来源于耐甲氧西林金黄色葡萄球菌噬菌体的宽谱内溶素及其应用的制作方法
本发明属于生物技术领域,具体涉及一种来源于耐甲氧西林金黄色葡萄球菌噬菌体的内溶素及其应用。
背景技术:
金黄色葡萄球菌(Staphylococcus aureus)为革兰氏阳性菌,是一种重要的食源性致病菌,广泛存在于自然界中,空气、土壤、水及食具上。食用了被该菌污染的食物可能会引起全身非特异性炎症反应,如恶心、呕吐、恶寒、胃部痉挛、肺炎、伪膜性肠炎、心包炎等疾病,严重时甚至会导致败血症、脓毒症等全身性感染,对人类的健康造成极大地潜在威胁。尤其是近几年来,金黄色葡萄球菌中出现了可以耐受甲氧西林的“超级细菌”(MRSA),感染者死亡率可达有31%,目前已经有50多万人死于“超级细菌”。
噬菌体内溶素作为一种新型的生物制剂,具有专一性强、安全性能高、不易产生抗性的特点,可以用于金黄色葡萄球菌的生物防控。尽管在1957年噬菌体内溶素裂解细菌的能力就被首次报道,但直到2001年Nelson等才证实纯化的重组内溶素可以作为抗菌剂有效控制大鼠感染A族链球菌。到目前为止,内溶素已被用来防控和检测食品中的食源性致病菌。与使用小分子抗生素用于抗菌治疗或预防相比,专一性的内溶素不易导致抗性菌株的快速出现。在细菌耐药性日益严重的现在,尤其在市场对新型抗菌剂的需求空前提高,内溶素不易产生抗性且裂解专一性的特点,使其作为新型抗菌剂具有一定的优势。
因此,为了更好地解决金黄色葡萄球菌日趋严重的耐药性及相关内溶素缺乏的问题,本发明开发出能够高效裂解金黄色葡萄球菌及某些革兰氏阴性菌的宽裂解谱内溶素或内溶素基因。
技术实现要素:
本发明提供一种来自耐甲氧西林金黄色葡萄球菌噬菌体的具有显著杀菌效果,并且具有宽裂解谱的内溶素。
本发明还提供所述的内溶素在制备杀菌剂中的应用。
本发明解决其技术问题所采用的技术方案是:
一种来源于耐甲氧西林金黄色葡萄球菌噬菌体的高效宽谱内溶素,其氨基酸序列为SEQ ID No:1所示的全部或部分氨基酸序列。经过序列结构分析,显示该酶为噬菌体中的能够裂解细菌细胞壁的裂解酶,经试验证实,该氨基酸序列形成的蛋白具有较好的杀菌活性。
编码所述的内溶素的基因,所述基因的核苷酸序列如SEQ ID No:2所示。
含有所述的基因的表达载体、工程菌或细胞系。
所述原核细胞表达载体为pET-28a。
所述工程菌为大肠杆菌BL21(DE3)。
一种所述的高效宽谱内溶素在制备杀菌剂中的应用。目前尚未有文献公开过耐甲氧西林金黄色葡萄球菌噬菌体qdsa002能够产生裂解金黄色葡萄球菌细胞壁的裂解酶,而本发明的内溶素具有该功能,对金黄色葡萄球菌及某些革兰氏阴性菌均具有有效的杀菌活性。
本发明的有益效果是:本发明的内溶素通过市场上已有的适合表达载体表达、纯化后,体外对金黄色葡萄球菌及某些革兰氏阴性菌表现出显著的杀菌效果,可用于金黄色葡萄球菌及某些革兰氏阴性菌杀菌剂的制备。
附图说明
图1为内溶素蛋白进行结构域分析预测图;
图2为内溶素蛋白在大肠杆菌中高效表达图谱,M为Protein marker;1为破碎大肠杆菌后得到的粗酶液,2为粗酶液上柱过程中的流出液,3,4为过柱过程中洗脱液,5为镍柱纯化后的蛋白。
图3为内溶素蛋白裂解耐甲氧西林金黄色葡萄球菌ATCC43300的吸光值变化图谱;
图4为溶菌酶裂解耐甲氧西林金黄色葡萄球菌ATCC43300的吸光值变化图谱;
图5为内溶素Lys qdsa002的在不同pH下的作用效果图。
具体实施方式
下面通过具体实施例,对本发明的技术方案作进一步的具体说明。应当理解,本发明的实施并不局限于下面的实施例,对本发明所做的任何形式上的变通和/或改变都将落入本发明保护范围。
在本发明中,若非特指,所有的份、百分比均为重量单位,所采用的设备和原料等均可从市场购得或是本领域常用的。下述实施例中的方法,如无特别说明,均为本领域的常规方法。
本发明试验涉及的菌株:金黄色葡萄球菌43300(保藏号为ATCC43300),购买于美国ATCC中心。
本发明涉及的金黄色葡萄球菌噬菌体,该噬菌体qdsa002保存于中国典型培养物保藏中心,地址:中国武汉、武汉大学,保藏编号:CCTCC NO:M 2015554,保藏日期为2015年9月17日,分类学命名:金黄色葡萄球菌噬菌体qdsa002(Staphylococcus aureus bacteriophage qdsa002)。
