水稻砷酸还原酶基因OsHAC4及其应用的制作方法
本发明属于植物基因工程领域。具体的说,本发明涉及一种图位克隆技术克隆的水稻OsHAC4基因的cDNA序列,该cDNA编码的蛋白属于结构上砷酸还原酶蛋白基因,转基因实验及砷酸盐处理后植物组织砷含量测定实验鉴定该基因调控水稻体内五价砷的还原。
背景技术:
砷在自然界中广泛存在,是地壳中含量最丰富的元素之一,平均浓度接近3mg·kg-1(Cullen and Reimer,1989)。通常情况下,砷通过自然反应和人为作用释放到环境中,例如火山排放、岩石风化、温泉释放、采矿、冶炼以及含砷的农药、除草剂,木材防腐剂和饲料添加剂的使用等(Zhao et al.,2010b)。目前,砷污染已然成为一个全球性的问题,主要涉及食物、空气、水体及土壤污染等方面。无机砷属于人类一级致癌物质,水体和食物为主要的砷污染来源(Smith et al.,2002;Tsuji et al.,2007)。报道显示有70多个国家的地下水受到砷的污染,对全世界约1亿5千万人的身体健康产生严重的危害,这些人中大约1亿1千万人来自于亚洲的十个国家,其中孟加拉国和印度已经大规模地遭受无机砷毒害(Brammer et al.,2009;Bhattacharjee,2007)。砷污染过的地下水用于灌溉,同样会对人体产生巨大的危害。所有植物都能吸收砷,但是相对于其他作物来说,水稻从土壤中同化砷的能力是高效的(Williams et al.,2007)。甚至在没有被砷污染过的土壤中,水稻中砷的含量也普遍很高,这可能与厌氧的环境下砷的生物利用率会提高相关(Xu et al.,2008)。由于特殊的生理结构和转运机制,砷会大量积累在稻粒中,通过食物链的作用被摄入到人体内,成为以水稻为主食的人类健康的潜在威胁(Meharg,2004;Zhu et al.,2008)。同时由于砷长期积累在被灌溉的土壤中,也会对这些地区的可持续发展造成一定影响(Ma et al.,2008)。目前砷的污染问题已经得到广泛的重视,并通过一些积极措施减轻危害。研究表明合理的管理运用灌溉水和选育低砷积累的品种是两种减少砷在水稻中积累的可行性方式(Yi et al.,2015)。
砷的毒性强度与存在形式有关,无机砷的毒性高于有机砷,无机砷中亚砷酸盐的毒性高于砷酸盐,但是当有机砷中的五价单甲基砷酸盐(MMA)和二甲基砷酸盐(DMA)还原成三价时,毒性会显著提高(Styblo et al.,2000)。砷对动植物的危害是巨大的,浓度超过一定的范围甚至会致死。例如长期暴露在砷环境下的植物会导致光合作用受阻,生长发育受到抑制,同时也会引起谷粒的减产(Jain and Gadre,1997)。人体在砷中毒后可能导致皮肤、消化系统、神经系统、呼吸系统、心血管系统等疾病(Kala et al.,2000;Mao et al.,2010)。研究表明砷酸盐会影响磷的代谢途径,导致DNA、RNA等物质的合成受阻(Hughes,2002)。亚砷酸盐会与一些功能基团如巯基(-SH)等发生亲和反应,使得某些酶和蛋白质功能丧失,最终导致细胞的代谢无法正常运行,从而产生细胞死亡(Quaghebeur and Rengel,2003),但是具体的致病机制仍不清楚。
目前发现,植物对砷的吸收形态主要是无机砷,包括砷酸(盐)(AS(V))和亚砷酸(盐)(AS(III)),有机砷在土壤中含量较少,而且吸收速率较慢(Raab et al.,2007)。植物体内砷的主要形态为为无机三价砷,如印度芥菜(Brassica juncea)根与地上部中三价砷占96-100%,拟南芥(Arabidopsis thaliana)叶片中97-100%为三价砷,水稻(Oryza sativa)和番茄(Lycopersicon esculentum)根中三价砷的比例为92-99%(Zhao et al.,2010;Pickering et al.,2000;Dhankher et al.,2002;Liu et al.,2010),说明五价砷吸收后很快被还原为三价砷,大多数植物都具有较强的砷酸还原能力。研究表明,五价砷的还原主要依靠砷酸还原酶,该还原酶还原反应需要还原型谷胱甘肽(GSH)作为电子供体。在拟南芥(A.thaliana)(Dhankher et al.,2006),绒毛草(H.lanatus)(Bleeker et al.,2006),水稻(O.sativa)(Duan et al.,2007)和砷超富集植物蜈蚣草(Pterisvittata)(Ellis et al.,2006)等植物体内都克隆到了砷酸还原酶ACR2基因,这些基因在大肠杆菌arsC缺失突变体中表达或者在酵母acr2p缺失突变体中表达都能回复对五价砷的抗性,说明植物ACR2基因具有还原五价砷的能力。然而对拟南芥ACR2基因研究发现,拟南芥acr2突变体无论砷的还原、外溢,还是砷的积累、抗性都与野生型没有差异,暗示AtACR2基因并不在拟南芥砷代谢过程中起作用。最近,两个研究组分别通过GWAS和近等基因系QTL的方法,克隆了拟南芥的另外一个砷酸还原酶基因AtHAC1(AtATQ1)(Chao et al.,2014;Sanchez-Bermejo et al.,2014)。HAC1在大肠杆菌arsC缺失突变体中表达也可以回复对五价砷的抗性。hac1突变体对砷的敏感性增加,根中五价砷含量大量增加,说明HAC1可能在拟南芥砷还原过程中有重要作用,起着在控制根部五价砷浓度的作用。