热激蛋白Hsp17.6CII在调控植物耐盐碱中的应用的制作方法
本发明涉及生物技术领域,尤其涉及热激蛋白hsp17.6cii在调控植物耐盐碱中的应用。
背景技术:
植物受到各种非生物胁迫时,细胞内的活性氧水平升高,通过信号级联系统传递信号,调控基因表达,促进植物对胁迫应答。但是,升高的活性氧需要及时被降解,以免对植物自身造成损伤。过氧化氢酶(cat)通过降解h2o2调控细胞内活性氧的含量,参与到植物对逆境的响应过程中。在多种植物中都发现,cat和盐胁迫有关,盐胁迫处理下,过氧化氢酶的活性升高,例如在水稻中过量表达大肠杆菌的过氧化氢酶kate会提高水稻对盐胁迫的耐受性。同时cat也参与到光胁迫的响应中。在光照强度较高时,过氧化氢酶cat2突变体中光呼吸作用大量产生的h2o2不能及时被清除,细胞内活性氧平衡遭到破坏,使cat2表现出明显的细胞死亡表型,叶片出现卷曲黄化;但提高co2浓度可以将cat2的表型完全恢复,表明过氧化氢酶cat2参与到拟南芥光呼吸过程中产生的h2o2的清除作用。此外,过氧化氢酶cat也参与到植物对干旱的响应过程中。在小麦中,严重的干旱会使过氧化氢酶cat的活性升高,同时,干旱会引起过氧化氢酶cat1和cat2转录下降,并且影响过氧化氢酶cat1和cat2的昼夜表达节律。以上研究结果都表明,cat参与了植物响应各种外界胁迫的过程中,并且逆境胁迫下过氧化氢酶活性的维持也同样十分重要。
小分子热激蛋白shsps是一组分子量为12-43kd,并且在c端含有一个保守的acd结构域的一类蛋白。小分子热激蛋白shsps具有分子伴侣的功能,在逆境胁迫条件下,小分子热激蛋白shsps会形成大量的寡聚物,这些shsps聚合体与目标蛋白结合以防止目标蛋白的损伤,小分子热激蛋白与hsp100/hsp70和hsp60相互作用,保持目标蛋白的三维构象,保持其生物学功能。小分子热激蛋白最初发现被热胁迫诱导,现在已表明它在生物体内广泛分布,多种非生物逆境胁迫都会诱导shsps的表达,shsps的表达水平与植物的多种抗逆性密切相关。
据报道,目前我国存在盐碱化的土地面积几乎与我国可耕地面积相当,每年造成经济损失不可估量,并且盐碱化土地每年还在以很高的速度增加,每年会有大量土地因为耕作不善而退化为不适宜作物生长的盐碱地。如何改良作物,使农作物可以耐受盐碱地并且不影响作物的产量和品质,成为了亟待解决的问题。
技术实现要素:
本发明的一个目的是提供如下1)-3)中任一种物质的用途。
本发明提供了如下1)-3)中任一种物质在如下a-d中至少一种中的应用:
a、调控植物耐盐性和/或耐碱性;
b、调控cat酶的活性;
c、调控cat酶在热胁迫下发生聚集;
d、维持cat酶在热胁迫下的折叠状态;
1)蛋白hsp17.6cii;
2)蛋白hsp17.6cii编码核酸分子;
3)含有蛋白hsp17.6cii编码核酸分子的重组载体、表达盒、转基因细胞系或重组菌。
本发明另一个目的是提供如下1)-3)中任一种物质的另一个用途。
本发明提供了如下1)-3)中任一种物质在培育耐盐性和/或耐碱性提高的植物中的应用:
1)蛋白hsp17.6cii;
2)蛋白hsp17.6cii编码核酸分子;
3)含有蛋白hsp17.6cii编码核酸分子的重组载体、表达盒、转基因细胞系或重组菌。
上述应用中,所述蛋白hsp17.6cii为如下(1)或(2):
(1)由序列表中序列2所示的氨基酸序列组成的蛋白质;
(2)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的由(1)衍生的蛋白质。
上述应用中,所述蛋白hsp17.6cii编码核酸分子是如下1)至3)中任一所述的dna分子:
1)序列表中序列1所示的dna分子;
2)在严格条件下与1)所限定的dna分子杂交且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子;
3)与1)所限定的dna分子至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子。
上述应用中,所述调控植物耐盐性和/或耐碱性为提高植物耐盐性和/或耐碱性;
或所述调控cat酶的活性为提高cat酶的活性,具体为提高cat酶的水解h2o2的活性。
或所述调控cat酶在热胁迫下发生聚集为抑制cat酶在热胁迫下发生聚集。
上述应用中,所述植物为单子叶植物或双子叶植物;
