提高大豆转化效率的方法与流程
本发明涉及一种植物转化的方法,特别是涉及一种通过外施筛选剂以提高大豆转化效率的方法。
背景技术:
随着转基因植物种类和种植面积的迅猛增加,转基因植物中选择标记基因的生物安全性已成为人们普遍关注的问题之一。合适的标记基因可以为获得真正的转化体提供有力的判断依据,在植物遗传转化过程中发挥着至关重要的作用。目前在植物遗传转化中广泛应用的标记基因有抗生素抗性基因(如nptⅱ基因、hpt基因等)和除草剂抗性基因(如pat基因、epsps基因、bar基因等),由于选择标记基因一旦转化成功便不再有用,甚至对生态环境和食品安全存在潜在威胁,所以对生物安全性标记基因的开发显得非常重要。
乙酰乳酸合成酶(acetolactatesynthase,以下称为“als”)存在于植物生长过程中,它能以高度专一性和极高的催化效率催化丙酮酸为乙酰乳酸,从而导致支链氨基酸的生物合成。亮氨酸、异亮氨酸、缬氨酸是植物体内3种必需的支链氨基酸,而als不仅是催化亮氨酸、缬氨酸和异亮氨酸生物合成过程中的关键酶,其活性也受产物缬氨酸和异亮氨酸的反馈调节。
als抑制剂是一类已知的除草剂,其通过抑制植物体内的als活性而阻止植物体内缬氨酸、亮氨酸和异亮氨酸的生物合成,导致蛋白质的合成受到破坏,从而使植物细胞的有丝分裂停止于g1阶段的s期(dna合成期)和g2阶段的m期,干扰了dna的合成,细胞因此不能完成有丝分裂,导致植物组织失绿、黄化,植株生长受抑,最终达到杀死生物个体的目的。因此,als抑制剂不仅具有超高活性,而且以其超高效、广谱、低毒、低残留、高选择性和良好的环境相容性备受欢迎,同时由于抑制als抑制剂的靶标不涉及人和动物,故其对人和动物十分安全,亦为标记基因和除草剂耐受性状提供了新的选择。
als抑制剂包括磺酰脲类除草剂、咪唑啉酮类除草剂、三唑并嘧啶类除草剂、嘧啶基硫代苯甲酸类除草剂或磺酰氨基-羰基-三唑啉酮类除草剂等。已报道了在植物转化过程中能以磺酰脲类除草剂水解酶基因作为选择标记物,但转化效率有待提高。
技术实现要素:
本发明的目的是提供一种提高大豆转化效率的方法,有效克服现有技术将als抑制剂配置在培养基中存在的假阳性植株率较高、转化效率低等技术缺陷,为规模化遗传转化及培育具有耐受除草剂性状的植物提供新的选择。
为实现上述目的,本发明提供了一种选择转化的植物细胞的方法,包括:
使用含有目标基因和编码磺酰脲类除草剂水解酶的基因的重组载体,转化植物细胞;
通过外施als抑制剂对上述转化后的植物细胞进行筛选培养,以编码磺酰脲类除草剂水解酶的基因作为选择标记物;
选择未被杀死和/或未被抑制的植物细胞。
进一步地,所述转化植物细胞为通过农杆菌介导过程转化植物细胞。
更进一步地,所述植物细胞为大豆细胞。
具体地,所述外施包括滴加、喷施或涂抹。
具体地,所述als抑制剂包括磺酰脲类化合物、咪唑啉酮类化合物、三唑并嘧啶类化合物、嘧啶基硫代苯甲酸类化合物或磺酰氨基-羰基-三唑啉酮类化合物。
更具体地,所述磺酰脲类化合物为苯磺隆、甲嘧磺隆、吡嘧磺隆、氯吡嘧磺隆、噻吩磺隆、苄嘧磺隆、甲磺隆、胺苯磺隆或氯嘧磺隆。
可选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆1-7μg。
优选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆3μg。
可选地,外施苯磺隆前在增殖培养基中培养5-9天。
优选地,外施苯磺隆前在增殖培养基中培养7天。
在上述技术方案的基础上,所述磺酰脲类除草剂水解酶包含:(a)具有seqidno:2所示的氨基酸序列组成的蛋白质;或(b)在(a)中的氨基酸序列经过取代和/或缺失和/或添加一个或几个氨基酸且具有芳氧基链烷酸酯双加氧酶活性的由(a)衍生的蛋白质;或(c)与seqidno:2具有至少90%序列同一性的氨基酸序列组成的蛋白质。
为实现上述目的,本发明还提供了一种由农杆菌介导且以编码磺酰脲类除草剂水解酶的基因作为选择标记物来制备转基因植物的方法,其中,包括:
制备至少包括能被农杆菌菌株转化的植物细胞的外植体;
将至少包括所述外植体中所述植物细胞的区域接触至少包含编码磺酰脲类除草剂水解酶的基因的农杆菌菌株;
通过外施als抑制剂对所述外植体进行筛选培养;
选择未被杀死和/或未被抑制的转化植物细胞;
所述转化植物细胞再生成植物。
进一步地,所述植物细胞为大豆细胞。
更进一步地,所述外植体为有子叶的外植体、半粒种子外植体或半胚胎种子外植体。
具体地,所述外施包括滴加、喷施或涂抹。
具体地,所述als抑制剂包括磺酰脲类化合物、咪唑啉酮类化合物、三唑并嘧啶类化合物、嘧啶基硫代苯甲酸类化合物或磺酰氨基-羰基-三唑啉酮类化合物。
更具体地,所述磺酰脲类化合物为苯磺隆、甲嘧磺隆、吡嘧磺隆、氯吡嘧磺隆、噻吩磺隆、苄嘧磺隆、甲磺隆、胺苯磺隆或氯嘧磺隆。
可选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆1-7μg。
优选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆3μg。
可选地,外施苯磺隆前在增殖培养基中培养5-9天。
优选地,外施苯磺隆前在增殖培养基中培养7天。
所述转化植物细胞再生成植物具体为通过外施als抑制剂使得所述转化植物细胞在分化培养基中培养分化成植物。
可选地,所述als抑制剂为苯磺隆时,每毫升分化培养基外施苯磺隆3-7μg。
优选地,所述als抑制剂为苯磺隆时,每毫升分化培养基外施苯磺隆7μg。
可选地,外施苯磺隆前在分化培养基中培养14-20天。
优选地,外施苯磺隆前在分化培养基中培养18天。
进一步地,所述农杆菌菌株还包括赋予除草剂耐受性的基因和/或昆虫抗性的基因。
具体地,所述赋予除草剂耐受性的基因编码如下除草剂耐受性蛋白质:5-烯醇丙酮酰莽草酸-3-磷酸合酶、草甘膦氧化还原酶、草甘膦-n-乙酰转移酶、草甘膦脱羧酶、草铵膦乙酰转移酶、α酮戊二酸依赖性双加氧酶、麦草畏单加氧酶、4-羟苯基丙酮酸双加氧酶、乙酰乳酸合酶、细胞色素类蛋白质和/或原卟啉原氧化酶。
具体地,所述昆虫抗性的基因包括cry类基因或vip类基因。
在上述技术方案的基础上,所述磺酰脲类除草剂水解酶包含:(a)具有seqidno:2所示的氨基酸序列组成的蛋白质;或(b)在(a)中的氨基酸序列经过取代和/或缺失和/或添加一个或几个氨基酸且具有芳氧基链烷酸酯双加氧酶活性的由(a)衍生的蛋白质;或(c)与seqidno:2具有至少90%序列同一性的氨基酸序列组成的蛋白质。
为实现上述目的,本发明还提供了一种转化大豆的方法,其中,包括:
大豆种子萌发后去除一片子叶和第一片真叶,获得带有一片子叶的裸露分生组织;
所述带有一片子叶的裸露分生组织接种到含有细胞分裂素的预处理培养基上进行预处理;
包含编码磺酰脲类除草剂水解酶的基因的农杆菌菌株侵染所述预处理后的分生组织块;
通过外施als抑制剂对侵染后的所述分生组织块进行筛选培养,以编码磺酰脲类除草剂水解酶的基因作为选择标记物;
选择未被杀死和/或未被抑制的植物细胞。
具体地,所述细胞分裂素为1mg/l6-苄基腺嘌呤和2mg/l玉米素的任意一种或任意组合。
进一步地,所述预处理培养基还包括乙酰丁香酮。
所述预处理还包括所述分生组织块创伤后进行超声波处理2-4分钟。
具体地,所述外施包括滴加、喷施或涂抹。
具体地,所述als抑制剂包括磺酰脲类化合物、咪唑啉酮类化合物、三唑并嘧啶类化合物、嘧啶基硫代苯甲酸类化合物或磺酰氨基-羰基-三唑啉酮类化合物。
更具体地,所述磺酰脲类化合物为苯磺隆、甲嘧磺隆、吡嘧磺隆、氯吡嘧磺隆、噻吩磺隆、苄嘧磺隆、甲磺隆、胺苯磺隆或氯嘧磺隆。