通过拟南芥砷积累的品种Krefeld(Kr)与砷非积累的品种Collumbia之间的嫁接实验证明,根中的AtHAC1基因的突变,是导致地上部砷积累的主要原因(Chao et al.,2014)。AtHAC1与AtARC2在蛋白序列上具有同源性,但缺失了酵母保守的HCX5R核心砷酸还原位点。在植物中,HAC1的蛋白序列具有高度保守性。研究还发现,虽然Athac1突变体中五价砷的含量增加,但是根中三价砷的含量并没有变化,说明还存在另外的砷酸还原系统,而在此途径中,GSH可能起重要作用。在大肠杆菌和酵母中,也存在这种功能冗余现象(Oden et al.,1994;Liu et al.,2012)。一种可能是依赖于GSH的缓慢的非酶促反应而导致的还原;另一种可能是以GSH作为还原剂,酶促剪切被砷酸化的底物而导致的还原(Gregus et al.,2009)。另外,拟南芥hac1arc2双突变体材料并没有上位或加性效应,暗示HAC1是参与拟南芥五价砷还原的主要还原酶(Ellis et al.,2006)。HAC1-GFP定位于根毛、表皮和中柱鞘细胞,表明其参与两个过程:1、在根表皮细胞内,砷酸盐被快速还原以外排;2、在中柱鞘细胞内,砷酸盐被快速还原并形成PCn-As络合物而被输送入液泡或直接被转运入木质部(Chao et al.,2014)。研究还发现,HAC1可以与三价砷外溢载体结合,从而促进三价砷的外溢。
对于水稻砷酸还原酶的研究有助于有助于加深对水稻砷酸盐吸收外排和耐受的分子机制的认识,可以通过基因改良减少水稻中砷的积累,对提高粮食安全具有重要意义。
上文中涉及的参考文献如下:
1、Bhattacharjee,Y.A Sluggish Response to Humanity's Biggest Mass Poisoning[J].Science,2007,315(5819):1659-1661(Bhattacharjee,Y.对人类最大规模的缓慢中毒反应[J].科学,2007,315(5819):1659-1661)。
2、Bleeker PM,Hakvoort HWJ,Bliek M,Souer E,Schat H.Enhanced arsenate reduction by a CDC25-like tyrosine phosphatase explains increased phytochelatin accumulation in arsenate-tolerant Holcus lanatus.Plant J,2006.45:917-929(Bleeker PM,Hakvoort HWJ,Bliek M,Souer E,Schat H.通过CDC25类酪氨酸磷酸酶增强的砷酸盐还原是砷酸盐耐受毛孔菌中的植物螯合素积累增加的原因。植物杂志,2006.45:917-929)。
3、Brammer,H.&Ravenscroft,P.Arsenic in groundwater:a threat to sustainable agriculture in South and South-east Asia.[J].Environment International,2009,35(3):647-654(Brammer,H.&Ravenscroft,P.地下水中的砷:对南亚和东南亚可持续农业的威胁。国际环境,2009,35(3):647-654)。
4、Chao DY,Chen Y,Chen J,Shi S,Chen Z,Wang C,Danku JM,Zhao FJ,Salt DE.Genome-wide association mapping identifies a new arsenate reductase enzyme critical for limiting arsenic accumulation in plants.PLoS Biol,2014.12:e1002009(Chao DY,Chen Y,Chen J,Shi S,Chen Z,Wang C,Danku JM,Zhao FJ,Salt DE.全基因组关联作图确定了一个限制植物中的砷积累新的关键砷酸还原酶酶。PLoS生物,2014.12:e1002009)。
5、Cullen,W.R.&Reimer,K.J.Arsenic speciation in the environment[J].Chemical Reviews,1989,89(1):713-764(Cullen,W.R.&Reimer,K.J.环境中的砷种类。化学评论,1989,89(1):713-764)。
6、Dhankher OP,Li YJ,Rosen BP,Rosen BP,Shi J,Salt DE,Senecoff JF,Sashti NA,Meagher RB.Engineering tolerance and hyperaccumulation of arsenic in plants by combining arsenate reductase and gamma-glutamyl cysteine synthetase expression.Nat Bioltechnol,2002.20:1140-1145(Dhankher OP,Li YJ,Rosen BP,Rosen BP,Shi J,Salt DE,Senecoff JF,Sashti NA,Meagher RB.通过基因工程手段组合砷酸还原酶和γ-谷氨酰半胱氨酸合成酶提高植物的砷耐受性和超积累。自然生物技术,2002.20:1140-1145)。