或,所述双子叶植物具体为豆科植物或十字花科植物。
本发明第三个目的是提供一种培育耐盐性和/或耐碱性提高的转基因植物的方法。
本发明提供的方法,为提高目的植物中蛋白hsp17.6cii活性,得到转基因植物;
所述转基因植物的耐盐性高于所述目的植物;
和/或,所述转基因植物的耐碱性高于所述目的植物。
上述方法中,所述提高目的植物中蛋白hsp17.6cii活性为将蛋白hsp17.6cii编码核酸分子导入目的植物中。
上述方法中,所述蛋白hsp17.6cii为如下(1)或(2):
(1)由序列表中序列2所示的氨基酸序列组成的蛋白质;
(2)将序列表中序列2所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的由(1)衍生的蛋白质;
或,所述蛋白hsp17.6cii编码核酸分子是如下1)至3)中任一所述的dna分子:
1)序列表中序列1所示的dna分子;
2)在严格条件下与1)所限定的dna分子杂交且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子;
3)与1)所限定的dna分子至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码由序列表中序列2所示的氨基酸序列组成的蛋白质的dna分子。
上述方法中,所述植物为单子叶植物或双子叶植物;
所述双子叶植物具体为豆科植物或十字花科植物。
本发明的实验证明,将hsp17.6cii基因过表达到野生型拟南芥col-0、突变体nca1-3或cat2cat3中得到的转基因植物,转入col-0的转基因植物对盐碱的耐性高于野生型拟南芥col-0,转入突变体nca1-3或cat2cat3中得到的转基因植物的表型与未转基因前的nca1-3或cat2cat3拟南芥无显著差异,从而证明,hsp17.6cii可以通过调控cat提高植物对盐碱的耐性;另外,热激蛋白hsp17.6cii在提高cat酶的活性且抑制cat酶在热胁迫下发生聚集。
附图说明
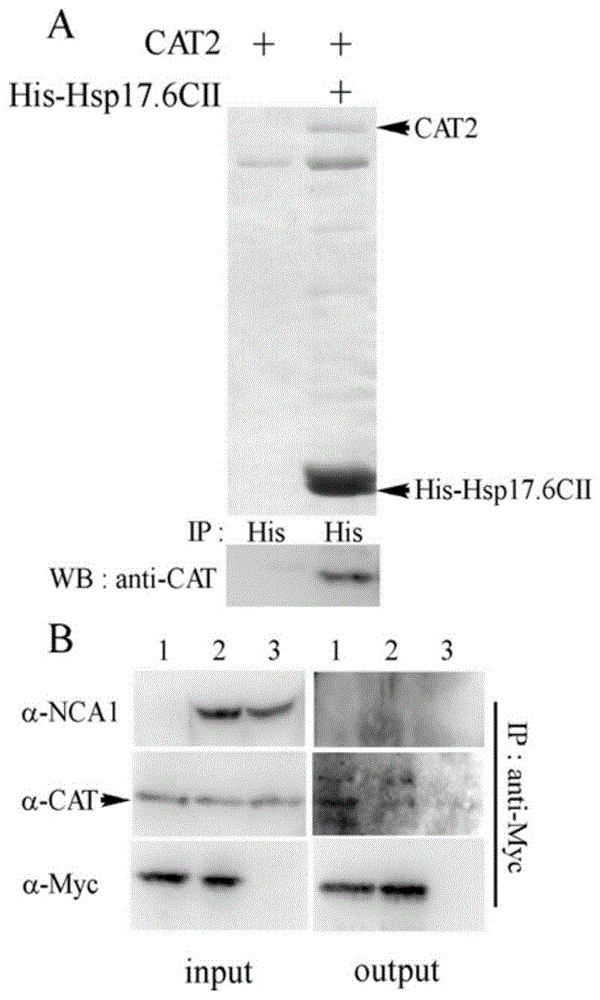
图1为热激蛋白hsp17.6cii与过氧化氢酶cat相互作用分析。
图2为热激蛋白hsp17.6cii保持cat2蛋白的活性和稳定性。
图3为hsp17.6cii基因在植物在盐碱胁迫下的表型分析。
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中,除特殊说明外,定量试验均设5个以上的重复实验。
实施例1、热激蛋白hsp17.6cii在调控过氧化氢酶活性中的应用
一、热激蛋白hsp17.6cii与过氧化氢酶cat相互作用
体外和体内实验验证hsp17.6cii与cat相互作用。
在大肠杆菌原核系统单独表达拟南芥过氧化氢酶cat,或者cat与his-hsp17.6cii共表达,然后用特异结合his标签的填料亲和层析hsp17.6cii蛋白,分别用anti-his和anti-cat2抗体检测hsp17.6cii和cat2蛋白,体外证明hsp17.6cii可以和过氧化氢酶cat相互作用(图1a)。
用免疫共沉淀的方法检测体内hsp17.6cii与cat相互作用。在hsp17.6cii-6myc过表达的拟南芥材料中用特异结合myc标签的beads免疫沉淀hsp17.6cii蛋白,分别用anti-his和anti-cat2抗体检测hsp17.6cii及共沉淀的cat2蛋白,体内证明hsp17.6cii可以和过氧化氢酶cat相互作用(图1b)。
二、热激蛋白hsp17.6cii保持cat2蛋白的活性和稳定性
1、hsp17.6cii可以激活cat2水解h2o2的活性
分别纯化在大肠杆菌原核系统表达的hsp17.6cii(氨基酸序列为序列2)与his-cat2重组蛋白(序列3),检测cat2蛋白降解h2o2的活性,以之前报道的过氧化氢酶cat激活重组蛋白his-nca1(序列4)为正对照。
使用碧云天过氧化氢酶检测试剂盒(产品编号:s0051)检测cat2蛋白降解h2o2的活性,具体方法如下:
配置过氧化氢溶液:
试剂盒中的过氧化氢浓度约为1m,由于过氧化氢易降解,使用前需要将此过氧化氢稀释100倍,测定a240。根据公式计算过氧化氢浓度(mm)=22.94×a240。根据计算出的过氧化氢实际浓度配置浓度为250mm、5mm的过氧化氢溶液备用。
