对PD1和TIM3特异性的双特异性抗体的制作方法
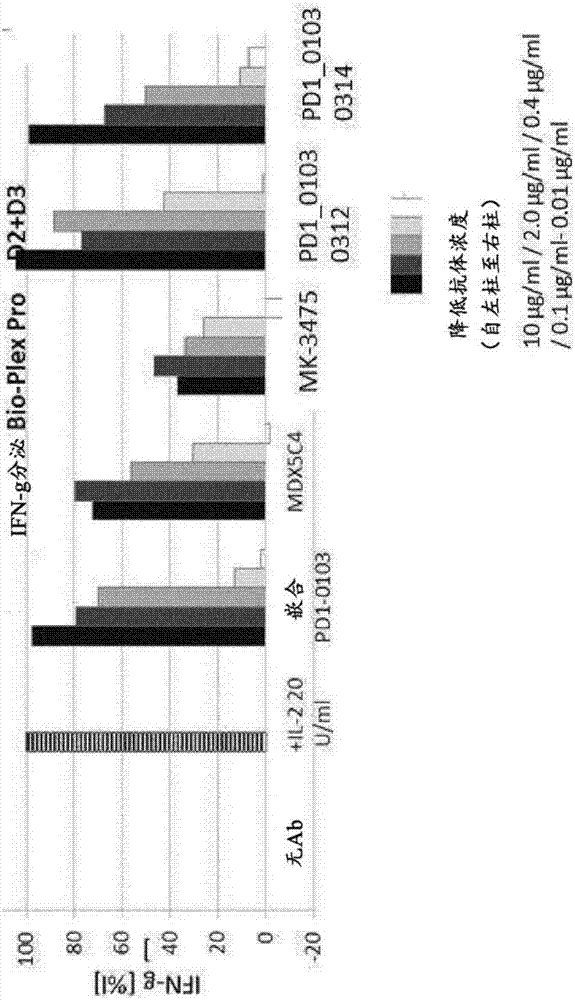
本发明涉及包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,特别是如下的双特异性抗体,其中该双特异性抗体以与对PD1的结合相比要低的结合亲和力结合TIM3。本发明进一步涉及生成这些分子的方法和使用它们的方法。发明背景免疫系统在针对癌症的保护中的重要性基于它检测和破坏异常细胞的能力。然而,一些肿瘤细胞能够通过造成免疫抑制的状态来逃避免疫系统(Zitvogeletal.,NatureReviewsImmunology6(2006),715-727)。携带肿瘤的宿主中存在的免疫抑制的机制的一个例子是促进T细胞功能障碍或耗尽。由于它们选择性识别自所有细胞隔室中的蛋白质衍生的肽的能力;它们直接识别和杀伤抗原表达性细胞(由CD8+效应T细胞;也称作细胞毒性T淋巴细胞(CTL)进行)的能力;和它们协调多种多样免疫应答(由CD4+辅助T细胞进行)(这整合获得性和先天效应器机制)的能力,T细胞已经是治疗性操作内源抗肿瘤免疫力的努力的主要焦点。耗尽的T细胞未能响应抗原刺激而增殖及发挥效应器功能诸如细胞毒性和细胞因子分泌。进一步的研究鉴定出耗尽的T细胞特征在于持续表达抑制性分子PD-1(编程性细胞死亡蛋白1)且阻断PD-1和PD-L1(PD-1配体)相互作用能在受到LCMV感染的小鼠中逆转T细胞耗尽并恢复抗原特异性T细胞应答(Barberetal.,Nature439(2006),682-687)。然而,单独靶向PD-1/PD-L1途径并不总是导致T细胞耗尽的逆转(Gehringetal.,Gastroenterology137(2009),682-690),这指示其它分子有可能涉及T细胞耗尽(Sakuishi,J.ExperimentalMed.207(2010),2187-2194)。TIM-3是最初鉴定为在分泌IFN-γ的Th1和Tc1细胞上选择性表达的一种分子(Monneyetal.,Nature415(2002),536-541)。TIM-3与它的配体,半乳凝集素-9的相互作用在TIM-3+T细胞中触发细胞死亡。如此,TIM-3和PD-1均能作为T细胞应答的负调节物发挥功能。已经显示了TIM-3在实体和血液学恶性二者的临床前模型中标志CD8+T细胞中最受遏制或最具功能障碍的群体(Sakuishi,J.ExperimentalMed.207(2010),2187-2194;Zhou,Blood117(2011),4501-4510;MajetiRetal.,PNAS,106(2009),3396-3401)。在这些模型中,所有CD8+TIM-3+T细胞共表达PD1,而且这些双重表达性细胞展现比单独表达PD1的细胞更大的细胞周期进展和效应细胞因子生成[白介素(IL)-2,TNF,和IFN-γ]二者中的缺陷。如此,TIM-3途径可能与PD-1途径合作来促进癌症中CD8+T细胞中严重功能障碍性表型的发生。因此预期组合靶向TIM-3和PD1途径在控制肿瘤生长方面高度有效。TIM3是一种属于免疫球蛋白超家族和蛋白TIM家族的人蛋白。在人中,与小鼠相似,TIM-3在T细胞以及吞噬性细胞诸如巨噬细胞和树突细胞上表达。TIM3对蛋白质配体(例如半乳凝集素-9)的结合能经由凋亡诱导的机制抑制Th1应答,并因此导致诸如对外周耐受的诱导。siRNA对人TIM3表达的降低或阻断性抗体对人TIM3的抑制提高来自CD4阳性T细胞的干扰素α分泌,支持TIM3在人T细胞中的抑制性作用。在吞噬细胞中,TIM3还作为识别凋亡细胞的受体发挥功能。对来自自身免疫病患者的临床样品的分析显示TIM3在CD4阳性细胞中不表达。特别地,与自正常健康人衍生的克隆相比,在自具有多发性硬化的患者的脑脊液衍生的T细胞克隆中,TIM3的表达水平更低且IFN-γ的分泌水平更高(KoguchiKetal.,JExpMed.203(2006)1413-1418)。有关于TIM-3与变应性疾病或哮喘的关系的报告(WO96/27603和WO2003/063792)。抗TIM3单克隆抗体的例子包括抗人TIM3大鼠单克隆抗体(克隆344823,由R&DSystems制造)和抗人TIM-3小鼠单克隆抗体(克隆F38-2E2,由R&DSystems制造)。WO2013/06490涉及显示快速内在化的抗TIM3抗体及其免疫缀合物,用于治疗癌症和减轻炎症。US2012/189617涉及展现更高效应器活性诸如抗体依赖性细胞的细胞毒性(ADCC活性)的抗TIM-3抗体,用于涉及人TIM3表达性细胞的疾病。编程性细胞死亡蛋白1(PD-1或CD279)是CD28受体家族的一个抑制性成员,该家族还包括CD28,CTLA-4,ICOS和BTLA。PD-1是细胞表面受体且在活化的B细胞,T细胞,和髓样细胞上表达(Okazakietal(2002)Curr.Opin.Immunol.14:391779-82;Bennettetal.(2003)JImmunol170:711-8)。PD-1的结构是一种单体1型跨膜蛋白,由一个免疫球蛋白可变样胞外域和一个含有免疫受体基于酪氨酸的抑制性基序(ITIM)和免疫受体基于酪氨酸的转换基序(ITSM)的胞质域组成。活化的T细胞瞬时表达PD1,但是PD1及其配体PDL1的持续超表达促进免疫耗尽,导致持久的病毒感染,肿瘤逃脱,升高的感染和死亡率。PD1表达是经由T细胞受体通过抗原识别来诱导的且它的表达是主要经由连续的T细胞受体信号传导来维持的。延长的抗原暴露后,PD1座未能再甲基化,这促进连续的超表达。阻断PD1途径能在癌症和慢性病毒感染中恢复耗尽的T细胞功能(Sheridan,NatureBiotechnology30(2012),729-730)。针对PD-1的单克隆抗体已有记载,例如WO2003/042402,WO2004/004771,WO2004/056875,WO2004/072286,WO2004/087196,WO2006/121168,WO2006/133396,WO2007/005874,WO2008/083174,WO2008/156712,WO2009/024531,WO2009/014708,WO2009/101611,WO2009/114335,WO2009/154335,WO2010/027828,WO2010/027423,WO2010/029434,WO2010/029435,WO2010/036959,WO2010/063011,WO2010/089411,WO2011/066342,WO2011/110604,WO2011/110621,WO2012/145493,WO2013/014668,WO2014/179664,和WO2015/112900。还已经显示了阻断PD1和TIM3二者能恢复抗细菌免疫应答,例如在具有急性酒精性肝炎(AAH)的患者中。由于慢性内毒素暴露,来自这些患者的淋巴细胞表达高水平的免疫抑制性受体,生成更低的水平的干扰素γ,且具有升高的IL10生成。这些效应能通过阻断PD1和TIM3来逆转,其提高T细胞和嗜中性细胞的抗微生物活性(Markwicketal,Gastroenterology148(2015),590-602)。WO2011/159877中已经记载了用于慢性免疫状况中的免疫疗法的针对TIM3和PD1的双特异性抗体。然而,需要提供新的双特异性抗体,其不仅同时结合PD1和TIM3并因此选择性靶向表达PD1和TIM3二者的T细胞,而且避免阻断其它细胞诸如先天免疫细胞,例如幼稚树突细胞(DC)和单核细胞上的TIM3。本发明的双特异性抗体不仅有效阻断过表达PD1和TIM3二者的T细胞上的PD1和Tim3,而且它们对于这些细胞具有很高选择性且由此可以避免施用高度有活性的TIM3抗体所致的副作用。发明概述在一个方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含(a)VH域,其包含(i)包含SEQIDNO:1的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:2的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:3的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:4或SEQIDNO:11或SEQIDNO:12的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:5的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:6的氨基酸序列的HVR-L3;或(b)VH域,其包含(i)包含SEQIDNO:17的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:18的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:19的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:20的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:21的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:22的氨基酸序列的HVR-L3;或(c)VH域,其包含(i)包含SEQIDNO:29的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:30的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:31的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:32的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:33的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:34的氨基酸序列的HVR-L3。在一个方面,该包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体是二价的。在另一个方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体以低亲和力结合TIM3且以高亲和力结合PD1。在一个特定的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体以与对PD1的结合相比要低至少50倍的结合亲和力,更特别的是以与对PD1的结合相比要低至少100倍的结合亲和力结合TIM3。在一个优选的实施方案中,该结合亲和力(KD)是用表面等离振子共振测定法(如例如实施例12中描述的)测定的。在又一个方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含(a)包含SEQIDNO:43的氨基酸序列的VH域和包含SEQIDNO:44的氨基酸序列的VL域,或(b)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,或(c)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:47的氨基酸序列的VL域,或(d)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:48的氨基酸序列的VL域,或(e)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:49的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含(a)包含SEQIDNO:7的氨基酸序列的VH域和包含SEQIDNO:8的氨基酸序列的VL域,或(b)包含SEQIDNO:9的氨基酸序列的VH域和包含SEQIDNO:10的氨基酸序列的VL域,或(c)包含SEQIDNO:13的氨基酸序列的VH域和包含SEQIDNO:14的氨基酸序列的VL域,或(d)包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域,或(e)包含SEQIDNO:23的氨基酸序列的VH域和包含SEQIDNO:24的氨基酸序列的VL域,或(f)包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域,或(g)包含SEQIDNO:27的氨基酸序列的VH域和包含SEQIDNO:28的氨基酸序列的VL域,或(h)包含SEQIDNO:35的氨基酸序列的VH域和包含SEQIDNO:36的氨基酸序列的VL域。在一个特定的方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域或包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。特别地,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含VH域,其包含(i)包含SEQIDNO:17的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:18的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:19的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:20的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:21的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:22的氨基酸序列的HVR-L3。更加特别地,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。在一个特定的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体以与对PD1的结合相比要低至少50倍的结合亲和力,更特别的是以与对PD1的结合相比要低至少100倍的结合亲和力结合TIM3。在又一个方面,该包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体是人,人源化或嵌合抗体。特别地,它是人源化或嵌合抗体。在另一个方面,本发明涉及一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体包含Fc域,包含该特异性结合PD1的抗原结合位点的第一Fab片段和包含该特异性结合TIM3的抗原结合位点的第二Fab片段。特别地,该Fc域是IgG域,更加特别是IgG1Fc域或IgG4Fc域。在一个方面,本发明涉及一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该Fc域包含一处或多处降低对Fc受体,特别是Fcγ受体的结合的氨基酸替代。特别地,该Fc域是人IgG1亚类的,具有氨基酸突变L234A,L235A和P329G(编号方式依照KabatEU索引)。在另一个方面,本发明涉及一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该Fc域包含促进Fc域的第一和第二亚基联合的修饰。在一个方面,该本发明涉及一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中依照节入穴方法,该Fc域的第一亚基包含节且该Fc域的第二亚基包含穴。在一个特定的方面,该Fc域的第一亚基包含氨基酸替代S354C和T366W(EU编号方式)且该Fc域的第二亚基包含氨基酸替代Y349C,T366S和Y407V(编号方式依照KabatEU索引)。在一个另外的方面,本发明涉及一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中在该Fab片段之一中可变域VL和VH彼此替换,使得VH域是轻链的一部分且VL域是重链的一部分。在一个特定的方面,该双特异性抗体是如下的,其中在包含该特异性结合PD1的抗原结合位点的第一Fab片段中,可变域VL和VH彼此替换。在又一个方面,本发明关注一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中在该Fab片段之一中在恒定域CL中位置124处的氨基酸被赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照KabatEU索引),且在恒定域CH1中位置147和213处的氨基酸被谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。在一个特定的方面,该双特异性抗体是如下的,其中在包含该特异性结合TIM3的抗原结合位点的第二Fab片段中在恒定域CL中位置124处的氨基酸被赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照KabatEU索引),且在恒定域CH1中位置147和213处的氨基酸被谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。在另一个方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含(a)包含与SEQIDNO:50的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:52的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:51的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:53的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(b)包含与SEQIDNO:54的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:56的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:55的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:57的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(c)包含与SEQIDNO:58的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:60的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:59的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:61的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(d)包含与SEQIDNO:62的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:64的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:63的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:65的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(e)包含与SEQIDNO:66的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:68的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:67的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:69的序列有至少95%序列同一性的氨基酸序列的第二轻链。在一个特定的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含(a)包含SEQIDNO:50的氨基酸序列的第一重链,包含SEQIDNO:52的氨基酸序列的第一轻链,包含SEQIDNO:51的氨基酸序列的第二重链,和包含SEQIDNO:53的氨基酸序列的第二轻链,或(b)包含SEQIDNO:54的氨基酸序列的第一重链,包含SEQIDNO:56的氨基酸序列的第一轻链,包含SEQIDNO:55的氨基酸序列的第二重链,和包含SEQIDNO:57的氨基酸序列的第二轻链,或(c)包含SEQIDNO:58的氨基酸序列的第一重链,包含SEQIDNO:60的氨基酸序列的第一轻链,包含SEQIDNO:59的氨基酸序列的第二重链,和包含SEQIDNO:61的氨基酸序列的第二轻链,或(d)包含SEQIDNO:62的氨基酸序列的第一重链,包含SEQIDNO:64的氨基酸序列的第一轻链,包含SEQIDNO:63的氨基酸序列的第二重链,和包含SEQIDNO:65的氨基酸序列的第二轻链,或(e)包含SEQIDNO:66的氨基酸序列的第一重链,包含SEQIDNO:68的氨基酸序列的第一轻链,包含SEQIDNO:67的氨基酸序列的第二重链,和包含SEQIDNO:69的氨基酸序列的第二轻链。依照本发明的另一个方面,提供编码如之前本文中描述的双特异性抗体的多核苷酸。本发明进一步提供包含本发明的多核苷酸的载体,特别是表达载体和包含本发明的多核苷酸或载体的原核或真核宿主细胞。在一些实施方案中,该宿主细胞是真核细胞,特别是哺乳动物细胞。在另一个方面,提供的是一种用于生成如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体的方法,其包括下述步骤:a)用包含编码所述双特异性抗体的多核苷酸的载体转化宿主细胞,b)在适合于该双特异性抗体表达的条件下培养该宿主细胞,并c)自培养物回收该双特异性抗体。本发明还涵盖通过本发明的方法生成的双特异性抗体。本发明进一步提供一种药物组合物,其包含如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,和至少一种药学可接受赋形剂。本发明还涵盖的是如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用作药物。在另一个方面,本发明提供如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用于i)调控免疫应答,诸如恢复T细胞活性,ii)刺激免疫应答或功能,iii)治疗感染,iv)治疗癌症,v)延迟癌症进展,vi)延长罹患癌症的患者的存活。在一个方面,提供的是如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用于在有所需要的个体中治疗疾病。在一个具体的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用于治疗癌症。在又一个具体的方面,提供了一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用于调控免疫应答。在另一个方面,提供了一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用于治疗慢性病毒感染。还提供的是如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体用于制造用于在有所需要的个体中治疗疾病的药物,特别是用于制造用于治疗癌症的药物的用途,以及一种在个体中治疗疾病的方法,其包括对所述个体施用治疗有效量的组合物,其包含药学可接受形式的如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体。在一个具体的方面,该疾病是癌症。在另一个具体的方面,该疾病是慢性病毒感染。在另一个方面,提供了一种在个体中调控免疫应答的方法,其包括对所述个体施用治疗有效量的组合物,其包含药学可接受形式的如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体。在任何上述方面,该个体优选是哺乳动物,特别是人。本发明还提供如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,或包含该双特异性抗体的药物组合物,其用于预防或治疗癌症,其中与化疗剂,放射和/或其它用于癌症免疫疗法的药剂组合施用该双特异性抗体。而且,提供的是一种在个体中抑制肿瘤细胞生长的方法,其包括对该个体施用有效量的如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体以抑制肿瘤细胞生长。该个体优选是哺乳动物,特别是人。附图简述图1:用嵌合PD1-0103阻断PD1强烈增强异基因的经刺激的原代人T细胞的IFNγ分泌。图2:用嵌合PD1-0103阻断PD1强烈提高异基因的经刺激的原代人T细胞的干扰素γ(IFN-γ)分泌。图3:用嵌合PD1-0103阻断PD1强烈提高异基因的经刺激的原代人T细胞的肿瘤坏死因子α(TNF)分泌。图4:4A)生成粒酶B的CD4T细胞的频率;4B)在渐增浓度的不同抗PD-1抗体存在下MLR上清液中通过吸光度(光密度,O.D.)检测的IFN-γ的量。图5:5A)在不同抗PD-1抗体存在下PD1/PD-L1阻断对受到遏制的T细胞受体信号传导的再活化的影响;5B)在不同抗PD-1抗体存在下PD1/PD-L1阻断对受到遏制的T细胞受体信号传导的再活化的影响。图6:抗PD1/Tim3双特异性抗体对重组细胞的同时结合的FRET测定法的示意图图7:处理后FRET的诱导/表达PD1和TIM3的细胞上不同双特异性PD1TIM3抗体的结合:将用PD1SNAPTim3CLIP双重转染的HEK293细胞在Tag-Lite缓冲液(Cisbio)中用100nMSNAP-Lumi4-Tb(Cisbio)和100nMClip-Red(Cisbio)于37℃染色1小时。清洗后,将标记细胞与所示双特异性抗PD1/Tim3抗体[0-10nM](图7B中显示的人源化双特异性变体)一起于4℃温育1小时,之后用BMGPherastar读数仪于665/620nm测量时间解析荧光(所描绘的是FRET信号的均值+/-SD[比率665/620nm*10,000],n=3)。7A:1+1格式(抗体PD1TIM3_0389和PD1TIM3_0168)与2+2构建物(PD1TIM3_0358+PD1TIM3_0359)比较,7B:人源化双特异性变体PD1TIM3_0476和PD1TIM3_0477。图8:抗PD1/TIM3双特异性抗体1+1PD1TIM3-0168的同时结合的FRET测定法:用100nMSNAP-Lumi4-Tb和100nMClip-Red标记带SNAP标签的PD1和带CLIP标签的TIM3细胞(如之前描述的)。清洗后,将标记细胞与双特异性抗PD1/TIM3抗体#0168[所示浓度]一起于4℃温育1小时,之后用BMGPherastar读数仪于665/620nm测量时间解析荧光(黑色线)。为了突显双特异性抗体诱导的FRET信号的特异性,添加抗PD1单克隆抗体(#0165;见图8A)或抗TIM3阻断性抗体(#0018,见图8B)任一进行竞争,导致几乎完全阻止FRET信号(灰色曲线)。单独的抗PD1抗体处理并不导致FRET诱导(点线,仅左边的图)。图9A:双特异性1+1PD1TIM3-0389显示与嵌合TIM3_0028(chi0028)和人源化TIM3-0438(0438)相比相同的对阳性CD4+T细胞(PD1+,TIM3+)的结合比,但是更少的对单核细胞,NK细胞和CD3+T细胞的结合。图9B:双特异性1+1PD1TIM3-0389显示与嵌合TIM3_0028(chi0028)和人源化Tim3-0438(0438)相比显著升高的对阳性CD4+T细胞(PD1+,TIM3+)的结合的MFI。图9C:双特异性1+1PD1TIM3-0168显示与嵌合TIM3_0018(Tim3-chi0018)和人源化TIM3-0434(0434)相比对阳性CD4+T细胞(PD1+,TIM3+)的结合没有差异。图9D:双特异性1+1PD1TIM3-0168显示仅仅略微升高的对阳性CD4+T细胞(PD1+,Tim3+)的结合的MFI。图9E和图9F:抗TIM3抗体TIM3-0038显示对单核细胞和CD4+T细胞二者的结合。图9G和图9H:双特异性1+1PD1TIM3-0166(基于嵌合PD1-0103/Tim3-0038)显示强烈降低的对单核细胞的结合(与亲本抗TIM3抗体TIM3_0038相比,见图4E和4F)同时保留强烈的对CD4+T细胞的结合。图10A至10D:双特异性1+1PD1TIM3-0166(基于嵌合PD1-0103/TIM3-0038)显示与双特异性2+2PD1TIM3-0321(也基于嵌合PD1-0103/TIM3-0038,但是具有两个针对PD的抗原结合位点和两个针对TIM3的)相比及与亲本TIM3-0038抗体相比降低的在活化的CD4+T细胞上的和在活化的NK细胞上的内在化图11A:随时间的分析显示与TIM3抗体的胞内聚簇相比更高的双特异性和PD1抗体二者的膜定位。图中的抗体名称:TIM3(chi18-A647=用AlexaA647标记的嵌合TIM3_0018),a-TIM3(chi28-A647=用AlexaA647标记的嵌合TIM3_0028),Bispec(0168-A647=用AlexaA647标记的1+1PD1TIM3_0168(基于嵌合PD1-0103/TIM3-0018)),Bispec(0389-A647=用Alexa647标记的1+1PD1TIM3_0389(基于嵌合PD1-0103/TIM3-0028))和a-PD1(0165-A488=用Alexa488标记的嵌合PD1-0103)。抗PD1和双特异性1+1PD1TIM3_0389(Bispec0389)只显示很慢的内在化,甚至在3小时后,而另一种双特异性1+1PD1TIM3_0168(Bispec0168)的内在化更强。aTIM3Ab0028显示更强的内在化,aTIM3-0018显示最内在化。图11B:嵌合PD1-0103(aPD1-0165)只显示较差的内在化,而高亲和力嵌合TIM3_0018(aTim3-chi18)在TIM3结合后强烈内在化,甚至在15分钟后。低亲和力结合物嵌合TIM3_0028(aTIM3-chi28)的内在化略微降低。双特异性1+1AB0168(由高亲和力结合物aPD1-0165和高亲和力aTIM3-0018构成)显示降低更多的内在化。双特异性1+1AB0389(由高亲和力结合物嵌合PD1-0103(aPD1-0165)和低亲和力嵌合TIM3_0028(aTIM3-0028)构成)显示降低很强的内在化。这可能是由于对PD1和TIM3的二价结合,其中对PD1的高亲和力结合将抗体保留在细胞表面。图12A:PD1-TIM3双特异性抗体1+1PD1TIM3_0168(基于嵌合PD1-0103/TIM3-0018(=AB0168)的功效,与嵌合PD1-0103(=PD1-0165)和嵌合TIM3_0018(=TIM3-chi18)及其组合比较。图12B:PD1-TIM3双特异性抗体1+1PD1TIM3_0389(基于嵌合PD1-0103/TIM3-0028(=BispecAB0389)的功效,与嵌合PD1-0103(=PD1-0165)和嵌合TIM3_0028(=TIM3-chi28)及其组合比较。图12C:PD1-TIM3双特异性抗体1+1PD1-0103/Ky8213(与实施例1中关于1+1CrossMab的描述类似生成的基于嵌合PD1-0103/来自US20120189617的抗TIM3Ky8213(见抗体8213,例如实施例33))的功效,与嵌合PD1-0103(=PD1-0165)和抗TIM3-Ky8213(来自US20120189617(见抗体8213,例如实施例33))及其组合比较。图12D:PD1-TIM3双特异性抗体1+1PD1TIM3_0389(基于嵌合PD1-0103/TIM3-0028(=BispecAB0389(1+1)))的功效,与PD1-TIM3双特异性抗体2+2PD1TIM3_0358(基于嵌合PD1-0103/TIM3-0028(=BispecAB0358(2+2))),和嵌合PD1-0103(=PD1-0165)和嵌合TIM3_0028(=TIIM3-chi28)及其组合比较。图13:与单独用PD1或TIM3抗体处理相比和甚至与用亲本抗体PD1_0376和抗体TIM3_0438的组合处理相比,PD1-TIM3双特异性抗体1+1PD1TIM3_0476处理显著提高CD4T细胞释放IFN-γ的能力。将CD4T细胞与表达MHCII的肿瘤细胞系一起共培养。针对PD1抗体aPD1_0376,MDX-1106(nivolumab)和MK-3475(pembrolizumab),针对TIM3抗体aTIM3_0438和Kyowa-8213(如WO2011/155697中公开的)及针对抗PD1抗体aPD1-0376和抗TIM3抗体aTIM3_0438的组合测试PD1-Tim3双特异性抗体1+1PD1TIM3_0476。图14A和14B:在用MKN45细胞攻击并提供有来自健康人供体的PBMC的受到免疫遏制的雌性小鼠(NOG)中比较PD1-TIM3双特异性抗体1+1(0476)与单独的PD1或TIM3抗体的功效实验的结果在图14A和14B中显示。该图呈现30天时段里肿瘤尺寸的测量均值(处理组内的),包括肿瘤尺寸均值的标准误差。具有实心圆圈的曲线对应于没有处理的肿瘤尺寸生长(媒介)。在图14A中显示更低剂量处理时的肿瘤生长(1.5mg/kg抗体PD1_0376,1.5mg/kgnivolumab,1.5mg/kg抗体TIM3_0438或3mg/kg双特异性抗体1+1PD1TIM3_0476),在图14B中显示更高剂量时的肿瘤生长(5mg/kg抗体PD1_0376,5mg/kgnivolumab,5mg/kg抗体Tim3_0438或10mg/kg双特异性抗体1+1PD1TIM3_0476)。发明详述定义除非另外定义,本文中使用的技术术语和科学术语具有与本发明所属领域中通常使用的相同的含义。出于解释本说明书的目的,以下定义将适用,并且在适当时,以单数形式使用的术语也将包括复数形式,反之亦然。如本文中使用的,术语“抗原结合分子”在其最广义上指的是特异性结合抗原决定簇的分子。抗原结合分子的实例是抗体,抗体片段和支架抗原结合蛋白。在本文中以最广义的方式使用术语“抗体”,其涵盖各种抗体结构,包括但不限于单克隆抗体,多克隆抗体,单特异性和多特异性抗体(例如,双特异性抗体)和抗体片段,只要它们表现出期望的抗原结合活性即可。如本文中使用的术语“单克隆抗体”是指从基本上同质的抗体的群获得的抗体,即包含该群的单独的抗体是同一的和/或结合同一表位,除了可能的变体抗体之外,例如,含有天然存在的突变的或在单克隆抗体制剂的产生过程中产生的抗体,这样变体通常以少量存在。与通常包括针对不同决定簇(表位)的不同抗体的多克隆抗体制剂相反,单克隆抗体制剂的每种单克隆抗体针对抗原上的单一决定簇。如本文中使用的术语“单特异性”抗体表示具有一个或多个结合位点的抗体,所述位点各自与相同抗原的相同表位结合。术语“双特异性”意指抗体能够特异性地与至少两个不同的抗原决定簇结合,例如,各自由结合不同抗原或相同抗原上的不同表位的一对抗体重链可变域(VH)和抗体轻链可变域(VL)形成的两个结合位点。此类双特异性抗体是1+1格式。其它双特异性抗体格式是2+1格式(包含两个针对第一抗原或表位的结合位点和一个针对第二抗原或表位的结合位点)或2+2格式(包含两个针对第一抗原或表位的结合位点和两个针对第二抗原或表位的结合位点)。通常,双特异性抗体包含两个抗原结合位点,每个抗原结合位点对于不同的抗原决定簇是特异性的。在本申请中使用的术语“价”意味着在抗原结合分子中特定数目的结合位点的存在。正因为因此,术语“二价”,“四价”和“六价”分别意味着在抗原结合分子中分别存在两个结合位点,四个结合位点和六个结合位点。依照本发明的双特异性抗体至少是“二价”的且可以是“三价”的或“多价”的(例如“四价”的或“六价”的)。在一个特定的方面,本发明的抗体具有两个或更多个结合位点且是双特异性的。就是说,抗体可以是双特异性,即使在有超过两个结合位点(即抗体四价或多价的)的情况中。特别地,本发明涉及具有针对它们特异性结合的每种抗原的一个结合位点的双特异性二价抗体。术语“全长抗体”,“完整抗体”和“全抗体”在本文中可互换使用,指的是具有与天然抗体结构基本相似的结构的抗体。“天然抗体”指的是具有不同结构的天然存在的免疫球蛋白分子。例如,天然IgG类抗体是约150,000道尔顿的异四聚体糖蛋白,由二硫键结合的两条轻链和两条重链组成。从N末端至C末端,每个重链具有可变区(VH),也称为可变重域或重链可变域,其后是三个恒定域(CH1,CH2,和CH3),也称为重链恒定区。类似地,从N末端至C末端,每个轻链具有可变区(VL),也称为可变轻域或轻链可变域,其后是轻链恒定域(CL),也称为轻链恒定区。抗体重链可被指定为五种类型中的一种,称为α(IgA),δ(IgD),ε(IgE),γ(IgG),或μ(IgM),其中一些可以进一步分为亚类,例如γ1(IgG1),γ2(IgG2),γ3(IgG3),γ4(IgG4),α1(IgA1)和α2(IgA2)。基于其恒定域的氨基酸序列,抗体轻链可被指定为两种类型之一,称为卡帕(κ)和兰布达(λ)。“抗体片段”指的是除了完整抗体之外的分子,其包含与完整抗体所结合的抗原结合的所述完整抗体的一部分。抗体片段的实例包括但不限于Fv,Fab,Fab',Fab’-SH,F(ab')2;双抗体,三抗体,四抗体,交叉Fab片段;线性抗体;单链抗体分子(例如scFv);自抗体片段形成的多特异性抗体和单域抗体。关于某些抗体片段的综述,参见Hudson等人,NatMed9,129-134(2003)。关于scFv片段的综述,参见例如Plückthun,引自ThePharmacologyofMonoclonalAntibodies,vol.113,RosenburgandMooreeds.,Springer-Verlag,NewYork,pp.269-315(1994);还参见WO93/16185;和美国专利Nos.5,571,894和5,587,458。关于包含补救受体结合表位残基并具有增加的体内半衰期的Fab和F(ab')2片段的讨论,参见美国专利No.5,869,046。双抗体是具有两个抗原结合位点的可以是二价或双特异性的抗体片段,参见,例如EP404,097;WO1993/01161;Hudson等人,NatMed9,129-134(2003);和Hollinger等人,ProcNatlAcadSciUSA90,6444-6448(1993)。Hudson等人在NatMed9,129-134(2003)中也描述了三抗体和四抗体。单域抗体是包含抗体的全部或部分重链可变域或全部或部分轻链可变域的抗体片段。在某些实施方案中,单域抗体是人单域抗体(Domantis,Inc.,Waltham,MA;参见例如美国专利No.6,248,516B1)。另外,抗体片段包含具有VH域特征或VL域特征的单链多肽,所述VH域特征即能够与VL域一起组装成功能性抗原结合位点,所述VL域特征即能够与VH域一起组装成功能性抗原结合位点,从而提供全长抗体的抗原结合特性。可以通过各种技术制造抗体片段,包括但不限于完整抗体的蛋白水解消化以及通过重组宿主细胞(例如大肠杆菌或噬菌体)产生,如本文中所述。完整抗体的木瓜蛋白酶消化产生两个完全相同的抗原结合片段,称为“Fab”片段,其各自含有重链和轻链可变域以及轻链的恒定域和重链的第一恒定域(CH1)。因此,如本文中使用的,术语“Fab片段”是指包含包括轻链的VL域和恒定域(CL)的轻链片段,和重链的VH域和第一恒定域(CH1)的抗体片段。Fab'片段因在重链CHl域的羧基末端增加了少数残基而与Fab片段有所不同,包括来自抗体铰链区的一个或多个半胱氨酸。Fab'-SH是其中恒定域的半胱氨酸残基具有游离巯基的Fab'片段。胃蛋白酶处理产生具有两个抗原结合位点(两个Fab片段)和Fc区的一部分的F(ab')2片段。术语“交叉Fab片段”或“xFab片段”或“交换Fab片段”是指其中重链和轻链的可变区或恒定区之间被交换的Fab片段。交换Fab分子的两种不同链组成是可能的并且包含在本发明的双特异性抗体中:一方面,Fab重链和轻链的可变区被交换,即交换Fab分子包含由轻链可变区(VL)和重链恒定区(CH1)组成的肽链,以及由重链可变区(VH)和轻链恒定区(CL)组成的肽链。这种交换Fab分子也被称为CrossFab(VLVH)。另一方面,当Fab重链和轻链的恒定区被交换时,交换Fab分子包含由重链可变区(VH)和轻链恒定区(CL)组成的肽链,以及由轻链可变区(VL)和重链恒定区(CH1)组成的肽链。这种交换Fab分子也被称为CrossFab(CLCH1)。“单链Fab片段”或“scFab”是由抗体重链可变域(VH),抗体恒定域1(CH1),抗体轻链可变域(VL),抗体轻链恒定域(CL)和接头组成的多肽,其中所述抗体域和所述接头在N末端至C末端方向具有以下顺序之一:a)VH-CH1-接头-VL-CL,b)VL-CL-接头-VH-CH1,c)VH-CL-接头-VL-CH1或d)VL-CH1-接头-VH-CL;并且其中所述接头是具有至少30个氨基酸,优选32至50个氨基酸之间的多肽。所述单链Fab片段经由CL域和CH1域之间的天然二硫键而被稳定化。另外,通过插入半胱氨酸残基(例如根据Kabat编号在可变重链中的位置44和可变轻链中的位置100)产生链间二硫键,这些单链Fab分子可以进一步被稳定化。“交换单链Fab片段”或“x-scFab”是由抗体重链可变域(VH),抗体恒定域1(CH1),抗体轻链可变域(VL),抗体轻链恒定域(CL)和接头组成的多肽,其中所述抗体域和所述接头在N末端至C末端方向具有以下顺序之一:a)VH-CL-接头-VL-CH1和b)VL-CH1-接头-VH-CL;其中VH和VL一起形成与抗原特异性结合的抗原结合位点,并且其中所述接头是具有至少30个氨基酸的多肽。另外,通过插入半胱氨酸残基(例如根据Kabat编号在可变重链中的位置44和可变轻链中的位置100)产生链间二硫键,这些x-scFab分子可以进一步被稳定化。“单链可变片段(scFv)”是用具有10个至约25个氨基酸的短接头肽连接的抗体的重链可变区(VH)和轻链可变区(VL)的融合蛋白。该接头通常富含有关柔性的甘氨酸,以及有关溶解性的丝氨酸或苏氨酸,并且可将VH的N末端与VL的C末端连接,或反之亦然。尽管去除了恒定区并引入了接头,但该蛋白质保留了原始抗体的特异性。scFv抗体例如描述于Houston,J.S.,MethodsinEnzymol.203(1991)46-96)。另外,抗体片段包含具有VH域特征或VL域特征的单链多肽,所述VH域特征即能够与VL域一起组装到功能性抗原结合位点上,所述VL域特征即能够与VH域一起组装到功能性抗原结合位点上,从而提供全长抗体的抗原结合特性。“支架抗原结合蛋白”在本领域中是已知的,例如,纤连蛋白和设计的锚蛋白重复蛋白(DARPin)已被用作抗原结合域的替代支架,参见例如Gebauer和Skerra,Engineeredproteinscaffoldsasnext-generationantibodytherapeutics.CurrOpinChemBiol13:245-255(2009)和Stumpp等人,Darpins:Anewgenerationofproteintherapeutics.DrugDiscoveryToday13:695-701(2008)。在本发明的一个方面,支架抗原结合蛋白选自CTLA-4(Evibody),脂质运载蛋白(Anticalin),蛋白A衍生的分子,如蛋白A的Z域(Affibody),A域(Avimer/Maxibody),血清转铁蛋白(trans-body);设计的锚蛋白重复蛋白(DARPin),抗体轻链或重链的可变域(单域抗体,sdAb),抗体重链的可变域(纳米抗体,aVH),VNAR片段,纤连蛋白(AdNectin),C型凝集素域(Tetranectin);新的抗原受体β-内酰胺酶的可变域(VNAR片段),人γ-晶体蛋白或泛素(Affilin分子);人蛋白酶抑制剂的kunitz型域,诸如来自knottin家族的蛋白质的微体,肽适体和纤连蛋白(adnectin)。CTLA-4(细胞毒性T淋巴细胞相关抗原4)是主要在CD4+T细胞上表达的CD28家族受体。其细胞外域具有类似可变域的Ig折叠。可用异源序列取代对应于抗体的CDR的环,以赋予不同的结合特性。经过工程化而具有不同结合特异性的CTLA-4分子也被称为Evibodies(例如,US7166697B1)。Evibodies的大小与抗体(例如,域抗体)的分离可变区大约相同。进一步的详情参见JournalofImmunologicalMethods248(1-2),31-45(2001)。脂质运载蛋白为细胞外蛋白质家族,其转运小的疏水性分子,如类固醇,胆红素,类视黄醇和脂质。它们具有刚性β-片层二级结构,在锥形结构的开口端具有可被工程化以结合不同的靶抗原的多个环。Anticalin的大小在160-180个氨基酸之间,并且衍生自脂质运载蛋白。进一步的详情参见BiochimBiophysActa1482:337-350(2000),US7250297B1和US20070224633。亲和体是衍生自金黄色葡萄球菌蛋白A的支架,其可被工程化以结合抗原。该域由具有约58个氨基酸的三螺旋束组成。通过表面残基的随机化生成了文库。进一步的详情参见ProteinEng.Des.Sel.2004,17,455-462和EP1641818A1。Avimers是衍生自A域支架家族的多域蛋白。具有约35个氨基酸的天然域采用定义的二硫键结构。多样性是通过改变A域家族所显示的自然变异产生的。进一步的详情参见NatureBiotechnology23(12),1556-1561(2005)和ExpertOpiniononInvestigationalDrugs16(6),909-917(June2007)。转铁蛋白是单体的血清转运糖蛋白。可以通过在允许的表面环中插入肽序列将转铁蛋白工程化以结合不同的靶抗原。工程化转铁蛋白支架的实例包括Trans-body。进一步的详情参见J.Biol.Chem274,24066-24073(1999)。设计的锚蛋白重复蛋白(DARPin)衍生自锚蛋白,锚蛋白是介导整合膜蛋白与细胞骨架附着的蛋白质家族。单个锚蛋白重复是由两个α-螺旋和β-转角组成的33个残基的基序。通过随机化每个重复的第一个α-螺旋和β-转角中的残基,它们可以被工程化以结合不同的靶抗原。通过增加模块的数量(亲和力成熟法)可以增加它们的结合界面。进一步的详情参见J.Mol.Biol.332,489-503(2003),PNAS100(4),1700-1705(2003)和J.Mol.Biol.369,1015-1028(2007)以及US20040132028A1。单域抗体是由单个单体的可变抗体域组成的抗体片段。第一个单域来源于来自骆驼科动物(camelids)抗体重链的可变域(纳米抗体或VHH片段)。此外,术语单域抗体包括自主的人重链可变域(aVH)或来源于鲨鱼的VNAR片段。纤连蛋白是一种可以被工程化为与抗原结合的支架。Adnectin由人纤连蛋白III型(FN3)的15个重复单元的第10个域的天然氨基酸序列的主链组成。可以将β-夹心一端的三个环工程化,使Adnectin能够特异性地识别感兴趣的治疗靶标。进一步的详情参见ProteinEng.Des.Sel.18,435-444(2005),US20080139791,WO2005056764和US6818418B1。肽适体是由恒定骨架蛋白组成的组合识别分子,所述骨架蛋白通常是含有在活性位点插入的约束可变肽环的硫氧还蛋白(TrxA)。进一步的详情参见ExpertOpin.Biol.Ther.5,783-797(2005)。微体源自长度为25-50个氨基酸的天然存在的微蛋白,其含有3-4个半胱氨酸桥-微蛋白的实例包括KalataBI以及芋螺毒素和半胱氨酸结肽(knottin)。微蛋白具有可以工程化以包括多达25个氨基酸而不影响微蛋白的整体折叠的环。工程化的半胱氨酸结肽域的更多详情参见WO2008098796。就参考分子而言“结合相同表位的抗原结合分子”是指在竞争测定中阻断参考分子与其抗原的结合的50%或以上的抗原结合分子,相反地,参考分子在竞争测定中阻断抗原结合分子与其抗原的结合的50%或以上。如本文中使用的,术语“抗原结合位点”是指抗原结合分子的一部分,其特异性结合抗原决定簇。更加特别地,术语“抗原结合位点”是指抗体的一部分,其包含与抗原的部分或全部特异性地结合且互补的区域。在大抗原的情况下,抗原结合分子可能仅与该抗原的特定部分结合,该部分称作表位。抗原结合位点可以由例如一个或多个可变域(也称为可变区)提供。优选地,抗原结合位点包含抗体轻链可变区(VL)和抗体重链可变区(VH)。在一个方面,抗原结合位点能够结合它的抗原并阻断或部分阻断它的功能。特异性结合PD1或TIM-3的抗原结合位点包括如本文中进一步定义的抗体及其片段。另外,抗原结合位点可包括支架抗原结合蛋白,例如基于设计的重复蛋白或设计的重复域的结合域(参见例如WO2002/020565)。如本文中使用的,术语“抗原决定簇”与“抗原”和“表位”同义,并且是指在抗原结合模块与其结合而形成抗原结合模块-抗原复合物的多肽大分子上的位点(例如氨基酸的连续段或由不连续氨基酸的不同区域构成的构象构型)。有用的抗原决定簇可以例如在肿瘤细胞的表面上,病毒感染细胞的表面上,其他病变的细胞表面上,免疫细胞表面上发现,在血清中和/或细胞外基质(ECM)中游离。用作本文中的抗原的蛋白质可以是来自任何脊椎动物来源的蛋白质的任何天然形式,所述脊椎动物包括哺乳动物如灵长类动物(例如人)和啮齿动物(例如小鼠和大鼠),除非另外指明。在一个具体实施方案中,抗原是人蛋白质。在提及本文中的特定蛋白质的情况下,该术语包括“全长”未加工的蛋白质以及通过在细胞中加工产生的任何形式的蛋白质。该术语还涵盖蛋白质的天然存在的变体,例如,剪接变体或等位基因变体。“特异性结合”意指结合对于抗原而言是选择性的,并且可以与不需要的或非特异性的相互作用区别开。抗原结合分子与特异性抗原结合的能力可以通过酶联免疫吸附测定(ELISA)或本领域技术人员熟悉的其他技术来测量,例如,表面等离子体共振(SPR)技术(在BIAcore仪器上分析)(Liljeblad等人,GlycoJ17,323-329(2000))和传统结合测定法(Heeley,EndocrRes28,217-229(2002))。在一个实施方案中,抗原结合分子与不相关蛋白的结合程度小于该抗原结合分子与抗原结合的约10%,正如通过例如SPR所测量的。在某些实施方案中,与抗原结合的分子具有≤1μM,≤100nM,≤10nM,≤1nM,≤0.1nM,≤0.01nM或≤0.001nM(例如10-7M或更小,例如10-7M至10-13M,例如10-9M至10-13M)的解离常数(Kd)。“亲和力”或“结合亲和力”是指分子(例如抗体)的单一结合位点与其结合伴侣(例如抗原)之间的非共价相互作用的总和的强度。除非另外指明,如本文中使用的,“结合亲和力”是指反映结合对的成员(例如抗体和抗原)之间的1:1相互作用的固有结合亲和力。分子X对其伴侣Y的亲和力通常可以通过解离常数(Kd)来表示,亲和力是解离速率常数与结合速率常数(分别为koff和kon)的比率。因此,等同亲和力可以包含不同的速率常数,只要速率常数的比率保持相同即可。可以通过本领域已知的常用方法测量亲和力,包括本文所述的那些方法。用于测量亲和力的特定方法是表面等离子体共振(SPR)。如本文中使用的,抗体的术语“高亲和力”指抗体具有针对靶抗原的10-9M或更小和甚至更加特别的是10-10M或更小的Kd。抗体的术语“低亲和力”指抗体具有10-8或更高的Kd。