双特异性抗体的改进的组装的制作方法
本申请是申请日为2012年10月11日、发明名称为“双特异性抗体的改进的组装”的中国专利申请201280060881.5的分案申请。
本发明涉及2011年10月11日提交的美国临时申请系列号61/545863、2011年10月12日提交的美国临时申请系列号61/546503、2011年11月16日提交的美国临时申请系列号61/560704和2012年7月27日提交的美国临时申请系列号61/676837,并要求其优先权。上文引用的临时申请中每一个的公开内容在此以其整体引入作为参考。
本发明公开涉及组装异源多聚体蛋白质诸如双特异性抗体的组合物及改进的方法。
背景技术:
igg类型的单克隆抗体包含两条相同的抗原结合臂和恒定结构域(fc)。在其结合臂中具有不同特异性的抗体通常不存在于自然界中,因此,必须借助化学工程(例如化学交联等)、重组dna和/或细胞融合技术来制备。
双特异性抗体可以同时结合两种不同的抗原。此特性使得能够发展对常规单克隆抗体而言不可能的治疗策略。已发展的一系列富于想象的双特异性抗体形式反映了对这些分子的浓厚兴趣。见bergj,lotschere,steimerks等,“bispecificantibodiesthatmediatekillingofcellsinfectedwithhumanimmunodeficiencyvirusofanystrain,”procnatlacadsciusa(1991)88(11):4723-4727及fischern和legero.,“biospecificantibodies:moleculesthatenablenoveltherapeuticstrategies,”pathobiology(2007)74:3-14。
另一类多特异性分子是重组融合蛋白。由免疫调节蛋白的胞外域和免疫球蛋白(ig)的恒定(fc)结构域组成的重组融合蛋白是一类正在不断壮大的人治疗剂。免疫黏附素将具有希望的特异性的蛋白质序列的结合区与抗体的效应子结构域组合。免疫黏附素具有对其作为治疗剂的潜能而言有意义的两种重要特性:靶标特异性和药代动力学稳定性(与抗体的体内半衰期相当的体内半衰期)。免疫黏附素可以用作拮抗剂来抑制或阻断有害的相互作用,或用作激动剂来模拟或增强生理反应。见chamowsm,zhangdz,tanxy等,“ahumanized,bispecificimmunoadhesin-antibodythatretargetscd3+effectorstokillhiv-1-infectedcells,”jhematother1995;4(5):439-446。
其他多特异性分子已在其他文献中讨论过。实例包括但不限于:fisher等,pathobiology(2007)74:3-14(多种双特异性形式的综述);feige等的2003年12月9日授权的美国专利号6,660,843(肽体(peptibody));2002年1月10日公开的美国专利公开号2002-004587(多特异性抗体);wu等的2009年11月3日授权的美国专利号7612181(双可变结构域形式);美国专利号6,534,628、nordk等,proteng(1995)8:601-608、nordk等,natbiotech(1997)15:772-777及
对于抗体,通常且尤其是对于上述多特异性分子,临床级物质的制备仍富有挑战。如上文所指出,存在许多产生具有混合结合臂(即相互不同的结合臂)的分子的途径。但这些方法中的每一种都有它的缺点。
化学交联为劳动密集型,因为可能还需从同二聚体及其他不想要的副产物中纯化相关物质。此外,化学修饰步骤可以改变蛋白质的完整性,从而导致稳定性差。因此,此方法常效率低,且可能导致抗体活性丧失。
细胞融合技术(例如杂交瘤)表达两条重链和两条轻链,其随机组装而导致产生10种抗体组合。期望的异源多聚体抗体只是这样产生的抗体中的一小部分。对期望的异源多聚体蛋白质的纯化会显著降低产率,并增加生产成本。
重组dna技术已用于产生不包含fc结构域的多种异源多聚体形式,例如单链fv、双抗体(diabody)等。这类抗体分子的主要缺点是缺乏fc结构域,因此缺乏抗体触发效应子功能(例如补体激活、fc受体结合等)的能力。因此,希望得到包含功能性fc结构域的双特异性抗体。
重组dna技术还已用于产生“杵入臼”(‘knobintohole’)双特异性抗体。见美国专利申请20030078385(arathoon等-genentech)。此策略的一个限制是,两种亲本抗体的轻链必须相同,以防止在同一细胞中表达时错配及形成不想要和/或无活性的分子。
此外,在退火和纯化过程中的限制事件之一是氧化还原效率。氧化的异二聚体在此步骤后通常仅占蛋白质的70-80%(bioanalyzer和ms-tof)。其余20-30%的抗体是同二聚体,且缺乏共价连接(sec-lls)。这可以去除,但显著影响总体产率。因此,仍然存在改进抗体生产(尤其是异二聚体)的总体产率的需要。本发明所描述的是可以改进双特异性抗体、异二聚体等的总体产率的方法。本发明的这些和其他方面及优势将从本文提供的发明详述显而易见。
发明概述
用目前的技术生产由两种或更多种含铰链多肽组装的异源多聚体蛋白质(例如从两种或更多种半抗体组装的多特异性抗体)具有缺点,包括产生混合产物、降低的产率和减少/消除的效应子功能等。此外,在制备每种含铰链多肽的过程中及在组装或退火异源多聚体的过程中常发生聚集和沉淀。聚集和沉淀可以大幅降低希望的异源多聚体的产率。因此,希望更有效和以更高的水平产生异源多聚体蛋白质。
本文公开,通过使用或调节一个或多个方面来经济地生产异源多聚体蛋白质(例如多特异性抗体)的有效生产方法/工艺,所述方面包括但不限于如下:稳定剂、加溶剂、还原条件、所选择的ph和所选择的温度等。本文描述的本发明方法减少了沉淀和/或聚集引起的蛋白质丢失,并提高了异源多聚体蛋白质生产(如双特异性抗体生产)的总体产率。
在一方面,本发明提供形成或产生异源多聚体蛋白质的方法,该方法包括:
a.在第一加溶剂的存在下,在ph4-9,优选5-9,提供第一含铰链多肽,其中该第一含铰链多肽包含异源多聚化结构域;
b.在第二加溶剂的存在下,在ph4-9,优选5-9,提供第二含铰链多肽,其中该第二含铰链多肽包含异源多聚化结构域;
c.在还原条件下混合该第一和第二含铰链多肽,以形成组装混合物;和
d.孵育该组装混合物,以形成或产生包含该第一和第二含铰链多肽的异源多聚体蛋白质,其中该第一含铰链多肽与该第二含铰链多肽在所述异源多聚化结构域处相互作用。
在这方面的某些实施方案中,步骤a和/或步骤b之前是纯化该第一和/或第二含铰链多肽的步骤。在某些具体实施方案中,通过蛋白a来纯化该第一和/或第二含铰链多肽。
在另一方面,本发明提供形成或产生双特异性抗体的方法,该方法包括:
a.在第一加溶剂的存在下,在ph4-9,优选5-9,提供第一半抗体,其中该第一半抗体包含异源多聚化结构域;
b.在第二加溶剂的存在下,在ph4-9,优选5-9,提供第二半抗体,其中该第二半抗体包含异源多聚化结构域;
c.在还原条件下混合该第一和第二半抗体,以形成组装混合物;和
d.孵育该组装混合物,以形成或产生包含该第一和第二半抗体的双特异性抗体,其中该第一半抗体与该第二半抗体在所述异源多聚化结构域处相互作用。
在这方面的某些实施方案中,步骤a和/或步骤b之前是纯化该第一和/或第二半抗体的步骤。在某些具体实施方案中,通过蛋白a来纯化该第一和/或第二半抗体。
在另一方面,本发明提供产生异源多聚体的方法,该方法包括提供包含精氨酸的含铰链多肽的混合物,该混合物具有4至9之间、优选5至9之间的ph,加入弱还原剂,并在便于产生异源多聚体的条件下孵育。
还在另一方面,本发明提供产生异源多聚体蛋白质的方法,该方法包括:
a.获得蛋白a纯化的第一含铰链多肽;
b.获得蛋白a纯化的第二含铰链多肽;
c.调节每种半抗体的ph至4和9之间;
d.混合该第一和第二含铰链多肽,以获得组装混合物;
e.向该组装混合物中加入摩尔过量的弱还原剂;和
f.孵育该组装混合物,以形成包含该第一和第二含铰链多肽的异源多聚体蛋白质。
在另一方面,本发明提供产生异源多聚体蛋白质的方法,该方法包括:
a.获得蛋白a纯化的第一含铰链多肽;
b.获得蛋白a纯化的第二含铰链多肽;
c.在l-精氨酸的存在下调节每种含铰链多肽的ph至4和9之间;
d.混合该第一和第二含铰链多肽,以获得混合的含铰链多肽池(pool);
e.孵育,以形成包含该第一和第二含铰链多肽的异源多聚体蛋白质。
在这方面的某些实施方案中,在还原条件下孵育该混合的含铰链多肽池。在某些实施方案中,含铰链多肽包括半抗体、免疫黏附素或其功能片段。在某些其他实施方案中,精氨酸以20mm-1m、20mm-200mm或50mm-200mm的浓度存在。在某些其他实施方案中,向步骤d或步骤e中加入pvp。在某些实施方案中,在混合后调节ph。
本申请人意外地发现,含铰链多肽(如半抗体)的中间ph保持可以促进构象转换,增强该含铰链多肽随后的组装。在某些实施方案中,该中间ph为ph4和9之间、优选5和9之间、或至少ph5、至少ph5.5、至少ph5.7、大于ph5、大于ph5.5、大于ph5.7、在5和9之间、5和8之间、5.5和8之间、5.5和9之间、5.7和8之间、5.7和9之间、6和8之间、6和9之间、7和8之间、7.5和8.5之间、或7和8.5之间。可以加入加溶剂来防止或最小化ph诱导的含铰链多肽的沉淀。在某些具体实施方案中,在中间ph保持之前加入加溶剂。在某些实施方案中,该第一加溶剂和第二加溶剂各自选自精氨酸、组氨酸和蔗糖,优选精氨酸和/或组氨酸。在某些其他实施方案中,精氨酸是精氨酸盐和/或组氨酸是组氨酸盐。在某些其他实施方案中,精氨酸是精氨酸衍生物和/或组氨酸是组氨酸衍生物。在某些其他实施方案中,精氨酸或组氨酸是l-精氨酸或l-组氨酸。在某些其他实施方案中,精氨酸或组氨酸是盐酸精氨酸或盐酸组氨酸。在某些其他实施方案中,精氨酸或组氨酸是磷酸精氨酸或磷酸组氨酸。在某些实施方案中,该第一和第二加溶剂是不同的,而在其他实施方案中,该第一和第二加溶剂是相同的。在某些优选实施方案中,该第一加溶剂和该第二加溶剂都包含精氨酸。在再其他实施方案中,精氨酸以20mm至1m、20mm至小于1m、20mm至100mm、20mm至200mm、20mm至300mm、20mm至400mm、50mm至100mm、50mm至150mm、50mm至200mm、50mm至250mm、或50mm至300mm、优选20mm至200mm之间的浓度存在。还在其他实施方案中,加溶剂包含精氨酸衍生物,包括但不限于乙酰精氨酸。在其他实施方案中,该第一加溶剂和第二加溶剂都包含以20mm至1m、20mm至小于1m、20mm至小于500mm、20mm至100mm、20mm至200mm、20mm至300mm、20mm至400mm、50mm至100mm、50mm至150mm、50mm至200mm、50mm至250mm、50mm至300mm、50mm至400mm、50mm至500mm、或50mm至600mm之间的浓度存在的组氨酸。在某些优选实施方案中,按50mm的浓度加入加溶剂。在某些其他实施方案中,按200mm的浓度加入加溶剂。在某些具体实施方案中,按20mm、50mm、100mm或200mm的浓度加入精氨酸或组氨酸。在某些其他具体实施方案中,在精氨酸和组氨酸二者的存在下提供该第一和/或第二含铰链多肽。在其他实施方案中,精氨酸和组氨酸各以20mm至1m、20mm至小于1m、20mm至100mm、20mm至200mm、20mm至300mm、20mm至400mm、50mm至100mm、50mm至150mm、50mm至200mm、50mm至250mm、或50mm至300mm、优选50mm至200mm的浓度存在。
在某些实施方案中,在中间ph保持(即ph调节)之前混合该第一和第二含铰链多肽。在某些其他实施方案中,于分别在第一含铰链多肽中和第二含铰链多肽中调节ph后,混合该第一和第二含铰链多肽。在某些实施方案中,在ph调节之前加入加溶剂。
在某些实施方案中,在混合之前分别纯化该第一含铰链多肽和该第二含铰链多肽;而在其他实施方案中,在混合后共纯化该第一含铰链多肽和该第二含铰链多肽。在某些具体实施方案中,含铰链多肽包含半抗体。在某些实施方案中,组装的异源多聚体蛋白质可以进行进一步纯化。
可以用任意适宜的方法进行纯化,其非限制性地包括通过蛋白a层析、蛋白g层析、疏水相互作用层析(hic)、免疫亲和柱上的分级分离、乙醇沉淀、硅石上或离子交换树脂(如deae)上的反向层析、层析聚焦、sds-page、硫酸铵沉淀、和使用例如sephadexg-75的凝胶过滤及其他类似的纯化方法、及其组合来纯化。
在某些实施方案中,通过蛋白a或蛋白g层析来纯化含铰链多肽,如半抗体。在另一实施方案中,在蛋白a纯化之前混合第一和第二含铰链多肽,并在蛋白a上共纯化。在某些实施方案中,在混合蛋白a纯化的多肽后调节ph。在其他实施方案中,在混合蛋白a纯化的多肽之前调节ph。在某些实施方案中,在ph调节之前加入加溶剂。
在某些其他实施方案中,通过hic或离子交换柱,纯化含铰链多肽。选择适宜的纯化方法在本领域技术人员的能力范围之内。例如,可以通过蛋白a柱后接着离子交换柱来纯化含铰链多肽;还可以通过蛋白a柱后接着凝胶过滤柱和/或hic来纯化含铰链多肽。在其他实例中,可以在通过蛋白a柱纯化之前通过一个或多个离子交换柱来纯化含铰链多肽。在某些实施方案中,在含铰链多肽的任何纯化步骤中所用的洗涤和/或洗脱缓冲液都不含精氨酸和/或组氨酸。
在其他实施方案中,将在酸性ph从蛋白a基质或其他柱基质洗脱的半抗体调节至中间ph。此后续ph调节(也称为中间ph保持)可以引起含铰链多肽(如半抗体)的沉淀,从而导致组装的异源多聚体蛋白质的产率降低。因此,在某些实施方案中,在ph调节之前在加溶剂的存在下提供在酸性ph下从蛋白a或蛋白g柱洗脱的半抗体。在ph调节步骤并非必要的情况下,在某些实施方案中,优选向纯化的含铰链多肽中加入加溶剂来防止或减少沉淀和/或聚集。
除中间ph保持外,本申请人意外地发现,加热可以增强含铰链多肽(如半抗体)的构象转换和/或组装。因此,在某些实施方案中,本发明方法的步骤a-d中的一个、多个或全部在15℃至39℃、15℃至42℃、18℃至37℃、20℃至42℃、20℃至25℃、25℃至42℃、25℃至35℃、25℃至39℃、30℃至35℃、32℃至35℃或32℃至37℃、优选35℃至37℃之间的温度加热至少30分钟。在某些实施方案中,孵育时间至多为72小时,尤其是在室温。在一些实施方案中,孵育时间为35℃3小时。在某些其他实施方案中,温度为或约为30℃、35℃或37℃。
但是,加热也可以增加聚集和/或沉淀。因此,在某些具体实施方案中,在加热之前向从蛋白a或蛋白g柱洗脱的半抗体中加入加溶剂。
在某些实施方案中,含铰链多肽包括半抗体、免疫黏附素或其功能片段。在某些具体实施方案中,含铰链多肽包含fc成分。
在某些具体实施方案中,第一和/或第二含铰链多肽包含半抗体。在某些实施方案中,半抗体是igg半抗体。在某些具体实施方案中,igg半抗体为igg1、igg4或igg2同种型。在某些有利的实施方案中,保留免疫球蛋白分子的信号肽,以便于该半抗体(尤其是在哺乳动物细胞中产生时)的分泌。在某些实施方案中,本发明的方法包括在浓度约为50mm的精氨酸的存在下,及备选地或此外在浓度约为200mm的组氨酸的存在下,在ph5-9提供第一和第二半抗体。在某些实施方案中,该第一和/或第二半抗体各包含对不同抗原或同一抗原上的不同表位特异的抗原结合结构域,组装的全抗体是双特异性抗体。在某些其他实施方案中,第一和第二半抗体属于同一同种型;而在其他实施方案中,第一和第二半抗体属于不同同种型。
半抗体可以包含vl结构域、vh结构域、铰链结构域、ch2结构域和/或ch3结构域。半抗体还可以是进一步包含系链(tether)的单链多肽,其中该单链多肽按n端至c端方向包含彼此相对位置如下的结构域:vl-系链-vh-铰链-ch2-ch3。在某些其他实施方案中,半抗体进一步包含cl结构域和ch1结构域;在其他实施方案中,半抗体可以是进一步包含系链的单链多肽,其中该单链多肽按n端至c端方向包含彼此相对位置如下的结构域:vl-cl-系链-vh-ch1-铰链-ch2-ch3。
系链可以包含一个或多个甘氨酸(g)和丝氨酸(s)残基。在其他实施方案中,系链包含ggs重复。系链的长度在例如15和50个氨基酸之间。在具体实施方案中,系链长度在20和32个氨基酸之间,例如长度为20、21、22、23、24、25、26、27、28、29、30、31或32个氨基酸。在某些实施方案中,系链是可切割的。在其他实施方案中,系链可以或不可以从蛋白质切下。在某些优选实施方案中,系链可以被同一个酶在位于或靠近系链的n端和c端的两个位点切割。在一个实施方案中,系链包含蛋白酶(如弗林蛋白酶(furin))的切割位点。在另一实施方案中,弗林蛋白酶在切割位点rxrxrr(seqidno:1)处切割系链,其中x是任意氨基酸。在一些实施方案中,第一含铰链多肽是半抗体,且第二含铰链多肽是单链半抗体。
在另一实施方案中,本发明提供包含系链和fc成分复合物的蛋白质,其中该系链可以或不可以从蛋白质切割。
在其他实施方案中,第一和第二含铰链多肽包含异源多聚化结构域。该异源多聚化结构域可以是杵入臼突变、亮氨酸拉链、静电等。该第一含铰链多肽可以包含杵,而该第二含铰链多肽可以包含臼。在某些实施方案中,含铰链多肽包含半抗体,第一半抗体包含杵,而第二半抗体包含臼。
在一些实施方案中,本发明方法包括在调节ph之前加入精氨酸至20mm至1m之间的终浓度。在一些实施方案中,加入精氨酸至50mm-600mm之间的终浓度。在一些实施方案中,加入精氨酸至50mm-100mm之间的终浓度。
在某些实施方案中,本发明方法包括在混合第一和第二半抗体之前在5至8之间的ph孵育各含铰链多肽蛋白a池(pool)。在其他实施方案中,混合该蛋白a池,然后调节ph到5至8之间。
在一些实施方案中,本文所述的方法包括在15℃-39℃之间、优选18℃-37℃之间、更优选20℃-25℃之间、更优选32℃-37℃之间的温度,孵育该混合的半抗体或混合的含铰链多肽池至少30分钟。
含铰链多肽可以通过例如细菌细胞、酵母细胞、昆虫细胞中的杆状病毒或哺乳动物细胞来产生。在某些实施方案中,通过细菌细胞,尤其是大肠杆菌(e.coli)来产生该含铰链多肽。在某些其他具体实施方案中,通过哺乳动物细胞,尤其是cho细胞来产生该含铰链多肽。在某些具体实施方案中,该含铰链多肽包含半抗体。
含铰链多肽可以通过异源二聚化结构域相互作用来形成二聚体或多聚体。在某些实施方案中,第一和第二含铰链多肽在异源二聚化结构域界面上的相互作用是凸起进入凹陷的相互作用、疏水相互作用和/或静电相互作用。在某些其他实施方案中,异源多聚化结构域包含杵(例如凸起)、臼(例如凹陷)、亮氨酸拉链、卷曲螺旋、或能够形成静电相互作用的极性氨基酸残基,或其组合。在某些实施方案中,第一含铰链多肽包含杵,第二含铰链多肽包含臼。在某些其他实施方案中,该相互作用涉及疏水相互作用和静电相互作用二者。在某些示例性实施方案中,第一和第二含铰链多肽各自的异源多聚化结构域都包含杵或臼和能够形成静电相互作用的氨基酸残基。本领域技术人员理解,异源二聚化结构域可以包含一种以上相互作用方式,例如杵和臼(k&h)及疏水相互作用、k&h和亮氨酸拉链等。在某些实施方案中,含铰链多肽进一步包含系链。在某些具体实施方案中,含铰链多肽包含半抗体。
在某些具体实施方案中,组装混合物存在于还原条件中,优选弱还原条件中,具有-50至-600mv、-100至-600mv、-200至-600mv、-100至-500mv、-150至-300mv之间、更优选-300至-500mv之间、最优选约-400mv的氧化电位,以及在促进异源多聚体蛋白质组装的条件下,如ph在7和9之间,温度在15℃和39℃之间。在某些实施方案中,在组装期间向步骤c或步骤d中加入还原剂来制备希望的还原条件。在某些其他实施方案中,还原剂选自二硫苏糖醇(dtt)、三(2-羧乙基)膦(tcep)、巯基乙酸、抗坏血酸、硫羟乙酸、谷胱甘肽(gsh)、β-巯基乙胺、半胱氨酸/胱氨酸、谷胱甘肽(gsh)、半胱胺/胱胺、甘氨酰半胱氨酸、和β-巯基乙醇,优选gsh。在某些优选实施方案中,还原剂,优选弱还原剂,选自谷胱甘肽(gsh)、β-巯基乙胺、半胱氨酸/胱氨酸、谷胱甘肽(gsh)/谷胱甘肽二硫化物(gssg)、半胱胺/胱胺、甘氨酰半胱氨酸、和β-巯基乙醇,且优选gsh。在某些实施方案中,还原剂不是dtt。
