水稻OsbZIP86基因及其编码蛋白在干旱胁迫中的应用的制作方法
本发明属于生物基因工程领域,具体涉及水稻中osbzip86基因及其编码转录因子,同时涉及该基因两个转录本的生化功能研究和在转基因水稻中的抗旱性应用。转基因水稻在种子萌发过程中对aba不敏感,幼苗期受干旱诱导表达,且能够通过促进aba合成关键限速酶基因osnced3的表达,导致aba积累增加,响应干旱胁迫中的功能研究。
背景技术:
世界上重要的粮食作物是水稻。目前全世界超过50%的人口以大米为主食。从19世纪70年代开始,水稻在中国和东南亚通过杂交水稻从而提高其产量。由于全球气候变暖及环境破坏加剧,导致高温、干旱频频发生,土地盐碱化日益严重。这些非生物逆境已经严重影响水稻产量增加。近年来对于耐旱和耐盐碱农作物的分子作用机制研究及新品种分子选育一直是热点研究领域。水稻由于拥有较小的基因组、完整的基因组测序以及有效的遗传转化技术和巨大的遗传信息而作为模式植物,特别是为单子叶植物的研究提供了基础。
植物对干旱胁迫的响应是一个非常复杂的信号调控过程。以往大量的研究热点集中于各种转录因子(dreb、nac、bzip、锌指蛋白基因、myb、wrky、tify等)和一些功能性蛋白(lea等)在干旱胁迫中的生物学功能,并分离鉴定了很多与耐旱相关的基因。这些基因能有效的改善植物的耐旱性,从而增加农作物的产量。bzip转录因子在真核生物细胞内普遍存在,广泛参与了植物生长发育及各种生物、非生物逆境胁迫过程。
碱性亮氨酸拉链(bzip)是在植物应激反应和激素信号转导中具有重要作用的一个转录因子家族。已经证实bzip蛋白存在所有真核生物中,包括酵母、脊椎动物和植物。人类有56个bzip蛋白。拟南芥中大约有100个bzip序列在其基因组中被预测并被分为10个亚家族。而在水稻中总共有89个bzip基因,被分为11组。bzip蛋白由保守的bzip结构域定义,bzip结构域的长度为60至80个氨基酸。bzip蛋白结构呈l型,具有2个结构域,包括高度保守的dna结合碱性区和更多样化的亮氨酸拉链二聚化区,通常以二聚体或者四聚体的形式发挥作用,碱性区具有~16个氨基酸残基。最引人注目的基本区域是存在不变的n-x7-r/k基序。亮氨酸拉链由leu的七肽重复组成,由重复的基序组成几种氨基酸或其他疏水性氨基酸的模式。由于其独特的氨基酸组成,leu拉链倾向于在每个七肽中形成具有2个螺旋匝的两亲性螺旋。大多数bzip蛋白显示对acgt基序的高结合亲和力,其包括cacgtg(g盒),gacgtc(c盒),tacgta(a盒),aacgtt(t盒)和gcn4基序,即tga(g/c)tca。少量的bzip因子如osobf1也可以识别回文序列。然而,其他,包括lip19,oszip-2a和oszip-2b,不结合dna序列。相反,这些bzip蛋白与其他bzips形成异二聚体来调节转录活性。很多bzip蛋白,如osbzip52/risbz5,osareb1/osabf2,osabi5/oreb1,osbz8能与aba反应元件abre结合调控aba依赖性基因的表达参与植株对非生物胁迫的应激过程。
bzip蛋白形成转录的超家族是介导植物胁迫反应的因子。目前bzip蛋白在多种生物过程中起作用,包括病原体防御、对非生物胁迫的反应、种子发育和发芽、衰老和对水杨酸、茉莉酸和水稻中的脱落酸信号过程中起作用。这些转录因子可能通过水杨酸途径参与了水稻的生物胁迫或者通过aba途径在非生物胁迫、胁迫应激过程及在种子发育中起着重要作用。
本发明涉及转录因子osbzip86转录因子osbzip86受干旱诱导表达,转基因水稻osbzip86-ox中能够诱导aba合成关键限速酶基因osnced3的表达升高,增加aba合成积累,因此对此基因的研究为进一步丰富bzip家族在aba合成信号途径中的作用研究提供依据。转录因子osbzip86将增强水稻干旱胁迫的耐受性,转基因水稻在种子萌发过程中对aba不敏感,且能够通过增加aba合成积累响应干旱胁迫中的功能研究。因此为水稻耐旱分子育种提供更为丰富的基因遗传资源。