“亲和力成熟的”抗体指与不拥有此类改变的亲本抗体相比,在一个或多个高变区(HVR)中具有一处或多处改变的抗体,此类改变导致该抗体对抗原的亲和力改善。术语“包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体”,“特异性结合PD1和TIM-3的双特异性抗体”,“对PD1和TIM-3特异性的双特异性抗原结合分子”在本文中可互换使用且指能够以足够的亲和力结合PD1和TIM-3,使得抗体可用作靶向PD1和TIM-3中的诊断和/或治疗剂的双特异性抗体。术语“PD1”,也称作编程性细胞死亡蛋白1,是一种288个氨基酸的I型膜蛋白,首次在1992年有描述(Ishidaetal.,EMBOJ.,11(1992),3887-3895)。PD-1是扩大的CD28/CTLA-4T细胞调节物家族的一个成员且具有两种配体,PD-L1(B7-H1,CD274)和PD-L2(B7-DC,CD273)。该蛋白质的结构包括一个胞外IgV域,接着是一个跨膜区和一个胞内尾。胞内尾含有位于免疫受体基于酪氨酸的抑制性基序和免疫受体基于酪氨酸的转换基序中的两个磷酸化位点,提示PD-1阴性负调节TCR信号。这与配体结合后SHP-1和SHP-2磷酸酶对PD-1的胞质尾的结合一致。虽然PD-1不在幼稚T细胞上表达,但是它在T细胞受体(TCR)介导的活化后上调且在活化的和耗尽的T细胞二者上观察到(Agataetal.,Int.Immunology8(1996),765-772)。这些耗尽的T细胞具有功能障碍性表型且不能够恰当地响应。虽然PD-1具有相对广泛的表达样式,但是它最重要的作用很可能是作为T细胞上的共抑制性受体(Chinaietal.,TrendsinPharmacologicalSciences36(2015),587-595)。当前的治疗办法因而聚焦于阻断PD-1与它的配体的相互作用以增强T细胞应答。术语“编程性死亡1”,“编程性细胞死亡1”,“蛋白PD-1”,“PD-1”,PD1”,PDCD1”,“hPD-1”和“hPD-I”可以互换使用,而且包括人PD-1的变体,同等型,物种同系物,和与PD-1具有至少一个共同表位的类似物。人PD1的氨基酸序列在UniProt(www.uniprot.org)登录号Q15116(SEQIDNO:89)中显示。术语“抗PD1抗体”和“包含结合PD1的抗原结合位点的抗体”指能够以足够的亲和力结合PD1,尤其是在细胞表面上表达的PD1多肽,使得抗体可用作靶向PD1中的诊断和/或治疗剂的抗体。在一个实施方案中,抗PD1抗体对无关的、非PD1蛋白的结合程度小于抗体对PD1的结合的约10%,如例如通过放射免疫测定法(RIA)或流式细胞术(FACS)或通过表面等离振子共振测定法使用生物传感器系统诸如系统测量的。在某些实施方案中,结合人PD1的抗原结合蛋白对于结合人PD1的结合亲和力具有≤1μM、≤100nM、≤10nM、≤1nM、≤0.1nM、≤0.01nM、或≤0.001nM(例如10-8M或更少,例如10-8M至10-13M,例如10-9M至10-13M)的KD值。在一个优选实施方案中,结合亲和力的相应KD值是在表面等离振子共振测定法中使用人PD1的胞外域(ECD)(PD1-ECD)(对于PD1结合亲和力)测定的。术语“抗PD1抗体”还涵盖能够结合PD1和第二抗原的双特异性抗体。术语“TIM3”,“含有T细胞免疫球蛋白和粘蛋白域的分子3”的缩写,也称作TIM-3,HAVCR2,KIM-3,TIMD3,和FLJ14428,指一种T辅助细胞1型特异性细胞表面蛋白,其调节巨噬细胞活化和炎性状况的严重程度。TIM3还与癌症,特别是癌干细胞有关。知道许多物种的TIM3的核苷酸和蛋白质序列。例如,人氨基酸序列可以在Uniprot登录号Q8TDQ0(SEQIDNO:93)下找到。人蛋白质特征在于一个胞外域,其包含Ig样域和粘蛋白域(进一步包含O连接的和N连接的糖基化位点),包含大约氨基酸22-202,一个跨膜域(氨基酸203-223),和一个胞内(胞质)域(氨基酸224-301)。对于作为SEQIDNO:93显示的人TIM3蛋白,胞外域包含大约氨基酸22-202,跨膜域包含大约氨基酸203-223,而胞质域包含大约氨基酸224-301。术语“TIM3”包括人TIM3的变体,同等型,物种同系物,和与TIM3具有至少一个共同表位的类似物。术语“抗TIM3抗体”和“包含结合TIM3的抗原结合位点的抗体”指能够以足够的亲和力结合TIM3,尤其是在细胞表面上表达的TIM3多肽,使得抗体可用作靶向TIM3中的诊断和/或治疗剂的抗体。在一个实施方案中,抗TIM3抗体对无关的、非TIM3蛋白的结合程度小于抗体对TIM3的结合的约10%,如例如通过放射免疫测定法(RIA)或流式细胞术(FACS)或通过表面等离振子共振测定法使用生物传感器系统诸如系统测量的。在某些实施方案中,结合人TIM3的抗原结合蛋白对于结合人TIM3的结合亲和力具有≤1μM、≤100nM、≤10nM、≤1nM、≤0.1nM、≤0.01nM、或≤0.001nM(例如10-8M或更少,例如10-7M至10-13M,例如10-9M至10-13M)的KD值。在一个优选实施方案中,结合亲和力的相应KD值是在表面等离振子共振测定法中使用人TIM3的胞外域(ECD)(TIM3-ECD)(对于TIM3结合亲和力)测定的。术语“抗TIM3抗体”还涵盖能够结合TIM3和第二抗原的双特异性抗体。“阻断性”抗体或“拮抗性”抗体是抑制或降低它结合的抗原的生物学活性的。在一些实施方案中,阻断性抗体或拮抗性抗体实质性或完全抑制抗原的生物学活性。例如,本发明的双特异性抗体阻断经由PD-1和TIM-3的信号传导从而自功能障碍性状态恢复T细胞对抗原刺激的功能性应答(例如增殖,细胞因子生成,靶细胞杀伤)。术语“可变区”或“可变域”是指涉及抗原结合分子与抗原结合的抗体重链或轻链的域。天然抗体的重链和轻链的可变域(分别为VH和VL)通常具有相似的结构,每个域包含四个保守的框架区(FR)和三个高变区(HVR)。参见例如Kindt等人,KubyImmunology,6thed.,W.H.Freeman和Co.,第91页(2007)。单个VH或VL域可足以赋予抗原结合特异性。如本文中使用的术语“高变区”或“HVR”是指在序列上高变/或形成结构上定义的环(“高变环”)的抗体可变域的每个区域。通常,天然四链抗体包含六个HVR:三个在VH中(H1,H2,H3)中,三个在VL中(L1,L2,L3)中。HVR通常包含来自高变环和/或来自“互补决定区”(CDR)的氨基酸残基,后者具有最高的序列变异性和/或涉及抗原识别。示例性的高变环发生在氨基酸残基26-32(L1),50-52(L2),91-96(L3),26-32(H1),53-55(H2),和96-101(H3)处。(Chothia和Lesk,J.Mol.Biol.196:901-917(1987).)示例性的CDR(CDR-L1,CDR-L2,CDR-L3,CDR-H1,CDR-H2,和CDR-H3)发生在L1的24-34,L2的50-56,L3的89-97,H1的31-35B,H2的50-65,和H3的95-102氨基酸残基处。(Kabat等人,SequencesofProteinsofImmunologicalInterest,5thEd.PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991).)高变区(HVR)也被称为互补决定区(CDR),并且在提到形成抗原结合区的可变区的部分时,这些术语在本文中可互换地使用。这个特定的区域已经由Kabat等人在U.S.Dept.ofHealthandHumanServices,"SequencesofProteinsofImmunologicalInterest"(1983)和Chothia等人在J.Mol.Biol.196:901-917(1987)中描述,其中定义包括当针对彼此进行比较时的氨基酸残基的重叠或子集。然而,提及抗体的CDR或其变体的任一定义的应用预期在本文中定义和使用的术语的范围内。涵盖如以上每一篇引用的参考文献所定义的CDR的适当氨基酸残基在以下表A中列出作为比较。涵盖特定CDR的确切残基数将取决于CDR的序列和大小而变化。考虑到抗体的可变区氨基酸序列,本领域技术人员可以常规地确定哪些残基包含特定的CDR。表A:CDR定义1CDRKabatChothiaAbM2VHCDR131-3526-3226-35VHCDR250-6552-5850-58VHCDR395-10295-10295-102VLCDR124-3426-3224-34VLCDR250-5650-5250-56VLCDR389-9791-9689-971表A中所有CDR定义的编号遵循Kabat等人提出的编号规则(见下文)。2具有如表A中使用的小写“b”的“AbM”是指OxfordMolecular的“AbM”抗体建模软件所定义的CDR。Kabat等人还定义了可应用于任何抗体的可变区序列的编号系统。本领域的普通技术人员可以将这个“Kabat编号”系统明确地指定给任何可变区序列,而不依赖于序列本身以外的任何实验数据。如本文中使用的,“Kabat编号”是指由Kabat等人在U.S.Dept.ofHealthandHumanServices,"SequenceofProteinsofImmunologicalInterest"(1983)中提出的编号系统。除非另有说明,抗体可变区中特定氨基酸残基位置编号的提及是根据Kabat编号系统。除了VH中的CDR1之外,CDR通常包含形成高变环的氨基酸残基。CDR还包含作为接触抗原的残基的“特异性测定残基”或“SDR”。SDR被包含在称为缩略CDR(或a-CDR)的CDR区域中。示例性的a-CDR(a-CDR-L1,a-CDR-L2,a-CDR-L3,a-CDR-H1,a-CDR-H2,和a-CDR-H3)发生在L1的31-34,L2的50-55,L3的89-96,H1的31-35B,H2的50-58,和H3的95-102氨基酸残基处。(参见Almagro和Fransson,Front.Biosci.13:1619-1633(2008).)除非另外指明,在本文中根据上文Kabat等人将可变域中的HVR残基和其他残基(例如FR残基)编号。“框架”或“FR”是指除了高变区(HVR)残基之外的可变域残基。可变域的FR通常由FR1,FR2,FR3和FR4四个FR域组成。因此,HVR和FR序列通常在VH(或VL)中以下列顺序出现:FR1-H1(L1)-FR2-H2(L2)-FR3-H3(L3)-FR4。出于本文的目的,“受体人框架”是包含衍生自人免疫球蛋白框架或人共有框架的轻链可变域(VL)框架或重链可变域(VH)框架的氨基酸序列的框架,如下所定义。“衍生自”人免疫球蛋白框架或人共有框架的受体人框架可以包含相同的氨基酸序列,或者其可以含有氨基酸序列变化。在一些实施方案中,氨基酸变化的数目为10个或更少,9个或更少,8个或更少,7个或更少,6个或更少,5个或更少,4个或更少,3个或更少,或2个或更少。在一些实施方案中,VL受体人框架在序列上与VL人免疫球蛋白框架序列或人共有框架序列相同。术语“嵌合”抗体是指其中重链和/或轻链的一部分衍生自特定来源或物种的抗体,同时重链和/或轻链的其余部分衍生自不同的来源或物种。抗体的“类”是指其重链所具有的恒定域或恒定区的类型。有五大类的抗体:IgA,IgD,IgE,IgG和IgM,其中几类可进一步分为亚类(同种型),例如,IgG1,IgG2,IgG3,IgG4,IgA1,和IgA2。对应于不同类型免疫球蛋白的重链恒定域分别称作α,δ,ε,γ,和μ。“人源化”抗体是指包含来自非人HVR的氨基酸残基和来自人FR的氨基酸残基的嵌合抗体。在某些实施方案中,人源化抗体将包含至少一个(通常两个)可变域中的基本上所有的可变域,其中所有或基本上所有的HVR(例如,CDR)对应于非人抗体的HVR,并且所有或基本上所有的FR对应于人抗体的FR。人源化抗体任选地可以包含衍生自人抗体的抗体恒定区的至少一部分。抗体例如非人抗体的“人源化形式”是指经过人源化的抗体。本发明涵盖的其他形式的“人源化抗体”是其中恒定区已经基于原始抗体的恒定区另外加以修饰或改变以产生根据本发明的特性特别是关于C1q结合和/或Fc受体(FcR)结合的那些抗体。“人”抗体是具有这样的氨基酸序列的抗体,所述氨基酸序列对应于人或人细胞产生的抗体或衍生自利用人抗体库或其他的人抗体编码序列的非人来源的抗体的氨基酸序列。人抗体的这个定义明确排除包含非人抗原结合残基的人源化抗体。如本文中使用的,术语“单克隆抗体”指从一群基本上同质的抗体获得的抗体,即构成群体的各个抗体是相同的和/或结合相同表位,除了例如含有天然存在的突变或在单克隆抗体制备物的生成期间发生的可能的变体抗体外,此类变体一般以极小量存在。与通常包含针对不同决定簇(表位)的不同抗体的多克隆抗体制备物不同,单克隆抗体制备物的每个单克隆抗体针对抗原上的单一决定簇。如此,修饰语“单克隆”指示抗体自一群基本上同质的抗体获得的特征,而不应解释为要求通过任何特定方法来生成抗体。例如,可以通过多种技术来生成要依照本发明使用的单克隆抗体,包括但不限于杂交瘤方法,重组DNA方法,噬菌体展示方法,和利用含有整个或部分人免疫球蛋白基因座的转基因动物的方法,本文中描述了用于生成单克隆抗体的此类方法和其它例示性方法。本文中使用的术语“Fc域”或“Fc区”用于定义含有恒定区的至少一部分的抗体重链的C末端区。该术语包括天然序列Fc区和变体Fc区。特别地,人IgG重链Fc区从Cys226或从Pro230延伸到重链的羧基末端。然而,Fc区的C末端赖氨酸(Lys447)可能存在或可能不存在。重链的氨基酸序列总是以C末端赖氨酸呈现,然而本发明中包括没有C末端赖氨酸的变体。IgGFc区包含IgGCH2和IgGCH3域。人IgGFc区的“CH2域”通常从约位置231的氨基酸残基延伸至约位置340的氨基酸残基。在一个实施方案中,碳水化合物链与CH2域附接。本文中的CH2域可以是天然序列CH2域或变体CH2域。“CH3域”包含在Fc区中的C末端到CH2域的残基段(即从IgG的约位置341的氨基酸残基到约位置447的氨基酸残基)。本文中的CH3区可以是天然序列CH3域或变体CH3域(例如,具有在其一条链中引入的“突起”(“节”)和在其另一条链中相应的引入的“腔”(“穴”)的CH3域;参见美国专利5,821,333,将其明确地通过提述并入本文)。如本文中所述,这种变体CH3域可用于促进两个不相同的抗体重链的异源二聚化。除非在本文中另外说明,Fc区或恒定区中氨基酸残基的编号是根据EU编号系统也称为EU索引进行的,如Kabat等人在SequencesofProteinsofImmunologicalInterest,5thEd.PublicHealthService,NationalInstitutesofHealth,Bethesda,MD,1991中描述。“节入穴”技术例如描述于US5,731,168;US7,695,936;Ridgway等人,ProtEng9,617-621(1996)和Carter,JImmunolMeth248,7-15(2001)中。通常,该方法包括在第一多肽的界面处引入突起(“节”)并且在第二多肽的界面中引入相应的腔(“穴”),使得该突起可以定位在该腔中,从而促进异二聚体形成并阻碍同二聚体形成。通过用较大的侧链(例如酪氨酸或色氨酸)替换第一多肽的界面的小氨基酸侧链而构建突起。通过用较小的氨基酸侧链(例如丙氨酸或苏氨酸)替换大的氨基酸侧链,在第二多肽的界面中产生与突起具有相同或相似大小的补偿腔。可以通过改变编码多肽的核酸,例如通过位点特异性诱变,或通过肽合成而产生突起和腔。在具体的实施方案中,节修饰包括Fc域的两个亚基之一中的氨基酸替代T366W,并且穴修饰包括Fc域的两个亚基中的另一个亚基中的氨基酸替代T366S,L368A和Y407V。在一个另外的具体实施方案中,包含节修饰的Fc域的亚基另外包含氨基酸替代S354C,并且包含穴修饰的Fc域的亚基另外包含氨基酸替代Y349C。引入这两个半胱氨酸残基导致在Fc区的两个亚基之间形成二硫桥,从而进一步稳定该二聚体(Carter,JImmunolMethods248,7-15(2001))。“等同于免疫球蛋白Fc区的区域”预期包括免疫球蛋白Fc区的天然存在的等位基因变体,以及具有产生取代,添加或缺失的改变但不实质性地降低免疫球蛋白介导效应器功能(如抗体依赖性细胞毒性)的能力的变体。例如,可以从免疫球蛋白Fc区的N末端或C末端缺失一个或多个氨基酸,而没有生物学功能的实质性丧失。可以根据本领域已知的一般规则来选择这些变体,以便对活性具有最小的影响(参见,例如Bowie,J.U.等人,Science247:1306-10(1990))。术语“效应器功能”是指可归因于抗体Fc区的那些生物活性,其随抗体同种型而变化。抗体效应器功能的实例包括:C1q结合和补体依赖性细胞毒性(CDC),Fc受体结合,抗体依赖性细胞介导的细胞毒性(ADCC),抗体依赖性细胞吞噬作用(ADCP),细胞因子分泌,免疫复合物介导的经由抗原呈递细胞的抗原摄取,细胞表面受体(例如B细胞受体)的下调,和B细胞活化。“活化性Fc受体”是被抗体Fc区接合之后引出信号传导事件的Fc受体,所述事件刺激带有该受体的细胞履行效应器功能。活化性Fc受体包括FcγRIIIa(CD16a),FcγRI(CD64),FcγRIIa(CD32),和FcαRI(CD89)。特定的活化性Fc受体是人FcγRIIIa(参见UniProt登录号P08637,版本141)。术语“肽接头”是指包含一个或多个氨基酸,通常约2-20个氨基酸的肽。肽接头在本领域中是已知的或描述于本文中。适合的非免疫原性接头肽例如是,(G4S)n,(SG4)n或G4(SG4)n肽接头,其中“n”通常为1至10之间的数字,通常为2至4,特别是2。“融合”或“连接”意指组分(例如抗原结合位点和Fc域)通过肽键直接连接或经由一个或多个肽接头连接。在本申请中使用的术语“氨基酸”表示包含丙氨酸(三字母代码:ala,单字母代码:A),精氨酸(arg,R),天冬酰胺(asn,N),天冬氨酸(asp,D),半胱氨酸(cys,C),谷氨酰胺(gln,Q),谷氨酸(glu,E),甘氨酸(gly,G),组氨酸(his,H),异亮氨酸(ile,I),亮氨酸(leu,L),赖氨酸(lys,K),甲硫氨酸(met,M),苯丙氨酸(phe,F),脯氨酸(pro,P),丝氨酸(ser,S),苏氨酸(thr,T),色氨酸(trp,W),酪氨酸(tyr,Y)和缬氨酸(val,V)的天然存在的羧基α-氨基酸的组。相对于参考多肽(蛋白质)序列的“氨基酸序列同一性百分比(%)”定义为在序列比对和引入空位(如果需要的话,达到最大序列同一性百分比,并且不考虑作为序列同一性的一部分的任何保守取代)之后候选序列中与参考多肽序列中的氨基酸残基相同的氨基酸残基的百分比。以本领域技术人员熟知的各种方法可实现为了确定氨基酸序列同一性百分比的比对,例如,利用可公开获得的计算机软件,如BLAST,BLAST-2,ALIGN.SAWI或Megalign(DNASTAR)软件。本领域技术人员可确定用于比对序列的适当参数,包括在所比较的序列的全长上实现最大比对所需要的任何算法。然而,出于本文的目的,通过使用序列比较计算机程序ALIGN-2生成了氨基酸序列同一性%值。ALIGN-2序列比较计算机程序由Genentech公司创作,并且源代码已在美国版权局(U.S.CopyrightOffice,WashingtonD.C.,20559)与用户文件一起备案,在美国版权局以美国版权登记号TXU510087登记。ALIGN-2程序可从加利福尼亚州南旧金山的Genentech公司公开获得,或者可以从源代码编译。ALIGN-2程序应当编译为供在包括数字UNIXV4.0D在内的UNIX操作系统上使用。所有序列比较参数均由ALIGN-2程序设定且没有变化。在采用ALIGN-2进行氨基酸序列比较的情形下,给定氨基酸序列A相对于(与,或针对)给定氨基酸序列B的氨基酸序列同一性%(或者可表述为给定氨基酸序列A相对于(与,或针对)给定氨基酸序列B具有或包含一定的氨基酸序列同一性%)计算如下:100乘以分数X/Y其中X为通过序列比对程序ALIGN-2在A与B的程序比对中评定为相同匹配的氨基酸残基的数目,且其中Y为B中的氨基酸残基的总数。应当理解的是,在氨基酸序列A的长度不等于氨基酸序列B的长度的情况下,A相对于B的氨基酸序列同一性%不会等于B相对于A的氨基酸序列同一性%。除非另外特别说明,否则使用ALIGN-2计算机程序如前一段所述获得在本文中使用的所有氨基酸序列同一性%值。在某些方面,考虑了本文提供的本发明的双特异性抗体的氨基酸序列变体。例如,可能希望改善双特异性抗体的结合亲和力和/或其他生物学性质。双特异性抗体的氨基酸序列变体可以通过向编码所述分子的核苷酸序列引入适当的修饰,或通过肽合成来制备。这些修饰包括,例如,抗体氨基酸序列中的残基的缺失和/或插入和/或取代。可以进行缺失,插入和取代的任何组合以得到最终的构建体,条件是最终构建体具有期望的特征,例如抗原结合。取代型诱变的感兴趣部位包括HVR和框架(FR)。在表B中的标题“优选取代”下提供了保守取代,并且在下文中参照氨基酸侧链类别(1)至(6)进一步加以描述。可以将氨基酸替代引入感兴趣的分子中,并筛选出具有期望活性的产物,所述活性例如保留/增强的抗原结合,降低的免疫原性,或增强的ADCC或CDC。表B可以根据常见的侧链性质将氨基酸分组:(1)疏水的:正亮氨酸,Met,Ala,Val,Leu,Ile;(2)中性亲水的:Cys,Ser,Thr,Asn,Gln;(3)酸性的:Asp,Glu;(4)碱性的:His,Lys,Arg;(5)影响链取向的残基:Gly,Pro;(6)芳香族的:Trp,Tyr,Phe。非保守取代需要将这些类之一的成员替换为另一类。术语“氨基酸序列变体”包括其中在亲本抗原结合分子(例如人源化或人抗体)的一个或多个高变区残基中存在氨基酸替代的实质性变体。通常,经过选择用于进一步研究的所得变体相对于亲本抗原结合分子在某些生物学特性(例如,增加的亲和力,降低的免疫原性)方面将具有改变(例如提高)和/或将具有基本上保留的亲本抗原结合分子的某些生物学特性。示例性的取代变体是亲和力成熟抗体,可以方便地例如使用例如本文所述的那些基于噬菌体展示的亲和力成熟技术来产生所述抗体。简言之,将一个或多个HVR残基突变,并且将变体抗原结合分子展示在噬菌体上并针对特定的生物活性(例如结合亲和力)进行筛选。在某些实施方案中,取代,插入或缺失可以在一个或多个HVR内进行,只要这些改变基本上不降低抗原结合分子结合抗原的能力即可。例如,可以在HVR中进行基本上不降低结合亲和力的保守改变(例如本文中提供的保守取代)。用于鉴定可能被靶向诱变的抗体残基或区域的有用方法被称为“丙氨酸扫描诱变”,如Cunningham和Wells(1989)Science,244:1081-1085所述。在该方法中,鉴定残基或一组靶残基(例如,带电残基,如Arg,Asp,His,Lys和Glu),并用中性或带负电的氨基酸(例如丙氨酸或聚丙氨酸)替换,以确定抗体与抗原的相互作用是否受到影响。可以在针对初始取代显示功能敏感性的氨基酸位置引入进一步的取代。替代地,或另外地,抗原-抗原结合分子复合物的晶体结构用于鉴定抗体和抗原之间的接触点。这样的接触残基和相邻残基可以作为取代候选物被靶向或消除。可以筛选变体以确定它们是否具备期望的特性。氨基酸序列插入包括长度从一个残基到含有一百个或更多个残基的多肽的氨基和/或羧基末端融合,以及单个或多个氨基酸残基的序列间插入。末端插入的实例包括具有N末端甲硫氨酰残基的双特异性抗体。分子的其他插入变体包括与多肽的N末端或C末端的融合,以增加双特异性抗体的血清半衰期。在某些方面,改变本文中提供的双特异性抗体以增加或降低抗体被糖基化的程度。通过改变氨基酸序列从而产生或去除一个或多个糖基化位点,可以方便地获得分子的糖基化变体,例如,可以改变附着于Fc域的碳水化合物。由哺乳动物细胞产生的天然抗体通常包含支链的双触角(biantennary)寡糖,其通常通过N键与Fc区的CH2域的Asn297连接。参见,例如Wright等人,TIBTECH15:26-32(1997)。寡糖可以包括各种碳水化合物,例如甘露糖,N-乙酰氨基葡萄糖(GlcNAc),半乳糖和唾液酸,以及与在双触角寡糖结构的“茎”中的GlcNAc附接的岩藻糖。在一些实施方案中,可以在本发明的双特异性抗体中进行寡糖修饰,以产生具有某些改良特性的变体。在一个方面,提供了具有缺乏与Fc区附接(直接或间接)的岩藻糖的碳水化合物结构的双特异性抗体的变体。这样的岩藻糖基化变体可具有增强的ADCC功能,参见,例如美国专利公开Nos.US2003/0157108(Presta,L.)或US2004/0093621(KyowaHakkoKogyoCo.,Ltd)。本发明的双特异性抗体的其他变体包括具有二等分寡糖的那些,例如,其中与Fc区附接的双触角寡糖被GlcNAc二等分。这些变体可以具有降低的岩藻糖基化和/或增强的ADCC功能,参见例如WO2003/011878(Jean-Mairet等人);美国专利No.6,602,684(Umana等人);和US2005/0123546(Umana等人)。还提供了与Fc区附接的寡糖中具有至少一个半乳糖残基的变体。这样的抗体变体可以具有增强的CDC功能,并被描述于例如WO1997/30087(Patel等人);WO1998/58964(Raju,S.);和WO1999/22764(Raju,S.)中。在某些方面,可能希望产生本发明的双特异性抗体的半胱氨酸工程化的变体,例如“thioMAb”,其中该分子的一个或多个残基被半胱氨酸残基取代。在特定实施方案中,取代的残基出现在分子的可接近位置。通过用半胱氨酸取代那些残基,由此反应性硫醇基团被定位在抗体的可接近位点,并且可以用于将抗体与其他模块(例如药物模块或接头-药物模块)缀合以产生免疫缀合物。在某些实施方案中,任何一个或多个以下残基可以被半胱氨酸取代:轻链的V205(Kabat编号);重链的A118(EU编号);和重链Fc区的S400(EU编号)。可以按照例如美国专利No.7,521,541中的描述产生半胱氨酸工程化的抗原结合分子。在某些方面,本文中提供的双特异性抗体可进一步被修饰,以含有另外的本领域已知并且易于获得的非蛋白质模块。适用于抗体衍生化的模块包括但不限于水溶性聚合物。水溶性聚合物的非限制性实例包括但不限于聚乙二醇(PEG),乙二醇/丙二醇共聚物,羧甲基纤维素,葡聚糖,聚乙烯醇,聚乙烯吡咯烷酮,聚-1,3-二氧戊环,聚-1,3,6-三烷,乙烯/马来酸酐共聚物,聚氨基酸(均聚物或无规共聚物)和葡聚糖或聚(N-乙烯基吡咯烷酮)聚乙二醇,丙二醇均聚物,聚氧化丙烯/氧化乙烯共聚物,聚氧乙烯化多元醇(例如甘油),聚乙烯醇及其混合物。聚乙二醇丙醛由于其在水中的稳定性而具有在制造方面的优点。聚合物可以具有任何分子量,并且可以是支链或非支链的。与抗体附接的聚合物的数量可以变化,并且如果附接一种以上的聚合物的话,它们可以是相同或不同的分子。通常,用于衍生化的聚合物的数量和/或类型可以基于以下考虑事项来确定,包括但不限于,待改进的抗体的特定性质或功能,是否将双特异性抗体衍生物在限定的条件下用于治疗中,等等。在另一个方面,提供了可通过暴露于辐射而选择性加热的抗体与非蛋白质模块的缀合物。在一个实施方案中,非蛋白质模块是碳纳米管(Kam,N.W.等人,Proc.Natl.Acad.Sci.USA102(2005)11600-11605)。辐射可以具有任何波长,并且包括但不限于不损害普通细胞的波长,但所述波长将非蛋白质模块加热到杀死抗体-非蛋白质模块附近的细胞的温度。“免疫缀合物”是与包括但不限于细胞毒性剂在内的一个或多个异源分子缀合的抗体。术语“多核苷酸”是指分离的核酸分子或构建体,例如,信使RNA(mRNA),病毒来源的RNA或质粒DNA(pDNA)。多核苷酸可以包含常规的磷酸二酯键或非常规键(例如,可见于肽核酸(PNA)中的酰胺键)。术语“核酸分子”是指任何一个或多个核酸区段,例如,存在于多核苷酸中的DNA或RNA片段。关于“分离的”核酸分子或多核苷酸,意指已经从其天然环境中移出的核酸分子,DNA或RNA。例如,编码包含在载体中的多肽的重组多核苷酸出于本发明的目的被认为是分离的。分离的多核苷酸的另外的例子包括保持在异源宿主细胞中的重组多核苷酸或溶液形式的纯化的(部分地或基本上纯化的)多核苷酸。分离的多核苷酸包括通常含有多核苷酸分子的细胞中所含的多核苷酸分子,但多核苷酸分子在染色体外存在或在与其天然染色体位置不同的染色体位置处存在。分离的RNA分子包括本发明的体内或体外RNA转录物,以及正链和负链形式以及双链形式。根据本发明的分离的多核苷酸或核酸还包括合成产生的此类分子。另外,多核苷酸或核酸可以是或可以包括调节元件,例如启动子,核糖体结合位点或转录终止子。关于具有至少例如与本发明的参考核苷酸序列95%“相同”的核苷酸序列的核酸或多核苷酸,意指除了该多核苷酸序列可以包括至多该参考核苷酸序列的每100个核苷酸的5个点突变之外,所述多核苷酸的核苷酸序列与参考序列相同。换句话说,为了获得具有与参考核苷酸序列至少95%相同的核苷酸序列的多核苷酸,参考序列中至多5%的核苷酸可以被缺失或被另一个核苷酸取代,或者,在参考序列中,总核苷酸的至多5%的多个核苷酸可被插入参考序列。参考序列的这些改变可以发生在参考核苷酸序列的5'或3'末端位置或那些末端位置之间的任何位置,独立散布在参考序列的残基之间或在参考序列内的一个基团或多个连续的基团中。实际情况是,使用例如上面针对多肽所讨论的已知的计算机程序(例如ALIGN-2),可以常规地确定任何特定的多核苷酸序列是否与本发明的核苷酸序列具有至少80%,85%,90%,95%,96%,97%,98%或99%同一性。术语“表达盒”是指重组或合成产生的多核苷酸,其具有一系列允许特定核酸在靶细胞中转录的特定核酸元件。重组表达盒可以掺入质粒,染色体,线粒体DNA,质体DNA,病毒或核酸片段中。通常,除了其他序列之外,表达载体的重组表达盒部分包括待转录的核酸序列和启动子。在某些实施方案中,本发明的表达盒包含编码本发明的双特异性抗原结合分子或其片段的多核苷酸序列。术语“载体”或“表达载体”与“表达构建体”是同义的,并且是指用于将与其可操作地相关联的特定基因引入靶细胞中并且指导所述特定基因的表达的DNA分子。该术语包括作为自身复制核酸结构的载体以及组入其已被引入的宿主细胞中的基因组中的载体。本发明的表达载体包括表达盒。表达载体允许大量稳定的mRNA的转录。一旦表达载体进入靶细胞内,则通过细胞转录和/或翻译机制产生由该基因编码的核糖核酸分子或蛋白质。在一个实施方案中,本发明的表达载体包括包含编码本发明的双特异性抗原结合分子或其片段的多核苷酸序列的表达盒。术语“宿主细胞”,“宿主细胞系”和“宿主细胞培养物”可互换使用,并且是指在其中引入了外源核酸的细胞,包括这些细胞的后代。宿主细胞包括“转化体”和“转化细胞”,其包括初级转化细胞和从其衍生的后代,而不考虑传代次数。后代在核酸含量方面与亲本细胞可能不完全相同,而可能含有突变。在本文中包括具有与在原始转化细胞中所筛选或选择相同的功能或生物活性的突变体后代。宿主细胞是可用于产生本发明的双特异性抗原结合分子的任何类型的细胞系统。特别地,宿主细胞是原核或真核宿主细胞。宿主细胞包括培养的细胞,例如培养的哺乳动物细胞,例如CHO细胞,BHK细胞,NS0细胞,SP2/0细胞,YO骨髓瘤细胞,P3X63小鼠骨髓瘤细胞,PER细胞,PER.C6细胞或杂交瘤细胞,酵母细胞,昆虫细胞和植物细胞,在此仅列举少数,还有包括在转基因动物,转基因植物或培养的植物或动物组织内的细胞。药剂的“有效量”是指导致其所施用的细胞或组织的生理变化所必需的量。药剂(例如药物组合物)的“治疗有效量”是指在剂量和时间段方面有效实现期望的治疗或预防结果所需的量。例如,治疗有效量的药剂消除,减少,延迟,最小化或预防疾病的不利影响。“个体”或“受试者”为哺乳动物。哺乳动物包括但不限于驯养动物(例如母牛,绵羊,猫,狗和马),灵长类动物(例如人类和非人灵长类动物如猴子),兔,和啮齿动物(例如小鼠和大鼠)。具体地说,个体或受试者是人。术语“药物组合物”是指其为这样的形式的制剂,所述形式允许其中所含的活性成分的生物活性是有效的,并且所述制剂不含另外的对该制剂所施用的受试者具有不可接受的毒性的组分。“药学上可接受的赋形剂”是指药物组合物中除了活性成分以外的对受试者无毒的成分。药学上可接受的赋形剂包括但不限于缓冲剂,稳定剂或防腐剂。术语“包装说明书”用于指通常包括在治疗产品的商业包装中的说明书,其包含关于使用这种治疗产品的适应症,用法,剂量,给药,联合疗法,禁忌症和/或警告的信息。如本文中使用的,“治疗/处理”(及其语法变化)是指试图改变被治疗个体的自然病程的临床干预,所述临床干预可用于预防或在临床病理过程中进行。期望的治疗效果包括但不限于预防疾病的发生或复发,症状减轻,疾病的任何直接或间接病理结果的削减,预防转移,降低疾病进展速度,改善或减轻疾病状态,以及缓解或改善预后。在一些实施方案中,本发明的分子用于延缓疾病的发展或减缓疾病的进展。如本文中使用的术语“癌症”是指增殖性疾病,例如淋巴瘤,淋巴细胞性白血病,肺癌,非小细胞肺癌(NSCL),支气管肺泡细胞肺癌,骨癌,胰腺癌,皮肤癌,头颈癌,皮肤或眼内黑素瘤,子宫癌,卵巢癌,直肠癌,肛门区癌,胃癌,胃的癌,结肠癌,乳腺癌,子宫癌,输卵管癌,子宫内膜癌,子宫颈癌,阴道癌,外阴癌,霍奇金病,食道癌,小肠癌,内分泌系统的癌症,甲状腺癌,甲状旁腺癌,肾上腺癌,软组织肉瘤,尿道癌,阴茎癌,前列腺癌,膀胱癌,肾癌或输尿管癌,肾细胞癌,肾盂癌,间皮瘤,肝细胞癌,胆道癌,中枢神经系统(CNS)肿瘤,脊索瘤,脑干神经胶质瘤,多形性胶质母细胞瘤,星形细胞瘤,神经鞘瘤,室管膜瘤,成神经管细胞瘤,脑膜瘤,鳞状细胞癌,垂体腺瘤和尤文氏肉瘤,包括任何上述癌症的难治性形式,或上述一种或多种癌症的组合。本发明的双特异性抗体本发明提供新颖的双特异性抗体,其包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点,具有特别有利的性质,诸如可生产性,稳定性,结合亲和力,生物学活性,对某些T细胞的特异性靶向,靶向效率和降低的毒性。特别地,这些是双特异性抗体,其中该双特异性抗体以高亲和力结合PD1且以低亲和力结合TIM3。A.例示性的结合PD1和TIM-3的双特异性抗体在一个方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1;(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含(a)VH域,其包含(i)包含SEQIDNO:1的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:2的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:3的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:4或SEQIDNO:11或SEQIDNO:12的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:5的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:6的氨基酸序列的HVR-L3;或(b)VH域,其包含(i)包含SEQIDNO:17的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:18的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:19的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:20的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:21的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:22的氨基酸序列的HVR-L3;或(c)VH域,其包含(i)包含SEQIDNO:29的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:30的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:31的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:32的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:33的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:34的氨基酸序列的HVR-L3。在一个特定的方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1;(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含VH域,其包含(i)包含SEQIDNO:1的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:2的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:3的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:12的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:5的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:6的氨基酸序列的HVR-L3。在另一个特定的方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1;(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含VH域,其包含(i)包含SEQIDNO:17的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:18的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:19的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:20的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:21的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:22的氨基酸序列的HVR-L3。在又一个方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含(a)包含SEQIDNO:43的氨基酸序列的VH域和包含SEQIDNO:44的氨基酸序列的VL域,或(b)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,或(c)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:47的氨基酸序列的VL域,或(d)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:48的氨基酸序列的VL域,或(e)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:49的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含(a)包含SEQIDNO:7的氨基酸序列的VH域和包含SEQIDNO:8的氨基酸序列的VL域,或(b)包含SEQIDNO:9的氨基酸序列的VH域和包含SEQIDNO:10的氨基酸序列的VL域,或(c)包含SEQIDNO:13的氨基酸序列的VH域和包含SEQIDNO:14的氨基酸序列的VL域,或(d)包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域,或(e)包含SEQIDNO:23的氨基酸序列的VH域和包含SEQIDNO:24的氨基酸序列的VL域,或(f)包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域,或(g)包含SEQIDNO:27的氨基酸序列的VH域和包含SEQIDNO:28的氨基酸序列的VL域,或(h)包含SEQIDNO:35的氨基酸序列的VH域和包含SEQIDNO:36的氨基酸序列的VL域。在又一个方面,该包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体是人,人源化或嵌合抗体。特别地,它是人源化抗体。在一个方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的人源化,双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含(a)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,或(b)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:47的氨基酸序列的VL域,或(c)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:48的氨基酸序列的VL域,或(d)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:49的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含(a)包含SEQIDNO:13的氨基酸序列的VH域和包含SEQIDNO:14的氨基酸序列的VL域,或(b)包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域,或(c)包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域,或(d)包含SEQIDNO:27的氨基酸序列的VH域和包含SEQIDNO:28的氨基酸序列的VL域。在一个特定的方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域或包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。特别地,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1;(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含VH域,其包含(i)包含SEQIDNO:17的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:18的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:19的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:20的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:21的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:22的氨基酸序列的HVR-L3。更加特别地,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含选自由SEQIDNO:46,SEQIDNO:47,SEQIDNO:48和SEQIDNO:49组成的组的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。更加具体地,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。在又一个方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含选自由SEQIDNO:46,SEQIDNO:47,SEQIDNO:48和SEQIDNO:49组成的组的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:27的氨基酸序列的VH域和包含SEQIDNO:28的氨基酸序列的VL域。在一个方面,该包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体是二价的。这意味着该双特异性抗体包含一个特异性结合PD1的抗原结合位点和一个特异性结合TIM3的抗原结合位点(1+1格式)。在另一个方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体以低亲和力结合TIM3且以高亲和力结合PD1。在一个特定的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体以与对PD1的结合相比低至少50倍的结合亲和力,更特别的是以与对PD1的结合相比低至少100倍的结合亲和力结合TIM3。在一个优选的实施方案中,该结合亲和力(KD)是用表面等离振子共振测定法(例如如实施例12中描述的)测定的。在一个方面,如此提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述以高亲和力特异性结合PD1的第一抗原结合位点包含(a)包含SEQIDNO:43的氨基酸序列的VH域和包含SEQIDNO:44的氨基酸序列的VL域,或(b)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,或(c)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:47的氨基酸序列的VL域,或(d)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:48的氨基酸序列的VL域,或(d)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:49的氨基酸序列的VL域,且所述以低亲和力特异性结合TIM3的第二抗原结合位点包含(a)包含SEQIDNO:23的氨基酸序列的VH域和包含SEQIDNO:24的氨基酸序列的VL域,或(c)包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域,或(d)包含SEQIDNO:27的氨基酸序列的VH域和包含SEQIDNO:28的氨基酸序列的VL域。在一个具体的方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述以高亲和力特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,且所述以低亲和力特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。在另一个方面,本发明涉及一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体包含Fc域,包含该特异性结合PD1的抗原结合位点的第一Fab片段和包含该特异性结合TIM3的抗原结合位点的第二Fab片段。特别地,该Fc域是IgG域,更加特别的是IgG1Fc域或IgG4Fc域。在另一个方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含(a)包含与SEQIDNO:50的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:52的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:51的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:53的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(b)包含与SEQIDNO:54的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:56的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:55的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:57的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(c)包含与SEQIDNO:58的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:60的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:59的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:61的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(d)包含与SEQIDNO:62的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:64的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:63的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:65的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(e)包含与SEQIDNO:66的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:68的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:67的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:69的序列有至少95%序列同一性的氨基酸序列的第二轻链。在一个特定的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含(a)包含SEQIDNO:50的氨基酸序列的第一重链,包含SEQIDNO:52的氨基酸序列的第一轻链,包含SEQIDNO:51的氨基酸序列的第二重链,和包含SEQIDNO:53的氨基酸序列的第二轻链,或(b)包含SEQIDNO:54的氨基酸序列的第一重链,包含SEQIDNO:56的氨基酸序列的第一轻链,包含SEQIDNO:55的氨基酸序列的第二重链,和包含SEQIDNO:57的氨基酸序列的第二轻链,或(c)包含SEQIDNO:58的氨基酸序列的第一重链,包含SEQIDNO:60的氨基酸序列的第一轻链,包含SEQIDNO:59的氨基酸序列的第二重链,和包含SEQIDNO:61的氨基酸序列的第二轻链,或(d)包含SEQIDNO:62的氨基酸序列的第一重链,包含SEQIDNO:64的氨基酸序列的第一轻链,包含SEQIDNO:63的氨基酸序列的第二重链,和包含SEQIDNO:65的氨基酸序列的第二轻链,或(e)包含SEQIDNO:66的氨基酸序列的第一重链,包含SEQIDNO:68的氨基酸序列的第一轻链,包含SEQIDNO:67的氨基酸序列的第二重链,和包含SEQIDNO:69的氨基酸序列的第二轻链。更加特别地,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含包含SEQIDNO:62的氨基酸序列的第一重链,包含SEQIDNO:64的氨基酸序列的第一轻链,包含SEQIDNO:63的氨基酸序列的第二重链,和包含SEQIDNO:65的氨基酸序列的第二轻链。