在某些其他实施方案中,相对于含铰链多肽的总量,向组装混合物中加入2-600x、2-200x、2-300x、2-400x、2-500x、2-20x、2-8x、20-50x、50-600x、50-200x或100-300x摩尔过量、优选50-400x、更优选100-300x和最优选200x摩尔过量的还原剂。在某些实施方案中,组装混合物具有7和9之间的ph,优选ph8.5。在某些实施方案中,含铰链多肽是半抗体。
在一些实施方案中,在混合前向第一和第二含铰链多肽中加入还原剂。优选地,该加入在混合前不到1小时、更优选不到15分钟、最优选不到5分钟进行。
在某些实施方案中,该方法进一步包括在一个或多个步骤中向反应中加入稳定剂,所述步骤非限制性地包括:中间ph保持步骤和组装步骤(在加热或不加热情况下)。例如,可以向含铰链多肽中加入稳定剂来防止或减少聚集。在其他实例中,可以向组装混合物中加入稳定剂来防止或减少在组装异源多聚体蛋白质期间的聚集。在某些具体实施方案中,含铰链多肽包含半抗体。
在某些具体实施方案中,稳定剂选自精氨酸、组氨酸和聚乙烯吡咯烷酮(pvp)。在某些实施方案中,精氨酸或组氨酸是精氨酸盐或组氨酸盐。在某些其他实施方案中,精氨酸或组氨酸是精氨酸衍生物或组氨酸衍生物。在某些其他实施方案中,精氨酸或组氨酸是盐酸精氨酸或盐酸组氨酸。在某些实施方案中,精氨酸不是磷酸精氨酸。
在某些其他实施方案中,本发明方法进一步包括在pvp的存在下孵育组装混合物的步骤。在相关实施方案中,加入pvp至不超过40%(w/v)。在某些其他实施方案中,pvp以2%-6%(w/v)、10%-20%、2%-10%、1%、1.3%、1.7%、2%、2.3%、2.7%、3%、3.3%、3.7%或4%、优选0.1%-10%、更优选2%-6%和最优选4%的浓度存在于组装混合物中。在某些实施方案中,pvp不超过100kd、不超过30kd,且优选10kd。在某些其他实施方案中,pvp以不到10%(w/w)或不到5%(w/w)存在。
在一些实施方案中,稳定剂是以20mm至1m、20mm至小于1m、20mm至100mm、20mm至200mm、20mm至300mm、20mm至400mm、20mm至50mm、50mm至100mm、50nm至150mm、50mm至200mm、50mm至250mm、或50mm至300mm之间的浓度存在的精氨酸。在其他实施方案中,稳定剂是以20mm至1m、20mm至小于1m、20mm至100mm、20mm至200mm、20mm至300mm、20mm至400mm、20mm至50mm、50mm至100mm、50nm至150mm、50mm至200mm、50mm至250mm、或50mm至300mm之间的浓度存在的组氨酸。在某些优选实施方案中,以50mm或200mm的浓度加入精氨酸或组氨酸。在某些其他实施方案中,以20mm至200mm、20mm至100mm、50mm至200mm、或50mm至100mm的浓度加入精氨酸和/或组氨酸。在某些其他实施方案中,含铰链多肽包含半抗体。
在再一方面,本发明提供表达含铰链多肽的宿主细胞。在某些实施方案中,含铰链多肽包含半抗体。
在另一方面,本发明提供产生双特异性抗体的方法,其包括步骤:(a)培养经改造以表达对第一抗原或抗原的第一表位特异的第一半抗体的第一宿主细胞;(b)培养经改造以表达对第二抗原或同一抗原的第二表位特异的第二半抗体的第二宿主细胞;(c)在第一加溶剂的存在下,在4-9、优选5-9之间的ph,从步骤a的培养物获得第一半抗体;(d)在第二加溶剂的存在下,在4-9、优选5-9之间的ph,从步骤b的培养物获得第二半抗体;(e)在还原条件中混合第一和第二半抗体,以形成组装混合物;和(f)孵育组装混合物,以形成包含第一和第二半抗体的双特异性抗体。
在一些实施方案中,在分开的培养物中培养第一宿主细胞和第二宿主细胞,并在混合前分别从第一和第二宿主细胞的培养物纯化第一半抗体和第二半抗体。在某些实施方案中,在分开的培养物中培养第一和第二宿主细胞,组合培养物,沉淀细胞,可选地匀浆和/或裂解,以及通过任何适宜的方法共纯化第一和第二半抗体。在某些实施方案中,通过蛋白a纯化来共纯化第一和第二半抗体。在其他实施方案中,在混合培养物中共培养第一和第二宿主细胞,并共纯化第一和第二半抗体。
宿主细胞可以是例如细菌细胞、酵母细胞、植物细胞、昆虫细胞或哺乳动物细胞。在某些具体实施方案中,通过诸如cho细胞的哺乳动物细胞来产生含铰链多肽或半抗体。在某些其他实施方案中,宿主细胞是细菌细胞,尤其是大肠杆菌。
在某些其他实施方案中,本发明的方法进一步包括步骤:回收步骤(d)中形成的异源多聚体蛋白质或双特异性抗体。组装的异源多聚体蛋白质可以通过本申请通篇描述的方法或本领域已知的适宜方法来进一步纯化。
在另一方面,本发明提供包含含铰链多肽和稳定剂的组合物,其中组合物的ph在ph4-ph9之间,优选ph5-9之间。在某些实施方案中,组合物的ph为至少ph5、至少ph5.5、至少ph5.7、大于ph5、大于ph5.5、大于ph5.7、5和9之间、5和8之间、5.5和8之间、5.5和9之间、5.7和8之间、5.7和9之间、6和8之间、6和9之间、7和8之间、7.5和8.5之间、或7和8.5之间。在某些实施方案中,稳定剂选自精氨酸、组氨酸和蔗糖,优选精氨酸和/或组氨酸。在某些其他实施方案中,精氨酸是精氨酸盐和/或组氨酸是组氨酸盐。在某些其他实施方案中,精氨酸是精氨酸衍生物和/或组氨酸是组氨酸衍生物。在某些其他实施方案中,精氨酸或组氨酸是l-精氨酸或l-组氨酸。在某些其他实施方案中,精氨酸或组氨酸是盐酸精氨酸或盐酸组氨酸。
在某些具体实施方案中,稳定剂是精氨酸或组氨酸。在某些其他实施方案中,精氨酸或组氨酸以20mm、50mm、200mm、400mm、20mm至1m之间、20mm至小于1m之间、20mm至200mm之间、20mm至400mm之间、20mm至100mm之间、50mm至100mm之间、50mm至200mm之间、50mm至300mm之间或50mm至400mm之间的浓度存在。在某些具体实施方案中,组合物包含各以20mm、50mm、200mm、400mm、20mm至1m之间、20mm至小于1m之间、20mm至200mm之间、20mm至400mm之间、20mm至100mm之间、50mm至100mm之间、50mm至200mm之间、50mm至300mm之间、或50mm至400mm之间的浓度存在的精氨酸和组氨酸二者。在某些其他实施方案中,组合物不包含盐酸胍或尿素。在某些实施方案中,组合物备选地或额外地包含稳定剂。
在某些具体实施方案中,含铰链多肽包含半抗体。在某些其他具体实施方案中,组合物仅包含一种类型的含铰链多肽或半抗体。在某些实施方案中,组合物仅包含一种类型的半抗体,即,杵半抗体。在某些其他实施方案中,组合物仅包含一种类型的半抗体,即,臼半抗体。
在某些具体实施方案中,组合物进一步包含第二含铰链多肽,其中第一含铰链多肽包含杵,而第二含铰链多肽包含臼。在某些实施方案中,含铰链多肽包含半抗体。在某些具体实施方案中,含铰链多肽是半抗体。在某些其他实施方案中,半抗体为igg1、igg2或igg4同种型。
除非文中清楚地另有说明,可以组合本文中公开的所有实施方案。此外,除非文中清楚地另有说明,上文所述的任一个和每一个实施方案都适用于本发明的任一个和每一个方面。
本发明的其他目的、特征和优势将从以下发明详述变得显而易见。但是,应理解,虽然指出了本发明的优选实施方案,但发明详述和具体实施例仅以举例说明的方式给出,对本领域技术人员而言,落入本发明的范围和精神之内的各种改变和修改将从本发明详述变得显而易见。
附图简述
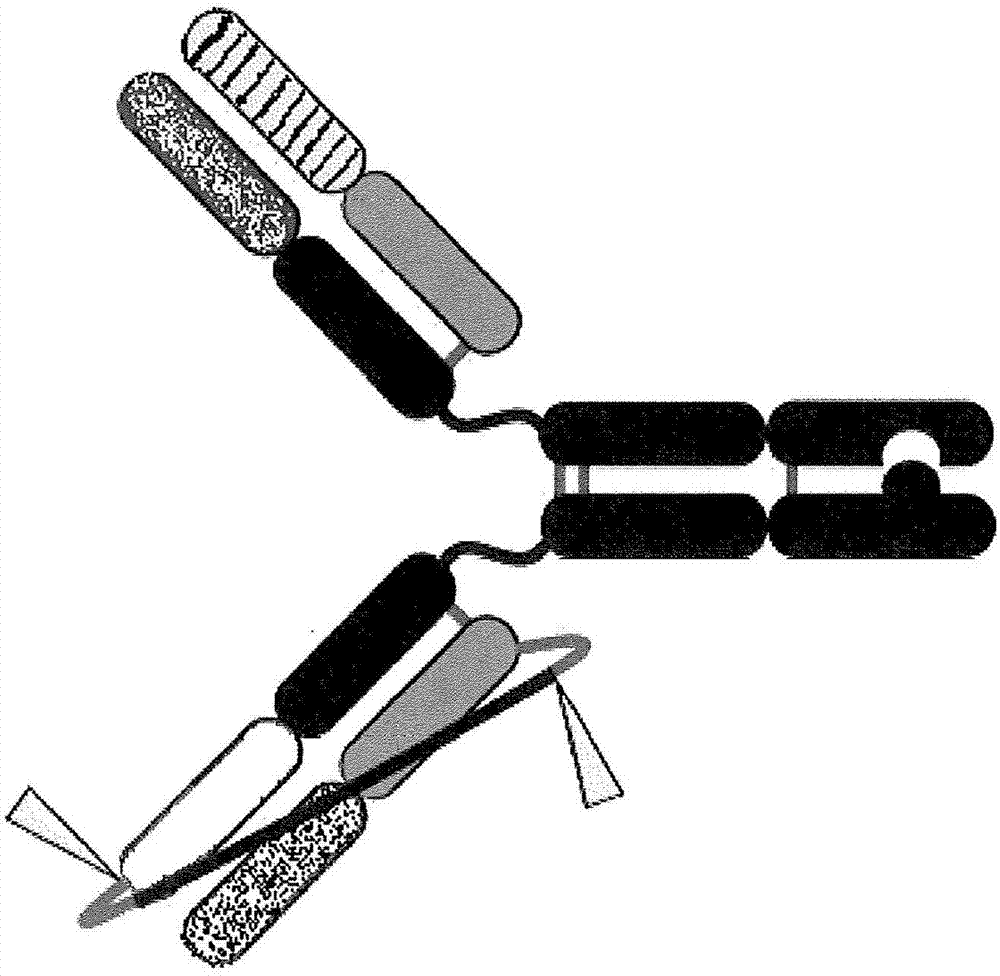
图1a显示完全氧化的半抗体。未显示“杵”或“臼”或其他异源二聚化结构域。此图中所示的半抗体是igg1同种型。本领域技术人员将理解,可以设想其他免疫球蛋白同种型作为具有相应的链间和链内键的半抗体。在完整抗体中,铰链半胱氨酸将形成链间二硫键。
图1b显示具有异源多聚化结构域的全长双特异性抗体。未显示铰链区中的重链间二硫键。所显示的异源多聚化结构域是杵入臼形式。
图1c是包含异源多聚化结构域(杵入臼)、弗林蛋白酶可切割系链和可选的额外二硫键(s354)的双特异性抗体的卡通示意图。还显示了铰链区中的重链间二硫键。三角形指示弗林蛋白酶切割位点。虽然弗林蛋白酶可切割系链显示在包含杵的半抗体上,但它也可以用在臼半抗体上或杵和臼半抗体二者上。
图2a-b显示复合大小排阻层析图,说明ph对半抗体构象转变的作用。图2a显示提高的ph诱导臼半抗体构象转变,导致更大的流体动力学半径。这种构象转变增强了组装期间的异二聚化。图2b显示提高的ph促进非共价杵半抗体同二聚体的形成。这种构象转变有利于组装期间的双特异性形成。参考实施例2。
图3a-3b显示这样的结果,其显示诸如盐酸精氨酸(图3a和b)或组氨酸(图3b)的稳定剂减少中间ph诱导的杵半抗体的沉淀。
图4a-b显示组成层析图,说明诸如谷胱甘肽的还原剂对聚集和双特异性抗体组装的作用。按2-200x摩尔过量加入谷胱甘肽。参考实施例3。
图5a的图显示温度对igg1双特异性抗体形成(组装)速率的影响。图5b显示,如通过反相层析分析的,在200x摩尔过量的谷胱甘肽存在下、在有或无ph保持下、提高的温度促进杵入臼双特异性igg1抗体的组装。图5b还显示,在组装前优化半抗体池的ph保持以驱动构象转变,提高了组装的速度和效率。参考实施例4。
图6a显示诸如pvp的稳定剂对稳定所形成的双特异性抗体和减少聚集体形成上的作用。
图6b显示,如通过反相层析分析,pvp和组氨酸在提高的温度下促进杵入臼双特异性igg4抗体的组装。下部曲线:室温组装,其中300x谷胱甘肽:ab比、ph=8.5、~400mm精氨酸;上部曲线:加热组装,其中200x谷胱甘肽:ab比、ph=8.0、4%pvp、50mm精氨酸、100mm组氨酸,在35℃。
图6c证明pvp减少igg4杵入臼双特异性抗体在37℃和ph8下加热组装6小时期间的聚集。参考实施例5。
图7显示组氨酸减少igg4臼半抗体在37℃和ph8下3小时由热诱导的聚集。
发明详述
现对本发明进行详细描述,使用以下定义和实施例。本文中提到的所有专利和出版物,包括这些专利和出版物中公开的所有序列,明确引入作为参考。
除非本文中另作定义,本文中使用的所有技术和科学术语具有本发明所属领域普通技术人员通常理解的含义。singleton等,dictionaryofmicrobiologyandmolecularbiology,第2版,johnwileyandsons,newyork(1994),和hale&marham,theharpercollinsdictionaryofbiology,harperperennial,ny(1991)为本领域技术人员提供了许多本发明中使用的术语的一般字典。本文描述了优选的方法和材料,但与本文中所述的那些方法和材料类似或等同的任意方法和材料都可以用于实施或测试本发明。数字范围包含定义该范围的数字。除非另有说明,按5’至3’的方向从左至右书写核酸;按氨基至羧基的方向从左至右书写氨基酸序列。对于本领域的定义和术语,从业者尤其可参考sambrook等,1989和ausubelfm等,1993。应理解,本发明不限于所述的具体方法、工艺和试剂,因为这些可以变动。
数字范围包含定义该范围的数字。
除非另有说明,按5’至3’的取向从左至右书写核酸;按氨基至羧基的取向从左至右书写氨基酸序列。
除非文中清楚地另有说明,本文中所用的单数形式“一”、“一个”和“该”包括复数指代物。例如,提到“一个宿主细胞”意指一个或多个宿主细胞。
本文中提供的标题不对本发明的各个方面或实施方案构成限制,本发明的方法和实施方案可参考说明书整体而得。因此,下文即将定义的术语通过参考说明书整体而得到更充分的定义。
除非文中清楚地另有说明,本文中公开的所有实施方案可以组合。此外,除非文中清楚地另有说明,下文所述的任一个和每一个实施方案适用于本发明的任一个和每一个方面。
i.定义
本发明提供产生包含第一含铰链多肽和第二含铰链多肽的异源多聚体蛋白质的方法。本文所用的术语“含铰链多肽”指,包含至少一个铰链区的多肽。在某些实施方案中,铰链区连接多个结构域,例如结合结构域和效应子结构域,并为多肽的二聚化或多聚化提供一定的结构柔性。作为一个实例,结合结构域可以是抗体的抗原结合结构域或受体的配体结合结构域,效应子结构域可以是抗体的fc成分。在某些实施方案中,第一含铰链多肽不同于第二含铰链多肽,得到的二聚体或多聚体是异二聚体或异源多聚体。在某些具体实施方案中,第一含铰链多肽和第二含铰链多肽结合同一靶蛋白上的两个不同表位。在某些其他实施方案中,第一含铰链多肽具有不同于第二含铰链多肽的靶标结合特异性,得到的异二聚体或异源多聚体结合两种或多种不同的靶蛋白。在某些实施方案中,含铰链多肽包含天然存在的或改造的异二聚化结构域。在某些具体实施方案中,含铰链多肽在铰链区中包含一个或多个半胱氨酸残基,其能够与另一含铰链多肽形成一个或多个二硫键。
含铰链多肽非限制性地包括半抗体、免疫黏附素及其功能片段。本文所用的术语“功能片段”指含铰链多肽的片段,即小于全长,其仍保留希望的功能,例如,保留靶标或抗原结合活性、fc效应子活性和/或二聚化/多聚化能力。在某些具体实施方案中,第一含铰链多肽和第二含铰链多肽各为具有不同抗原结合特异性的半抗体,得到的二聚体或多聚体是双特异性或多特异性抗体。在某些实施方案中,得到的异源多聚体蛋白质包含半抗体和免疫黏附素。
术语“多特异性抗体”以最广泛的含义使用,特别涵盖具有多表位特异性的抗体。这类多特异性抗体包括但不限于:包含重链可变结构域(vh)和轻链可变结构域(vl)的抗体,其中vhvl单元具有多表位特异性;具有两个或多个vl和vh结构域的抗体,每个vhvl单位结合不同的表位;具有两个或多个单可变结构域的抗体,每个单可变结构域结合不同的表位;全长抗体;抗体片段,如fab、fv、dsfv、scfv;双抗体;双特异性双抗体和三抗体;共价或非共价连接的抗体片段。“多表位特异性”指特异性结合一个或多个相同或不同靶标上的两个或多个不同表位的能力。“单特异性”指仅结合一种表位的能力。根据一个实施方案,多特异性抗体是以5μm至0.001pm、3μm至0.001pm、1μm至0.001pm、0.5μm至0.001pm、或0.1μm至0.001pm的亲和力结合各表位的igg抗体。
天然存在的基本4链抗体单位是由两条相同的轻(l)链和两条相同的重(h)链组成的异源四聚体糖蛋白(igm抗体由5个基本的异源四聚体单位连同称为j链的附加多肽组成,因此包含10个抗原结合部位,而分泌性iga抗体可以多聚化形成包含2-5个基本4链单位连同j链的多价集聚体)。在igg的情况下,4链单位通常为约150,000道尔顿。每个l链通过一个共价二硫键与h链连接,而取决于h链同种型,两条h链通过一个或多个二硫键相互连接。每条h和l链还具有规则间隔开的链内二硫键。每条h链在n端具有可变结构域(vh),对于α和γ链,后随三个恒定结构域(ch),对于μ和ε同种型,后随四个ch结构域。每条l链在n端具有可变结构域(vl),后随c端的恒定结构域(cl)。vl与vh对齐,而cl与重链的第一恒定结构域(ch1)对齐。认为特定的氨基酸残基形成轻链和重链可变结构域之间的界面。vh和vl配对在一起形成单个抗原结合部位。对于不同种类抗体的结构和性质,见例如basicandclinicalimmunology,第8版,danielp.stites,abbai.ter和tristramg.parslow(编辑),appleton&lange,norwalk,ct,1994,第71页和第6章。
可以将来自任意脊椎动物物种的l链,根据其恒定结构域的氨基酸序列,分至两个明显不同的类型之一,称为κ和λ。取决于其重链恒定结构域(ch)的氨基酸序列,可以将免疫球蛋白分至不同的类或同种型。存在五类免疫球蛋白:iga、igd、ige、igg和igm,其分别具有称为α、δ、ε、γ和μ的重链。根据ch序列和功能上相对小的差异,将γ和α类进一步划分为亚类,例如,人表达以下亚类:igg1、igg2、igg3、igg4、iga1和iga2。
术语“可变的”指可变结构域的某些区段在抗体间在序列上十分不同的事实。v结构域介导抗原结合,并限定特定抗体对其特定抗原的特异性。但是,可变性并非在可变区的110个氨基酸的跨度内均匀分布。相反,v区由15-30个氨基酸的称为构架区(fr)的相对不变的区段组成,构架区由长度各为9-12个氨基酸的称为“高变区”的极端可变性的较短区域分开。天然重链和轻链的可变结构域各包含四个fr,其大部分采用β-折叠构象,通过三个高变区连接,高变区形成连接β-折叠结构的环,且在一些情况下形成β-折叠结构的部分。每条链中的高变区通过fr紧靠在一起,并与来自另一条链的高变区一起促成抗体的抗原结合部位的形成(见kabat等,sequencesofproteinsofimmunologicalinterest,第5版publichealthservice,nationalinstitutesofhealth,bethesda,md(1991))。恒定结构域不直接参与抗体与抗原的结合,但显示多种效应子功能,如在抗体依赖性细胞毒性(adcc)中抗体的参与。
在本文中使用时,术语“高变区”、“hvr”或“hv”指抗体可变结构域中在序列上高变和/或形成结构上限定的环的区域。通常,抗体包含六个hvr;三个在vh(h1、h2、h3)中,三个在vl(l1、l2、l3)中。在天然抗体中,h3和l3显示六个hvr中最大的多样性,尤其h3被认为在赋予抗体良好的特异性中发挥独特作用。见例如xu等,immunity13:37-45(2000);johnson和wu,inmethodsinmolecularbiology248:1-25(lo,ed.,humanpress,totowa,nj,2003)。事实上,仅由重链组成的天然驼类抗体在缺乏轻链的情况下具有功能且稳定。见例如hamers-casterman等,nature363:446-448(1993);sheriff等,naturestruct.biol.3:733-736(1996)。
许多hvr划界法在使用,并为本文所涵盖。kabat互补决定区(cdr)基于序列可变性,且最常用((kabat等,sequencesofproteinsofimmunologicalinterest,第5版publichealthservice,nationalinstitutesofhealth,bethesda,md.(1991))。而chothia指结构环的位置(chothia和leskj.mol.biol.196:901-917(1987))。abmhvr代表了在kabathvr和chothia结构环之间的折中,且为oxfordmolecular'sabm抗体模拟软件所使用。“接触”hvr所基于的是对可用的复合物晶体结构的分析。下文指出了来自这些hvr的每一个的残基。
hvr可以包含以下“延长的hvr”:vl中24-36或24-34(l1)、46-56或50-56(l2)和89-97或89-96(l3);vh中26-35(h1)、50-65或49-65(h2)和93-102、94-102或95-102(h3)。对于这些定义中的每一个,按照kabat等编号可变结构域残基。
“构架区”(fr)是除cdr残基外的那些可变结构域残基。每个可变结构域通常具有标识为fr1、fr2、fr3和fr4的四个fr。如果按照kabat定义cdr,则轻链fr残基位于大约残基1-23(lcfr1)、35-49(lcfr2)、57-88(lcfr3)和98-107(lcfr4),重链fr残基位于大约重链残基中的残基1-30(hcfr1)、36-49(hcfr2)、66-94(hcfr3)和103-113(hcfr4)。如果cdr包含来自高变环的氨基酸残基,则轻链fr残基位于轻链中大约残基1-25(lcfr1)、33-49(lcfr2)、53-90(lcfr3)和97-107(lcfr4),重链fr残基位于在重链残基中大约残基1-25(hcfri)、33-52(hcfr2)、56-95(hcfr3)和102-113(hcfr4)。在一些情况下,在cdr包含来自kabat定义的cdr和高变环的氨基酸时,将相应地调整fr残基。例如,在cdrh1包括氨基酸h26-h35时,重链fr1残基位于1-25位,fr2残基位于36-49位。