技术实现要素:
本发明的第一个目的是提供一个能增加水稻抗旱性以及影响种子发育的转录因子osbzip86及其该基因osbzip86的两个转录本,即:osbzip86-2和osbzip86-3的应用。
本发明的所阐述的水稻转录因子osbzip86,其氨基酸序列如序列表中seqidno.1和seqidno.2所示,其两个转录本的cdna阅读框核苷酸序列如序列表中seqidno.3和seqidno.4所示在提高植物对抗逆性中的应用,特别地,所述抗逆性是指抗旱胁迫。
本发明的另一目的是提供水稻转录因子基因osbzip86的两个转录本的转录激活活性功能和转录因子的亚细胞定位。
插入有osbzip86基因的cdna阅读框序列的酵母重组表达载体,所述osbzip86基因的cdna为如seqidno.3和seqidno.4所示的核苷酸序列,或为与seqidno.3和seqidno.4互补配对的核苷酸序列,或为编码序列氨基酸序列如seqidno.1和seqidno.2的核苷酸序列。
插入有osbzip86基因的cdna阅读框序列的gfp瞬时和稳定表达载体,所述osbzip86基因的cdna为如seqidno.3和seqidno.4所示的核苷酸序列,或为与seqidno.3和seqidno.4互补配对的核苷酸序列,或为编码序列氨基酸序列如seqidno.1和seqidno.2的核苷酸序列。
插入有osbzip86基因的cdna阅读框序列的超表达和干扰表达载体,所述osbzip86基因的cdna为如seqidno.3所示的核苷酸序列,或为与seqidno.3互补配对的核苷酸序列,或为编码序列氨基酸序列如seqidno.1的核苷酸序列。
本发明的另一目的是提供osbzip86基因在植物育种中的应用。
osbzip86基因,或权利要求2所述酵母重组表达载体,或权利要求3所述的gfp瞬时表达和稳定表达载体,或权利要求4所述的超表达和干扰表达载体在改良水稻对干旱的耐受性的遗传育种中的应用,所述osbzip86基因的cdna为如seqidno.3和seqidno.4所示的核苷酸序列,或为与seqidno.3和seqidno.4互补配对的核苷酸序列,或为编码序列氨基酸序列如seqidno.1和seqidno.2的核苷酸序列。
本发明利用酵母双杂交体系自激活原理,构建酵母目标载体并转化酵母。分析osbzip86-2和osbzip86-3的转录激活活性。
构建osbzip86-2::sgfp、osbzip86-3::sgfp的瞬时表达载体转化原生质体和稳定表达载体转化水稻愈伤,利用激光扫描共聚焦显微镜观察细胞。对照组中gfp蛋白在细胞内均有分布,且与mcherry的定位不完全一致。而瞬时表达和稳定表达的实验结果都明显表明osbzip86-2和osbzip86-3与mcherry共定位,且于细胞核中。
本发明的另一目的是一种将上述水稻转录因子基因osbzip86应用于植物基因工程中,得到osbzip86的超表达和干扰的水稻转基因植株,用于改变水稻对干旱的耐受性。
本发明公开了水稻转录因子osbzip86的干旱胁迫诱导表达特征,表明了该基因参与干旱胁迫的应答,论证了该基因为水稻应答干旱胁迫的重要基因,进一步延伸了该基因作为影响植物体内aba积累和响应干旱胁迫的候选功能基因。同时改变该基因的表达能影响植株对干旱的抗性。
本发明的另一目的是提供一种提高水稻的抗旱胁迫的生物制剂。
一种提高水稻的抗旱胁迫的生物制剂,其活性成分来源于权利要求3所述酵母重组表达载体或权利要求4所述的超表达和干扰表达载体,或其活性成分含有调控osbzip86基因表达的生物制品,所述osbzip86基因的cdna为如seqidno.3和seqidno.4所示的核苷酸序列,或为与seqidno.3和seqidno.4互补配对的核苷酸序列,或为编码序列氨基酸序列如seqidno.1和seqidno.2的核苷酸序列。
本发明的另一目的是提供一种调控水稻的抗旱胁迫的方法。