在另一个特定的方面,提供的是一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含包含SEQIDNO:66的氨基酸序列的第一重链,包含SEQIDNO:68的氨基酸序列的第一轻链,包含SEQIDNO:67的氨基酸序列的第二重链,和包含SEQIDNO:69的氨基酸序列的第二轻链。在另一个方面,该包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体是四价的。在一个方面,该双特异性抗体包含两个特异性结合PD1的抗原结合位点和两个特异性结合TIM3的抗原结合位点(2+2格式)。在一个方面,本发明的双特异性抗体包含(a)包含两个如下Fab片段的抗体的两条轻链和两条重链,该Fab片段包含特异性结合TIM3的抗原结合位点,和(b)两个如下另外的Fab片段,该另外的Fab片段包含特异性结合PD1的抗原结合位点的,其中所述另外的Fab片段各自经由肽接头连接至(a)的重链的C端。在一个特定的方面,该肽接头是(G4S)4。在另一个方面,该两个包含特异性结合PD1的抗原结合位点的另外的Fab片段是交换Fab片段,其中可变域VL和VH彼此替换且VL-CH链各自经由肽接头连接至(a)的重链的C端。在一个特定的方面,本发明提供一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其包含(a)各自包含SEQIDNO:70的氨基酸序列的两条重链,包含SEQIDNO:71的氨基酸序列的第一轻链,和包含SEQIDNO:72的氨基酸序列的第二轻链,或(b)各自包含SEQIDNO:73的氨基酸序列的两条重链,包含SEQIDNO:74的氨基酸序列的第一轻链,和包含SEQIDNO:75的氨基酸序列的第二轻链,或(c)各自包含SEQIDNO:76的氨基酸序列的两条重链,包含SEQIDNO:77的氨基酸序列的第一轻链,和包含SEQIDNO:78的氨基酸序列的第二轻链。降低Fc受体结合和/或效应器功能的Fc域修饰在某些方面,可以将一处或多处氨基酸修饰引入本文中提供的抗体的Fc区中,由此生成Fc区变体。Fc区变体可以包含在一个或多个氨基酸位置包含氨基酸修饰(例如替代)的人Fc区序列(例如人IgG1,IgG2,IgG3或IgG4Fc区)。下一节描述本发明的包含降低Fc受体结合和/或效应器功能的Fc域修饰的双特异性抗原结合分子的优选方面。在一个方面,本发明涉及包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中Fc域包含一处或多处降低对Fc受体,特别是Fcγ受体的结合的氨基酸替代。特别地,该Fc域是人IgG1亚类的,其具有氨基酸突变L234A,L235A和P329G(编号方式依照KabatEU索引)。Fc域赋予本发明的双特异性抗体以有利的药动学特性,包括长血清半衰期,其有助于在靶组织中的较好积累和有利的组织-血液分配比。然而,同时它可能导致不想要的本发明的双特异性抗体对表达Fc受体的细胞而非优选的携带抗原的细胞的靶向。因而,在具体的实施方案中,本发明的双特异性抗体的Fc域展现出与天然IgGFc域,特别是IgG1Fc域或IgG4Fc域相比降低的对Fc受体的结合亲和力和/或降低的效应器功能。更加特别地,Fc域是IgG1Fc域。在一个此类方面,Fc域(或包含所述Fc域的本发明的双特异性抗原结合分子)展现出与天然IgG1Fc域(或包含天然IgG1Fc域的本发明的双特异性抗原结合分子)相比少于50%,优选少于20%,更优选少于10%且最优选少于5%的对Fc受体的结合亲和力,和/或与天然IgG1Fc域(或包含天然IgG1Fc域的本发明的双特异性抗原结合分子)相比少于50%,优选少于20%,更优选少于10%且最优选少于5%的效应器功能。在一个方面,Fc域(或包含所述Fc域的本发明的双特异性抗原结合分子)没有实质性结合Fc受体和/或诱导效应器功能。在一个特定的方面,所述Fc受体是Fcγ受体。在一个方面,所述Fc受体是人Fc受体。在一个方面,所述Fc受体是活化性Fc受体。在一个具体的方面,所述Fc受体是活化性人Fcγ受体,更具体地是人FcγRIIIa,FcγRI或FcγRIIa,最具体地是人FcγRIIIa。在一个方面,所述Fc受体是抑制性Fc受体。在一个具体的方面,所述Fc受体是抑制性人Fcγ受体,更具体地是人FcγRIIB。在一个方面,效应器功能是CDC,ADCC,ADCP,和细胞因子分泌中的一项或多项。在一个特定的方面,所述效应器功能是ADCC。在一个方面,所述Fc域展现出与天然IgG1Fc域相比基本相似的对新生儿Fc受体(FcRn)的结合亲和力。当Fc域(或包含所述Fc域的本发明的双特异性抗原结合分子)展现出超过约70%,特别是超过约80%,更特别是超过约90%的天然IgG1Fc域(或包含天然IgG1Fc域的本发明的双特异性抗原结合分子)对FcRn的结合亲和力时,实现基本相似的对FcRn的结合。在一个特定的方面,Fc域工程化改造为具有与非工程化Fc域相比降低的对Fc受体的结合亲和力和/或降低的效应器功能。在一个特定的方面,本发明的双特异性抗原结合分子的Fc域包含一处或多处降低Fc域对Fc受体的结合亲和力和/或效应器功能的氨基酸突变。通常,Fc域的两个亚基的每一个中存在相同的一处或多处氨基酸突变。在一个方面,所述氨基酸突变降低Fc域对Fc受体的结合亲和力。在另一个方面,所述氨基酸突变将Fc域对Fc受体的结合亲和力降低至少2倍,至少5倍,或至少10倍。在一个方面,包含工程化Fc域的本发明的双特异性抗原结合分子展现出与包含非工程化Fc域的本发明的双特异性抗体相比少于20%,特别是少于10%,更特别是少于5%的对Fc受体的结合亲和力。在一个特定的方面,所述Fc受体是Fcγ受体。在其它方面,所述Fc受体是人Fc受体。在一个方面,所述Fc受体是抑制性Fc受体。在一个具体的方面,所述Fc受体是抑制性人Fcγ受体,更具体地是人FcγRIIB。在一些方面,所述Fc受体是活化性Fc受体。在一个具体的方面,所述Fc受体是活化性人Fcγ受体,更特别是人FcγRIIIa,FcγRI或FcγRIIa,最特别是人FcγRIIIa。优选地,对这些受体的每一种的结合是降低的。在一些方面,对补体成分的结合亲和力,特别是对C1q的结合亲和力也是降低的。在一个方面,对新生儿Fc受体(FcRn)的结合亲和力没有降低。当Fc域(或包含所述Fc域的本发明的双特异性抗原结合分子)展现出非工程化形式的Fc域(或包含所述非工程化形式的Fc域的本发明的双特异性抗原结合分子)对FcRn的结合亲和力的超过约70%时,实现基本相似的对FcRn的结合,即保留该Fc域对所述受体的结合亲和力。Fc域或包含所述Fc域的本发明的双特异性抗原结合分子可以展现出超过约80%和甚至超过约90%的此类亲和力。在某些实施方案中,本发明的双特异性抗原结合分子的Fc域工程化改造为具有与非工程化Fc域相比降低的效应器功能。所述降低的效应器功能可包括但不限于下列一项或多项:降低的补体依赖性细胞毒性(CDC),降低的抗体依赖性细胞介导的细胞毒性(ADCC),降低的抗体依赖性细胞吞噬作用(ADCP),降低的细胞因子分泌,降低的免疫复合物介导的抗原呈递细胞的抗原摄取,降低的对NK细胞的结合,降低的对巨噬细胞的结合,降低的对单核细胞的结合,降低的对多形核细胞的结合,降低的诱导凋亡的直接信号传导,降低的树突细胞成熟,或降低的T细胞引发。具有降低的效应器功能的抗体包括那些具有Fc区残基238,265,269,270,297,327和329中的一个或多个的替代的(美国专利No.6,737,056)。此类Fc突变体包括在氨基酸位置265,269,270,297和327中的两处或更多处具有替代的Fc突变体,包括残基265和297替代成丙氨酸的所谓的“DANA”Fc突变体(美国专利No.7,332,581)。描述了具有改善的或降低的对FcR的结合的某些抗体变体(见例如美国专利No.6,737,056;WO2004/056312,及Shields,R.L.etal.,J.Biol.Chem.276(2001)6591-6604)。在本发明的一个方面,Fc域包含在E233,L234,L235,N297,P331和P329位置处的氨基酸替代。在一些方面,Fc域包含氨基酸替代L234A和L235A(“LALA”)。在一个此类实施方案中,Fc域是IgG1Fc域,特别是人IgG1Fc域。在一个方面,Fc域包含在位置P329处的氨基酸替代。在一个更加具体的方面,氨基酸替代是P329A或P329G,特别是P329G。在一个实施方案中,Fc域包含在位置P329处的氨基酸替代和又一处选自下组的氨基酸替代:E233P,L234A,L235A,L235E,N297A,N297D或P331S。在更具体的实施方案中,所述Fc域包含氨基酸突变L234A,L235A和P329G(“P329GLALA”)。氨基酸替代组合“P329GLALA”几乎完全消除了人IgG1Fc域的Fcγ受体结合,如记载于PCT专利申请No.WO2012/130831A1。所述文件还描述了制备此类突变体Fc域的方法和用于测定其特性(诸如Fc受体结合或效应器功能)的方法。此类抗体是具有突变L234A和L235A或具有突变L234A,L235A和P329G(编号方式依照Kabatetal,Kabatetal.,SequencesofProteinsofImmunologicalInterest,5thEd.PublicHealthService,NationalInstitutesofHealth,Bethesda,MD,1991的EU索引)的IgG1。在一个方面,本发明的双特异性抗体包含(所有位置依照Kabat的EU索引)(i)人IgG1亚类的同二聚体Fc区,任选具有突变P329G,L234A和L235A,或(ii)人IgG4亚类的同二聚体Fc区,任选具有突变P329G,S228P和L235E,或(iii)人IgG1亚类的同二聚体Fc区,任选具有突变P329G,L234A,L235A,I253A,H310A,和H435A,或任选具有突变P329G,L234A,L235A,H310A,H433A,和Y436A,或(iv)异二聚体Fc区,其中一条Fc区多肽包含突变T366W,且另一条Fc区多肽包含突变T366S,L368A和Y407V,或其中一条Fc区多肽包含突变T366W和Y349C,且另一条Fc区多肽包含突变T366S,L368A,Y407V,和S354C,或其中一条Fc区多肽包含突变T366W和S354C,且另一条Fc区多肽包含突变T366S,L368A,Y407V和Y349C,或(v)人IgG1亚类的异二聚体Fc区,其中两条Fc区多肽均包含突变P329G,L234A和L235A且一条Fc区多肽包含突变T366W,且另一条Fc区多肽包含突变T366S,L368A和Y407V,或其中一条Fc区多肽包含突变T366W和Y349C,且另一条Fc区多肽包含突变T366S,L368A,Y407V,和S354C,或其中一条Fc区多肽包含突变T366W和S354C,且另一条Fc区多肽包含突变T366S,L368A,Y407V和Y349C。在一个方面,Fc域为IgG4Fc域。在一个更具体的实施方案中,Fc域是包含在位置S228(Kabat编号)处的氨基酸替代具体地为氨基酸替代S228P的IgG4Fc域。在一个更具体的实施方案中,Fc域是包含氨基酸替代L235E和S228P和P329G的IgG4Fc域。这种氨基酸替代降低体内IgG4抗体的Fab臂交换(参见Stubenrauchetal.,DrugMetabolismandDisposition38,84-91(2010))。如此,在一个方面,提供的是一种双特异性抗体,其包含(所有位置依照Kabat的EU索引)人IgG4亚类的异二聚体Fc区,其中两条Fc区多肽均包含突变P329G,S228P和L235E且一条Fc区多肽包含突变T366W,且另一条Fc区多肽包含突变T366S,L368A和Y407V,或其中一条Fc区多肽包含突变T366W和Y349C,且另一条Fc区多肽包含突变T366S,L368A,Y407V,和S354C,或其中一条Fc区多肽包含突变T366W和S354C,且另一条Fc区多肽包含突变T366S,L368A,Y407V和Y349C。具有延长的半衰期和改善的对新生儿Fc受体(FcRn)的结合的抗体记载于US2005/0014934,新生儿Fc受体(FcRn)负责将母体IgG转移至胎儿(Guyer,R.L.etal.,J.Immunol.117(1976)587-593及Kim,J.K.etal.,J.Immunol.24(1994)2429-2434)。那些抗体包含其中具有改善Fc区对FcRn结合的一处或多处替代的Fc区。此类Fc变体包括那些在Fc区残基238,256,265,272,286,303,305,307,311,312,317,340,356,360,362,376,378,380,382,413,424或434中的一处或多处具有替代,例如,Fc区残基434的替代的(美国专利No.7,371,826)。还可见Duncan,A.R.andWinter,G.,Nature322(1988)738-740;US5,648,260;US5,624,821;及WO94/29351,其关注Fc区变体的其它例子。可以例如通过ELISA或通过使用诸如BIAcore仪(GEHealthcare)的标准仪器的表面等离子体共振(SPR)容易地确定与Fc受体的结合,并且可以例如通过重组表达获得Fc受体。适合的此类结合测定描述于本文中。或者,可以使用已知表达特定Fc受体的细胞系例如表达FcγIIIa受体的人NK细胞来评估Fc域或包含Fc域的细胞活化性双特异性抗原结合分子针对Fc受体的结合亲和力。可以通过本领域已知的方法测量Fc域或包含Fc域的本发明的双特异性抗体的效应器功能。适合的用于测量ADCC的测定法描述于本文中。评估感兴趣分子的ADCC活性的体外测定法的其他实例描述于美国专利No.5,500,362;Hellstrom等人,Proc.NatlAcadSciUSA83,7059-7063(1986)和Hellstrom等人,ProcNatlAcadSciUSA82,1499-1502(1985);美国专利No.5,821,337;Bruggemann等人,JExpMed166,1351-1361(1987)。或者,可以采用非放射性测定方法(参见,例如ACTITMnon-radioactivecytotoxicityassayforflowcytometry(CellTechnology,Inc.MountainView,CA);CytoToxnon-radioactivecytotoxicityassay(Promega,Madison,WI))。用于这种测定法的有用的效应细胞包括外周血单核细胞(PBMC)和自然杀伤(NK)细胞。替代地,或另外地,可以在体内,例如,在Clynes等人,ProcNatlAcadSciUSA95,652-656(1998)中公开的动物模型中评估感兴趣分子的ADCC活性。促进异源二聚化的Fc域修饰本发明的双特异性抗原结合分子包含与Fc域的两个亚基中的一个或另一个融合的不同抗原结合位点,因此该Fc域的两个亚基可包含在两条不相同的多肽链中。这些多肽的重组共表达和随后的二聚化导致两种多肽的数种可能的组合。为了在重组生产中提高本发明的双特异性抗体的产率和纯度,因此在本发明的双特异性抗原结合分子的Fc域中引入促进期望多肽的联合的修饰会是有利的。因此,在特定的方面,本发明涉及双特异性抗体,其包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点,其中该Fc域包含促进Fc域的第一和第二亚基联合的修饰。人IgGFc域的两个亚基之间最广泛的蛋白质-蛋白质相互作用的位点在Fc域的CH3域中。因此,在一个方面,所述修饰在Fc域的CH3域中。在一个具体的方面,所述修饰是所谓的“节入穴”修饰,包括在Fc域的两个亚基之一中的“节”修饰和Fc域的两个亚基的另一个中的“穴”修饰。因此,本发明涉及双特异性抗体,其包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点,其中依照该节入穴方法,该Fc域的第一亚基包含节且该Fc域的第二亚基包含穴。在一个特定的方面,该Fc域的第一亚基包含氨基酸替代S354C和T366W(EU编号方式)且该Fc域的第二亚基包含氨基酸替代Y349C,T366S和Y407V(编号方式依照KabatEU索引)。“节入穴”技术例如描述于US5,731,168;US7,695,936;Ridgway等人ProtEng9,617-621(1996)andCarter,JImmunolMeth248,7-15(2001)中。通常,该方法包括在第一多肽的界面处引入突起(“节”)并且在第二多肽的界面中引入相应的腔(“穴”),使得该突起可以定位在该腔中,从而促进异二聚体形成并阻碍同二聚体形成。通过用较大的侧链(例如酪氨酸或色氨酸)替换第一多肽的界面的小氨基酸侧链而构建突起。通过用较小的氨基酸侧链(例如丙氨酸或苏氨酸)替换大的氨基酸侧链,在第二多肽的界面中产生与突起具有相同或相似大小的补偿腔。因此,在一个方面,在本发明的双特异性抗原结合分子的Fc域的第一亚基的CH3结构域中,将氨基酸残基替换为具有较大侧链体积的氨基酸残基,由此在第一亚基的CH3结构域内产生可定位于第二亚基的CH3结构域内的腔中的突起;并且在Fc域的第二亚基的CH3结构域中,将氨基酸残基替换为具有较小侧链体积的氨基酸酸残基,由此在第二亚基的CH3结构域内产生可将第一亚基的CH3结构域内的突起定位在其中的腔。可以通过改变编码多肽的核酸,例如通过位点特异性诱变,或通过肽合成而产生突起和腔。在一个具体方面,在Fc域的第一亚基的CH3结构域中,第366位的苏氨酸残基替换为色氨酸残基(T366W),而在Fc域的第二亚基的CH3结构域中,第407位酪氨酸残基替换为缬氨酸残基(Y407V)。在一个方面,在Fc域的第二亚基中,另外将366位的苏氨酸残基替换为丝氨酸残基(T366S),将368位的亮氨酸残基替换为丙氨酸残基(L368A)。在还有又一个方面,在Fc域的第一亚基中,另外将第354位的丝氨酸残基替换为半胱氨酸残基(S354C),而在Fc域的第二亚基中,另外将第349位酪氨酸残基替换为半胱氨酸残基(Y349C)。引入这两个半胱氨酸残基导致在Fc域的两个亚基之间形成二硫桥,其进一步使该二聚体稳定(Carter(2001),JImmunolMethods248,7-15)。在一个特定的方面,该Fc域的第一亚基包含氨基酸替代S354C和T366W(EU编号方式)且该Fc域的第二亚基包含氨基酸替代Y349C,T366S和Y407V(编号方式依照KabatEU索引)。但是,或者/另外,也可以使用EP1870459记载的其它节入穴技术。在一个实施方案中,该多特异性抗体包含“节链”的CH3域中的突变R409D和K370E和“穴-链”的CH3域中的突变D399K和E357K(编号方式依照KabatEU索引)。在一个方面,该双特异性抗体包含“节链”的CH3域中的T366W突变和“穴链”的CH3域中的突变T366S,L368A和Y407V和另外地“节链”的CH3域中的突变R409D和K370E和“穴链”的CH3域中的突变D399K和E357K(编号方式依照KabatEU索引)。在一个方面,该双特异性抗体包含两个CH3域之一中的突变Y349C和T366W和两个CH3域之另一中的突变S354C,T366S,L368A和Y407V,或该多特异性抗体包含两个CH3域之一中的突变Y349C和T366W和两个CH3域之另一中的突变S354C,T366S,L368A和Y407V和另外地“节链”的CH3域中的突变R409D和K370E和“穴链”的CH3域中的突变D399K和E357K(编号方式依照KabatEU索引)。在一个替代方面,促进Fc域的第一和第二亚基的联合的修饰包括介导静电操纵效应的修饰,例如,如PCT公开WO2009/089004中所述。通常,这种方法包括通过将两个Fc域亚基的界面处的一个或多个氨基酸残基替换为带电氨基酸残基,使得在静电方面不利于同二聚体形成,但在静电方面有利于异二聚化。在“节入穴技术”以外,用于修饰多特异性抗体的重链的CH3域以加强异二聚化的其它技术是本领域已知的。本文中涵盖这些技术作为与双特异性抗体组合的“节入穴技术”的备选,尤其是WO96/27011,WO98/050431,EP1870459,WO2007/110205,WO2007/147901,WO2009/089004,WO2010/129304,WO2011/90754,WO2011/143545,WO2012/058768,WO2013/157954和WO2013/096291中记载的。在一个方面,在双特异性抗体中,使用EP1870459中记载的办法来支持多特异性抗体的第一重链和第二重链的异二聚化。这种办法基于在第一和第二重链二者之间的CH3/CH3域界面中的特定氨基酸位置处引入具有相反电荷的带电荷的氨基酸。因而,在这个方面,在多特异性抗体的三级结构中,第一重链的CH3域和第二重链的CH3域形成位于各自抗体CH3域之间的界面,其中第一重链的CH3域的各自氨基酸序列和第二重链的CH3域的氨基酸序列均包含位于抗体的三级结构中的所述界面内的一组氨基酸,其中自位于一条重链的CH3域中的界面中的该组氨基酸,第一氨基酸用带正电荷的氨基酸替代且自位于另一条重链的CH3域中的界面中的该组氨基酸,第二氨基酸用带负电荷的氨基酸替代。依照这个方面的双特异性抗体在本文中也称作“CH3(+/-)工程化双特异性抗体”(其中缩写+/-”代表在各自CH3域中引入的带相反电荷的氨基酸)。在一个方面,在CH3(+/-)工程化双特异性抗体中,带正电荷的氨基酸选自K,R和H,且带负电荷的氨基酸选自E或D。在一个方面,在CH3(+/-)工程化双特异性抗体中,带正电荷的氨基酸选自K和R,且带负电荷的氨基酸选自E或D。在一个方面,在CH3(+/-)工程化双特异性抗体中,带正电荷的氨基酸是K,且带负电荷的氨基酸是E。在一个方面,在CH3(+/-)工程化双特异性抗体中,在一条重链的CH3域中位置409处的氨基酸R用D替代且位置370处的氨基酸K用E替代,且在另一条重链的CH3域中位置399处的氨基酸D用K替代且位置357处的氨基酸E用K替代(编号方式依照KabatEU索引)。在一个方面,使用WO2013/157953中记载的办法来支持多特异性抗体的第一重链和第二重链的异二聚化。在一个实施方案中,在一条重链的CH3域中位置366处的氨基酸T用K替代,且在另一条重链的CH3域中位置351处的氨基酸L用D替代(编号方式依照KabatEU索引)。在另一个实施方案中,在一条重链的CH3域中位置366处的氨基酸T用K替代且位置351处的氨基酸L用K替代,且在另一条重链的CH3域中位置351处的氨基酸L用D替代(编号方式依照KabatEU索引)。在另一个方面,在一条重链的CH3域中位置366处的氨基酸T用K替代且位置351处的氨基酸L用K替代,且在另一条重链的CH3域中位置351处的氨基酸L用D替代(编号方式依照KabatEU索引)。另外,在另一条重链的CH3域中包含下述替代中至少一项:位置349处的氨基酸Y用E替代,位置349处的氨基酸Y用D替代且位置368处的氨基酸L用E替代(编号方式依照KabatEU索引)。在一个实施方案中,位置368处的氨基酸L用E替代(编号方式依照KabatEU索引)。在一个方面,使用WO2012/058768中记载的办法来支持多特异性抗体的第一重链和第二重链的异二聚化。在一个方面,在一条重链的CH3域中位置351处的氨基酸L用Y替代且位置407处的氨基酸Y用A替代,且在另一条重链的CH3域中位置366处的氨基酸T用A替代且位置409处的氨基酸K用F替代(编号方式依照KabatEU索引)。在另一个实施方案中,在上述替代以外,在另一条重链的CH3域中还替代氨基酸位置411(最初是T),399(最初是D),400(最初是S),405(最初是F),390(最初是N)和392(最初是K)中的至少一个(编号方式依照KabatEU索引)。优选的替代是:-用选自N,R,Q,K,D,E和W的氨基酸替代位置411处的氨基酸T(编号方式依照KabatEU索引),-用选自R,W,Y,和K的氨基酸替代位置399处的氨基酸D(编号方式依照KabatEU索引),-用选自E,D,R和K的氨基酸替代位置400处的氨基酸S(编号方式依照KabatEU索引),-用选自I,M,T,S,V和W的氨基酸替代位置405处的氨基酸F(编号方式依照KabatEU索引;-用选自R,K和D的氨基酸替代位置390处的氨基酸N(编号方式依照KabatEU索引;和-用选自V,M,R,L,F和E的氨基酸替代位置392处的氨基酸K(编号方式依照KabatEU索引)。在另一个方面,依照WO2012/058768改造双特异性抗体,即在一条重链的CH3域中位置351处的氨基酸L用Y替代且位置407处的氨基酸Y用A替代,且在另一条重链的CH3域中位置366处的氨基酸T用V替代且位置409处的氨基酸K用F替代(编号方式依照KabatEU索引)。在多特异性抗体的另一个实施方案中,在一条重链的CH3域中位置407处的氨基酸Y用A替代,且在另一条重链的CH3域中位置366处的氨基酸T用A替代且位置409处的氨基酸K用F替代(编号方式依照KabatEU索引)。在最后一个上述实施方案中,在另一条重链的CH3域中位置392处的氨基酸K用E替代,且位置411处的氨基酸T用E替代,且位置399处的氨基酸D用R替代且位置400处的氨基酸S用R替代(编号方式依照KabatEU索引)。在一个方面,使用WO2011/143545中记载的办法来支持多特异性抗体的第一重链和第二重链的异二聚化。在一个方面,在位置368和/或409处引入两条重链的CH3域中的氨基酸修饰(编号方式依照KabatEU索引)。在一个方面,使用WO2011/090762中记载的办法来支持双特异性抗体的第一重链和第二重链的异二聚化。WO2011/090762涉及依照“节入穴”(KiH)技术的氨基酸修饰。在一个实施方案中,在一条重链的CH3域中位置366处的氨基酸T用W替代,且在另一条重链的CH3域中位置407处的氨基酸Y用A替代(编号方式依照KabatEU索引)。在另一个实施方案中,在一条重链的CH3域中位置366处的氨基酸T用Y替代,且在另一条重链的CH3域中位置407处的氨基酸Y用T替代(编号方式依照KabatEU索引)。在一个方面,使用WO2009/089004中记载的办法来支持双特异性抗体的第一重链和第二重链的异二聚化。在一个实施方案中,在一条重链的CH3域中位置392处的氨基酸K或N用带负电荷的氨基酸(在一个实施方案中,用E或D,在一个优选的实施方案中,用D)替代,且在另一条重链的CH3域中位置399处的氨基酸D位置356处的氨基酸E或D或位置357处的氨基酸E用带正电荷的氨基酸替代(在一个实施方案中,用K或R,在一个优选的实施方案中,用K,在一个优选的实施方案中,位置399或356处的氨基酸用K替代)(编号方式依照KabatEU索引)。在一个进一步的实施方案,在上述替代以外,在一条重链的CH3域中位置409处的氨基酸K或R用带负电荷的氨基酸(在一个实施方案中,用E或D,在一个优选的实施方案中,用D)替代(编号方式依照KabatEU索引)。在一个甚至进一步的方面,作为上述替代的补充或替代,在一条重链的CH3域中位置439处的氨基酸K和/或位置370处的氨基酸K彼此独立地用带负电荷的氨基酸(在一个实施方案中,用E或D,在一个优选的实施方案中,用D)替代(编号方式依照KabatEU索引)。在一个方面,使用WO2007/147901中记载的办法来支持多特异性抗体的第一重链和第二重链的异二聚化。在一个实施方案中,在一条重链的CH3域中位置253处的氨基酸K用E替代,位置282处的氨基酸D用K替代且位置322处的氨基酸K用D替代,且在另一条重链的CH3域中位置239处的氨基酸D用K替代,位置240处的氨基酸E用K替代和且位置292处的氨基酸K用D替代(编号方式依照KabatEU索引)。本文中报告的双特异性抗体的重链的C端可以是以氨基酸残基PGK结尾的完整C端。重链的C端可以是缩短的C端,其中去除了C端氨基酸残基中的一个或两个。在一个优选的方面,重链的C端是以PG结尾的缩短的C端。在本文中报告的所有方面的一个方面,包含包括本文中规定的C端CH3域的重链的双特异性抗体包含C端甘氨酸-赖氨酸二肽(G446和K447,编号方式依照KabatEU索引)。在本文中报告的所有方面的一个实施方案中,包含包括本文中规定的C端CH3域的重链的双特异性抗体包含C端甘氨酸残基(G446,编号方式依照KabatEU索引)。Fab域中的修饰在一个方面,本发明涉及一种双特异性抗体,其包含特异性结合PD1的第一Fab片段和特异性结合TIM3的第二Fab片段,其中在该Fab片段之一中可变域VH和VL或恒定域CH1和CL任一是交换的。该双特异性抗体依照Crossmab技术来制备。具有一个结合臂中的域替换/交换的多特异性抗体(CrossMabVH-VL或CrossMabCH-CL)详细记载于WO2009/080252和Schaefer,W.etal,PNAS,108(2011)11187-1191。它们清楚地减少由针对第一抗原的轻链与错误的针对第二抗原的重链错配引起的副产物(与没有此类域交换的办法相比)。在一个特定的方面,本发明涉及一种双特异性抗体,其包含特异性结合PD1的第一Fab片段和特异性结合TIM3的第二Fab片段,其中在该Fab片段之一中可变域VL和VH彼此替换使得该VH域是轻链的一部分且该VL域是重链的一部分。在一个特定的方面,该双特异性抗体是这样一种,其中在包含该特异性结合PD1的抗原结合位点的第一Fab片段中可变域VL和VH彼此替换。在另一个方面及为了进一步提高正确配对,包含特异性结合PD1的第一Fab片段和特异性结合TIM3的第二Fab片段的双特异性抗体可以含有不同的带电荷的氨基酸替代(所谓的“带电荷的残基”)。在交叉的或非交叉的CH1和CL域中引入这些修饰。在一个特定的方面,本发明关注包含特异性结合PD1的第一Fab片段和特异性结合TIM3的第二Fab片段的双特异性抗体,其中在该Fab片段之一中在恒定域CL中位置124处的氨基酸用赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照KabatEU索引),且在恒定域CH1中位置147和213处的氨基酸用谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。在一个特定的方面,该双特异性抗体是如下的一种,其中在包含特异性结合TIM3的抗原结合位点的第二Fab片段中在恒定域CL中位置124处的氨基酸用赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照KabatEU索引),且在恒定域CH1中位置147和213处的氨基酸用谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。在一个特定的方面,本发明涉及包含特异性结合PD1的第一Fab片段和特异性结合TIM3的第二Fab片段的双特异性抗体,其中在CL域之一中位置123(EU编号方式)处的氨基酸用精氨酸(R)替换且位置124(EU编号方式)处的氨基酸用赖氨酸(K)替代且其中在CH1域之一中位置147(EU编号方式)处的和位置213(EU编号方式)处的氨基酸用谷氨酸(E)替代。在一个特定的方面,该双特异性抗体是如下的一种,其中在包含特异性结合TIM3的抗原结合位点的第二Fab片段中位置123(EU编号方式)处的氨基酸用精氨酸(R)替换且位置124(EU编号方式)处的氨基酸用赖氨酸(K)替代且其中在CH1域之一中位置147(EU编号方式)处的和位置213(EU编号方式)处的氨基酸用谷氨酸(E)替代。在又一个方面,该双特异性抗体是一种二价抗体,其包含a)特异性结合第一抗原的抗体的第一轻链和第一重链,和b)特异性结合第二抗原的抗体的第二轻链和第二重链,其中该第二轻链和该第二重链的可变域VL和VH彼此替换。a)下的抗体不含b)下报告的修饰和该重链且a)下的轻链是分离的链。在b)下的抗体中在轻链内可变轻链域VL用所述抗体的可变重链域VH替换,且在重链中可变重链域VH用所述抗体的可变轻链域VL替换。在一个方面,(i)在a)下的第一轻链的恒定域CL中位置124处的氨基酸(编号方式依照Kabat)用带正电荷的氨基酸替代,且其中在a)下的第一重链的恒定域CH1中位置147处的氨基酸或位置213处的氨基酸(编号方式依照KabatEU索引)用带负电荷的氨基酸替代,或(ii)在b)下的第二轻链的恒定域CL中位置124处的氨基酸(编号方式依照Kabat)用带正电荷的氨基酸替代,且其中在b)下的第二重链的恒定域CH1中位置147处的氨基酸或位置213处的氨基酸(编号方式依照KabatEU索引)用带负电荷的氨基酸替代。在另一个方面,(i)在a)下的第一轻链的恒定域CL中位置124处的氨基酸用赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照Kabat)(在一个优选的实施方案中,用赖氨酸(K)或精氨酸(R)独立替代),且其中在a)下的第一重链的恒定域CH1中位置147处的氨基酸或位置213处的氨基酸用谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引),或(ii)在b)下的第二轻链的恒定域CL中位置124处的氨基酸用赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照Kabat)(在一个优选的实施方案中,用赖氨酸(K)或精氨酸(R)独立替代),且其中在b)下的第二重链的恒定域CH1中位置147处的氨基酸或位置213处的氨基酸用谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。在一个方面,在第二重链的恒定域CL中位置124和123处的氨基酸用K替代(编号方式依照KabatEU索引)。在一个方面,在第二重链的恒定域CL中位置123处的氨基酸用R替代且位置124处的氨基酸用K替代(编号方式依照KabatEU索引)。在一个方面,在第二轻链的恒定域CH1中位置147和213处的氨基酸用E替代(编号方式依照Kabat的EU索引)。在一个方面,在第一轻链的恒定域CL中位置124和123处的氨基酸用K替代,且在第一重链的恒定域CH1中位置147和213处的氨基酸用E替代(编号方式依照KabatEU索引)。在一个方面,在第一轻链的恒定域CL中位置123处的氨基酸用R替代且位置124处的氨基酸用K替代,且在第一重链的恒定域CH1中位置147和213处的氨基酸均用E替代(编号方式依照KabatEU索引)。在一个方面,在第二重链的恒定域CL中位置124和123处的氨基酸用K替代,且其中在第二轻链的恒定域CH1中位置147和213处的氨基酸用E替代,且在第一轻链的可变域VL中位置38处的氨基酸用K替代,在第一重链的可变域VH中位置39处的氨基酸用E替代,在第二重链的可变域VL中位置38处的氨基酸用K替代,且在第二轻链的可变域VH中位置39处的氨基酸用E替代(编号方式依照KabatEU索引)。在一个方面,该双特异性抗体是一种二价抗体,其包含a)特异性结合第一抗原的抗体的第一轻链和第一重链,和b)特异性结合第二抗原的抗体的第二轻链和第二重链,其中该第二轻链和该第二重链的可变域VL和VH彼此替换,且其中该第二轻链和该第二重链的恒定域CL和CH1彼此替换。a)下的抗体不含b)下报告的修饰且a)下的重链和轻链是分离的链。在b)下的抗体中在轻链中可变轻链域VL用所述抗体的可变重链域VH替换,且恒定轻链域CL用所述抗体的恒定重链域CH1替换;且在重链内可变重链域VH用所述抗体的可变轻链域VL替换,且恒定重链域CH1用所述抗体的恒定轻链域CL替换。在一个方面,该双特异性抗体是一种二价抗体,其包含a)特异性结合第一抗原的抗体的第一轻链和第一重链,和b)特异性结合第二抗原的抗体的第二轻链和第二重链,其中该第二轻链和该第二重链的恒定域CL和CH1彼此替换。a)下的抗体不含b)下的报告的修饰且a)下的重链和轻链是分离的链。在b)下的抗体中在轻链内恒定轻链域CL用所述抗体的恒定重链域CH1替换;且在重链内恒定重链域CH1用所述抗体的恒定轻链域CL替换。在一个方面,该多特异性抗体是一种多特异性抗体,其包含a)特异性结合第一抗原且由两条抗体重链和两条抗体轻链组成的全长抗体,和b)特异性结合一种至四种别的抗原(即第二和/或第三和/或第四和/或第五抗原,优选特异性结合一种别的抗原,即第二抗原)的一个,两个,三个或四个单链Fab片段,其中所述b)下的单链Fab片段在所述全长抗体的重或轻链的C或N端经由肽接头融合至所述a)下的全长抗体。在一个方面,一个或两个相同的结合第二抗原的单链Fab片段在所述全长抗体的重或轻链的C端经由肽接头融合至该全长抗体。在一个方面,一个或两个相同的结合第二抗原的单链Fab片段在所述全长抗体的重链的C端经由肽接头融合至该全长抗体。在一个方面,一个或两个相同的结合第二抗原的单链Fab片段在所述全长抗体的轻链的C端经由肽接头融合至该全长抗体。在一个方面,两个相同的结合第二抗原的单链Fab片段在所述全长抗体的每一条重或轻链的C端经由肽接头融合至该全长抗体。在一个方面,两个相同的结合第二抗原的单链Fab片段在所述全长抗体的每一条重链的C端经由肽接头融合至该全长抗体。在一个方面,两个相同的结合第二抗原的单链Fab片段在所述全长抗体的每一条轻链的C端经由肽接头融合至该全长抗体。在一个方面,该双特异性抗体是一种三价抗体,其包含a)特异性结合第一抗原且由两条抗体重链和两条抗体轻链组成的全长抗体,b)由下述组成的第一多肽:ba)抗体重链可变域(VH),或bb)抗体重链可变域(VH)和抗体恒定域1(CH1),其中所述第一多肽以它的VH域的N端经由肽接头融合至所述全长抗体的两条重链之一的C端,c)由下述组成的第二多肽:ca)抗体轻链可变域(VL),或cb)抗体轻链可变域(VL)和抗体轻链恒定域(CL),其中所述第二多肽以VL域的N端经由肽接头融合至所述全长抗体的两条重链之另一的C端,且其中该第一多肽的抗体重链可变域(VH)和该第二多肽的抗体轻链可变域(VL)一起形成特异性结合第二抗原的抗原结合位点。在一个方面,通过在下述位置之间引入二硫键,b)下的多肽的抗体重链可变域(VH)和c)下的多肽的抗体轻链可变域(VL)经由链间二硫桥连接和稳定化:(i)重链可变域位置44对轻链可变域位置100,或(ii)重链可变域位置105对轻链可变域位置43,或(iii)重链可变域位置101对轻链可变域位置100(编号方式始终依照KabatEU索引)。为了稳定化而引入非天然二硫桥的技术记载于例如WO94/029350;Rajagopal,V.,etal.,Prot.Eng.(1997)1453-1459;Kobayashi,H.,etal.,Nucl.Med.Biol.25(1998)387-393;和Schmidt,M.,etal.,Oncogene18(1999)1711-1721。在一个实施方案中,任选的b)和c)下的多肽的可变域之间的二硫键在重链可变域位置44和轻链可变域位置100之间。在一个实施方案中,该任选的b)和c)下的多肽的可变域之间的二硫键在重链可变域位置105和轻链可变域位置43之间(编号方式始终依照Kabat)。在一个实施方案中,没有所述任选的单链Fab片段的可变域VH和VL之间的二硫化物稳定化的三价,双特异性抗体是优选的。在一个方面,该双特异性抗体是一种三特异性或四特异性抗体,其包含a)特异性结合第一抗原的全长抗体的第一轻链和第一重链,和b)特异性结合第二抗原的全长抗体的第二(经修饰)轻链和第二(经修饰)重链,其中可变域VL和VH彼此替换,和/或其中恒定域CL和CH1彼此替换,且c)其中特异性结合一种或两种别的抗原(即第三和/或第四抗原)的一个至四个抗原结合肽经由肽接头融合至a)和/或b)的轻链或重链的C或N端。a)下的抗体不含b)下报告的修饰且a)下的重链和该轻链是分离的链。在一个方面,该三特异性或四特异性抗体在c)下包含特异性结合一种或两种别的抗原的一个或两个抗原结合肽。在一个方面,该抗原结合肽选自scFv片段和scFab片段的组。在一个方面,该抗原结合肽是scFv片段。在一个方面,该抗原结合肽是scFab片段。在一个方面,该抗原结合肽融合至a)和/或b)的重链的C端。在一个方面,该三特异性或四特异性抗体在c)下包含特异性结合一种别的抗原的一个或两个抗原结合肽。在一个方面,该三特异性或四特异性抗体在c)下包含两个相同的特异性结合第三抗原的抗原结合肽。在一个优选的实施方案中,此类两个相同的抗原结合肽均经由相同的肽接头融合至a)和b)的重链的C端。在一个优选的实施方案中,该两个相同的抗原结合肽任一是scFv片段或scFab片段。在一个方面,该三特异性或四特异性抗体在c)下包含特异性结合第三和第四抗原的两个抗原结合肽。在一个实施方案中,所述两个抗原结合肽均经由相同的肽连接头融合至a)和b)的重链的C端。在一个优选的实施方案中,所述两个抗原结合肽任一是scFv片段或scFab片段。在一个方面,该双特异性抗体是一种双特异性,四价抗体,其包含a)特异性结合第一抗原(且包含两个Fab片段)的抗体的两条轻链和两条重链,b)特异性结合第二抗原的抗体的两个另外的Fab片段,其中所述另外的Fab片段均经由肽接头融合至a)的重链的C或N端任一,且其中在该Fab片段中实施下述修饰:(i)在a)的两个Fab片段中,或在b)的两个Fab片段中,可变域VL和VH彼此替换,和/或恒定域CL和CH1彼此替换,或(ii)在a)的两个Fab片段中可变域VL和VH彼此替换,且恒定域CL和CH1彼此替换,且在b)的两个Fab片段中可变域VL和VH彼此替换,或恒定域CL和CH1彼此替换,或(iii)在a)的两个Fab片段中可变域VL和VH彼此替换,或恒定域CL和CH1彼此替换,且在b)的两个Fab片段中可变域VL和VH彼此替换,且恒定域CL和CH1彼此替换,或(iv)在a)的两个Fab片段中可变域VL和VH彼此替换,且在b)的两个Fab片段中恒定域CL和CH1彼此替换,或(v)在a)的两个Fab片段中恒定域CL和CH1彼此替换,且在b)的两个Fab片段中可变域VL和VH彼此替换。在一个方面,所述另外的Fab片段均经由肽接头融合至a)的重链的C端或a)的重链的N端任一。在一个方面,所述另外的Fab片段均经由肽接头融合至a)的重链的C端任一。在一个方面,所述另外的Fab片段均经由肽连接头融合至a)的重链的N端。在一个方面,在该Fab片段中实施下述修饰:在a)的两个Fab片段中,或在b)的两个Fab片段中,可变域VL和VH彼此替换,和/或恒定域CL和CH1彼此替换。在一个方面,该双特异性抗体是一种四价抗体,其包含:a)特异性结合第一抗原且包含第一VH-CH1域对的第一抗体的(经修饰)重链,其中所述第一抗体的第二VH-CH1域对的N端经由肽接头融合至所述重链的C端,b)所述a)的第一抗体的两条轻链,c)特异性结合第二抗原且包含第一VH-CL域对的第二抗体的(经修饰)重链,其中所述第二抗体的第二VH-CL域对的N端经由肽接头融合至所述重链的C端,和d)所述c)的第二抗体的两条(经修饰)轻链,每一条包含CL-CH1域对。在一个方面,该双特异性抗体包含:a)特异性结合第一抗原的第一全长抗体的重链和轻链,和b)特异性结合第二抗原的第二全长抗体的重链和轻链,其中该重链的N端经由肽接头连接至该轻链的C端。a)下的抗体不含b)下报告的修饰且该重链和该轻链是分离的链。在一个方面,该双特异性抗体包含a)特异性结合第一抗原且由两条抗体重链和两条抗体轻链组成的全长抗体,和b)特异性结合第二抗原且包含VH2域和VL2域的Fv片段,其中两个域经由二硫桥彼此连接,其中只有该VH2域或该VL2域任一经由肽接头融合至该特异性结合第一抗原的全长抗体的重或轻链。在该双特异性抗体中a)下的重链和轻链是分离的链。在一个方面,该VH2域或该VL2域之另一并未经由肽接头融合至该特异性结合第一抗原的全长抗体的重或轻链。在本文中报告的所有方面,第一轻链包含VL域和CL域且第一重链包含VH域,CH1域,铰链区,CH2域和CH3域。在一个方面,该双特异性抗体是一种三价抗体,其包含a)特异性结合第一抗原的两个Fab片段,b)特异性结合第二抗原的一个CrossFab片段,其中CH1和CL域彼此交换,c)包含第一Fc区重链和第二Fc区重链的一个Fc区,其中该两个Fab片段的CH1域的C端连接至该重链Fc区多肽的N端,且其中该CrossFab片段的CL域的C端连接至该Fab片段之一的VH域的N端。在一个方面,该双特异性抗体是一种三价抗体,其包含a)特异性结合第一抗原的两个Fab片段,b)特异性结合第二抗原的一个CrossFab片段,其中CH1和CL域彼此交换,c)包含第一Fc区重链和第二Fc区重链的一个Fc区,其中第一Fab片段的CH1域的C端连接至该重链Fc区多肽之一的N端且该CrossFab片段的CL域的C端连接至另一条重链Fc区多肽的N端,且其中该第二Fab片段的CH1域的C端连接至该第一Fab片段的VH域的N端或该CrossFab片段的VH域的N端。在一个方面,该双特异性抗体包含a)特异性结合第一抗原且由两条抗体重链和两条抗体轻链组成的全长抗体,和b)特异性结合第二抗原且包含VH2域和VL2域包含重链片段和轻链片段的Fab片段,其中在该轻链片段中该可变轻链域VL2用所述抗体的可变重链域VH2替换,且在该重链片段中该可变重链域VH2用所述抗体的可变轻链域VL2替换,其中该重链Fab片段插入该全长抗体的重链之一的CH1域和该全长抗体的各自Fc区之间,且该轻链Fab片段的N端连接至该全长抗体中与该全长抗体中插入有该重链Fab片段的重链配对的轻链的C端。在一个方面,该双特异性抗体包含a)特异性结合第一抗原且由两条抗体重链和两条抗体轻链组成的全长抗体,和b)特异性结合第二抗原且包含VH2域和VL2域包含重链片段和轻链片段的Fab片段,其中在该轻链片段内该可变轻链域VL2用所述抗体的可变重链域VH2替换,且在该重链片段内该可变重链域VH2用所述抗体的可变轻链域VL2替换且其中该Fab片段的重链片段的C端连接至该全长抗体的重链之一的N端且该Fab片段的该轻链片段的C端连接至该全长抗体中与该全长抗体中缀合有该Fab片段的重链片段的重链配对的轻链的N端。多核苷酸本发明进一步提供了编码如本文中描述的双特异性抗体或其片段的分离的多核苷酸。在某些实施方案中,该多核苷酸或核酸是DNA。在其它实施方案中,本发明的多核苷酸是RNA,例如处于信使RNA(mRNA)的形式。本发明的RNA可以是单链或双链的。B.重组方法可使用重组方法和组合物来生成本文中提供的双特异性抗体,例如如记载于美国专利No.4,816,567的。在一个方面,提供编码本文所述双特异性抗体的分离的核酸。此类核酸可编码分别包含特异性结合PD1和TIM-3的抗原结合位点的VL的氨基酸序列和/或包含特异性结合PD1和TIM-3的抗原结合位点的VH的氨基酸序列(例如在抗体的轻和/或重链中)。在又一个方面,提供包含此类核酸的一种或多种载体(例如表达载体)。在又一个方面,提供包含此类核酸的宿主细胞。在一个此类方面,宿主细胞包含(例如已经用下述各项转化):(1)包含核酸的载体,所述核酸编码包含抗体VL的氨基酸序列和包含抗体VH的氨基酸序列,或(2)第一载体和第二载体,所述第一载体包含编码包含抗体VL的氨基酸序列的核酸,所述第二载体包含编码包含抗体VH的氨基酸序列的核酸。在一个方面,宿主细胞是真核的,例如中国仓鼠卵巢(CHO)细胞或淋巴样细胞(例如Y0,NS0,Sp20细胞)。在一个方面,提供生成双特异性抗体的方法,其中该方法包括在适合于抗体表达的条件下培养包含编码抗体的核酸的宿主细胞,如上文提供的,并任选自宿主细胞(或宿主细胞培养液)回收抗体。对于如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体的重组生成,分离编码双特异性抗体的核酸(例如如上文所描述的),并插入一种或多种载体中,用于在宿主细胞中进一步克隆和/或表达。可使用常规规程将此类核酸容易地分离并测序(例如通过使用寡核苷酸探针来进行,所述寡核苷酸探针能够特异性结合编码抗体重和轻链的基因)。适合于克隆或表达编码抗体的载体的宿主细胞包括本文所述原核或真核细胞。例如,可以在细菌中生成抗体,特别是在不需要糖基化和Fc效应器功能时。对于抗体片段和多肽在细菌中的表达,参见例如US5,648,237;US5,789,199;及US5,840,523。还可参见Charlton,K.A.,In:MethodsinMolecularBiology,Vol.248,Lo,B.K.C.(ed.),HumanaPress,Totowa,NJ(2003),pp.245-254,其描述抗体片段在大肠杆菌中的表达。表达后,可以在可溶性级分中自细菌细胞浆分离抗体,并可以进一步纯化。在原核生物之外,真核微生物诸如丝状真菌或酵母也是适合于编码抗体的载体的克隆或表达宿主,包括其糖基化途径已经“人源化”,导致生成具有部分或完全人的糖基化样式的抗体的真菌和酵母菌株。参见Gerngross,T.U.,Nat.Biotech.22:1409-1414(2004);及Li,H.etal.,Nat.Biotech.24:210-215(2006)。适合于表达糖基化抗体的宿主细胞也是自多细胞生物体(无脊椎动物和脊椎动物)衍生的。无脊椎动物细胞的例子包括植物和昆虫细胞。已经鉴定出许多杆状病毒株,其可以与昆虫细胞联合使用,特别是用于转染草地夜蛾(Spodopterafrugiperda)细胞。也可利用植物细胞培养物作为宿主。参见例如美国专利No.5,959,177;No.6,040,498;No.6,420,548;No.7,125,978;及No.6,417,429(其描述用于在转基因植物中生成抗体的PLANTIBODIESTM技术)。也可使用脊椎动物细胞作为宿主。例如,适应悬浮生长的哺乳动物细胞系可能是有用的。有用哺乳动物宿主细胞系的其它例子有经SV40转化的猴肾CV1系(COS-7);人胚肾系(293或293细胞,如记载于例如Graham,F.L.etal.,J.GenVirol.36:59-74(1977));幼仓鼠肾细胞(BHK);小鼠塞托利(sertoli)细胞(TM4细胞,如记载于例如Mather,J.P.,Biol.Reprod.23:243-252(1980));猴肾细胞(CV1);非洲绿猴肾细胞(VERO-76);人宫颈癌细胞(HELA);犬肾细胞(MDCK);牛鼠(buffalorat)肝细胞(BRL3A);人肺细胞(W138);人肝细胞(HepG2);小鼠乳房肿瘤(MMT060562);TRI细胞,如记载于例如Mather,J.P.etal.,AnnalsN.Y.Acad.Sci.383:44-68(1982);MRC5细胞;和FS4细胞。其它有用哺乳动物宿主细胞系包括中国仓鼠卵巢(CHO)细胞,包括DHFR-CHO细胞(Urlaub,G.etal.,Proc.Natl.Acad.Sci.USA77:4216-4220(1980));和骨髓瘤细胞系诸如Y0,NS0和Sp2/0。关于适合于抗体生成的某些哺乳动物宿主细胞系的综述参见例如Yazaki,P.andWu,A.M.,MethodsinMolecularBiology,Vol.248,Lo,B.K.C.(ed.),HumanaPress,Totowa,NJ(2004),pp.255-268。C.测定法可以通过本领域已知的各种测定法来鉴定,筛选或通过其物理/化学性质和/或生物学活性来表征在本文中提供的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体。1.亲和力测定法根据实施例中提出的通过表面等离子体共振(SPR)的方法,使用标准仪器如仪器(GEHealthcare),以及可以通过诸如重组表达获得的受体或靶蛋白,可以测定在本文中提供的双特异性抗原结合分子,抗体和抗体片段与相应的抗原的亲和力。在实施例1b,5或12中描述了用于测量结合亲和力的具体的说明性和示例性实施方案。根据一个方面,使用T100仪(GEHealthcare)通过表面等离子体共振在25℃测量KD。2.结合测定法和其它测定法在一个方面,对本发明的抗体测试它的抗原结合活性,例如通过已知方法诸如ELISA,Western印迹,等。本文中提供的双特异性抗体对相应的重组抗原或表达抗原的细胞的结合可以通过实施例12中描述的ELISA来评估。在另一个方面,本发明提供一种基于细胞的TR-FRET测定法,其用于测定双特异性抗体格式对一种细胞上存在的两种不同受体的同时结合。所选择的Tag-lite技术是经典的TR-FRET(时间解析荧光共振能量转移)和SNAP-tag技术(例如NewEnglandBiolabs,CISBIO)的组合,其容许用荧光供体或受体染料标记细胞表面上存在的抗原。