配制显色工作液:
将试剂盒中的显色底物在冰上溶解,分装,按照1:1000的比例加入过氧化物酶,混合均匀得到显色工作液。
样品制备:
将重组蛋白his-nca1,his-cat2和hsp17.6cii蛋白用ni-ntaagarose进行纯化。所有蛋白均换至50mmtris-hcl(ph8.0)缓冲液中,使样品蛋白终浓度定到1μg/μl。
测定过氧化氢标准曲线:
(1)取0、12.5、25、50、70μl配制好的5mm过氧化氢溶液至1.5ml离心管中,分别加入检测缓冲液,至最终体积为100μl,混匀。
(2)将上步配置的过氧化氢溶液各取4μl,加入到96孔板中。再加入200μl显色工作液。25℃至少孵育15-45分钟,测定a520。
样品测定:
(1)取1-10μl样品至1.5ml离心管中,加入检测缓冲液,最终体积为40μl,混匀。再加入10μl250mm过氧化氢溶液,用移液器迅速混匀。室温反应3分钟。
(2)加入450μl反应终止液,颠倒混匀以终止反应。在终止反应后15分钟内完成下面的步骤。
(3)在一个新的离心管内加入40μl检测缓冲液,再加入10μl上一步中的混合液,混匀。
(4)从上一步骤的50μl体系中取10μl加入到96孔板内。加入200μl显色工作液。
(5)25℃至少孵育15分钟后测定a520,但孵育时间不能超过45分钟。
计算:
(1)计算出标准曲线。
(2)根据标准曲线计算出样品中残余的过氧化氢摩尔数。
(3)样品过氧化氢酶酶活力(unit)=反应消耗过氧化氢量(μmol)×250/(3×样品体积×蛋白浓度)。1个酶活力单位(1unit)指在25℃,ph7.0的条件下,在1分钟内可以催化分解1微摩尔过氧化氢。
结果如图2a所示,与his-cat重组蛋白相比,hsp17.6cii可以激活cat2水解h2o2的活性。
2、热激蛋白hsp17.6cii保持cat2蛋白的稳定性
hsp17.6cii可以激活cat2水解h2o2的活性,那么hsp17.6cii是否通过与cat相互作用影响了cat的蛋白折叠状态或热聚集,具体检测方法。
检测方法:将重组蛋白his-nca1(序列4),his-cat2(序列3)和hsp17.6cii蛋白(序列2)均溶解于50mmtris-hcl(ph8.0)缓冲液中,得到不同的蛋白溶液,各个溶液的浓度均为1μmol。
具体分为如下几组:
cat2组:将10μl浓度为1μmol的his-cat2蛋白溶液加在190μl的反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
cat2+bsa组(1:10):将10μl浓度为1μmol的his-cat2蛋白溶液和10μl浓度为10μmol的bsa加在180μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
cat2+nca1(1:1)组:将10μl浓度为1μmol的nca1蛋白溶液和10μl浓度为1μmol的his-cat2蛋白溶液加在180μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
cat2+nca1(1:5)组:将10μl浓度为1μmol的nca1蛋白溶液和50μl浓度为1μmol的his-cat2蛋白溶液加在140μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
cat2+hsp17.6cii组(1:1):将10μl浓度为1μmol的hsp17.6cii蛋白溶液和10μl浓度为1μmol的his-cat2蛋白溶液加在180μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
cat2+hsp17.6cii(1:5)组:将10μl浓度为1μmol的hsp17.6cii蛋白溶液和50μl浓度为1μmol的his-cat2蛋白溶液加在140μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
hsp17.6cii组:将10μl浓度为1μmol的hsp17.6cii蛋白溶液加在190μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值;