7、Dhankher OP,Rosen BP,McKinney EC,Meagher RB.Hyperaccumulation of arsenic in the shoots of Arabidopsis silenced for arsenate reductase(ACR2).Proc Nat Acad Sci USA,2006.103:5413-5418(Dhankher OP,Rosen BP,McKinney EC,Meagher RB.通过沉默ACR2基因导致拟南芥地上部砷超级累.美国科学院院报,2006.103:5413-5418)。
8、Duan GL,Zhou Y,Tong YP,Mukhopadhyay R,Rosen BP,Zhu YG.A CDC25homologue from rice functions as an arsenate reductase.New Phytol,2007.174:311-321(Duan GL,Zhou Y,Tong YP,Mukhopadhyay R,Rosen BP,Zhu YG.一个水稻CDC25同源基因具有砷酸还原酶活性.新植物学家,2007.174:311-321)。
9、Ellis DR,Gumaelius L,Indriolo E,Pickering IJ,Banks JA,Salt DE.A novel arsenate reductase from the arsenic hyperaccumulating fern Pterisvittata.Plant Physiol,2006.141:1544-1554(Ellis DR,Gumaelius L,Indriolo E,Pickering IJ,Banks JA,Salt DE.蕨类植物Pterisvittata中发现的新的砷酸还原酶基因.植物生理,2006.141:1544-1554)。
10、Hughes,M.F.Arsenic toxicity and potential mechanisms of action[J].Toxicology Letters,2002,133(1):1–16(Hughes,M.F.砷毒性和作用机制[J].毒理学通讯,2002,133(1):1–16)。
11、Jain,M.&Gadre,R.Effect of As on Chlorophyll and Protein Contents and enzymic Activities in Greening Maize Tissues[J].Water,1997,volume 93(1-4):109-115(7)(Jain,M.&Gadre,R.As对玉米绿色组织中叶绿素、蛋白质含量和的酶活性的影响[J].水科学,1997,volume 93(1-4):109-115(7))。
12、Kala,S.V.,Neely,M.W.,Kala,G.,et al.The MRP2/cMOAT transporter and arsenic-glutathione complex formation are required for biliary excretion of arsenic[J].Journal of Biological Chemistry,2000,275(43):33404-33408(Kala,S.V.,Neely,M.W.,Kala,G.,et al.MRP2/cMOAT转运蛋白和砷-谷胱甘肽复合物形成是胆汁排泄砷所必需的[J].生物化学杂志,2000,275(43):33404-33408)。
13、Liu W,Schat H,Bliek M,Chen Y,McGrath SP,George G,Salt DE,Zhao FJ.Knocking out ACR2 does not affect arsenic redox status in Arabidopsis thaliana:implications for as detoxification and accumulation in plants.PLoS One,2012.7:e42408(Liu W,Schat H,Bliek M,Chen Y,McGrath SP,George G,Salt DE,Zhao FJ.敲除ACR2不影响拟南芥中的砷氧化还原状态:作为解毒和植物中的积累的影响。PLoS One,2012.7:e42408)。
14、Liu WJ,Wood BA,Raab A,McGrath SP,Zhao FJ,Feldmann J.Complexation of arsenite with phytochelatins reduces arsenite efflux and translocation from roots to shoots in arabidopsis.Plant Physiol,2010.152:2211-2221(Liu WJ,Wood BA,Raab A,McGrath SP,Zhao FJ,Feldmann J.亚砷酸盐与植物螯合素的络合减少了拟南芥中砷的外流和从根到地上部的转移。植物生理,2010.152:2211-2221)。