该测定法在实施例13中描述。在又一个方面,在结合测定法中使用新鲜的外周血单个核细胞(PBMC)来显示对不同外周血单个核细胞(PBMC)诸如单核细胞,NK细胞和T细胞的结合。在另一个方面,可以使用竞争测定法来鉴定分别与特异性抗体或抗原结合位点竞争对靶物的结合的抗体。在某些实施方案中,此类竞争性抗体结合依照本发明的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体所结合的相同表位(例如线性或构象表位)。用于定位抗体所结合的表位的详细的例示性方法在Morris(1996)“EpitopeMappingProtocols,”inMethodsinMolecularBiologyvol.66(HumanaPress,Totowa,NJ)中提供。在一种例示性竞争测定法中,在包含第一经标记抗体(其结合PD1或TIM3)和第二未标记抗体(其要测试与第一抗体竞争对PD1或TIM3的结合的能力)的溶液中温育固定化的PD1或TIM3。第二抗体可存在于杂交瘤上清液中。作为对照,在包含该第一经标记抗体但不包含第二未标记抗体的溶液中温育固定化的PD1或TIM3。在允许第一抗体结合PD1或TIM3的条件下温育后,去除过量的未结合抗体,并测量与固定化的PD1或TIM3联合的标记物的量。如果测试样品中与固定化的PD1或TIM3联合的标记物的量相对于对照样品实质性降低,那么这指示第二抗体与第一抗体竞争对PD1或TIM3的结合。参见HarlowandLane(1988)Antibodies:ALaboratoryManualch.14(ColdSpringHarborLaboratory,ColdSpringHarbor,NY)。3.活性测定法在一个方面,提供了用于鉴定具有生物学活性的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体的测定法。生物学活性可以包括例如增强不同免疫细胞,尤其是T细胞的活化和/或增殖,免疫调控性细胞因子诸如IFNγ或TNF-α的分泌,阻断PD1途径,阻断TIM3途径,杀伤肿瘤细胞的能力。还提供了在体内和/或在体外具有此类生物学活性的抗体。在某些方面,对本发明的抗体测试此类生物学活性。在一个方面,提供的是一种免疫细胞测定法,其测量来自一个个体(供体X)的淋巴细胞对来自另一个个体(供体Y)的淋巴细胞的活化。混合淋巴细胞反应(MLR)能证明阻断PD1途径对淋巴细胞效应细胞的影响。在本发明的双特异性抗体存在或缺失下对该测定法中的T细胞测试活化和它们的IFN-γ分泌。该测定法在实施例16中更加详细的描述。D.免疫缀合物本发明还提供包含本发明的双特异性抗体的免疫缀合物,该抗体与一种或多种细胞毒剂诸如化疗剂或药物,生长抑制剂,毒素(例如蛋白质毒素,细菌,真菌,植物,或动物起源的酶活性毒素,或其片段),或放射性同位素缀合。在一个方面,免疫缀合物是抗体-药物缀合物(ADC),其中抗体与一种或多种药物缀合,包括但不限于美登木生物碱(maytansinoid)(参见美国专利No.5,208,020,5,416,064和欧洲专利EP0425235B1);auristatin诸如单甲基auristatin药物模块DE和DF(MMAE和MMAF)(参见美国专利No.5,635,483和5,780,588,和7,498,298);多拉司他汀(dolastatin);加利车霉素(calicheamicin)或其衍生物(参见美国专利No.5,712,374,5,714,586,5,739,116,5,767,285,5,770,701,5,770,710,5,773,001,和5,877,296;Hinman等,CancerRes.53:3336-3342(1993);和Lode等,CancerRes.58:2925-2928(1998));蒽环类抗生素诸如道诺霉素(daunomycin)或多柔比星(doxorubicin)(参见Kratz等,CurrentMed.Chem.13:477-523(2006);Jeffrey等,Bioorganic&Med.Chem.Letters16:358-362(2006);Torgov等,Bioconj.Chem.16:717-721(2005);Nagy等,Proc.Natl.Acad.Sci.USA97:829-834(2000);Dubowchik等,Bioorg.&Med.Chem.Letters12:1529-1532(2002);King等,J.Med.Chem.45:4336-4343(2002);和美国专利No.6,630,579);甲氨蝶呤;长春地辛(vindesine);紫杉烷诸如多西他赛(docetaxel),帕利他赛(paclitaxel),larotaxel,tesetaxel,和ortataxel;单端孢菌素(trichothecene);和CC1065。在另一个实施方案中,免疫缀合物包含本文所述抗体,该抗体与酶活性毒素或其片段缀合,包括但不限于白喉毒素A链,白喉毒素的非结合活性片段,外毒素A链(来自铜绿假单胞菌Pseudomonasaeruginosa),蓖麻毒蛋白(ricin)A链,相思豆毒蛋白(abrin)A链,蒴莲根毒蛋白(modeccin)A链,α-帚曲霉素(sarcin),油桐(Aleuritesfordii)毒蛋白,香石竹(dianthin)毒蛋白,美洲商陆(Phytolacaamericana)毒蛋白(PAPI,PAPII和PAP-S),苦瓜(Momordicacharantia)抑制物,麻疯树毒蛋白(curcin),巴豆毒蛋白(crotin),肥皂草(sapaonariaofficinalis)抑制物,白树毒蛋白(gelonin),丝林霉素(mitogellin),局限曲菌素(restrictocin),酚霉素(phenomycin),依诺霉素(enomycin)和单端孢菌素(tricothecenes)。在另一个实施方案中,免疫缀合物包含本文所述抗体,该抗体与放射性原子缀合以形成放射缀合物。多种放射性同位素可用于生成放射缀合物。实例包括At211,I131,I125,Y90,Re186,Re188,Sm153,Bi212,P32,Pb212和Lu的放射性同位素。在将放射缀合物用于检测时,它可包含放射性原子用于闪烁照相研究,例如Tc99m或I123,或自旋标记物用于核磁共振(NMR)成像(也称为磁共振成像,mri),诸如再一次的碘-123,碘-131,铟-111,氟-19,碳-13,氮-15,氧-17,钆,锰或铁。可使用多种双功能蛋白质偶联剂来制备抗体和细胞毒剂的缀合物,诸如N-琥珀酰亚氨基-3-(2-吡啶基二硫代)丙酸酯(SPDP),琥珀酰亚氨基-4-(N-马来酰亚氨基甲基)环己烷-1-羧酸酯(SMCC),亚氨基硫烷(IT),亚氨酸酯(诸如盐酸己二酰亚氨酸二甲酯),活性酯类(诸如辛二酸二琥珀酰亚氨基酯),醛类(诸如戊二醛),双叠氮化合物(诸如双(对-叠氮苯甲酰基)己二胺),双重氮衍生物(诸如双(对-重氮苯甲酰基)乙二胺),二异氰酸酯(诸如甲苯2,6-二异氰酸酯),和双活性氟化合物(诸如1,5-二氟-2,4-二硝基苯)的双功能衍生物。例如,可如Vitetta等,Science238:1098(1987)中所述制备蓖麻毒蛋白免疫毒素。碳-14标记的1-异硫氰酸苯甲基-3-甲基二亚乙基三胺五乙酸(MX-DTPA)是用于将放射性核苷酸与抗体缀合的例示性螯合剂。参见WO94/11026。接头可以是便于在细胞中释放细胞毒药物的“可切割接头”。例如,可使用酸不稳定接头,肽酶敏感接头,光不稳定接头,二甲基接头,或含二硫化物接头(Chari等,CancerRes.52:127-131(1992);美国专利No.5,208,020)。本文中的免疫缀合物或ADC明确涵盖但不限于用下列交联剂制备的此类缀合物,包括但不限于:商品化(如购自PierceBiotechnologyInc.,Rockford,IL,U.S.A)的BMPS,EMCS,GMBS,HBVS,LC-SMCC,MBS,MPBH,SBAP,SIA,SIAB,SMCC,SMPB,SMPH,sulfo-EMCS,sulfo-GMBS,sulfo-KMUS,sulfo-MBS,sulfo-SIAB,sulfo-SMCC,和sulfo-SMPB,和SVSB(琥珀酰亚胺基-(4-乙烯基砜)苯甲酸酯)。E.用于诊断和检测的方法和组合物在某些方面,本文中提供的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的任何双特异性抗体对于检测生物学样品中PD1和TIM3二者的存在可能是有用的。如本文中使用的,术语“检测”涵盖定量或定性检测。在某些实施方案中,生物学样品包括细胞或组织,诸如AML干癌细胞。在一个方面,提供在诊断或检测方法中使用的双特异性抗体。在又一方面,提供检测生物学样品中PD1和TIM3二者的存在的方法。在某些实施方案中,该方法包括在允许双特异性抗体结合PD1和TIM3二者的条件下使生物学样品与本文所述双特异性抗体接触,并检测是否在双特异性抗体与两种抗原之间形成复合物。此类方法可以是体外或体内方法。在一个实施方案中,使用双特异性抗体来选择适合用包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体治疗的受试者,例如其中PD1和TIM3是用于选择患者的生物标志物。在某些方面,提供经标记的双特异性抗体。标记物包括但不限于直接检测的标记物或模块(诸如荧光,发色,电子致密,化学发光,和放射性标记物),以及例如经由酶促反应或分子相互作用间接检测的模块,诸如酶或配体。例示性标记物包括但不限于放射性同位素32P,14C,125I,3H,和131I,荧光团诸如稀土螯合物或荧光素及其衍生物,罗丹明(rhodamine)及其衍生物,丹酰,伞形酮,萤光素酶例如萤火虫萤光素酶和细菌萤光素酶(美国专利No.4,737,456),萤光素,2,3-二氢酞嗪二酮,辣根过氧化物酶(HRP),碱性磷酸酶,β-半乳糖苷酶,葡糖淀粉酶,溶菌酶,糖氧化酶例如葡萄糖氧化酶,半乳糖氧化酶,和葡萄糖-6-磷酸脱氢酶,杂环氧化酶诸如尿酸酶和黄嘌呤氧化酶(其与采用过氧化氢氧化染料前体的酶诸如HRP偶联),乳过氧化物酶,或微过氧化物酶,生物素/亲合素,自旋标记物,噬菌体标记物,或稳定的自由基。F.药物组合物,配制剂和给药途径在又一个方面,本发明提供了包含本文中提供的任一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体的药物组合物,以供例如在任何下列治疗方法中使用。在一个实施方案中,药物组合物包含本文中提供的任一种双特异性抗体和至少一种药学上可接受的赋形剂。在另一个实施方案中,药物组合物包含本文中提供的任一种双特异性抗体和至少一种另外的例如如下文所述的治疗剂。本发明的药物组合物包含治疗有效量的一种或多种溶解或分散在药学上可接受的赋形剂中的双特异性抗体。短语“药学的或药学上可接受的”是指在采用的剂量和浓度下通常对接受者无毒的分子实体和组合物,即在适当施用于动物时例如人时不产生不利的,过敏的或其他不良的反应。根据本公开内容,包含至少一种双特异性抗体和任选地另外的活性成分的药物组合物的制备将是本领域技术人员已知的,如Remington'sPharmaceuticalSciences第18版,MackPrintingCompany,1990所例示,通过提述将其并入本文。具体地说,组合物是冻干配制剂或水溶液。如本文中使用的,正如本领域普通技术人员已知,“药学上可接受的赋形剂”包括溶剂,缓冲剂,分散介质,包衣,表面活性剂,抗氧化剂,防腐剂(例如抗菌剂,抗真菌剂),等渗剂,盐,稳定剂中任一种和全部及其组合。肠胃外组合物包括为通过注射给药而设计的那些,例如皮下,皮内,病灶内,静脉内,动脉内,肌内,鞘内或腹膜内注射。为了注射,可以将本发明的双特异性抗体配制在水溶液中,优选在生理学相容的缓冲液如汉克斯溶液,林格氏溶液或生理盐水缓冲液中。溶液可以含有配制剂,例如悬浮剂,稳定剂和/或分散剂。或者,双特异性抗体在使用前可处于粉末形式以与适合的载体(例如无菌无热原水)组构。根据需要,通过将在适当溶剂中的本发明的融合蛋白以所要求的量并入下文列举的多种其他成分中来制备无菌注射溶液。可以例如通过无菌过滤膜进行过滤而容易地实现无菌性。通常,通过将各种灭菌的活性成分并入无菌载体中来制备分散体,所述无菌载体含有基础分散介质和/或其他成分。在用于制备无菌注射溶液,悬浮液或乳状液的无菌粉末的情况下,优选制备方法为真空干燥和冷冻干燥技术,从而产生来自其先前无菌过滤的液体介质的活性成分加上任何另外的期望的成分的粉末。必要时应该适当缓冲该液体介质并且在注射之前首先用足够盐水或葡萄糖使液体稀释剂变得等张。在制造和储存条件下该组合物必须是稳定的,并且保持对抗微生物诸如细菌和真菌的污染作用。应当理解,内毒素污染应该最低限度地保持在安全水平,例如低于0.5ng/mg蛋白质。适合的药学上可接受的赋形剂包括但不限于:诸如磷酸盐,柠檬酸盐,和其他有机酸的缓冲剂;包括抗坏血酸和甲硫氨酸在内的抗氧化剂;防腐剂(如十八烷基二甲基苄基氯化铵;氯化六甲双铵;苯扎氯铵,苄索氯铵;苯酚,丁醇或苄醇;烷基对羟基苯甲酸酯,如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯;儿茶酚;间苯二酚;环己醇;3-戊醇;和间甲酚);低分子量(少于约10个残基)多肽;蛋白质,如血清白蛋白,明胶,或免疫球蛋白;亲水性聚合物,如聚乙烯吡咯烷酮;氨基酸,如甘氨酸,谷氨酰胺,天冬酰胺,组氨酸,精氨酸,或赖氨酸;单糖类,二糖类,和其他碳水化合物类,包括葡萄糖,甘露糖,或糊精;螯合剂,如EDTA;糖类,如蔗糖,甘露醇,海藻糖或山梨糖醇;成盐反离子,如钠;金属复合物(例如,Zn-蛋白质复合物);和/或非离子型表面活性剂,如聚乙二醇(PEG)。水性注射悬液可含有增加该悬液粘度的化合物,例如羧甲基纤维素钠,山梨糖醇,或葡聚糖等。任选地,悬液还可以含有适合的稳定剂或增加化合物的溶解度以允许制备高度浓缩的溶液的药剂。另外,这些活性化合物的悬液可制备为适当的油性注射悬液。适合的亲脂性溶剂或载体包括脂肪油(如芝麻油),合成脂肪酸酯(如油酸乙酯或甘油三酯),或脂质体。活性成分可以包埋在例如通过凝聚技术或通过界面聚合制备的微胶囊中,例如分别是羟甲基纤维素或明胶微胶囊和聚甲基丙烯酸甲酯微胶囊,在胶体药物递送系统(例如,脂质体,白蛋白微球,微乳液,纳米颗粒和纳米胶囊)中或在粗乳液中。这样的技术公开于Remington'sPharmaceuticalSciences(18thEd.MackPrintingCompany,1990)中。可以制备持续释放配制剂。持续释放配制剂的适合实例包括含有该多肽的固体疏水聚合物的半透性基质,所述基质呈成形物品的形式,例如薄膜,或微胶囊。在特定实施方案中,通过在组合物中使用延迟吸收的药剂,例如单硬脂酸铝,明胶或其组合,可延长注射组合物的吸收。本文中的示例性药学上可接受的赋形剂还包括诸如可溶性中性活性透明质酸酶糖蛋白(sHASEGP)的间质药物分散剂,例如人可溶性PH-20透明质酸酶糖蛋白,例如rHuPH20(BaxterInternational,Inc.)。在美国专利公开No.2005/0260186和2006/0104968中描述了包括rHuPH20的某些示例性的sHASEGP和使用方法。在一个方面,将sHASEGP与一种或多种另外的糖胺聚糖酶如软骨素酶组合。示例性的冻干抗体配制剂描述于美国专利No.6,267,958中。水性抗体配制剂包括美国专利No.6,171,586和WO2006/044908中描述的那些,后一种配制剂包括组氨酸-乙酸盐缓冲液。除前述组合物之外,双特异性抗体还可配制为贮库配制剂。此类长效配制剂可通过植入(例如皮下或肌内)或通过肌内注射给予。因此,例如,双特异性抗体可以用适合的聚合材料或疏水材料(例如作为可接受的油中的乳液)或离子交换树脂配制,或配制为微溶的衍生物,例如微溶盐。包含本发明的双特异性抗体的药物组合物可以通过常规的混合,溶解,乳化,包封,包埋或冻干过程进行制造。可以使用有利于将蛋白质加工成药学上可使用的配制剂的一种或多种生理学上可接受的载体,稀释剂,赋形剂或助剂以常规方式配制药物组合物。适当的配制剂取决于所选择的给药途径。可以将双特异性抗体以游离酸或碱,中性或盐形式配制成组合物。药学上可接受的盐是基本保留游离酸或碱的生物学活性的盐。这些盐包括酸加成盐,例如由蛋白质组合物的游离氨基形成的或与无机酸例如盐酸或磷酸形成的那些,或与诸如乙酸,草酸,酒石酸或扁桃酸的有机酸形成的那些。与游离羧基形成的盐也可以衍生自无机碱,例如氢氧化钠,氢氧化钾,氢氧化铵,氢氧化钙或氢氧化铁;或诸如异丙胺,三甲胺,组氨酸或普鲁卡因的有机碱。与相应的游离碱形式相比,药物盐在水性溶剂和其他质子溶剂中的溶解度更高。在被治疗的特殊适应症需要时,本文中的组合物还可含有一种以上的活性成分,优选地,所述活性成分具有彼此无不良影响的互补活性。这样的活性成分以对预期目的有效的量适当地组合存在。用于体内给药的配制剂通常是无菌的。可以例如通过无菌过滤膜进行过滤而容易地实现无菌性。G.治疗方法和组合物本文中提供的任何包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体均可在治疗方法中使用。为了在治疗方法中使用,如本文中之前定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体可以以符合良好医学实践的方式配制,给药和施用。在这种情况下考虑的因素包括正在治疗的具体病症,被治疗的具体哺乳动物,个体患者的临床状况,病症的原因,药剂的递送部位,给药方法,给药方案,以及执业医师已知的其他因素。在一个方面,提供了如本文中定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体,其用作药物。在又一些方面,提供了如本文中定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体,其用于治疗疾病,特别是用于治疗癌症。在某些实施方案中,提供了包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体,其用于治疗方法。在一个实施方案中,本发明提供如本文中描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体,其用于在有所需要的个体中治疗疾病。在某些实施方案中,本发明提供包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体,其用于治疗具有疾病的个体的方法,该方法包括对该个体施用治疗有效量的该双特异性抗体。在某些实施方案中,要治疗的疾病是癌症。在另一个方面,要治疗的疾病是慢性病毒感染,像HIV,HBV,HCV,HSV1,CMV,LCMV或EBV。需要治疗的受试者,患者,或“个体”通常是哺乳动物,更加具体地是人。在又一个方面,本发明提供如本文中之前定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体制造或制备用于在有所需要的个体中治疗疾病的药物中的用途。在一个实施方案中,该药物用于治疗疾病的方法,该方法包括对具有疾病的个体施用治疗有效量的该药物。在某些方面,要治疗的疾病是增殖性病症,特别是癌症。癌症的实例包括膀胱癌,脑癌,头和颈癌,胰腺癌,肺癌,乳腺癌,卵巢癌,子宫癌,子宫颈癌,子宫内膜癌,食道癌,结肠癌,结肠直肠癌,直肠癌,胃癌,前列腺癌,血癌,皮肤癌,鳞状细胞癌,骨癌,和肾癌。可以使用依照本发明的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体进行治疗的其它细胞增殖性病症包括但不限于位于腹部,骨骼,乳腺,消化系统,肝脏,胰腺,腹膜,内分泌腺(肾上腺,甲状旁腺,垂体,睾丸,卵巢,胸腺,甲状腺),眼,头和颈,神经系统(中枢和周围),淋巴系统,盆腔,皮肤,软组织,脾脏,胸部,和泌尿生殖系统的赘生物。还包括癌前状态或病变和癌症转移。在某些方面,癌症选自由肾细胞癌,皮肤癌,肺癌,结肠直肠癌,乳腺癌,脑癌,头和颈癌组成的组。在又一些方面,该癌症选自癌瘤,淋巴瘤(例如霍奇金(Hodgkin)氏和非霍奇金氏淋巴瘤),母细胞瘤,肉瘤和白血病。在另一个方面,要治疗的癌症选自鳞状细胞癌,小细胞肺癌,非小细胞肺癌,肺的腺癌,肺的鳞癌,腹膜癌,肝细胞癌,胃肠癌,胰腺癌,胶质瘤,宫颈癌,卵巢癌,肝癌,膀胱癌,肝瘤,乳腺癌,结肠癌,结肠直肠癌,子宫内膜癌或子宫癌,唾液腺癌,肾癌,肝癌,前列腺癌,外阴癌,甲状腺癌,肝的癌,白血病和其它淋巴增殖性病症,及各种类型的头和颈癌。在又一个方面,要治疗的疾病是慢性病毒感染。术语“慢性病毒感染”指受试者受到慢性病毒折磨或感染。慢性病毒感染的例子有人免疫缺陷病毒(HIV),乙型肝炎病毒感染(HBV),丙型肝炎病毒感染(HCV),单纯疱疹病毒1(HSV1),巨细胞病毒(CMV),淋巴细胞性脉络丛脑膜炎病毒(LCMV)或Epstein-Barr病毒(EBV)。技术人员容易认识到,在许多情况下,双特异性抗体可能不提供治愈,而可能仅提供部分益处。在一些实施方案中,具有一些益处的生理变化也被认为是治疗有益的。因此,在一些实施方案中,提供生理变化的双特异性抗体的量被认为是“有效量”或“治疗有效量”。在又一个方面,本发明提供了治疗个体中的疾病的方法,所述方法包括向所述个体施用治疗有效量的本发明的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体。在一个实施方案中,向所述个体施用呈药学上可接受的形式的包含本发明的双特异性抗体的组合物。在某些实施方案中,待治疗的疾病是增殖性病症。在一个特定的实施方案,该疾病是癌症。在某些实施方案中,该方法进一步包括向个体施用治疗有效量的至少一种另外的治疗剂,例如,如果待治疗的疾病是癌症,则为抗癌剂。在另一个方面,该疾病是慢性病毒感染。根据任何上述实施方案的“个体”可以是哺乳动物,优选人。为了预防或治疗疾病,本发明的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体的适当剂量(当单独使用或与一种或多种其他另外的治疗剂组合使用时)将取决于待治疗疾病的类型,给药途径,患者的体重,融合蛋白的类型,疾病的严重程度和病程,给予双特异性抗体是用于预防还是治疗目的,先前或同时的治疗干预,患者的临床病史,和对融合蛋白的反应,和主治医师的决定。在任何情况下,负责给药的医师将决定组合物中活性成分的浓度和个体受试者的适当剂量。本文中考虑到各种给药方案,包括但不限于在各个时间点上的单次或多次给药,推注给药和脉冲输注。一次性地或以一系列治疗的方式适当地向患者施用如本文中定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体。根据疾病的类型和严重程度,约1μg/kg至15mg/kg(例如0.1mg/kg-10mg/kg)的双特异性抗体可以是向患者施用的初始候选剂量,例如无论是通过一次或多次分开给药,还是通过连续输注。取决于上面提到的因素,一个典型的日剂量可能在约1μg/kg至100mg/kg或更高的范围内。对于数天或更长时间的反复给药,根据病情,治疗通常将持续到出现期望的疾病症状的抑制时为止。双特异性抗体的一个示例性剂量将在约0.005mg/kg至约10mg/kg的范围内。在其他实例中,剂量还可以包含每次给药从约1μg/kg体重,约5μg/kg体重,约10μg/kg体重,约50μg/kg体重,约100μg/kg体重,约200μg/kg体重,约350μg/kg体重,约500μg/kg体重,约1mg/kg体重,约5mg/kg体重,约10mg/kg体重,约50mg/kg体重,约100mg/kg体重,约200mg/kg体重,约350mg/kg体重,约500mg/kg体重,至约1000mg/kg体重或更多,以及其中可推导的任何范围。在本文中所列数字的可推导范围的实例中,可以基于上述数字以约5mg/kg体重至约100mg/kg体重,约5μg/kg体重至约500mg/kg体重等范围进行施用。因此,可向患者施用约0.5mg/kg,2.0mg/kg,5.0mg/kg或10mg/kg(或其任何组合)的一个或多个剂量。这样的剂量可以间歇施用,例如,每周或每三周一次(例如使得患者接受约2个至约20个,或例如约6个剂量的融合蛋白)。可以施用初始较高的负荷剂量,然后施用一个或多个较低的剂量。然而,其他剂量方案可能是有用的。通过常规技术和测定法易于监测这种疗法的进展。通常会以有效实现预期目的的量使用如本文中定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体。为了用于治疗或预防疾病状况,以治疗有效量施用或应用本发明的双特异性抗体或其药物组合物。治疗有效量的确定完全在本领域技术人员的能力范围内,尤其是在根据本文中提供的详细公开内容的情况下。对于全身给药,可以根据体外测定法(例如细胞培养物测定法)初步估计治疗有效剂量。然后可以在动物模型中配制剂量以达到包括在细胞培养中确定的IC50在内的循环浓度范围。这类信息可以用来更精确地确定用于人的剂量。也可以使用本领域熟知的技术根据体内数据(例如,动物模型)估计初始剂量。根据动物数据,本领域普通技术人员可以容易地优化对人的给药。可以个别调整剂量和间隔以提供足以维持治疗效果的双特异性抗体的血浆水平。通常注射给药的患者剂量为约0.1至50mg/kg/天,一般为约0.5至1mg/kg/天。可以通过每天施用多个剂量来实现治疗有效的血浆水平。可以例如通过HPLC测量血浆中的水平。在局部给药或选择性吸收的情况下,双特异性抗体的有效局部浓度可能与血浆浓度无关。本领域技术人员将能够优化治疗有效的局部剂量而无需过度实验。本文中描述的双特异性抗体的治疗有效剂量通常将提供治疗益处而不引起实质性毒性。可以通过细胞培养或实验动物中的标准药物程序来确定融合蛋白的毒性和治疗效果。可使用细胞培养物测定法和动物研究来确定LD50(对群体的50%致死的剂量)和ED50(在50%的群体中治疗有效的剂量)。毒性与疗效之间的剂量比为治疗指数,它可以被表示为比率LD50/ED50。表现出较大治疗指数的双特异性抗体是优选的。在一个实施方案中,依照本发明的双特异性抗体表现出高治疗指数。从细胞培养物测定法和动物研究中获得的数据可以在配制适合于在人类中使用的剂量范围中加以使用。剂量优选地处在循环浓度范围内,其包括具有很小或没有毒性的ED50。取决于多种因素,例如采用的剂型,所用给药途径,受试者的状况等,剂量可以在这个范围内变化。考虑到患者的状况,个体医师可以选择确切的配方,给药途径和剂量(参见,例如Fingl等人,1975,in:ThePharmacologicalBasisofTherapeutics,Ch.1,p.1,通过提述将其完整并入本文)。用本发明的双特异性抗体治疗患者的主治医师知道如何并且何时由于毒性,器官功能障碍等而终止,中断或调整给药。相反,如果临床反应不足(排除毒性),主治医师也会知道将治疗调整到更高水平。在感兴趣病症的管理中,施用剂量的大小将随着待治疗病症的严重程度,给药途径等而变化。可以例如部分地通过标准预后评价方法来评价病症的严重程度。此外,剂量和可能的剂量频率也将根据个体患者的年龄,体重和反应而变化。其它药剂和治疗如本文中之前描述的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体可以在治疗中与一种或多种其它药剂联合给药。例如,本发明的融合蛋白可以与至少一种另外的治疗剂共同给药。术语“治疗剂”涵盖可用于治疗需要这种治疗的个体中的症状或疾病的任何药剂。这种另外的治疗剂可包含适合于被治疗的特定适应症的任何活性成分,优选地,所述活性成分具有彼此无不良影响的互补活性。在某些实施方案中,另外的治疗剂是另一种抗癌剂。此类其它药剂以对预期目的有效的量适当地组合存在。此类其它药剂的有效量取决于所使用的融合蛋白的量,病症或治疗的类型,以及以上讨论的其它因素。通常以与本文中所述相同的剂量和给药途径,或本文中所述剂量的约1%至99%,或以在经验/临床上确定为适当的任何剂量并通过任何途径使用本发明的抗原结合分子。上述此类联合疗法涵盖联合给药(其中两种或更多种治疗剂包括在同一个或分开的组合物中)和分开给药,在分开给药的情况下,双特异性抗体的给药可以发生在给予另外的治疗剂和/或佐剂之前,同时和/或之后。H.制品在本发明的另一方面,提供一种制品,其包含对于治疗,预防和/或诊断上文所述病症有用的材料。制品包括容器和容器上或与容器联合的标签或包装插页。合适的容器包括例如瓶,管形瓶,注射器,IV溶液袋,等。容器可以自多种材料诸如玻璃或塑料形成。容器装有单独或与另一种组合物组合有效治疗,预防和/或诊断疾患的组合物,并且可具有无菌存取口(例如,容器可以是具有由皮下注射针可刺穿的塞子的管形瓶或静脉内溶液袋)。组合物中的至少一种活性剂是如本文中之前定义的包含特异性结合PD1的第一抗原结合位点和特异性结合TIM-3的第二抗原结合位点的双特异性抗体。标签或包装插页指示使用组合物来治疗选择的状况。此外,制品可包括(a)其中装有组合物的第一容器,其中该组合物包含本发明的双特异性抗体;和(b)其中装有组合物的第二容器,其中该组合物包含别的细胞毒性或其它方面治疗性的药剂。在本发明的这个实施方案中的制品可进一步包括包装插页,其指示可使用组合物来治疗特定状况。或者/另外,制品可进一步包括第二(或第三)容器,其装有药学可接受缓冲液,诸如抑菌性注射用水(BWFI),磷酸盐缓冲盐水,林格(Ringer)氏溶液和右旋糖溶液。它可进一步包括从商业和用户立场看期望的其它材料,包括其它缓冲液,稀释剂,滤器,针,和注射器。表C(序列):下面列出本发明的具体实施方案:1.一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含VH域,其包含(i)包含SEQIDNO:37的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:38的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:39的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:40的氨基酸序列的HVR-L1;(ii)包含SEQIDNO:41的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:42的氨基酸序列的HVR-L3;且所述特异性结合TIM3的第二抗原结合位点包含(a)VH域,其包含(i)包含SEQIDNO:1的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:2的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:3的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:4或SEQIDNO:11或SEQIDNO:12的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:5的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:6的氨基酸序列的HVR-L3;或(b)VH域,其包含(i)包含SEQIDNO:17的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:18的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:19的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:20的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:21的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:22的氨基酸序列的HVR-L3;或(c)VH域,其包含(i)包含SEQIDNO:29的氨基酸序列的HVR-H1,(ii)包含SEQIDNO:30的氨基酸序列的HVR-H2,和(iii)包含SEQIDNO:31的氨基酸序列的HVR-H3;和VL域,其包含(i)包含SEQIDNO:32的氨基酸序列的HVR-L1,(ii)包含SEQIDNO:33的氨基酸序列的HVR-L2,和(iii)包含SEQIDNO:34的氨基酸序列的HVR-L3。2.一种包含特异性结合PD1的第一抗原结合位点和特异性结合TIM3的第二抗原结合位点的双特异性抗体,其中该双特异性抗体以与对PD1的结合相比低至少50倍的结合亲和力,更加特别的是以与对PD1的结合相比低至少100倍的结合亲和力结合TIM3。3.如本文中之前定义的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含(a)包含SEQIDNO:43的氨基酸序列的VH域和包含SEQIDNO:44的氨基酸序列的VL域,或(b)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,或(c)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:47的氨基酸序列的VL域,或(d)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:48的氨基酸序列的VL域,或(e)包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:49的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含(a)包含SEQIDNO:7的氨基酸序列的VH域和包含SEQIDNO:8的氨基酸序列的VL域,或(b)包含SEQIDNO:9的氨基酸序列的VH域和包含SEQIDNO:10的氨基酸序列的VL域,或(c)包含SEQIDNO:13的氨基酸序列的VH域和包含SEQIDNO:14的氨基酸序列的VL域,或(d)包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域,或(e)包含SEQIDNO:23的氨基酸序列的VH域和包含SEQIDNO:24的氨基酸序列的VL域,或(f)包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域,或(g)包含SEQIDNO:27的氨基酸序列的VH域和包含SEQIDNO:28的氨基酸序列的VL域,或(h)包含SEQIDNO:35的氨基酸序列的VH域和包含SEQIDNO:36的氨基酸序列的VL域。4.如本文中之前定义的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,且所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:15的氨基酸序列的VH域和包含SEQIDNO:16的氨基酸序列的VL域或包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。5.如本文中之前定义的双特异性抗体,其中所述特异性结合PD1的第一抗原结合位点包含包含SEQIDNO:45的氨基酸序列的VH域和包含SEQIDNO:46的氨基酸序列的VL域,和所述特异性结合TIM3的第二抗原结合位点包含包含SEQIDNO:25的氨基酸序列的VH域和包含SEQIDNO:26的氨基酸序列的VL域。6.如本文中之前定义的双特异性抗体,其中该双特异性抗体是人,人源化或嵌合抗体。7.如本文中之前定义的双特异性抗体,其中该双特异性抗体包含Fc域,包含该特异性结合PD1的抗原结合位点的第一Fab片段和包含该特异性结合TIM3的抗原结合位点的第二Fab片段。8.如本文中之前定义的双特异性抗体,其中该Fc域是IgG,特别是IgG1Fc域或IgG4Fc域。9.如本文中之前定义的双特异性抗体,其中该Fc域包含一处或多处降低对Fc受体,特别是Fcγ受体的结合的氨基酸替代。10.如本文中之前定义的双特异性抗体,其中该Fc域是人IgG1亚类的,具有氨基酸突变L234A,L235A和P329G(编号方式依照KabatEU索引)。11.如本文中之前定义的双特异性抗体,其中该Fc域包含促进Fc域的第一和第二亚基联合的修饰。12.如本文中之前定义的双特异性抗体,其中依照节入穴方法,该Fc域的第一亚基包含节且该Fc域的第二亚基包含穴。13.如本文中之前定义的双特异性抗体,其中该Fc域的第一亚基包含氨基酸替代S354C和T366W(EU编号方式)且该Fc域的第二亚基包含氨基酸替代Y349C,T366S和Y407V(编号方式依照KabatEU索引)。14.如本文中之前定义的双特异性抗体,其中在该Fab片段之一中可变域VL和VH彼此替换,使得VH域是轻链的一部分且VL域是重链的一部分。15.如本文中之前定义的双特异性抗体,其中在包含该特异性结合PD1的抗原结合位点的第一Fab片段中,可变域VL和VH彼此替换。16.如本文中之前定义的双特异性抗体,其中在该Fab片段之一中在恒定域CL中位置124处的氨基酸被赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照KabatEU索引),且在恒定域CH1中位置147和213处的氨基酸被谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。17.如本文中之前定义的双特异性抗体,其中在包含该特异性结合TIM3的抗原结合位点的第二Fab片段中在恒定域CL中位置124处的氨基酸被赖氨酸(K),精氨酸(R)或组氨酸(H)独立替代(编号方式依照KabatEU索引),且在恒定域CH1中位置147和213处的氨基酸被谷氨酸(E)或天冬氨酸(D)独立替代(编号方式依照KabatEU索引)。18.如本文中之前定义的双特异性抗体,其包含(a)包含与SEQIDNO:50的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:52的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:51的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:53的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(b)包含与SEQIDNO:54的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:56的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:55的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:57的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(c)包含与SEQIDNO:58的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:60的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:59的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:61的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(d)包含与SEQIDNO:62的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:64的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:63的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:65的序列有至少95%序列同一性的氨基酸序列的第二轻链,或(e)包含与SEQIDNO:66的序列有至少95%序列同一性的氨基酸序列的第一重链,包含与SEQIDNO:68的序列有至少95%序列同一性的氨基酸序列的第一轻链,包含与SEQIDNO:67的序列有至少95%序列同一性的氨基酸序列的第二重链,和包含与SEQIDNO:69的序列有至少95%序列同一性的氨基酸序列的第二轻链。19.如本文中之前定义的双特异性抗体,其包含(a)包含SEQIDNO:50的氨基酸序列的第一重链,包含SEQIDNO:52的氨基酸序列的第一轻链,包含SEQIDNO:51的氨基酸序列的第二重链,和包含SEQIDNO:53的氨基酸序列的第二轻链,或(b)包含SEQIDNO:54的氨基酸序列的第一重链,包含SEQIDNO:56的氨基酸序列的第一轻链,包含SEQIDNO:55的氨基酸序列的第二重链,和包含SEQIDNO:57的氨基酸序列的第二轻链,或(c)包含SEQIDNO:58的氨基酸序列的第一重链,包含SEQIDNO:60的氨基酸序列的第一轻链,包含SEQIDNO:59的氨基酸序列的第二重链,和包含SEQIDNO:61的氨基酸序列的第二轻链,或(d)包含SEQIDNO:62的氨基酸序列的第一重链,包含SEQIDNO:64的氨基酸序列的第一轻链,包含SEQIDNO:63的氨基酸序列的第二重链,和包含SEQIDNO:65的氨基酸序列的第二轻链,或(e)包含SEQIDNO:66的氨基酸序列的第一重链,包含SEQIDNO:68的氨基酸序列的第一轻链,包含SEQIDNO:67的氨基酸序列的第二重链,和包含SEQIDNO:69的氨基酸序列的第二轻链。20.一种多核苷酸,其编码如本文中之前定义的双特异性抗体。21.一种载体,特别是表达载体,其包含如本文中之前定义的多核苷酸。22.一种原核或真核宿主细胞,其包含如本文中之前定义的多核苷酸或如本文中之前定义的载体。23.一种生成如本文中之前定义的双特异性抗体的方法,其包括下述步骤:a)用包含编码所述双特异性抗体的多核苷酸的载体转化宿主细胞,b)在适合于该双特异性抗体表达的条件下培养该宿主细胞,并c)自该培养物回收该双特异性抗体。24.一种药物组合物,其包含如本文中之前定义的双特异性抗体和至少一种药学可接受赋形剂。25.如本文中之前定义的双特异性抗体或如本文中之前定义的药物组合物,其用作药物。26.如本文中之前定义的双特异性抗体或如本文中之前定义的药物组合物,其用于i)调控免疫应答,诸如恢复T细胞活性,ii)刺激免疫应答或功能,iii)治疗感染,iv)治疗癌症,v)延迟癌症进展,vi)延长罹患癌症的患者的存活。27.如本文中之前定义的双特异性抗体或如本文中之前定义的药物组合物,其用于预防或治疗癌症。28.如本文中之前定义的双特异性抗体或如本文中之前定义的药物组合物,其用于治疗慢性病毒感染。29.如本文中之前定义的双特异性抗体或如本文中之前定义的药物组合物,其用于预防或治疗癌症,其中与化疗剂,放射和/或其它用于癌症免疫疗法的药剂组合施用该双特异性抗体。30.一种在个体中抑制肿瘤细胞生长的方法,其包括对该个体施用有效量的如本文中之前定义的双特异性抗体以抑制肿瘤细胞生长。实施例以下是本发明的方法和组合物的实施例。应当理解,根据上面提供的一般描述,可以实践各种其它实施方案。材料和通用方法关于人免疫球蛋白轻和重链的核苷酸序列的一般信息见Kabat,E.A.,etal.,SequencesofProteinsofImmunologicalInterest,5thed.,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991)。依照上文定义的依照Kabat的编号系统(Kabat,E.A.,etal.,SequencesofProteinsofImmunologicalInterest,5thed.,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991))对抗体链的氨基酸编号及提述。重组DNA技术使用标准方法操作DNA,如Sambrook,J.etal.,MolecularCloning:Alaboratorymanual;ColdSpringHarborLaboratoryPress,ColdSpringHarbor,NewYork,1989中所述。按照制造商的说明使用分子生物试剂。基因合成从通过化学合成生成的寡核苷酸制备期望的基因区段。包括PCR扩增在内,通过退火和连接寡核苷酸装配侧翼为单一限制性内切核酸酶切割位点的600-1800bp长基因区段,并随后经由指定的限制性位点例如KpnI/SacI或AscI/PacI克隆入基于pPCRScript(Stratagene)的pGA4克隆载体。通过DNA测序确认亚克隆基因片段的DNA序列。依照Geneart(Regensburg,Germany)给出的说明书定购基因合成片段。DNA序列测定通过在MediGenomixGmbH(Martinsried,Germany)或SequiserveGmbH(Vaterstetten,Germany)实施的双链测序测定DNA序列。DNA和蛋白质序列分析和序列数据管理使用该GCG's(GeneticsComputer组,Madison,Wisconsin)软件包版本10.2和Infomax的VectorNT1Advance套组版本8.0进行序列创建,定位,分析,注释和说明。表达载体为了表达描述的抗体,将基于有或无CMV-内含子A启动子的cDNA组织或基于有CMV启动子的基因组组织的表达质粒的变体应用于瞬时表达(例如HEK293)细胞。在抗体表达盒以外,载体还包含:-复制起点,其容许这种质粒在大肠杆菌中复制,和-β-内酰胺酶基因,其在大肠杆菌中赋予氨苄青霉素抗性。抗体基因的转录单元由下述元件构成:-在5’端的一个或多个独特限制性位点,-来自人巨细胞病毒的立即早期增强子和启动子,-接着是在cDNA组织的情况中,内含子A序列,-人抗体基因的5’非翻译区,-免疫球蛋白重链信号序列,-作为cDNA或作为基因组组织具有免疫球蛋白外显子-内含子组织的人抗体链(野生型或具有域交换),-具有多腺苷酸化信号序列的3’非翻译区,和-在3’端的一个或多个独特限制性位点。通过PCR和/或基因合成产生包含如下文所述抗体链的融合基因,并通过已知重组方法和技术通过连接(例如使用相应载体中的独特限制性位点)相应的核酸区段进行装配。通过DNA测序验证亚克隆的核酸序列。为了瞬时转染,通过来自经过转化的大肠杆菌培养物的质粒制备物(NucleobondAX,Macherey-Nagel)制备较大量的质粒。细胞培养技术使用如CurrentProtocolsinCellBiology(2000),Bonifacino,J.S.,Dasso,M.,Harford,J.B.,Lippincott-Schwartz,J.和Yamada,K.M.(eds.),JohnWiley&Sons,Inc.中所述的标准细胞培养技术。通过如下文所述在贴壁生长的HEK293-EBNA中或在悬浮生长的HEK29-F细胞中瞬时共转染各自的表达质粒来表达多特异性抗体。HEK293系统中的瞬时转染通过使用Freestyle系统(ThermoFisher)瞬时转染293F细胞来生成所有抗体和双特异性抗体。这里,在F17培养基中培养293F细胞,用293Free(Novagene)转染并在4小时后补料VPA4mM和补料7和16小时后补料0.6%葡萄糖。并且使用Expi293FTM表达系统试剂盒(ThermoFisher)。这里,在Expi293TM表达培养基中培养Expi293FTM细胞并使用ExpiFectamineTM293转染试剂盒依照制造商的说明书转染。由于具有另外引入的CH1/CL界面(别的详情见各自序列中的位置)中的带电荷的氨基酸对的CrossMAbVh-VL双特异性抗体的改善的稳定性和纯度和降低的聚集趋势,没有采用质粒比率的调整。因此,使用相对质粒比1:1:1:1(用于1+1CrossMab)或1:1:1(用于2+2CrossMab)进行LC,HC,交叉的LC和交叉的HC质粒的共转染。7天后收获细胞上清液并通过标准方法纯化。蛋白质测定使用依照Pace,etal.,ProteinScience,1995,4,2411-1423根据氨基酸序列计算的摩尔消光系数,通过测定280nm处的光密度(OD)来测定纯化的抗体和衍生物的蛋白质浓度。上清液中的抗体浓度测定用蛋白A琼脂糖珠(Roche)通过免疫沉淀来评估细胞培养物上清液中抗体和衍生物的浓度。将60μL蛋白A琼脂糖珠在TBS-NP40(50mMTris,pH7.5,150mMNaCl,1%Nonidet-P40)中清洗三次。随后,将1-15mL细胞培养物上清液应用至在TBS-NP40中预平衡的蛋白A琼脂糖珠。于室温温育1小时之后,将珠在Ultrafree-MC-滤器柱(Amicon)上用0.5mLTBS-NP40清洗一次,用0.5mL2x磷酸盐缓冲盐水(2xPBS,Roche)清洗两次,并用0.5mL100mM柠檬酸钠pH5,0简单清洗四次。通过添加35μlLDS样品缓冲液(Invitrogen)来洗脱结合的抗体。分别将一半样品与样品还原剂组合或保持不还原,并于70℃加热10min。因此,将5-30μl应用至4-12%Bis-TrisSDS-PAGE(Invitrogen)(MOPS缓冲液用于非还原SDS-PAGE,带抗氧化剂运行缓冲液添加剂(Invitrogen)的MES缓冲液用于还原SDS-PAGE),并用考马斯蓝染色。通过亲和HPLC层析定量测量细胞培养物上清液中抗体和衍生物的浓度。简言之,在AgilentHPLC1100系统上,将含有结合蛋白A的抗体和衍生物的细胞培养物上清液应用至200mMKH2PO4,100mM柠檬酸钠,pH7.4中的AppliedBiosystemsPorosA/20柱,并用200mMNaCl,100mM柠檬酸,pH2,5自基质洗脱。通过UV吸光度和峰面积积分来定量洗脱的蛋白质。纯化的标准IgG1抗体充当标准。或者,通过三明治式IgG-ELISA测量细胞培养物上清液中抗体和衍生物的浓度。简言之,将StreptaWell高结合链霉亲合素A-96孔微量滴定板(Roche)用100μL/孔0.1μg/mL生物素化抗人IgG捕捉分子F(ab’)2<h-Fcγ>BI(Dianova)于室温包被1小时或于4℃包被过夜,随后用200μL/孔PBS,0.05%Tween(PBST,Sigma)清洗三次。将100μL/孔各含有抗体的细胞培养物上清液在PBS(Sigma)中的稀释系列添加至孔,并在微量滴定板摇床上于室温温育1-2小时。将孔用200μL/孔PBST清洗三次,并在微量滴定板摇床上用100μl0.1μg/mLF(ab’)2<hFcγ>POD(Dianova)作为检测抗体于室温检测结合的抗体1-2小时。三次用200μL/孔PBST洗掉未结合的检测抗体,并通过添加100μLABTS/孔来检测结合的检测抗体。在Tecan荧光分光仪上实施吸光度测定,测量波长405nm(参照波长492nm)。蛋白质纯化参考标准方案,从过滤的细胞培养物上清液中纯化蛋白质。简言之,将抗体施加到蛋白A琼脂糖凝胶柱(GEHealthcare)上并用PBS洗涤。在pH2.8实现抗体的洗脱,然后立即中和样品。通过尺寸排阻色谱法(Superdex200,GEHealthcare)在PBS或在20mM组氨酸、150mMNaCl(pH6.0)中从单体抗体分离聚集的蛋白质。合并单体抗体级分,使用例如MILLIPOREAmiconUltra(30MWCO)离心浓缩机进行浓缩(如果需要的话),在-20℃或-80℃冷冻并储存。提供部分样品用于随后例如通过SDS-PAGE、尺寸排阻色谱法(SEC)或质谱术进行蛋白质分析和分析表征。SDS-PAGE根据制造商的说明使用Pre-Cast凝胶系统(Invitrogen)。具体地说,使用10%或4-12%的Bis-TRISPre-Cast凝胶(pH6.4)和MES(还原凝胶,具有抗氧化剂运行缓冲液添加剂)或MOPS(非还原凝胶)运行缓冲液。分析尺寸排阻色谱法通过HPLC色谱法进行尺寸排阻色谱法(SEC),用于测定抗体的聚集和寡聚状态。简言之,将蛋白A纯化的抗体施加到在AgilentHPLC1100系统上的300mMNaCl、50mMKH2PO4/K2HPO4(pH7.5)中的TosohTSKgelG3000SW柱上,或者在DionexHPLC系统上的2xPBS中的Superdex200柱(GEHealthcare)上。