“人共有构架”是指这样的构架,其代表了一组选择的人免疫球蛋白vl或vh构架序列中最常见的氨基酸残基。通常,选择的该组人免疫球蛋白vl或vh序列来自一个可变结构域序列亚组。通常,该序列亚组是kabat中的亚组。在一个实施方案中,对于vl,该亚组是kabat中的亚组κi。在一个实施方案中,对于vh,该亚组是kabat中的亚组iii。
“完整”抗体的一个实例是包含抗原结合部位以及cl和至少重链恒定结构域ch1、ch2和ch3的抗体。恒定结构域可以是天然序列恒定结构域(例如人天然序列恒定结构域)或其氨基酸序列变体。
“抗体片段”包含完整抗体的部分,优选完整抗体的抗原结合区或可变区。抗体片段的实例包括fab、fab’、f(ab’)2和fv片段;双抗体(db);串联双抗体(tadb);线性抗体(例如美国专利号5,641,870,实施例2;zapata等,proteineng.8(10):1057-1062(1995));单臂抗体;单可变结构域抗体;微抗体;单链抗体分子;自抗体片段形成的多特异性抗体(例如,包括但不限于db-fc、tadb-fc、tadb-ch3和(scfv)4-fc)。
本文所用的术语“半抗体”指与一条免疫球蛋白轻链结合的一条免疫球蛋白重链。图1a中显示示例性半抗体。本领域技术人员将易于理解,半抗体可以涵盖其片段,并且也可以具有由单可变结构域组成的抗原结合结构域(例如源自驼科(camelidae))。
本发明人意外地发现,对低ph下从蛋白a柱或其他基质洗脱的半抗体的ph优化或调节诱导含铰链多肽诸如半抗体的构象转变。ph优化至中间ph(本公开通篇中有时称为ph保持或中间ph保持)可以导致半抗体的沉淀或聚集。因此,在某些实施方案中,本发明产生异源多聚体蛋白质的方法包括步骤:分别在第一或第二加溶剂的存在下、在ph5-9下提供第一或第二含铰链多肽。
本文中所用的加溶剂定义为:防止或减少含铰链多肽(如半抗体)沉淀的试剂。适宜的加溶剂包括但不限于精氨酸和组氨酸,或其盐或衍生物,及蔗糖。在某些实施方案中,加溶剂是精氨酸和/或组氨酸。在某些实施方案中,加溶剂防止或减少由中间ph保持和/或加热诱导的沉淀。在某些具体实施方案中,在中间ph保持(即在调节至中间ph之前)和/或加热之前加入加溶剂。在某些实施方案中,精氨酸或组氨酸是精氨酸盐或组氨酸盐。在某些其他实施方案中,精氨酸或组氨酸是精氨酸衍生物或组氨酸衍生物。沉淀的减少可以导致希望的组装终产物的产率提高。
对于一般的蛋白质制备和纯化,咪唑和胍一直用来溶解蛋白质。但是,意外地发现,咪唑和胍单独,在没有组氨酸或精氨酸的情况下,不足以提高本文所述的组装的异源多聚体蛋白质(如双特异性抗体)的总体产率。在某些实施方案中,咪唑和鸟苷可以变性蛋白质。
类似地,诸如盐酸胍和尿素的去垢剂一般常用来减少聚集/沉淀,但它们可以彻底变性蛋白质。因此,在某些实施方案中,本发明加溶剂防止或减少沉淀而不变性目的蛋白质。因此,在某些具体实施方案中,本发明加溶剂不是盐酸胍、胍、咪唑或尿素。在某些其他实施方案中,本发明的组合物不包含盐酸胍或尿素。在某些其他实施方案中,加溶剂不是吐温(tween)或peg。
此外,可以例如在各半抗体的中间ph保持期间或在组装期间于混合含铰链多肽或半抗体时或紧接混合后,加入稳定剂。可以在组装之前、组装期间和/或组装之后,在本发明方法的一个或多个或全部步骤的反应中加入稳定剂来防止或减少含铰链多肽或半抗体的聚集。
聚集体可以作为高分子量物质被检测到,在半抗体的情况下,可以作为分子量大于150kda的高分子量物质被检测到。可以通过例如大小排阻层析或其他适宜的方法来检测和定量聚集体。在某些其他实施方案中,通过大小排阻层析检测到的聚集体可以通过0.2μm除菌滤器。另一方面,沉淀的蛋白质可以由变性蛋白质或聚集蛋白质组成,其可形成非常巨大的复合物。已测试了几种试剂,并确定其用作稳定剂是无效的或不合适的,例如咪唑、3-(n-吗啉代)丙磺酸(mops)、2-(n-吗啉代)乙磺酸(mes)、环糊精、cuso4和naoac。因此,在某些实施方案中,稳定剂不包括无机盐或过渡金属。适宜的稳定剂包括但不限于pvp、组氨酸和精氨酸。聚集体的减少可以导致希望的组装终产物的产率提高。
pvp是具有吡咯烷酮基团的水溶性不带电荷聚合物。在某些实施方案中,其他不带电荷的极性聚合物、其他试剂或化合物、尤其是与本文中所述的适宜稳定剂具有相似的结构和特性的化合物,可以是适用于本发明的稳定剂。本领域技术人员能够通过本领域已知的方法(包括本文中提供的方法),分析化合物对聚集水平的影响,从而确定适宜的稳定剂。
试剂可以具有加溶剂和稳定剂二者的特征。例如,精氨酸可以在中间ph保持和/或加热期间用作加溶剂来减少半抗体的沉淀,并可以在组装步骤期间用作稳定剂来减少聚集。类似地,组氨酸可以在中间ph保持和/或加热期间,用作加溶剂来减少半抗体的沉淀以及用作稳定剂来减少半抗体的聚集。不受限于任何具体机制,在某些实施方案中,加溶剂和稳定剂二者都可以通过防止蛋白质表面上可以导致聚集的疏水斑块的相互作用来发挥作用。在其他实施方案中,加溶剂和稳定剂二者都可以通过形成笼形物来防止不希望的蛋白质相互作用,从而发挥作用。
本文所用的术语“单链半抗体”指包含vl结构域、可选的cl结构域、系链、vh结构域、可选的ch1结构域、铰链结构域、ch2结构域和ch3结构域的单链多肽,其中按n端至c端方向所述结构域彼此的相对位置如下:vl-系链-vh-铰链-ch2-ch3或vl-cl-系链-vh-ch1-铰链-ch2-ch3。
表述“单结构域抗体”(sdab)或“单可变结构域(svd)抗体”一般指,其中单个可变结构域(vh或vl)可以赋予抗原结合的抗体。换言之,为了识别靶抗原,单可变结构域无需与另一可变结构域相互作用。单结构域抗体的实例包括:衍生自驼类(羊驼和骆驼)和软骨鱼(例如护士鲨)的抗体、以及通过重组方法自人和小鼠抗体衍生的那些(nature(1989)341:544-546;devcompimmunol(2006)30:43-56;trendbiochemsci(2001)26:230-235;trendsbiotechnol(2003):21:484-490;wo2005/035572;wo03/035694;febslett(1994)339:285-290;wo00/29004;wo02/051870)。
表述“线性抗体”一般指zapata等,proteineng.8(10):1057-1062(1995)中所述的抗体。简言之,这些抗体包含一对串联fd区段(vh-ch1-vh-ch1),其与互补轻链多肽一起形成一对抗原结合区。线性抗体可以是双特异性或单特异性的。
本文中提到的“杵入臼”或“knh”技术指,通过在两条多肽相互作用的界面上,在一条多肽中引入凸起(杵),在另一条多肽中引入凹陷(臼),从而指导两条多肽在体外或体内配对在一起的技术。例如,已在抗体的fc:fc结合界面、cl:ch1界面或vh/vl界面中引入knh(例如us2007/0178552;wo96/027011;wo98/050431;zhu等(1997)proteinscience6:781-788)。这尤其可以用于在多特异性抗体制备过程中驱动两条不同的重链配对在一起。例如,在其fc区中具有knh的多特异性抗体可以进一步包含与各fc区连接的单可变结构域,或进一步包含与相似或不同的轻链可变结构域配对的不同重链可变结构域。knh技术还可以用来使两个不同的受体胞外域或包含不同靶标识别序列的任何其他多肽序列(例如,包括亲和体、肽体和其他fc融合物)配对在一起。
木瓜蛋白酶消化抗体产生两个相同的抗原结合片段(称为“fab”片段)和剩余的“fc”片段(该命名反映其易结晶的能力)。fab片段由整个l链连同一条重链的可变区结构域(vh)和该h链的第一恒定结构域(ch1)组成。胃蛋白酶处理抗体产生单个大的f(ab’)2片段,其大致对应于两个由二硫键连接的fab片段,具有二价抗原结合活性,且仍能够交联抗原。fab’片段与fab片段的不同在于,在ch1结构域的羧基端具有几个额外的残基,包括来自抗体铰链区的一个或多个半胱氨酸。fab’-sh是本文对恒定结构域的一个或多个半胱氨酸残基具有自由巯基的fab’的命名。f(ab’)2抗体片段最初作为其间具有铰链半胱氨酸的fab’片段对产生。还已知抗体片段的其他化学偶联。
fc片段包含通过二硫键靠在一起的两条h链的羧基端部分。抗体的效应子功能由fc区中的序列决定;此区域也是存在于某些类型的细胞上的fc受体(fcr)所识别的部分。
“fv”由紧密非共价结合的一个重链可变区结构域和一个轻链可变区结构域的二聚体组成。从这两个结构域的折叠,生出六个高变环(自h链和l链各产生三个环),其贡献用于抗原结合的氨基酸残基并赋予抗体抗原结合特异性。但是,甚至单可变结构域(或仅包含对抗原具有特异性的三个cdr的半个fv)也具有识别和结合抗原的能力,虽然亲和力常低于整个结合部位。
“单链fv”(也缩写为“sfv”或“scfv”)是包含连接为单条多肽链的vh和vl抗体结构域的抗体片段。优选地,sfv多肽进一步在vh和vl结构域之间包含多肽接头,其使得sfv能够形成希望的结构用于抗原结合。sfv的综述见pluckthun,thepharmacologyofmonoclonalantibodies,113卷,rosenburg和moore编辑,springer-verlag,纽约,269-315页(1994);malmborg等,j.immunol.methods183:7-13,1995。
术语“双抗体”指通过以下制备的小抗体片段:构建在vh和vl结构域之间具有短接头(约5-10个残基)的sfv片段(见前一段),使得达到v结构域的链间配对而不是链内配对,产生二价片段,即具有两个抗原结合部位的片段。双特异性双抗体是两个“交换”sfv片段的异二聚体,其中该两个抗体的vh和vl结构域存在于不同的多肽链上。双抗体更充分地描述于例如ep404,097;wo93/11161;和hollinger等,proc.natl.acad.sci.usa90:6444-6448(1993)中。
术语“单臂抗体”指,包含(1)通过肽键连接至含有ch2结构域、ch3结构域或ch2-ch3结构域的多肽上的可变结构域及(2)第二ch2、ch3或ch2-ch3结构域的抗体,其中可变结构域不通过肽键连接至含有该第二ch2、ch3或ch2-ch3结构域的多肽。在一个实施方案中,单臂抗体包含3条多肽:(1)含有可变结构域(例如vh)、ch1、ch2和ch3的第一多肽;(2)含有可变结构域(例如vl)和cl结构域的第二多肽;及(3)含有ch2和ch3结构域的第三多肽。在一个实施方案中,该第三多肽不包含可变结构域。在另一实施方案中,单臂抗体具有部分铰链区,该部分铰链区包含两个半胱氨酸残基,其形成二硫键而连接恒定重链。在一个实施方案中,单臂抗体的可变结构域形成抗原结合区。在另一实施方案中,单臂抗体的可变结构域是单可变结构域,其中每个单可变结构域都是抗原结合区。
本发明的抗体可以是“嵌合”抗体,其中部分重链和/或轻链与衍生自特定物种或隶属于特定抗体种类或亚类的抗体中对应的序列相同或同源,而所述链(一条或多条)的其余部分与衍生自另一物种或隶属于另一抗体种类或亚类的抗体中对应的序列相同或同源,以及这类抗体的片段,只要它们显示希望的生物学活性(美国专利号4,816,567;和morrison等,proc.natl.acad.sci.usa81:6851-6855(1984))。本文中的目的嵌合抗体包括含有衍生自非人灵长类(例如旧大陆猴、猿等)的可变结构域抗原结合序列和人恒定区序列的灵长类源化抗体。
非人(例如啮齿动物)抗体的“人源化”形式是含有衍生自非人抗体的最小序列的嵌合抗体。就大部分而言,人源化抗体是人免疫球蛋白(受体抗体),其中来自受体高变区的残基被替换为来自具有期望抗体特异性、亲和力和能力的非人物种(供体抗体)(如小鼠、大鼠、兔或非人灵长类)的高变区的残基。在一些情况下,用对应的非人残基替换所述人免疫球蛋白的构架区(fr)残基。此外,人源化抗体可以包含不存在于受体抗体中或供体抗体中的残基。进行这些修饰来进一步精化抗体性能。通常,人源化抗体将包含至少一个、通常两个可变结构域的基本上全部,其中高变环的全部或基本上全部对应于非人免疫球蛋白的高变环,而fr的全部或基本上全部是人免疫球蛋白序列的fr。人源化抗体还可以任选地包含至少部分免疫球蛋白恒定区(fc),通常是人免疫球蛋白的恒定区。进一步的细节见jones等,nature321:522-525(1986);riechmann等,nature332:323-329(1988);及presta,curr.op.struct.biol.2:593-596(1992)。
本文中所用的“复合物”或“复合的”指,通过并非肽键的键和/或力(例如范德瓦尔斯力、疏水力、亲水力)而彼此相互作用的两个或多个分子的结合。在一个实施方案中,复合物是异源多聚体。应理解,本文中所用的术语“蛋白质复合物”或“多肽复合物”包括这样的复合物,在该蛋白质复合物中具有缀合至蛋白质的非蛋白质实体(例如,包括但不限于化学分子,如毒素或检测剂)。
本文中所用的术语“异源多聚体”或“异源多聚体的”描述通过非肽共价键(例如二硫键)和/或非共价相互作用(例如氢键、离子键、范德瓦尔斯力和疏水相互作用)彼此相互作用的两个或更多个多肽,其中这些分子中的至少两个具有彼此不同的序列。
本文中所用的术语“异源多聚化结构域”指,为促进异源多聚体形成和阻碍同源多聚体形成,对生物分子的改变或添加。具有形成异二聚体超过同二聚体的强烈偏好的任意异源二聚化结构域都在本发明的范围之内。举例说明性实例包括但不限于,例如美国专利申请20030078385(arathoon等-genentech;描述杵入臼);wo2007147901(
本文所用的术语“免疫黏附素”指,将异源蛋白质(“黏附素”)的结合特异性与免疫球蛋白恒定结构域的效应子功能相组合的分子。在结构上,免疫黏附素包含具有希望的结合特异性的氨基酸序列(该氨基酸序列不同于抗体的抗原识别和结合部位(即,相对于抗体的恒定区,是“异源的”))和免疫球蛋白恒定结构域序列(例如igg的ch2和/或ch3序列)的融合物。示例性黏附素序列包括含有与目的蛋白质结合的受体或配体的部分的连续氨基酸序列。黏附素序列还可以是结合目的蛋白质但不是受体或配体序列的序列(例如肽体中的黏附素序列)。这类多肽序列可以通过包括噬菌体展示技术和高通量分选方法的多种方法来选择或鉴定。免疫黏附素中的免疫球蛋白恒定结构域序列可以获自任意免疫球蛋白,如igg-1、igg-2、igg-3或igg-4亚型、iga(包括iga-1和iga-2)、ige、igd或igm。
本发明的“结合”目的抗原的抗体是这样的抗体,其以足够的亲和力结合抗原,使得抗体可以作为诊断剂和/或治疗剂用于靶向蛋白质或表达抗原的细胞或组织,而不与其他蛋白质发生显著交叉反应。在这类实施方案中,通过荧光激活细胞分选(facs)分析或放射免疫沉淀(ria)或elisa测定,抗体与“非靶”蛋白结合的程度将小于抗体与其特定靶蛋白的结合的约10%。就抗体与靶分子的结合而言,术语“特异性结合”或“特异地结合”或“特异于”特定多肽或特定多肽靶标上的表位,是指该结合可测量地不同于非特异性相互作用(例如非特异性相互作用可以是与牛血清白蛋白或酪蛋白的结合)。特异性结合可以例如通过与对照分子的结合相比,测定分子的结合来测量。例如,特异性结合可以通过与类似于靶标的对照分子(例如过量的未标记靶标)的竞争来确定。在这种情况下,如果过量的未标记靶标竞争性地抑制标记靶标与探针的结合,则指示特异性结合。本文中,术语“特异性结合”或“特异地结合”或“特异于”特定多肽或特定多肽靶标上的表位,可以表现为:例如,分子对靶标具有至少约200nm、备选地至少约150nm、备选地至少约100nm、备选地至少约60nm、备选地至少约50nm、备选地至少约40nm、备选地至少约30nm、备选地至少约20nm、备选地至少约10nm、备选地至少约8nm、备选地至少约6nm、备选地至少约4nm、备选地至少约2nm、备选地至少约1nm,或更高的kd。在一个实施方案中,术语“特异性结合”指这样的结合,其中分子与特定多肽或特定多肽上的表位结合,而与任何其他多肽或多肽表位无实质性结合。
“结合亲和力”一般指,分子(例如抗体)的单个结合部位与它的结合配偶体(例如抗原)之间非共价相互作用的总和的强度。除非另有说明,本文所用的“结合亲和力”指固有的结合亲和力,其反映结合对的成员(例如抗体和抗原)之间的1:1相互作用。分子x对它的配偶体y的亲和力一般可通过解离常数(kd)来表示。例如,kd可以是约200nm、150nm、100nm、60nm、50nm、40nm、30nm、20nm、10nm、8nm、6nm、4nm、2nm、1nm或更强。亲和力可以通过本领域公知的方法来测量,包括本文中描述的那些。低亲和力抗体一般缓慢地结合抗原并倾向于容易解离,而高亲和力抗体一般更快地结合抗原并倾向于保持更长久的结合。多种测量结合亲和力的方法为本领域已知,其中的任一种都可以用于本发明的目的。
在一个实施方案中,通过表面等离振子共振测定法,使用biacoretm-2000或biacoretm-3000(biacore,inc.,piscataway,nj),在25℃,用固定抗原的cm5芯片,以~10响应单位(ru),测量本发明的“kd”或“kd值”。简言之,按照厂家的说明,用n-乙基-n’-(3-二甲基氨基丙基)-碳二亚胺盐酸盐(edc)和n-羟基琥珀酰亚胺(nhs),活化羧甲基化葡聚糖生物传感芯片(cm5,biacoreinc.)。用10mm乙酸钠ph4.8将抗原稀释为5μg/ml(~0.2μm),然后按5μl/分钟的流速注射,以达到约10个响应单位(ru)的偶联蛋白质。注入抗原后,注入1m乙醇胺来封闭未反应的基团。对于动力学测量,按约25μl/分钟的流速在25℃注入在含0.05%tween20的pbs(pbst)中fab的两倍系列稀释物(例如0.78nm至500nm)。通过同时拟合结合和解离传感图,用简单的一对一langmuir结合模型(biacoreevaluationsoftwareversion3.2),计算结合速率(kon)和解离速率(koff)。将平衡解离常数(kd)计算为比值koff/kon。见例如chen等,j.mol.biol.293:865-881(1999)。如果通过以上表面等离振子共振测定的结合速率超过106m-1s-1,则可以通过使用荧光淬灭技术来确定结合速率,荧光淬灭技术测量在浓度递增的抗原存在下、pbsph7.2中的20nm抗-抗原抗体(fab形式)在25℃的荧光发射强度(激发=295nm;发射=340nm;16nm带通)的增加或减少,如在分光光度计,如配有停流装置的分光光度计(avivinstruments)或具有搅拌比色杯的8000系列slm-aminco分光光度计(thermospectronic)中测量。
本发明的“结合速率”或“kon”也可以用与上文所述相同的表面等离振子共振技术,使用biacoretm-2000或biacoretm-3000(biacore,inc.,piscataway,nj),用按~10个响应单位(ru)固定抗原的cm5芯片、在25℃测定。简言之,按照厂家的说明,用n-乙基-n’-(3-二甲基氨基丙基)-碳二亚胺盐酸盐(edc)和n-羟基琥珀酰亚胺(nhs)活化羧甲基化葡聚糖生物传感芯片(cm5,biacoreinc.)。用10mm乙酸钠ph4.8将抗原稀释为5μg/ml(~0.2μm),然后按5μl/分钟的流速注入,以达到约10个响应单位(ru)的偶联蛋白质。注入抗原后,注入1m乙醇胺来封闭未反应的基团。对于动力学测量,按约25μl/分钟的流速在25℃在含0.05%tween20的pbs(pbst)中注入fab的两倍系列稀释物(例如0.78nm至500nm)。通过同时拟合结合和解离传感图,用简单的一对一langmuir结合模型(biacoreevaluationsoftwareversion3.2),计算结合速率(kon)和解离速率(koff)。将平衡解离常数(kd)计算为比值koff/kon。见例如chen等,j.mol.biol.293:865-881(1999)。但是,如果通过以上表面等离振子共振测定的结合速率超过106m-1s-1,则优选通过使用荧光淬灭技术来测定结合速率,其中,如在分光光度计,如配有停流装置的分光光度计(avivinstruments)或具有搅拌比色杯的8000系列slm-aminco分光光度计(thermospectronic)中测量,荧光淬灭技术测量在浓度递增的抗原存在下、pbsph7.2中的20nm抗-抗原抗体(fab形式)在25℃的荧光发射强度(激发=295nm;发射=340nm;16nm带通)的增加或减少。