15、Ma,J.F.,Yamaji,N.,Mitani,N.,et al.Transporters of arsenite in rice and their role in arsenic accumulation in rice grain[J].Proceedings of the National Academy of Sciences of the United States of America,2008,105(29):9931-9935(Ma,J.F.,Yamaji,N.,Mitani,N.,et al.水稻中亚砷酸盐的转运蛋白及其在稻谷中砷积累的作用[J].美国科学院院报,2008,105(29):9931-9935)。
16、Mao,G.,Guo,X.,Kang,R.,et al.Prevalence of disability in an arsenic exposure area in Inner Mongolia,China[J].Chemosphere,2010,80(9):978–981(Mao,G.,Guo,X.,Kang,R.,et al.中国内蒙古的砷暴露区残疾的流行[J].化学圈,2010,80(9):978–981)。
17、Meharg,A.A.Arsenic in rice:understanding a new disaster for South-East Asia[J].Trends in Plant Science,2004,9(9):415-417(Meharg,A.A.水稻砷:理解东南亚的新灾难[J].植物科学进展,2004,9(9):415-417)。
18、Oden KL,Gladysheva TB,Rosen BP.Arsenate reduction mediated by the plasmid-encoded ArsC protein is coupled to glutathione.Molecular Microbiology,1994.12:301-306(Oden KL,Gladysheva TB,Rosen BP.由质粒编码的ArsC蛋白介导的砷酸盐还原与谷胱甘肽偶联.微生物分子生物学,1994.12:301-306)。
19、Pickering IJ,Prince RC,George MJ.Reduction and coordination of arsenic in Indian mustard.Plant Physiol,2000.122:1171-1177(Pickering IJ,Prince RC,George MJ.减少和协调印度芥菜中的砷。植物生理,2000.122:1171-1177)。
20、Quaghebeur,M.&Rengel,Z.The distribution of arsenate and arsenite in shoots and roots of Holcus lanatus is influenced by arsenic tolerance and arsenate and phosphate supply[J].Plant Physiology,2003,132(3):1600-9(Quaghebeur,M.&Rengel,Z.绒毛的芽和根中的砷酸盐和亚砷酸盐的分布受砷耐受性和砷酸盐和磷酸盐供应的影响。植物生理,2003,132(3):1600-9)。
21、Raab A,Williams PN,Meharg A,Feldmann J.Uptake and translocation of inorganic and methylated arsenic species by plants.Environ Chem,2007.4:197-203(Raab A,Williams PN,Meharg A,Feldmann J.植物吸收和转移无机和甲基化砷种类。环境化学,2007.4:197-203)。
22、Sanchez-Bermejo E,Castrillo G,del Llano B,Navarro C,Zarco-Fernandez S,Martinez-Herrera DJ,Leo-del Puerto Y,Munoz R,Camara C,Paz-Ares J,Alonso-Blanco C,Leyva A.Natural variation in arsenate tolerance identifies an arsenate reductase in Arabidopsis thaliana.Nat Commun,2014.5:4617(Sanchez-Bermejo E,Castrillo G,del Llano B,Navarro C,Zarco-Fernandez S,Martinez-Herrera DJ,Leo-del Puerto Y,Munoz R,Camara C,Paz-Ares J,Alonso-Blanco C,Leyva A.通过砷酸盐耐受性的自然变异发现拟南芥中的砷酸盐还原酶.自然通讯,014.5:4617)。
23、Smith,A.H.&Steinmaus,C.M.Arsenic Epidemiology and Drinking Water Standards[J].Science,2002,296(5576):2145-2146(Smith,A.H.&Steinmaus,C.M.砷流行病学和饮用水标准[J].科学,2002,296(5576):2145-2146)。