TSB培养基,北京陆桥技术有限责任公司;
LB液体培养基,北京陆桥技术有限责任公司;
质粒Pet-28a,美国默克公司;
大肠杆菌BL21(DE3),北京康为世纪公司;
Ni-IDA填料,生工生物工程(上海)股份有限公司;
异丙基-β-D-硫代半乳糖苷(IPTG),北京索莱宝科技有限公司。
溶菌酶,北京索莱宝科技有限公司。
实施例1:内溶素蛋白功能预测
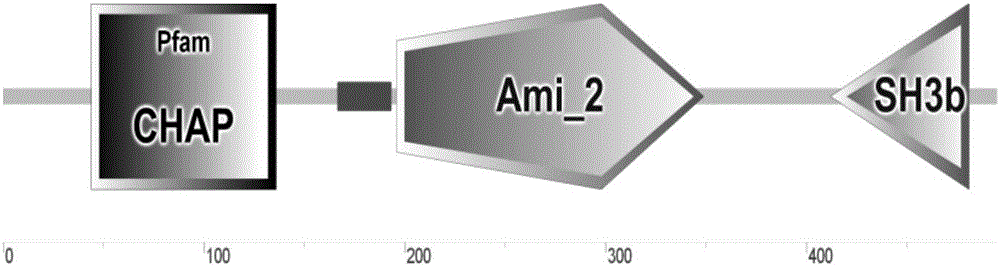
本发明人对从污水中分离出金黄色葡萄球菌噬菌体qdsa002进行全基因组测序和分析,鉴定该噬菌体ORF84编码的蛋白在氨基酸序列上与多株金黄色葡萄球菌噬菌体内溶素具有一定的相似度。使用SMART软件对ORF84编码的蛋白Lys qdsa002进行结构域预测分析。结构域预测结果如图1所示,Lys qdsa002为金黄色葡萄球菌噬菌体内溶素典型的双酶裂解系统的模块结构,即在N端具有两个催化区域:CHAP区域和Amidase-2区域,Lys qdsa002的29-160氨基酸区间属于CHAP域,196-369氨基酸区间属于Amidase-2域,以上两个区域与内溶素的裂解细胞壁肽聚糖相关;Lys qdsa002的409-481氨基酸区间属于SH3b家族,该家族与细胞壁肽聚糖结合有关。
实施例2:内溶素在大肠杆菌中的高效表达
1、重组质粒的构建
根据内溶素基因序列(SEQ ID No:2),在不改变氨基酸序列的前提下,对内溶素基因序列的前后两端分别设定限制性内切酶NcoI和XhoⅠ,得到将要进行合成内溶素基因序列。按得到的序列设计70条引物进行PCR扩增,完成后进行电泳,将PCR回收产物克隆入pET-28a载体的多克隆位点NcoI和XhoⅠ之间,得到重组质粒,转化至大肠杆菌BL21(DE3)中。
2、重组蛋白的制备
重组质粒命名为pET-28a-Lys qdsa002转化大肠杆菌BL21(DE3),筛选得到可表达内溶素的工程菌BL21(Lys qdsa002),接种单菌落于2.5mL LB液体培养基(含30μg/mL卡那霉素)中,37℃,150r/min,过夜振荡培养,次日,按1:100比例接种于4L的LB液体培养基(含30μg/mL卡那霉素)中,37℃,220rpm培养约3h,当OD值达到0.6左右时,添加终浓度为0.5mM IPTG,20℃,220rpm,诱导过夜,离心收集细胞菌体。
将收集的细菌菌体用破碎Buffer(50mM Tris,500mM NaCl,pH 8.0)溶解,冰浴中超声破碎菌体后离心,上清液过0.22μm无菌滤膜,获得粗酶液。采用Ni-IDA进行纯化,利用透析进行脱盐处理获得纯度较高的重组蛋白。结果如图2所示,重组蛋白Lys qdsa002得到高效表达。
图2中M为marker,1为未经IPTG诱导的细菌总蛋白,2、3、4、5过柱过程中收集的洗脱液,6为镍柱纯化后的融合蛋白,7为超滤浓缩脱盐后的融合蛋白,8为经过诱导的细菌总蛋白
实施例3:以金黄色葡萄球菌ATCC43300为靶细菌测试内溶素Lys qdsa002抑菌效果
挑取单菌落ATCC43300于50ml TSB液体培养基中,过夜培养,细菌离心用0.9%无菌生理盐水清洗2次。测定活性之前,将细菌沉淀用缓冲液(20mM PB,150mM NaCl,20%甘油,pH 7.4复融),调整OD590至1.5左右。在96孔板中先加入100μL细菌复融液,再加入100μl重组蛋白溶液,每隔一定时间,测定一次OD590值,将缓冲液和溶菌酶作为空白组和阳性对照组,相同条件下培养,用酶标仪测定OD590值,每组3个平行。吸光值下降反映细菌被裂解。
结果如图3和图4所示,重组蛋白Lys qdsa002作用持续时间可长达3天,并且裂解细菌的效果也非常好,可以使浊度下降1.3左右,证明重组蛋白Lys qdsa002具有活性;而作为阳性对照的溶菌酶的菌液浊度仅下降0.4左右,其裂解细菌的效果远远不及重组蛋白Lys qdsa002的裂解效果。
检测Lys qdsa002对多株金黄色葡萄球菌及多种革兰氏阴性菌株的抑菌活性,表1所示,Lys qdsa002能够裂解其余多株金黄色葡萄球菌以及革兰氏阴性菌,具有较宽的裂解谱。(注:OD590下降值为内溶素过夜作用后的下降值,并不是最终的下降值,最终下降值要比表中的下降值大,在此,仅利用下降值表明对作用的细菌具有一定的裂解性)