一种调控水稻的抗旱胁迫的方法,该方法包括调控osbzip86基因表达,所述osbzip86基因的cdna为如seqidno.3和seqidno.4所示的核苷酸序列,或为与seqidno.3和seqidno.4互补配对的核苷酸序列,或为编码序列氨基酸序列如seqidno.1和seqidno.2的核苷酸序列。
本发明构建osbzip86的超表达(osbzip86-ox)和干扰表达载体(osbzip86-rnai),并通过农杆菌eha105介导法的遗传转化方法,将超表达和干扰表达载体转化至正常粳稻品种zh11,最后获得osbzip86-ox和osbzip86-rnai的转基因水稻纯合体。经过多次重复逆境处理实验,osbzip86-ox表现在种子萌发过程中对aba不敏感,且诱导aba合成基因osnced3的表达,通过增加aba合成积累,响应干旱胁迫中的功能研究。osbzip86-rnai则表现相反。
本发明的有益效果如下:在本发明中,我们获得的一个水稻转录因子osbzip86并进行应用性探索,阐述了该基因以下应用方式:
(1)本发明的水稻转录因子osbzip86具有两个转录本,即:osbzip86-2和osbzip86-3。且通过酵母双杂交体系探究:osbzip86-3以及osbzip86-3△c片段具有转录激活活性。这有助于探究剪切产生不同的转录本的功能差异性研究。
(2)本发明涉及转录因子osbzip86受干旱诱导表达的特征,表明该基因具有干旱胁迫应答的基本功能,可将该基因应用于水稻针对干旱的遗传育种,改良水稻对干旱的耐受性。
(3)转录因子osbzip86将增强水稻干旱胁迫的耐受性,在转基因水稻中能够诱导aba合成关键限速酶基因osnced3的表达,且通过增加aba合成积累响应干旱胁迫中的功能研究,因此对此基因的研究为进一步丰富bzip家族在aba合成信号途径中的作用研究提供依据,为水稻耐旱分子育种提供更为丰富的基因遗传资源。
附图说明
图1示osbzip86基因的cdna阅读框序列的pcr扩增电泳分析,dna分子量:2000bp;1-4为osbzip86-2重复样;5-8为osbzip86-3重复样。
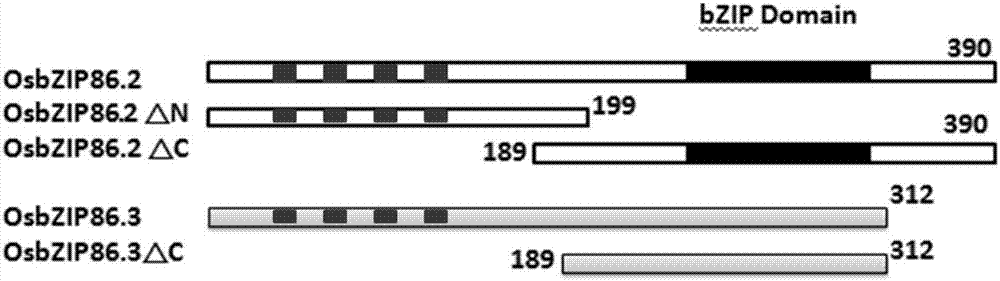
图2示osbzip86-2和osbzip86-3根据bzipdomain位置分段示意图。
图3示酵母双杂交自激活实验,osbzip86-3及osbzip86-3△c具有转录激活活性。
图4示osbzip86::sgfp原生质体瞬时表达具有明显的核定位。
图5示osbzip86::sgfp稳定表达的转基因水稻根系具有明显的核定位。
图6示osbzip86的表达显著的受到干旱诱导zh11水稻2周幼苗,不同时间脱水及复水处理osbzip86基因表达。
图7示osbzip86-ox转基因水稻种子萌发过程中对aba不敏感性,其中osbzip86-ox、osbzip86-rnai和zh11水稻种子于aba浓度为0、3、5、10μm的1/2ms中,处理10天;1:zh11;2:osbzip86-rnai;3:osbzip86-ox;ck:对照组。
图8示osbzip86-ox转基因水稻植株抗旱性更强,其中osbzip86-ox、osbzip86-rnai和zh11水稻2周幼苗,脱水处理8小时图;1:zh11;2:osbzip86-rnai;3:osbzip86-ox。
图9示aba合成基因osnced3在干旱胁迫osbzip86-ox转基因水稻中被诱导表达;osbzip86-ox、osbzip86-rnai和zh11水稻2周幼苗干旱处理0、1、2、3小时后osnecd3表达量图;ck:处理0小时对照组的放大图。
图10示干旱胁迫osbzip86-ox转基因水稻中aba含量升高;其中,osbzip86-ox、osbzip86-rnai和zh11水稻2周幼苗干旱处理4小时后aba含量图;ck:对照组。
具体实施方式
