本发明涉及一种陆地棉非特异性磷脂酶C基因GhNPC3a及其在提高棉花抗逆性中的应用。属于植物基因工程领域。
背景技术:
棉花是世界上最重要的天然纤维,干旱、低磷等非生物胁迫影响棉花的生长,最终导致棉花产量降低。在耕地面积只减不增的情况下,一些地方出现了粮棉争地的现象。在人类文明发展的长河中,穿衣和吃饭同等重要。解决粮棉争地的最好办法是培育抗逆性较强的优良棉花品种,及高效开发利用干旱区、盐碱地或土壤贫瘠化土地。
非特异性磷脂酶C,主要以磷脂酰胆碱为底物,生成二酰甘油和磷酸胆碱。拟南芥中,非特异性磷脂酶C基因家族有6个成员(Nakamura et al.2005)。2005年至今,相继有文献报导植物非特异性磷脂酶C参与植物激素的信号转导以及抗逆过程(Nakamura et al.2005;Peters et al.2010;Wimalasekera et al.2010;Kocourkova et al.2011;Pokotylo et al.2013;Nakamura 2014;Peters et al.2014;et al.2015;Pejchar et al.2015;Hong et al.2016)。然而,研究对象大多是模式生物拟南芥,其他物种中非特异性磷脂酶C的功能研究甚少。为了更好地了解植物非特异性磷脂酶C的功能,对棉花非特异性磷脂酶C基因家族进行研究是非常必要的,但检索发现迄今为止,没有任何棉花非特异性磷脂酶C的相关报导。
技术实现要素:
针对现有技术的不足,本发明的目的是提供一种陆地棉非特异性磷脂酶C基因GhNPC3a及其在提高棉花抗逆性中的应用。
本发明所述的陆地棉非特异性磷脂酶C基因,其特征在于:该基因命名为陆地棉非特异性磷脂酶C基因GhNPC3a,该基因的cDNA核苷酸序列如SEQ ID No.1所示,其编码的氨基酸序列如SEQ ID No.2所示。
本发明首次克隆出陆地棉非特异性磷脂酶C基因GhNPC3a,实验分析证实:GhNPC3a基因定位在A13染色体上,氨基酸的数目为508,含有4个外显子,3个内含子。MEME分析发现,GhNPC3a有16个motif。对得到的motif进一步在ExPASy中分析,发现有6个motif注释为磷酸酯结构域。用NCBI Conserved Domain search和SMART在线分析GhNPC3a的保守结构域,发现GhNPC3a含有磷酸酯结构域。对GhNPC3a进行启动子顺式作用元件分析,发现GhNPC3a的启动子中含有非常多的与胁迫相关或激素相关的顺式作用元件。如HSE(热胁迫相关),MBS(干旱胁迫相关),LTR(低温胁迫相关),TC-rich repeats(防御与胁迫相关),ARE(厌氧诱导相关),Box-W1(真菌诱导相关),ABRE(脱落酸相关),TGACG-motif和CGTCA-motif(茉莉酮酸甲酯相关),GARE-motif(赤霉素相关),AuxRR-core和TGA-element(生长素相关)和ERE(乙烯相关)。
对GhNPC3a进行不同器官的表达谱分析,实时荧光定量PCR结果表明,GhNPC3a主要在根中表达。对四叶期棉花分别进行低磷(5μM)、盐(200mM NaCl)、干旱(20%PEG6000)和ABA(200μM)的胁迫处理,分别在处理0,1,2,3,6,9和12h时进行取材,取材部位为根部。提取RNA并反转为cDNA,实时荧光定量PCR结果表明,低磷胁迫1h时,GhNPC3a表达水平上调,但是处理1h后表达水平下调;盐胁迫下,GhNPC3a表达水平下调;干旱胁迫处理1h和2h时表达水平分别上调5.7和2.9倍,随之表达水平表现出轻微下调的现象。ABA胁迫处理1h时,GhNPC3a表达水平上调3.6倍,2h和3h时上调幅度有所下降,6h之后表达水平与0h相比差异不显著。GhNPC3a基因的表达受低磷、干旱和ABA的胁迫诱导。