通过UV吸光度和峰面积的积分将洗脱的蛋白质定量。以BioRad凝胶过滤标准151-1901作为标准。质谱术本节描述了具有VH/VL交换(VH/VLCrossMabs)的多特异性抗体的表征,重点在于它们的正确装配。通过去糖基化的完整CrossMabs和去糖基化的/纤溶酶消化的或替代地去糖基化的/限制性LysC消化的CrossMabs的电喷雾电离质谱术(ESI-MS)分析预期的一级结构。在37℃下以1mg/ml的蛋白质浓度将VH/VLCrossMabs用磷酸盐或Tris缓冲液中的N-糖苷酶F进行去糖基化达17个小时。用Tris缓冲液(pH8)中的100μg去糖基化的VH/VLCrossMabs,分别在室温下进行120小时和在37℃下进行40分钟的纤溶酶或限制性LysC(Roche)消化。在进行质谱分析之前,将样品通过HPLC在SephadexG25柱(GEHealthcare)上脱盐。在装备有TriVersaNanoMate来源(Advion)的maXis4GUHR-QTOFMS系统(BrukerDaltonik)上,通过ESI-MS测定总质量。使用表面等离振子共振(SPR)(BIACORE)测定多特异性抗体对各自抗原的结合和结合亲和力使用BIACORE仪器(GEHealthcareBiosciencesAB,Uppsala,Sweden)通过表面等离振子共振来调查生成的抗体对各自抗原的结合。简言之,为了亲和力测量,经胺偶联在CM5芯片上固定化山羊抗人IgG,JIR109-005-098抗体以呈现针对各自抗原的抗体。在HBS缓冲液(HBS-P(10mMHEPES,150mMNaCl,0.005%Tween20,ph7.4)中于25℃(或者于37℃)测量结合。在溶液中以各种浓度添加抗原(R&DSystems或内部纯化)。通过80秒钟至3分钟的抗原注射来测量结合;通过用HBS缓冲液清洗芯片表面3-10分钟来测量解离,并使用1:1Langmuir结合模型来估算KD值。自样品曲线减去阴性对照数据(例如缓冲液曲线)以修正系统内在基线漂移及降低噪声信号。使用各自Biacore评估软件来分析传感图及计算亲和力数据。TIM3抗体实施例1a:抗TIM3抗体的生成小鼠的免疫接种通过皮内应用100ug载体DNA(质粒15304_hTIM3-fl),接着电穿孔(2个1000V/cm的方形脉冲,持续时间0.1ms,间隔0.125s;接着4个287.5V/cm的方形脉冲,持续时间10ms,间隔0.125s),使用编码全长人Tim-3的质粒表达载体对NMRI小鼠进行基因免疫接种。小鼠在第0,14,28,42,56,70,和84天接受6次连续免疫接种任一。在第36,78和92天采集血液并制备血清,用于通过ELISA进行的滴度测定(见下文)。选择具有最高滴度的动物在第96天通过静脉内注射50ug重组人Tim-3人Fc嵌合物进行加强,并通过杂交瘤技术,通过在加强后3天融合脾细胞与骨髓瘤细胞系来分离单克隆抗体。血清滴定的测定(ELISA)以0.3ug/ml,100μl/孔,在PBS中在96孔NUNCMaxisorp板上固定化人重组Tim-3人Fc嵌合物,接着:用含2%CroteinC的PBS,200μl/孔封闭板;一式两份,在含0.5%CroteinC的PBS中,100μl/孔应用抗血清的连续稀释液;用HRP缀合的山羊抗小鼠抗体(JacksonImmunoresearch/Dianova115-036-071;1/16000)检测。对于所有步骤,将板于37℃温育1小时。在所有步骤之间,将板用含0.05%Tween20的PBS清洗3次。通过添加可溶性BMBluePOD底物(Roche),100ul/孔来产生信号;并通过添加1MHCl,100ul/孔来停止信号。针对作为参照的690nm,读取450nm处的吸光度。滴度定义为产生半最大信号的抗血清的稀释度。实施例1b:抗TIM3抗体的表征针对TIM3的ELISA将Nunc-MaxiSorp链霉亲合素板(MicroCoat#11974998/MC1099)用25μl/孔Tim3-ECD-His-生物素(用BirA连接酶生物素化)包被并于室温温育1小时同时以400rpm旋转摇动。清洗(3x90μl/孔PBST缓冲液)后添加25μlaTim3样品或稀释的(1:2梯级)参照抗体aTim3F38-2E2(Biolegend)并于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后添加25μl/孔1:9000稀释度的绵羊抗小鼠POD(GENA9310V)并于室温温育1小时同时以400rpm旋转摇动。清洗(4x90μl/孔PBST缓冲液)后添加25μl/孔TMB底物(Calbiochem,#CL07)并温育直至OD1.5–2.5。然后通过添加25μl/孔1NHCl溶液来停止反应。于370/492nm进行测量。ELISA结果作为EC50值[ng/ml]在下文汇总表1中列出。针对TIM3的细胞ELISA以1.2x10E6个细胞/ml的浓度将用编码全长人Tim3的质粒15312_hTIM3-fl_pUC_Neo稳定转染且用G418(质粒上的新霉素抗性标志物)选择的贴壁CHO-K1细胞系接种入384孔平底板并生长过夜。次日添加25μl/孔Tim3样品或aTim3参照抗体F38-2E2无叠氮化物(Biolegend,354004)并于4℃温育2小时(以避免内在化)。清洗(3x90μl/孔PBST(BIOTEK清洗仪:程序29,1x90)后通过弹出残留缓冲液并添加50μl/孔0.05%戊二醛:25%戊二醛(Sigma目录号:G5882)在1xPBS缓冲液中的1:500稀释液并于室温温育1小时来固定细胞。清洗(3x90μl/孔PBST(BIOTEK清洗仪:程序21,3x90GreinLysin)后添加25μl/孔二抗进行检测(绵羊抗小鼠-POD;辣根POD连接的F(ab')2片段;GENA9310),接着于室温温育2小时同时以400rpm摇动。清洗(3x90μl/孔PBST(BIOTEK清洗仪:程序21,3x90GreinLysin)后添加25μl/孔TMB底物溶液(Roche11835033001)并温育直至OD1.5–2.5。然后通过添加25μl/孔1NHCl溶液来停止反应。于370/492nm进行测量。细胞ELISA结果作为“EC50CHO-Tim3”值[ng/ml]在下文汇总表1中列出。表1:例示性抗体的结合亲和力(ELISA和BIACORE)TIM3抗体的BIAcore表征基于表面等离振子共振(SPR)的测定法已经用于测定数种鼠Tim3结合物以及商业性人Tim3结合参照之间的结合的动力学参数。因此,通过胺偶联将抗小鼠IgG固定化至(BIAcore)CM5传感器芯片的表面。然后捕捉样品并使单体hu/cyTim3-ECD以及带Fc标签的人Tim3-ECD二聚体结合至它们。每个分析循环后再生传感器芯片表面。最后通过将数据拟合1:1朗格缪尔(Langmuir)相互作用模型来获取平衡常数KD。使用由GEHealthcare供应的胺偶联试剂盒于pH5.0将约12000个响应单位(RU)的30μg/ml抗小鼠IgG(GEHealthcare#BR-1008-38)偶联到BIAcoreB4000中的CM5传感器芯片的流动室1-4的点1,2,4和5上(点1,5是有活性的而点2,4是参照点)。样品和运行缓冲液是HBS-EP+(0.01MHEPES,0.15MNaCl,3mMEDTA,0.05%v/v表面活性剂P20,pH7.4)。流动室温度设为25℃且样品隔室温度设为12℃。用运行缓冲液引发系统。样品以200μg/ml的浓度注射30秒并结合至每个流动室的点1和5,容许平行测量8份样品。然后一整套不同(单体食蟹猴,单体人和huFc融合的二聚体人Tim3-ECD)浓度在每份样品上注射240秒,接着是解离时间30/1800秒。然后用30秒长注射甘氨酸-HClpH1.7再生每个分析循环(样品捕捉,点1和5–Tim3ECD注射)。整个运行的流速设为30μl/min。最后,用BIAcoreB4000评估软件将双重参照数据拟合1:1朗格缪尔相互作用模型。所得对单体人,食蟹猴Tim3和huFc融合的二聚体人Tim3的亲和力在表2a中显示。对huTim3二聚体的亲和力更有可能受到亲合作用影响,因此表观上比对单体huTim3的亲和力要强。表2a:通过BIAcore测定的结合亲和力-通过动力学SPR测量获取的KD值。-n.f.表示没有可能的拟合,最有可能是由于没有结合或结合弱经由SPR对Tim3的亲和力的测定(嵌合TIM3-0016变体(0018)和人源化型式)通过胺偶联将蛋白A固定化至(Biacore)CM5传感器芯片的表面。然后捕捉样品并使huTim3-ECD结合至它们。每个分析周期后再生传感器芯片表面。最后通过将数据拟合1:1朗格缪尔相互作用模型来获取平衡常数和动力学速率常数。使用由GEHealthcare供应的胺偶联试剂盒将约2000个响应单位(RU)的20μg/ml蛋白A偶联到BiacoreB4000仪器中的CM5传感器芯片的所有流动室的点1,2,4和5上。样品和运行缓冲液是HBS-EP+(0.01MHEPES,0.15MNaCl,3mMEDTA,0.05%v/v表面活性剂P20,pH7.4)。流动室温度设为25℃且样品隔室温度设为12℃。用运行缓冲液引发系统。不同样品以10nM的浓度注射30秒并序贯结合至所有流动室的点1和5。然后一整套单体人Tim3-ECD稀释液(600nM,200nM,66.7nM,2×22.2nM,7.4nM,2.5nM和2×0nM)在每份样品上序贯注射300秒。每次抗原注射接着解离时间12秒/1000秒和两个30秒使用甘氨酸-HClpH1.5溶液的再生步骤,其中最后一个在20秒注射后含有稳定期。最后,使用BiacoreB4000评估软件将双重参照数据拟合1:1朗格缪尔相互作用模型。所得KD值在表2b中显示。经由SPR对Tim3的亲和力的测定(嵌合TIM3-0028和人源化型式)通过胺偶联将抗人FcIgG固定化至(Biacore)CM5传感器芯片的表面。然后捕捉样品并使huTim3-ECD结合至它们。每个分析周期后再生传感器芯片表面。最后通过将数据拟合1:1朗格缪尔相互作用模型来获取平衡常数和动力学速率常数。使用由GEHealthcare供应的胺偶联试剂盒将约2500个响应单位(RU)的10μg/ml抗人FcIgG(GEHealthcare#BR-1008-39)偶联到BiacoreB4000仪器中的CM5传感器芯片的所有流动室的点1,2,4和5上。样品和运行缓冲液是HBS-EP+(0.01MHEPES,0.15MNaCl,3mMEDTA,0.05%v/v表面活性剂P20,pH7.4)。流动室温度设为25℃且样品隔室温度设为12℃。用运行缓冲液引发系统。不同样品以10nM的浓度注射30秒并序贯结合至所有流动室的点1和5。然后一整套单体人Tim3-ECD稀释液(600nM,200nM,66.7nM,2×22.2nM,7.4nM,2.5nM和2×0nM)在每份样品上序贯注射300秒。每次抗原注射接着解离时间12秒/700秒和两个30秒使用3MMgCl2溶液的再生步骤,其中最后一个含有使用运行缓冲液的“注射后额外清洗”。最后,使用BiacoreB4000评估软件将双重参照数据拟合1:1朗格缪尔相互作用模型。所得KD值在表2b中显示。表2b:通过BIAcore测定的结合亲和力-通过动力学SPR测量获取的KD值样品huTim3KD(25℃)[M]嵌合TIM3-0016变体(0018)2.78E-09TIM3-04335.74E-09TIM3-04345.76E-09嵌合TIM3-00282.35E-07TIM3-04383.05E-07TIM3-04432.87E-07实施例2:抗TIM3抗体衍生物的生成嵌合抗体衍生物如下生成嵌合Tim3抗体,即经由PCR扩增抗TIM3小鼠抗体Tim3-0016,Tim3-0016变体(0018),Tim3-0021,Tim3-0022,Tim3-0026,Tim3-0028,Tim3-0030,和Tim3-0033,Tim3-0038的可变重和轻链区并将它们作为与人IgG1主链/人CH1-铰链-CH2-CH3(具有消除效应器功能的LALA和PG突变(亮氨酸234变成丙氨酸,亮氨酸235变成丙氨酸,脯氨酸329变成甘氨酸))的融合蛋白克隆入重链表达载体及作为与人C-κ的融合蛋白克隆入轻链表达载体。然后将LC和HC质粒共转染入HEK293并在7天后通过用于抗体纯化的标准方法自上清液纯化。去除糖基化位点NYT:通过用Q或S替代N来修饰Tim3-0016,Tim3_0016变体(称作0018或Tim3_0018)中的1个HVR-L1位置使用Agilent“QuickChangeLightning定点诱变试剂盒”依照制造商的说明书通过体外诱变生成Tim3_0016和Tim3_0016变体(0018)的可变轻链区内的突变。通过这种方法,轻链HVR-L1(SEQIDNO:4)中的糖基化位点基序NYT的天冬酰胺(N)用谷氨酰胺(Q)替换(产生SEQIDNO:11=Tim3_0016_HVR-L1变体1_NQ),或者,天冬酰胺(N)用丝氨酸(S)替换(产生SEQIDNO:12=Tim3_0016_HVR-L1变体2_NS)。在二者中,糖基化位点基序NYT均成功修饰。然后将编码变体的LC和HC质粒共转染入HEK293并在7天后通过用于抗体纯化的标准方法自上清液纯化。通过人Tim3上的ELISA,食蟹猴Tim3上的ELISA和表达全长人Tim3的贴壁CHO-K1细胞上的细胞ELISA测试所生成的突变体。表3:发现所生成的所有突变体均显示分别比亲本抗体Tim3_0016或Tim3_0016抗体变体Tim3_0018甚至更具功能性的对CHO细胞上的人TIM3(人),食蟹猴TIM3(cyno)或人TIMR的结合。人源化抗体衍生物鼠抗Tim3-0016变体(0018)和抗Tim3_0028的VH和VL域的人源化基于a)抗Tim3抗体Tim3_0016变体(0018)(具有6个HVR的氨基酸序列,其中在轻链中使用HVR-L1变体2_NS(通过N变成S的突变来去除糖基化位点))的VH和VL域的氨基酸序列生成了人源化抗Tim3抗体变体Tim3-0433和Tim3-0434,而基于b)抗Tim3抗体Tim3_0028的VH和VL域的氨基酸序列,生成了人源化抗Tim3抗体变体Tim3-0438和Tim3-0443。将重和轻链可变区的人源化氨基酸序列倒退翻译成DNA并合成所得cNDA(GenArt),然后作为与具有消除效应器功能的LALA和PG突变(亮氨酸234变成丙氨酸,亮氨酸235变成丙氨酸,脯氨酸329变成甘氨酸)的人IgG1主链/人CH1-铰链-CH2-CH3的融合蛋白克隆入重链表达载体或作为与人C-κ的融合蛋白克隆入轻链表达载体。然后将LC和HC质粒共转染入HEK293并在7天后通过用于抗体纯化的标准方法自上清液纯化。所得人源化Tim3-抗体命名如下:表4:人源化抗体的VH和VL序列Tim3_0016变体(0018)的人源化抗体VH/SEQIDNO:VL/SEQIDNO:Tim3-0433SEQIDNO:13SEQIDNO:14Tim3-0434SEQIDNO:15SEQIDNO:16Tim3_0028的人源化抗体VH/SEQIDNO:VL/SEQIDNO:Tim3-0438SEQIDNO:25SEQIDNO:26Tim3-0443SEQIDNO:27SEQIDNO:28表5:人源化抗体的HVR序列实施例3:混合淋巴细胞反应(MLR)中人抗TIM-3抗体对细胞因子生成的影响使用混合淋巴细胞反应来展现阻断TIM-3途径对淋巴细胞效应细胞的影响。在抗TIM-3单抗存在或缺失下对测定法中的T细胞测试活化和IFN-γ分泌。使用Leukosep(GreinerBioOne,227288)通过密度梯度离心自健康供体的外周血分离人淋巴细胞。简言之,用3倍体积的PBS稀释肝素化血液并将稀释血液的25ml小样在50mlLeukosep管中分层。于室温以800xg离心15分钟(无中断)后收获含有淋巴细胞的级分,在PBS中清洗并在功能测定法中直接使用或以1.0E+07个细胞/ml在冷冻培养基(10%DMSO,90%FCS)中重悬浮并在液氮中保存。在MLR测定法中使用1:1靶细胞/响应者细胞比率(即每份MLR培养物在总体积200μl中含有-2.0E+05个来自每位供体的PBMC。以不同抗体浓度将抗TIM3单克隆抗体Tim3_0016,Tim3_0016变体(Tim3_0018),Tim3_0021,Tim3_0022,Tim3_0026,Tim3_0028,Tim3_0030,Tim3_0033,Tim3_0038和F38-2E2(BioLegend)添加至每份培养物。使用无抗体或同种型对照抗体任一作为阴性对照而使用重组huIL-2(20EU/ml)作为阳性对照。将细胞于37℃培养6天。第6天后自每份培养物取出100μl培养基用于细胞因子测量。使用OptEIAELISA试剂盒(BDBiosciences)测量IFN-γ的水平。结果在表6中显示(IFN-g分泌/释放)。抗TIM-3单克隆抗体以浓度依赖性方式促进T细胞活化和IFN-γ分泌。抗TIM3抗体Tim3_0021,Tim3_0022,Tim3_0028,和Tim3_0038比F38-2E2抗体更多地降低炎性细胞因子IFN-γ)的释放。比较F38-2E2抗体时Tim3_0016,Tim3_0016变体(Tim3_0018),Tim3_0033和Tim3_0038显示相似的释放。与之对比,含有同种型对照抗体的培养物并不显示IFN-γ分泌的升高。表6a:抗Tim3抗体诱导的IFNγ释放的百分比,与作为阳性对照的rechuIL-2(20EU/ml)(=100%)和作为阴性对照的无抗体比较在进一步的实验中,测量与0.1μg/ml抗PD1单抗组合的下述嵌合和人源化抗体(如上所述生成的)的EC50值:用不同淋巴细胞供体混合物(分别为D2和D3,或D1和D5)测量嵌合chi_Tim3_018抗体及其人源化型式Tim3-433和Tim3-434,嵌合chi_Tim3_028抗体及其人源化型式Tim3-438和Tim3-443。结果在表6b中显示。表6b:抗Tim3抗体诱导的(IFN-g分泌/释放)的EC50抗体供体D2+D3的EC50[nM]供体D1+D5的EC50[nM]chi_Tim3_0183.14.2Tim3-4333.02.4Tim3-4341.72.6chi_Tim3_0282.96.4Tim3-4381.92.7Tim3-4433.04.7PD1抗体实施例4:抗PD-1抗体的生成小鼠的免疫接种对NMRI小鼠进行遗传免疫接种,其使用编码全长人PD-1的质粒表达载体通过皮内应用100ug载体DNA(质粒15300_hPD1-fl),接着电穿孔(2个1000V/cm矩形脉冲,持续0.1ms,间隔0.125s;接着4个287.5V/cm矩形脉冲,持续10ms,间隔0.125s)进行。小鼠在第0,14,28,42,56,70,和84天连续接受6次免疫接种。在第36,78和92天采集血液并制备血清,用于通过ELISA进行的滴度测定(见下文)。选择具有最高滴度的动物在第96天通过静脉内注射50ug重组人PD1人Fc嵌合物进行强化,并通过在强化后3天融合脾细胞与骨髓瘤细胞系通过杂交瘤技术分离单克隆抗体。血清滴度的测定(ELISA)在PBS中以0.3ug/ml,100ul/孔在96孔NUNCMaxisorp板上固定化人重组PD1人Fc嵌合物,接着是:用含2%CroteinC的PBS,200ul/孔封闭板;应用抗血清的连续稀释液,一式两份,在含0.5%CroteinC的PBS中,100ul/孔;用HRP缀合的山羊抗小鼠抗体(JacksonImmunoresearch/Dianova115-036-071;1/16000)检测。对于所有步骤,将板于37℃温育1小时。在所有步骤之间,将板用含0.05%Tween20的PBS清洗3次。通过添加BMBluePOD底物溶液(Roche),100ul/孔来发生信号;并通过添加1MHCl,100ul/孔来终止。于450nm读取吸光度,针对690nm作为参照。滴度定义为产生半最大信号的抗血清的稀释度。实施例5:抗PD1抗体的表征/抗PD1抗体对人PD1的结合针对人PD1的ELISA将NuncMaxisorp链霉亲合素包被板(MicroCoat#11974998001)用25μl/孔生物素化PD1-ECD-AviHis包被并于4℃温育过夜。清洗(3x90μl/孔PBST缓冲液)后,添加25μl抗PD1样品或参照抗体(人抗PD1;Roche/小鼠抗PD1;Biolegend;目录:329912)并于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,以1:2000/1:1000稀释度添加25μl/孔山羊抗人H+L-POD(JIR,JIR109-036-088)/绵羊抗小鼠-POD(GEHealthcare;NA9310)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche目录号11835033001)并温育直至OD2–3。于370/492nm进行测量。在下文汇总表7和8中作为EC50值[ng/ml]列出ELISA结果。针对PD1的细胞ELISA以0.01x10E6细胞/ml的浓度将用编码全长人PD1的质粒15311_hPD1-fl_pUC_Neo稳定转染并用G418(质粒上的新霉素抗性标志物)选择的贴壁CHO-K1细胞系接种入384孔平底板并生长过夜。次日,添加25μl/孔PD1样品或人抗PD1(Roche)/小鼠抗PD1(Biolegend;目录:329912)参照抗体并于4℃温育2小时(以避免内在化)。小心清洗(1x90μl/孔PBST)后,通过添加30μl/孔在1xPBS缓冲液中稀释的0.05%戊二醛(Sigma,目录号:G5882,25%)并于室温温育10分钟来固定细胞。清洗(3x90μl/孔PBST)后,添加25μl/孔二抗进行检测:山羊抗人H+L-POD(JIR,JIR109-036-088)/绵羊抗小鼠-POD(GENA9310),接着在摇床上于室温温育1小时。清洗(3x90μl/孔PBST)后,添加25μl/孔TMB底物溶液(Roche11835033001)并温育直至OD1.0–2.0。于370/492nm测量板。下文汇总表8中作为“EC50CHO-PD1”值[ng/ml]列出细胞ELISA结果。针对食蟹猴PD1的ELISA将NuncMaxisorp链霉亲合素包被板(MicroCoat#11974998001)用25μl/孔生物素化cynoPD1-ECD-生物素包被并于4℃温育过夜。清洗(3x90μl/孔PBST缓冲液)后,添加25μl抗PD1样品或参照抗体(人抗PD1;Roche)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,以1:1000稀释度添加25μl/孔山羊抗人H+L-POD(JIR,JIR109-036-088)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche,11835033001)并温育直至OD2–3。于370/492nm进行测量。在下文汇总表7和8中作为EC50值[ng/ml]列出ELISA结果。PD配体1置换测定法将NuncMaxisorp链霉亲合素包被板(MicroCoat#11974998001)用25μl/孔生物素化PD1-ECD-AviHis包被并于4℃温育过夜。清洗(3x90μl/孔PBST缓冲液)后,添加25μl抗PD1样品或参照抗体(小鼠抗PD1;Biolegend;目录:329912)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔PD-L1(重组人B7-H1/PD-L1Fc嵌合物;156-B7,R&D)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,以1:1000稀释度添加25μl/孔山羊抗人H+L-POD(JIR,109-036-088)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche,11835033001)并温育直至OD2–3。于370/492nm进行测量。在下文汇总表7中作为IC50值[ng/ml]列出ELISA结果。PD配体2置换测定法将NuncMaxisorp链霉亲合素包被板(MicroCoat#11974998001)用25μl/孔生物素化PD1-ECD-AviHis包被并于4℃温育过夜。清洗(3x90μl/孔PBST缓冲液)后,添加25μl抗PD1样品或参照抗体(小鼠抗huPD1;Roche)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔PD-L2(重组人B7-DC/PD-L2Fc嵌合物;1224-PL-100,R&D)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,以1:2000稀释度添加25μl/孔山羊抗人H+L-POD(JIR,109-036-088)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche,11835033001)并温育直至OD2–3。于370/492nm进行测量。在下文汇总表7中作为IC50值[ng/ml]列出ELISA结果。表位作图ELISA/结合竞争测定法将NuncMaxisorp板(Nunc#464718)用25μl/孔捕捉抗体(山羊抗小鼠IgG;JIR;115-006-071)包被并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,将板用含有2%BSA的PBS缓冲液在摇床上于室温封闭1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl小鼠抗PD1样品并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,将捕捉抗体用30μl/孔小鼠IgG(JIR;015-000-003)在摇床上于室温封闭1小时。同时,将生物素化PD1-ECD-AviHis与第二样品抗体一起在摇床上于室温预温育1小时。清洗测定板(3x90μl/孔PBST缓冲液)后,将PD1抗体混合物转移至测定板并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,以1:4000稀释度添加25μl/孔链霉亲合素POD(Roche,#11089153001)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche,11089153001)并温育直至OD1.5–2.5。于370/492nm进行测量。通过针对参照抗体的等级聚簇来定义表位组。表7:例示性抗体的结合,PD-L1抑制和表位区分组(ELISA)表8:自亲本小鼠抗体PD1-0103衍生的人源化PD1抗体的生化和细胞结合(ELISA)人源化抗PD-1抗体的Biacore表征已经使用基于表面等离振子共振(SPR)的测定法在数种鼠PD1结合物以及商业性人PD1结合参照之间测定结合的动力学参数。因此,通过胺偶联将抗人IgG固定化至(Biacore)CM5传感器芯片的表面。然后捕捉样品并使人PD1-ECD结合至它们。在每个分析循环后再生传感器芯片表面。最终通过将数据拟合至1:1Langmuir相互作用模型来得到平衡常数和动力学速率常数。通过使用胺偶联试剂盒(GEHealthcare)于pH5.0将约2000个响应单位(RU)的20μg/ml抗人IgG(GEHealthcare#BR-1008-39)偶联到BiacoreT200中的CM5传感器芯片的流动室1和2(或者3和4)上。样品和运行缓冲液是HBS-EP+(0.01MHEPES,0.15MNaCl,3mMEDTA,0.05%v/v表面活性剂P20,pH7.4)。流动室温度设定为25℃,而样品隔室温度设定为12℃。用运行缓冲液引发系统。样品以10nM的浓度注射20秒,并结合至第二流动室。然后在每份样品上注射一整套人PD1-ECD浓度(144nM,48nM,16nM,5.33nM,1.78nM,0.59nM,0.20nM和0nM)120秒,接着是解离时间30/300秒和3MMgCl2溶液的两个20秒再生步骤,其中最后一个含有运行缓冲液的“注射后额外清洗”。最终,用BiacoreT200评估软件将双重参照数据拟合至1:1Langmuir相互作用模型。所得KD,ka和kd值在表9中显示。表9:通过Biacore测定的嵌合PD1-0103和人源化PD1抗体的动力学速率常数和平衡常数如表9中显示的,嵌合PD1-0103的所有人源化型式(生成见实施例6)展示与亲本抗体(嵌合PD1-0103)相似的动力学特性。动力学将S系列CM5传感器安置到Biacore4000系统中并依照制造商的说明书对检测点进行流体动力学编址。在流动室1,2,3和4中的检测点1和5上以10000Ru固定化多克隆家兔IgG抗体<IgGFCγM>R(JacksonImmunoResearchLaboratoriesInc.)。经由EDC/NHS化学依照制造商的说明书进行偶联。流动室中的剩余点充当参照。样品缓冲液是补充有1mg/ml羧甲基葡聚糖的系统缓冲液。在一个实施方案中,于25℃驱动测定法。在另一个实施方案中,于37℃驱动测定法。通过以10μl/min注射1分钟,在传感器表面上捕捉50nM每一种鼠单克隆抗体。随后,对于4分钟结合期时间,以30μl/min注射相应抗原的100nM,2x33nM,11nM,4nM,1nM浓度系列和系统缓冲液0nM。监测解离另外4分钟。使用以30μl/min注射10mM甘氨酸pH1.5达3分钟来再生捕捉系统。使用Biacore评估软件依照制造商的说明书计算相关动力学数据。表位作图对Biacore4000仪器安置BiacoreCAP传感器并像制造商推荐的那样做好准备。仪器缓冲液是HBS-ET(10mMHEPESpH7.4,150mMNaCl,3mMEDTA,0.005%w/vTween20)。于25℃运行仪器。在系统缓冲液中稀释所有样品。通过在流动室1,2,3和4中在点1和5中以30μl/min注射1分钟,以200RU在CAP传感器表面上捕捉35kDa生物素化抗原PD1-ECD-AviHis。点2,3和4充当参照。在另一个实施方案中,以相同方式以200RU在CAP传感器上捕捉35kDa生物素化抗原PD1-ECD-AviHis。随后,以30μl/min注射100nM一抗达3分钟,接着以30μl/min注射100nM二抗达3分钟。注射一抗直至表面呈递抗原完全饱和。在一抗和二抗注射期结束时,设置报告点“晚结合”(BL)来监测相应抗体的结合响应。计算二抗结合响应“BL2”和一抗响应“BL1”之间的商,摩尔比。使用该摩尔比作为抗原早就被一抗复合时,二抗的抗原可及性的指标。如制造商推荐的那样,通过以30μl/min注射2M盐酸胍250mMNaOH再生缓冲液达2分钟,自传感器表面完全去除复合物,接着以30μl/min注射系统缓冲液达1分钟。实施例6:在混合淋巴细胞反应(MLR)中不同抗PD-1抗体对细胞因子生成的影响3A)混合淋巴细胞反应(MLR)是一种免疫细胞测定法,其测量来自一名个体(供体X)的淋巴细胞对来自另一名个体(供体Y)的淋巴细胞的活化。使用混合淋巴细胞反应来证明阻断PD1途径对淋巴细胞效应细胞的影响。在抗PD1单抗存在或缺失下对该测定法中的T细胞测试活化和它们的IFNγ分泌。为了实施异基因MLR,使用Leukosep(GreinerBioOne,227288)通过密度梯度离心分离来自HLA类型未知的至少四名健康供体的外周血单个核细胞(PBMC)。简言之,用3倍体积的PBS稀释肝素化血液样品并在50mlLeukosep管中铺放稀释血液的25ml等分试样。于室温以800xg离心15分钟(无中断)后,收获含有淋巴细胞的级分,在PBS中清洗并直接用于功能测定法或以1.0E+07个细胞/ml在冷冻介质(10%DMSO,90%FCS)中重悬浮并在液氮中保存。如下设置个体两路MLR反应,即以1:1刺激者/响应者细胞比混合来自两名不同供体的PBMC并至少一式两份在平底96孔板中在不同浓度范围的纯化的抗PD1单克隆抗体PD1-0050,PD1-0069,PD1-0073,PD1-0078,PD1-0098,PD1-0102,PD1-0103存在或缺失下于37℃,5%CO2进行共培养6天。作为参照抗PD1抗体,合成包含纳武单抗(也称作MDX-5C4或MDX-1106)或帕姆单抗(也称作MK-3475或Org1.09A)的VH和VL域并与人IgG1主链(具有突变L234A,L235A和P329G(Kabat的EU索引))一起克隆的抗体。使用无抗体或同种型对照抗体作为阴性对照并使用重组人IL-2(20EU/ml)作为阳性对照。第6天后,自每一份培养物取出100μl培养液用于测量细胞因子。使用OptEIAELISA试剂盒(BDBiosciences)测量IFNγ的水平。结果在表10中显示(IFN-g分泌/释放)。抗PD1单克隆抗体以浓度依赖性方式促进T细胞活化和IFNγ分泌。IFNg分泌的%升高值与不添加任何阻断性单抗的MLR(基础异基因刺激诱导的IFNg值作为E-c)和有添加20EU/ml重组人IL-2的MLR(阳性对照=100%IFNg值作为E+c)的IFN-g生成相比计算并依照下面的公式计算:相对刺激[%]=((E样品-E-c)/(E+c-E-c))*100表10:异基因刺激和抗PD-1抗体处理后IFNγ分泌的百分比,与作为阳性对照的重组人IL-2处理(20EU/ml)(=100%升高)的效果比较浓度(μg/ml)1:121:1201:1200在MLR中的效果PD1-0050441369633+++PD1-006960767155+++PD1-0073431036338++PD1-007864997221++通过增强干扰素γ(IFN-g)的分泌(没有显示所有抗体的数据),数种PD1阻断性抗体PD1-0050,PD1-0069,PD1-0073,PD1-0078,PD1-0098,PD1-0102,PD1-0103展现强免疫调控活性。3B)在又一项实验中,评估嵌合PD1-0103(具有突变L234A,L235A和P329G(Kabat的EU索引)的人IgG1同种型)。用嵌合PD1-0103阻断PD1强烈增强异基因的经刺激的原代人T细胞的IFNγ分泌。嵌合PD1-0103比参照抗PD1抗体更加有力(见图1)。为了比较,使用合成包含纳武单抗(也称作MDX5C4或MDX-1106)和帕姆单抗(也称作MK-3475或Org1.09A)的VH和VL域并与人IgG1主链(具有突变L234A,L235A和P329G(Kabat的EU索引))一起克隆的参照抗PD1抗体。3C)在另外的实验中,在上文所述MLR中评估抗PD-1抗体PD1-0103的人源化变体(人源化抗体PD1-0103-0312,PD1-0103-0314,在图2和3中,还可见下文实施例9)的免疫调控活性,a)IFN释放(分泌)和b)TNFα释放(分泌)。将嵌合PD1-0103抗体及其人源化型式的效果与包含纳武单抗(也称作MDX5C4或MDX-1106)和帕姆单抗(也称作MK-3475或Org1.09A)的VH和VL域且具有人IgG1主链(具有突变L234A,L235A和P329G(Kabat的EU索引))的参照抗PD1抗体进行比较。MLR培养6天后,取出50μl上清液并使用Bio-PlexProTM人细胞因子Th1/Th2测定法(Bio-RadLaboratoriesInc.)在一份培养物中测量多种细胞因子(没有显示所有细胞因子的数据)。嵌合PD1-0103抗体及其人源化型式(PD1-0103_0312和PD1-0103_0314)与参照抗PD1抗体相比在增强T细胞活化和IFNγ分泌方面更加有力(见图2)。而且,嵌合PD1-0103抗体及其人源化变体提高抗原呈递细胞的肿瘤坏死因子α(TNFα)(见图3)和IL-12(数据未显示)分泌且增强单核细胞/巨噬细胞或抗原呈递细胞刺激T细胞的能力。实施例7:抗PD-1阻断对与异基因成熟树突细胞共培养的人CD4T细胞的细胞毒性粒酶B释放和IFN-γ分泌的影响为了进一步调查抗PD-1处理在异基因设置中的效果,我们开发了一种测定法,其中将新鲜纯化的CD4T细胞在单核细胞衍生的异基因成熟树突细胞(mDC)存在下共培养5天。1周前,经由塑胶粘附,接着去除非粘附细胞,自新鲜PBMC分离单核细胞。然后,通过将它们在含有GM-CSF(50ng/ml)和IL-4(100ng/ml)的培养基中培养5天,我们自单核细胞生成未成熟DC。为了诱导iDC成熟,我们对培养中的培养基添加TNF-α,IL-1β和IL-6(各50ng/ml)再过2天。然后,通过经由流式细胞术(LSRFortessa,BDBiosciences)测量它们II类主要组织相容性复合物(MHCII),CD80,CD83和CD86的表面表达,我们评估DC成熟。在最小限度混合淋巴细胞反应(mMLR)那天,经由微珠试剂盒(MiltenyiBiotec)自从无关供体获得的108个PBMC富集CD4T细胞。培养前,用5μM羧基-荧光素-琥珀酰亚胺基酯(CFSE)标记CD4T细胞。然后在浓度为10μg/ml(若图中没有不同标示)的阻断性抗PD1抗体(PD1-0103,嵌合PD1-0103,或人源化抗体PD1-0103-0312,PD1-0103-0313,PD1-0103-0314,PD1-0103-0315,在图4A和4B中缩写为0312,0313,0314,0315)存在或缺失下在96孔板中与成熟异-DC一起(5:1)铺放105个CD4T细胞。5天后,我们收集细胞培养物上清液,稍后用于通过ELISA(R&Dsystems)测量IFNγ水平,并将细胞在GolgiPlug(布雷菲德菌素A)和GolgiStop(莫能菌素)存在下于37℃再放置5小时。然后,清洗细胞,在表面上用抗人CD4抗体和活/死可固定染料Aqua(Invitrogen)染色,之后用固定/透化缓冲液(BDBioscience)固定/透化。我们实施了针对粒酶B(BDBioscience),IFNγ和IL-2(二者均来自eBioscience)的细胞内染色。我们还测试了不同浓度的人源化变体PD1-0103(人源化抗体PD1-0103-0312,PD1-0103-0313,PD1-0103-0314,PD1-0103-0315,在图中缩写为0312,0313,0314,0315,还可见下文实施例9)并发现它们在增强粒酶B和干扰素γ方面同等好。DP47是一种非结合性人IgG,具有Fc部分中的LALA突变以避免受到FcγR识别且用作阴性对照。结果在图4A和4B中显示。实施例8:嵌合抗体衍生物生成了嵌合PD1抗体,其通过经由PCR扩增抗PD1小鼠抗体PD1-0098,PD1-0103的可变重和轻链区并将它们作为与具有消除效应器功能的突变L234A,L235A和P329G(Kabat的EU索引))(亮氨酸234变成丙氨酸,亮氨酸235至丙氨酸,脯氨酸329至甘氨酸)的人IgG1主链/人CH1-铰链-CH2-CH3的融合蛋白克隆入重链表达载体及作为与人Cκ的融合蛋白克隆入轻链表达载体。然后将LC和HC质粒共转染入HEK293并在7天后通过抗体纯化的标准方法自上清液纯化。将嵌合PD1抗体重命名为嵌合chiPD1-0098(chiPD1-0098)和嵌合PD1-0103(chiPD1-0103)。为了比较,使用合成包含纳武单抗(也称作MDX-5C4或MDX-1106)和帕姆单抗(也称作MK-3475或Org1.09A)的VH和VL域并与人IgG1主链(具有突变L234A,L235A和P329G(Kabat的EU索引))一起克隆的参照抗PD1抗体。实施例9:抗PD1抗体PD-0103的人源化变体(huMabPD-0103)的生成,表达和纯化及表征鼠抗PD1抗体0103的VH和VL域的人源化基于鼠抗PD1抗体PD1-0103的鼠VH和VL域的氨基酸序列(SEQIDNO:43和44),生成了人源化抗PD1抗体变体。VH的人源化变体基于人种系IMGT_hVH_3_23,与具有数项突变的人J-元件种系IGHJ5-01组合(产生SEQIDNO:45)。VL的人源化变体基于人种系IMGT_hVK_4_1,IMGT_hVK_2_30,IMGT_hVK_3_11和IMGT_hVK_1_39,与人J-元件种系IGKJ1-01组合。不同突变产生人源化变体SEQIDNO:46至SEQIDNO:49。将PD1-0103的重和轻链可变区的人源化氨基酸序列逆翻译成DNA并合成(GenArt)所得cNDA,然后作为与具有消除效应器功能的LALA和PG突变(亮氨酸234变成丙氨酸,亮氨酸235变成丙氨酸,脯氨酸329变成甘氨酸)的人IgG1主链/人CH1-铰链-CH2-CH3的融合蛋白克隆入重链表达载体或作为与人Cκ的融合蛋白克隆入轻链表达载体。然后将LC和HC质粒共转染入HEK293并在7天后通过抗体纯化的标准方法自上清液纯化。所得人源化PD1抗体命名如下:表11:PD1-0103的人源化变体抗体的VH和VL序列表12:PD1-0103的人源化变体抗体的HVR序列如上文所述表征人源化PD1-0103抗体变体和亲本嵌合PD1-0103。结果在表13中显示。表13:人源化PD1-0103抗体变体和亲本嵌合PD1-0103的结果的汇总实施例10:PD-1抗体的中和效力为了测试内部生成的PD-1抗体在体外在模拟恢复受到遏制的T细胞应答中的中和效力,使用可商购的PD1/PD-L1报告物测定法(Promega)。这个系统由PD1+NFATJurkat细胞和PD-L1+CHO对应物(其还给出活化信号)组成。原理上,报告物系统基于三个步骤:(1)TCR介导的NFAT活化,(2)活化后PD-1/PD-L1轴对NFAT信号的抑制和(3)PD-1阻断性抗体对NFAT信号的恢复。材料和方法:·PD-L1培养基:PANBiotech(#P04-03609);FBS(10%)和L-Gln(4mM)·测定培养基:RPMI1640(#31870;Invitrogen),25mMHEPES,2mML-Gln,FBS(2%)·用于这项测定法的细胞(两种细胞类型均购自Promega):PD-L1+CHO细胞(批号#139147):2-3x104个细胞/96孔PD-1+NFATJurkat细胞(批号#133024:3.5x104个细胞/孔第1天,融化PD-L1+细胞,以所示细胞浓度在上文所述培养基中接种并于37℃和5%CO2培养过夜。次日,去除培养基并将PD-L1+细胞与所示浓度的制备的抗体一起温育(在测定培养基中)。平行地,融化PD-1+NFATJurkat细胞并将上文所述细胞数转移至PD-L1+细胞并共培养。于37℃和5%CO2温育6小时后,温热Bio-Glo底物至室温(在添加前1-2小时)。自温箱取出细胞培养板并调节至室温(10分钟),之后添加每孔80μlBio-Glo溶液,温育5-10分钟,之后在TecanInfinite读数仪上依照试剂盒制造商的推荐测量发光。结果可以在图5A和5B中看到,其中显示了TCR刺激后不同PD-1抗体对PD-1/PD-L1介导的NFAT信号遏制的恢复:图5A:嵌合PD1-0103显示可再现的与参照抗体相比卓越的效果。作为参照,合成包含纳武单抗(也称作MDX-5C4或MDX-1106)的VH和VL域并与人IgG1主链(具有突变L234A,L235A和P329G(Kabat的EU索引))一起克隆的抗PD1抗体。图5B:PD1-0103的四种人源化变体展现与先导抗体相似的体外效力且略微优于参照抗体。双特异性PD1/TIM3抗体实施例11A具有一个结合臂中的VH/VL域交换/替换(CrossMAbVh-VL)和CH1/CL界面中的单一带电荷氨基酸替代的结合PD1和TIM3的多特异性抗体的生成和表达在一个实施例中,通过经典的分子生物学技术如通用方法一节中描述的生成结合人PD1和人TIM3的多特异性抗体并如上文描述的在Expi293F细胞的293F中瞬时表达。多特异性1+1CrossMAbVh-Vl抗体还在WO2009/080252中描述。使用含有编码表14a中描绘的氨基酸序列的核酸的表达质粒表达多特异性抗体。表14a:具有VH/VL域交换/替换(1+1CrossMAbVh-Vl)的轻链(LC)和重链(HC)的氨基酸序列1+1抗体HC1HC2LC1LC2PD1TIM3_0389SEQIDNO:50SEQIDNO:51SEQIDNO:52SEQIDNO:53PD1TIM3-0168SEQIDNO:54SEQIDNO:55SEQIDNO:56SEQIDNO:57PD1TIM3-0476SEQIDNO:62SEQIDNO:63SEQIDNO:64SEQIDNO:65PD1TIM3-0477SEQIDNO:66SEQIDNO:67SEQIDNO:68SEQIDNO:69PD1TIM3_0166SEQIDNO:58SEQIDNO:59SEQIDNO:60SEQIDNO:61对于所有构建物,使用节入穴异二聚化技术,第一CH3域中有典型的节(T366W)替代且第二CH3域中有相应的穴替代(T366S,L368A和Y410V)(以及两个另外引入的半胱氨酸残基S354C/Y349’C)(含在上文描绘的各自相应的重链(HC)序列中)。实施例11B具有两个结合臂中的VH/VL域交换/替换(2+2CrossMAbVh-VL)和CH1/CL界面中的单一带电荷氨基酸替代的结合PD1和TIM3的多特异性抗体的生成和表达在一个实施例中,通过经典的分子生物学技术如通用方法一节中描述的生成结合人PD1和人TIM3的多特异性抗体并如上文描述的在Expi293F细胞的293F中瞬时表达。多特异性2+2CrossMAbVH-VL抗体还在WO2010/145792中描述。使用含有编码表14b中描绘的氨基酸序列的核酸的表达质粒表达多特异性抗体。表14b:具有VH/VL域交换/替换(2+2CrossMAbVh-VL)的轻链(LC)和重链(HC)的氨基酸序列2+2抗体HCLC1LC2PD1TIM3_0358SEQIDNO:70SEQIDNO:71SEQIDNO:72PD1TIM3_0359SEQIDNO:73SEQIDNO:74SEQIDNO:75PD1TIM3_0321SEQIDNO:76SEQIDNO:77SEQIDNO:80实施例11C结合PD1和TIM3的多特异性抗体的纯化和表征通过蛋白A亲和层析和大小排阻层析的组合自上清液纯化上文表达的多特异性抗体。所有多特异性抗体均能以较好的产量生成且是稳定的。对所获得的产物表征同一性(通过质谱术)和分析性特性诸如纯度(通过SDS-PAGE),单体含量和稳定性。质谱术通过去糖基化完整CrossMabs和去糖基化/经纤溶酶消化的或去糖基化/经有限LysC消化的CrossMabs的电喷射离子化质谱术(ESI-MS)来分析预期的一级结构。以1mg/ml的蛋白质浓度将VH/VLCrossMab在磷酸盐或Tris缓冲液中用N-糖苷酶F于37℃去糖基化长至17小时。纤溶酶或有限LysC(Roche)消化用100μg去糖基化VH/VLCrossMab在Tris缓冲液pH8中分别于室温实施120小时和于37℃实施40分钟。在质谱术之前,将样品经由SephadexG25柱(GEHealthcare)上的HPLC脱盐。经由配备有TriVersaNanoMate源(Advion)的maXis4GUHR-QTOFMS系统(BrukerDaltonik)上的ESI-MS测定总质量。多特异性抗体的稳定性为了评估抗体构建物的稳定性,依照下述规程评估热稳定性以及聚集肇始温度。在20mM组氨酸/组氨酸氯化物,140mMNaCl,pH6.0中以1mg/mL的浓度制备所示抗体的样品,转移入10μL微量杯阵列并用Optim1000仪器(AvactaInc.)记录静态光散射数据以及用266nm激光激发后的荧光数据,同时以0.1℃/min的速率自25℃至90℃加热样品。聚集肇始温度(Tagg)定义为散射光强度开始升高时的温度。熔解温度(Tm)定义为荧光强度对波长图的拐点。结果在表15中显示。表15实施例12抗PD1-TIM3多特异性抗体的表征结合ELISA针对huPD1的ELISA将NuncMaxisorp链霉亲合素包被板(MicroCoat#11974998001)用25μl/孔500ng/ml浓度的生物素化PD1-ECD-AviHis包被并于4℃温育过夜。清洗(3x90μl/孔PBST缓冲液)后,添加25μl递增浓度的抗PD1抗体样品并于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔1:5000稀释度的山羊抗人H+L-POD(JIR,JIR109-036-098)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche,11835033001)并温育直至OD2–3。于370/492nm进行测量。针对huTIM3的ELISA将NuncMaxisorp链霉亲合素包被板(MicroCoat#11974998001)用25μl/孔60ng/ml浓度的生物素化TIM3-ECD-AviHis包被并于4℃温育过夜。清洗(3x90μl/孔PBST缓冲液)后,添加25μl递增浓度的抗PD1抗体样品并于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔1:5000稀释度的山羊抗人H+L-POD(JIR,JIR109-036-098)并在摇床上于室温温育1小时。清洗(3x90μl/孔PBST缓冲液)后,添加25μl/孔TMB底物(Roche,11835033001)并温育直至OD2–3。于370/492nm进行测量。ELISA结果作为EC50值[nM]在表16中列出。表16:抗PD1-TIM3双特异性抗体的生化和细胞结合(ELISA)对于对Tim3呈二价的抗体(嵌合TIM3-0018,2+2PD1TIM3-0359,嵌合TIM3-0028,2+2PD1TIM3-0358)可以检测亲合结合(即两个臂的结合)。1+1CrossMab的较高EC50值源自针对所包被的抗原的单价(针对Tim3),非亲合结合。对于PD1结合没有检测到亲合效应。对于二价和四价格式,EC50值是相当的。结合Biacore结合PD1和TIM3的多特异性抗体的抗原结合特性通过评估多特异性抗体对它们各自靶抗原,即PD1和TIM3的结合。依照下述规程评估PD1结合:通过胺偶联将抗人FcIgG固定化至(Biacore)CM5传感器芯片的表面。然后捕捉样品并使huPD1-ECD结合至它们。每个分析周期后再生传感器芯片表面。最后通过将数据拟合1:1朗格缪尔相互作用模型来获取平衡常数和动力学速率常数。使用由GEHealthcar供应的胺偶联试剂盒将约10,000个响应单位(RU)的20μg/ml抗人IgG(GEHealthcare#BR-1008-39)偶联到BiacoreT200中的CM5传感器芯片的所有流动室上。样品和运行缓冲液是HBS-EP+(0.01MHEPES,0.15MNaCl,3mMEDTA,0.05%v/v表面活性剂P20,pH7.4)。流动室温度设为25℃且样品隔室温度设为12℃。用运行缓冲液引发系统。不同样品以10nM的浓度注射15秒并序贯结合至流动室2,3和4。然后一整套人PD1-ECD浓度(300nM,100nM,2×33.3nM,11.1nM,3.7nM,1.2nM和2×0nM)在每份样品上注射300秒,接着是解离时间10/600秒和两个30秒使用3MMgCl2的再生步骤,其中最后一个含有使用运行缓冲液的“注射后额外清洗”。最后,使用BiacoreT200评估软件将双重参照数据拟合1:1朗格缪尔相互作用模型。所得KD,ka和kd值在表17中显示。依照下述规程评估TIM3结合:通过胺偶联将抗人FabIgG固定化至(Biacore)CM5传感器芯片的表面。然后捕捉样品并使huTim3-ECD结合至它们。每个分析周期后再生传感器芯片表面。最后通过将数据拟合1:1朗格缪尔相互作用模型来获取平衡常数和动力学速率常数。使用由GEHealthcare供应的胺偶联试剂盒将约10,000个响应单位(RU)的20μg/ml抗人FabIgG(GEHealthcare#28-9583-25)偶联到BiacoreT200中的CM5传感器芯片的所有流动室上。样品和运行缓冲液是HBS-EP+(0.01MHEPES,0.15MNaCl,3mMEDTA,0.05%v/v表面活性剂P20,pH7.4)。流动室温度设为25℃且样品隔室温度设为12℃。用运行缓冲液引发系统。不同样品以10nM的浓度注射30秒并序贯结合至流动室2,3和4。