就本发明的多肽,如抗体、片段或其衍生物而言,除非另有说明,“具有生物活性”和“生物活性”和“生物特征”意指具有与生物分子结合的能力。
“肽体”(peptibody)指随机产生的肽与fc结构域的融合物。见2003年12月9日授权给feige等的美国专利号6,660,843(以其整体引入作为参考)。它们包括与n端、c端、氨基酸侧链、或这些位点之一个以上连接的一个或多个肽。肽体技术使得能够设计其中掺入了靶向一种或多种配体或受体的肽、肿瘤归巢肽、膜转运肽等的治疗剂。已证明,肽体技术可以用于设计许多这类分子,包括线性肽和二硫键限制肽、“串联肽多聚体”(即于单链的fc结构域上一个以上肽)。见例如美国专利号6,660,843;2003年10月16日公开的美国专利申请号2003/0195156(对应于2002年11月21日公开的wo02/092620);2003年9月18日公开的美国专利申请号2003/0176352(对应于2003年4月17日公开的wo03/031589);1999年10月22日提交的美国系列号09/422,838(对应于2000年5月4日公开的wo00/24770);2003年12月11日公开的美国专利申请号2003/0229023;2003年7月17日公开的wo03/057134;2003年12月25日公开的美国专利申请号2003/0236193(对应于2004年4月8日提交的pct/us04/010989);2003年9月18日提交的美国系列号10/666,480(对应于2004年4月1日公开的wo04/026329);每篇文献在此以其整体引入作为参考。
“亲和体”(affibody)指,通过肽键与fc区连接的蛋白质的用途,其中该蛋白质用作支架来提供靶分子的结合表面。所述蛋白质通常是天然存在的蛋白质,如葡萄球菌蛋白a或igg结合性b结构域,或从其衍生的z蛋白(见nilsson等(1987),proteng1,107-133和美国专利号5,143,844),或其片段或衍生物。例如,可以从对一种或多种靶分子具有改变的结合亲和力的z蛋白变体产生亲和体,其中为产生能够结合靶分子的变体文库,该z蛋白的一个区段已通过随机诱变而发生突变。亲和体的实例包括美国专利号6,534,628、nordk等,proteng8:601-608(1995)和nordk等,natbiotech15:772-777(1997)、biotechnolapplbiochem.2008年6月;50(pt2):97-112。
“分离的”异源多聚体或复合物意指,已从其天然细胞培养环境的成分中分开和/或回收的异源多聚体或复合物。其天然环境中的污染成分是将干扰异源多聚体的诊断或治疗用途的物质,且可以包括酶、激素和其他蛋白质性或非蛋白质性的溶质。在优选实施方案中,纯化异源多聚体至:(1)通过lowry法测定,蛋白质为大于95%重量,且最优选大于99%重量;(2)足以通过使用旋杯式测序仪获得n端或内部氨基酸序列的至少15个残基的程度;或(3)使用考马斯蓝染色或优选银染,在还原或非还原条件下的sds-page显示的同质性。
本发明的异源多聚体通常纯化至基本同质。短语“基本同质”、“基本上同质的形式”和“基本上同质的”用来指示产物基本上不含源自不希望的多肽组合的副产物(例如同源多聚体)。
用纯度表示的话,基本同质意指,副产物的量不超过10wt%、9wt%、8wt%、7wt%、6wt%、4wt%、3wt%、2wt%或1wt%,或少于1wt%。在一个实施方案中,副产物低于5%。
“生物分子”指核酸、蛋白质、糖、脂质及其组合。在一个实施方案中,生物分子存在于自然界中。
在用来描述本文公开的各种抗体时,“分离的”意指,该抗体已从表达它的细胞或细胞培养物中获得鉴定及分离和/或回收。其天然环境中的污染成分是通常将干扰所述多肽的诊断或治疗用途的物质,且可以包括酶、激素和其他蛋白质性或非蛋白质性的溶质。在优选实施方案中,将抗体纯化至:(1)足以通过使用旋杯式测序仪获得n端或内部氨基酸序列的至少15个残基的程度;或(2)使用考马斯蓝染色或优选银染、通过非还原或还原条件下的sds-page显示的同质。分离的抗体包括在重组细胞内的原位抗体,因为该多肽的天然环境中的至少一种成分不存在。但是,通常,通过至少一个纯化步骤来制备分离的抗体。
本文所用的“连接的”或“连接”意指,第一和第二氨基酸序列之间直接的肽键连接,或涉及第三氨基酸序列的连接,其中所述第三氨基酸序列与第一和第二氨基酸序列通过肽键连接于其之间。例如,与一个氨基酸序列的c端以及与另一个氨基酸序列的n端通过肽键连接的接头。
本文所用的“接头”意指,长度为两个或更多个氨基酸的氨基酸序列。接头可以由中性极性或非极性氨基酸组成。接头可以是例如长度2至100个氨基酸,如长度在2至50个氨基酸之间,例如长度为3、5、10、15、20、25、30、35、40、45或50个氨基酸。接头可以例如通过自切割或酶切割或化学切割来“切割”。氨基酸序列中的切割位点及在这类位点切割的酶和化学品为本领域公知,并还在本文中描述。
本文所用的“系链”意指,连接另外两个氨基酸序列的氨基酸接头。本文所述的系链可以将免疫球蛋白重链可变结构域的n端与免疫球蛋白轻链恒定结构域的c端连接。在具体实施方案中,系链长度在约15至50个氨基酸之间,例如,长度在20至26个氨基酸之间(例如长度为20、21、22、23、24、25或26个氨基酸)。系链可以是“可切割的”,例如,用本领域的方法和试剂标准,自切割或酶切割或化学切割。在某些实施方案中,系链包含gly-gly-ser重复序列。
“接头”或“系链”的酶切割可以涉及内肽酶的使用,例如lys-c、asp-n、arg-c、v8、glu-c、胰凝乳蛋白酶、胰蛋白酶、胃蛋白酶、木瓜蛋白酶、凝血酶、genenase、因子xa、tev(烟草蚀刻病毒半胱氨酸蛋白酶)、肠激酶、hrvc3(人鼻病毒c3蛋白酶)、激肽原酶以及枯草蛋白酶样前蛋白转化酶(例如弗林蛋白酶(pc1)、pc2或pc3)或二碱基n-精氨酸转化酶。化学切割可以涉及例如羟胺、n-氯代琥珀酰亚胺、n-溴代琥珀酰亚胺或溴化氰的使用。
本文所用的“lys-c内肽酶切割位点”是氨基酸序列中可以被lys-c内肽酶在c端侧切割的赖氨酸残基。lys-c内肽酶在赖氨酸残基的c端侧切割。在某些实施方案中,半抗体还在铰链区中包含k222a突变以去除该内源性lys-c内肽酶切割位点,从而在用lys-c内肽酶切割系链后保留半抗体或组装的双特异性抗体的结构。
“铰链区”一般定义为:从人igg1的glu216延伸至pro230(burton,molec.immunol.22:161-206(1985))。可以通过将形成重链间s-s键的第一个和最后一个半胱氨酸残基放置在相同的位置上,将其他igg同种型的铰链区与igg1序列对齐。
fc区的“铰链下区”通常定义为紧接铰链区c端的残基段,即fc区的残基233至239。在本发明之前,fcγr结合通常被归因于iggfc区的铰链下区中的氨基酸残基。
人iggfc区的“ch2结构域”通常从igg的约残基231延伸至约残基340。ch2结构域是独特的,因为它不与另一结构域紧密配对。相反,两个n连接的分枝糖链插在完整的天然igg分子的两个ch2结构域之间。推测该糖可能提供结构域-结构域配对的替代,帮助稳定ch2结构域。burton,molec.immunol.22:161-206(1985)。
“ch3结构域”包含fc区中ch2结构域c端的残基段(即从igg的约氨基酸残基341至约氨基酸残基447)。
本文的术语“fc区”用来定义免疫球蛋白重链的c端区域,包括天然序列fc区和变体fc区。虽然免疫球蛋白重链fc区的分界可以不同,但人igg重链fc区通常被定义为从cys226位或从pro230位的氨基酸残基延伸至其羧基端的区段。fc区的c端赖氨酸(eu编号系统的残基447)可以例如在生产或纯化抗体期间去除,或通过重组改造编码抗体重链的核酸来去除。因此,完整抗体的组合物可以包括所有k447残基均被去除了的抗体群体、未去除k447残基的抗体群体、及包含带k447残基和不带k447残基的抗体的混合物的抗体群体。
“功能性fc区”具有天然序列fc区的“效应子功能”。示例性“效应子功能”包括c1q结合、cdc、fc受体结合、adcc、吞噬作用、细胞表面受体(例如b细胞受体;bcr)的下调等。这类效应子功能一般需要与结合结构域(例如,抗体可变结构域)组合的fc区,且可以用多种测定法例如本文定义中公开的测定法来评估。
“天然序列fc区”包含与见于自然界中的fc区的氨基酸序列相同的氨基酸序列。天然序列人fc区包括天然序列人igg1fc区(非a和a同种异型);天然序列人igg2fc区;天然序列人igg3fc区;天然序列人igg4fc区;及其天然存在的变体。
“变体fc区”包含由于至少一个氨基酸修饰(优选一个或多个氨基酸取代)而不同于天然序列fc区的氨基酸序列的氨基酸序列。优选地,与天然序列fc区或与亲本多肽的fc区相比,变体fc区在天然序列fc区中或在亲本多肽的fc区中具有至少一个氨基酸取代,例如从约一个至约十个氨基酸取代,且优选从约一个至约五个氨基酸取代。本文的变体fc区将优选与天然序列fc区和/或与亲本多肽的fc区具有至少约80%同源性,最优选与之具有至少约90%同源性,更优选与之具有至少约95%同源性。
本文所用的“fc复合物”指相互作用在一起的fc区的两个ch2结构域和/或相互作用在一起的fc区的两个ch3结构域,其中该ch2结构域和/或该ch3结构域通过非肽键的键和/或力(例如范德瓦尔斯力、疏水力、亲水力)相互作用。
本文所用的“fc成分”指fc区的铰链区、ch2结构域或ch3结构域。
本文所用的“fcch成分”或“fcch”指包含fc区的ch2结构域、ch3结构域或ch2和ch3结构域的多肽。
抗体“效应子功能”指可归因于抗体的fc区(天然序列fc区或氨基酸序列变体fc区)的那些生物学活性,其随抗体同种型而变。抗体效应子功能的实例包括:c1q结合和依赖补体的细胞毒性、fc受体结合、依赖抗体的细胞介导的细胞毒性(adcc)、吞噬作用、细胞表面受体(例如b细胞受体)的下调、和b细胞激活。
在本发明中,“还原条件”根据反应中(例如组装混合物中)的氧化还原电位来定义,其指该反应的氧化还原电位为负(-)。还原条件下的反应的氧化还原电位优选在约-50至-600mv、-100至-600mv、-200至-600mv、-100至-500mv、-150至-300mv之间,更优选在约-300至-500mv之间,最优选约-400mv。
可以用任意适宜的方法来制备期望的还原条件。例如,可以通过向反应(如本发明的组装混合物)中加入还原剂来制备期望的还原条件。适宜的还原剂包括但不限于二硫苏糖醇(dtt)、三(2-羧乙基)膦(tcep)、巯基乙酸、抗坏血酸、硫羟乙酸、谷胱甘肽(gsh)、β-巯基乙胺、半胱氨酸/胱氨酸、gsh/谷胱甘肽二硫化物(gssg)、半胱胺/胱胺、甘氨酰半胱氨酸、和β-巯基乙醇,优选gsh。在某些具体实施方案中,还原剂是弱还原剂,包括但不限于gsh、β-巯基乙胺、半胱氨酸/胱氨酸、gsh/gssg、半胱胺/胱胺、甘氨酰半胱氨酸和β-巯基乙醇,且优选gsh。在某些优选实施方案中,还原剂是gsh。本领域普通技术人员能够选择在适宜浓度下和适宜实验条件下的适宜还原剂以在反应中达到期望的还原条件。例如,在20℃,在具有10g/l双特异性抗体蛋白质浓度的溶液中10mml-还原型谷胱甘肽将产生约-400mv的起始氧化还原电位。本领域技术人员可以用任意适宜的方法来测量给定反应中的氧化还原电位。
可以用本领域已知的任意适宜方法,估计和测量反应的还原条件。例如,可以用刃天青指示剂(在还原条件下从蓝色脱色至无色)测量还原条件。对于更精确的测量,可以使用氧化还原电位计(如broadley
备选地,可以通过在约10mmhg或更小、优选约5mmhg或更小、更优选约3mmhg或更小的减小压力下去除溶解的气体、尤其是溶解的氧,约1至60分钟、优选约5至40分钟,以制备还原条件。
在本发明中,优选从一经混合该第一和第二含铰链多肽(如半抗体)后至组装步骤都保持还原条件。在某些实施方案中,优选在约50%或更长、更优选约70%或更长、还更优选约90%或更长的反应时间,保持反应或组装混合物在还原条件中。尤其优选,在约50%或更多、更优选约70%或更多、还更优选约90%或更多的反应时间内,将反应介质的氧化还原电位保持在约-200至-600mv、更优选-300至-500mv之间、最优选约-400mv。
在某些具体实施方案中,还原条件是弱还原条件。本文所用的术语“弱还原剂”或“弱还原条件”指,在25℃具有负氧化电位的还原剂或由该还原剂制备的还原条件。在ph为7至9、温度在15℃和39℃之间时,还原剂的氧化电位优选在-50至-600mv、-100至-600mv、-200至-600mv、-100至-500mv、-150至-300mv之间、更优选约-300至-500mv之间、最优选-400mv。本领域技术人员将能够选择适宜的还原剂来制备希望的还原条件。本领域技术人员明了,可以以较低的浓度使用强还原剂,即在相同的浓度、ph和温度下比上述还原剂具有更负的氧化电位的还原剂。在优选实施方案中,在上述条件下孵育时,蛋白质将能够在还原剂的存在下形成二硫键。弱还原剂的实例包括但不限于谷胱甘肽、β-巯基乙胺、半胱氨酸/胱氨酸、gsh/gssg、半胱胺/胱胺、甘氨酰半胱氨酸和β-巯基乙醇。在某些实施方案中,可以用与200x摩尔比的gsh:抗体的氧化电位类似的氧化电位作为弱还原条件的参考点,在该点,预期可用其他还原剂实现有效组装。
“组装混合物”是包含第一含铰链多肽、第二含铰链多肽的溶液。在某些实施方案中,组装混合物存在于还原条件中。在一些实施方案中,组装混合物存在于弱还原条件中。在某些其他实施方案中,组装混合物进一步包含弱还原剂。ph在7和9之间、温度在15℃和39℃之间时,组装混合物的氧化电位为-50至-600mv、-100至-600mv、-200至-600mv、-100至-500mv、-150至-300mv之间、更优选在约-300至-500mv之间、最优选约-400mv。
除非另有说明,实施例中提到的市售试剂按照厂家的说明使用。以下实施例和说明书通篇中用atcc保藏号标识的那些细胞的来源是美国典型培养物保藏中心(americantypeculturecollection,manassas,va)。除非另有说明,本发明使用重组dna技术的标准方法,如上文及以下教科书中所述的那些:sambrook等,上引文;ausubel等,currentprotocolsinmolecularbiology(greenpublishingassociatesandwileyinterscience,ny,1989);innis等,pcrprotocols:aguidetomethodsandapplications(academicpress,inc.,ny,1990);harlow等,antibodies:alaboratorymanual(coldspringharborpress,coldspringharbor,1988);gait,oligonucleotidesynthesis(irlpress,oxford,1984);freshney,animalcellculture,1987;coligan等,currentprotocolsinimmunology,1991。
在本说明书和权利要求通篇中,词语“包含”或其变化形式将理解为暗示包括所述整数或整数组,但不排除任何其他整数或整数组。
ii.异源多聚体蛋白质的构建
典型地,本文所述的异源多聚体蛋白质将包含抗体fc区的显著部分。但是,在其他方面,重链仅包含ch1、ch2和/或ch3结构域的部分。
异源多聚化结构域
异源多聚体蛋白质包含异源多聚化结构域。为了产生异二聚体分子的基本上同质的群体,异源二聚化结构域必须具有形成异二聚体超过同二聚体的强烈倾向。虽然本文中示例的异源多聚体蛋白质用杵入臼技术以便于异源多聚化,但本领域技术人员应理解,其他异源多聚化结构域也可用于本发明。
杵入臼
杵入臼作为产生多特异性抗体的方法的用途为本领域公知。见1998年3月24日授权的、转让给genentech的美国专利号5,731,168;2009年7月16日公开、转让给amgen的pct公开号wo2009089004;及2009年7月16日公开、转让给novonordiska/s的美国专利公开号20090182127。还见marvin和zhu,actapharmacologicasincia(2005)26(6):649-658及kontermann(2005)actapharacol.sin.,26:1-9。本文提供简要的讨论。
“凸起”指至少一个氨基酸侧链,其从第一多肽的界面凸出来,因此可置入相邻界面(即第二多肽的界面)中的互补凹陷内来稳定异源多聚体,从而例如相比同源多聚体形成,更利于异源多聚体形成。凸起可以存在于原始界面上或可以经合成引入(例如通过改变编码界面的核酸)。通常,改变编码第一多肽的界面的核酸来编码凸起。为此,将编码第一多肽界面中的至少一个“原始”氨基酸残基的核酸替代为编码至少一个“输入”氨基酸残基的核酸,其中输入氨基酸残基具有比原始氨基酸残基大的侧链体积。应理解,可以存在一个以上原始残基和对应的输入残基。取代的原始残基的数目的上限是第一多肽界面中的残基总数。下表中显示各种氨基残基的侧链体积。
表1
氨基酸残基的特性
a氨基酸分子量减去水的分子量。来自handbookofchemistryandphysics,第43版cleveland,chemicalrubberpublishingco.,1961的值。
b来自a.a.zamyatnin,prog.biophys.mol.biol.24:107-123,1972的值。
c来自c.chothia,j.mol.biol.105:1-14,1975的值。可及表面积在此参考文献的图6-20中定义。
优选的用于形成凸起的输入残基一般是天然存在的氨基酸残基,且优选选自精氨酸(r)、苯丙氨酸(f)、酪氨酸(y)和色氨酸(w)。最优选的是色氨酸和酪氨酸。在一个实施方案中,对于凸起的形成,原始残基具有小的侧链体积,如丙氨酸、天冬酰胺、天冬氨酸、甘氨酸、丝氨酸、苏氨酸或缬氨酸。ch3结构域中用于形成凸起的示例性氨基酸取代包括但不限于t366w取代。
“凹陷”指至少一个氨基酸侧链,其从第二多肽的界面凹入,从而可以容纳相邻的第一多肽界面上的对应凸起。凹陷可以存在于原始界面中,或者可以通过合成引入(例如通过改变编码界面的核酸)。通常,改变编码第二多肽界面的核酸来编码凹陷。为此,用编码至少一个“输入”氨基酸残基的dna取代编码第二多肽界面中的至少一个“原始”氨基酸残基的核酸,其中输入氨基酸残基具有比原始氨基酸残基小的侧链体积。应理解,可以存在一个以上原始残基和对应的输入残基。取代的原始残基的数目的上限是第二多肽界面中的残基的总数。上文表1中显示多种氨基残基的侧链体积。优选的用于形成凹陷的输入残基通常是天然存在的氨基酸残基,且优选选自丙氨酸(a)、丝氨酸(s)、苏氨酸(t)和缬氨酸(v)。最优选的是丝氨酸、丙氨酸或苏氨酸。在一个实施方案中,对于形成凹陷,原始残基具有大的侧链体积,如酪氨酸、精氨酸、苯丙氨酸或色氨酸。ch3结构域中用于产生凹陷的示例性氨基酸取代包括但不限于,t366s、l368a、y407a、y407t和y407v取代。在某些实施方案中,杵半抗体包含t366w取代,而臼半抗体包含t366s/l368a/y407v取代。