表1内溶素Lys qdsa002抑菌效果分析
实施例4:以金黄色葡萄球菌ATCC43300为靶细菌测试内溶素Lys qdsa002在不同pH下的作用效果
挑取金黄色葡萄球菌单菌落至TSB培养基中,过夜培养,菌体离心,用生理盐水清洗,得细菌沉淀,将沉淀用pH 1.0-12.0蛋白保存液复融获得不同pH的细菌悬浮液;将重组蛋白用pH 1.0-12.0蛋白保存液稀释。然后将不同pH值条件下的等量的重组蛋白与细菌复融液进行充分混匀,作用一段时间后,测定OD590的数值。
为更直观表示,各个实验组的酶活力=各个组的OD下降值/实验组中OD下降最多值×100%。结果如图5所示,内溶素Lys qdsa002在pH 8-10表现出较好的裂解活性,说明其作用的最适pH为8-10。
以上所述的实施例只是本发明的一种较佳的方案,并非对本发明作任何形式上的限制,在不超出权利要求所记载的技术方案的前提下还有其它的变体及改型。
SEQUENCE LISTING
<110> 中国海洋大学
<120> 来源于耐甲氧西林金黄色葡萄球菌噬菌体的宽谱内溶素及其应用
<130> ZGHY-201701
<160> 2
<170> PatentIn version 3.3
<210> 1
<211> 495
<212> PRT
<213> 人工序列
<400> 1
Met Ala Lys Thr Gln Ala Glu Ile Asn Lys Arg Leu Asp Ala Tyr Ala
1 5 10 15
Lys Gly Thr Val Asp Ser Pro Tyr Arg Ile Lys Lys Ala Thr Ser Tyr
20 25 30
Asp Pro Ser Phe Gly Val Met Glu Ala Gly Ala Ile Asp Ala Asp Gly
35 40 45
Tyr Tyr His Ala Gln Cys Gln Asp Leu Ile Thr Asp Tyr Val Leu Trp
50 55 60
Leu Thr Asp Asn Lys Val Arg Thr Trp Gly Asn Ala Lys Asp Gln Ile
65 70 75 80
Lys Gln Ser Tyr Gly Thr Gly Phe Lys Ile His Glu Asn Lys Pro Ser
85 90 95
Thr Val Pro Lys Lys Gly Trp Ile Ala Val Phe Thr Ser Gly Ser Tyr
100 105 110
Gln Gln Trp Gly His Ile Gly Ile Val Tyr Asp Gly Gly Asn Thr Ser
115 120 125
Thr Phe Thr Ile Leu Glu Gln Asn Trp Asn Gly Tyr Ala Asn Lys Lys
130 135 140
Pro Thr Lys Arg Val Asp Asn Tyr Tyr Gly Leu Thr His Phe Ile Glu
145 150 155 160
Ile Pro Val Lys Ala Gly Thr Thr Val Lys Lys Glu Thr Ala Lys Lys
165 170 175
Ser Ala Ser Lys Thr Pro Ala Pro Lys Lys Lys Ala Thr Leu Lys Val
180 185 190
Ser Lys Asn His Ile Asn Tyr Thr Met Asp Lys Arg Gly Lys Lys Pro
195 200 205
Glu Gly Met Val Ile His Asn Asp Ala Gly Arg Ser Ser Gly Gln Gln
210 215 220
Tyr Glu Asn Ser Leu Ala Asn Ala Gly Tyr Ala Arg Tyr Ala Asn Gly
225 230 235 240
Ile Ala His Tyr Tyr Gly Ser Glu Gly Tyr Val Trp Glu Ala Ile Asp