24、Styblo,M.,Razo,L.M.D.,Vega,L.,et al.Comparative toxicity of trivalent and pentavalent inorganic and methylated arsenicals in rat and human cells[J].Archives of Toxicology,2000,74(6):289-299(Styblo,M.,Razo,L.M.D.,Vega,L.,et al.三价和五价无机和甲基化砷在大鼠和人类细胞中的比较毒性[J].毒理学档案,2000,74(6):289-299)。
25、Tsuji,J.S.,Yost,L.J.,Barraj,L.M.,et al.Use of background inorganic arsenic exposures to provide perspective on risk assessment results[J].Regulatory Toxicology&Pharmacology Rtp,2007,48(1):59–68(Tsuji,J.S.,Yost,L.J.,Barraj,L.M.,et al.使用背景无机砷暴露以提供风险评估结果的观点[J].毒理学和药理学控制,2007,48(1):59–68)。
26、Williams,P.N.,Villada,A.,Deacon,C.,et al.Greatly enhanced arsenic shoot assimilation in rice leads to elevated grain levels compared to wheat and barley[J].Environmental Science&Technology,2007,41(19):6854-6859(Williams,P.N.,Villada,A.,Deacon,C.,et al.与小麦和大麦相比,水稻地上部砷积累要大大提高谷粒砷水平[J].环境科学与技术,2007,41(19):6854-6859)。
27、Xu,X.Y.,Mcgrath,S.P.,Meharg,A.A.,et al.Growing rice aerobically markedly decreases arsenic accumulation[J].Environmental Science&Technology,2008,42(15):5574-5579(Xu,X.Y.,Mcgrath,S.P.,Meharg,A.A.,et al.旱种水稻显着减少砷积累[J].环境科学与技术,2008,42(15):5574-5579)。
28、Yi,C.,Moore,K.L.,Miller,A.J.,et al.The role of nodes in arsenic storage and distribution in rice[J].Journal of Experimental Botany,2015(Yi,C.,Moore,K.L.,Miller,A.J.,et al.节在水稻砷储存和分布中的作用[J].实验生物学报,2015)。
29、Zhao,F.J.,Mcgrath,S.P.,Meharg,A.A.,et al.Arsenic as a food chain contaminant:mechanisms of plant uptake and metabolism and mitigation strategies[J].Annual Review of Plant Biology,2010b,61(4),535-59.(Zhao,F.J.,Mcgrath,S.P.,Meharg,A.A.,et al.砷作为食物链污染物:植物吸收和代谢机制以及缓解策略[J].植物生物学年报,2010b,61(4),535-59)。
30、Zhu,Y.G.,Williams,P.N.&Meharg,A.A.Exposure to inorganic arsenic from rice:a global health issue[J].Environmental Pollution,2008,154(2):169–171(Zhu,Y.G.,Williams,P.N.&Meharg,A.A.暴露于来自稻米的无机砷:全球健康问题[J].环境污染,2008,154(2):169–171)。
技术实现要素:
本发明要解决的技术问题是提供一种水稻砷酸还原酶基因OsHAC4及其在食品安全方面的应用。
为了解决上述技术问题,本发明提供一种水稻具有砷酸还原活性的砷酸还原酶基因OsHAC4编码的蛋白质,具有SEQ ID NO:2所示的氨基酸序列;该蛋白是一个砷酸还原酶蛋白基因。
作为本发明的蛋白质的改进:所述氨基酸序列还包括在SEQ ID NO:2所示的氨基酸序列中添加、取代、插入和缺失一个或多个氨基酸生成的衍生物。
本发明还同时提供了一种编码上述蛋白或其功能类似物的基因,具有SEQ ID NO:1所示的核苷酸序列。
作为本发明的基因的改进:核苷酸序列还包括在SEQ ID NO:1所示的核苷酸序列中添加、取代、插入和缺失一个或多个核苷酸生成的突变体、等位基因和衍生物。
本发明还同时提供了上述基因的用途:用于降低水稻(特别是在砷污染地区的水稻)体内砷含量。
作为本发明的基因用途的改进:利用含有所述基因的植物表达载体来转化水稻,从而降低水稻砷浓度。
本发明的发明过程如下:从本实验室发展的EMS诱变的phf1突变体库为研究群体,在含20μM Na3AsO4的水稻全培养液中培养5天,筛选到一株对砷的敏感程度回复到野生型的突变体,根据后续研究将该突变体命名为phf1/hac4。与phf1相比,phf1/hac4突变体根受砷的毒害表型随着砷浓度的增加而逐渐加重,在20μM Na3AsO4的条件下回复到野生型表型,但地上部变化不明显。(图1)。