可选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆1-7μg。
优选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆3μg。
可选地,外施苯磺隆前在增殖培养基中培养5-9天。
优选地,外施苯磺隆前在增殖培养基中培养7天。
进一步地,所述农杆菌菌株还包括赋予除草剂耐受性的基因和/或昆虫抗性的基因。
具体地,所述赋予除草剂耐受性的基因编码如下除草剂耐受性蛋白质:5-烯醇丙酮酰莽草酸-3-磷酸合酶、草甘膦氧化还原酶、草甘膦-n-乙酰转移酶、草甘膦脱羧酶、草铵膦乙酰转移酶、α酮戊二酸依赖性双加氧酶、麦草畏单加氧酶、4-羟苯基丙酮酸双加氧酶、乙酰乳酸合酶、细胞色素类蛋白质和/或原卟啉原氧化酶。
具体地,所述昆虫抗性的基因包括cry类基因或vip类基因。
在上述技术方案的基础上,所述磺酰脲类除草剂水解酶包含:(a)具有seqidno:2所示的氨基酸序列组成的蛋白质;或(b)在(a)中的氨基酸序列经过取代和/或缺失和/或添加一个或几个氨基酸且具有芳氧基链烷酸酯双加氧酶活性的由(a)衍生的蛋白质;或(c)与seqidno:2具有至少90%序列同一性的氨基酸序列组成的蛋白质。
为实现上述目的,本发明还提供了一种生产稳定转化的大豆植物的方法,其中,包括:
大豆种子萌发后去除一片子叶和第一片真叶,获得带有一片子叶的裸露分生组织;
所述带有一片子叶的裸露分生组织接种到含有细胞分裂素的预处理培养基上进行预处理;
包含编码磺酰脲类除草剂水解酶的基因的农杆菌菌株侵染所述预处理后的分生组织块;
侵染后的所述分生组织块与所述农杆菌菌株共培养;
通过外施als抑制剂对上述共培养后的分生组织块进行筛选培养,并以编码磺酰脲类除草剂水解酶的基因作为选择标记物选择转化的抗性组织;
转化的抗性组织再生为大豆植物。
具体地,所述细胞分裂素为1mg/l6-苄基腺嘌呤和2mg/l玉米素的任意一种或任意组合。
进一步地,所述预处理培养基还包括乙酰丁香酮。
所述预处理还包括所述分生组织块创伤后进行超声波处理2-4分钟。
具体地,所述外施包括滴加、喷施或涂抹。
具体地,所述als抑制剂包括磺酰脲类化合物、咪唑啉酮类化合物、三唑并嘧啶类化合物、嘧啶基硫代苯甲酸类化合物或磺酰氨基-羰基-三唑啉酮类化合物。
更具体地,所述磺酰脲类化合物为苯磺隆、甲嘧磺隆、吡嘧磺隆、氯吡嘧磺隆、噻吩磺隆、苄嘧磺隆、甲磺隆、胺苯磺隆或氯嘧磺隆。
可选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆1-7μg。
优选地,所述磺酰脲类化合物为苯磺隆时,每毫升增殖培养基外施苯磺隆3μg。
可选地,外施苯磺隆前在增殖培养基中培养5-9天。
优选地,外施苯磺隆前在增殖培养基中培养7天。
所述转化的抗性组织再生为大豆植物具体为通过外施als抑制剂使得所述转化的抗性组织在分化培养基中培养分化成大豆植物。
可选地,所述als抑制剂为苯磺隆时,每毫升分化培养基外施苯磺隆3-7μg。
优选地,所述als抑制剂为苯磺隆时,每毫升分化培养基外施苯磺隆7μg。
可选地,外施苯磺隆前在分化培养基中培养14-20天。
优选地,外施苯磺隆前在分化培养基中培养18天。
进一步地,所述农杆菌菌株还包括赋予除草剂耐受性的基因和/或昆虫抗性的基因。
具体地,所述赋予除草剂耐受性的基因编码如下除草剂耐受性蛋白质:5-烯醇丙酮酰莽草酸-3-磷酸合酶、草甘膦氧化还原酶、草甘膦-n-乙酰转移酶、草甘膦脱羧酶、草铵膦乙酰转移酶、α酮戊二酸依赖性双加氧酶、麦草畏单加氧酶、4-羟苯基丙酮酸双加氧酶、乙酰乳酸合酶、细胞色素类蛋白质和/或原卟啉原氧化酶。
具体地,所述昆虫抗性的基因包括cry类基因或vip类基因。
在上述技术方案的基础上,所述磺酰脲类除草剂水解酶包含:(a)具有seqidno:2所示的氨基酸序列组成的蛋白质;或(b)在(a)中的氨基酸序列经过取代和/或缺失和/或添加一个或几个氨基酸且具有芳氧基链烷酸酯双加氧酶活性的由(a)衍生的蛋白质;或(c)与seqidno:2具有至少90%序列同一性的氨基酸序列组成的蛋白质。
本发明中的克隆基因、表达盒、载体(例如质粒)、蛋白和蛋白片段、以及转化的细胞核植物均可使用标准方法生产。
本发明可用于在植物中表达任何目的基因。目的基因可以是耐除草剂基因、抗病基因或抗虫基因,或者是选择或评价标记,并含有植物可操作的启动子、编码区和终止子区。耐除草剂基因包括对咪唑啉酮或磺酰脲除草剂耐受的ahas基因、对草丁膦除草剂耐受的pat或bar基因、对草甘膦除草剂耐受的epsps基因,对2,4-d除草剂耐受的aad基因,对hppd抑制剂耐受的hppd基因等。抗病基因包括抗生素合成酶基因,例如硝吡咯菌素合成酶基因,植物来源的抗性基因等。抗虫基因包括苏云金芽孢杆菌杀虫基因。目的基因也可以编码与生物化学途径有关的酶,该酶的表达可改变食物、饲料、营养药和/或药物生产中重要的性状。目的基因可以位于质粒上。适合本发明使用的质粒可以含有一个以上目的基因和/或农杆菌可含有带有不同目的基因的不同质粒。
本发明中所述植物可以为大豆,所述“大豆”是指glycinemax,所述方法是以农杆菌介导的目的基因传输到大豆细胞,之后再生为转化的大豆植物为基础的。本发明的方法独立于栽培品种。
本发明中所述“选择标记物”是指这样的基因或多核苷酸,其表达允许鉴别已经用含有所述基因或多核苷酸dna构建体或载体转化的细胞。选择标记物可以提供对毒性化合物的抗性,例如对抗生素、除草剂等。
本发明中“乙酰乳酸合酶或乙酰乳酸合成酶(als)”是指具有根据iubmb酶命名法ec2.2.1.6定义的活性的酶。酶催化两个丙酮酸分子之间的反应,产生2-乙酰乳酸和co2。酶需要二磷酸硫胺素,并且还可以称为乙酰羟酸合酶(ahas)。
本发明中“als抑制剂”是指抑制野生型als蛋白质的化合物,对含有野生型als的细胞是毒性的。此类化合物包括已知的除草剂,主要包括磺酰脲类、咪唑啉酮类、三唑并嘧啶类、嘧啶基硫代苯甲酸类或磺酰氨基-羰基-三唑啉酮类化合物。
可以在本发明中使用的磺酰脲类化合物包括:1)苯基磺酰脲类,包括氯嘧磺隆、氟嘧磺隆、3-(4-乙基-6-甲氧基-1,3,5-三嗪-2-基)-1-(2,3-二氢-1,1-二氧代-2-甲基苯并[b]噻吩-7-磺酰基)-脲、3-(4-乙氧基-6-乙基-1,3,5-三嗪-2-基)-1-(2,3-二氢-1,1-二氧代-2-甲基苯并[b]噻吩-7-磺酰基)-脲、苯磺隆、甲磺隆、氯磺隆、醚苯磺隆和甲嘧磺隆;2)噻吩基磺酰脲类,如噻吩磺隆;3)吡唑基磺酰脲类,包括吡嘧磺隆和甲基3-氯代-5-(4,6-二甲氧基嘧啶-2-基氨基甲酰基氨磺酰)-1-甲基-吡唑-4-羧酸酯;4)砜肼衍生物,包括酰嘧磺隆和结构类似物;5)吡啶基磺酰脲基,包括烟嘧磺隆和dpx-e9636;6)苯氧基磺酰脲类。
本发明中所述的植物、植物组织或植物细胞的基因组,是指植物、植物组织或植物细胞内的任何遗传物质,且包括细胞核和质体和线粒体基因组。
本发明中所述的“植物繁殖体”包括但不限于植物有性繁殖体和植物无性繁殖体。所述植物有性繁殖体包括但不限于植物种子;所述植物无性繁殖体是指植物体的营养器官或某种特殊组织,其可以在离体条件下产生新植株;所述营养器官或某种特殊组织包括但不限于根、茎和叶,例如:以根为无性繁殖体的植物包括草莓和甘薯等;以茎为无性繁殖体的植物包括甘蔗和马铃薯(块茎)等;以叶为无性繁殖体的植物包括芦荟和秋海棠等。
本发明中所述“抗性”是可遗传的,并允许植物在除草剂对给定植物进行一般除草剂有效处理的情况下生长和繁殖。正如本领域技术人员所认可的,即使植物受到除草剂处理的一定损伤程度明显,植物仍可被认为“抗性”。本发明中术语“耐性”或“耐受性”比术语“抗性”更广泛,并包括“抗性”,以及特定植物具有的抵抗除草剂诱导的各种程度损伤的提高的能力,而在同样的除草剂剂量下一般导致相同基因型野生型植物损伤。