245 250 255
Ala Lys Asn Gln Ile Ala Trp His Thr Gly Asp Gly Thr Gly Ala Asn
260 265 270
Ser Gly Asn Phe Arg Phe Ala Gly Ile Glu Val Cys Gln Ser Met Ser
275 280 285
Ala Ser Asp Ala Gln Phe Leu Lys Asn Glu Gln Ala Val Phe Gln Phe
290 295 300
Thr Ala Glu Lys Phe Lys Glu Trp Gly Leu Thr Pro Asn Arg Lys Thr
305 310 315 320
Val Arg Leu His Met Glu Phe Val Pro Thr Ala Cys Pro His Arg Ser
325 330 335
Met Val Leu His Thr Gly Phe Asn Pro Val Thr Gln Gly Arg Pro Ser
340 345 350
Gln Ala Ile Met Asn Lys Leu Lys Asp Tyr Phe Ile Lys Gln Ile Lys
355 360 365
Asn Tyr Met Asp Lys Gly Thr Ser Ser Ser Thr Val Val Lys Asp Gly
370 375 380
Lys Thr Ser Ser Ala Ser Thr Pro Ala Thr Arg Pro Val Thr Gly Ser
385 390 395 400
Trp Lys Lys Asn Gln Tyr Gly Thr Trp Tyr Lys Pro Glu Asn Ala Thr
405 410 415
Phe Val Asn Gly Asn Gln Pro Ile Val Thr Arg Ile Gly Ser Pro Phe
420 425 430
Leu Asn Ala Pro Val Gly Gly Asn Leu Pro Ala Gly Ala Thr Ile Val
435 440 445
Tyr Asp Glu Val Cys Ile Gln Ala Gly His Ile Trp Ile Gly Tyr Asn
450 455 460
Ala Tyr Asn Gly Asn Arg Val Tyr Cys Pro Val Arg Thr Cys Gln Gly
465 470 475 480
Val Pro Pro Asn His Ile Pro Gly Val Ala Trp Gly Val Phe Lys
485 490 495
<210> 2
<211> 1485
<212> DNA
<213> 人工序列
<400> 2
atggctaaga ctcaagcaga aataaataaa cgtttagacg cttatgcaaa aggtacagta 60
gacagtcctt atagaattaa aaaagctaca agctatgacc catcgtttgg tgtaatggaa 120
gcaggagcaa ttgacgcaga tggttactat catgcacagt gccaagactt aattactgat 180
tatgtattat ggttaacaga taataaagtt agaacttggg gtaatgctaa agaccaaatc 240
aaacaaagtt atggtactgg atttaaaata catgaaaata aaccttctac agtacctaaa 300
aaaggatgga ttgctgtatt tacatccggt agttatcagc aatggggtca cataggtatt 360
gtatatgatg gaggtaatac ttctacattt actattttag agcaaaactg gaacggttac 420
gctaataaaa aacctacaaa acgtgtagat aattattacg gattaactca ttttattgag 480
atacctgtaa aagcaggaac tactgttaaa aaagaaacag ctaagaaaag tgcaagtaaa 540