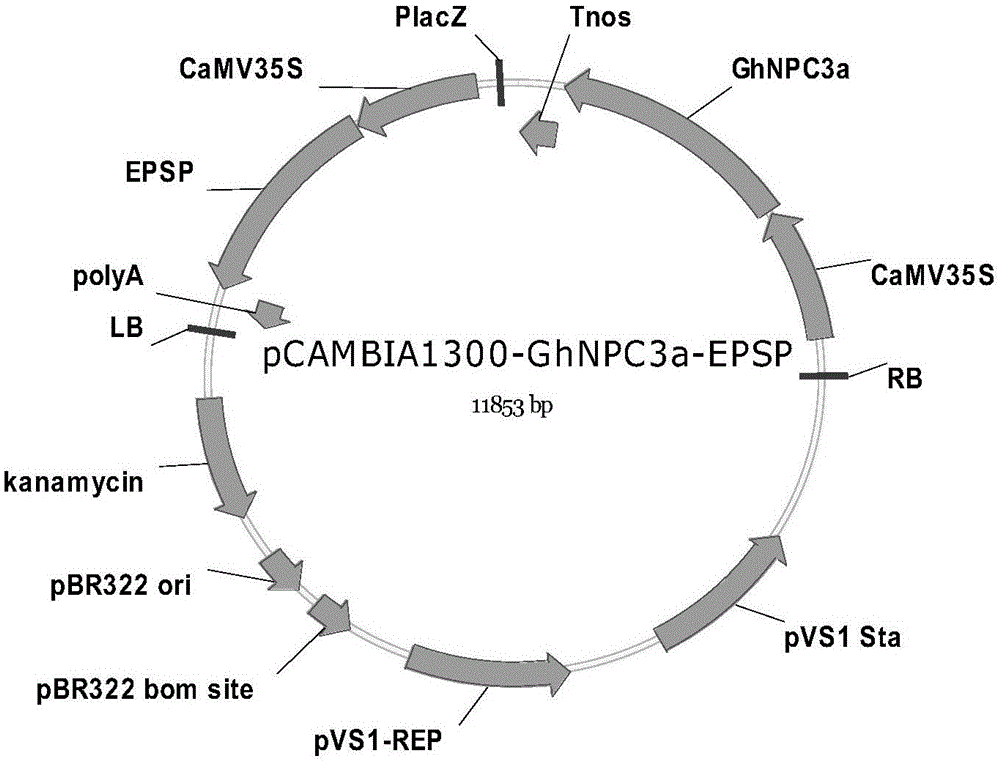
本发明还提供了一种含有上述陆地棉非特异性磷脂酶C基因GhNPC3a的植物表达载体pCAMBIA1300-GhNPC3a-EPSP。
本发明所述陆地棉非特异性磷脂酶C基因GhNPC3a在提高棉花抗逆性中的应用。
本发明所述植物表达载体pCAMBIA1300-GhNPC3a-EPSP在提高棉花抗逆性中的应用。
其中:所述抗逆性是指抗旱和/或耐低磷。所述棉花品种是春棉、夏棉、常规棉或杂交棉。
在构建GhNPC3a过表达结构时,将GhNPC3a的cDNA核苷酸序列插入到植物表达载体pCAMBIA1300-EPSP中,获得质粒pCAMBIA1300-GhNPC3a-EPSP。酶切鉴定正确后,将质粒导入根瘤农杆菌AGL1或LBA4404中,用于植物遗传转化。
通过农杆菌介导的转基因方法,以棉花的茎尖分生组织细胞为受体将目标基因转入棉花细胞中,即获得转基因植株。通过分子检测和植株抗逆性测定选出目的基因稳定表达且植株抗性明显提高的转基因植株。通过盆栽试验和大田抗逆性测定试验,即可筛选出抗逆性明显提高的转基因育种材料。获得的优异材料有望用于棉花育种或生产实践。
本发明的有益效果是:
本发明首次从陆地棉中克隆出非特异性磷脂酶C基因GhNPC3a。该基因的表达受低磷、干旱和ABA胁迫诱导。同时本发明成功构建了pCAMBIA1300-GhNPC3a-EPSP植物表达载体,将该植物表达载体转化到棉花中证实,本发明所述基因可以显著提高转基因棉花的抗逆特性。预示,GhNPC3a基因应用于培育抗逆转基因农作物具有极大的潜力。
附图说明
图1:质粒pCAMBIA1300-GhNPC3a-EPSP的的结构示意图。
图2:GhNPC3a基因组织特异性分析
其中:TR,主根;LR,侧根;SM,茎;CL,子叶;SL,衰老叶片;EL,幼嫩叶片;ST,茎尖;BR,苞片;PE,花瓣;OV,胚珠。
图3:GhNPC3a基因在非生物胁迫下的表达模式分析。
具体实施方式
以下叙述本发明的实施例。需说明的是,本发明的实施例只对本发明起说明作用而没有限制作用。如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为常规生化试剂。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。
实施例1.高保真PCR扩增GhNPC3a目的基因
取陆地棉中9807四叶期幼嫩的叶片和根,参照植物总RNA提取试剂盒(TIANGEN,北京)说明书,提取植物总RNA,用PrimeScript RT Reagent Kit(TaKaRa,大连)反转录试剂盒反转录得到cDNA。以特异引物进行高保真PCR扩增。特异引物序列如下:
GhNPC3a-F:GGGGTACCATGGCAGTTGAAACAAGCT
GhNPC3a-R:CGAGCTCTCAACGATCACAAACCAAA
PCR反应总体积为25μL,反应体系为:5μL 5×PS Buffer,2μL dNTP,0.5μL正向引物,0.5μL反向引物,0.25μL Prime STARase,1μL稀释十倍的cDNA模板,ddH2O补齐至25μL。
PCR扩增条件,预变性95℃3min,35个循环:98℃,10sec;56℃,5sec;72℃,90sec,最后72℃延伸6min。
将PCR产物经琼脂糖凝胶DNA回收试剂盒回收(TIANGEN,北京)纯化后,连接到pEASY-Blunt Cloning vector(TRANSGEN,北京)上。然后转化大肠杆菌TransT1(TRANSGEN,北京),进行测序分析。通过上述步骤,获得了陆地棉非特异性磷脂酶C基因GhNPC3a的cDNA核苷酸序列。