本发明中所述的多核苷酸和/或核苷酸形成完整“基因”,在所需宿主细胞中编码蛋白质或多肽。本领域技术人员很容易认识到,可以将本发明的多核苷酸和/或核苷酸置于目的宿主中的调控序列控制下。
本领域技术人员所熟知的,dna典型的以双链形式存在。在这种排列中,一条链与另一条链互补,反之亦然。由于dna在植物中复制产生了dna的其它互补链。这样,本发明包括对序列表中示例的多核苷酸及其互补链的使用。本领域常使用的“编码链”指与反义链结合的链。为了在体内表达蛋白质,典型将dna的一条链转录为一条mrna的互补链,它作为模板翻译出蛋白质。mrna实际上是从dna的“反义”链转录的。“有义”或“编码”链有一系列密码子(密码子是三个核苷酸,一次读三个可以产生特定氨基酸),其可作为开放阅读框(orf)阅读来形成目的蛋白质或肽。本发明还包括与示例的dna有相当功能的rna。
本发明中多核苷酸或核酸分子或其片段在严格条件下与本发明磺酰脲类除草剂水解酶基因杂交。任何常规的核酸杂交或扩增方法都可以用于鉴定本发明磺酰脲类除草剂水解酶基因的存在。核酸分子或其片段在一定情况下能够与其他核酸分子进行特异性杂交。本发明中,如果两个核酸分子能形成反平行的双链核酸结构,就可以说这两个核酸分子彼此间能够进行特异性杂交。如果两个核酸分子显示出完全的互补性,则称其中一个核酸分子是另一个核酸分子的“互补物”。本发明中,当一个核酸分子的每一个核苷酸都与另一个核酸分子的对应核苷酸互补时,则称这两个核酸分子显示出“完全互补性”。如果两个核酸分子能够以足够的稳定性相互杂交从而使它们在至少常规的“低度严格”条件下退火且彼此结合,则称这两个核酸分子为“最低程度互补”。类似地,如果两个核酸分子能够以足够的稳定性相互杂交从而使它们在常规的“高度严格”条件下退火且彼此结合,则称这两个核酸分子具有“互补性”。从完全互补性中偏离是可以允许的,只要这种偏离不完全阻止两个分子形成双链结构。为了使一个核酸分子能够作为引物或探针,仅需保证其在序列上具有充分的互补性,以使得在所采用的特定溶剂和盐浓度下能形成稳定的双链结构。
本发明中,基本同源的序列是一段核酸分子,该核酸分子在高度严格条件下能够和相匹配的另一段核酸分子的互补链发生特异性杂交。促进dna杂交的适合的严格条件,例如,大约在45℃条件下用6.0×氯化钠/柠檬酸钠(ssc)处理,然后在50℃条件下用2.0×ssc洗涤,这些条件对本领域技术人员是公知的。例如,在洗涤步骤中的盐浓度可以选自低度严格条件的约2.0×ssc、50℃到高度严格条件的约0.2×ssc、50℃。此外,洗涤步骤中的温度条件可以从低度严格条件的室温约22℃,升高到高度严格条件的约65℃。温度条件和盐浓度可以都发生改变,也可以其中一个保持不变而另一个变量发生改变。优选地,本发明所述严格条件可为在6×ssc、0.5%sds溶液中,在65℃下与本发明磺酰脲类除草剂水解酶的核苷酸序列发生特异性杂交,然后用2×ssc、0.1%sds和1×ssc、0.1%sds各洗膜1次。
因此,具有除草剂耐受性活性并在严格条件下与本发明磺酰脲类除草剂水解酶的核苷酸序列杂交的序列包括在本发明中。这些序列与本发明序列至少大约40%-50%同源,大约60%、65%或70%同源,甚至至少大约75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更大的序列同源性。
本发明提供功能蛋白质。“功能活性”(或“活性”)在本发明中指本发明用途的蛋白质/酶(单独或与其它蛋白质组合)具有降解除草剂的能力。产生本发明蛋白质的植物优选产生“有效量”的蛋白质,从而在用除草剂处理植物时,蛋白质表达的水平足以给予植物对除草剂(若无特别说明则为一般用量)完全或部分的抗性或耐性。可以以通常杀死靶植物的用量、正常的大田用量和浓度使用除草剂。优选地,本发明的转化植物和植物细胞优选具有磺酰脲类除草剂的抗性或耐性,即转化的植物和植物细胞能在有效量的磺酰脲类除草剂存在下生长。
本发明中所述的基因和蛋白质不但包括特定的示例序列,还包括保存了所述特定示例的蛋白质的除草剂耐受性活性特征的部分和/片段(包括与全长蛋白质相比在内和/或末端缺失)、变体、突变体、取代物(有替代氨基酸的蛋白质)、嵌合体和融合蛋白。所述“变体”或“变异”是指编码同一蛋白或编码有除草剂抗性活性的等价蛋白的核苷酸序列。所述“等价蛋白”是指与权利要求的蛋白具有相同或基本相同的除草剂耐受性的生物活性的蛋白。
本发明中所述的dna分子或蛋白序列的“片段”或“截短”是指涉及的原始dna或蛋白序列(核苷酸或氨基酸)的一部分或其人工改造形式(例如适合植物表达的序列),前述序列的长度可存在变化,但长度足以确保(编码)蛋白质为除草剂耐受性蛋白质。
由于遗传密码子的丰余性,多种不同的dna序列可以编码相同的氨基酸序列。产生这些编码相同或基本相同的蛋白的可替代dna序列正在本领域技术人员的技术水平内。这些不同的dna序列包括在本发明的范围内。所述“基本上相同的”序列是指有氨基酸取代、缺失、添加或插入但实质上不影响除草剂耐受性活性的序列,亦包括保留除草剂耐受性活性的片段。
本发明中氨基酸序列的取代、缺失或添加是本领域的常规技术,优选这种氨基酸变化为:小的特性改变,即不显著影响蛋白的折叠和/或活性的保守氨基酸取代;小的缺失,通常约1-30个氨基酸的缺失;小的氨基或羧基端延伸,例如氨基端延伸一个甲硫氨酸残基;小的连接肽,例如约20-25个残基长。
保守取代的实例是在下列氨基酸组内发生的取代:碱性氨基酸(如精氨酸、赖氨酸和组氨酸)、酸性氨基酸(如谷氨酸和天冬氨酸)、极性氨基酸(如谷氨酰胺、天冬酰胺)、疏水性氨基酸(如亮氨酸、异亮氨酸和缬氨酸)、芳香氨基酸(如苯丙氨酸、色氨酸和酪氨酸),以及小分子氨基酸(如甘氨酸、丙氨酸、丝氨酸、苏氨酸和甲硫氨酸)。通常不改变特定活性的那些氨基酸取代在本领域内是众所周知的,并且已由,例如,n.neurath和r.l.hill在1979年纽约学术出版社(academicpress)出版的《protein》中进行了描述。最常见的互换有ala/ser,val/ile,asp/glu,thu/ser,ala/thr,ser/asn,ala/val,ser/gly,tyr/phe,ala/pro,lys/arg,asp/asn,leu/ile,leu/val,ala/glu和asp/gly,以及它们相反的互换。
对于本领域的技术人员而言显而易见地,这种取代可以在对分子功能起重要作用的区域之外发生,而且仍产生活性多肽。对于由本发明的多肽,其活性必需的并因此选择不被取代的氨基酸残基,可以根据本领域已知的方法,如定点诱变或丙氨酸扫描诱变进行鉴定(如参见,cunningham和wells,1989,science244:1081-1085)。后一技术是在分子中每一个带正电荷的残基处引入突变,检测所得突变分子的除草剂抗性活性,从而确定对该分子活性而言重要的氨基酸残基。底物-酶相互作用位点也可以通过其三维结构的分析来测定,这种三维结构可由核磁共振分析、结晶学或光亲和标记等技术测定(参见,如devos等,1992,science255:306-312;smith等,1992,j.mol.biol224:899-904;wlodaver等,1992,febsletters309:59-64)。