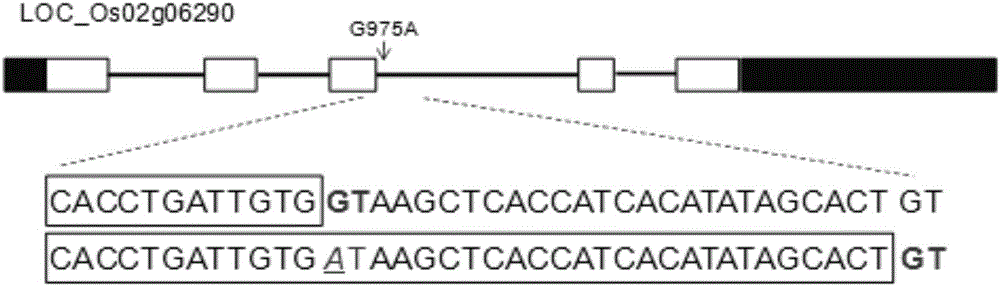
为了克隆OsHAC4基因,本发明采用二代基因组重测序方法。取phf1/hac4与phf1杂交群体F2代中25株砷敏感表型(与phf1/Oshac4相似的表型)单株进行基因组重测序,测序结果发现,在第2号染色体的基因组第3142971个碱基位置发生突变(由G变为A),并且此位点两侧的SNP呈现连锁现象。通过进一步根据水稻基因注释网站Rice Genome Annotation Project(http://rice.plantbiology.msu.edu/index.shtml)研究发现,OsHAC4基因组序列为2669bp,由五个外显子和四个内含子组成,CDS序列为390bp,蛋白序列由129个氨基酸组成。突变体Oshac4从基因编码序列开始第975位碱基由G变为A(图2),由于突变位点在内含子上,会导致基因在转录过程中无法正常剪切,从而发生移码突变。
为了进一步确定OsHAC4的突变引起了突变体phf1/hac4表型的变化,用转基因回复实验进行验证。构建含有OsHAC4基因启动子和基因组序列的载体,转入到phf1/hac4突变体的愈伤组织中,经过继代培养、共培养、选择培养、诱导分化和移栽鉴定后,利用转基因苗T2代纯合种子,在20μM Na3AsO4的半营养液条件下进行培养,5天后观察表型(图3)。转基因苗phf1/hac4-OsHAC4-1和phf1/hac4-OsHAC4-2与phf1相比,地上部和地下部均没有明显的区别,说明OsHAC4基因突变导致了phf1/hac4突变体对砷的超敏感。
为了进一步说明OsHAC4的功能,由35S强启动子启动的完整的OsHAC4ORF(SEQ ID NO:1)克隆到双元植物转基因载体pCAMBIA1300中(图4)。将构建好的回复载体通过农杆菌介导转化日本晴的愈伤,进行诱导。T1的转基因苗21天时在10μM Na3AsO4的处理48小时后,测定地上部和根部的总砷含量,结果发现,OsHAC4超表达株系,总砷含量较野生型明显减少(图5)。
由这些结果表明,本发明克隆的水稻OsHAC4基因具有一定的应用价值。我们可以通过利用该基因对作物品种进行转基因改造,如通过外源导入超表达OsHAC4基因来降低在砷污染地区水稻体内砷含量,提高食品安全性。
综上所述,本发明提供一种从水稻五价砷敏感突变体hac4中克隆的新基因OsHAC4,如SEQ ID NO:1所示的cDNA序列,也包括与SEQ ID NO:1所示的cDNA序列至少有80%同源性的基因序列。本发明中的SEQ ID NO:2所示的蛋白质与AtHAC1类似,其中进行一个或几个替换,插入或缺失所获得的功能类似物。另外,也包括在SEQ ID NO:1,SEQ ID NO:2中添加、取代、插入或缺失一个或多个核苷酸而生成的突变体、等位基因或衍生物,具有相同功能的序列也能达到本发明的目的。本发明的另一个目的是提供一种用OsHAC4基因进行高效的植物转化的方法,具体地说,本发明提供了具有SEQ ID NO:1所示的序列的基因片段的载体。
本发明还提供了一种利用植物表达载体转化植物降低水稻砷浓度的方法。本发明的转基因植物株系,为包含上述核酸的转基因植物株系。
附图说明
下面结合附图对理解本发明作进一步的详细描述,但并非对发明作限定。
图1是水稻磷吸收突变体hac4的表型图;
A为不同Na3AsO4浓度处理5天的野生型(黑粳2,HJ2)、phf1和phf1/hac4的全株照;
B为A图中野生型(HJ2)、phf1和phf1/hac4的根长;
C为A图中野生型(HJ2)、phf1和phf1/hac4的株高。Bar=2cm。统计样本为5株。
图2是OsHAC4的基因结构和突变位点。
图3是功能互补实验图;
A-B为水稻半营养液(A)和添加20μM Na3AsO4的半营养液(B)条件下培养5天的phf1、突变体phf1/hac4和转基因回复材料phf1/hac4-HAC4-1和phf1/hac4-HAC4-2的全株照;
C-D为A-B图中phf1、phf1/hac4、转基因回复材料phf1/hac4-HAC4-1和phf1/hac4-HAC4-2的根长(C)和株高(D)。Bar=2cm。统计样本为5株。
图4是超表达转化载体pCAMBIA1300改的结构示意图。
图5是超表达OsHAC4株系RT-PCR鉴定(A)和总砷含量(B)的对比图。
具体实施方式
下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明范围。
在本发明中,所需的各种培养液,例如水稻全营养液、半营养液等,均属于公知配方,例如可按照文献《Zhongchang Wu,Hongyan Ren,Steve P.McGrath,Ping Wu,and Fang-Jie Zhao.Investigating the Contribution of the Phosphate Transport Pathway to Arsenic Accumulation in Rice.Plant Physiology 2011,157:498-508》(Zhongchang Wu,Hongyan Ren,Steve P.McGrath,Ping Wu,and Fang-Jie Zhao.水稻磷酸盐转运途径对砷积累贡献的研究。植物生理,2011,157:498-508)所述的配方。