GhNPC3a基因cDNA核苷酸序列克隆的结果如下:
通过高保真酶扩增获得GhNPC3a基因的cDNA核苷酸序列为1527bp,编码508个氨基酸,GhNPC3a的cDNA核苷酸序列如SEQ ID No.1所示,其编码的GhNPC3a蛋白的氨基酸序列如SEQ ID No.2所示。
实施例2.GhNPC3a基因生物信息学分析
使用ExPASy proteomics server database估算GhNPC3a分子量、等电点等信息;使用Gene Structure Display Server 2.0分析GhNPC3a基因结构;使用MEME分析GhNPC3a的保守motif,并用ExPASy中的ScanProsite工具对获得的motif进行注释;使用NCBI Conserved Domain search和SMART在线分析GhNPC3a的保守结构域,使用Plant CARE分析GhNPC3a的启动子。
经过生物信息学分析,发现GhNPC3a基因,定位在A13染色体上,氨基酸的数目为508,GhNPC3a的理论分子量为56.96kD,理论等电点为5.18。含有4个外显子,3个内含子。MEME分析发现,GhNPC3a有16个motif。对得到的motif进一步去ExPASy中分析,发现有6个motif注释为磷酸酯结构域。用NCBI Conserved Domain search和SMART在线分析GhNPC3a的保守结构域,发现GhNPC3a含有磷酸酯结构域。对GhNPC3a进行启动子顺式作用元件分析,发现GhNPC3a启动子中与胁迫相关的顺式作用元件数目非常多。包括HSE(热胁迫相关),MBS(干旱胁迫相关),LTR(低温胁迫相关),TC-rich repeats(防御与胁迫相关),ARE(厌氧诱导相关),Box-W1(真菌诱导相关),ABRE(脱落酸相关),TGACG-motif和CGTCA-motif(茉莉酮酸甲酯相关),GARE-motif(赤霉素相关),AuxRR-core和TGA-element(生长素相关)和ERE(乙烯相关)。
实施例3.GhNPC3a组织特异性分析
陆地棉(品种为中9807)种子经消毒后种植在温室中,生长条件为:30℃/25℃,相对湿度为60-70%,光照14h,黑暗10h,光子通量密度为800μmol m-2s-1。生长至四叶期时,取主根、侧根、茎、茎尖、子叶、衰老叶片和幼嫩叶片于液氮中,-80℃保存;生长至开花期,取开花第一天时的苞片、花瓣和胚珠于液氮中,-80℃保存。分别提取植物总RNA,并反转录得到cDNA,稀释10倍后用于实时荧光定量PCR分析。陆地棉Histone基因(GenBank accession number NC_006639)为内标。荧光定量PCR反应试剂盒为TAKARA的SYBR Premix Ex TaqⅡ试剂盒,反应在96 System荧光定量仪中进行,进行3次生物学重复,实验结果用相对定量2ΔCt法计算GhNPC3a基因的表达量(Schmittgen and Livak 2008)。引物序列为:
QGhNPC3a-F:GCAGTTGAAACAAGCTCTGCAACT
QGhNPC3a-R:GAGTTGTACGTTCAGCATTTTGTAC
结果:GhNPC3a主要在根中表达(图1)。
实施例4.GhNPC3a基因在非生物胁迫下的表达模式分析
