布YPDSZ筛选平板,置于30℃培养箱中培养至长出单菌落,随机挑取约100个单菌落分别溶于10uL无菌水中,用无菌牙签沾取菌液分别一一对应接种于含0.5mg/mL和1.0mg/mLZeocin的两块YPDZ平板,将平板置于30℃培养箱中培养,通过高浓度的Zeocin(博来霉素)YPDZ平板筛选具有目的基因高拷贝的毕赤酵母转化子。毕赤酵母转化子的高抗筛选结果如图4所示。(4)pIFN-γ-C毕赤酵母分泌表达菌株的诱导发酵:将上述步骤(3)中得到的抗1.0mg/mLZeocin的酵母表达菌株接种于BMGY液体培养基,30℃、230rpm震荡培养至OD595nm值5-6时,按照无菌操作要求,离心收集菌体沉淀,按4:1或5:1的比例(如40或50mLBMGY液体培养基培养物转入10mLBMMY液体培养基中),将菌体沉淀转接于含1%甲醇的BMMY液体培养基,28℃、230rpm诱导发酵48小时,每隔24小时向BMMY液体培养基补加甲醇至1%,48小时后离心收集诱导发酵上清。诱导发酵上清SDS-PAGE电泳结果如图5所示;诱导发酵上清Western-blot结果如图6所示,Western-blot所用一抗为抗His标签鼠单克隆抗体,二抗为羊抗鼠IgG-HRP抗体。(5)诱导发酵上清中pIFN-γ-C的Ni亲和层析:按照氨酸亲和层析树脂填料Ni-NTAHis-Bind要求,对上述诱导发酵上清中的pIFN-γ-C进行纯化,Ni亲和层析结果如图7所示。(6)优化后的pIFN-γ的发酵产量和活性对比,以未优化的SEQIDNO:2序列通过质粒转入毕赤酵母并表达为对照,在5升发酵罐内发酵后,pIFN-γ和pIFN-γ-C的表达水平和活性分别如表1和2所示。表1为pIFN-γ和pIFN-γ-C5升发酵罐发酵水平实验组表达量(mg/L)pIFN-γ303pIFN-γ-C1921表2为pIFN-γ和pIFN-γ-C5升发酵罐发酵产品活性实验组活性单位(U/ml)pIFN-γ3.47×106pIFN-γ-C9.28×108由表1、2的结果可知,经优化的pIFN-γ-C的表达量和活性远高于未优化的pIFN-γ。上述结果说明调整pIFN-γ的氨基酸序列有得于毕赤酵母的表达及提高其活性。实施例2天然未经过改造的pIFN-γ肽链序列如SEQIDNO:2所示,本发明利用本领域常用的基因突变方法将天然pIFN-γ肽链序列第129和131位氨基酸残基由精氨酸(R)定向突变为组氨酸(H),得到SEQIDNO:1所示的优化的pIFN-γ肽链。(1)将SEQIDNO:1所示氨基酸序列的pIFN-γ肽链插入质粒pPICZαA,转入与毕赤酵母GS115野生型感受态细胞,利用质粒上的抗性基因,通过高抗筛选得到具有目的基因高拷贝的毕赤酵母重组子,可以获得能高效分泌表达具有高活性pIFN-γ的毕赤酵母工程菌。(2)优化后的pIFN-γ的发酵产量和活性对比,以未优化的SEQIDNO:2序列通过质粒转入毕赤酵母并表达为对照,在5升发酵罐内发酵后,pIFN-γ和pIFN-γ-C的表达水平和活性分别如表3和4所示。表3为pIFN-γ和pIFN-γ-C5升发酵罐发酵水平实验组表达量(mg/L)pIFN-γ248pIFN-γ-C1681表4为pIFN-γ和pIFN-γ-C5升发酵罐发酵产品活性实验组活性单位(U/ml)pIFN-γ1.77×106pIFN-γ-C6.94×108由表3、4的结果可知,经优化的pIFN-γ-C的表达量和活性远高于未优化的pIFN-γ。上述结果说明调整pIFN-γ的氨基酸序列有得于毕赤酵母的表达及提高其活性。实施例3天然未经过改造的pIFN-γ肽链序列如SEQIDNO:2所示,本发明利用本领域常用的基因突变方法将天然pIFN-γ肽链序列第129和131位氨基酸残基由精氨酸(R)定向突变为组氨酸(H),得到SEQIDNO:1所示的优化的pIFN-γ肽链。