phf1即为phf1-1,在已经公开发表的《Jieyu Chen,Yu Liu,Jun Ni,Yifeng Wang,Youhuang Bai,Jing Shi,Jian Gan,Zhongchang Wu,and Ping Wu.OsPHF1Regulates the Plasma Membrane Localization of Low-and High-Affinity Inorganic Phosphate Transporters and Determines Inorganic Phosphate Uptake and Translocation in Rice.Plant Physiology 2011,157:269-278》(Jieyu Chen,Yu Liu,Jun Ni,Yifeng Wang,Youhuang Bai,Jing Shi,Jian Gan,Zhongchang Wu,and Ping Wu.OsPHF1通过调控水稻低亲和和高亲和磷酸盐转运体的质膜定位来调控磷的吸收和转运。植物生理,2011,157:269-278)中有相应告知。
实施例1、突变体的筛选和表型
以本实验室所有的phf1(粳稻黑粳2(HJ2)背景)为原始材料,利用经EMS(Ethyl methylsulfonate)诱变后的群体作为对象进行突变体的筛选;所述EMS(Ethyl methylsulfonate)诱变方法具体为:选取饱满、无病虫害、大小均匀、发芽率95-100%的phf1种子,置于网袋中(每袋250克),用蒸馏水浸24小时。凉干后,用EMS溶液处理10小时后,自来水冲洗4小时。处理好的种子繁殖一代,单株收种用于突变体筛选。
种子烘干后,用超纯水冲洗干净,然后用1%HNO3(v/v)破休眠处理16小时,37℃温箱中暗处理催芽约两天至露白,每隔12小时更换一次超纯水。将露白的种子播于含20M Na3AsO4水稻培养液的尼龙网纱上,在水稻生长室中培养5天后根据根的表型变化进行突变体的筛选,从中筛选到一株根长显著短于phf1的株系,即,phf1/hac4突变体。与phf1相比,phf1/hac4突变体根受砷的毒害表型随着砷浓度的增加而逐渐加重,在20μM Na3AsO4的条件下回复到野生型表型,但地上部变化不明显(图1)。
将水稻培养液中Na3AsO4的浓度由“20μM As5+”改成“0μM As5+”、“10μM As5+”,其余方法同上,所得结果如图1所述;
根据图1,我们得知:phf1/hac4突变体根受砷的毒害表型随着砷浓度的增加而逐渐加重,说明Oshac4基因突变后,水稻对砷的敏感性增加。
实施例2、基因克隆
将phf1/hac4突变体做母本,phf1做父本进行杂交,得到F2代的种子培养在含有20μM Na3AsO4的水稻全营养液中,挑出25株砷敏感表型(与phf1/hac4相似的表型,即满足20μM Na3AsO4的条件下,根长与野生型HJ2相同)单株进行提取基因组DNA。取大约0.1g水稻幼嫩叶片,经液氮冷冻,在1.5ml的离心管中将叶片磨成粉末,提取总DNA,获得的DNA溶解于200μl无菌水中。25个DNA等量混合,送公司进行基因重测序,并进行SNP分析,测序结果发现,在第2号染色体的基因组第3142971个碱基位置发生突变(由G变为A),并且此位点两侧的SNP呈现连锁现象(表1)。
表1基因组重测序分析
水稻具有砷酸还原活性的砷酸还原酶基因OsHAC4,具有SEQ ID NO:1所示的核苷酸序列;其编码的蛋白质,具有SEQ ID NO:2所示的氨基酸序列。该蛋白是一个砷酸还原酶蛋白基因。
实施例3、突变体回复验证
为了进一步确定OsHAC4的突变引起了突变体phf1/Oshac4表型的变化,用转基因回复实验进行验证。根据OsHAC4基因启动子和基因组序列,设计引物如下:
上游序列:CCATGATTACGAATTCATTTTCTCTCATATAATAGCAT
下游序列:CGACTCTAGAGGATCCTTACTGTTGATGAGCTGCAGGT
以粳稻HJ2基因组DNA为模板,扩增出OsHAC4启动子和基因组序列,用INFUSION(clontech)方法连入pCAMBIA1300载体,连接产物热激转化大肠杆菌DH5α感受态细胞。酶切检测正确后备用。pCAMBIA1300为本实验室改造载体,该载体是以pCAMBIA-1300载体为基本骨架,在多克隆位点插入35S启动子和Nos终止子而得到的增强表达载体(图4)。将得到的正确克隆的质粒通过农杆菌株系EHA105介导的水稻遗传转化体系导入到突变体phf1/hac4中,经过预培养、侵染、共培养、筛选具有潮霉素抗性的愈伤、分化、生根、练苗移栽,得到2个转基因植株phf1/hac4-HAC4-1和phf1/hac4-HAC4-2。农杆菌(EHA105)介导的水稻遗传转化体系主要应用Hiei等人(1994)报道的方法基础上进行优化。
利用转基因苗T2代纯合种子,在20μM Na3AsO4的半营养液条件下进行培养,5天后观察表型(图3)。转基因苗phf1/hac4-HAC4-1和phf1/hac4-HAC4-2与phf1相比,地上部和地下部均没有明显的区别,说明OsHAC4基因突变导致了phf1/hac4突变体对砷的超敏感。
实施例4、OsHAC4基因超表达株系砷浓度分析。
根据OsHAC4基因ORF序列,设计引物如下:
上游引物:CAGggtaccATGGCGTCTGCGGCGATGAGC
下游引物:CAGtctagaTTACTGTTGATGAGCTGCAGG