本发明描述了水稻转录因子osbzip86在生物工程方面的应用。本发明首先克隆了水稻转录因子osbzip86两个转录本的cdna阅读框全长。以正常粳稻品种zh11为材料,提取总rna,逆转录成cdna。以cdna为模板,克隆osbzip86-2和osbzip86-3的全长cdna。将osbzip86-2和osbzip86-3根据bzipdomain位置(图2)分为两段,扩增osbzip86全长以及片段序列,分别与载体pgbkt7连接,构建用于酵母pgbkt7的融合载体,之后将这一重组载体转化酵母,若其表达产生的bd(bindingdomain)单独与uas结合也可以引起下游报告基因的转录,那么此片段具有转录激活活性。此实验证明只有osbzip86-3和osbzip86-3△c具有转录活性。该基因可用于剪切产生不同的转录本功能差异性研究,以及作为转录因子的两个转录本在参与抗逆性信号途径中转录激活活性的功能差异性研究。
构建osbzip86-2::sgfp、osbzip86-3::sgfp的瞬时表达和稳定表达载体,瞬时表达载体转化水稻原生质体,利用激光扫描共聚焦显微镜zeiss-lsm780观察细胞。利用农杆菌eha105介导法的遗传转化方法,将稳定表达载体转化至正常粳稻品种zh11获得osbzip86-2::sgfp、osbzip86-3::sgfp的稳定表达转基因材料,以及负对照zh11,正对照puc18-gfp,待根生长至约2cm。利用激光扫描共聚焦显微镜zeiss-lsm750观察细胞。结果表明,对照gfp蛋白在细胞内均有分布,且与mcherry的定位不完全一致。而osbzip86-2和osbzip86-3则与mcherry共定位于细胞核中,与网站预测结果一致。稳定表达的转基因水稻也得到一致性结果。
构建osbzip86的超表达(osbzip86-ox)和干扰载体(osbzip86-rnai),并通过农杆菌eha105介导法的遗传转化方法,将超表达和干扰载体转化至正常粳稻品种zh11,最后获得osbzip86-ox和osbzip86-rnai的转基因水稻纯合体。进一步对zh11、osbzip86-rnai和osbzip86-ox的水稻种子进行aba、nacl的敏感性实验以及幼苗进行耐盐、耐旱实验。结果表明超表达osbzip86增强了水稻种子萌发时对aba的耐受性,表现出aba的不敏感性,而幼苗期进行干旱胁迫处理时,osbzip86-ox水稻幼苗对干旱耐受性及干旱条件下的保水性都有显著性增强,osbzip86-rnai水稻幼苗对干旱耐受性及干旱条件下都有显著性降低。说明osbzip86不仅对水稻的抗旱反应是必须的,过量表达会增强水稻的抗旱反应。同时发现干旱处理下osbzip86-ox中osnced3在1、2、3h都表现出显著性升高,而osbzip86-rnai中osnced3在1、2h时表现出显著下降趋势,本结果表明osbzip86可能直接参与osnced3的转录表达。osnced3基因是aba合成途径关键限速酶基因,而这个结果和aba含量测定结果表现出一致性。因此此基因可作为丰富bzip家族在aba合成信号途径中的作用做进一步研究,也有为水稻耐旱分子育种提供更丰富的基因遗传资源的潜力。
seqidno.1
氨基酸序列
>osbzip86-2protein
mgssgadaptktskasapqeqqppassstatpavypdwanfqgyppipphgffpspvasspqghpymwgaqpmippygtppppyvmyppgvyahpsmppgahpftpyamaspngnadptgttttaaaaaagetdgkssegkekspikrskgslgslnmitgknstehgktsgasangaisqsgesgsesssegseansqndshhkesgqeqdgevrssqngvsrspsqaklnqtmaimpmtssgpvpapttnlnigmdywantasstpaihgkatptaapgsmvpgeqwvqderelkrqrrkqsnresarrsrlrkqaeceelaqraevlkqentslrdevnrirkeydellsknsslkekledkqhktdeagvdnklqhsgddsqkkgn*.