然后一整套人Tim3-ECD浓度(600nM,200nM,2×66.7nM,22.2nM,7.4nM和2×0nM)在每份样品上注射200秒,接着是解离时间10/600秒和两个30秒使用甘氨酸HClpH2.1的再生步骤,其中最后一个含有使用运行缓冲液的“注射后额外清洗”。最后,使用BiacoreT200评估软件将双重参照数据拟合1:1朗格缪尔相互作用模型。所得KD,ka和kd值在表17中显示。结果在表17中标示。表17:PD1-Tim3双特异性抗体的亲和力样品PD1臂KD[nM]Tim3臂KD[nM]PD1TIM3-0389(0357)1.3245PD1TIM3-0358(2+2)0.3240PD1TIM3-01681.210.3PD1TIM3-0359(2+2)1.82.3PD1TIM3-04760.1332PD1TIM3-0477<0.112所有测试抗体均特异性结合两种靶物,PD1和TIM3,且展现纳摩尔范围中的抗原亲和力。实施例13:抗PD1/TIM3双特异性抗体对重组细胞的同时结合的FRET测定法这个实施例描述用于测定双特异性抗体格式对一种细胞上存在的两种不同受体的同时结合的基于细胞的TR-FRET测定法的开发。所选择的Tag-lite技术是经典的TR-FRET(时间解析荧光共振能量转移)和SNAP-tag技术(例如NewEnglandBiolabs,CISBIO)的组合,其容许用荧光供体或受体染料标记细胞表面上存在的抗原。这项技术评估的目的这项测定法意图展现抗PD1/Tim3双特异性抗体对作为由给定受体的胞外域(ECD)和标签(荧光染料能对之结合)组成的重组融合蛋白表达PD1和Tim3受体二者的细胞的同时结合。在能结合两种经标记受体的PD1-Tim3双特异性抗体存在下,蛋白质会变成密切接近以容许两种FRET染料之间的能量转移(见图6)。重组PD1+TIM3+HEK细胞的生成使用标准方法生成DNA,如Sambrooketal.,Molecularcloning:Alaboratorymanual;ColdSpringHarborLaboratoryPress,ColdSpringHarbor,NewYork,1989中描述的。为了克隆PD1和TIM3变体,接近受体的TM区插入SNAP或CLIP并除了7个氨基酸以外去除胞质域并用Flag标签替换。瞬时转染一次用15μg总量的DNA使用两种质粒在30ml培养物体积中用转染试剂293free(Novagen)和IReducedSerumMedia(LifeTechnologies)共转染HEK293细胞。简言之,如别处描述的用编码由PD1或Tim3ECD和SNAP或CLIP标签组成的融合蛋白的下述质粒瞬时转染HEK293细胞:质粒ID和提述,例如PD1-SNAP,Tim3-CLIP(还构建并表达PD1-CLIP和Tim3-SNAP的组合,但是只导致较低的FRET信号)。质粒:a)PD1-SNAP(5’至3’方向):-编码包括信号肽的人PD1胞外域(SEQIDNO:89的残基1-170(30)的核酸,-编码GGGGS间隔物(SEQIDNO:90)的核酸,-编码来自pSNAP-tag(T7)2的SNAP(没有N末端甲硫氨酸残基)(它是O6-烷基鸟嘌呤-DNA-烷基转移酶(hAGT)的人基因的突变体形式,与野生型hAGT相比,SNAP标签蛋白含有突变C26A,K125A,A127T,R128A,G131K,G132T,M134L,R135S,C150S,N157G,S159E,且在G182之后截短)(SEQIDNO:91)的核酸,-编码GGGGS间隔物(SEQIDNO:90)的核酸,-编码人PD1跨膜和胞质域(SEQIDNO:89的残基171-191)的核酸,-编码GGGGS间隔物(SEQIDNO:90)的核酸,-编码Flag标签(DYKDDDDK;SEQIDNO:92)的核酸。b)Tim3-CLIP(5’至3’方向):-编码包括信号肽的人Tim3胞外域(SEQIDNO:93的残基1-202)的核酸,-编码GGGGS间隔物(SEQIDNO:90)的核酸,-编码来自pCLIPf的CLIP(没有N末端甲硫氨酸残基)(它是O6-烷基鸟嘌呤-DNA-烷基转移酶(hAGT)的人基因的突变体形式)(SEQIDNO:94)的核酸,-编码GGGGS间隔物(SEQIDNO:90)的核酸,-编码人Tim3跨膜和胞质域(SEQIDNO:93的残基203-230)的核酸,-编码GGGGS间隔物(SEQIDNO:90)的核酸,-编码Flag标签(DYKDDDDK;SEQIDNO:92)的核酸。转染后,在摇瓶中温育细胞直至最终用于FACS(转染后24-48小时)或FRET实验(48小时后)。瞬时转染的HEK293细胞上PD1和Tim3表达的确认(FACS)瞬时转染Hek293细胞后24-48小时,对细胞分析PD1和Tim3表达:通常,将1-3x105个单一或双重转染细胞以10μg/ml在冰上染色30分钟,用PBS/2%FCS清洗两次并使用下述在FacsCantoII上分析:·PD1-FITCBiolegend329904,克隆EH12.2H7)或PD1-PE(R&D#FAB1086P)和/或·Tim3-PE(R&DFAB2365P,克隆344823)和/或使用抗PD1/TIM3双特异性抗体的细胞标记和FRET测定法的描述使用抗PD1/TIM3双特异性抗体的细胞标记和FRET测定法的描述:将转染细胞沉降并以1x106个细胞/ml的密度在Tag-lite缓冲液(Cisbio)中重悬浮。然后,将细胞在Tag-Lite缓冲液(Cisbio)中用100nMSNAP-Lumi4-Tb(Cisbio)和100nMClip-Red(Cisbio)于37℃染色1小时。清洗并在PBS/2%FCS中重悬浮后,将约50,000个细胞(在50μl体积中)接种入96孔平底白色板(Costar)并以0.001-10nM的终浓度将对照(例如单特异性,同种型参照)或双特异性抗体添加至细胞。在一些实验中,经由山羊抗人Fc交联亲本单克隆抗体(20nM终浓度,数据未显示)。于4℃或室温温育1小时后,用BMGPherastar读数仪或TecanInfiniteM1000Pro使用销售商提供的标准设置作为665/620nm之比测量时间解析荧光。任选地,将SNAP-Lumi4-Tb和100nMClip-Red标记细胞保存于-80℃或液氮中并为了FRET实验新鲜融化。结果:不同双特异性抗体和抗体格式关于通过FRET诱导展现的同时受体结合和交联的表征。如上文描述的处理表达PD1和TIM3的HEK细胞,经由与滴定量的不同双特异性抗体(0.12-10nM)一起温育来测量同时受体结合后的FRET信号。所有双特异性抗体均以剂量依赖性方式在表达PD-1-TIM3的细胞中诱导FRET信号。1+1格式(抗体#389和168)与2+2构建物(358+359)之间没有显著差异,如图7A中能看出的。另外,还对两种基于人源化PD1TIM3抗体的双特异性格式(#476和477)评估了它们在处理后在细胞中诱导FRET的能力。如图7B中展现的,两种构建物均在PD1+TIM3+HEK细胞中诱导显著的FRET信号,以功能性方式突显了同时结合。为了显示由双特异性抗体的同时结合诱导的FRET信号的特异性,添加只有一种特异性的单克隆IgG进行竞争。用100nMSNAP-Lumi4-Tb和100nMClip-Red标记带SNAP标签的PD1和带CLIP标签的TIM3细胞(如之前描述的)。清洗后,将标记细胞与双特异性抗PD1/TIM3抗体#0168[以所示浓度]一起于4℃温育1小时,之后用BMGPherastar读数仪于665/620nm测量时间解析荧光(黑线)。为了突显双特异性抗体处理后的FRET信号的特异性,添加抗PD1单克隆抗体(#0165,见图8A,灰色曲线)或抗Tim-3单克隆抗体(#0018,见图8B,灰色曲线)进行竞争,导致几乎完全阻止FRET信号。亲本(单特异性)抗PD-1抗体单独并不诱导FRET(点线)。在与双特异性抗体平行添加的TIM3亲本抗体(0018;灰色曲线)存在下FRET信号的诱导也得到阻止(图8B)。实施例14:抗体对不同外周血单个核细胞(PBMC)的结合结合测定法将新鲜分离的PBMC或3天多克隆活化的(板结合的抗CD3和可溶性抗CD28抗体,各1ug/ml,均来自BDPharmingen)CD4T细胞用Alexa647直接缀合的抗TIM-3或抗TIM-3/抗PD-1双特异性抗体任一于4℃染色1小时。然后清洗细胞以消除未结合的抗体并针对表面标志物于4℃染色30分钟以区分单核细胞(CD14+(BDPharmingen)),NK细胞(CD16+(eBioscience),CD56+(BioLegend)和CD3-)和T细胞(CD3+(eBioscience)),之后用BDCellFix固定。在LSRFortessa,BDBiosciences获取细胞。与抗Tim3抗体相比双特异性抗体的结果在图9A至9H中显示。实施例15:内在化实施例15A)将先前用1mg/ml板结合的抗CD3和1mg/ml可溶性抗CD28抗体培养的3天多克隆活化的CD4T细胞在抗TIM-3或抗TIM-3/抗PD-1双特异性抗体(一式两份)任一存在下于4℃温育30分钟。然后清洗细胞,分成两组,一个于37℃温育另外3小时,另一个立即用经标记二抗(eBioscience)染色,之后用BDCellFix固定。温育3小时后,将第二组细胞用经标记二抗染色,之后固定。在LSRFortessa(BDBiosciences)获取细胞并在两个组之间比较细胞表面上可检测抗体的表达水平。结果在图10A至10D中显示。双特异性1+1PD1TIM3-0166(基于嵌合PD1-0103/TIM3-0038)显示在活化的CD4+T细胞上和在活化的NK细胞上与双特异性2+2PD1TIM3-0321(也基于嵌合PD1-0103/TIM3-0038,但是具有两个针对PD的抗原结合位点和两个针对TIM3的)相比且与亲本TIM3-0038抗体相比降低的内在化。实施例15B)通过荧光共焦显微术显现抗体定位和内在化将活化的CD4阳性细胞用CMFDA(MolecularProbes,Lifetechnologies)染色,除了在用a-PD1抗体染色时以外,并在用聚L-赖氨酸(Sigma)处理的圆形盖玻片上分配。容许细胞于37℃粘附30分钟,之后直接将带荧光标签的抗体(1ug/mL:a-TIM3(chi18-A647=用AlexaA647标记的嵌合Tim3_0018),a-TIM3(chi28-A647=用AlexaA647标记的嵌合Tim3_0028),Bispec(0168-A647=用AlexaA647标记的1+1PD1TIM3_0168(基于嵌合PD1-0103/Tim3-0018))和Bispec(0389-A647=用Alexa647标记的1+1PD1TIM3_0389(基于嵌合PD1-0103/Tim3-0028))和a-PD1(0165-A488=用Alexa488标记的嵌合PD1-0103)添加入生长培养基达不同持续时间(15分钟,1小时,2小时,3小时)。使用冷PBS(Lonza)来淬灭反应及清洗掉未结合的抗体。然后将细胞固定(BDCytofix)20分钟并用清洗缓冲液(BD染色缓冲液)清洗两次。将盖玻片转移至干表面后,然后用封固介质(FluoromountG,eBioscience)将它们封固在载玻片上并在成像前在黑暗中保持于4℃。将来自高度靶定细胞的膜ROI的荧光信号的强度除以来自相同细胞的胞质ROI的荧光信号的强度,得到在方框图中展示的比。为了比较样品,使用单因素ANOVA分析(*=p<0.05;**=p<0.001)。用60倍油镜用来自Zeiss的倒置LSM700实施荧光共焦显微术。使用与显微镜偶联的Zen软件(Zeiss)收集图像。用Imaris软件(Bitplane;OxfordInstrument)实施图像分析并通过GraphPadPrism(GraphpadSoftware)实施统计分析。显示与TIM3抗体的胞内聚簇相比双特异性和PD1抗体二者中更高的膜定位的随时间的分析在图11A和11B中显示。抗PD1和Bispec0389只显示很慢的内在化,甚至在3小时后,而另一种Bispec0168的内在化更强。aTim3Ab0028显示更强的内在化,aTim3-0018显示最内在化。实施例16:经由混合淋巴细胞反应(MLR)测定法的T细胞活化混合淋巴细胞反应(MLR)是一种免疫细胞测定法,其测量来自一名个体(供体X)的淋巴细胞对来自另一名个体(供体Y)的淋巴细胞的活化。使用混合淋巴细胞反应来证明阻断PD1途径对淋巴细胞效应细胞的影响。在抗PD1/TIM3双特异性单抗存在或缺失下对该测定法中的T细胞测试活化和它们的IFNγ分泌。为了实施异基因MLR,使用Leukosep(GreinerBioOne,227288)通过密度梯度离心分离来自HLA类型未知的至少四名健康供体的外周血单个核细胞(PBMC)。简言之,用3倍体积的PBS稀释肝素化血液样品并在50mlLeukosep管中铺放稀释血液的25ml等分试样。于室温以800xg离心15分钟(无中断)后,收获含有淋巴细胞的级分,在PBS中清洗并直接用于功能测定法或以1.0E+07个细胞/ml在冷冻介质(10%DMSO,90%FCS)中重悬浮并在液氮中保存。如下设置个体两路MLR反应,即以1:1刺激者/响应者细胞比混合来自两名不同供体的PBMC并至少一式两份在平底96孔板中在不同浓度范围的纯化的双特异性PD1-TIM3抗体或它们的亲本单特异性抗体(单独地或组合地)存在或缺失下于37℃,5%CO2进行共培养6天。使用无抗体或同种型对照抗体作为阴性对照并使用重组人IL-2(20EU/ml)作为阳性对照。第6天后,自每一份培养物取出100μl培养液用于测量细胞因子。使用OptEIAELISA试剂盒(BDBiosciences)测量IFNγ的水平。结果在表18A至18D中显示(IFN-γ分泌/释放)。双特异性PD1TIM3抗体以浓度依赖性方式促进T细胞活化和IFN-γ分泌。IFNγ分泌的%升高值与不添加任何阻断性单抗的MLR(基础异基因刺激诱导的IFNγ值作为E-c)和有添加20EU/ml重组人IL-2的MLR(阳性对照=100%IFNγ值作为E+c)的IFNγ生成相比依照下面的公式计算:相对刺激[%]=((实施例-E-c)/(E+c-E-c))*100。实施了四项分开的实验:在实验1中与嵌合PD1-0103(=PD1-0165)和嵌合TIM3_0018(=Tim3-chi18)及其组合相比评估PD1-TIM3双特异性抗体1+1PD1TIM3_0168(基于嵌合PD1-0103/TIM3-0018(=AB0168)的功效。结果在图12A和表18A中显示。表18A在实验2中与嵌合PD1-0103(=PD1-0165)和嵌合TIM3_0028(=TIM3-chi28)及其组合相比评估PD1-TIM3双特异性抗体1+1PD1TIM3_0389(基于嵌合PD1-0103/TIM3-0028(=BispecAB0389)的功效。结果在图12B和表18B中显示。表18B在实验3中与嵌合嵌合PD1-0103(=PD1-0165)和抗TIM3-Ky8213(来自US2012/0189617,见抗体8213,例如实施例33)及其组合相比评估与实施例1中关于1+1CrossMab的描述类似生成的PD1-TIM3双特异性抗体1+1PD1-0103/Ky8213(基于嵌合PD1-0103/来自US2012/0189617的抗TIM3Ky8213(见抗体8213,例如实施例33))的功效。结果在图12C和表18C中显示。表18C在实验4中与PD1-TIM3双特异性抗体2+2PD1TIM3_0358基于嵌合PD1-0103/TIM3-0028(=BispecAB0358(2+2)),和嵌合PD1-0103(=PD1-0165)和嵌合TIM3_0028(=TIM3-chi28)及其组合相比评估PD1-TIM3双特异性抗体1+1PD1TIM3_0389(基于嵌合PD1-0103/TIM3-0028(=BispecAB0389(1+1)))的功效。结果在图12D和表18D中显示。表18D表19:观察到的特性/结果的汇总实施例17:抗原特异性CD4T细胞与B细胞成淋巴细胞样细胞系ARH77的共培养为了调查在表达MHCII的肿瘤细胞系存在下抗PD-1阻断对CD4T细胞的影响,我们开发了一种测定法,其中在EBV永生化B细胞成淋巴细胞细胞系(ARH77)存在下将新鲜纯化的CD4T细胞共培养5天。在最小限度混合淋巴细胞反应(mMLR)那天,经由微珠试剂盒(MiltenyiBiotec)自从健康供体获得的108个PBMC富集CD4T细胞。培养前,用5mM羧基-荧光素-琥珀酰亚胺基酯(CFSE)标记CD4T细胞。然后在浓度为10μg/ml的阻断性抗PD1抗体(人源化PD-10376,nivolumab或pembrolizumab),抗TIM3抗体(人源化抗TIM30438或Kyowa-8213)或抗PD-1/TIM3双特异性抗体(人源化0476)存在或缺失下在96孔板中与B细胞系一起(5:1)铺放105个CD4T细胞。5天后,我们收集细胞培养物上清液,用于通过ELISA(R&Dsystems)测量IFN-γ水平。如图13中所示,我们有趣地观察到与未处理CD4T细胞(短划线)相比抗PD-1处理显著提高CD4T细胞生成IFN-γ的能力。在这项测定法中,抗PD-1抗体0376在诱导CD4T细胞的IFN-γ分泌方面与基准抗体具有同等能力,而单独的抗TIM3抗体0438只具有边缘效应,即使比基准抗体Kyowa-8213更强。令人惊讶地,双特异性抗体0476在驱动CD4T细胞的IFN-γ分泌方面比亲本抗体的组合,单独的抗PD1抗体0376和基准抗体更好(P<0.01,单因素ANOVA)。实施例18:在体内PD1-TIM3双特异性抗体的增强的功效在第0天在1:1比率的Matrigel存在下用106个MKN45细胞(人胃癌细胞系,表达高水平的CEA)皮下攻击实验开始时6-8周龄的受到免疫遏制的雌性小鼠(NOG)。在第7天,分离来自健康人供体的PBMC。在磷酸盐缓冲盐水(PBS)(Gibco)中约2:1稀释人肝素化血液并转移入准备好的50mlLeucosep管,每个装有15mlHistopaque-1077(SigmaAldrich)。离心(30分钟,450xg,室温,无中断)后,用5ml移液器收集PBMC条带。将细胞转移入50ml管并用PBS清洗(以350xg离心10分钟)。重复清洗步骤(以300xg离心10分钟)。离心(10分钟,350xg)后,在RPMI培养基中重悬浮细胞。在NOG小鼠中静脉内注射107个PBMC,产生小鼠-人嵌合模型。在第10天开始每周制定疗法(媒介或用选自抗PD1(0376),Nivolumab,抗TIM3(0438)或抗PD1-TIM3(0476)的化合物处理)且通过腹膜内注射来给予。将PD1-Tim3双特异性抗体(3或10mg/kg;空心三角形)处理与等摩尔(1.5或5mg/kg)浓度的单一药剂PD1抗体(0376),Nivolumab和Tim3抗体(0438)比较。在30天时段里每2-3天以mm计用测径器测量肿瘤尺寸。在图14A和14B中,作为该组小鼠内的均值体积显示肿瘤体积的测量。所有处理显示与媒介处理组相比控制肿瘤生长的能力。仅抑制PD1(通过PD1抗体(0376)或基准Nivolumab)和仅抑制TIM3(抗体0438)在控制肿瘤生长方面引起相似的功效。这显示了通过阻断PD1或TIM3任一有可能增强抗肿瘤应答。然而,当PD1和TIM3二者受到PD1-TIM3双特异性抗体结合时,能观察到肿瘤生长抑制的升高。虽然在低浓度在双特异性抗体和另一种处理之间观察不到肿瘤生长的差异,但是在更高剂量通过PD1-TIM3双特异性抗体处理抑制PD1和TIM3二者导致对肿瘤生长的强抑制。***序列表<110>豪夫迈·罗氏有限公司(F.Hoffmann-LaRocheAG)<120>对PD1和TIM3特异性的双特异性抗体<130>P33115-WO<150>EP15188036.6<151>2015-10-02<160>94<170>PatentInversion3.5<210>1<211>9<212>PRT<213>小鼠(Musmusculus)<400>1GlyPheSerLeuSerThrSerGlyMet15<210>2<211>3<212>PRT<213>小鼠(Musmusculus)<400>2LeuAsnAsp1<210>3<211>8<212>PRT<213>小鼠(Musmusculus)<400>3AsnGlyTyrLeuTyrAlaLeuAsp15<210>4<211>6<212>PRT<213>小鼠(Musmusculus)<400>4SerSerSerValAsnTyr15<210>5<211>3<212>PRT<213>小鼠(Musmusculus)<400>5AspAlaPhe1<210>6<211>7<212>PRT<213>小鼠(Musmusculus)<400>6TrpSerSerTyrProTrpThr15<210>7<211>120<212>PRT<213>小鼠(Musmusculus)<400>7GlnValThrLeuLysGluSerGlyProGlyIleLeuGlnProSerGln151015ThrLeuArgLeuThrCysSerPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProSerGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysSerArgLeuThrIleSerLysAspThrSerAsnAsnGlnVal65707580PheLeuGlnIleAlaSerValValThrAlaAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyThrSerValThrValSerSer115120<210>8<211>106<212>PRT<213>小鼠(Musmusculus)<400>8GlnIleValLeuThrGlnSerProAlaIleMetSerAlaSerProGly151015GlnLysValThrIleThrCysSerAlaSerSerSerValAsnTyrThr202530GlnTrpTyrGlnGlnLysLeuGlySerSerProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyValProAlaArgPheSerGlySer505560GlyThrGlyThrSerTyrSerLeuThrIleSerSerMetGluAlaGlu65707580AspAlaAlaSerTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlyGlyThrLysLeuGluIleLys100105<210>9<211>120<212>PRT<213>小鼠(Musmusculus)<400>9GlnValThrLeuLysGluSerGlyProGlyIleLeuGlnProSerGln151015ThrLeuSerLeuThrCysSerPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProSerGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysArgArgLeuThrIleSerLysAspThrSerAsnAsnGlnVal65707580PheLeuGlnIleAlaSerValValThrAlaAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyIleSerValThrValSerSer115120<210>10<211>106<212>PRT<213>小鼠(Musmusculus)<400>10GlnIleValLeuThrGlnSerProAlaIleMetSerAlaSerProGly151015GlnLysValThrIleThrCysSerAlaSerSerSerValAsnTyrThr202530GlnTrpTyrGlnGlnLysLeuGlySerSerProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyValProAlaArgPheSerGlySer505560GlyThrGlyThrSerTyrSerLeuThrIleSerSerMetGluAlaGlu65707580AspAlaAlaSerTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlyGlyThrLysLeuGluIleLys100105<210>11<211>6<212>PRT<213>小鼠(Musmusculus)<400>11SerSerSerValGlnTyr15<210>12<211>6<212>PRT<213>小鼠(Musmusculus)<400>12SerSerSerValSerTyr15<210>13<211>120<212>PRT<213>人工的<220><223>Tim3_0016变体(0018)的VH人源化型式(=Tim3-0433)<400>13GlnIleThrLeuLysGluSerGlyProThrLeuValLysProThrGln151015ThrLeuThrLeuThrCysThrPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProProGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysSerArgLeuThrIleThrLysAspThrSerLysAsnGlnVal65707580ValLeuThrMetThrAsnMetAspProValAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyThrLeuValThrValSerSer115120<210>14<211>106<212>PRT<213>人工的<220><223>Tim3_0016变体(0018)的VL人源化型式(=Tim3-0433)<400>14GluThrThrLeuThrGlnSerProAlaPheMetSerAlaThrProGly151015AspLysValAsnIleAlaCysSerAlaSerSerSerValSerTyrThr202530GlnTrpTyrGlnGlnLysProGlyGluAlaProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyIleProProArgPheSerGlySer505560GlyTyrGlyThrAspPheThrLeuThrIleAsnAsnIleGluSerGlu65707580AspAlaAlaTyrTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlnGlyThrLysLeuGluIleLys100105<210>15<211>120<212>PRT<213>人工的<220><223>Tim3_0016变体(0018)的VH人源化型式(=Tim3-0434)<400>15GlnIleThrLeuLysGluSerGlyProThrLeuValLysProThrGln151015ThrLeuThrLeuThrCysThrPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProProGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysSerArgLeuThrIleThrLysAspThrSerLysAsnGlnVal65707580ValLeuThrMetThrAsnMetAspProValAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyThrLeuValThrValSerSer115120<210>16<211>106<212>PRT<213>人工的<220><223>Tim3_0016变体(0018)的VL人源化型式(=Tim3-0434)<400>16AspIleGlnLeuThrGlnSerProSerPheLeuSerAlaSerValGly151015AspArgValThrIleThrCysSerAlaSerSerSerValSerTyrThr202530GlnTrpTyrGlnGlnLysProGlyLysAlaProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyValProSerArgPheSerGlySer505560GlySerGlyThrGluPheThrLeuThrIleSerSerLeuGlnProGlu65707580AspPheAlaThrTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlnGlyThrLysLeuGluIleLys100105<210>17<211>7<212>PRT<213>小鼠(Musmusculus)<400>17GlyPheAsnIleLysThrThr15<210>18<211>3<212>PRT<213>小鼠(Musmusculus)<400>18AlaAspAsp1<210>19<211>8<212>PRT<213>小鼠(Musmusculus)<400>19PheGlyTyrValAlaTrpPheAla15<210>20<211>7<212>PRT<213>小鼠(Musmusculus)<400>20SerGlnSerValAspAsnTyr15<210>21<211>3<212>PRT<213>小鼠(Musmusculus)<400>21TyrAlaSer1<210>22<211>6<212>PRT<213>小鼠(Musmusculus)<400>22HisTyrSerSerProTyr15<210>23<211>119<212>PRT<213>小鼠(Musmusculus)<400>23GluValGlnLeuGlnGlnSerValAlaGluLeuValArgProGlyAla151015SerValLysLeuSerCysThrAlaSerGlyPheAsnIleLysThrThr202530TyrMetHisTrpValLysGlnArgProGluGlnGlyLeuGluTrpIle354045GlyArgIleAspProAlaAspAspAsnThrLysTyrAlaProLysPhe505560GlnGlyLysAlaThrIleThrAlaAspThrSerSerAsnThrAlaTyr65707580LeuGlnLeuSerSerLeuThrSerGluAspAlaAlaIleTyrTyrCys859095ValArgAspPheGlyTyrValAlaTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrPheSerAla115<210>24<211>107<212>PRT<213>小鼠(Musmusculus)<400>24AsnIleValMetThrProThrProLysPheLeuProValSerSerGly151015AspArgValThrMetThrCysArgAlaSerGlnSerValAspAsnTyr202530ValAlaTrpTyrGlnGlnLysProGlyGlnSerProLysLeuLeuIle354045TyrTyrAlaSerAsnArgTyrIleGlyValProAspArgPheThrGly505560SerGlySerGlyThrAspPheThrPheThrIleSerSerValGlnVal65707580GluAspLeuAlaValTyrPheCysGlnGlnHisTyrSerSerProTyr859095ThrPheGlySerGlyThrLysLeuGluIleLys100105<210>25<211>119<212>PRT<213>人工的<220><223>Tim3-0028的VH人源化型式(=Tim3-0438)<400>25GluValGlnLeuValGluSerGlyGlyGlyLeuValGlnProGlyGly151015SerLeuArgLeuSerCysAlaAlaSerGlyPheAsnIleLysThrThr202530TyrMetHisTrpValArgGlnAlaProGlyLysGlyLeuGluTrpVal354045GlyArgIleAspProAlaAspAspAsnThrLysTyrAlaProLysPhe505560GlnGlyLysAlaThrIleSerAlaAspThrSerLysAsnThrAlaTyr65707580LeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCys859095ValArgAspPheGlyTyrValAlaTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrValSerSer115<210>26<211>107<212>PRT<213>人工的<220><223>Tim3-0028的VL人源化型式(=Tim3-0438)<400>26AspIleValMetThrGlnSerProLeuSerLeuProValThrProGly151015GluProAlaSerIleSerCysArgAlaSerGlnSerValAspAsnTyr202530ValAlaTrpTyrLeuGlnLysProGlyGlnSerProGlnLeuLeuIle354045TyrTyrAlaSerAsnArgTyrIleGlyValProAspArgPheSerGly505560SerGlySerGlyThrAspPheThrLeuLysIleSerArgValGluAla65707580GluAspValGlyValTyrTyrCysGlnGlnHisTyrSerSerProTyr859095ThrPheGlyGlnGlyThrLysValGluIleLys100105<210>27<211>119<212>PRT<213>人工的<220><223>Tim3-0028的VH人源化型式(=Tim3-0443)<400>27GluValGlnLeuValGluSerGlyGlyGlyLeuValGlnProGlyGly151015SerLeuArgLeuSerCysAlaAlaSerGlyPheAsnIleLysThrThr202530TyrMetHisTrpValArgGlnAlaProGlyLysGlyLeuGluTrpVal354045GlyArgIleAspProAlaAspAspAsnThrLysTyrAlaProLysPhe505560GlnGlyLysAlaThrIleSerAlaAspThrSerLysAsnThrAlaTyr65707580LeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCys859095ValArgAspPheGlyTyrValAlaTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrPheSerSer115<210>28<211>107<212>PRT<213>人工的<220><223>Tim3-0028的VL人源化型式(=Tim3-0443)<400>28AspIleValMetThrGlnSerProLeuSerLeuProValThrProGly151015GluProAlaSerIleSerCysArgAlaSerGlnSerValAspAsnTyr202530ValAlaTrpTyrLeuGlnLysProGlyGlnSerProGlnLeuLeuIle354045TyrTyrAlaSerAsnArgTyrIleGlyValProAspArgPheSerGly505560SerGlySerGlyThrAspPheThrLeuLysIleSerArgValGluAla65707580GluAspValGlyValTyrTyrCysGlnGlnHisTyrSerSerProTyr859095ThrPheGlyGlnGlyThrLysValGluIleLys100105<210>29<211>7<212>PRT<213>小鼠(Musmusculus)<400>29GlyPheAsnIleLysAspTyr15<210>30<211>3<212>PRT<213>小鼠(Musmusculus)<400>30GluAspGly1<210>31<211>8<212>PRT<213>小鼠(Musmusculus)<400>31HisGlyTyrValGlyTrpPheAla15<210>32<211>8<212>PRT<213>小鼠(Musmusculus)<400>32AlaSerGluAsnValAspThrTyr15<210>33<211>3<212>PRT<213>小鼠(Musmusculus)<400>33GlyAlaSer1<210>34<211>6<212>PRT<213>小鼠(Musmusculus)<400>34SerTyrSerTyrProTrp15<210>35<211>119<212>PRT<213>小鼠(Musmusculus)<400>35GluValGlnLeuGlnGlnSerGlyAlaGluProLeuLysProGlyAla151015SerValLysLeuThrCysThrThrSerGlyPheAsnIleLysAspTyr202530TyrIleHisTrpValLysGlnArgSerAspGlnGlyLeuGluTrpIle354045GlyArgIleAspProGluAspGlyGluLeuIleTyrAlaProLysPhe505560GlnAspLysAlaThrIleThrValAspThrSerSerAsnIleAlaTyr65707580LeuGlnLeuAsnSerLeuThrSerGluAspThrAlaValTyrTyrCys859095SerArgAspHisGlyTyrValGlyTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrValSerAla115<210>36<211>107<212>PRT<213>小鼠(Musmusculus)<400>36AsnValValMetThrGlnSerProLysSerMetIleMetSerValGly151015GlnArgValThrLeuAsnCysLysAlaSerGluAsnValAspThrTyr202530ValSerTrpTyrGlnGlnLysProGluGlnSerProLysLeuLeuIle354045TyrGlyAlaSerAsnArgTyrThrGlyValProAspArgPheThrGly505560SerArgSerAlaThrAspPheThrLeuThrIleSerSerValGlnAla65707580GluAspLeuAlaValTyrTyrCysGlyGlnSerTyrSerTyrProTrp859095ThrPheGlyGlyGlyThrLysLeuGluPheArg100105<210>37<211>7<212>PRT<213>小鼠(Musmusculus)<400>37GlyPheSerPheSerSerTyr15<210>38<211>3<212>PRT<213>小鼠(Musmusculus)<400>38GlyGlyArg1<210>39<211>9<212>PRT<213>小鼠(Musmusculus)<400>39ThrGlyArgValTyrPheAlaLeuAsp15<210>40<211>11<212>PRT<213>小鼠(Musmusculus)<400>40SerGluSerValAspThrSerAspAsnSerPhe1510<210>41<211>3<212>PRT<213>小鼠(Musmusculus)<400>41ArgSerSer1<210>42<211>6<212>PRT<213>小鼠(Musmusculus)<400>42AsnTyrAspValProTrp15<210>43<211>120<212>PRT<213>小鼠(Musmusculus)<400>43GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSer115120<210>44<211>111<212>PRT<213>小鼠(Musmusculus)<400>44LysIleValLeuThrGlnSerProAlaSerLeuProValSerLeuGly151015GlnArgAlaThrIleSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnArgProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAla505560ArgPheSerGlySerGlySerArgThrAspPheThrLeuThrIleAsp65707580ProValGluAlaAspAspValAlaThrTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlyGlyThrLysLeuGluIleLys100105110<210>45<211>120<212>PRT<213>人工的<220><223>PD1-0103_01的人源化变体-重链可变域VH<400>45GluValGlnLeuLeuGluSerGlyGlyGlyLeuValGlnProGlyGly151015SerLeuArgLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnAlaProGlyLysGlyLeuGluTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnSerLysAsnThrLeuTyr65707580LeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrLeuValThrValSerSer115120<210>46<211>111<212>PRT<213>人工的<220><223>PD1-0103_01的人源化变体-轻链可变域VL<400>46AspIleValMetThrGlnSerProAspSerLeuAlaValSerLeuGly151015GluArgAlaThrIleAsnCysLysAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnLysProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAsp505560ArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSer65707580SerLeuGlnAlaGluAspValAlaValTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlnGlyThrLysValGluIleLys100105110<210>47<211>111<212>PRT<213>人工的<220><223>PD1-0103_02的人源化变体-轻链可变域VL<400>47AspValValMetThrGlnSerProLeuSerLeuProValThrLeuGly151015GlnProAlaSerIleSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnArgProGlyGlnSerPro354045ArgLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAsp505560ArgPheSerGlySerGlySerGlyThrAspPheThrLeuLysIleSer65707580ArgValGluAlaGluAspValGlyValTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlnGlyThrLysValGluIleLys100105110<210>48<211>111<212>PRT<213>人工的<220><223>PD1-0103_03的人源化变体-轻链可变域VL<400>48GluIleValLeuThrGlnSerProAlaThrLeuSerLeuSerProGly151015GluArgAlaThrLeuSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnLysProGlyGlnSerPro354045ArgLeuLeuIleTyrArgSerSerThrLeuGluSerGlyIleProAla505560ArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSer65707580SerLeuGluProGluAspPheAlaValTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlnGlyThrLysValGluIleLys100105110<210>49<211>111<212>PRT<213>人工的<220><223>PD1-0103_04的人源化变体-轻链可变域VL<400>49GluIleValLeuThrGlnSerProAlaThrLeuSerLeuSerProGly151015GluArgAlaThrLeuSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnLysProGlyGlnSerPro354045ArgLeuLeuIleTyrArgSerSerThrLeuGluSerGlyIleProAla505560ArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSer65707580SerLeuGluProGluAspPheAlaValTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlnGlyThrLysValGluIleLys100105110<210>50<211>443<212>PRT<213>人工的<220><223>1+1PD1TIM3_0389(基于嵌合PD1-0103/Tim3-0028)的重链1<400>50LysIleValLeuThrGlnSerProAlaSerLeuProValSerLeuGly151015GlnArgAlaThrIleSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnArgProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAla505560ArgPheSerGlySerGlySerArgThrAspPheThrLeuThrIleAsp65707580ProValGluAlaAspAspValAlaThrTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlyGlyThrLysLeuGluIleLysSer100105110SerAlaSerThrLysGlyProSerValPheProLeuAlaProSerSer115120125LysSerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAsp130135140TyrPheProGluProValThrValSerTrpAsnSerGlyAlaLeuThr145150155160SerGlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyr165170175SerLeuSerSerValValThrValProSerSerSerLeuGlyThrGln180185190ThrTyrIleCysAsnValAsnHisLysProSerAsnThrLysValAsp195200205LysLysValGluProLysSerCysAspLysThrHisThrCysProPro210215220CysProAlaProGluAlaAlaGlyGlyProSerValPheLeuPhePro225230235240ProLysProLysAspThrLeuMetIleSerArgThrProGluValThr245250255CysValValValAspValSerHisGluAspProGluValLysPheAsn260265270TrpTyrValAspGlyValGluValHisAsnAlaLysThrLysProArg275280285GluGluGlnTyrAsnSerThrTyrArgValValSerValLeuThrVal290295300LeuHisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSer305310315320AsnLysAlaLeuGlyAlaProIleGluLysThrIleSerLysAlaLys325330335GlyGlnProArgGluProGlnValTyrThrLeuProProCysArgAsp340345350GluLeuThrLysAsnGlnValSerLeuTrpCysLeuValLysGlyPhe355360365TyrProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGlu370375380AsnAsnTyrLysThrThrProProValLeuAspSerAspGlySerPhe385390395400PheLeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGly405410415AsnValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyr420425430ThrGlnLysSerLeuSerLeuSerProGlyLys435440<210>51<211>449<212>PRT<213>人工的<220><223>1+1PD1TIM3_0389的重链2<400>51GluValGlnLeuGlnGlnSerValAlaGluLeuValArgProGlyAla151015SerValLysLeuSerCysThrAlaSerGlyPheAsnIleLysThrThr202530TyrMetHisTrpValLysGlnArgProGluGlnGlyLeuGluTrpIle354045GlyArgIleAspProAlaAspAspAsnThrLysTyrAlaProLysPhe505560GlnGlyLysAlaThrIleThrAlaAspThrSerSerAsnThrAlaTyr65707580LeuGlnLeuSerSerLeuThrSerGluAspAlaAlaIleTyrTyrCys859095ValArgAspPheGlyTyrValAlaTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrPheSerAlaAlaSerThrLysGlyProSerValPhe115120125ProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeu130135140GlyCysLeuValGluAspTyrPheProGluProValThrValSerTrp145150155160AsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaValLeu165170175GlnSerSerGlyLeuTyrSerLeuSerSerValValThrValProSer180185190SerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysPro195200205SerAsnThrLysValAspGluLysValGluProLysSerCysAspLys210215220ThrHisThrCysProProCysProAlaProGluAlaAlaGlyGlyPro225230235240SerValPheLeuPheProProLysProLysAspThrLeuMetIleSer245250255ArgThrProGluValThrCysValValValAspValSerHisGluAsp260265270ProGluValLysPheAsnTrpTyrValAspGlyValGluValHisAsn275280285AlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArgVal290295300ValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLysGlu305310315320TyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGluLys325330335ThrIleSerLysAlaLysGlyGlnProArgGluProGlnValCysThr340345350LeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeuSer355360365CysAlaValLysGlyPheTyrProSerAspIleAlaValGluTrpGlu370375380SerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProValLeu385390395400AspSerAspGlySerPhePheLeuValSerLysLeuThrValAspLys405410415SerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHisGlu420425430AlaLeuHisAsnArgPheThrGlnLysSerLeuSerLeuSerProGly435440445Lys<210>52<211>227<212>PRT<213>人工的<220><223>1+1PD1TIM3_0389的轻链1<400>52GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>53<211>214<212>PRT<213>人工的<220><223>1+1PD1TIM3_0389的轻链2<400>53AsnIleValMetThrProThrProLysPheLeuProValSerSerGly151015AspArgValThrMetThrCysArgAlaSerGlnSerValAspAsnTyr202530ValAlaTrpTyrGlnGlnLysProGlyGlnSerProLysLeuLeuIle354045TyrTyrAlaSerAsnArgTyrIleGlyValProAspArgPheThrGly505560SerGlySerGlyThrAspPheThrPheThrIleSerSerValGlnVal65707580GluAspLeuAlaValTyrPheCysGlnGlnHisTyrSerSerProTyr859095ThrPheGlySerGlyThrLysLeuGluIleLysArgThrValAlaAla100105110ProSerValPheIlePheProProSerAspArgLysLeuLysSerGly115120125ThrAlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAla130135140LysValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGln145150155160GluSerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSer165170175SerThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyr180185190AlaCysGluValThrHisGlnGlyLeuSerSerProValThrLysSer195200205PheAsnArgGlyGluCys210<210>54<211>443<212>PRT<213>人工的<220><223>1+1PD1TIM3_0168(基于嵌合PD1-0103/Tim3-0018)的重链1<400>54LysIleValLeuThrGlnSerProAlaSerLeuProValSerLeuGly151015GlnArgAlaThrIleSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnArgProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAla505560ArgPheSerGlySerGlySerArgThrAspPheThrLeuThrIleAsp65707580ProValGluAlaAspAspValAlaThrTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlyGlyThrLysLeuGluIleLysSer100105110SerAlaSerThrLysGlyProSerValPheProLeuAlaProSerSer115120125LysSerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAsp130135140TyrPheProGluProValThrValSerTrpAsnSerGlyAlaLeuThr145150155160SerGlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyr165170175SerLeuSerSerValValThrValProSerSerSerLeuGlyThrGln180185190ThrTyrIleCysAsnValAsnHisLysProSerAsnThrLysValAsp195200205LysLysValGluProLysSerCysAspLysThrHisThrCysProPro210215220CysProAlaProGluAlaAlaGlyGlyProSerValPheLeuPhePro225230235240ProLysProLysAspThrLeuMetIleSerArgThrProGluValThr245250255CysValValValAspValSerHisGluAspProGluValLysPheAsn260265270TrpTyrValAspGlyValGluValHisAsnAlaLysThrLysProArg275280285GluGluGlnTyrAsnSerThrTyrArgValValSerValLeuThrVal290295300LeuHisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSer305310315320AsnLysAlaLeuGlyAlaProIleGluLysThrIleSerLysAlaLys325330335GlyGlnProArgGluProGlnValTyrThrLeuProProCysArgAsp340345350GluLeuThrLysAsnGlnValSerLeuTrpCysLeuValLysGlyPhe355360365TyrProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGlu370375380AsnAsnTyrLysThrThrProProValLeuAspSerAspGlySerPhe385390395400PheLeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGly405410415AsnValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyr420425430ThrGlnLysSerLeuSerLeuSerProGlyLys435440<210>55<211>450<212>PRT<213>人工的<220><223>1+1PD1TIM3_0168的重链2<400>55GlnValThrLeuLysGluSerGlyProGlyIleLeuGlnProSerGln151015ThrLeuSerLeuThrCysSerPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProSerGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysArgArgLeuThrIleSerLysAspThrSerAsnAsnGlnVal65707580PheLeuGlnIleAlaSerValValThrAlaAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyIleSerValThrValSerSerAlaSerThrLysGlyProSerVal115120125PheProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAla130135140LeuGlyCysLeuValGluAspTyrPheProGluProValThrValSer145150155160TrpAsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaVal165170175LeuGlnSerSerGlyLeuTyrSerLeuSerSerValValThrValPro180185190SerSerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLys195200205ProSerAsnThrLysValAspGluLysValGluProLysSerCysAsp210215220LysThrHisThrCysProProCysProAlaProGluAlaAlaGlyGly225230235240ProSerValPheLeuPheProProLysProLysAspThrLeuMetIle245250255SerArgThrProGluValThrCysValValValAspValSerHisGlu260265270AspProGluValLysPheAsnTrpTyrValAspGlyValGluValHis275280285AsnAlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArg290295300ValValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLys305310315320GluTyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGlu325330335LysThrIleSerLysAlaLysGlyGlnProArgGluProGlnValCys340345350ThrLeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeu355360365SerCysAlaValLysGlyPheTyrProSerAspIleAlaValGluTrp370375380GluSerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProVal385390395400LeuAspSerAspGlySerPhePheLeuValSerLysLeuThrValAsp405410415LysSerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHis420425430GluAlaLeuHisAsnArgPheThrGlnLysSerLeuSerLeuSerPro435440445GlyLys450<210>56<211>227<212>PRT<213>人工的<220><223>1+1PD1TIM3_0168的轻链1<400>56GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>57<211>213<212>PRT<213>人工的<220><223>1+1PD1TIM3_0168的轻链2<400>57GlnIleValLeuThrGlnSerProAlaIleMetSerAlaSerProGly151015GlnLysValThrIleThrCysSerAlaSerSerSerValAsnTyrThr202530GlnTrpTyrGlnGlnLysLeuGlySerSerProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyValProAlaArgPheSerGlySer505560GlyThrGlyThrSerTyrSerLeuThrIleSerSerMetGluAlaGlu65707580AspAlaAlaSerTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlyGlyThrLysLeuGluIleLysArgThrValAlaAlaPro100105110SerValPheIlePheProProSerAspArgLysLeuLysSerGlyThr115120125AlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLys130135140ValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGlu145150155160SerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSer165170175ThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAla180185190CysGluValThrHisGlnGlyLeuSerSerProValThrLysSerPhe195200205AsnArgGlyGluCys210<210>58<211>443<212>PRT<213>人工的<220><223>1+1PD1TIM3_0166:(基于嵌合PD1-0103/Tim3-0038)的重链1<400>58LysIleValLeuThrGlnSerProAlaSerLeuProValSerLeuGly151015GlnArgAlaThrIleSerCysArgAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnArgProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAla505560ArgPheSerGlySerGlySerArgThrAspPheThrLeuThrIleAsp65707580ProValGluAlaAspAspValAlaThrTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlyGlyThrLysLeuGluIleLysSer100105110SerAlaSerThrLysGlyProSerValPheProLeuAlaProSerSer115120125LysSerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAsp130135140TyrPheProGluProValThrValSerTrpAsnSerGlyAlaLeuThr145150155160SerGlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyr165170175SerLeuSerSerValValThrValProSerSerSerLeuGlyThrGln180185190ThrTyrIleCysAsnValAsnHisLysProSerAsnThrLysValAsp195200205LysLysValGluProLysSerCysAspLysThrHisThrCysProPro210215220CysProAlaProGluAlaAlaGlyGlyProSerValPheLeuPhePro225230235240ProLysProLysAspThrLeuMetIleSerArgThrProGluValThr245250255CysValValValAspValSerHisGluAspProGluValLysPheAsn260265270TrpTyrValAspGlyValGluValHisAsnAlaLysThrLysProArg275280285GluGluGlnTyrAsnSerThrTyrArgValValSerValLeuThrVal290295300LeuHisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSer305310315320AsnLysAlaLeuGlyAlaProIleGluLysThrIleSerLysAlaLys325330335GlyGlnProArgGluProGlnValTyrThrLeuProProCysArgAsp340345350GluLeuThrLysAsnGlnValSerLeuTrpCysLeuValLysGlyPhe355360365TyrProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGlu370375380AsnAsnTyrLysThrThrProProValLeuAspSerAspGlySerPhe385390395400PheLeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGly405410415AsnValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyr420425430ThrGlnLysSerLeuSerLeuSerProGlyLys435440<210>59<211>449<212>PRT<213>人工的<220><223>1+1PD1TIM3_0166的重链2<400>59GluValGlnLeuGlnGlnSerGlyAlaGluProLeuLysProGlyAla151015SerValLysLeuThrCysThrThrSerGlyPheAsnIleLysAspTyr202530TyrIleHisTrpValLysGlnArgSerAspGlnGlyLeuGluTrpIle354045GlyArgIleAspProGluAspGlyGluLeuIleTyrAlaProLysPhe505560GlnAspLysAlaThrIleThrValAspThrSerSerAsnIleAlaTyr65707580LeuGlnLeuAsnSerLeuThrSerGluAspThrAlaValTyrTyrCys859095SerArgAspHisGlyTyrValGlyTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrValSerAlaAlaSerThrLysGlyProSerValPhe115120125ProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeu130135140GlyCysLeuValGluAspTyrPheProGluProValThrValSerTrp145150155160AsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaValLeu165170175GlnSerSerGlyLeuTyrSerLeuSerSerValValThrValProSer180185190SerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysPro195200205SerAsnThrLysValAspGluLysValGluProLysSerCysAspLys210215220ThrHisThrCysProProCysProAlaProGluAlaAlaGlyGlyPro225230235240SerValPheLeuPheProProLysProLysAspThrLeuMetIleSer245250255ArgThrProGluValThrCysValValValAspValSerHisGluAsp260265270ProGluValLysPheAsnTrpTyrValAspGlyValGluValHisAsn275280285AlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArgVal290295300ValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLysGlu305310315320TyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGluLys325330335ThrIleSerLysAlaLysGlyGlnProArgGluProGlnValCysThr340345350LeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeuSer355360365CysAlaValLysGlyPheTyrProSerAspIleAlaValGluTrpGlu370375380SerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProValLeu385390395400AspSerAspGlySerPhePheLeuValSerLysLeuThrValAspLys405410415SerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHisGlu420425430AlaLeuHisAsnArgPheThrGlnLysSerLeuSerLeuSerProGly435440445Lys<210>60<211>227<212>PRT<213>人工的<220><223>1+1PD1TIM3_0166的轻链1<400>60GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>61<211>214<212>PRT<213>人工的<220><223>1+1PD1TIM3_0166的轻链2<400>61AsnValValMetThrGlnSerProLysSerMetIleMetSerValGly151015GlnArgValThrLeuAsnCysLysAlaSerGluAsnValAspThrTyr202530ValSerTrpTyrGlnGlnLysProGluGlnSerProLysLeuLeuIle354045TyrGlyAlaSerAsnArgTyrThrGlyValProAspArgPheThrGly505560SerArgSerAlaThrAspPheThrLeuThrIleSerSerValGlnAla65707580GluAspLeuAlaValTyrTyrCysGlyGlnSerTyrSerTyrProTrp859095ThrPheGlyGlyGlyThrLysLeuGluPheArgArgThrValAlaAla100105110ProSerValPheIlePheProProSerAspArgLysLeuLysSerGly115120125ThrAlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAla130135140LysValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGln145150155160GluSerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSer165170175SerThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyr180185190AlaCysGluValThrHisGlnGlyLeuSerSerProValThrLysSer195200205PheAsnArgGlyGluCys210<210>62<211>443<212>PRT<213>人工的<220><223>1+1PD1TIM3_0476:(基于人源化PD1-0103_0312/Tim3-0438)的重链1<400>62AspIleValMetThrGlnSerProAspSerLeuAlaValSerLeuGly151015GluArgAlaThrIleAsnCysLysAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnLysProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAsp505560ArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSer65707580SerLeuGlnAlaGluAspValAlaValTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlnGlyThrLysValGluIleLysSer100105110SerAlaSerThrLysGlyProSerValPheProLeuAlaProSerSer115120125LysSerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAsp130135140TyrPheProGluProValThrValSerTrpAsnSerGlyAlaLeuThr145150155160SerGlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyr165170175SerLeuSerSerValValThrValProSerSerSerLeuGlyThrGln180185190ThrTyrIleCysAsnValAsnHisLysProSerAsnThrLysValAsp195200205LysLysValGluProLysSerCysAspLysThrHisThrCysProPro210215220CysProAlaProGluAlaAlaGlyGlyProSerValPheLeuPhePro225230235240ProLysProLysAspThrLeuMetIleSerArgThrProGluValThr245250255CysValValValAspValSerHisGluAspProGluValLysPheAsn260265270TrpTyrValAspGlyValGluValHisAsnAlaLysThrLysProArg275280285GluGluGlnTyrAsnSerThrTyrArgValValSerValLeuThrVal290295300LeuHisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSer305310315320AsnLysAlaLeuGlyAlaProIleGluLysThrIleSerLysAlaLys325330335GlyGlnProArgGluProGlnValTyrThrLeuProProCysArgAsp340345350GluLeuThrLysAsnGlnValSerLeuTrpCysLeuValLysGlyPhe355360365TyrProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGlu370375380AsnAsnTyrLysThrThrProProValLeuAspSerAspGlySerPhe385390395400PheLeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGly405410415AsnValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyr420425430ThrGlnLysSerLeuSerLeuSerProGlyLys435440<210>63<211>449<212>PRT<213>人工的<220><223>1+1PD1TIM3_0476的重链2<400>63GluValGlnLeuValGluSerGlyGlyGlyLeuValGlnProGlyGly151015SerLeuArgLeuSerCysAlaAlaSerGlyPheAsnIleLysThrThr202530TyrMetHisTrpValArgGlnAlaProGlyLysGlyLeuGluTrpVal354045GlyArgIleAspProAlaAspAspAsnThrLysTyrAlaProLysPhe505560GlnGlyLysAlaThrIleSerAlaAspThrSerLysAsnThrAlaTyr65707580LeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCys859095ValArgAspPheGlyTyrValAlaTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrValSerSerAlaSerThrLysGlyProSerValPhe115120125ProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeu130135140GlyCysLeuValGluAspTyrPheProGluProValThrValSerTrp145150155160AsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaValLeu165170175GlnSerSerGlyLeuTyrSerLeuSerSerValValThrValProSer180185190SerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysPro195200205SerAsnThrLysValAspGluLysValGluProLysSerCysAspLys210215220ThrHisThrCysProProCysProAlaProGluAlaAlaGlyGlyPro225230235240SerValPheLeuPheProProLysProLysAspThrLeuMetIleSer245250255ArgThrProGluValThrCysValValValAspValSerHisGluAsp260265270ProGluValLysPheAsnTrpTyrValAspGlyValGluValHisAsn275280285AlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArgVal290295300ValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLysGlu305310315320TyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGluLys325330335ThrIleSerLysAlaLysGlyGlnProArgGluProGlnValCysThr340345350LeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeuSer355360365CysAlaValLysGlyPheTyrProSerAspIleAlaValGluTrpGlu370375380SerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProValLeu385390395400AspSerAspGlySerPhePheLeuValSerLysLeuThrValAspLys405410415SerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHisGlu420425430AlaLeuHisAsnHisTyrThrGlnLysSerLeuSerLeuSerProGly435440445Lys<210>64<211>227<212>PRT<213>人工的<220><223>1+1PD1TIM3_0476的轻链1<400>64GluValGlnLeuLeuGluSerGlyGlyGlyLeuValGlnProGlyGly151015SerLeuArgLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnAlaProGlyLysGlyLeuGluTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnSerLysAsnThrLeuTyr65707580LeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrLeuValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>65<211>214<212>PRT<213>人工的<220><223>1+1PD1TIM3_0476的轻链2<400>65AspIleValMetThrGlnSerProLeuSerLeuProValThrProGly151015GluProAlaSerIleSerCysArgAlaSerGlnSerValAspAsnTyr202530ValAlaTrpTyrLeuGlnLysProGlyGlnSerProGlnLeuLeuIle354045TyrTyrAlaSerAsnArgTyrIleGlyValProAspArgPheSerGly505560SerGlySerGlyThrAspPheThrLeuLysIleSerArgValGluAla65707580GluAspValGlyValTyrTyrCysGlnGlnHisTyrSerSerProTyr859095ThrPheGlyGlnGlyThrLysValGluIleLysArgThrValAlaAla100105110ProSerValPheIlePheProProSerAspArgLysLeuLysSerGly115120125ThrAlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAla130135140LysValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGln145150155160GluSerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSer165170175SerThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyr180185190AlaCysGluValThrHisGlnGlyLeuSerSerProValThrLysSer195200205PheAsnArgGlyGluCys210<210>66<211>443<212>PRT<213>人工的<220><223>1+1PD1TIM3_0477:(基于人源化PD1-0103_0312/Tim3-0434)的重链1<400>66AspIleValMetThrGlnSerProAspSerLeuAlaValSerLeuGly151015GluArgAlaThrIleAsnCysLysAlaSerGluSerValAspThrSer202530AspAsnSerPheIleHisTrpTyrGlnGlnLysProGlyGlnSerPro354045LysLeuLeuIleTyrArgSerSerThrLeuGluSerGlyValProAsp505560ArgPheSerGlySerGlySerGlyThrAspPheThrLeuThrIleSer65707580SerLeuGlnAlaGluAspValAlaValTyrTyrCysGlnGlnAsnTyr859095AspValProTrpThrPheGlyGlnGlyThrLysValGluIleLysSer100105110SerAlaSerThrLysGlyProSerValPheProLeuAlaProSerSer115120125LysSerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAsp130135140TyrPheProGluProValThrValSerTrpAsnSerGlyAlaLeuThr145150155160SerGlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyr165170175SerLeuSerSerValValThrValProSerSerSerLeuGlyThrGln180185190ThrTyrIleCysAsnValAsnHisLysProSerAsnThrLysValAsp195200205LysLysValGluProLysSerCysAspLysThrHisThrCysProPro210215220CysProAlaProGluAlaAlaGlyGlyProSerValPheLeuPhePro225230235240ProLysProLysAspThrLeuMetIleSerArgThrProGluValThr245250255CysValValValAspValSerHisGluAspProGluValLysPheAsn260265270TrpTyrValAspGlyValGluValHisAsnAlaLysThrLysProArg275280285GluGluGlnTyrAsnSerThrTyrArgValValSerValLeuThrVal290295300LeuHisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSer305310315320AsnLysAlaLeuGlyAlaProIleGluLysThrIleSerLysAlaLys325330335GlyGlnProArgGluProGlnValTyrThrLeuProProCysArgAsp340345350GluLeuThrLysAsnGlnValSerLeuTrpCysLeuValLysGlyPhe355360365TyrProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGlu370375380AsnAsnTyrLysThrThrProProValLeuAspSerAspGlySerPhe385390395400PheLeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGly405410415AsnValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyr420425430ThrGlnLysSerLeuSerLeuSerProGlyLys435440<210>67<211>450<212>PRT<213>人工的<220><223>1+1PD1TIM3_0477的重链2<400>67GlnIleThrLeuLysGluSerGlyProThrLeuValLysProThrGln151015ThrLeuThrLeuThrCysThrPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProProGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysSerArgLeuThrIleThrLysAspThrSerLysAsnGlnVal65707580ValLeuThrMetThrAsnMetAspProValAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyThrLeuValThrValSerSerAlaSerThrLysGlyProSerVal115120125PheProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAla130135140LeuGlyCysLeuValGluAspTyrPheProGluProValThrValSer145150155160TrpAsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaVal165170175LeuGlnSerSerGlyLeuTyrSerLeuSerSerValValThrValPro180185190SerSerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLys195200205ProSerAsnThrLysValAspGluLysValGluProLysSerCysAsp210215220LysThrHisThrCysProProCysProAlaProGluAlaAlaGlyGly225230235240ProSerValPheLeuPheProProLysProLysAspThrLeuMetIle245250255SerArgThrProGluValThrCysValValValAspValSerHisGlu260265270AspProGluValLysPheAsnTrpTyrValAspGlyValGluValHis275280285AsnAlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArg290295300ValValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLys305310315320GluTyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGlu325330335LysThrIleSerLysAlaLysGlyGlnProArgGluProGlnValCys340345350ThrLeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeu355360365SerCysAlaValLysGlyPheTyrProSerAspIleAlaValGluTrp370375380GluSerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProVal385390395400LeuAspSerAspGlySerPhePheLeuValSerLysLeuThrValAsp405410415LysSerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHis420425430GluAlaLeuHisAsnHisTyrThrGlnLysSerLeuSerLeuSerPro435440445GlyLys450<210>68<211>227<212>PRT<213>人工的<220><223>1+1PD1TIM3_0477的轻链1<400>68GluValGlnLeuLeuGluSerGlyGlyGlyLeuValGlnProGlyGly151015SerLeuArgLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnAlaProGlyLysGlyLeuGluTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnSerLysAsnThrLeuTyr65707580LeuGlnMetAsnSerLeuArgAlaGluAspThrAlaValTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrLeuValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>69<211>213<212>PRT<213>人工的<220><223>1+1PD1TIM3_0477的轻链2<400>69AspIleGlnLeuThrGlnSerProSerPheLeuSerAlaSerValGly151015AspArgValThrIleThrCysSerAlaSerSerSerValSerTyrThr202530GlnTrpTyrGlnGlnLysProGlyLysAlaProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyValProSerArgPheSerGlySer505560GlySerGlyThrGluPheThrLeuThrIleSerSerLeuGlnProGlu65707580AspPheAlaThrTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlnGlyThrLysLeuGluIleLysArgThrValAlaAlaPro100105110SerValPheIlePheProProSerAspArgLysLeuLysSerGlyThr115120125AlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLys130135140ValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGlu145150155160SerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSer165170175ThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAla180185190CysGluValThrHisGlnGlyLeuSerSerProValThrLysSerPhe195200205AsnArgGlyGluCys210<210>70<211>685<212>PRT<213>人工的<220><223>2+2PD1TIM3_0358:嵌合PD1-0103/Tim3-0028的重链<400>70GluValGlnLeuGlnGlnSerValAlaGluLeuValArgProGlyAla151015SerValLysLeuSerCysThrAlaSerGlyPheAsnIleLysThrThr202530TyrMetHisTrpValLysGlnArgProGluGlnGlyLeuGluTrpIle354045GlyArgIleAspProAlaAspAspAsnThrLysTyrAlaProLysPhe505560GlnGlyLysAlaThrIleThrAlaAspThrSerSerAsnThrAlaTyr65707580LeuGlnLeuSerSerLeuThrSerGluAspAlaAlaIleTyrTyrCys859095ValArgAspPheGlyTyrValAlaTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrPheSerAlaAlaSerThrLysGlyProSerValPhe115120125ProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeu130135140GlyCysLeuValGluAspTyrPheProGluProValThrValSerTrp145150155160AsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaValLeu165170175GlnSerSerGlyLeuTyrSerLeuSerSerValValThrValProSer180185190SerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysPro195200205SerAsnThrLysValAspGluLysValGluProLysSerCysAspLys210215220ThrHisThrCysProProCysProAlaProGluAlaAlaGlyGlyPro225230235240SerValPheLeuPheProProLysProLysAspThrLeuMetIleSer245250255ArgThrProGluValThrCysValValValAspValSerHisGluAsp260265270ProGluValLysPheAsnTrpTyrValAspGlyValGluValHisAsn275280285AlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArgVal290295300ValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLysGlu305310315320TyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGluLys325330335ThrIleSerLysAlaLysGlyGlnProArgGluProGlnValTyrThr340345350LeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeuThr355360365CysLeuValLysGlyPheTyrProSerAspIleAlaValGluTrpGlu370375380SerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProValLeu385390395400AspSerAspGlySerPhePheLeuTyrSerLysLeuThrValAspLys405410415SerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHisGlu420425430AlaLeuHisAsnHisTyrThrGlnLysSerLeuSerLeuSerProGly435440445GlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerGly450455460GlyGlyGlySerLysIleValLeuThrGlnSerProAlaSerLeuPro465470