以粳稻HJ2的根部cDNA为模板,扩增出OsHAC4基因的编码区(即,如SEQ ID NO:1所述),用Kpn I和Xba I双酶切,连入用Kpn I和Xba I双酶切的pCAMBIA1300改载体中,连接产物热激转化大肠杆菌DH5α感受态细胞。酶切检测正确后备用。将得到的正确克隆的质粒通过农杆菌株系EHA105介导的水稻遗传转化体系导入到粳稻HJ2中,经过预培养、侵染、共培养、筛选具有潮霉素抗性的愈伤、分化、生根、练苗移栽,得到2个转基因植株OsHAC4-OVER-1、OsHAC4-OVER-2。
为了检测叶片有效磷得到回复的转基因阳性植株是否超表达了OsHAC4基因,采用Trizol法分别提取HJ2和2个转基因回复株系叶片的总RNA(我们构建的回复载体为CaMV 35S组成型表达启动子,如果为转基因阳性植株,那么植株体会整体超表达OsHAC4基因),逆转录成cDNA,并以定量PCR反应测定转基因植株表达丰度。
试剂及qRT-PCR体系和程序:
Master为PROMEGA公司的SYBR Premix Ex TaqTM。
反应体系(5μl)
定量PCR反应仪器为Roche LightCycle480定量PCR仪。
PCR程序
用参照基因(如Actin)作为标准进行相对定量,优点是能准确量化初始材料的载量,缺点是这个方法要求得到一个或多个在所有测试样本中恒定表达的已知参照基因。用相对定量比较多个样本,样本之一常被选为参照。在其它所有样本中目的基因的表达都相对于参照上调或下调,通常用未处理的或基准样本作为参照样本,一般选择野生型样本。
进行相对基因表达分析普遍采取2-ΔΔCT法,条件是目的基因和参照基因扩增效率都接近100%且相互效率偏差在5%以内。定量结果如图5,OSHAC4超表达株系OsHAC4在转录水平上相对表达量明显高于野生型HJ2。
HJ2、OsHAC4-OVER-1、OsHAC4-OVER-2在水稻全营养液中培养20天后,第21天时选取长势一致的水稻苗进行砷处理,处理时不加磷。在新换的水稻培养液中加入10μM的Na3AsO4,处理时间为48小时,每个处理4个重复。处理完毕后及时取出样品,用自来水洗净根表面的砷,然后置于事先配好的洗脱液中,洗脱约10分钟后取出,用超纯水再冲洗一遍,最后用吸水纸吸干表面水分,装入信封放入65℃烘箱中,完全烘干后(约72小时)消解。消解后通过ICP-MS测定砷含量。结果发现OsHAC4超表达株系,总砷含量较野生型明显减少。说明可以通过基因工程手段,降低水稻体内砷含量(图5)。
最后,还需要注意的是,以上列举的仅是本发明的若干个具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。
<110> 浙江大学
<120> 水稻砷酸还原酶基因OsHAC4及其应用
<160> 2
<210> 1
<211> 402
<212> DNA
<213> 水稻(Oryza sativa)
<400> 1
ATGGCGTCTG CGGCGATGAG CAGCAACAAG AAGGAGCTGG AAGCCCTCCC AATCGTCGAC 60
GCCGGCGAGG TTCGTGAGCT CATGAGCTCC GGCCACCATT ACCTCGATGT CAGGCTTGGG 120
AAGGATTTTG ACAAGGCGCA TGCTGACGGT GCTCGCAACA TTTCCTACTA CCTCTCAGTG 180
ACTCCCAGTG GGAAGGAGAA GAACCCACAC TTTGTAGATG AAGTGGCTTC ACTCTTCGGC 240
AAGGATGAGC ACCTGATTGT GGCTTGCAAC ACAGGTGTGC GATCCAGGCT CGCGACTAAG 300
GACCTTTTGG ACGCGGGATT CAAGAATGTG AGGAACCTGA AAGGTGGTTA CCAATCATTT 360
CTCCGAAGTG AAAGCCAGCA ACCTGCAGCT CATCAACAGT AA 402
<210> 2
<211> 133
<212> PRT
<213> 水稻(Oryza sativa)
<400> 2
Met Ala Ser Ala Ala Met Ser Ser Asn Lys Lys Glu Leu Glu Ala Leu Pro Ile Val Asp
Ala Gly Glu Val Arg Glu Leu Met Ser Ser Gly His His Tyr Leu Asp Val Arg Leu Gly
Lys Asp Phe Asp Lys Ala His Ala Asp Gly Ala Arg Asn Ile Ser Tyr Tyr Leu Ser Val
Thr Pro Ser Gly Lys Glu Lys Asn Pro His Phe Val Asp Glu Val Ala Ser Leu Phe Gly
Lys Asp Glu His Leu Ile Val Ala Cys Asn Thr Gly Val Arg Ser Arg Leu Ala Thr Lys
Asp Leu Leu Asp Ala Gly Phe Lys Asn Val Arg Asn Leu Lys Gly Gly Tyr Gln Ser Phe
Leu Arg Ser Glu Ser Gln Gln Pro Ala Ala His Gln Gln *
- 还没有人留言评论。精彩留言会获得点赞!