“原始”氨基酸残基是被“输入”残基取代的氨基酸残基,输入残基可以具有比原始残基小或大的侧链体积。输入氨基酸残基可以是天然存在的或非天然存在的氨基酸残基,但优选前者。“天然存在的”氨基酸残基是由遗传密码编码并在上文表1中列出的那些残基。“非天然存在的”氨基酸残基意指这样的残基,其不由遗传密码编码,但其能够在多肽链中共价结合邻近的一个或多个氨基酸残基。非天然存在的氨基酸残基的实例是正亮氨酸、鸟氨酸、正缬氨酸、高丝氨酸及其他氨基酸残基类似物,如描述于例如ellman等,meth.enzym.202:301-336(1991)中的那些。为了产生这类非天然存在的氨基酸残基,可以使用noren等science244:182(1989)和ellman等,同上引文的方法。简言之,这涉及用非天然存在的氨基酸残基化学活化阻抑型trna,然后进行rna的体外转录和翻译。本发明的方法涉及取代至少一个原始残基,但可以取代一个以上的原始残基。通常,取代的原始氨基酸残基将不超过第一或第二多肽的界面中的总残基。典型地,取代的原始残基是“埋藏的”。“埋藏的”意指溶剂基本上不可接近该残基。通常,输入残基不是半胱氨酸,以防止可能的氧化或二硫键的错配。
凸起“可置入”凹陷中,意指凸起和凹陷各自在第一多肽和第二多肽界面上的空间位置及凸起和凹陷的大小,使得凸起可以置入凹陷中而不显著扰乱第一和第二多肽在界面上的正常结合。由于诸如tyr、phe和trp的凸起通常不从界面的轴垂直伸出且具有优选的构象,故凸起与对应的凹陷的对齐依赖于根据三维结构(如通过x射线晶体分析法或核磁共振(nmr)获得的三维结构)模拟凸起/凹陷对。这可以用本领域广泛接受的技术来达到。
“原始或模板核酸”意指编码目的多肽的核酸,其可以被“改变”(即遗传改造或突变)以编码凸起或凹陷。原始或起始核酸可以是天然存在的核酸,或者可以包含之前已进行了改变的核酸(例如人源化抗体片段)。“改变”核酸意指,通过插入、缺失或取代至少一个编码目的氨基酸残基的密码子来突变原始核酸。通常,用编码输入残基的密码子替代编码原始残基的密码子。用于以这种方式遗传修饰dna的技术综述于mutagenesis:apracticalapproach,m.j.mcpherson编辑,(irlpress,oxford,uk.(1991)中,且包括例如位点定向诱变、盒诱变和聚合酶链反应(pcr)诱变。通过突变原始/模板核酸,从而相应地改变原始/模板核酸编码的原始/模板多肽。
可以通过合成手段,例如通过重组技术、体外肽合成、之前描述的用于引入非天然存在的氨基酸残基的那些技术,通过肽的酶偶联或化学偶联,或这些技术的一些组合,将凸起或凹陷“引入”第一或第二多肽的界面。因此,“引入的”凸起或凹陷是“非天然存在的”或“非天然的”,意指它不存在于自然界中或原始多肽(例如人源化单克隆抗体)中。
通常,用于形成凸起的输入氨基酸残基具有相对小数目的“旋转异构体”(例如约3-6)。“旋转异构体”是氨基酸侧链在能量上有利的构象。多种氨基酸残基的旋转异构体的数目综述于ponders和richards,j.mol.biol.193:775-791(1987)中。
iii.载体、宿主细胞和重组方法
为了重组产生本发明的异源多聚体蛋白质(例如双特异性抗体),分离编码它的核酸,并插入可复制载体用于进一步克隆(dna的扩增)或用于表达。编码抗体的dna易于用常规方法(例如通过使用能够特异性结合编码抗体重链和轻链的基因的寡核苷酸探针)分离和测序。许多载体可用。载体的选择部分取决于要使用的宿主细胞。通常,优选的宿主细胞为原核生物或真核生物(通常为哺乳动物,但也包括真菌(例如酵母)、昆虫、植物和来自其他多细胞生物的有核细胞)来源。应理解,任何同种型的恒定区都可以用于此目的,包括igg、igm、iga、igd和ige恒定区,且这些恒定区可以获自任何人或动物物种。在某些实施方案中,恒定区来自igg,尤其是igg1、igg2或igg4。
改造宿主细胞,使得它表达包含异源二聚化结构域的含铰链多肽,其中宿主细胞不表达包含第二异源二聚化结构域的含铰链多肽。
a.用原核宿主细胞生产异源多聚体蛋白质
i.载体构建
编码本发明的异源多聚体蛋白质(例如抗体)的多肽成分的多核苷酸序列可以用标准重组技术获得。可以从例如产生抗体的细胞(如杂交瘤细胞)分离和测序希望的多核苷酸序列。备选地,可以用核苷酸合成仪或pcr技术合成多核苷酸。一旦获得,即可以将编码多肽的序列插入能够在原核宿主中复制和表达异源多核苷酸的重组载体。本领域可得和已知的许多载体可以用于本发明的目的。适宜载体的选择将主要取决于待插入载体的核酸的大小及待用载体转化的具体宿主细胞。取决于载体的功能(扩增或表达异源多核苷酸,或二者)和它与它所驻留的具体宿主细胞的相容性,每个载体包含多种成分。载体成分通常包括但不限于:复制起点、选择标记基因、启动子、核糖体结合位点(rbs)、信号序列、异源核酸插入片段和转录终止序列。
通常,将包含源自与宿主细胞相容的物种的复制子和控制序列的质粒载体与这些宿主结合使用。载体通常携带复制位点、以及能够在转化细胞中提供表型选择的标记序列。例如,通常用衍生自大肠杆菌物种的质粒pbr322转化大肠杆菌。pbr322包含编码氨苄青霉素(amp)和四环素(tet)抗性的基因,从而提供用于鉴定转化细胞的容易手段。pbr322、其衍生物、或其他微生物质粒或噬菌体也可以包含,或经修饰以包含,能够被微生物用于表达内源蛋白质的启动子。可以用于表达具体抗体的pbr322衍生物的实例详细描述于carter等的美国专利号5,648,237中。
此外,含有与宿主生物相容的复制子和控制序列的噬菌体载体可以用作转化载体,与这些宿主组合。例如,诸如λgem.tm.-11的噬菌体可以用于制备能够用来转化诸如大肠杆菌le392的易感宿主细胞的重组载体。
本发明的表达载体可以包含两个或更多个启动子-顺反子对,分别编码各多肽成分。启动子是位于顺反子上游(5’)的非翻译调节序列,调节顺反子的表达。原核启动子典型地分为两类:诱导型和组成型。诱导型启动子是响应培养条件的变化(例如营养物的存在或缺乏或温度的变化)而起始处于其控制下的顺反子的提高水平的转录的启动子。
多种潜在的宿主细胞识别的大量启动子为本领域公知。可以通过限制酶消化从来源dna移出启动子并将该分离的启动子插入本发明的载体,将所选择的启动子有效连接至编码例如轻链或重链的顺反子dna。天然启动子序列和许多异源启动子都可以用来指导靶基因的扩增和/或表达。在一些实施方案中,利用异源启动子,因为与天然靶多肽启动子相比,它们通常允许所表达的靶基因获得更大的转录和更高的产率。
适合用于原核宿主的启动子包括phoa启动子、β-galactamase和乳糖启动子系统、色氨酸(trp)启动子系统和杂合启动子,如tac或trc启动子。但是,在细菌中具有功能的其他启动子(如其他已知的细菌或噬菌体启动子)也是适宜的。它们的核苷酸序列已公开,从而使得技术人员能够用接头或衔接子提供所需的任何限制位点,将它们有效连接至编码异源多聚体蛋白质(例如靶轻链和重链)的基因的顺反子(siebenlist等,(1980)cell20:269)。
在本发明的一个方面,重组载体内的每个顺反子都包含指导所表达的多肽跨膜转运的分泌信号序列成分。通常,该信号序列可以是载体的成分,或者它可以是插入载体的靶多肽dna的部分。选择用于本发明的目的的信号序列应是可被宿主细胞识别和加工(即通过信号肽酶切割)的信号序列。对于不识别和加工异源多肽的天然信号序列的原核宿主细胞,用选自例如碱性磷酸酶、青霉素酶、lpp或热稳定肠毒素ii(stii)前导序列、lamb、phoe、pelb、ompa和mbp的原核信号序列取代该天然信号序列。在本发明的一个实施方案中,用于表达系统的两个顺反子中的信号序列是stii信号序列或其变体。
在另一方面,本发明的免疫球蛋白的产生可以发生在宿主细胞的胞质中,因此无需在每个顺反子内存在分泌信号序列。在这方面,免疫球蛋白轻链和重链在胞质内表达、折叠并组装形成功能性免疫球蛋白。某些宿主菌株(例如大肠杆菌trxb-菌株)提供有利于二硫键形成的胞质条件,从而允许所表达的蛋白质亚基的正确折叠和组装。见proba和pluckthungene,159:203(1995)。
适合用于表达本发明的异源多聚体蛋白质(例如抗体)的原核宿主细胞包括古细菌(archaebacteria)和真细菌(eubacteria),如革兰氏阴性或革兰氏阳性生物。有用的细菌的实例包括埃希氏菌属(escherichia)(例如大肠杆菌)、芽孢杆菌属(bacilli)(例如枯草芽孢杆菌(b.subtilis))、肠细菌、假单胞菌属物种(pseudomonasspecies)(例如铜绿假单胞菌(p.aeruginosa))、鼠伤寒沙门氏菌(salmonellatyphimurium)、粘质沙雷氏菌(serratiamarcescans)、克雷伯氏菌属(klebsiella)、变形杆菌属(proteus)、志贺氏菌属(shigella)、根瘤菌属(rhizobia)、透明颤菌属(vitreoscilla)或副球菌属(paracoccus)。在一个实施方案中,使用革兰氏阴性细胞。在一个实施方案中,用大肠杆菌作为本发明的宿主。大肠杆菌菌株的实例包括菌株w3110(bachmann,cellularandmolecularbiology,第2卷(washington,d.c.:americansocietyformicrobiology,1987),1190-1219页;atcc保藏号27,325)及其衍生菌株,包括具有基因型w3110δfhua(δtona)ptr3laciqlacl8δomptδ(nmpc-fepe)degp41kanr的菌株33d3(美国专利号5,639,635)。其他菌株及其衍生菌株,诸如大肠杆菌294(atcc31,446)、大肠杆菌b、大肠杆菌λ1776(atcc31,537)和大肠杆菌rv308(atcc31,608)也是适宜的。在一个实施方案中,大肠杆菌δlpp尤其有用。这些实例是举例说明性的而不是限制性的。用于构建任意上述细菌的具有确定基因型的衍生菌株的方法为本领域已知,并描述于例如bass等,proteins,8:309-314(1990)中。通常有必要考虑复制子在细菌细胞中的可复制性来选择适当的细菌。例如,在用众所周知的质粒诸如pbr322、pbr325、pacyc177或pkn410来提供复制子时,可以适宜地用大肠杆菌、沙雷氏菌属(serratia)或沙门氏菌属(salmonella)物种作为宿主。通常,宿主细胞应分泌最小量的蛋白水解酶,且可能期望将额外的蛋白酶抑制剂掺入细胞培养物中。
ii.多肽生产
用上述表达载体转化宿主细胞,并培养在根据诱导启动子、选择转化体或扩增编码期望序列的基因的需要改进的常规营养培养基中。
转化意指将dna引入原核宿主,使得该dna可作为染色体外元件或通过染色体整合而复制。取决于所使用的宿主细胞,用适于该细胞的标准技术进行转化。利用氯化钙的钙处理通常用于含有实质性细胞壁屏障的细菌细胞。另一转化方法利用聚乙二醇/dmso。另一常用的技术是电穿孔。
将用来产生本发明多肽的原核细胞培养在本领域已知且适合用于培养所选择的宿主细胞的培养基中。适宜的培养基的实例包括加入了必要的营养补充物的luria培养基(lb)。在一些实施方案中,培养基还包含根据表达载体的构建而选择的选择剂,以选择性地允许含有该表达载体的原核细胞生长。例如,向培养基中加入氨苄青霉素,用于生长表达氨苄青霉素抗性基因的细胞。
除碳、氮和无机磷酸来源外,还可以包括适当浓度的任何必要的补充物,其可以单独引入或作为与另一补充物或培养基(如复合氮源)的混合物引入。可选地,培养基可以包含选自谷胱甘肽、半胱氨酸、胱胺、巯基乙酸、二硫赤藓糖醇和二硫苏糖醇的一种或多种还原剂。
在适宜的温度下培养原核宿主细胞。对于大肠杆菌生长,例如,优选的温度范围从约20℃至约39℃,更优选从约25℃至约37℃,甚至更优选在约30℃。主要取决于宿主生物,培养基的ph可以是从约5至约9的任何ph。对于大肠杆菌,ph优选从约6.8至约7.4,更优选约7.0。
如果在本发明的表达载体中使用诱导型启动子,则在适用于激活该启动子的条件下诱导蛋白质表达。在本发明的一个方面,用phoa启动子来控制多肽的转录。因此,将转化的宿主细胞培养在磷酸限制培养基中进行诱导。优选地,该磷酸限制培养基是c.r.a.p培养基(见例如simmons等,j.immunol.methods(2002),263:133-147)。如本领域已知,根据所利用的载体构建体,可以使用多种其他诱导物。
在一个实施方案中,分别培养第一和第二含铰链多肽的宿主细胞,所表达的本发明多肽分泌进入宿主细胞的周质,并分别从宿主细胞的周质回收。在第二实施方案中,分别培养第一和第二含铰链多肽的宿主细胞,在分离含铰链多肽之前,将两种宿主细胞培养物混合在一起并沉淀细胞。在第三实施方案中,分别培养第一和第二含铰链多肽的宿主细胞,分别离心和重悬,然后在分离含铰链多肽之前混合在一起。在第四实施方案中,在同一培养容器中一起培养第一和第二含铰链多肽的宿主细胞。蛋白质回收通常涉及破裂微生物细胞膜,一般通过诸如渗透压休克、超声处理或裂解的手段。一旦破裂细胞,即可以通过离心或过滤来去除细胞碎片或完整细胞。可以例如通过亲和树脂层析,进一步纯化蛋白质。备选地,蛋白质可以转运或分泌入培养基,并从其中分离。带有外源序列标签(或表位标签)表达的重组蛋白质可以便于纯化步骤。克隆和纯化含有外源序列标签(包括但不限于his标签和gst标签)的蛋白质的技术为本领域公知。可以从培养物去除细胞,过滤并浓缩培养物上清用于进一步纯化所产生的蛋白质。可以用诸如聚丙烯酰胺凝胶电泳(page)和western印迹测定等公知方法,进一步分离和鉴定所表达的多肽。分离的多肽用来产生异源多聚体蛋白质。
在本发明的一个方面中,通过发酵方法,大量生产异源多聚体蛋白质(例如抗体)。多种大规模补料分批发酵方法可用于生产重组蛋白质。大规模发酵具有至少1000升的容量,优选约1,000至100,000升的容量。这些发酵罐用搅拌桨来分布氧气和营养物,尤其是葡萄糖(优选的碳/能量来源)。小规模发酵一般指体积容量不超过约100升的发酵罐中的发酵,该发酵罐可以在约1升至约100升的范围内。
在发酵方法中,通常将细胞在适宜条件下培养至希望的密度(例如约180-220的od550,细胞在该阶段处于早期平台期)后,起始蛋白质表达的诱导。如本领域已知和本文所述,根据所利用的载体构建体,可以使用多种诱导物。细胞可以在诱导前培养更短的时期。通常诱导细胞约12-50小时,但也可以使用更长或更短的诱导时间。
为了改善本发明多肽的产率和质量,可以修饰多种发酵条件。例如,为了改善分泌性异源多聚体蛋白质(例如抗体)的正确组装和折叠,可以用过表达诸如dsb蛋白质(dsba、dsbb、dsbc、dsbd和/或dsbg)或fkpa(具有分子伴侣活性的肽酰脯氨酰顺反异构酶)的分子伴侣蛋白质的额外载体来共转化宿主原核细胞。已证明分子伴侣蛋白质有利于细菌宿主细胞中产生的异源蛋白质的正确折叠和溶解。chen等(1999)jbiochem274:19601-19605;georgiou等,美国专利号6,083,715;georgiou等,美国专利号6,027,888;bothmann和pluckthun(2000)j.biol.chem.275:17100-17105;ramm和pluckthun(2000)j.biol.chem.275:17106-17113;arie等(2001)mol.microbiol.39:199-210。
为了最小化所表达的异源蛋白质(尤其是对蛋白酶解敏感的那些蛋白质)的蛋白酶解,可以将蛋白水解酶缺陷的某些宿主菌株用于本发明。例如,可以修饰宿主细胞菌株,在编码已知的细菌蛋白酶(如蛋白酶iii、ompt、degp、tsp、蛋白酶i、蛋白酶mi、蛋白酶v、蛋白酶vi及其组合)的基因中产生一个或多个遗传突变。一些大肠杆菌蛋白酶缺陷菌株可得,并描述于例如,joly等(1998)proc.natl.acad.sci.usa95:2773-2777;georgiou等的美国专利号5,264,365;georgiou等,美国专利号5,508,192;hara等,microbialdrugresistance,2:63-72(1996)中。
在一个实施方案中,将蛋白水解酶缺陷并用过表达一种或多种分子伴侣蛋白质的质粒转化的大肠杆菌菌株用作本发明的表达系统中的宿主细胞。在第二实施方案中,大肠杆菌菌株为外膜脂蛋白缺陷(δlpp)。
iii.异源多聚体蛋白质纯化
在一个实施方案中,进一步纯化本文中产生的异源多聚体蛋白质,以获得基本上同质的制品用于其他测定和用途。可以利用本领域已知的标准蛋白质纯化方法。以下方法是适宜的纯化方法的示例:在免疫亲和或离子交换柱上分级分离、乙醇沉淀、反相hplc、硅石上或诸如deae的离子交换树脂上的层析、层析聚焦、sds-page、硫酸铵沉淀和使用例如sephadexg-75的凝胶过滤。
在一方面,用固定在固相上的蛋白a来进行例如本发明的半抗体或全长抗体产物的免疫亲和纯化。蛋白a是以高亲和力结合抗体fc区的来自金黄色葡萄球菌(staphylococcusaureus)的41kd细胞壁蛋白质。lindmark等(1983)j.immunol.meth.62:1-13。固定蛋白a的固相优选是含有玻璃或硅石表面的柱,更优选可控孔度玻璃柱或硅酸柱。在一些应用中,已用诸如甘油的试剂包被柱,以图防止污染物的非特异性附着。
作为纯化的第一步,将源自上述细胞培养物的制备物上样至固定了蛋白a的固相上,以允许目的抗体与蛋白a特异性结合。然后洗涤固相,去除非特异性结合于固相的污染物。通过洗脱,从固相回收异源多聚体蛋白质(例如抗体)。
b.用真核宿主细胞生产异源多聚体蛋白质
载体成分通常包括但不限于以下的一个或多个:信号序列、复制起点、一个或多个标记基因、增强子元件、启动子和转录终止序列。
i.信号序列成分
用于真核宿主细胞的载体也可以包含在成熟蛋白质或目的多肽的n端具有特异性切割位点的信号序列或其他多肽。所选择的异源信号序列优选是宿主细胞识别和加工(即通过信号肽酶切割)的信号序列。在哺乳动物细胞表达中,可用哺乳动物信号序列、以及病毒分泌前导序列,例如单纯疱疹gd信号。将该前体区的dna符合读框地连接至编码一种或多种希望的异源多聚体蛋白质(例如抗体)的dna。
ii.复制起点
通常,哺乳动物表达载体无需复制起点成分。例如,sv40起点可以典型地被使用,但仅是因为它包含早期启动子。
iii.选择基因成分
表达载体和克隆载体可以包含选择基因,也称为选择标记。典型的选择基因编码这样的蛋白质,该蛋白质(a)赋予对抗生素或其他毒素(例如氨苄青霉素、新霉素、氨甲喋呤或四环素)的抗性;(b)相关时,弥补营养缺陷;或(c)提供不能从复杂培养基获得的关键营养物。
选择方案的一个实例利用药物来阻滞宿主细胞的生长。用异源基因成功转化的那些细胞产生赋予药物抗性的蛋白质,从而在该选择方案下存活。这种显性选择的实例使用药物新霉素、霉酚酸和潮霉素。
适用于哺乳动物细胞的选择标记的另一实例是使得能够鉴定有能力摄取抗体核酸的细胞的那些选择标记,如dhfr、胸苷激酶、金属硫蛋白-i和-ii(优选灵长类金属硫蛋白基因)、腺苷脱氨酶、鸟氨酸脱羧酶等。
例如,首先通过在包含氨甲喋呤(mtx)(dhfr的竞争性拮抗剂)的培养基中培养所有转化体,鉴定转化了dhfr选择基因的细胞。在利用野生型dhfr时,适当的宿主细胞是缺乏dhfr活性的中国仓鼠卵巢(cho)细胞系(例如atcccrl-9096)。
备选地,用编码抗体、野生型dhfr蛋白质和另一选择标记(如氨基糖苷3’-磷酸转移酶(aph))的dna序列转化或共转化的宿主细胞(尤其是包含内源dhfr的野生型宿主),可以通过在包含用于该选择标记的选择剂(如氨基糖苷抗生素,例如卡那霉素、新霉素或g418)的培养基中培养细胞而选择。见例如美国专利号4,965,199。
iv.启动子成分
表达载体和克隆载体通常包含宿主生物可识别的启动子,其与一个或多个期望的含铰链多肽(例如抗体)核酸有效连接。用于真核生物的启动子序列是已知的。基本上所有真核基因都在距离转录起始位点上游约25至30个碱基的位置具有富含at的区域。见于许多基因的转录起点上游70至80个碱基的另一序列是cncaat区,其中n可以是任意核苷酸。大多数真核基因的3’端是aataaa序列,其可以是将polya尾加至编码序列的3’端的信号。所有这些序列都适宜插入真核表达载体。
可以例如通过获自病毒(例如多瘤病毒、禽痘病毒、腺病毒(如腺病毒2)、牛乳头状病毒、禽肉瘤病毒、巨细胞病毒、反转录病毒、乙型肝炎病毒和猿猴病毒40(sv40))基因组、获自异源哺乳动物启动子(例如肌动蛋白启动子或免疫球蛋白启动子)、或获自热休克启动子的启动子,控制哺乳动物宿主细胞中希望的一种或多种含铰链多肽(例如半抗体)从载体的转录,条件是启动子与宿主细胞系统相容。