本发明中,编码磺酰脲类除草剂水解酶的氨基酸序列包括但不限于本发明序列表中涉及的序列,与其具有一定同源性的氨基酸序列也包括在本发明中。这些序列与本发明序列类似性/相同性典型的大于60%,优选的大于75%,更优选的大于80%,甚至更优选的大于90%,并且可以大于95%。也可以根据更特定的相同性和/或类似性范围定义本发明的优选的多核苷酸和蛋白质。例如与本发明示例的序列有49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的相同性和/或类似性。
本发明中所述调控序列包括但不限于启动子、转运肽、终止子、增强子、前导序列、内含子以及其它可操作地连接到所述磺酰脲类除草剂水解酶基因的调节序列。
所述启动子为植物中可表达的启动子,所述的“植物中可表达的启动子”是指确保与其连接的编码序列在植物细胞内进行表达的启动子。植物中可表达的启动子可为组成型启动子。指导植物内组成型表达的启动子的示例包括但不限于,来源于花椰菜花叶病毒的35s启动子、玉米ubi启动子、水稻gos2基因的启动子等。备选地,植物中可表达的启动子可为组织特异的启动子,即该启动子在植物的一些组织内如在绿色组织中指导编码序列的表达水平高于植物的其他组织(可通过常规rna试验进行测定),如pep羧化酶启动子。备选地,植物中可表达的启动子可为创伤诱导启动子。创伤诱导启动子或指导创伤诱导的表达模式的启动子是指当植物经受机械或由昆虫啃食引起的创伤时,启动子调控下的编码序列的表达较正常生长条件下有显著提高。创伤诱导启动子的示例包括但不限于,马铃薯和西红柿的蛋白酶抑制基因(pinⅰ和pinⅱ)和玉米蛋白酶抑制基因(mpi)的启动子。
所述转运肽(又称分泌信号序列或导向序列)是指导转基因产物到特定的细胞器或细胞区室,对受体蛋白质来说,所述转运肽可以是异源的,例如,利用编码叶绿体转运肽序列靶向叶绿体,或者利用‘kdel’保留序列靶向内质网,或者利用大麦植物凝集素基因的ctpp靶向液泡。
所述前导序列包含但不限于,小rna病毒前导序列,如emcv前导序列(脑心肌炎病毒5’非编码区);马铃薯y病毒组前导序列,如mdmv(玉米矮缩花叶病毒)前导序列;人类免疫球蛋白质重链结合蛋白质(bip);苜蓿花叶病毒的外壳蛋白质mrna的不翻译前导序列(amvrna4);烟草花叶病毒(tmv)前导序列。
所述增强子包含但不限于,花椰菜花叶病毒(camv)增强子、玄参花叶病毒(fmv)增强子、康乃馨风化环病毒(cerv)增强子、木薯脉花叶病毒(csvmv)增强子、紫茉莉花叶病毒(mmv)增强子、夜香树黄化曲叶病毒(cmylcv)增强子、木尔坦棉花曲叶病毒(clcumv)、鸭跖草黄斑驳病毒(coymv)和花生褪绿线条花叶病毒(pclsv)增强子。
对于单子叶植物应用而言,所述内含子包含但不限于,玉米hsp70内含子、玉米泛素内含子、adh内含子1、蔗糖合酶内含子或水稻act1内含子。对于双子叶植物应用而言,所述内含子包含但不限于,cat-1内含子、pkannibal内含子、piv2内含子和“超级泛素”内含子。
所述终止子可以为在植物中起作用的适合多聚腺苷酸化信号序列,包括但不限于,来源于农杆菌(agrobacteriumtumefaciens)胭脂碱合成酶(nos)基因的多聚腺苷酸化信号序列、来源于蛋白酶抑制剂ⅱ(pinⅱ)基因的多聚腺苷酸化信号序列、来源于豌豆ssrubiscoe9基因的多聚腺苷酸化信号序列和来源于α-微管蛋白(α-tubulin)基因的多聚腺苷酸化信号序列。
本发明中所述“有效连接”表示核酸序列的联结,所述联结使得一条序列可提供对相连序列来说需要的功能。在本发明中所述“有效连接”可以为将启动子与感兴趣的序列相连,使得该感兴趣的序列的转录受到该启动子控制和调控。当感兴趣的序列编码蛋白并且想要获得该蛋白的表达时“有效连接”表示:启动子与所述序列相连,相连的方式使得得到的转录物高效翻译。如果启动子与编码序列的连接是转录物融合并且想要实现编码的蛋白的表达时,制造这样的连接,使得得到的转录物中第一翻译起始密码子是编码序列的起始密码子。备选地,如果启动子与编码序列的连接是翻译融合并且想要实现编码的蛋白的表达时,制造这样的连接,使得5’非翻译序列中含有的第一翻译起始密码子与启动子相连结,并且连接方式使得得到的翻译产物与编码想要的蛋白的翻译开放读码框的关系是符合读码框的。可以“有效连接”的核酸序列包括但不限于:提供基因表达功能的序列(即基因表达元件,例如启动子、5’非翻译区域、内含子、蛋白编码区域、3’非翻译区域、聚腺苷化位点和/或转录终止子)、提供dna转移和/或整合功能的序列(即t-dna边界序列、位点特异性重组酶识别位点、整合酶识别位点)、提供选择性功能的序列(即抗生素抗性标记物、生物合成基因)、提供可计分标记物功能的序列、体外或体内协助序列操作的序列(即多接头序列、位点特异性重组序列)和提供复制功能的序列(即细菌的复制起点、自主复制序列、着丝粒序列)。
本发明中所述“宿主细胞”是指在宿主细胞基因组中,或者在独立于宿主细胞的基因组而自主复制的染色体外载体中含有目标重组核酸的细胞。宿主细胞可以是任何细胞类型。
本发明中所述“转化”是指将dna引入细胞,从而dna以染色体外元件或染色体整合体形式维持在细胞内。
多核苷酸可以被整合到宿主细胞的基因组中,或者存在于在宿主细胞内自主复制的载体上。
本发明中,将外源dna导入植物,如将编码所述磺酰脲类除草剂水解酶的基因或表达盒或重组载体导入植物细胞,常规的转化方法包括但不限于,农杆菌介导的转化、微量发射轰击、直接将dna摄入原生质体、电穿孔或晶须硅介导的dna导入。
能使用不同农杆菌菌株,包括但不限于根癌农杆菌和毛根农杆菌。优选使用可转化态菌株,合适根癌农杆菌菌株包括菌株a208、菌株eha101、lba4404。合适毛根农杆菌包括菌株k599。可转化态农杆菌载体的构建为本领域所熟知的。
在选择剂存在的情况下培养转化的植物细胞。优选地,用磺酰脲类除草剂水解酶(sule)基因转化,并且转化的基因在磺酰脲类除草剂存在的情况下培养。在含有磺酰脲类除草剂为选择剂的培养基中,sule基因转化的植物细胞选择性生长。
含有异源核酸(即含有按照本发明的方法转化的细胞或组织)的转基因植物以及通过该转基因植物产生的种子和后代是本发明所涉及的。将转化的细胞培养成有用栽培品种的方法是本领域技术人员公知的。植物组织体外培养技术和整个植株再生技术也是公知的。相应的,所述“种子”包括这些转化植物的种子以及转化植物后代产生的种子。所述“植物”不仅包括转化和再生的植物,还包括通过本发明的方法产生的转化和再生植物的后代。
可以从本发明的方法产生的植物中筛选成功的转化植物。为了开发改良的植物和种子品系,可以持续地筛选和选择本发明再生植物的种子和子代植物以使转基因和整合的核酸序列持续存在。因此,可以将所需要的转基因核酸序列移入(即渐渗或交配)其它的遗传品系如某些原种或商业上有用的品系或品种中。渐渗目的基因进入遗传植物品系的方法可通过本领域公知的多种技术来实现,包括通过传统的育种、原生质体融合、细胞核转移以及染色体转移。育种方法和技术也是本领域公知的。根据本发明获得的转基因植物和自交系可用于生产商业上有价值的杂交植物和农作物。
本发明提供了一种提高大豆转化效率的方法,具有以下优点:
1、与现有技术将选择剂配置于培养基中不同,本发明首次提出在植物转化过程中以外施的方式(特别是滴加)将选择剂(如苯磺隆)加入增殖培养基和分化培养基中,其后代阳性植株率和转化效率显著提高,为在植物转化过程中使用选择剂提供了新的思路。
2、优化了筛选体系。本发明采用农杆菌介导的转化方法,不仅提供了磺酰脲类除草剂作为选择剂时新的使用方式和筛选方式,并优化了有效的筛选浓度范围,从而获得了对als抑制剂具有耐受性的转基因植株。
3、转化效率高,降低转化成本。本发明以磺酰脲类除草剂作为选择剂时,以优化的筛选浓度通过外施的方式(特别是滴加)加入选择剂进行筛选,其后代获得阳性植株的比例显著升高,转化效率可高达20%以上,同时以外施的方式(特别是滴加)可以使选择剂得到有效利用,降低了植物遗传转化的成本。