acacctgcac ctaaaaagaa agcaacacta aaagtttcta agaaccatat taactataca 600
atggataaac gtggtaagaa acctgaagga atggtaatac acaacgatgc aggtcgttct 660
tcagggcaac aatacgagaa ttcattagct aacgcaggtt atgctagata tgctaatggt 720
attgctcatt actatggctc tgaaggttat gtatgggaag caatagatgc taagaatcaa 780
attgcttggc acacaggaga tggaacagga gcaaactcag gtaactttag atttgcaggt 840
attgaagtct gtcaatcaat gagtgctagt gatgctcaat tccttaaaaa cgaacaagca 900
gtattccaat ttactgcaga gaaatttaaa gaatggggtc ttactcctaa tcgtaaaact 960
gtaagattgc atatggaatt tgttccaaca gcttgtcctc atcgttctat ggttcttcat 1020
acaggattta atccagtaac acaaggaaga ccatctcaag caataatgaa taaactaaaa 1080
gattatttca ttaaacaaat taaaaactac atggataaag gaacttcaag ttctacagta 1140
gttaaagacg gtaaaacaag tagcgcaagt acaccggcaa ctagaccagt aacaggctct 1200
tggaaaaaga accagtacgg aacttggtac aaaccggaaa atgcaacatt tgttaatggt 1260
aaccaaccta tagtaactag aataggttct ccattcttaa atgctccagt aggaggtaac 1320
ttaccggcag gagctacaat tgtatatgac gaagtttgta tccaagcagg tcacatttgg 1380
ataggttaca atgcttacaa tggtaacaga gtatattgcc ctgttagaac ttgtcaagga 1440
gttccaccta atcatatacc tggggttgct tggggagtat tcaaa 1485
- 还没有人留言评论。精彩留言会获得点赞!
- 抗甲氧西林金黄色葡萄球菌的检测的制作方法
- 对甲氧西林抗性金黄色葡萄球菌特异的抗体及将其用于甲氧西林抗性葡萄球菌的检测方 ...的制作方法
- 甲氧滴涕的制备方法
- 抗耐甲氧西林金黄色葡萄球菌耐药基因的反义核酸的制作方法
- 耐甲氧西林金黄色葡萄球菌重组基因工程疫苗候选抗原I<sub>12</sub>C制备中的纯化方法
- 耐甲氧西林金黄色葡萄球菌(MRSA)疫苗重组蛋白抗原HI<sub>2</sub>的纯化方法
- 甲氧虫酰肼的合成方法
- 抗耐甲氧西林金黄色葡萄球菌耐药基因mecA的反义核酸的制作方法
- 耐甲氧西林金黄色葡萄球菌(mrsa)重组多亚单位基因工程疫苗及其制备方法
- 使甲氧西林抗性金黄色葡萄球菌对苯唑西林敏感的组合物和方法
- 用于治疗细菌感染的稠合螺环杂芳族化合物的制造方法与工艺
- 来源于耐甲氧西林金黄色葡萄球菌噬菌体的宽谱内溶素及其应用的制造方法与工艺
- 一种甲氧卡那霉素药物中间体γ‑邻苯二甲酰亚氨基‑α‑羟基丁酸的合成方法与制造工艺
- 一种2‑甲基‑5‑(5‑溴吡啶‑2‑基)四氮唑的制备方法与制造工艺
- 具有免疫原性的模拟金黄色葡萄球菌LTA表位的多肽及其应用的制造方法与工艺
- 使用单剂量奥利万星的治疗方法与制造工艺
- 泰地唑胺合成中间体的晶型的制造方法与工艺
- 含有环己基的不对称二硫醚类化合物及其在制备抗菌药物中的用图
- 一种能够防治犊牛肺炎链球菌病的闽南牛专用饲料添加剂及其制备方法
- 一种能够防治犊牛肺炎链球菌病的安格斯牛专用饲料添加剂及其制备方法