sv40病毒的早期和晚期启动子可以作为sv40限制片段方便地获得,该限制片段还包含sv40病毒复制起点。人巨细胞病毒的立即早期启动子可以作为hindiiie限制片段方便地获得。美国专利号4,419,446中公开了用牛乳头状病毒作为载体在哺乳动物宿主中表达dna的系统。美国专利号4,601,978中描述了对此系统的修改。还见reyes等,nature297:598-601(1982),其涉及人β-干扰素cdna在来自单纯疱疹病毒的胸苷激酶启动子的控制下在小鼠细胞中表达。备选地,可以用劳斯肉瘤病毒长末端重复序列作为启动子。
v.增强子元件成分
将增强子序列插入载体,可以增加高等真核生物对编码希望的一种或多种含铰链多肽(例如抗体)的dna的转录。现已知来自哺乳动物基因(例如珠蛋白、弹性蛋白酶、清蛋白、甲胎蛋白和胰岛素基因)的许多增强子序列。另外,还可以使用来自真核细胞病毒的增强子。实例包括位于复制起点晚期一侧(bp100-270)的sv40增强子、巨细胞病毒早期启动子增强子、复制起点晚期一侧的多瘤增强子、和腺病毒增强子。关于用于增强真核启动子的激活的元件的描述,还见yaniv,nature297:17-18(1982)。只要可以实现增加作用,增强子可以拼接在载体中抗体多肽编码序列5’或3’的位置,但通常在启动子5’的位置。
vi.转录终止成分
用于真核宿主细胞的表达载体通常还将包含终止转录和稳定mrna所必需的序列。通常可从真核或病毒dna或cdna的5’和偶尔地3’非翻译区获得这类序列。这些区域包含在编码抗体的mrna的非翻译部分中转录为的聚腺苷酸化片段的核苷酸区段。一种有用的转录终止成分是牛生长激素聚腺苷酸化区域。见wo94/11026及其中公开的表达载体。
vii.宿主细胞的选择和转化
适合用于克隆或表达本文载体中的dna的宿主细胞包括本文所述的高等真核细胞,包括脊椎动物宿主细胞。在培养物(组织培养物)中繁殖脊椎动物细胞已成为常规方法。有用的哺乳动物宿主细胞系的实例是sv40转化的猴肾cv1细胞系(cos-7,atcccrl1651);人胚肾细胞系(293或为在悬浮培养物中生长而亚克隆的293细胞,graham等,j.genvirol.36:59(1977));幼仓鼠肾细胞(bhk,atccccl10);中国仓鼠卵巢细胞/-dhfr(cho,urlaub等,proc.natl.acad.sci.usa77:4216(1980));小鼠支持细胞(tm4,mather,biol.reprod.23:243-251(1980));猴肾细胞(cvlatccccl70);非洲绿猴肾细胞(vero-76,atcccrl-1587);人宫颈癌细胞(hela,atccccl2);犬肾细胞(mdck,atccccl34);buffalo大鼠肝细胞(brl3a,atcccrl1442);人肺细胞(w138,atccccl75);人肝细胞(hepg2,hb8065);小鼠乳腺肿瘤(mmt060562,atccccl51);tri细胞(mather等,annalsn.y.acad.sci.383:44-68(1982));mrc5细胞;fs4细胞;和人肝癌细胞系(hepg2)。
为生产期望的含铰链多肽(例如抗体,一种或多种),用上述表达或克隆载体转化宿主细胞,并在根据诱导启动子、选择转化体或扩增期望序列编码基因的需要改进的常规营养培养基中培养。
viii.培养宿主细胞
可以在多种培养基中培养用于产生期望的本发明一种或多种含铰链多肽(例如抗体)的宿主细胞。诸如ham'sf10(sigma)、minimalessentialmedium(mem)(sigma)、rpmi-1640(sigma)和dulbecco'smodifiedeagle'smedium(dmem,sigma)等市售培养基适用于培养宿主细胞。此外,可以将ham等,meth.enz.58:44(1979);barnes等,anal.biochem.102:255(1980);美国专利号4,767,704;4,657,866;4,927,762;4,560,655;或5,122,469;wo90/03430;wo87/00195;或美国专利再版30,985中所述的任何培养基用作宿主细胞的培养基。可以根据需要向任意这些培养基中补充激素和/或其他生长因子(如胰岛素、转铁蛋白或表皮生长因子)、盐(如氯化钠、钙、镁和磷酸盐)、缓冲液(如hepes)、核苷酸(如腺苷和胸苷)、抗生素(如gentamycintm药物)、痕量元素(定义为通常以微摩尔范围的终浓度存在的无机化合物)和葡萄糖或等同的能量来源。还可以按适当浓度包含本领域技术人员已知的任何其他必需的补充物。诸如温度、ph等培养条件可以是选择用于表达的宿主细胞之前所用的那些条件,且对普通技术人员而言显而易见。
ix.异源多聚体蛋白质的纯化
在使用重组技术时,含铰链多肽可以在胞内产生,或者直接分泌进入培养基。如果含铰链多肽在胞内产生,作为第一步,例如通过离心或超滤,去除颗粒碎片(宿主细胞或裂解片段)。在含铰链多肽分泌进入培养基时,通常首先用市售蛋白质浓缩滤器(例如amicon或milliporepellicon超滤单元)浓缩来自这类表达系统的上清。可以在任意前述步骤中包含蛋白酶抑制剂(如pmsf)来抑制蛋白酶解,且可以包含抗生素来防止外来污染物的生长。
可以用例如离子交换、疏水相互作用层析、羟基磷灰石层析、凝胶电泳、透析和亲和层析来纯化从细胞制备的异源多聚体组合物,亲和层析是优选的纯化技术。还考虑上述技术的组合。蛋白a作为亲和配体的适合性取决于抗体中存在的任何免疫球蛋白fc结构域的种类和同种型。可以用a蛋白来纯化基于人γ1、γ2或γ4重链的抗体(lindmark等,j.immunol.meth.62:1-13(1983))。建议g蛋白用于所有小鼠同种型和人γ3(guss等,emboj.5:15671575(1986))。亲和配体所附着的基质最常是琼脂糖,但其他基质也可用。与用琼脂糖可以达到的流速和处理时间相比,机械稳定的基质(如可控孔度玻璃或聚(苯乙烯二乙烯)苯)允许更快的流速和更短的处理时间。在抗体包含ch3结构域时,可以用bakerbondabxtm树脂(j.t.baker,phillipsburg,nj)进行纯化。取决于待回收的抗体,也可用其他用于蛋白质纯化的技术,如离子交换柱上的分级分离、乙醇沉淀、反相hplc、硅石上的层析、肝素sepharosetm上的层析、阴离子或阳离子交换树脂(如聚天冬氨酸柱)上的层析、层析聚焦、sds-page和硫酸铵沉淀。
任何一个或多个初步纯化步骤后,可以对包含目的抗体和污染物的混合物进行低ph疏水作用层析,该层析使用ph在约2.5-4.5之间的洗脱缓冲液,优选在低盐浓度(例如约0-0.25m盐)下进行。(对于任意前述具体方法)备选地或额外地,异源多聚体蛋白质的产生可以包括透析含有多肽混合物的溶液。
x.使用杆状病毒生产抗体
可以通过用例如lipofectin(可从gibco-brl购得),将编码抗体或抗体片段的质粒和baculogoldtm病毒dna(pharmingen)共转染入昆虫细胞诸如草地贪夜蛾(spodopterafrugiperda)细胞(例如sf9细胞;atcccrl1711)或黑腹果蝇(drosophilamelanogaster)s2细胞,产生重组杆状病毒。在一个具体实例中,将抗体序列融合在包含于杆状病毒表达载体内的表位标签(epitopetag)上游。这类表位标签包括聚组氨酸标签。可以利用多种质粒,包括衍生自诸如pvl1393(novagen)或pacgp67b(pharmingen)的市售质粒的质粒。简言之,可以用与5’和3’区域互补的引物通过pcr来扩增编码抗体或其片段的序列。5’引物可以掺入侧翼(选择的)限制酶位点。然后可以用所选择的限制酶消化产物,并亚克隆入表达载体。
用表达载体转染后,在28℃孵育宿主细胞(例如sf9细胞)4-5天,收获释放的病毒,并用于进一步扩增。病毒感染和蛋白质表达可以按例如o’reilley等(baculovirusexpressionvectors:alaboratorymanual.oxford:oxforduniversitypress(1994))所述进行。
然后可以例如按以下方式,通过ni2+-螯合亲和层析来纯化所表达的带聚组氨酸标签的抗体。可以按rupert等(nature362:175-179(1993))所述,从重组病毒转染的sf9细胞制备提取物。简言之,洗涤sf9细胞,重悬在超声处理缓冲液(25mlhepesph7.9;12.5mmmgcl2;0.1mmedta;10%甘油;0.1%np-40;0.4mkcl)中,冰上超声处理两次20秒。通过离心澄清超声处理产物,将上清50倍稀释在上样缓冲液(50mm磷酸;300mmnacl;10%甘油ph7.8)中,滤过0.45μm滤器。制备柱床体积为5ml的ni2+-nta琼脂糖柱(可从qiagen购得),用25ml水洗涤,并用25ml上样缓冲液平衡。按每分钟0.5ml的流速将经过滤的细胞提取物上样至柱上。用上样缓冲液洗涤柱至基线a280,在此点开始级分收集。然后,用洗脱非特异性结合的蛋白质的第二洗涤缓冲液(50mm磷酸;300mmnacl;10%甘油ph6.0),洗涤柱。再次达到a280基线后,用含0至500mm咪唑梯度的第二洗涤缓冲液,洗柱。收集1ml级分,并通过sds-page和银染或使用ni2+-nta缀合的碱性磷酸酶(qiagen)以western印迹来分析。混合含有洗脱的带his10标签的抗体的级分,并对上样缓冲液透析。
备选地,可以用包括例如蛋白a或蛋白g柱层析的已知层析技术进行抗体的纯化。在一个实施方案中,可以通过洗脱入含有离液剂或温和去垢剂的溶液中来从柱的固相回收目的抗体。示例性离液剂和温和去垢剂包括但不限于盐酸胍、尿素、高氯酸锂(lithiumperclorate)、精氨酸、组氨酸、sds(十二烷基硫酸钠)、tween、triton和np-40,其全都是市售可得的。
iv.靶分子
本发明的异源多聚体蛋白质可以靶向的分子的实例包括但不限于可溶性血清蛋白质和它们的受体及其他膜结合的蛋白质(例如黏附素)。见wo2011/133886,其在此以其整体引入作为参考。
在另一实施方案中,本发明的异源多聚体蛋白质能够结合选自以下的一种、两种或多种细胞因子、细胞因子相关蛋白质和细胞因子受体:bmpi、bmp2、bmp3b(gdfio)、bmp4、bmp6、bmp8、csfi(m-csf)、csf2(gm-csf)、csf3(g-csf)、epo、fgfi(afgf)、fgf2(bfgf)、fgf3(int-2)、fgf4(hst)、fgf5、fgf6(hst-2)、fgf7(kgf)、fgf9、fgf10、fgf11、fgf12、fgf12b、fgf14、fgf16、fgf17、fgf19、fgf20、fgf21、fgf23、igf1、igf2、ifnai、ifna2、ifna4、ifna5、ifna6、ifna7、ifnbi、ifng、ifnwi、feli、feli(epselon)、feli(zeta)、il1a、il1b、il2、il3、il4、il5、il6、il7、il8、il9、il10、il11、il12a、il12b、il13、il14、il15、il16、il17、il17b、il18、il19、il20、il22、il23、il24、il25、il26、il27、il28a、il28b、il29、il30、pdgfa、pdgfb、tgfa、tgfb1、tgfb2、tgfb3、lta(tnf-b)、ltb、tnf(tnf-a)、tnfsf4(ox40配体)、tnfsf5(cd40配体)、tnfsf6(fasl)、tnfsf7(cd27配体)、tnfsf8(cd30配体)、tnfsf9(4-1bb配体)、tnfsfio(trail)、tnfsf1i(trance)、tnfsf12(apo3l)、tnfsf13(april)、tnfsf13b、tnfsf14(hvem-l)、tnfsf15(vegi)、tnfsf18、hgf(vegfd)、vegf、vegfb、vegfc、ilir1、il1r2、il1rl1、il1rl2、il2ra、il2rb、il2rg、il3ra、il4r、il5ra、il6r、il7r、il8ra、il8rb、il9r、ili0ra、ili0rb、il11ra、il12rb1、il12rb2、il13ra1、il13ra2、il15ra、il17r、il18r1、il20ra、il21r、il22r、il1hy1、il1rap、il1rapl1、il1rapl2、il1rn、il6st、il18bp、il18rap、il22ra2、aifi、hgf、lep(瘦蛋白)、ptn和thpo。
在另一实施方案中,靶分子是选自以下的趋化因子、趋化因子受体或趋化因子相关蛋白质:ccli(i-309)、ccl2(mcp-1/mcaf)、ccl3(mip-la)、ccl4(mip-lb)、ccl5(rantes)、ccl7(mcp-3)、ccl8(mcp-2)、cclh(eotaxin)、ccl13(mcp-4)、ccl15(mip-ld)、ccl16(hcc-4)、ccl17(tarc)、ccl18(parc)、ccl19(mdp-3b)、ccl20(mip-3a)、ccl21(slc/exodus-2)、ccl22(mdc/stc-i)、ccl23(mpif-i)、ccl24(mpif-2/eotaxin-2)、ccl25(teck)、ccl26(eotaxin-3)、ccl27(ctack/ilc)、ccl28、cxcli(groi)、cxcl2(gro2)、cxcl3(gr03)、cxcl5(ena-78)、cxcl6(gcp-2)、cxcl9(mig)、cxcl10(ip10)、cxcl11(l-tac)、cxcl12(sdfi)、cxcl13、cxcl14、cxcl16、pf4(cxcl4)、ppbp(cxcl7)、cx3cl1(scydi)、scyei、xcli(lymphotactin)、xcl2(scm-lb)、blri(mdr15)、ccbp2(d6/jab61)、ccr1(ckri/hm145)、ccr2(mcp-irb/ra)、ccr3(ckr3/cmkbr3)、ccr4、ccr5(cmkbr5/chemr13)、ccr6(cmkbr6/ckr-l3/strl22/dry6)、ccr7(ckr7/ebii)、ccr8(cmkbr8/teri/ckr-li)、ccr9(gpr-9-6)、ccrli(vshki)、ccrl2(l-ccr)、xcri(gpr5/ccxcri)、cmklri、cmkori(rdci)、cx3cr1(v28)、cxcr4、gpr2(ccrio)、gpr31、gpr81(fksg80)、cxcr3(gpr9/ckr-l2)、cxcr6(tymstr/strl33/bonzo)、hm74、il8ra(il8ra)、il8rb(il8rb)、ltb4r(gpr16)、tcpio、cklfsf2、cklfsf3、cklfsf4、cklfsf5、cklfsf6、cklfsf7、cklfsf8、bdnf、c5r1、csf3、grccio(cio)、epo、fy(darc)、gdf5、hdfia、dl8、prl、rgs3、rgs13、sdf2、slit2、tlr2、tlr4、tremi、trem2和vhl。
在另一实施方案中,本发明的异源多聚体蛋白质能够结合选自以下的一种或多种靶标:abcfi;acvri;acvrib;acvr2;acvr2b;acvrli;ad0ra2a;aggrecan;agr2;aicda;aifi;aigi;akapi;akap2;amh;amhr2;angpti;angpt2;angptl3;angptl4;anpep;apc;apoci;ar;azgpi(锌-a-糖蛋白);b7.1;b7.2;bad;baff(blys);bagi;bah;bcl2;bcl6;bdnf;blnk;blri(mdr15);bmpi;bmp2;bmp3b(gdfio);bmp4;bmp6;bmp8;bmpria;bmprib;bmpr2;bpagi(网蛋白);brcai;c19orflo(il27w);c3;c4a;c5;c5r1;canti;casp1;casp4;cavi;ccbp2(d6/jab61);ccli(1-309);cclii(eotaxin);ccl13(mcp-4);ccl15(mip-id);ccl16(hcc-4);ccl17(tarc);ccl18(parc);ccl19(mip-3b);ccl2(mcp-1);mcaf;ccl20(mip-3a);ccl21(mtp-2);slc;exodus-2;ccl22(mdc/stc-i);ccl23(mpif-1);ccl24(mpif-2/eotaxin-2);ccl25(teck);ccl26(eotaxin-3);ccl27(ctack/ilc);ccl28;ccl3(mtp-la);ccl4(mdp-lb);ccl5(rantes);ccl7(mcp-3);ccl8(mcp-2);ccnai;ccna2;ccndi;ccnei;ccne2;ccri(ckri/hm145);ccr2(mcp-irb/ra);ccr3(ckr3/cmkbr3);ccr4;ccr5(cmkbr5/chemr13);ccr6(cmkbr6/ckr-l3/strl22/dry6);ccr7(ckr7/ebii);ccr8(cmkbr8/teri/ckr-li);ccr9(gpr-9-6);ccrli(vshki);ccrl2(l-ccr);cd164;cd19;cdic;cd20;cd200;cd22;cd24;cd28;cd3;cd37;cd38;cd3e;cd3g;cd3z;cd4;cd40;cd40l;cd44;cd45rb;cd52;cd69;cd72;cd74;cd79a;cd79b;cd8;cd80;cd81;cd83;cd86;cdhi(e-钙黏着蛋白);cdh10;cdh12;cdh13;cdh18;cdh19;cdh20;cdh5;cdh7;cdh8;cdh9;cdk2;cdk3;cdk4;cdk5;cdk6;cdk7;cdk9;cdknia(p21wapl/cipl);cdknib(p27kipl);cdknic;cdkn2a(p16ink4a);cdkn2b;cdkn2c;cdkn3;cebpb;ceri;chga;chgb;几丁质酶;chst10;cklfsf2;cklfsf3;cklfsf4;cklfsf5;cklfsf6;cklfsf7;cklfsf8;cldn3;cldn7(claudin-7);cln3;clu(簇蛋白);cmklri;cmkori(rdci);cnri;col18a1;coliai;col4a3;col6a1;cr2;crp;csfi(m-csf);csf2(gm-csf);csf3(gcsf);ctla4;ctnnbi(b-连环蛋白);ctsb(组织蛋白酶b);cx3cl1(scydi);cx3cr1(v28);cxcli(groi);cxcl10(ip-10);cxclii(l-tac/ip-9);cxcl12(sdfi);cxcl13;cxcl14;cxcl16;cxcl2(gro2);cxcl3(gro3);cxcl5(ena-78/lix);cxcl6(gcp-2);cxcl9(mig);cxcr3(gpr9/ckr-l2);cxcr4;cxcr6(tymstr/strl33/bonzo);cyb5;cyci;cysltri;dab2ip;des;dkfzp451j0118;dncli;dpp4;e2f1;ecgfi;edgi;efnai;efna3;efnb2;egf;egfr;elac2;eng;eno1;eno2;eno3;ephb4;epo;erbb2(her-2);ereg;erk8;esri;esr2;f3(tf);fadd;fasl;fasn;fceria;fcer2;fcgr3a;fgf;fgfi(afgf);fgf10;fgf11;fgf12;fgf12b;fgf13;fgf14;fgf16;fgf17;fgf18;fgf19;fgf2(bfgf);fgf20;fgf21;fgf22;fgf23;fgf3(int-2);fgf4(hst);fgf5;fgf6(hst-2);fgf7(kgf);fgf8;fgf9;fgfr3;figf(vegfd);feli(epsilon);fili(zeta);flj12584