4、商业价值高。磺酰脲类除草剂为内吸型除草剂,本发明获得的转基因植株对磺酰脲类除草剂的耐受性较高,且其后代能稳定遗传,可直接开发成耐受磺酰脲类作物用于产品的开发。
下面通过附图和实施例,对本发明的技术方案做进一步的详细描述。
附图说明
图1为本发明提高大豆转化效率的方法的重组克隆载体dbn01-t构建流程图;
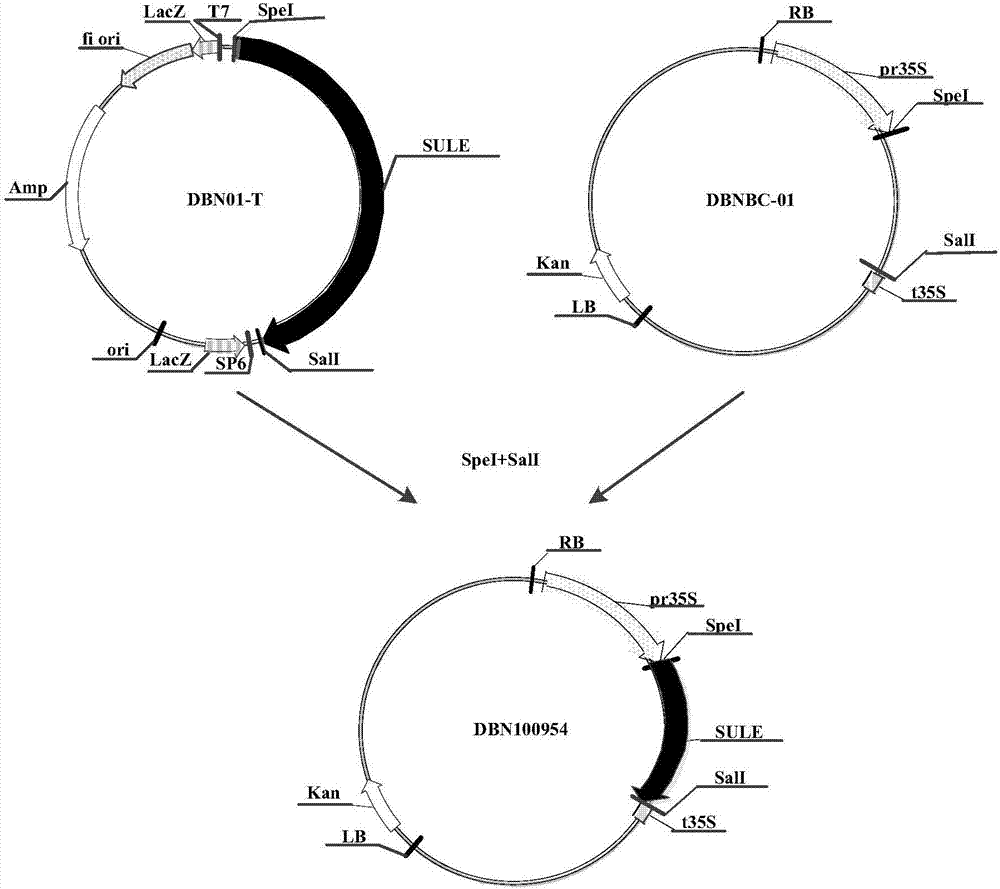
图2为本发明提高大豆转化效率的方法的重组表达载体dbn100954构建流程图;
图3为本发明提高大豆转化效率的方法的转化大豆组织的效果图。
具体实施方式
下面通过具体实施例进一步说明本发明提高大豆转化效率的方法的技术方案。
第一实施例、重组表达载体的构建及重组表达载体转化农杆菌
1、构建含有目的基因的重组克隆载体
将sule核苷酸序列连入克隆载体pgem-t(promega,madison,usa,cat:a3600)上,操作步骤按promega公司产品pgem-t载体说明书进行,得到重组克隆载体dbn01-t,其构建流程如图1所示(其中,amp表示氨苄青霉素抗性基因;f1表示噬菌体f1的复制起点;lacz为lacz起始密码子;sp6为sp6rna聚合酶启动子;t7为t7rna聚合酶启动子;sule为磺酰脲类除草剂水解酶基因核苷酸序列(seqidno:1);mcs为多克隆位点)。
然后将重组克隆载体dbn01-t用热激方法转化大肠杆菌t1感受态细胞(transgen,beijing,china,cat:cd501),其热激条件为:50μl大肠杆菌t1感受态细胞、10μl质粒dna(重组克隆载体dbn01-t),42℃水浴30秒;37℃振荡培养1小时(100rpm转速下摇床摇动),在表面涂有iptg(异丙基硫代-β-d-半乳糖苷)和x-gal(5-溴-4-氯-3-吲哚-β-d-半乳糖苷)的氨苄青霉素(100mg/l)的lb平板(胰蛋白胨10g/l,酵母提取物5g/l,nacl10g/l,琼脂15g/l,用naoh调ph至7.5)上生长过夜。挑取白色菌落,在lb液体培养基(胰蛋白胨10g/l,酵母提取物5g/l,nacl10g/l,氨苄青霉素100mg/l,用naoh调ph至7.5)中于温度37℃条件下培养过夜。碱法提取其质粒:将菌液在12000rpm转速下离心1min,去上清液,沉淀菌体用100μl冰预冷的溶液i(25mmtris-hcl,10mmedta(乙二胺四乙酸),50mm葡萄糖,ph8.0)悬浮;加入200μl新配制的溶液ii(0.2mnaoh,1%sds(十二烷基硫酸钠)),将管子颠倒4次,混合,置冰上3-5min;加入150μl冰冷的溶液iii(3m醋酸钾,5m醋酸),立即充分混匀,冰上放置5-10min;于温度4℃、转速12000rpm条件下离心5min,在上清液中加入2倍体积无水乙醇,混匀后室温放置5min;于温度4℃、转速12000rpm条件下离心5min,弃上清液,沉淀用浓度(v/v)为70%的乙醇洗涤后晾干;加入30μl含rnase(20μg/ml)的te(10mmtris-hcl,1mmedta,ph8.0)溶解沉淀;于温度37℃下水浴30min,消化rna;于温度-20℃保存备用。
提取的质粒经apai和ecorv酶切鉴定后,对阳性克隆进行测序验证,结果表明重组克隆载体dbn01-t中插入的sule基因序列为序列表中seqidno:1所示的核苷酸序列。
2、构建含有目的基因的重组表达载体
用限制性内切酶spei和sali分别酶切重组克隆载体dbn01-t和表达载体dbnbc-01(载体骨架:pcambia2301(cambia机构可以提供)),将切下的sule基因序列插到表达载体dbnbc-01的spei和sali位点之间,利用常规的酶切方法构建载体是本领域技术人员所熟知的,构建成重组表达载体dbn100954,其构建流程如图2所示(kan:卡那霉素基因;rb:右边界;pr35s:花椰菜花叶病毒35s基因的启动子(seqidno:3);sule:磺酰脲类除草剂水解酶基因核苷酸序列(seqidno:1,其氨基酸序列如seqidno:2所示);t35s:花椰菜花叶病毒35s基因的终止子(seqidno:4);lb:左边界)。
将重组表达载体dbn100954用热激方法转化大肠杆菌t1感受态细胞,其热激条件为:50μl大肠杆菌t1感受态细胞、10μl质粒dna(重组表达载体dbn100954),42℃水浴30秒;37℃振荡培养1小时(200rpm转速下摇床摇动);然后在含50mg/l卡那霉素(kanamycin)的lb固体平板(胰蛋白胨10g/l,酵母提取物5g/l,nacl10g/l,琼脂15g/l,用naoh调ph至7.5)上于温度37℃条件下培养12小时,挑取白色菌落,在lb液体培养基(胰蛋白胨10g/l,酵母提取物5g/l,nacl10g/l,卡那霉素50mg/l,用naoh调ph至7.5)中于温度37℃条件下培养过夜。碱法提取其质粒。将提取的质粒用限制性内切酶spei和sali酶切后鉴定,并将阳性克隆进行测序鉴定,结果表明重组表达载体dbn100954在spei和sali位点间的核苷酸序列为序列表中seqidno:1所示核苷酸序列,即sule核苷酸序列。
3、重组表达载体转化农杆菌
对己经构建正确的重组表达载体dbn100954用液氮法转化到农杆菌lba4404(invitrgen,chicago,usa,cat:18313-015)中,其转化条件为:100μl农杆菌lba4404、3μl质粒dna(重组表达载体);置于液氮中10分钟,37℃温水浴10分钟;将转化后的农杆菌lba4404接种于lb试管中于温度28℃、转速为200rpm条件下培养2小时,涂于含50mg/l的利福平(rifampicin)和100mg/l的卡那霉素(kanamycin)的lb平板上直至长出阳性单克隆,挑取单克隆培养并提取其质粒,用限制性内切酶apali和ecorv酶切后进行酶切验证,结果表明重组表达载体dbn100954结构完全正确。
第二实施例、转基因大豆植株的获得
大豆种子消毒:取完全干透的成熟大豆种子(中黄13)放在培养皿中,数量约占培养皿的1/3。将其放在通风橱的干燥器中,干燥器中放置250ml大烧杯,里面装有120ml次氯酸钠。沿着烧杯壁逐滴加入6ml浓盐酸,关闭干燥器并密封盖好,同时关闭通风橱的玻璃,使大豆种子暴露于通风橱内的氯气中灭菌3小时;盖好培养皿盖后将其拿出摇晃2-3分钟。重复上述过程一次,并将装有大豆种子的培养皿放置通风橱过夜灭菌。
大豆种子萌发:将消毒后的大豆种子15粒接种在大豆萌发培养基(b5盐3.1g/l、b5维他命、蔗糖20g/l、琼脂8g/l,ph5.6)上进行萌发,培养条件为:温度25±1℃,光周期为16/8h,大豆种子的种脐一侧插入培养基,接种后用保鲜膜将培养皿包好。
在本实施例的萌发培养基中,b5盐还可以为n6盐(浓度为3.95g/l)或ms盐(浓度为4.3g/l),蔗糖的浓度可以为5-100g/l;上述成分均可以在其浓度范围内进行任意组合,但以所述萌发培养基(b5盐3.1g/l、b5维他命、蔗糖20g/l、琼脂8g/l,ph5.6)为优选。
预处理外植体:萌发1天后,去除1片子叶和第1片真叶,将该裸露的分生组织接种到含有细胞分裂素的预处理培养基(ms盐4.3g/l、b5维他命、蔗糖20g/l、琼脂8g/l、2-吗啉乙磺酸(mes)4g/l、玉米素(zt)2mg/l、6-苄基腺嘌呤(6-bap)1mg/l、乙酰丁香酮(as)40mg/l,ph5.3),在此步骤中加入了细胞分裂素,使分生组织区的细胞处于活跃状态,同时在培养基还加入了as以促进外源基因的整合,预处理2-5天,优选为3天,将预处理后的分生组织块用解剖刀的刀背进行创伤(至少3刀,优选为5刀),创伤后进行超声波处理2-5分钟,优选为3分钟。