nca1组:将10μl浓度为1μmol的nca1蛋白溶液加在190μl反应缓冲液(40mmhepes,ph7.5)中,检测45℃下340nm处的吸收值。
340nm处的吸收值与蛋白聚集成正相关。
吸收值越高,则在热胁迫下的cat的聚集作用越大。
结果如图2b所示,可以看出,当不加hsp17.6cii时,cat聚集最快;当加入hsp17.6cii时,热诱导的cat的聚集作用减弱,并且这种减弱作用有剂量依赖效应,随着hsp17.6cii量的增加而加强;表明,热诱导的cat2的聚集作用能够被hsp17.6cii抑制,并且同样具有剂量依赖效应。这些结果说明hsp17.6cii具有分子伴侣活性,能够与cat2相互作用维持cat2的蛋白折叠状态。
实施例2、hsp17.6cii基因在植物耐盐碱中的应用
一、转hsp17.6cii拟南芥的获得及鉴定
1、重组质粒的制备
人工合成序列表中序列1所示的hsp17.6cii基因,将其插入pcambia1307载体(cambiavzc0323)的bamhi酶切位点间,得到重组质粒pcambia1307-hsp17.6cii,编码蛋白hsp17.6cii(氨基酸序列为序列2)。
2、重组菌的制备
将重组质粒pcambia1307-hsp17.6cii用电击法导入农杆菌gv3101,得到重组菌。
提取重组菌的质粒,经过测序,该质粒为pcambia1307-hsp17.6cii,将含有该质粒的重组菌命名为gv3101/pcambia1307-hsp17.6cii,即为重组农杆菌。
3、转hsp17.6cii拟南芥的获得
将重组农杆菌gv3101/pcambia1307-hsp17.6cii培养至对数期,然后用抽真空法将其转化哥伦比亚生态型拟南芥(col-0)(种子购自arabidopsisbiologicalresourcecenter(abrc))花中,经培育后收获种子,将种子播于含潮霉素(50mg/l)的ms筛选培养基上,待筛选得到的t1代植株长至6叶时移到蛭石上生长,收获t1代单株,各单株种子分别播种,用相同的ms筛选培养基继续筛选以观察t2代的分离情况,如此重复数代直至获得遗传稳定的转基因纯合株系,获得t3代转hsp17.6cii拟南芥col-0。
采用同样的方法将重组农杆菌gv3101/pcambia1307-hsp17.6cii转化突变体nca1-3(nca1-3又名hphd1,突变体种子于2014年09月16日保藏在中国微生物菌种保藏管理委员会普通微生物中心(简称cgmcc,地址:北京市朝阳区北辰西路1号院3号,中国科学院微生物研究所,邮编100101),保藏号cgmccno.9599,分类命名为拟南芥(arabidopsisthaliana)),突变体cat2cat3(cat2(salk_076998)和cat3(salk_092911)单突变体从tair(thearabidopsisinformationresource),即拟南芥信息资源网站订购,突变体号为cat2(salk_076998)和cat3(salk_092911),双突变体cat2cat3是将cat2(salk_076998)和cat3(salk_092911)单突变体进行杂交获得的),将重组农杆菌gv3101/pcambia1307-hsp17.6cii转化突变体cat2cat3突变体的花,得到t3代转hsp17.6cii拟南芥nca1-3和t3代转hsp17.6cii拟南芥cat2cat3。
采用同样的方法将空载体pcambia1307载体转化哥伦比亚生态型拟南芥(col-0),得到t3代转空载体拟南芥。
4、转hsp17.6cii拟南芥的鉴定
分别提取t3代转空载体拟南芥、t3代转hsp17.6cii拟南芥col-0、t3代转hsp17.6cii拟南芥nca1-3和t3代转hsp17.6cii拟南芥cat2cat3的基因组dna作为模板,用hsp17.6cii基因特异引物前端引物5’atggatttaggaaggtttccaataat3’,反端引物5’tcaagcaacttgaacttgaattg3’进行rt-pcr扩增。
以actin基因为内参基因,内参引物为actin前端引物5’gatgcccagaagtcttgttcc3’,反端引物5’accaccgatccagacactgtacttcc3’。
以野生型拟南芥(col-0)、拟南芥nca1-3和拟南芥cat2cat3为对照。