陆地棉(品种为中9807)种子经消毒后种植在温室中,生长条件为:30℃/25℃,相对湿度为60-70%,光照14h,黑暗10h,光子通量密度为800μmol m-2s-1。生长至四叶期时,分别进行低磷(5μM)、盐(200mM NaCl)、干旱(20%PEG6000)的胁迫处理,分别在处理0,1,2,3,6,9和12h时进行取材,取材部位为根部。提取总RNA并反转为cDNA,稀释10倍后用于实时荧光定量PCR分析。陆地棉Histone基因(GenBank accession number NC_006639)为内标。荧光定量PCR反应试剂盒为TAKARA的SYBR Premix Ex TaqⅡ试剂盒,反应在96 System荧光定量仪中进行,进行3次生物学重复,实验结果用相对定量2-ΔΔCt法计算GhNPC3a基因的表达量(Schmittgen and Livak 2008)。引物序列为:
QGhNPC3a-F:GCAGTTGAAACAAGCTCTGCAACT
QGhNPC3a-R:GAGTTGTACGTTCAGCATTTTGTAC
结果表明,低磷胁迫1h时,GhNPC3a表达水平上调,但是处理1h后表达水平下调;盐胁迫下,GhNPC3a表达水平下调;干旱胁迫处理1h和2h时表达水平分别上调5.7和2.9倍,随之表达水平表现出轻微下调的现象。ABA胁迫处理1h时,GhNPC3a表达水平上调3.6倍,2h和3h时上调幅度有所下降,6h之后表达水平与0h相比差异不显著。GhNPC3a基因的表达受低磷、干旱和ABA的胁迫诱导。
实施例5.转GhNPC3a基因创造棉花抗逆材料
一:pCAMBIA1300-GhNPC3a-EPSP植物表达载体的构建
在构建GhNPC3a过表达结构时,将GhNPC3a的cDNA核苷酸序列插入到植物表达载体pCAMBIA1300-EPSP中,获得质粒pCAMBIA1300-GhNPC3a-EPSP。酶切鉴定正确后,将质粒导入根瘤农杆菌AGL1或LBA4404中,用于植物遗传转化。其中植物表达载体pCAMBIA1300-EPSP为全人工合成EPSP基因(两端分别加BamHⅠ、KpnI酶切位点)链接到pCAMBIA1300的多克隆位点上获得。植物表达载体pCAMBIA1300-EPSP中的pCAMBIA1300载体为CAMBIA公司产品,GenBank注册号AF134296;EPSP基因注册号为AY573186.1
二:棉花转基因植株的获得
棉花种子经硫酸脱绒后,用70%乙醇浸泡1min、0.1%的升汞浸泡15min,然后用无菌水洗涤5遍。消毒后种子放入铺有三层滤纸的无菌瓶中,加入适量无菌水,于28℃温箱中萌发。2~3天后下胚轴长到1~2cm,将萌发种子插到MS固体培养基中继续培养,株高7~10cm时用于转化。
将带有载体(质粒pCAMBIA1300-GhNPC3a-EPSP带有除草剂抗性基因EPSP和目的基因GhNPC3a)的根瘤农杆菌(AGL1或LBA4404)在附加抗生素的YEP培养基中28℃下震荡培养,震荡速率为110r/min,使细菌处于对数生长期。然后在3000r/min下离心10分钟,弃上清液。菌体用1/2MS改良液体培养基洗涤,离心收集。再将菌体用添加100mg/L乙酰丁香酮的1/2MS改良液体培养基悬浮,稀释5~20倍用于转化。在转化试验中,加入1%的表面活性剂Silwet L-77(Lehle Seeds,USA)或Tween80。
当种皮脱落或快脱落时,去掉种皮和一片子叶,裸露出小苗茎尖。若苗端已有小叶出现,则用镊子剥去。轻轻划伤顶端分生组织后,将蘸有农杆菌菌液的棉球(由灭菌脱脂棉制备)放到小苗顶端,持续24h后取下棉团,用无菌滤纸吸去感染部位的残留菌液。小苗浸染时置于真空干燥器中,辅以0.5×105Pa的负压处理。浸染过的幼苗于21℃左右暗培养3天,然后在光下培养2~3天后移栽入花盆。花盆下部为土壤,上部为6~8cm厚的蛭石,隔天浇灌1/2MS改良无机盐溶液。
三:转化植株筛选与定植