;flj25530;flrti(纤连蛋白);flti;fos;fosli(fra-i);fy(darc);gabrp(gabaa);gagebi;gageci;galnac4s-6st;gata3;gdf5;gfi1;ggt1;gm-csf;gnasi;gnrhi;gpr2(ccrio);gpr31;gpr44;gpr81(fksg80);grccio(cio);grp;gsn(凝溶胶蛋白);gstpi;havcr2;hdac4;hdac5;hdac7a;hdac9;hgf;hifia;hdpi;组胺和组胺受体;hla-a;hla-dra;hm74;hmoxi;humcyt2a;iceberg;icosl;id2;ifn-a;ifnai;ifna2;ifna4;ifna5;ifna6;ifna7;ifnb1;ifnγ;dfnwi;igbpi;igfi;igfir;igf2;igfbp2;igfbp3;igfbp6;il-i;il10;il10ra;il10rb;il11;il11ra;il-12;il12a;il12b;il12rb1;il12rb2;il13;il13ra1;il13ra2;il14;il15;il15ra;il16;il17;il17b;il17c;il17r;il18;il18bp;il18r1;il18rap;il19;il1a;il1b;ilif10;il1f5;il1f6;il1f7;il1f8;il1f9;il1hyi;il1ri;il1r2;il1rap;il1rapl1;il1rapl2;il1rl1;il1rl2;ilirn;il2;il20;il20ra;il21r;il22;il22r;il22ra2;il23;il24;il25;il26;il27;il28a;il28b;il29;il2ra;il2rb;il2rg;il3;il30;il3ra;il4;il4r;il5;il5ra;il6;il6r;il6st(糖蛋白130);el7;el7r;el8;il8ra;dl8rb;il8rb;dl9;dl9r;dlk;inha;inhba;insl3;insl4;iraki;erak2;itgai;itga2;itga3;itga6(a6整联蛋白);itgav;itgb3;itgb4(b4整联蛋白);jagi;jaki;jak3;jun;k6hf;kaii;kdr;kitlg;klf5(gcboxbp);klf6;klkio;klk12;klk13;klk14;klk15;klk3;klk4;klk5;klk6;klk9;krt1;krt19(角蛋白19);krt2a;khthb6(毛发特异性h型角蛋白);lamas;lep(瘦蛋白);lingo-p75;lingo-troy;lps;lta(tnf-b);ltb;ltb4r(gpr16);ltb4r2;ltbr;macmarcks;mag或omgp;map2k7(c-jun);mdk;mibi;midkine;mef;mip-2;mki67;(ki-67);mmp2;mmp9;ms4a1;msmb;mt3(metallothionectin-lll);mtssi;muci(黏蛋白);myc;myd88;nck2;神经蛋白聚糖;nfkbi;nfkb2;ngfb(ngf);ngfr;ngr-lingo;ngr-nogo66(nogo);ngr-p75;ngr-troy;nmei(nm23a);n0x5;nppb;nrobi;nr0b2;nridi;nr1d2;nr1h2;nr1h3;nr1h4;nr1i2;nr1i3;nr2c1;nr2c2;nr2e1;nr2e3;nr2f1;nr2f2;nr2f6;nr3c1;nr3c2;nr4a1;nr4a2;nr4a3;nr5a1;nr5a2;nr6a1;nrpi;nrp2;nt5e;ntn4;odzi;oprdi;p2rx7;pap;parti;pate;pawr;pca3;pcna;pdgfa;pdgfb;pecami;pf4(cxcl4);pgf;pgr;phosphacan;pias2;pik3cg;plau(upa);plg;plxdci;ppbp(cxcl7);ppid;pri;prkcq;prkdi;prl;proc;prok2;psap;psca;ptafr;pten;ptgs2(cox-2);ptn;rac2(p21rac2);rarb;rgsi;rgs13;rgs3;rnfiio(znf144);robo2;s100a2;scgb1d2(亲脂素b);scgb2a1(乳腺珠蛋白2);scgb2a2(乳腺珠蛋白1);scyei(内皮单核细胞激活细胞因子);sdf2;serpinai;serpina3;serp1nb5(maspin);serpinei(pai-i);serpdmf1;shbg;sla2;slc2a2;slc33a1;slc43a1;slit2;sppi;sprrib(sprl);st6gal1;stabi;stat6;steap;steap2;tb4r2;tbx21;tcpio;tdgfi;tek;tgfa;tgfbi;tgfbiii;tgfb2;tgfb3;tgfbi;tgfbri;tgfbr2;tgfbr3;thil;thbsi(血小板反应蛋白-1);thbs2;thbs4;thpo;tie(tie-1);tmp3;组织因子;tlrio;tlr2;tlr3;tlr4;tlr5;tlr6;tlr7;tlr8;tlr9;tnf;tnf-a;tnfaep2(b94);tnfaip3;tnfrsfiia;tnfrsfia;tnfrsfib;tnfrsf21;tnfrsf5;tnfrsf6(fas);tnfrsf7;tnfrsf8;tnfrsf9;tnfsfio(trail);tnfsfi1(trance);tnfsf12(apo3l);tnfsf13(april);tnfsf13b;tnfsf14(hvem-l);tnfsf15(vegi);tnfsf18;tnfsf4(ox40配体);tnfsf5(cd40配体);tnfsf6(fasl);tnfsf7(cd27配体);tnfsf8(cd30配体);tnfsf9(4-1bb配体);tollip;toll样受体;top2a(拓扑异构酶ea);tp53;tpmi;tpm2;tradd;trafi;traf2;traf3;traf4;traf5;traf6;tremi;trem2;trpc6;tslp;tweak;vegf;vegfb;vegfc;多能聚糖;vhlc5;vla-4;xcli(lymphotactin);xcl2(scm-lb);xcri(gpr5/ccxcri);yyi;及zfpm2。
本发明所涵盖的抗体的优选靶分子包括cd蛋白质,如cd3、cd4、cd8、cd16、cd19、cd20、cd34;cd64、cd200;erbb受体家族的成员,如egf受体、her2、her3或her4受体;细胞黏附分子,如lfa-1、mad、p150.95、vla-4、icam-1、vcam、α4/β7整联蛋白和αv/β3整联蛋白,包括其α或β亚基(例如抗-cd11a、抗-cd18或抗-cd11b抗体);生长因子,如vegf-a、vegf-c;组织因子(tf);α干扰素(αlfn);tnfα;白细胞介素,如il-1β、il-3、il-4、il-5、il-8、il-9、il-13、il17a/f、il-18、il-13rα1、il13rα2、il-4r、il-5r、il-9r、ige;血型抗原;flk2/flt3受体;肥胖(ob)受体;mpl受体;ctla-4;rankl、rank、rsvf蛋白、蛋白c等。
在一个实施方案中,本发明的异源多聚体蛋白质结合低密度脂蛋白受体相关蛋白质(lrp-1)或lrp-8或转铁蛋白受体、和至少一个选自以下的靶标:1)β-分泌酶(bace1或bace2)、2)α-分泌酶、3)γ-分泌酶、4)tau-分泌酶、5)淀粉样蛋白前体蛋白(app)、6)死亡受体6(dr6)、7)淀粉样蛋白β肽、8)α-突触核蛋白、9)帕金蛋白、10)亨廷顿蛋白、11)p75ntr和12)胱天蛋白酶-6。
在一个实施方案中,本发明的异源多聚体蛋白质结合选自以下的至少两种靶分子:il-1α和il-1β;il-12和il-18;il-13和il-9;il-13和il-4;il-13和il-5;il-5和il-4;il-13和il-1β;il-13和il-25;il-13和tarc;il-13和mdc;il-13和mef;il-13和tgf-β;il-13和lhr激动剂;il-12和tweak;il-13和cl25;il-13和sprr2a;il-13和sprr2b;il-13和adam8;il-13和ped2;il17a和il17f;cd3和cd19;cd138和cd20;cd138和cd40;cd19和cd20;cd20和cd3;cd38和cd138;cd38和cd20;cd38和cd40;cd40和cd20;cd-8和il-6;cd20和br3;tnfα和tgf-β;tnfα和il-1β;tnfα和il-2;tnfα和il-3;tnfα和il-4;tnfα和il-5;tnfα和il6;tnfα和il8;tnfα和il-9;tnfα和il-10;tnfα和il-11;tnfα和il-12;tnfα和il-13;tnfα和il-14;tnfα和il-15;tnfα和il-16;tnfα和il-17;tnfα和il-18;tnfα和il-19;tnfα和il-20;tnfα和il-23;tnfα和ifnα;tnfα和cd4;tnfα和vegf;tnfα和mif;tnfα和icam-1;tnfα和pge4;tnfα和peg2;tnfα和rank配体;tnfα和te38;tnfα和baff;tnfα和cd22;tnfα和ctla-4;tnfα和gp130;tnfa和il-12p40;vegf和her2;vegf-a和her2;vegf-a和pdgf;her1和her2;vegf-a和vegf-c;vegf-c和vegf-d;her2和dr5,vegf和il-8;vegf和met;vegfr和met受体;vegfr和egfr;her2和cd64;her2和cd3;her2和cd16;her2和her3;egfr(heri)和her2;egfr和her3;egfr和her4;il-13和cd40l;il4和cd40l;tnfr1和il-1r;tnfr1和il-6r;tnfr1和il-18r;epcam和cd3;mapg和cd28;egfr和cd64;cspgs和rgma;ctla-4和btno2;igf1和igf2;igf1/2和erb2b;mag和rgma;ngr和rgma;nogoa和rgma;omgp和rgma;pdl-i和ctla-4;及rgma和rgmb
可以用可选地缀合至其他分子的可溶性抗原或其片段作为免疫原来产生抗体。对于跨膜分子如受体,可以用这些的片段(例如受体的胞外域)作为免疫原。备选地,可以用表达跨膜分子的细胞作为免疫原。这类细胞可以衍生自天然来源(例如癌细胞系)或可以是已通过重组技术转化以表达跨膜分子的细胞。用于制备抗体的其他抗原及其形式对本领域技术人员而言显而易见。
v.实施方案
本发明提供下文所述的其它实施方案。在第一实施方案中,提供产生异源多聚体蛋白质的方法,该方法包括:获得蛋白a纯化的第一含铰链多肽;获得蛋白a纯化的第二含铰链多肽;调节各半抗体的ph至4和9之间;混合该第一和第二含铰链多肽来获得混合的含铰链多肽池;向该混合的含铰链多肽池加入摩尔过量的弱还原剂;用弱还原剂孵育该混合的含铰链多肽池,以形成包含该第一和第二含铰链多肽的异源多聚体蛋白质。
在第二实施方案中,根据第一实施方案,该第一和第二含铰链多肽可以选自半抗体、免疫黏附素及其片段。在第三实施方案中,根据第一实施方案,该第一含铰链多肽是半抗体。在第四实施方案中,根据第一实施方案,该第二含铰链多肽是fc成分。在第五实施方案中,根据第三实施方案,该半抗体包含vl结构域、cl结构域、vh结构域、ch1结构域、铰链结构域、ch2结构域和ch3结构域。在第六实施方案中,根据第五实施方案,该半抗体是进一步包含系链的单条多肽链,其中按n端至c端所述结构域的相对彼此位置如下:vl-cl-系链-vh-ch1-铰链-ch2-ch3。在第七实施方案中,根据第一实施方案,在蛋白a纯化之前混合该第一和第二含铰链多肽,并在蛋白a上共纯化。在第八实施方案中,根据第一实施方案,该第一和第二含铰链多肽包含异源多聚化结构域。在第九实施方案中,根据第八实施方案,该异源多聚化结构域选自杵入臼突变、亮氨酸拉链、静电等。在第十实施方案中,根据第九实施方案,该第一含铰链多肽包含杵,而该第二含铰链多肽包含臼。在第十一实施方案中,根据第一实施方案,在混合后调节ph。在第十二实施方案中,根据第一或第十一实施方案,该方法进一步包括在调节ph之前加入l-精氨酸到20mm至1m之间的终浓度。在第十三实施方案中,根据第一实施方案,该方法进一步包括在15℃至39℃之间的温度孵育该混合池至少30分钟。在第十四实施方案中,根据第一实施方案,组装混合物具有-200至-600mv之间、更优选-300至-500mv之间、最优选约-400mv的氧化电位。在第十五实施方案中,根据第一实施方案,该弱还原剂选自gsh、β-巯基乙胺、半胱氨酸/胱氨酸、gsh/gssg、半胱胺/胱胺、甘氨酰半胱氨酸、和β-巯基乙醇。在第十六实施方案中,根据第一实施方案,按50-600摩尔过量加入该弱还原剂。在第十七实施方案中,根据第一实施方案,在混合之前加入该弱还原剂。在第十八实施方案中,根据第十七实施方案,在混合前不到一小时进行该加入。在第十九实施方案中,根据第一实施方案,在聚乙烯吡咯烷酮(pvp)的存在下在15℃至39℃之间的温度进行孵育组装混合物的步骤。在第二十实施方案中,根据第十九实施方案,在pvp之前、与pvp同时或在pvp之后加入组氨酸。在第二十一实施方案中,根据第十九实施方案,加入pvp至不超过40%(w/v)。
在第二十二实施方案中,本发明提供产生双特异性抗体的方法,该方法包括:
a.获得蛋白a纯化的第一半抗体;
b.获得蛋白a纯化的第二半抗体;
c.向各半抗体中加入l-精氨酸溶液;
d.调节各半抗体的ph至4和9之间;
e.混合该第一和第二半抗体池以获得混合的半抗体池;
f.向该混合的半抗体池加入摩尔过量的弱还原剂;
g.在pvp的存在下在15℃至39℃之间的温度孵育该混合的半抗体池;
由此产生包含该第一和第二半抗体的双特异性抗体。
在第二十三实施方案中,本发明提供产生异源多聚体的方法,该方法包括:(a)提供至少两种不同的含铰链多肽的含l-精氨酸的混合物,其中该混合物具有7至9之间的ph;(b)加入摩尔过量的弱还原剂;和(c)在产生异源多聚体的条件下孵育该混合物。
在第二十四实施方案中,根据第二十二实施方案,第一半抗体是包含以下的单链多肽:(a)含有vl结构域和cl结构域的全长轻链;(b)系链;(c)含有vh结构域、ch1结构域、铰链、ch2结构域和ch3结构域的全长重链;该多肽包含按n端至c端相对彼此位置如下的轻链和重链结构域:vl-cl-cl/vh-系链-vh-ch1-铰链-ch2-ch3。在第二十五实施方案中,根据第二十四实施方案,单链多肽进一步包含异源多聚化结构域。在第二十六实施方案中,根据第二十五实施方案,异源多聚化结构域是臼(例如凹陷)或杵(例如凸起)。在第二十七实施方案中,根据第二十六实施方案,在第一半抗体包含杵时,该第二半抗体包含臼。在第二十八实施方案中,根据第二十六实施方案,在第一半抗体包含臼时,该第二半抗体包含杵。在第二十九实施方案中,根据第二十四实施方案,系链包含ggs重复序列。在第三十实施方案中,根据第二十四实施方案,系链为15-50个氨基酸。
在第三十一实施方案中,本发明提供产生异源多聚体蛋白质的方法,该方法包括:(a)获得蛋白a纯化的第一含铰链多肽;(b)获得蛋白a纯化的第二含铰链多肽;(c)在l-精氨酸的存在下调节各含铰链多肽的ph至4和9之间;(d)混合该第一和第二含铰链多肽来获得混合的半抗体池,并孵育以形成包含该第一和第二含铰链多肽的异源多聚体蛋白质。
在第三十二实施方案中,根据第一或第三十一实施方案,至少一个半抗体是包含以下的单链多肽:(a)包含vl结构域和cl结构域的全长轻链;(b)系链;(c)包含vh结构域、ch1结构域、铰链、ch2结构域和ch3结构域的全长重链。
在第三十三实施方案中,根据第一实施方案,第一含铰链多肽是包含按n端至c端方向彼此相对位置如下的轻链和重链结构域的单链多肽:vl-cl-vh-ch1-铰链-ch2-ch3。在第三十四实施方案中,根据第三十三实施方案,该单链多肽进一步包含系链,其中按n端至c端方向所述结构域彼此相对位置如下:vl-cl-系链-vh-ch1-铰链-ch2-ch3。
在第三十五实施方案中,本发明提供产生异源多聚体的方法,该方法包括提供包含精氨酸的含铰链多肽的混合物,该混合物具有4至9之间的ph,加入弱还原剂并在产生异源多聚体的条件下孵育。
在第三十六实施方案中,本发明提供已改造来表达半抗体的宿主细胞,其中该半抗体是包含系链、vl结构域、cl结构域、vh结构域、ch1结构域、铰链结构域、ch2结构域和ch3结构域的单链多肽,其中按n端至c端方向所述结构域的相对彼此位置如下:vl-cl-系链-vh-ch1-铰链-ch2-ch3。在第三十七实施方案中,根据第三十六实施方案,单链多肽进一步包含异源多聚化结构域。在第三十八实施方案中,根据第三十六或第三十七实施方案,该宿主细胞选自原核细胞、真核细胞、哺乳动物细胞或植物细胞。在第三十九实施方案中,根据第三十八实施方案,该宿主细胞是原核细胞。在第四十实施方案中,根据第三十九实施方案,该原核细胞是大肠杆菌细胞。在第四十一实施方案中,根据第四十实施方案,该大肠杆菌细胞为lpp缺陷。在第四十二实施方案中,根据第三十八实施方案,该宿主细胞是哺乳动物细胞。在第四十三实施方案中,根据第四十一实施方案,该哺乳动物细胞是cho细胞。在第四十四实施方案中,根据第三十六或三十七实施方案,该宿主细胞包含编码该单链半抗体的载体。在第四十五实施方案中,根据第36或第37实施方案,半抗体进一步包含异源多聚化结构域。在第46实施方案中,根据第45实施方案,异源多聚化结构域选自杵、臼、界面内在静电上不利于同二聚体形成但在静电上有利于异二聚体形成的一个或多个带电荷氨基酸、改变来增强分子内离子相互作用的一个或多个氨基酸、卷曲螺旋和亮氨酸拉链。
在第47实施方案中,本发明提供包含改造来表达第一单链半抗体的第一宿主细胞和改造来表达fc成分的第二宿主细胞的宿主细胞混合物。在第48实施方案中,根据第36实施方案,通过宿主细胞产生的半抗体包含异源多聚化结构域。
在以下实验公开内容中,应用以下缩写:eq(等同物)、m(摩尔浓度)、μm(微摩尔浓度)、n(正常)、mol(摩尔)、mmol(毫摩尔)、μmol(微摩尔)、nmol(纳摩尔)、g(克)、mg(毫克)、kg(千克)、μg(微克)、l(升)、ml(毫升)、μl(微升)、cm(厘米)、mm(毫米)、μm(微米)、nm(纳米)、℃(摄氏度)、h(小时)、min(分钟)、sec(秒)、msec(毫秒)。
实施例
在以下实施例中更详细地描述本发明,这些实施例并非旨在以任何方式限制本发明所要求保护的范围。附图意在视为本发明的说明书和描述的整体部分。本文引用的所有参考文献在此明确就其中所述的所有内容引入作为参考。
实施例1:表达和纯化
此实施例举例说明半抗体的表达和纯化。
在大肠杆菌中构建和表达半抗体的示例性方法可以见于例如共同未决申请u.s.2011/0287009中,在此以其整体引入作为参考。改变和调整培养和表达条件处于本领域技术人员的能力范围之内。
在大肠杆菌细胞中表达半抗体
表达质粒的构建
将重链和轻链dna编码序列都克隆入表达质粒,该表达质粒包含用于各序列的分开的启动子元件及用于选择含有该表达质粒的细菌细胞的抗生素抗性。该载体构建体还编码用于将抗体多肽输出到细菌细胞周质空间的热稳定肠毒素ii(stii)分泌信号(picken等,1983,infect.immun.42:269-275和lee等,1983,infect.immun.42:264-268)。各链的转录由phoa启动子(kikuchi等,1981,nucleicacidsres.,9:5671-5678)控制,翻译控制由之前描述过的具有经测量的相对翻译强度的stii信号序列变体提供,所述变体在翻译起始区(tif)中包含沉默密码子改变(simmons和yansura,1996,naturebiotechnol.14:629-634及simmons等,2002,j.immunol.methods,263:133-147)。