结果,与各自对照相比,t3代转hsp17.6cii拟南芥、t3代转hsp17.6cii拟南芥nca1-3和t3代转hsp17.6cii拟南芥cat2cat3中hsp17.6cii的表达量提高。
t3代转空载体拟南芥与野生型拟南芥(col-0)结果无显著差异。
二、转hsp17.6cii拟南芥的功能研究
将拟南芥nca1-3突变体、拟南芥cat2cat3突变体、野生型拟南芥col-0、t3代转空载体拟南芥、t3代转hsp17.6cii拟南芥col-0(hsp17.6ciioeincol-0)株系5#、16#、t3代转hsp17.6cii拟南芥nca1-3和t3代转hsp17.6cii拟南芥cat2cat3种子点在普通ms培养基上生长7天后分别转移到ph7.3的ms培养基和含有75mmnacl的ms培养基上,光箱中培养12天后观察表型。
ph7.3的ms培养基为将ph值为5.8ms培养基的调节为ph为7.3,得到的培养基。
含有75mmnacl的ms培养基为将nacl和ms培养基混匀,得到培养基,且nacl在培养基中的浓度为75mm,ph值为5.8。
1、表型检测
检测表型,结果如图3a-3f所示,在ph值为7.3的ms培养基和含有75mmnacl的ms培养基上,与野生型拟南芥col-0相比,t3代转hsp17.6cii拟南芥col-0(hsp17.6ciioeincol-0)株系5#、16#的根长增加,表明hsp17.6cii增加植物的抗盐碱能力;
而在nca1-3或cat2cat3突变体中过表达hsp17.6cii不能回补nca1-3或cat2cat3的盐碱敏感表型,表明hsp17.6cii增加植物的抗盐碱能力是通过nca1和cat起作用的。
2、鲜重
称取各个株系的鲜重,结果如图3g-l所示,在ph值为7.3的ms培养基和含有75mmnacl的ms培养基上,与野生型拟南芥col-0相比,t3代转hsp17.6cii拟南芥col-0(hsp17.6ciioeincol-0)株系5#、16#的鲜重增加,表明hsp17.6cii增加植物的抗盐碱能力;
而在nca1-3或cat2cat3突变体中过表达hsp17.6cii不能回补nca1-3或cat2cat3的盐碱敏感表型,表明hsp17.6cii增加植物的抗盐碱能力是通过nca1和cat起作用的。
序列表
<110>中国农业大学
<120>热激蛋白hsp17.6cii在调控植物耐盐碱中的应用
<160>4
<170>patentinversion3.5
<210>1
<211>155
<212>prt
<213>artificialsequence
<400>1
metaspleuglyargpheproileileserileleugluaspmetleu
151015
gluvalprogluasphisasnasnglulysthrargasnasnproser
202530
argvaltyrmetargaspalalysalametalaalathrproalaasp
354045
valilegluhisproasnalatyralaphevalvalaspmetprogly
505560
ilelysglyaspgluilelysvalglnvalgluasnaspasnvalleu
65707580
valvalserglygluargglnarggluasnlysgluasngluglyval
859095
lystyrvalargmetgluargargmetglylysphemetarglysphe
100105110
glnleuprogluasnalaaspleuasplysileseralavalcyshis
115120125
aspglyvalleulysvalthrvalglnlysleuproproproglupro
130135140
lyslysprolysthrileglnvalglnvalala
145150155
<210>2
<211>468
<212>dna
<213>artificialsequence
<400>2
atggatttaggaaggtttccaataatctcaatcctcgaagacatgcttgaagtccccgaa60
gaccacaacaacgagaagacccgcaacaacccttcacgagtttacatgcgagacgctaag120
gcaatggctgctacacctgctgacgtcatcgagcaccctaacgcatatgcattcgtcgtg180