待转化植株长出3片真叶后,喷洒1.6~1.8‰除草剂草甘膦(转基因选择标记基因为EPSP)水溶液,以未转化植株为对照。喷洒量以植株掉液滴为宜。未转化植株在喷洒后3天停止生长,7~10天左右开始死亡。转化处理后的植株,一些个体的变化与对照植株相似,另一些个体则持续生长,无明显变化。植株长到4~5叶期,将其定植到田间。
四:转基因植株的鉴定和利用
转基因植株开花后自交或姊妹交结实。种子播种在大田或温室,在植株四叶期时取叶片提取DNA,采用PCR技术检测子代植株是否携带外源基因GhNPC3a,并统计外源基因在子代中的分离比例。
结果表明,以无菌苗茎尖分生组织为转基因受体组织,能有效获得转基因植株。并且,在部分转基因株系后代中,外源基因遗传方式符合孟德尔定律,及在传代中稳定。转基因纯合系经田间性状选择和抗性测定,选出保持受体品种基本特性且抗逆性有明显提高的转基因株系,并用于棉花品种改良。
以上详细描述了本发明的较佳具体实施例。应当理解,本领域的普通技术无需创造性劳动就可以根据本发明的构思做出诸多修改和变化。因此,凡本技术领域中技术人员依本发明的构思在现有技术的基础上通过逻辑分析、推理或者有限的实验可以得到的技术方案,皆应在由权利要求书所确定的保护范围内。
序列表
<110>山东大学
<120>一种陆地棉非特异性磷脂酶C基因GhNPC3a及其应用
<141>2016-12-7
<160>2
<210>1
<211>1527
<212>cDNA
<213>棉花(Gossypiumspp)
<221>陆地棉非特异性磷脂酶C基因GhNPC3a的cDNA核苷酸序列
<222>(1)…(1527)
<400>1
atggcagttg aaacaagctc tgcaactcca tctccagtca aaacagtggt tgttttggtt 60
caagagaacc gttcctttga ccacatgtta ggctggttca aaactataaa cccagaaatc 120
gacggtgtca caggctccga atccaacccc atttccacct ccgaccccaa ctccacccaa 180
atcaccttca aggacaccgc cggctacgtt gatcccgacc ccgaccactc tttccaagcc 240
atatacgaac aggtatccgg caaaacgtgg gataccagca acccggatcc gaacccgggg 300
ataaaaatga acggttttgt acaaaatgct gaacgtacaa ctccggggct gtcggagacc 360
gtaatgaatg ggtttaaacc ggaggctgtg ccggtgttta agcagctagt gacggagttc 420
gcagtgtgtg atcggtggtt tgcgtcgttg ccggcgtcga cgcagcctaa caggctttac 480
gtacactcag cgacatcgca tggtgccatg agcaacaaca cacaacagct tatcgaagga 540
tttcctcaaa aaactatatt tgaatcattg gaagagaatg gatatagctt tgggatttat 600
tatcaatctt ttccatctac gcttttttac aggaagctta ggcacttgaa atatgtggac 660
aatttccatc aatacgatct aagcttcaag cgtcactgta aggatgggaa gctaccaaat 720
tatgtggtga ttgagcccag atattttgac attttaacag ctgctgcaaa cgacgaccac 780