seqidno.2
>osbzip86-3protein
mgssgadaptktskasapqeqqppassstatpavypdwanfqqgyppipphgffpspvasspqghpymwgaqpmippygtppppyvmyppgvyahpsmppgahpftpyamaspngnadptgttttaaaaaagetdgkssegkekspikrskgslgslnmitgknstehgktsgasangaisqsgesgsesssegseansqndshhkesgqeqdgevrssqngvsrspsqaklnqtmaimpmtssgpvpapttnlnigmdywantasstpaihgkatptaapgsmvpgeqwvqvcsispslqepctcniffpl*.
cdna阅读框核苷酸序列
seqidno.3
>osbzip86-3
atgggtagcagtggcgcagacgcaccgactaagacaagcaaggcatctgcacctcaggagcaacagccacctgctagttcaagcactgcaacaccggctgtttacccagattgggccaactttcagggatatcctccaattccaccacatggcttctttccatcacctgtggcgtcaagcccacagggtcatccttacatgtggggagctcagcctatgataccaccatatggaaccccaccaccaccatatgtcatgtatcctccaggagtatatgctcacccatctatgcctccggcgcacacccatttactccatatgccatggcctctccaaatggcaatgctgatcctactggaactacaactactgctgctgctgctgctgctggtgagacggatggcaaatcctctgaaggaaaagaaaaaagtcccattaaaagatctaaaggaagtttaggtagcttgaatatgattacagggaagaactctactgaacatggtaaaacctctggggcatcagctaatggagccatttctcaaagtggggaaagtggaagtgaaagttctagtgaaggaagtgaagcaaattctcagaatgattcacatcacaaggaaagtggacaagagcaagatggagaggttcgaagttcccagaatggtgtatcacgttcaccatcccaggcaaagttgaatcaaactatggcaatcatgccaatgacatcaagtggtccagtacctgctccaactacaaacttgaacataggaatggactactgggccaacacagctagctccactccagcaatacatggtaaagcaaccccaactgcagctccagggtctatggttccaggagagcagtgggtgcaggtttgttctattagcccttcccttcaagaaccgtgcacttgtaatattttcttcccgttgtag.