gacatgcctggaatcaaaggagatgagatcaaggttcaggtcgagaacgacaatgtgctt240
gtggtgagtggagagaggcagagagagaacaaagaaaacgaaggtgtgaagtatgtgagg300
atggagagaagaatgggtaagttcatgaggaagtttcagttgcctgaaaatgcagatttg360
gacaagatctctgctgtttgtcatgacggtgtgttgaaggttactgttcagaaacttcct420
cctccggaaccaaagaaaccaaagacaattcaagttcaagttgcttga468
<210>3
<211>492
<212>prt
<213>artificialsequence
<400>3
metaspprotyrlystyrargproalasersertyrasnserprophe
151015
phethrthrasnserglyalaprovaltrpasnasnasnsersermet
202530
thrvalglyproargglyproileleuleugluasptyrhisleuval
354045
glulysleualaasnpheasparggluargileprogluargvalval
505560
hisalaargglyalaseralalysglyphephegluvalthrhisasp
65707580
ileserasnleuthrcysalaasppheleuargalaproglyvalgln
859095
thrprovalilevalargpheserthrvalilehisgluargglyser
100105110
progluthrleuargaspproargglyphealavallysphetyrthr
115120125
arggluglyasnpheaspleuvalglyasnasnpheprovalphephe
130135140
ileargaspglymetlyspheproaspmetvalhisalaleulyspro
145150155160
asnprolysserhisileglngluasntrpargileleuaspphephe
165170175
serhishisprogluserleuasnmetphethrpheleupheaspasp
180185190
ileglyileproglnasptyrarghismetaspglyserglyvalasn
195200205
thrtyrmetleuileasnlysalaglylysalahistyrvallysphe
210215220
histrplysprothrcysglyvallysserleuleuglugluaspala
225230235240
ileargvalglyglythrasnhisserhisalathrglnaspleutyr
245250255
aspserilealaalaglyasntyrproglutrplysleupheilegln
260265270
ileileaspproalaaspgluasplyspheasppheaspproleuasp
275280285
valthrlysthrtrpprogluaspileleuproleuglnprovalgly
290295300
argmetvalleuasnlysasnileaspasnphephealagluasnglu
305310315320
glnleualaphecysproalaileilevalproglyilehistyrser
325330335
aspasplysleuleuglnthrargvalphesertyralaaspthrgln
340345350
arghisargleuglyproasntyrleuglnleuprovalasnalapro
355360365
lyscysalahishisasnasnhishisgluglyphemetasnphemet
370375380
hisargaspglugluvalasntyrpheproserargtyraspglnval
385390395400
arghisalaglulystyrprothrproproalavalcysserglylys