(1)将SEQIDNO:1所示氨基酸序列的pIFN-γ肽链插入质粒pPICZαA,转入与毕赤酵母KM71野生型感受态细胞,利用质粒上的抗性基因,通过高抗筛选得到具有目的基因高拷贝的毕赤酵母重组子,可以获得能高效分泌表达具有高活性pIFN-γ的毕赤酵母工程菌。(2)优化后的pIFN-γ的发酵产量和活性对比,以未优化的SEQIDNO:2序列通过质粒转入毕赤酵母并表达为对照,在5L发酵罐内发酵后,pIFN-γ和pIFN-γ-C的表达水平和活性分别如表5和6所示。表5为pIFN-γ和pIFN-γ-C5升发酵罐发酵水平实验组表达量(mg/L)pIFN-γ189pIFN-γ-C843表6为pIFN-γ和pIFN-γ-C5升发酵罐发酵产品活性实验组活性单位(U/ml)pIFN-γ3.59×106pIFN-γ-C8.42×108由表5、6的结果可知,经优化的pIFN-γ-C的表达量和活性远高于未优化的pIFN-γ。上述结果说明调整pIFN-γ的氨基酸序列有得于毕赤酵母的表达及提高其活性。SEQUENCELISTING<110>华南农业大学<120>一种优化的pIFN-γ肽链及在提高毕赤酵母分泌表达猪IFN-γ产量和活性中的应用<130><160>2<170>PatentInversion3.3<210>1<211>143<212>PRT<213>改造后的pIFN-γ肽链序列<400>1GlnAlaProPhePheLysGluIleThrIleLeuLysAspTyrPheAsn151015AlaSerThrSerAspValProAsnGlyGlyProLeuPheLeuGluIle202530LeuLysAsnTrpLysGluGluSerAspLysLysIleIleGlnSerGln354045IleValSerPheTyrPheLysPhePheGluIlePheLysAspAsnGln505560AlaIleGlnArgSerMetAspValIleLysGlnAspMetPheGlnArg65707580PheLeuAsnGlySerSerGlyLysLeuAsnAspPheGluLysLeuIle859095LysIleProValAspAsnLeuGlnIleGlnArgLysAlaIleSerGlu100105110LeuIleLysValMetAsnAspLeuSerProArgSerAsnLeuArgLys115120125HisLysHisSerGlnThrMetPheGlnGlyGlnArgAlaSerLys130135140<210>2<211>143<212>PRT<213>未改造pIFN-γ肽链序列<400>2GlnAlaProPhePheLysGluIleThrIleLeuLysAspTyrPheAsn151015AlaSerThrSerAspValProAsnGlyGlyProLeuPheLeuGluIle202530LeuLysAsnTrpLysGluGluSerAspLysLysIleIleGlnSerGln354045IleValSerPheTyrPheLysPhePheGluIlePheLysAspAsnGln505560AlaIleGlnArgSerMetAspValIleLysGlnAspMetPheGlnArg65707580PheLeuAsnGlySerSerGlyLysLeuAsnAspPheGluLysLeuIle859095LysIleProValAspAsnLeuGlnIleGlnArgLysAlaIleSerGlu100105110LeuIleLysValMetAsnAspLeuSerProArgSerAsnLeuArgLys115120125ArgLysArgSerGlnThrMetPheGlnGlyGlnArgAlaSerLys130135140当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1