在本实施例的预处理培养基中,ms盐还可以为n6盐(浓度为3.95g/l)或b5盐(浓度为3.1g/l),蔗糖的浓度可以为5-100g/l,mes的浓度可以为0.1-5g/l,zt的浓度可以为0.1-5mg/l,6-bap的浓度可以为0.1-5g/l,as的浓度可以为10-50mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述预处理培养基(ms盐4.3g/l、b5维他命、蔗糖20g/l、琼脂8g/l、mes4g/l、zt2mg/l、6-bap1mg/l、as40mg/l,ph5.3)为优选。
农杆菌菌液的制备:将农杆菌菌株从-80℃冰箱中取出,挑取含有dbn100954的农杆菌单菌落,用枪头在加有卡那霉素(kanamycin)的固体yp培养板(酵母提取物5g/l、蛋白胨10g/l、氯化钠5g/l、琼脂8g、卡那霉素25mg/l,ph7.0)上划线,在28℃黑暗条件下培养2-3天。刮取yp培养板上的菌斑,在新的yp培养板上再培养1天;刮取培养3-4天的菌落,放入装有30ml侵染液(即侵染培养基(ms盐2.15g/l、b5维他命、蔗糖20g/l、葡萄糖10g/l、as40mg/l、mes4g/l、zt2mg/l,ph5.3))的50ml离心管中,不断摇晃以使菌体完全稀释在侵染液中,将稀释好的农杆菌菌液倒入装有150ml侵染液的250ml玻璃瓶(灭菌)中,定容,将农杆菌菌液的浓度调整为od660=0.5-0.8,待用。
在本实施例的侵染培养基中,ms盐还可以为n6盐(浓度为3.95g/l)或b5盐(浓度为3.1g/l),蔗糖的浓度可以为5-100g/l,葡萄糖的浓度可以为5-100g/l,as的浓度可以为10-50mg/l,mes的浓度可以为0.1-5g/l,zt的浓度可以为0.1-5mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述侵染培养基(ms盐2.15g/l、b5维他命、蔗糖20g/l、葡萄糖10g/l、as40mg/l、mes4g/l、zt2mg/l,ph5.3)为优选。
农杆菌侵染大豆分生组织块:将农杆菌菌液(10-15ml)接触上述超声波处理后的大豆分生组织块的子叶节组织一起放置至少3小时,优选为5小时;侵染结束后将农杆菌菌液吸出,并用滤纸将附着在大豆分生组织块上的农杆菌菌液彻底吸干。
农杆菌与大豆分生组织块共培养:将吸干农杆菌菌液的分生组织块转移到共培养培养基(ms盐4.3g/l、b5维他命、蔗糖20g/l、葡萄糖10g/l、mes4g/l、zt2mg/l、琼脂8g/l,ph5.6)上,培养基中放置一张滤纸,并且子叶近轴面向上,每皿15粒,在22℃恒温暗培养箱中共培养2-5天,优选为3天。
在本实施例上述共培养培养基中,蔗糖的浓度可以为5-100g/l,葡萄糖的浓度可以为5-100g/l,mes的浓度可以为0.1-5g/l,zt的浓度可以为0.1-5mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述共培养培养基(ms盐4.3g/l、b5维他命、蔗糖20g/l、葡萄糖10g/l、mes4g/l、zt2mg/l、琼脂8g/l,ph5.6)为优选。
大豆分生组织块的恢复:将共培养后的分生组织块的伸长的胚轴切掉,然后将切掉胚轴后的分生组织块转移到恢复培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、zt2mg/l、琼脂8g/l、头孢霉素150mg/l、谷氨酸100mg/l、天冬氨酸100mg/l,ph5.6)上,恢复2-5天,优选为3天,以消除农杆菌并为侵染细胞提供恢复期。
在本实施例上述恢复培养基中,mes的浓度可以为0.1-5g/l,蔗糖的浓度可以为5-100g/l,zt的浓度可以为0.1-5mg/l,头孢霉素100-300mg/l,谷氨酸50-200mg/l,天冬氨酸50-200mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述恢复培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、zt2mg/l、琼脂8g/l、头孢霉素150mg/l、谷氨酸100mg/l、天冬氨酸100mg/l,ph5.6)为优选。
大豆分生组织块的筛选:恢复期结束后,将分生组织块转移到无选择剂(苯磺隆)的60ml增殖培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、6-bap1mg/l、琼脂8g/l、头孢霉素150mg/l、谷氨酸100mg/l、天冬氨酸100mg/l,ph5.6)上,能使大豆的分生组织块快速的生长,在温度24℃、光周期(光/暗)16:8的条件下筛选培养5-9天,优选为7天。上述大豆分生组织块增殖5-9天后,分别按每毫升增殖培养基外施1、3、5和7μg苯磺隆除草剂(95%苯磺隆可湿性粉剂)的浓度,用移液器向所述增殖培养基中滴加稀释后的苯磺隆除草剂溶液,从而导致转化的细胞选择性生长,在温度24℃、光周期(光/暗)16:8的条件下筛选培养2周。
在本实施例所述增殖培养基中,b5盐还可以为n6盐(浓度为3.95g/l)或ms盐(浓度为4.3g/l),mes的浓度可以为0.1-5g/l,蔗糖的浓度可以为5-100g/l,6-bap的浓度可以为0.1-5mg/l,头孢霉素100-300mg/l,谷氨酸50-200mg/l,天冬氨酸50-200mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述增殖培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、6-bap1mg/l、琼脂8g/l、头孢霉素150mg/l、谷氨酸100mg/l、天冬氨酸100mg/l,ph5.6)为优选。
大豆抗性组织块再生成植物:分别将上述4种浓度苯磺隆筛选处理后的抗性组织块从含苯磺隆的所述增殖培养基中取出,切掉其上死去的组织和附着在上面的子叶,将抗性组织块转移到(斜插入)无选择剂(苯磺隆)的b5分化培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、zt1mg/l、琼脂8g/l、头孢霉素150mg/l、谷氨酸50mg/l、天冬氨酸50mg/l、赤霉素1mg/l、生长素1mg/l,ph5.6)上,在温度24℃、光周期(光/暗)16:8的条件下培养分化14-20天,优选为18天。上述4种筛选处理后大豆抗性组织块分化生长14-20天后,分别按每毫升b5分化培养基外施3、5和7μg苯磺隆除草剂(95%苯磺隆可湿性粉剂)的浓度,用移液器向所述b5分化培养基中滴加稀释后的苯磺隆除草剂溶液,在温度24℃、光周期(光/暗)16:8的条件下继续筛选培养直到抗性组织块(转化的细胞)再生成植物或已再生的植株能够存活。
在本实施例上述b5分化培养基中,mes的浓度可以为0.1-5g/l,蔗糖的浓度可以为5-100g/l,zt的浓度可以为0.1-5mg/l,头孢霉素100-300mg/l,谷氨酸50-200mg/l、天冬氨酸50-200mg/l,赤霉素0.1-5mg/l,生长素0.1-5mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述b5分化培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、zt1mg/l、琼脂8g/l、头孢霉素150mg/l、谷氨酸50mg/l、天冬氨酸50mg/l、赤霉素1mg/l、生长素1mg/l,ph5.6)为优选。