475480ValSerLeuGlyGlnArgAlaThrIleSerCysArgAlaSerGluSer485490495ValAspThrSerAspAsnSerPheIleHisTrpTyrGlnGlnArgPro500505510GlyGlnSerProLysLeuLeuIleTyrArgSerSerThrLeuGluSer515520525GlyValProAlaArgPheSerGlySerGlySerArgThrAspPheThr530535540LeuThrIleAspProValGluAlaAspAspValAlaThrTyrTyrCys545550555560GlnGlnAsnTyrAspValProTrpThrPheGlyGlyGlyThrLysLeu565570575GluIleLysSerSerAlaSerThrLysGlyProSerValPheProLeu580585590AlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeuGlyCys595600605LeuValLysAspTyrPheProGluProValThrValSerTrpAsnSer610615620GlyAlaLeuThrSerGlyValHisThrPheProAlaValLeuGlnSer625630635640SerGlyLeuTyrSerLeuSerSerValValThrValProSerSerSer645650655LeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysProSerAsn660665670ThrLysValAspLysLysValGluProLysSerCysAsp675680685<210>71<211>200<212>PRT<213>人工的<220><223>2+2PD1TIM3_0358的轻链1<400>71AsnIleValMetThrProThrProLysPheLeuProValSerSerGly151015AspArgValThrMetThrCysArgAlaSerGlnSerValAspAsnTyr202530ValAlaTrpTyrGlnGlnLysProGlyGlnSerProLysLeuLeuIle354045TyrTyrAlaSerAsnArgTyrIleGlyValProAspArgPheThrGly505560SerGlySerGlyThrAspPheThrPheThrIleSerSerValGlnVal65707580GluAspLeuAlaValTyrPheCysGlnGlnHisTyrSerSerProTyr859095ThrPheGlySerGlyThrLysLeuGluIleLysArgThrValAlaAla100105110ProSerValPheIlePheProProSerAspArgLysLeuLysSerGly115120125ThrAlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAla130135140LysValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGln145150155160GluSerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSer165170175SerThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyr180185190AlaCysGluValThrHisGlnGly195200<210>72<211>227<212>PRT<213>人工的<220><223>2+2PD1TIM3_0358的轻链2<400>72GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>73<211>686<212>PRT<213>人工的<220><223>2+2PD1TIM3_0359:嵌合PD1-0103/Tim3-0018的重链<400>73GlnValThrLeuLysGluSerGlyProGlyIleLeuGlnProSerGln151015ThrLeuSerLeuThrCysSerPheSerGlyPheSerLeuSerThrSer202530GlyMetSerValGlyTrpIleArgGlnProSerGlyLysGlyLeuGlu354045TrpLeuAlaHisIleTrpLeuAsnAspAspValPhePheAsnProAla505560LeuLysArgArgLeuThrIleSerLysAspThrSerAsnAsnGlnVal65707580PheLeuGlnIleAlaSerValValThrAlaAspThrAlaThrTyrTyr859095CysValArgAlaAsnGlyTyrLeuTyrAlaLeuAspTyrTrpGlyGln100105110GlyIleSerValThrValSerSerAlaSerThrLysGlyProSerVal115120125PheProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAla130135140LeuGlyCysLeuValGluAspTyrPheProGluProValThrValSer145150155160TrpAsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaVal165170175LeuGlnSerSerGlyLeuTyrSerLeuSerSerValValThrValPro180185190SerSerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLys195200205ProSerAsnThrLysValAspGluLysValGluProLysSerCysAsp210215220LysThrHisThrCysProProCysProAlaProGluAlaAlaGlyGly225230235240ProSerValPheLeuPheProProLysProLysAspThrLeuMetIle245250255SerArgThrProGluValThrCysValValValAspValSerHisGlu260265270AspProGluValLysPheAsnTrpTyrValAspGlyValGluValHis275280285AsnAlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArg290295300ValValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLys305310315320GluTyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGlu325330335LysThrIleSerLysAlaLysGlyGlnProArgGluProGlnValTyr340345350ThrLeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeu355360365ThrCysLeuValLysGlyPheTyrProSerAspIleAlaValGluTrp370375380GluSerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProVal385390395400LeuAspSerAspGlySerPhePheLeuTyrSerLysLeuThrValAsp405410415LysSerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHis420425430GluAlaLeuHisAsnHisTyrThrGlnLysSerLeuSerLeuSerPro435440445GlyGlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySer450455460GlyGlyGlyGlySerLysIleValLeuThrGlnSerProAlaSerLeu465470475480ProValSerLeuGlyGlnArgAlaThrIleSerCysArgAlaSerGlu485490495SerValAspThrSerAspAsnSerPheIleHisTrpTyrGlnGlnArg500505510ProGlyGlnSerProLysLeuLeuIleTyrArgSerSerThrLeuGlu515520525SerGlyValProAlaArgPheSerGlySerGlySerArgThrAspPhe530535540ThrLeuThrIleAspProValGluAlaAspAspValAlaThrTyrTyr545550555560CysGlnGlnAsnTyrAspValProTrpThrPheGlyGlyGlyThrLys565570575LeuGluIleLysSerSerAlaSerThrLysGlyProSerValPhePro580585590LeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeuGly595600605CysLeuValLysAspTyrPheProGluProValThrValSerTrpAsn610615620SerGlyAlaLeuThrSerGlyValHisThrPheProAlaValLeuGln625630635640SerSerGlyLeuTyrSerLeuSerSerValValThrValProSerSer645650655SerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysProSer660665670AsnThrLysValAspLysLysValGluProLysSerCysAsp675680685<210>74<211>213<212>PRT<213>人工的<220><223>2+2PD1TIM3_0359的轻链1<400>74GlnIleValLeuThrGlnSerProAlaIleMetSerAlaSerProGly151015GlnLysValThrIleThrCysSerAlaSerSerSerValAsnTyrThr202530GlnTrpTyrGlnGlnLysLeuGlySerSerProLysLeuTrpIleTyr354045AspAlaPheLysLeuAlaProGlyValProAlaArgPheSerGlySer505560GlyThrGlyThrSerTyrSerLeuThrIleSerSerMetGluAlaGlu65707580AspAlaAlaSerTyrPheCysHisGlnTrpSerSerTyrProTrpThr859095PheGlyGlyGlyThrLysLeuGluIleLysArgThrValAlaAlaPro100105110SerValPheIlePheProProSerAspArgLysLeuLysSerGlyThr115120125AlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLys130135140ValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGlu145150155160SerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSer165170175ThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAla180185190CysGluValThrHisGlnGlyLeuSerSerProValThrLysSerPhe195200205AsnArgGlyGluCys210<210>75<211>227<212>PRT<213>人工的<220><223>2+2PD1TIM3_0359的轻链2<400>75GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>76<211>685<212>PRT<213>人工的<220><223>2+2PD1TIM3_0321:嵌合PD1-0103/Tim3-0038的重链<400>76GluValGlnLeuGlnGlnSerGlyAlaGluProLeuLysProGlyAla151015SerValLysLeuThrCysThrThrSerGlyPheAsnIleLysAspTyr202530TyrIleHisTrpValLysGlnArgSerAspGlnGlyLeuGluTrpIle354045GlyArgIleAspProGluAspGlyGluLeuIleTyrAlaProLysPhe505560GlnAspLysAlaThrIleThrValAspThrSerSerAsnIleAlaTyr65707580LeuGlnLeuAsnSerLeuThrSerGluAspThrAlaValTyrTyrCys859095SerArgAspHisGlyTyrValGlyTrpPheAlaTyrTrpGlyGlnGly100105110ThrLeuValThrValSerAlaAlaSerThrLysGlyProSerValPhe115120125ProLeuAlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeu130135140GlyCysLeuValGluAspTyrPheProGluProValThrValSerTrp145150155160AsnSerGlyAlaLeuThrSerGlyValHisThrPheProAlaValLeu165170175GlnSerSerGlyLeuTyrSerLeuSerSerValValThrValProSer180185190SerSerLeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysPro195200205SerAsnThrLysValAspGluLysValGluProLysSerCysAspLys210215220ThrHisThrCysProProCysProAlaProGluAlaAlaGlyGlyPro225230235240SerValPheLeuPheProProLysProLysAspThrLeuMetIleSer245250255ArgThrProGluValThrCysValValValAspValSerHisGluAsp260265270ProGluValLysPheAsnTrpTyrValAspGlyValGluValHisAsn275280285AlaLysThrLysProArgGluGluGlnTyrAsnSerThrTyrArgVal290295300ValSerValLeuThrValLeuHisGlnAspTrpLeuAsnGlyLysGlu305310315320TyrLysCysLysValSerAsnLysAlaLeuGlyAlaProIleGluLys325330335ThrIleSerLysAlaLysGlyGlnProArgGluProGlnValTyrThr340345350LeuProProSerArgAspGluLeuThrLysAsnGlnValSerLeuThr355360365CysLeuValLysGlyPheTyrProSerAspIleAlaValGluTrpGlu370375380SerAsnGlyGlnProGluAsnAsnTyrLysThrThrProProValLeu385390395400AspSerAspGlySerPhePheLeuTyrSerLysLeuThrValAspLys405410415SerArgTrpGlnGlnGlyAsnValPheSerCysSerValMetHisGlu420425430AlaLeuHisAsnHisTyrThrGlnLysSerLeuSerLeuSerProGly435440445GlyGlyGlyGlySerGlyGlyGlyGlySerGlyGlyGlyGlySerGly450455460GlyGlyGlySerLysIleValLeuThrGlnSerProAlaSerLeuPro465470475480ValSerLeuGlyGlnArgAlaThrIleSerCysArgAlaSerGluSer485490495ValAspThrSerAspAsnSerPheIleHisTrpTyrGlnGlnArgPro500505510GlyGlnSerProLysLeuLeuIleTyrArgSerSerThrLeuGluSer515520525GlyValProAlaArgPheSerGlySerGlySerArgThrAspPheThr530535540LeuThrIleAspProValGluAlaAspAspValAlaThrTyrTyrCys545550555560GlnGlnAsnTyrAspValProTrpThrPheGlyGlyGlyThrLysLeu565570575GluIleLysSerSerAlaSerThrLysGlyProSerValPheProLeu580585590AlaProSerSerLysSerThrSerGlyGlyThrAlaAlaLeuGlyCys595600605LeuValLysAspTyrPheProGluProValThrValSerTrpAsnSer610615620GlyAlaLeuThrSerGlyValHisThrPheProAlaValLeuGlnSer625630635640SerGlyLeuTyrSerLeuSerSerValValThrValProSerSerSer645650655LeuGlyThrGlnThrTyrIleCysAsnValAsnHisLysProSerAsn660665670ThrLysValAspLysLysValGluProLysSerCysAsp675680685<210>77<211>214<212>PRT<213>人工的<220><223>2+2PD1TIM3_0321的轻链1<400>77AsnValValMetThrGlnSerProLysSerMetIleMetSerValGly151015GlnArgValThrLeuAsnCysLysAlaSerGluAsnValAspThrTyr202530ValSerTrpTyrGlnGlnLysProGluGlnSerProLysLeuLeuIle354045TyrGlyAlaSerAsnArgTyrThrGlyValProAspArgPheThrGly505560SerArgSerAlaThrAspPheThrLeuThrIleSerSerValGlnAla65707580GluAspLeuAlaValTyrTyrCysGlyGlnSerTyrSerTyrProTrp859095ThrPheGlyGlyGlyThrLysLeuGluPheArgArgThrValAlaAla100105110ProSerValPheIlePheProProSerAspArgLysLeuLysSerGly115120125ThrAlaSerValValCysLeuLeuAsnAsnPheTyrProArgGluAla130135140LysValGlnTrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGln145150155160GluSerValThrGluGlnAspSerLysAspSerThrTyrSerLeuSer165170175SerThrLeuThrLeuSerLysAlaAspTyrGluLysHisLysValTyr180185190AlaCysGluValThrHisGlnGlyLeuSerSerProValThrLysSer195200205PheAsnArgGlyGluCys210<210>78<211>227<212>PRT<213>人工的<220><223>2+2PD1TIM3_0321的轻链2<400>78GluValIleLeuValGluSerGlyGlyGlyLeuValLysProGlyGly151015SerLeuLysLeuSerCysAlaAlaSerGlyPheSerPheSerSerTyr202530ThrMetSerTrpValArgGlnThrProGluLysArgLeuAspTrpVal354045AlaThrIleSerGlyGlyGlyArgAspIleTyrTyrProAspSerVal505560LysGlyArgPheThrIleSerArgAspAsnAlaLysAsnThrLeuTyr65707580LeuGluMetSerSerLeuMetSerGluAspThrAlaLeuTyrTyrCys859095ValLeuLeuThrGlyArgValTyrPheAlaLeuAspSerTrpGlyGln100105110GlyThrSerValThrValSerSerAlaSerValAlaAlaProSerVal115120125PheIlePheProProSerAspGluGlnLeuLysSerGlyThrAlaSer130135140ValValCysLeuLeuAsnAsnPheTyrProArgGluAlaLysValGln145150155160TrpLysValAspAsnAlaLeuGlnSerGlyAsnSerGlnGluSerVal165170175ThrGluGlnAspSerLysAspSerThrTyrSerLeuSerSerThrLeu180185190ThrLeuSerLysAlaAspTyrGluLysHisLysValTyrAlaCysGlu195200205ValThrHisGlnGlyLeuSerSerProValThrLysSerPheAsnArg210215220GlyGluCys225<210>79<211>107<212>PRT<213>人(HomoSapiens)<400>79ArgThrValAlaAlaProSerValPheIlePheProProSerAspGlu151015GlnLeuLysSerGlyThrAlaSerValValCysLeuLeuAsnAsnPhe202530TyrProArgGluAlaLysValGlnTrpLysValAspAsnAlaLeuGln354045SerGlyAsnSerGlnGluSerValThrGluGlnAspSerLysAspSer505560ThrTyrSerLeuSerSerThrLeuThrLeuSerLysAlaAspTyrGlu65707580LysHisLysValTyrAlaCysGluValThrHisGlnGlyLeuSerSer859095ProValThrLysSerPheAsnArgGlyGluCys100105<210>80<211>105<212>PRT<213>人(HomoSapiens)<400>80GlnProLysAlaAlaProSerValThrLeuPheProProSerSerGlu151015GluLeuGlnAlaAsnLysAlaThrLeuValCysLeuIleSerAspPhe202530TyrProGlyAlaValThrValAlaTrpLysAlaAspSerSerProVal354045LysAlaGlyValGluThrThrThrProSerLysGlnSerAsnAsnLys505560TyrAlaAlaSerSerTyrLeuSerLeuThrProGluGlnTrpLysSer65707580HisArgSerTyrSerCysGlnValThrHisGluGlySerThrValGlu859095LysThrValAlaProThrGluCysSer100105<210>81<211>330<212>PRT<213>人(HomoSapiens)<400>81AlaSerThrLysGlyProSerValPheProLeuAlaProSerSerLys151015SerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAspTyr202530PheProGluProValThrValSerTrpAsnSerGlyAlaLeuThrSer354045GlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyrSer505560LeuSerSerValValThrValProSerSerSerLeuGlyThrGlnThr65707580TyrIleCysAsnValAsnHisLysProSerAsnThrLysValAspLys859095LysValGluProLysSerCysAspLysThrHisThrCysProProCys100105110ProAlaProGluLeuLeuGlyGlyProSerValPheLeuPheProPro115120125LysProLysAspThrLeuMetIleSerArgThrProGluValThrCys130135140ValValValAspValSerHisGluAspProGluValLysPheAsnTrp145150155160TyrValAspGlyValGluValHisAsnAlaLysThrLysProArgGlu165170175GluGlnTyrAsnSerThrTyrArgValValSerValLeuThrValLeu180185190HisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSerAsn195200205LysAlaLeuProAlaProIleGluLysThrIleSerLysAlaLysGly210215220GlnProArgGluProGlnValTyrThrLeuProProSerArgAspGlu225230235240LeuThrLysAsnGlnValSerLeuThrCysLeuValLysGlyPheTyr245250255ProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGluAsn260265270AsnTyrLysThrThrProProValLeuAspSerAspGlySerPhePhe275280285LeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGlyAsn290295300ValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyrThr305310315320GlnLysSerLeuSerLeuSerProGlyLys325330<210>82<211>330<212>PRT<213>人(HomoSapiens)<400>82AlaSerThrLysGlyProSerValPheProLeuAlaProSerSerLys151015SerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAspTyr202530PheProGluProValThrValSerTrpAsnSerGlyAlaLeuThrSer354045GlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyrSer505560LeuSerSerValValThrValProSerSerSerLeuGlyThrGlnThr65707580TyrIleCysAsnValAsnHisLysProSerAsnThrLysValAspLys859095LysValGluProLysSerCysAspLysThrHisThrCysProProCys100105110ProAlaProGluAlaAlaGlyGlyProSerValPheLeuPheProPro115120125LysProLysAspThrLeuMetIleSerArgThrProGluValThrCys130135140ValValValAspValSerHisGluAspProGluValLysPheAsnTrp145150155160TyrValAspGlyValGluValHisAsnAlaLysThrLysProArgGlu165170175GluGlnTyrAsnSerThrTyrArgValValSerValLeuThrValLeu180185190HisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSerAsn195200205LysAlaLeuProAlaProIleGluLysThrIleSerLysAlaLysGly210215220GlnProArgGluProGlnValTyrThrLeuProProSerArgAspGlu225230235240LeuThrLysAsnGlnValSerLeuThrCysLeuValLysGlyPheTyr245250255ProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGluAsn260265270AsnTyrLysThrThrProProValLeuAspSerAspGlySerPhePhe275280285LeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGlyAsn290295300ValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyrThr305310315320GlnLysSerLeuSerLeuSerProGlyLys325330<210>83<211>330<212>PRT<213>人(HomoSapiens)<400>83AlaSerThrLysGlyProSerValPheProLeuAlaProSerSerLys151015SerThrSerGlyGlyThrAlaAlaLeuGlyCysLeuValLysAspTyr202530PheProGluProValThrValSerTrpAsnSerGlyAlaLeuThrSer354045GlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyrSer505560LeuSerSerValValThrValProSerSerSerLeuGlyThrGlnThr65707580TyrIleCysAsnValAsnHisLysProSerAsnThrLysValAspLys859095LysValGluProLysSerCysAspLysThrHisThrCysProProCys100105110ProAlaProGluAlaAlaGlyGlyProSerValPheLeuPheProPro115120125LysProLysAspThrLeuMetIleSerArgThrProGluValThrCys130135140ValValValAspValSerHisGluAspProGluValLysPheAsnTrp145150155160TyrValAspGlyValGluValHisAsnAlaLysThrLysProArgGlu165170175GluGlnTyrAsnSerThrTyrArgValValSerValLeuThrValLeu180185190HisGlnAspTrpLeuAsnGlyLysGluTyrLysCysLysValSerAsn195200205LysAlaLeuGlyAlaProIleGluLysThrIleSerLysAlaLysGly210215220GlnProArgGluProGlnValTyrThrLeuProProSerArgAspGlu225230235240LeuThrLysAsnGlnValSerLeuThrCysLeuValLysGlyPheTyr245250255ProSerAspIleAlaValGluTrpGluSerAsnGlyGlnProGluAsn260265270AsnTyrLysThrThrProProValLeuAspSerAspGlySerPhePhe275280285LeuTyrSerLysLeuThrValAspLysSerArgTrpGlnGlnGlyAsn290295300ValPheSerCysSerValMetHisGluAlaLeuHisAsnHisTyrThr305310315320GlnLysSerLeuSerLeuSerProGlyLys325330<210>84<211>327<212>PRT<213>人(HomoSapiens)<400>84AlaSerThrLysGlyProSerValPheProLeuAlaProCysSerArg151015SerThrSerGluSerThrAlaAlaLeuGlyCysLeuValLysAspTyr202530PheProGluProValThrValSerTrpAsnSerGlyAlaLeuThrSer354045GlyValHisThrPheProAlaValLeuGlnSerSerGlyLeuTyrSer505560LeuSerSerValValThrValProSerSerSerLeuGlyThrLysThr65707580TyrThrCysAsnValAspHisLysProSerAsnThrLysValAspLys859095ArgValGluSerLysTyrGlyProProCysProSerCysProAlaPro100105110GluPheLeuGlyGlyProSerValPheLeuPheProProLysProLys115120125AspThrLeuMetIleSerArgThrProGluValThrCysValValVal130135140AspValSerGlnGluAspProGluValGlnPheAsnTrpTyrValAsp145150155160GlyValGluValHisAsnAlaLysThrLysProArgGluGluGlnPhe165170175AsnSerThrTyrArgValValSerValLeuThrValLeuHisGlnAsp180185190TrpLeuAsnGlyLysGluTyrLysCysLysValSerAsnLysGlyLeu195200205ProSerSerIleGluLysThrIleSerLysAlaLysGlyGlnProArg210215220GluProGlnValTyrThrLeuProProSerGlnGluGluMetThrLys225230235240AsnGlnValSerLeuThrCysLeuValLysGlyPheTyrProSerAsp245250255IleAlaValGluTrpGluSerAsnGlyGlnProGluAsnAsnTyrLys260265270ThrThrProProValLeuAspSerAspGlySerPhePheLeuTyrSer275280285ArgLeuThrValAspLysSerArgTrpGlnGluGlyAsnValPheSer290295300CysSerValMetHisGluAlaLeuHisAsnHisTyrThrGlnLysSer305310315320LeuSerLeuSerLeuGlyLys325<210>85<211>280<212>PRT<213>人(HomoSapiens)<400>85SerGluValGluTyrArgAlaGluValGlyGlnAsnAlaTyrLeuPro151015CysPheTyrThrProAlaAlaProGlyAsnLeuValProValCysTrp202530GlyLysGlyAlaCysProValPheGluCysGlyAsnValValLeuArg354045ThrAspGluArgAspValAsnTyrTrpThrSerArgTyrTrpLeuAsn505560GlyAspPheArgLysGlyAspValSerLeuThrIleGluAsnValThr65707580LeuAlaAspSerGlyIleTyrCysCysArgIleGlnIleProGlyIle859095MetAsnAspGluLysPheAsnLeuLysLeuValIleLysProAlaLys100105110ValThrProAlaProThrArgGlnArgAspPheThrAlaAlaPhePro115120125ArgMetLeuThrThrArgGlyHisGlyProAlaGluThrGlnThrLeu130135140GlySerLeuProAspIleAsnLeuThrGlnIleSerThrLeuAlaAsn145150155160GluLeuArgAspSerArgLeuAlaAsnAspLeuArgAspSerGlyAla165170175ThrIleArgIleGlyIleTyrIleGlyAlaGlyIleCysAlaGlyLeu180185190AlaLeuAlaLeuIlePheGlyAlaLeuIlePheLysTrpTyrSerHis195200205SerLysGluLysIleGlnAsnLeuSerLeuIleSerLeuAlaAsnLeu210215220ProProSerGlyLeuAlaAsnAlaValAlaGluGlyIleArgSerGlu225230235240GluAsnIleTyrThrIleGluGluAsnValTyrGluValGluGluPro245250255AsnGluTyrTyrCysTyrValSerSerArgGlnGlnProSerGlnPro260265270LeuGlyCysArgPheAlaMetPro275280<210>86<211>181<212>PRT<213>人(HomoSapiens)<400>86SerGluValGluTyrArgAlaGluValGlyGlnAsnAlaTyrLeuPro151015CysPheTyrThrProAlaAlaProGlyAsnLeuValProValCysTrp202530GlyLysGlyAlaCysProValPheGluCysGlyAsnValValLeuArg354045ThrAspGluArgAspValAsnTyrTrpThrSerArgTyrTrpLeuAsn505560GlyAspPheArgLysGlyAspValSerLeuThrIleGluAsnValThr65707580LeuAlaAspSerGlyIleTyrCysCysArgIleGlnIleProGlyIle859095MetAsnAspGluLysPheAsnLeuLysLeuValIleLysProAlaLys100105110ValThrProAlaProThrArgGlnArgAspPheThrAlaAlaPhePro115120125ArgMetLeuThrThrArgGlyHisGlyProAlaGluThrGlnThrLeu130135140GlySerLeuProAspIleAsnLeuThrGlnIleSerThrLeuAlaAsn145150155160GluLeuArgAspSerArgLeuAlaAsnAspLeuArgAspSerGlyAla165170175ThrIleArgIleGly180<210>87<211>268<212>PRT<213>人(HomoSapiens)<400>87ProGlyTrpPheLeuAspSerProAspArgProTrpAsnProProThr151015PheSerProAlaLeuLeuValValThrGluGlyAspAsnAlaThrPhe202530ThrCysSerPheSerAsnThrSerGluSerPheValLeuAsnTrpTyr354045ArgMetSerProSerAsnGlnThrAspLysLeuAlaAlaPheProGlu505560AspArgSerGlnProGlyGlnAspCysArgPheArgValThrGlnLeu65707580ProAsnGlyArgAspPheHisMetSerValValArgAlaArgArgAsn859095AspSerGlyThrTyrLeuCysGlyAlaIleSerLeuAlaProLysAla100105110GlnIleLysGluSerLeuArgAlaGluLeuArgValThrGluArgArg115120125AlaGluValProThrAlaHisProSerProSerProArgProAlaGly130135140GlnPheGlnThrLeuValValGlyValValGlyGlyLeuLeuGlySer145150155160LeuValLeuLeuValTrpValLeuAlaValIleCysSerArgAlaAla165170175ArgGlyThrIleGlyAlaArgArgThrGlyGlnProLeuLysGluAsp180185190ProSerAlaValProValPheSerValAspTyrGlyGluLeuAspPhe195200205GlnTrpArgGluLysThrProGluProProValProCysValProGlu210215220GlnThrGluTyrAlaThrIleValPheProSerGlyMetGlyThrSer225230235240SerProAlaArgArgGlySerAlaAspGlyProArgSerAlaGlnPro245250255LeuArgProGluAspGlyHisCysSerTrpProLeu260265<210>88<211>150<212>PRT<213>人(HomoSapiens)<400>88ProGlyTrpPheLeuAspSerProAspArgProTrpAsnProProThr151015PheSerProAlaLeuLeuValValThrGluGlyAspAsnAlaThrPhe202530ThrCysSerPheSerAsnThrSerGluSerPheValLeuAsnTrpTyr354045ArgMetSerProSerAsnGlnThrAspLysLeuAlaAlaPheProGlu505560AspArgSerGlnProGlyGlnAspCysArgPheArgValThrGlnLeu65707580ProAsnGlyArgAspPheHisMetSerValValArgAlaArgArgAsn859095AspSerGlyThrTyrLeuCysGlyAlaIleSerLeuAlaProLysAla100105110GlnIleLysGluSerLeuArgAlaGluLeuArgValThrGluArgArg115120125AlaGluValProThrAlaHisProSerProSerProArgProAlaGly130135140GlnPheGlnThrLeuVal145150<210>89<211>288<212>PRT<213>人(HomoSapiens)<400>89MetGlnIleProGlnAlaProTrpProValValTrpAlaValLeuGln151015LeuGlyTrpArgProGlyTrpPheLeuAspSerProAspArgProTrp202530AsnProProThrPheSerProAlaLeuLeuValValThrGluGlyAsp354045AsnAlaThrPheThrCysSerPheSerAsnThrSerGluSerPheVal505560LeuAsnTrpTyrArgMetSerProSerAsnGlnThrAspLysLeuAla65707580AlaPheProGluAspArgSerGlnProGlyGlnAspCysArgPheArg859095ValThrGlnLeuProAsnGlyArgAspPheHisMetSerValValArg100105110AlaArgArgAsnAspSerGlyThrTyrLeuCysGlyAlaIleSerLeu115120125AlaProLysAlaGlnIleLysGluSerLeuArgAlaGluLeuArgVal130135140ThrGluArgArgAlaGluValProThrAlaHisProSerProSerPro145150155160ArgProAlaGlyGlnPheGlnThrLeuValValGlyValValGlyGly165170175LeuLeuGlySerLeuValLeuLeuValTrpValLeuAlaValIleCys180185190SerArgAlaAlaArgGlyThrIleGlyAlaArgArgThrGlyGlnPro195200205LeuLysGluAspProSerAlaValProValPheSerValAspTyrGly210215220GluLeuAspPheGlnTrpArgGluLysThrProGluProProValPro225230235240CysValProGluGlnThrGluTyrAlaThrIleValPheProSerGly245250255MetGlyThrSerSerProAlaArgArgGlySerAlaAspGlyProArg260265270SerAlaGlnProLeuArgProGluAspGlyHisCysSerTrpProLeu275280285<210>90<211>5<212>PRT<213>人工序列<220><223>GGGGS间隔物<400>90GlyGlyGlyGlySer15<210>91<211>192<212>PRT<213>人工序列<220><223>SNAP标签<400>91AspLysAspCysGluMetLysArgThrThrLeuAspSerProLeuGly151015LysLeuGluLeuSerGlyCysGluGlnGlyLeuHisGluIleLysLeu202530LeuGlyLysGlyThrSerAlaAlaAspAlaValGluValProAlaPro354045AlaAlaValLeuGlyGlyProGluProLeuMetGlnAlaThrAlaTrp505560LeuAsnAlaTyrPheHisGlnProGluAlaIleGluGluPheProVal65707580ProAlaLeuHisHisProValPheGlnGlnGluSerPheThrArgGln859095ValLeuTrpLysLeuLeuLysValValLysPheGlyGluValIleSer100105110TyrGlnGlnLeuAlaAlaLeuAlaGlyAsnProAlaAlaThrAlaAla115120125ValLysThrAlaLeuSerGlyAsnProValProIleLeuIleProCys130135140HisArgValValSerSerSerGlyAlaValGlyGlyTyrGluGlyGly145150155160LeuAlaValLysGluTrpLeuLeuAlaHisGluGlyHisArgLeuGly165170175LysProGlyLeuGlyProAlaGlyGlySerProGlyLeuGluValAsn180185190<210>92<211>8<212>PRT<213>人工序列<220><223>Flag标签<400>92AspTyrLysAspAspAspAspLys15<210>93<211>300<212>PRT<213>人(HomoSapiens)<400>93MetPheSerHisLeuProPheAspCysValLeuLeuLeuLeuLeuLeu151015LeuLeuThrArgSerSerGluValGluTyrArgAlaGluValGlyGln202530AsnAlaTyrLeuProCysPheTyrThrProAlaAlaProGlyAsnLeu354045ValProValCysTrpGlyLysGlyAlaCysProValPheGluCysGly505560AsnValValLeuArgThrAspGluArgAspValAsnTyrTrpThrSer65707580ArgTyrTrpLeuAsnGlyAspPheArgLysGlyAspValSerLeuThr859095IleGluAsnValThrLeuAlaAspSerGlyIleTyrCysCysArgIle100105110GlnIleProGlyIleMetAsnAspGluLysPheAsnLeuLysLeuVal115120125IleLysProAlaLysValThrProAlaProThrArgGlnArgAspPhe130135140ThrAlaAlaPheProArgMetLeuThrThrArgGlyHisGlyProAla145150155160GluThrGlnThrLeuGlySerLeuProAspIleAsnLeuThrGlnIle165170175SerThrLeuAlaAsnGluLeuArgAspSerArgLeuAlaAsnAspLeu180185190ArgAspSerGlyAlaThrIleArgIleGlyIleTyrIleGlyAlaGly195200205IleCysAlaGlyLeuAlaLeuAlaLeuIlePheGlyAlaLeuIlePhe210215220LysTrpTyrSerHisSerLysGluLysIleGlnAsnLeuSerLeuIle225230235240SerLeuAlaAsnLeuProProSerGlyLeuAlaAsnAlaValAlaGlu245250255GlyIleArgSerGluGluAsnIleTyrThrIleGluGluAsnValTyr260265270GluValGluGluProAsnGluTyrTyrCysTyrValSerSerArgGln275280285GlnProSerGlnProLeuGlyCysArgPheAlaMet290295300<210>94<211>181<212>PRT<213>人工序列<220><223>Clip标签<400>94AspLysAspCysGluMetLysArgThrThrLeuAspSerProLeuGly151015LysLeuGluLeuSerGlyCysGluGlnGlyLeuHisArgIleIlePhe202530LeuGlyLysGlyThrSerAlaAlaAspAlaValGluValProAlaPro354045AlaAlaValLeuGlyGlyProGluProLeuIleGlnAlaThrAlaTrp505560LeuAsnAlaTyrPheHisGlnProGluAlaIleGluGluPheProVal65707580ProAlaLeuHisHisProValPheGlnGlnGluSerPheThrArgGln859095ValLeuTrpLysLeuLeuLysValValLysPheGlyGluValIleSer100105110GluSerHisLeuAlaAlaLeuValGlyAsnProAlaAlaThrAlaAla115120125ValAsnThrAlaLeuAspGlyAsnProValProIleLeuIleProCys130135140HisArgValValGlnGlyAspSerAspValGlyProTyrLeuGlyGly145150155160LeuAlaValLysGluTrpLeuLeuAlaHisGluGlyHisArgLeuGly165170175LysProGlyLeuGly180当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