各半抗体具有按美国专利号7,642,228中所述改造入重链中的杵(凸起)或臼(凹陷)。简言之,首先产生ch3杵突变体。然后通过将靠近配偶体ch3结构域上杵的残基366、368和407进行随机化,产生ch3臼突变体文库。在以下实施例中,在igg1或igg4主链中,杵突变是t366w,而臼具有突变t366s、l368a和y407v。本领域技术人员可以产生在其他免疫球蛋白同种型中的等同突变。此外,本领域技术人员易于理解,优选用于双特异性抗体的两个半抗体是相同的同种型。
表达和纯化
通过在细菌宿主细胞(例如大肠杆菌)中表达重链和轻链构建体,在分开的培养物中产生含有杵或臼突变的半抗体。将表达质粒引入大肠杆菌宿主菌株33d3(ridgway等(1999)59(11):2718)或64b4(w3110.delta.fhua.delta.phoailvg+.delta.prcspr43h1.delta.degp.delta.manalaci.sup.q.delta.ompt),并在含有羧苄青霉素的lb平板上选择转化体。然后用转化体来接种含有羧苄青霉素的lb起子培养物,将此培养物30℃震荡培养过夜。将起子培养物100x稀释在含有羧苄青霉素的磷酸限制培养基c.r.a.p.(simmons等,2002,j.immunol.methods,263:133-147)中,30℃震荡培养该培养物24小时。离心培养物,冷冻细胞沉淀,直至开始抗体纯化。融化沉淀,并按每5克细胞沉淀100mlteb的体积重量比,重悬在含有用盐酸调节至ph7.5的25mmtris碱、125mmnacl和5mmedta的提取缓冲液(teb或tris提取缓冲液)中,使重悬的混合物通过microfluidicscorporationmodel110fmicrofluidizer(newton,mass.)三次来用微流技术破碎细胞进行提取。然后通过15,000xg离心20分钟来澄清细菌细胞提取物,收集上清,并在纯化之前滤过0.22微米醋酸滤膜。
分别通过蛋白a亲和层析纯化各半抗体。将来自杵半抗体的澄清的细胞提取物按2ml/分钟的流速上样在来自gehealthcare(pistcataway,n.j.)的1mlhitrapmabselectsuretm柱上。上样后,用10柱体积(cv)的40mm醋酸钠ph6.0、125mm氯化钠和5mmedta,然后用5柱体积的20mm柠檬酸钠ph6.0洗涤柱,以便于通过阳离子交换柱捕获。用10柱体积(cv)的0.2mm醋酸(ph2-3)洗脱亲和捕获的半抗体。
在cho细胞中表达半抗体
表达质粒的构建
重链和轻链cdna都处在巨细胞病毒立即早期基因启动子和增强子(cmv)的控制之下。各cmv转录起始位点后随剪接供体和受体序列,其限定了从最终的转录物去除的内含子(lucas等,high-levelproductionofrecombinantproteinsinchocellsusingadicistronicdhfrintronexpressionvector.nucl.acidres.(1996)24:1774-9)。谷氨酰胺合成酶(gs)基因作为选择标记用于稳定细胞系开发(sanders等,amplificationandcloningofthechinesehamsterglutaminesynthetasegene.theemboj(1984)3:65-71),并处于sv40早期启动子和增强子的控制之下。
细胞培养
在37℃和5%co2下,在摇瓶中在基于dmem/f12的专利培养基中培养cho细胞。每三至四天,按3x105/ml的接种密度传代细胞。
稳定转染
用lipofectamine2000cd,按照厂家的建议(invitrogen,carlsbad,ca),转染cho细胞。离心所转染的细胞,并接种入含有多种浓度的蛋氨酸亚砜亚胺(msx)的基于dmem/f-12的选择(无谷氨酰胺)培养基。接种后约三周,将单克隆挑入96孔板。通过取上清进行elisa分析来针对抗体生产评价所挑取的克隆。将排序靠前的克隆扩大,并根据抗体效价、有利的代谢(主要是乳酸消耗)和可接受的产品质量属性,进行评价。
表达
在cho细胞中表达每种半抗体。培养2l培养物并收获。
半抗体的纯化
将每种半抗体捕获在mabselectsuretm柱上。然后用4柱体积(cv)的以下缓冲液洗涤柱:由50mmtrisph8.0、150mmnacl组成的平衡缓冲液,由0.4m磷酸钾ph7.0组成的洗涤缓冲液。将每种臂洗脱入0.15m醋酸钠ph2.9中。
上述表达和纯化半抗体的方法通用于不同同种型的igg。
实施例2:加溶剂和ph保持
以下实施例详述在中间ph孵育半抗体如何驱动构象转变和提高组装效率,及加入加溶剂诸如精氨酸和组氨酸如何减少中间ph诱导的半抗体沉淀。
由于ch2和ch3结构域的内表面(其可能包含正常情况下处于抗体的非溶剂暴露表面中的疏水斑块)暴露,半抗体蛋白a池是固有不稳定的。因此,在调节至大于4的ph后,半抗体蛋白a池趋向于沉淀。本发明人发现,存在小浓度的l-精氨酸(在某些实施例中≥50mm)时,半抗体被稳定,并在ph调节时保留在溶液中。诸如精氨酸的加溶剂的加入保持了抗体处于溶液中,降低了ph调节时的浊度,并提高了双特异性组装产率。见图3。精氨酸还保护双特异性半抗体和纯化的双特异性抗体免于在冷冻期间形成聚集。在组氨酸中也观察到类似的沉淀减少作用。
用实施例1中从蛋白a柱回收的蛋白质作为本实施例的起始物质。
向蛋白a纯化的蛋白质中加入l-精氨酸(1m,ph9)至50-600mm的终浓度。随后根据需要用1.5mtris碱ph11将溶液滴定至更高的ph。从蛋白a柱酸性洗脱后提高至中间ph的步骤称为中间ph保持。
由于ch3结构域中的杵和臼突变,与标准抗体相比,双特异性抗体具有不同的柔性程度。由于此独特的柔性及ch2和ch3结构域暴露的内表面,半抗体看起来在ph调节时经历构象转变。见图2。
在本实验中,在将ph调节至大于4的ph时,杵经历了从单体至非共价连接的同二聚体的转变。见图2b。根据大小排阻层析保留时间,臼经历了从较小的流体动力学半径至较大的流体动力学半径的构象转变。该转变在ph5开始发生,ph≥7驱动该转变完成。见图2a。
通过hplc层析进行了用于测定抗体的聚集和寡聚状态的大小排阻层析(sec)。简言之,在agilenthplc1200系统上将蛋白a纯化的抗体加样至tosohtskgelsw2000柱。按0.5ml/分钟的流速用0.2mk3po40.25mkclph6.2洗脱蛋白质(在大肠杆菌中产生的igg1半抗体)。通过uv吸光度和峰面积积分来定量洗脱的蛋白质。见图2a和b。由于半抗体,尤其是ch3结构域中具有突变的杵和臼半抗体更高的柔性,此转变可以为由ph变化诱导的折叠中间体。
但是,中间ph保持可以导致半抗体沉淀。如图3a中所示,精氨酸的存在降低了蛋白a纯化的igg1杵半抗体的ph诱导的浊度。在此实验中,用1m精氨酸来滴定大肠杆菌产生的igg1半抗体蛋白a池中的ph。最终的精氨酸浓度在ph滴定至5.5时为约50mm,在ph7.5时为约200mm,在ph8.5时为约400mm(见图3a)。精氨酸的存在还使杵入臼双特异性抗体的组装产率提高了15%(数据未显示)。
类似地,组氨酸能够降低ph诱导的由沉淀引起的浊度。在mabselectsuretm柱上从大肠杆菌匀浆物纯化“杵”igg1半抗体,得到半抗体浓度为12g/l的蛋白a池。加入四分之一体积的盐酸精氨酸或盐酸组氨酸至最终的添加剂浓度为200mm,加入等体积的纯化水作为“空白”对照。用逐滴加入的浓氢氧化钠(50%w/vnaoh溶液或19.1n)提高样品ph,并记录数据点。用orionross81-03微探头,测量ph。用hach2100实验室浊度计,测量溶液的浊度。
图3b中的数据显示,精氨酸(200mm)和组氨酸(200mm)都减少了分离自大肠杆菌的igg1中ph诱导的沉淀。总结起来,中间ph诱导了有利于双特异性组装的半抗体构象转变,加至中间ph保持步骤的加溶剂减少了ph诱导的沉淀。
实施例3:还原
以下实施例详述如何使用还原条件减少聚集而导致形成更多希望的异源多聚体,例如双特异性抗体。例如,向组装混合物中加入的谷胱甘肽创造了对杵入臼双特异性组装有利的弱还原条件。诸如bmea(β-巯基乙胺)的其他类似种类的还原剂可以具有类似作用。
聚集可以在杵和臼半抗体组装形成双特异性抗体的过程中发生。提高谷胱甘肽水平最小化了组装期间聚集的量。相反,诸如dtt的强还原剂在高浓度有时可以增加聚集。不受限于具体机制,谷胱甘肽似乎作为正确二硫键形成的催化剂发挥作用来改组二硫键,而不是永久性还原二硫键。对于谷胱甘肽组装,为了在目的双特异性产物中形成铰链区二硫键,无需进行缓冲液更换,而这是使用强还原剂时再氧化所需的。在使用诸如谷胱甘肽的弱还原剂时,也无需加入化学再氧化剂。
谷胱甘肽浓度可以表示为体积摩尔浓度或表示为相对于存在于组装混合物中的含铰链多肽或半抗体的量的摩尔比。针对组装混合物中的蛋白质浓度,用靶摩尔比的还原剂控制;这防止由可变的蛋白质浓度引起的过度还原或还原不足。
在此实施例中,按2至200x摩尔过量向混合的半抗体中加入谷胱甘肽。室温孵育样品46小时。在rp-hplc(反相高效液相层析)中,用0.1%三氟乙酸将所有样品稀释至1.0mg/ml的最大浓度。通过280nm光度测量来确定蛋白质浓度。在样品分析之前注入0.1%三氟乙酸的四个样品。这确保柱完全平衡。将蛋白a纯化的大肠杆菌产生的igg1半抗体加样至agilenthplc1200系统上的porosr2/202.1mmdx20mml。按0.5ml/分钟的流速在20分钟用38–49%缓冲液a至0.09%三氟乙酸80%乙腈(缓冲液b)的线性梯度洗脱蛋白质。通过uv吸光度和峰面积积分来定量所洗脱的蛋白质。如图4a中所示,双特异性性形成的水平随谷胱甘肽:ab摩尔比的提高而提高。见图4a。
进行了用于测定抗体的聚集和寡聚状态的大小排阻层析(sec)。简言之,将蛋白a纯化的大肠杆菌产生的igg1半抗体加样至agilenthplc1200系统上的tosohtskgelsw2000柱。按0.5ml/分钟的流速用0.2mk3po40.25mkclph6.2洗脱蛋白质。通过uv吸光度和峰面积积分来定量所洗脱的蛋白质。确认了所观察到的150kd峰由双特异性抗体的形成引起。见图4b。
可以观察到,存在从不想要的单体(即半抗体,杵或臼)和同二聚体向异源多聚体(即双特异性抗体)的峰迁移(图4a和b)。总结起来,数据显示,提高谷胱甘肽与半抗体的摩尔比减少了聚集,并提高双特异性形成(图4b)。
实施例4:温度
此实施例举例说明温度对半抗体稳定性和异源多聚体组装的影响。
半抗体溶液的温度对组装速率具有剧烈影响。图5a中显示大肠杆菌产生的igg1半抗体在较高温度下增强的组装的一个实例。
显示温度对双特异性组装的影响的另一实例显示在图5b中。在此实验中,按实施例1中所述,在大肠杆菌中产生两个igg1半抗体并在蛋白a上纯化。组合半抗体并分为四个等分试样,用于在有或无加热和/或中间ph保持的不同条件下测试双特异性组装。
如图5b中所示,200摩尔过量的谷胱甘肽在不同条件下增强了双特异性igg1抗体形成的速率。对照条件(室温,约20℃,半抗体保持在ph4,无中间ph保持)允许组装双特异性,但速率较慢。在不到达更高温度的情况下,室温保持蛋白a纯化的半抗体于中间ph(杵半抗体为ph5;臼半抗体为ph7)16小时提高了双特异性抗体形成的速率(在图5b中“ph优化”中,组装混合物在ph8.5)。提高温度至37℃而无中间ph保持步骤提高了双特异性抗体组装的速率超过对照,它相对于室温下进行的ph保持步骤和组装也更快。但是,在中间ph保持蛋白a纯化的半抗体(如上文),然后在提高的温度(即37℃)下在ph8.5组装时,观察到了最快的组装速率。在此条件下,在仅约6小时内达到了约80%的双特异性组装(图5b)。总结起来,加热提高了总体组装速率,ph保持和加热对组装具有协同作用。
在igg4双特异性抗体中也观察到了加热增强的组装。图6b的结果显示,在pvp和组氨酸的存在下,加热的由大肠杆菌培养物产生的igg4半抗体达到了与大肠杆菌产生的igg1半抗体的组装相似的组装结果。用上述反相hplc分析了双特异性的量。总结起来,数据显示,加热有利于双特异性形成。
实施例5:稳定剂
以下实施例详述稳定剂可以如何减少由组装和/或中间ph保持期间加热和/或提高的ph引起的聚集。
聚乙烯吡咯烷酮(pvp)是具有吡咯烷酮基团的水溶性不带电荷聚合物。pvp减少了在加热的组装期间的聚集。不受限于具体机制,pvp可能通过与双特异性抗体的疏水斑块相互作用,发挥作用来稳定该双特异性抗体的折叠中间体或保护半抗体免于聚集。
在实施例3中所述的条件下用sec分析了pvp对聚集体形成的影响。与不加入pvp时4%nmws相比,加入pvp使存在于组装池中的高分子量物(hmws)最小化至12%hmws(4%pvp(w/v))。见图6a。所有样品都是在200mm精氨酸存在下加热的大肠杆菌产生的igg1。
随后,测试了igg4双特异性抗体的组装。如图6b中所示,pvp和组氨酸存在下的加热组装使大肠杆菌产生的igg4双特异性组装极大地提高至与图5a中所示的大肠杆菌产生的igg1的加热组装类似的水平。精氨酸作为加溶剂和ph滴定剂存在于加热样品和室温样品二者中。除pvp外,在加热的中间ph保持步骤之前加入另一稳定剂组氨酸,以在此步骤中稳定半抗体。结果显示,pvp和组氨酸都使igg4双特异性组装提高至与igg1双特异性组装类似的水平(图6b与5b比较)。
图6c显示另一实例,其中pvp最小化igg4双特异性抗体的加热组装期间的hmws形成。
加热半抗体蛋白a池加速了半抗体在中间ph孵育时的构象转变。但尤其是对于自大肠杆菌产生的igg4半抗体组装igg4杵和臼双特异性抗体,加热可以引起聚集。因此,在组装期间加热igg4半抗体时,加入额外的加溶剂和/或稳定剂。
室温孵育48小时后检测了大肠杆菌igg4臼半抗体的构象转变。在37℃孵育igg4臼半抗体时,在约三小时内检测到了构象转变(数据未显示)。但是,如通过sec测定,加热导致了聚集的增加。见图7,左图。在此实验中,在加热的中间ph保持步骤中加入组氨酸,测试其对减少半抗体聚集的作用。如图7中所示,加热期间组氨酸的存在使聚集水平从11%高分子量物(hmws,无组氨酸)最小化至6%hmws(200mm组氨酸)而不影响半抗体的构象转变。因此,结果显示,稳定剂在双特异性抗体组装以及半抗体的中间ph保持期间都减少聚集体形成。
实施例6-组装
此实施例提供用于两种示例性免疫球蛋白同种型(即igg1和igg4)的方案。在两种不同宿主细胞(即大肠杆菌或cho)之一中,产生了半抗体。应理解,本文所述的方法可以经调整而适用于在相同来源或其他来源中产生的其他抗体同种型。根据本领域的知识和本申请中公开的教导,通过常规实验来修改该方案处于本领域技术人员的能力范围之内。
此外,应理解,抗体的形成包括,在第一宿主细胞(例如cho)中产生的半抗体可以与在第二宿主细胞(例如大肠杆菌)中产生的互补半抗体组装(数据未显示)。因此,例如,在cho中产生的杵半抗体可以与在大肠杆菌中产生的臼半抗体组装,或者反之亦然。
此实施例中所述的全部四种组装方法都导致了组装,该组装在4小时后达到平台,并产生了少于10%的聚集体和组装期间最小的聚集。
a.来自大肠杆菌的igg1
半抗体的构象变化:如果蛋白a池于小于7的ph,则用1m精氨酸(ph9)调节两个半抗体池的ph至ph7,并在37℃孵育两个池3小时。备选地,在室温孵育了这些池48小时。如果这些池已经于ph7中48小时或更长,则省却此步骤。定量使该溶液达到ph7所加入的精氨酸的量。
组合这些(仍然温暖的)池,用1m精氨酸(ph9)调节ph至ph8.5。定量在此阶段加入的精氨酸的量。
加入2m精氨酸(ph8.5),直至精氨酸的终浓度为0.5m。
加入含200mm还原型谷胱甘肽(gsh)的0.5m精氨酸(最终ph8.5),直至gsh:ab比为200x(例如,每毫克半抗体加入6.88μl该谷胱甘肽溶液)。
37℃孵育该谷胱甘肽半抗体溶液4小时,以允许该半抗体组装为双特异性抗体。
b.来自cho的igg1:
按上文针对大肠杆菌所述组装。对于中间ph保持,可能无需将ph提高至ph7或以上。加入谷胱甘肽后的组装时间可以在2小时后完成。
c.来自cho的igg4:
在与上文(来自cho的igg1)相同的条件下组装。可能无需加入组氨酸和pvp。
d.来自大肠杆菌的igg4:
用以上方案组装来自大肠杆菌的igg4导致了高(即~35%)聚集体水平。这使得有必要修改组装条件:
据测定,包含0.2m组氨酸和50mm精氨酸的蛋白a池组合物产生了可接受的结果(数据未显示)。因此,考察了提供最终结果的几种方式,并确定其提供可接受的结果。
第一种方法在蛋白a纯化期间使用改变的洗脱(ph3)和洗涤缓冲液(ph7),使其包含0.2m组氨酸和50mm精氨酸。这导致包含0.2m组氨酸和50mm精氨酸、ph4的最终蛋白a池,然后用1.5mtris碱(ph11)滴定至ph8。
第二种备选方法利用盐酸组氨酸的0.8m溶液(其具有0.8m的溶解度)。向蛋白a池(一个或多个)加入三分之一体积的盐酸组氨酸,达到0.2m组氨酸的终浓度。然后加入1/40体积的2m精氨酸。
第三种备选方法是对蛋白a池进行缓冲液更换,换入0.2m组氨酸和50mm精氨酸缓冲液(优选ph8)中。
最后一种备选方法是直接向蛋白a池(一个或多个)加入组氨酸(31.03g/l),然后加入1/40体积的2m精氨酸。
向蛋白a池(一个或多个)加入四分之一体积spectrumpvpk-15在0.2m组氨酸和50mm精氨酸中的20%w/v溶液。
应指出,pvp在低谷胱甘肽条件下也使igg1组装期间的聚集最小化。
半抗体的构象变化:用含0.2m组氨酸、50mm精氨酸和4%pvpk-15的1.5mtris碱(ph11)调节蛋白a池(一个或多个)的ph至ph8.0,并在37℃孵育3小时。备选地,可以在室温孵育该池48小时。
加入含200mm还原型谷胱甘肽(gsh)的0.2m组氨酸、4%pvp、50mm精氨酸、最终ph8.0,直至gsh:ab比为200x(例如,每毫克半抗体加入6.88μl谷胱甘肽溶液)。如果半抗体池尚未组合,则在此时组合它们。
37℃孵育混合的半抗体4小时,以允许形成双特异性抗体。此时,百分比双特异性抗体已达到平台。
一旦双特异性抗体的量已达到平台,即可以将溶液保存在低温下或调节至更低的ph,用于在随后的层析步骤上处理。
本文公开的方法可以用于制备诸如双特异性抗体的治疗性蛋白质。
应理解,本文所述的实施例和实施方案仅是为了举例说明的目的,鉴于其,本领域技术人员知晓对本发明的多种修改或改变,其也将包含在本申请的精神和范围以及所附权利要求的范围之内。本文引用的所有出版物、专利和专利申请在此为了所有目的以其整体引入作为参考。
序列表
<110>弗·哈夫曼-拉罗切有限公司(f.hoffmann-larocheag)
<120>双特异性抗体的改进的组装
<130>p4764r1-wo
<150>61/676837
<151>2012-07-27
<150>61/560704
<151>2011-11-16
<150>61/546503
<151>2011-10-12
<150>61/545863
<151>2011-10-11
<160>1
<170>patentinversion3.5
<210>1
<211>6
<212>prt
<213>人工序列
<220>
<223>合成序列
<220>
<221>misc_feature
<222>(2)..(2)
<223>x是任何氨基酸残基
<220>
<221>misc_feature
<222>(4)..(4)
<223>x是任何氨基酸残基
<400>1
argxaaargxaaargarg
15
- 还没有人留言评论。精彩留言会获得点赞!
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺
- 利用特异性挥发物筛选昆虫相关嗅觉蛋白的方法与制造工艺
- 使用与表面偶联的单特异性四聚抗体复合物组合物从样品中分离出靶实体的方法与制造工艺
- 抗-WT1/HLA双特异性抗体的制造方法与工艺