在本实施例上述大豆分生组织块的筛选步骤和大豆抗性组织块再生成植物步骤中,按每毫升增殖培养基或b5分化培养基外施苯磺隆除草剂(95%苯磺隆可湿性粉剂)的浓度,向增殖培养基或b5分化培养基中滴加稀释后的苯磺隆除草剂溶液的示例如表1所示。对增殖后的大豆分生组织块进行苯磺隆除草剂耐受性筛选,12种处理的具体信息如表2所示,每个处理的筛选结果如表3所示,每个处理设3次重复。
表1、外施苯磺隆除草剂的示例
注:苯磺隆除草剂滴加质量(μg/皿)=稀释后苯磺隆除草剂溶液浓度(μg/ml)×稀释后苯磺隆除草剂溶液的滴加体积(ml);苯磺隆除草剂滴加浓度(μg/ml培养基)=苯磺隆除草剂滴加质量(μg/皿)/培养基体积(ml/皿)。
表2、对大豆分生组织块进行12种不同苯磺隆筛选处理的具体信息
将上述12种处理分化出来的小苗转移到b5生根培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、琼脂8g/l、头孢霉素150mg/l、吲哚-3-丁酸(iba)1mg/l,ph5.6)上,在25℃下培养至约10cm高,移至温室培养至结实。在温室中,每天于26℃下培养16小时,再于20℃下培养8小时,可以获得转基因植株。
在本实施例上述b5生根培养基中,mes的浓度可以为0.1-5g/l,蔗糖的浓度可以为5-100g/l,头孢霉素100-300mg/l,iba的浓度可以为0.1-5mg/l;上述成分均可以在其浓度范围内进行任意组合,但以所述b5生根培养基(b5盐3.1g/l、b5维他命、mes1g/l、蔗糖30g/l、琼脂8g/l、头孢霉素150mg/l、iba1mg/l,ph5.6)为优选。
在本实施例中,除苯磺隆外,其它均采用优选方案。
第三实施例、用taqman验证转入sule核苷酸序列的大豆植株
分别取上述12种处理后的转入sule核苷酸序列的大豆植株的叶片约100mg作为样品,用qiagen的dneasyplantmaxikit提取其基因组dna,通过taqman探针荧光定量pcr方法检测sule基因的拷贝数。同时以野生型大豆植株作为对照,按照上述方法进行检测分析。实验设3次重复,取平均值。
检测sule基因拷贝数的具体方法如下:
步骤11、分别取上述12种处理后的转入sule核苷酸序列的大豆植株和野生型大豆植株的叶片各100mg,分别在研钵中用液氮研成匀浆,每个样品取3个重复;
步骤12、使用qiagen的dneasyplantminikit提取上述样品的基因组dna,具体方法参考其产品说明书;
步骤13、用nanodrop2000(thermoscientific)测定上述样品的基因组dna浓度;
步骤14、调整上述样品的基因组dna浓度至同一浓度值,所述浓度值的范围为80-100ng/μl;
步骤15、采用taqman探针荧光定量pcr方法鉴定样品的拷贝数,以经过鉴定已知拷贝数的样品作为标准品,以野生型大豆植株的样品作为对照,每个样品3个重复,取其平均值;荧光定量pcr引物和探针序列分别是:
以下引物和探针用来检测sule核苷酸序列:
引物1:tgggagaggaaggggtaacat,如序列表中seqidno:5所示;
引物2:tatctctcacccaggcacctt,如序列表中seqidno:6所示;
探针1:acggacctttcggacagttggagga,如序列表中seqidno:7所示;
pcr反应体系为:
所述50×引物/探针混合物包含1mm浓度的每种引物各45μl,100μm浓度的探针50μl和860μl1×te缓冲液,并且在4℃,贮藏在琥珀试管中。
pcr反应条件为:
利用sds2.3软件(appliedbiosystems)分析数据。
实验结果表明,sule核苷酸序列整合到部分所检测的大豆植株的染色体组(即阳性植株)中,12种处理的实验结果如表3和图3所示。
表3、12种处理的实验结果
注:阳性植株率(%)=阳性植株数(株)/出苗数(株)×100%;
转化效率(%)=阳性植株数(株)/起始萌发大豆数量(个)×100%。
由表3可得,在转化过程的大豆分生组织块的筛选步骤和大豆抗性组织块再生成植物步骤中,按每毫升增殖培养基或b5分化培养基外施苯磺隆除草剂(95%苯磺隆可湿性粉剂)的浓度,通过向增殖培养基或b5分化培养基中滴加稀释后的苯磺隆除草剂溶液的方式进行筛选,上述12种处理均能获得较高的阳性植株率和转化效率,尤以处理6为佳,转化效率可高达20%以上。
综上所述,与现有技术将选择剂配置于培养基中不同,本发明首次提出在植物转化过程中以外施的方式(特别是滴加)将选择剂(如苯磺隆)加入增殖培养基和分化培养基中,其后代阳性植株率和转化效率显著提高,为在植物转化过程中使用选择剂提供了新的思路;本发明采用农杆菌介导的转化方法,不仅提供了磺酰脲类除草剂作为选择剂时新的使用方式和筛选方式,并优化了有效的筛选浓度范围,从而获得了对als抑制剂具有耐受性的转基因植株;本发明以磺酰脲类除草剂作为选择剂时,以优化的筛选浓度通过外施的方式(特别是滴加)加入选择剂进行筛选,其后代获得阳性植株的比例显著升高,转化效率可高达20%以上,同时以外施的方式(特别是滴加)可以使选择剂得到有效利用,降低了植物遗传转化的成本;同时本发明使用磺酰脲类除草剂水解酶基因作为选择标记物的转化获得的转基因植株商业价值高、抗性好且遗传稳定。
最后所应说明的是,以上实施例仅用以说明本发明的技术方案而非限制,尽管参照较佳实施例对本发明进行了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的精神和范围。
sequencelisting
<110>北京大北农生物技术有限公司
<120>提高大豆转化效率的方法
<130>dbnbc121
<160>7
<170>patentinversion3.5
<210>1
<211>1110
<212>dna
<213>artificialsequence
<220>
<223>磺酰脲类除草剂水解酶基因核苷酸序列
<400>1
atggagactgataacgtggaactcgcccaatctaagagaaaggtggtgctggctgaacaa60
gggtcattttacatagggggtaggactgttactggtcctggcaagtttgatccatccaaa120
cctgtgataccctacagtaacgaaggagcaacattctatattaaccaaatgtatgttaac180
ttccaggccccagtgagacctaggggacttccattggttttctggcatggaggtggcttg240
actggtcacatctgggagtctacacctgacggcagacccgggtttcaaaccctcttcgtt300
caggataggcataccgtgtacactattgaccaacctgggagaggaaggggtaacatccca360
acttttaacggacctttcggacagttggaggaagagagtattgttaacactgtgacagga420
aattcttcaaaggaaggtgcctgggtgagagataggcttggccctgctcccgggcaattt480
ttcgagaactctcagtttcctagaggctatgaagacaattactttaaggagatgggattc540