405410415
arggluargcysileileglulysgluasnasnphelysgluprogly
420425430
gluargtyrargthrphethrprogluargglngluargpheilegln
435440445
argtrpileaspalaleuseraspproargilethrhisgluilearg
450455460
seriletrpilesertyrtrpserglnalaasplysserleuglygln
465470475480
lysleualaserargleuasnvalargproserile
485490
<210>4
<211>405
<212>prt
<213>artificialsequence
<400>4
metthrthrthrservalcyspropheserlysalaalaargproasp
151015
aspglyserthrarglysglnglygluilethralaserglycyspro
202530
pheserlysalaalaargproaspaspalaseralaarglysglngly
354045
gluthrthralaserglycyspropheserlysseralaargproasp
505560
gluasnglyserlysglycysprogluglngluglyasnleuasnlys
65707580
aspserthraspseralathrvalproalalyscyspropheglytyr
859095
aspserglnthrphelysleuglyprophesercysmetleucysgln
100105110
alaleuleutyrgluserserargcysvalprocysthrhisvalphe
115120125
cyslysvalcysleuthrargphelysaspcysproleucysglyala
130135140
aspilegluserilegluvalaspgluasnleuglnlysmetvalasp
145150155160
glnpheilegluglyhisalaargilelysargservalvalasngly
165170175
thrglulysglugluvalgluasnaspasnlyslysvaliletyrala
180185190
aspvalsermetgluargglyserpheleuvalglnglnalametarg
195200205
alapheseralaglnasntyrgluseralalysserargleualamet
210215220
cysthrgluaspileargaspglnleuglyarggluglyasnthrpro
225230235240
gluleucysserglnleuglyalavalleuglymetleuglyaspcys
245250255
serargalametglyaspserserseralavallyshisphegluglu
260265270
servalglupheleumetlysleuproleuasnaspleugluilethr
275280285
histhrleuservalserleuasnlysileglyaspleulystyrtyr
290295300
aspgluaspleuglnalaalaargsertyrtyraspargalaleuasn
305310315320
valargargaspalametlyshishisproasnalaproserglnile
325330335
leuaspvalalavalserleualalysvalalaaspileaspargthr
340345350
leuglnasngluvalalaalathraspglyphelysgluglymetarg
355360365
leuleugluserleulysleuaspsergluaspseralaleuglugln
370375380
argargleuservalleuglupheleulyslysglnvalgluthrasp
385390395400
alagluthralaleu
405
- 还没有人留言评论。精彩留言会获得点赞!