ccttcccatg acgtctcaga gggccagaag cttgtgaagg aaatctatga agcgctcaga 840
tcaagtcctc aatggaatga aatcttgttc ctggtcatat atgatgaaca tggtggtttc 900
tatgaccatg ttccgacacc aaccggagtc cctagccccg atgatattgt cggtcctgag 960
ccttataact tcaagtttga tcgtcttggt tgcagggttc ctgccattat ggtttcccct 1020
tggattgagc ctggaacagt gttgcatagg ccatcagggc cagatcctac atcagagttc 1080
gagcattcct ccattgcagc aacacttaag aagattttca atctcaaaga atttctaaca 1140
aagcgtgatg cgtgggctgg ttcctttgat attgttgtca atcgaagcac cccaagaaca 1200
gactgtccag aaaaactggc agagccagtg aaaatgagag acagtgatgc aaaagaaaca 1260
gcaaaactaa gcgattttca agaagagcta gtgcaagcag cagcagcatt gaaaggagat 1320
ccattcaatc ttgtcgaaaa catgacagtt tcatctggtc tcaagtacgt tgaagatgcc 1380
ttcaaaaaat tctatgatga cggccagaaa gctaaggaaa tcaatgaagt tgaagatact 1440
gtttcagctg atgcatcaac taggaggaca acagcttcca aaactttcat gcagaaagtt 1500
ttctcctgtt tggtttgtga tcgttga 1527
<210>2
<211>508
<212>PRT
<213>人工序列
<221>陆地棉非特异性磷脂酶C基因GhNPC3a编码的氨基酸序列
<222>(1)…(508)
<400>2
MAVETSSATP SPVKTVVVLV QENRSFDHML GWFKTINPEI DGVTGSESNP ISTSDPNSTQ 60
ITFKDTAGYV DPDPDHSFQA IYEQVSGKTW DTSNPDPNPG IKMNGFVQNA ERTTPGLSET 120
VMNGFKPEAV PVFKQLVTEF AVCDRWFASL PASTQPNRLY VHSATSHGAM SNNTQQLIEG 180
FPQKTIFESL EENGYSFGIY YQSFPSTLFY RKLRHLKYVD NFHQYDLSFK RHCKDGKLPN 240
YVVIEPRYFD ILTAAANDDH PSHDVSEGQK LVKEIYEALR SSPQWNEILF LVIYDEHGGF 300
YDHVPTPTGV PSPDDIVGPE PYNFKFDRLG CRVPAIMVSP WIEPGTVLHR PSGPDPTSEF 360
EHSSIAATLK KIFNLKEFLT KRDAWAGSFD IVVNRSTPRT DCPEKLAEPV KMRDSDAKET 420
AKLSDFQEEL VQAAAALKGD PFNLVENMTV SSGLKYVEDA FKKFYDDGQK AKEINEVEDT 480
VSADASTRRT TASKTFMQKV FSCLVCDR 508