agcccatctatatccagtgatgaaattgttgacgctgttgtgaaactcgtgacccatatt600
ggtccttgtgttctggtgactcactcagcatccggcgttcttgggatgagagtggctaca660
cacgcaaagaatgttaggggaattgtggcctatgaaccagctacctcaatcttccccaag720
ggaaaagttccagagataccacctctcgctgataagaaaagccaaatctttcccccattc780
gaaatacaggagtcttactttaagaaacttgccaagattccaatccaatttgttttcgga840
gataacatccccaagaatccaaaatcagcatattggttcctggactggtggagagtgaca900
agatacgcacatagtctcagcctggaggccataaacaaattggggggacaagcttccctt960
ttggatcttcctactgcaggattgagaggtaatacacactttcccttcaccgataggaac1020
aatgttcaggtggcttctctcctgtcagactttctgggtaaacacggtctggatcaaaat1080
gagagcaaactcgccgccgccctggaatga1110
<210>2
<211>369
<212>prt
<213>artificialsequence
<220>
<223>磺酰脲类除草剂水解酶基因氨基酸序列
<400>2
metgluthraspasnvalgluleualaglnserlysarglysvalval
151015
leualagluglnglyserphetyrileglyglyargthrvalthrgly
202530
proglylyspheaspproserlysprovalileprotyrserasnglu
354045
glyalathrphetyrileasnglnmettyrvalasnpheglnalapro
505560
valargproargglyleuproleuvalphetrphisglyglyglyleu
65707580
thrglyhisiletrpgluserthrproaspglyargproglyphegln
859095
thrleuphevalglnasparghisthrvaltyrthrileaspglnpro
100105110
glyargglyargglyasnileprothrpheasnglypropheglygln
115120125
leugluglugluserilevalasnthrvalthrglyasnserserlys
130135140
gluglyalatrpvalargaspargleuglyproalaproglyglnphe
145150155160
phegluasnserglnpheproargglytyrgluaspasntyrphelys
165170175
glumetglypheserproserileserseraspgluilevalaspala
180185190
valvallysleuvalthrhisileglyprocysvalleuvalthrhis
195200205
seralaserglyvalleuglymetargvalalathrhisalalysasn
210215220
valargglyilevalalatyrgluproalathrserilepheprolys
225230235240
glylysvalprogluileproproleualaasplyslysserglnile
245250255
pheproprophegluileglnglusertyrphelyslysleualalys
260265270
ileproileglnphevalpheglyaspasnileprolysasnprolys
275280285
seralatyrtrppheleuasptrptrpargvalthrargtyralahis
290295300
serleuserleuglualaileasnlysleuglyglyglnalaserleu
305310315320
leuaspleuprothralaglyleuargglyasnthrhispheprophe
325330335
thraspargasnasnvalglnvalalaserleuleuserasppheleu
340345350
glylyshisglyleuaspglnasngluserlysleualaalaalaleu
355360365
glu
<210>3
<211>530
<212>dna
<213>cauliflowermosaicvirus
<400>3
ccatggagtcaaagattcaaatagaggacctaacagaactcgccgtaaagactggcgaac60
agttcatacagagtctcttacgactcaatgacaagaagaaaatcttcgtcaacatggtgg120
agcacgacacgcttgtctactccaaaaatatcaaagatacagtctcagaagaccaaaggg180
caattgagacttttcaacaaagggtaatatccggaaacctcctcggattccattgcccag240
ctatctgtcactttattgtgaagatagtggaaaaggaaggtggctcctacaaatgccatc300
attgcgataaaggaaaggccatcgttgaagatgcctctgccgacagtggtcccaaagatg360
gacccccacccacgaggagcatcgtggaaaaagaagacgttccaaccacgtcttcaaagc420
aagtggattgatgtgatatctccactgacgtaagggatgacgcacaatcccactatcctt480
cgcaagacccttcctctatataaggaagttcatttcatttggagaggaca530
<210>4
<211>195
<212>dna
<213>cauliflowermosaicvirus
<400>4
ctgaaatcaccagtctctctctacaaatctatctctctctataataatgtgtgagtagtt60
cccagataagggaattagggttcttatagggtttcgctcatgtgttgagcatataagaaa120
cccttagtatgtatttgtatttgtaaaatacttctatcaataaaatttctaattcctaaa180
accaaaatccagtgg195
<210>5
<211>21
<212>dna
<213>artificialsequence
<220>
<223>引物1
<400>5
tgggagaggaaggggtaacat21
<210>6
<211>21
<212>dna
<213>artificialsequence
<220>
<223>引物2
<400>6
tatctctcacccaggcacctt21
<210>7
<211>25
<212>dna
<213>artificialsequence
<220>
<223>探针1
<400>7
acggacctttcggacagttggagga25
- 还没有人留言评论。精彩留言会获得点赞!