seqidno.4
>osbzip86-2
atgggtagcagtggcgcagacgcaccgactaagacaagcaaggcatctgcacctcaggagcaacagccacctgctagttcaagcactgcaacaccggctgtttacccagattgggccaactttcagggatatcctccaattccaccacatggcttctttccatcacctgtggcgtcaagcccacagggtcatccttacatgtggggagctcagcctatgataccaccatatggaaccccaccaccaccatatgtcatgtatcctccaggagtatatgctcacccatctatgcctccgggcgcacacccatttactccatatgccatggcctctccaaatggcaatgctgatcctactggaactacaactactgctgctgctgctgctgctggtgagacggatggcaaatcctctgaaggaaaagaaaaaagtcccattaaaagatctaaaggaagtttaggtagcttgaatatgattacagggaagaactctactgaacatggtaaaacctctggggcatcagctaatggagccatttctcaaagtggggaaagtggaagtgaaagttctagtgaaggaagtgaagcaaattctcagaatgattcacatcacaaggaaagtggacaagagcaagatggagaggttcgaagttcccagaatggtgtatcacgttcaccatcccaggcaaagttgaatcaaactatggcaatcatgccaatgacatcaagtggtccagtacctgctccaactacaaacttgaacataggaatggactactgggccaacacagctagctccactccagcaatacatggtaaagcaaccccaactgcagctccagggtctatggttccaggagagcagtgggtgcaggatgaacgggaactcaaaaggcagagaagaaaacaatccaacagggagtctgctcgcagatctaggttgcgtaagcaggctgaatgtgaggagttggctcaacgtgctgaagttttaaagcaagaaaatacttcacttagagatgaagtgaaccggatcagaaaggagtatgatgagcttctatcaaagaatagctcattaaaggaaaaacttgaggacaaacaacacaaaactgatgaggcaggagttgacaataagctgcaacattctggtgatgacagccagaaaaaaggaaactaa.
cdna阅读框核苷酸序列
seqidno.5
>osbzip86-3△c
tctagtgaaggaagtgaagcaaattctcagaatgattcacatcacaaggaaagtggacaagagcaagatggagaggttcgaagttcccagaatggtgtatcacgttcaccatcccaggcaaagttgaatcaaactatggcaatcatgccaatgacatcaagtggtccagtacctgctccaactacaaacttgaacataggaatggactactgggccaacacagctagctccactccagcaatacatggtaaagcaaccccaactgcagctccagggtctatggttccaggagagcagtgggtgcaggtttgttctattagcccttcccttcaagaaccgtgcacttgtaatattttcttcccgttgtag.
pcr扩增转录因子osbzip86-2阅读框序列的引物核苷酸序列:
osbzip86-2f:5’-atgggtagcagtg-3’seqidno.6
osbzip86-2r:5’-ttagtttccttttttctggc-3’seqidno.7。
pcr扩增转录因子osbzip86-3阅读框序列的引物核苷酸序列:
osbzip86-3f:5’-atgggtagcagtg-3’seqidno.8
osbzip86-3r:5’-ctacaacgggaagaaaatattacaa-3’seqidno.9。
pcr扩增构建酵母载体的osbzip86-2阅读框序列的引物核苷酸序列:
osbzip86-2-f:acggaattcatgggtagcagtgseqidno.10
osbzip86-2-r:cgggatccttagtttccttttttctggcseqidno.11
pcr扩增构建酵母载体的osbzip86-3阅读框序列的引物核苷酸序列:
osbzip86-3-f:acggaattcatgggtagcagtgseqidno.12
osbzip86-3-r:cgggatccctacaacgggaagaaaatattacaaseqidno.13。
pcr扩增构建酵母载体的osbzip86-2△n阅读框序列的引物核苷酸序列:
osbzip86-2△n-f:acggaattcatgggtagcagtgseqidno.14
osbzip86-2△n-r:cgggatccagaatttgcttcacttccttcacseqidno.15。
pcr扩增构建酵母载体的osbzip86-2△c阅读框序列的引物核苷酸序列:
osbzip86-2△c-f:cggaattctctagtgaaggaagtgaagcaseqidno.16
osbzip86-2△c-r:cgggatccttagtttccttttttctggcseqidno.17。
pcr扩增构建酵母载体的osbzip86-3△c阅读框序列的引物核苷酸序列:
osbzip86-3△c-f:cggaattctctagtgaaggaagtgaagcaseqidno.18
osbzip86-3△c-r:cgggatccctacaacgggaagaaaatattacaaseqidno.19。
realtimert-pcr检测水稻中osbzip86的表达的引物核苷酸序列:
osbzip86-qrt/f:5’-tgctccaactacaaacttgaacata-3’seqidno.20
osbzip86-qrt/r:5’-gggaagggctaatagaacaaacc-3’seqidno.21。
realtimert-pcr检测水稻内参基因eef-1a的表达的引物核苷酸序列:
eef-1a-qrt/f:5’-gcacgctcttcttgctttc-3’seqidno.22
eef-1a-qrt/r:5’-agggaatcttgtcagggttg-3’seqidno.23。
realtimert-pcr检测水稻中aba合成途径基因osnced3的表达的引物核苷酸序列:
osnced3-qrt/f:5’-cccctcccaaaccatccaaaccga-3’seqidno.24
osnced3-qrt/r:5’-tgtgagcatatcctggcgtcgtga-3’seqidno.25。
为了便于理解本发明,下面将对本发明进行更全面的描述。本发明可以以许多不同的形式来实现,并不限于本文所描述的实施例。相反地,提供这些实施例的目的是使对本发明公开内容的理解更加透彻全面。
下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如sambrook等人,分子克隆:实验室手册(newyork:coldspringharborlaboratorypress,1989)中所述的条件,或按照制造厂商所建议的条件。实施例中所用到的各种常用化学试剂,均为市售产品。
除非另有定义,本发明所使用的所有的技术和科学术语与属于本发明的技术领域的技术人员通常理解的含义相同。本发明的说明书中所使用的术语只是为了描述具体的实施例的目的,不用于限制本发明。本发明所使用的术语“和/或”包括一个或多个相关的所列项目的任意的和所有的组合。
实施例1:水稻转录因子osbzip86基因cdna序列的rt-pcr扩增
正常粳稻品种zh11为材料,trizol裂解法提取总rna。对提取的叶片总rna进行b-500biophotometer核酸蛋白检测仪浓度检测。采用两步法以总rna为模板进行逆转录反应获取cdna第一链。cdna单链的合成依照takaraclontech公司
主要试剂
其它常规试剂为进口或国产分析纯试剂。
以水稻叶片的cdna单链为模板,采用高保真dna聚合酶
pcr反应体积为50μl。pcr扩增反应体系为:
pcr产物通过1%的琼脂糖凝胶电泳检测,可获得单一的dna条带(1173bp),osbzip86-3克隆步骤同上,可获得单一的dna条带(936bp)。即为pcr扩增得到的osbzip86基因片段(见图1所示)。osbzip86片段依照magen公司hipuregelpurednakits说明书进行琼脂糖凝胶电泳回收。
实施例2:水稻转录因子osbzip86生化功能的探究
酵母表达载体pgbkt7经bamhi和ecori双酶切处理,回收线性化载体。osbzip86-2和osbzip86-3(添加酶切位点bamhi和ecori)的pcr片段与线性化pgbkt7载体经nanodrop公司紫外分光光度计测定浓度,采用takara公司的t4dna连接酶进行dna片段和载体的同源重组连接。反应体系为:载体dna片段~0.03pmol,目的基因dna片段~0.3pmol,t4dna连接酶1μl,10×t4dna连接酶buffer1μl,ddh2o至总体积10μl。按照说明书的方法将反应产物转化大肠杆菌top10感受态菌株。挑取单克隆,提取质粒(质粒提取方法依照magen公司hipureplasmidmicrokitp1001-03说明书进行)。经测序鉴定为正确的阳性克隆后,保存质粒备用。经测序分析后,osbzip86-2和osbzip86-3的cdna阅读框核苷酸序列如seqidno.3和seqidno.4所示,所编码的蛋白质氨基酸序列如seqidno.1和seqidno.2所示。
采用引物对(表1)扩增osbzip86全长以及片段序列,分别与载体pgbkt7连接(连接步骤和方法同上),之后将这一重组载体转化进入酵母,酵母转化方法采用takaraclontech公司matchmakertmgoldyeasttwo-hybridsystemusermanual进行,若其表达产生的bd单独与uas结合也可以引起下游报告基因的转录,那么此片段具有转录激活活性。
列表1osbzip86-2和osbzip86-3分片段克隆引物序列
通过酵母双杂交体系中的自激活现象探究:通过转化酵母,同时转化pgbkt7空载体作为负对照。osbzip86-3及osbzip86-3△c具有转录活性(见图3)。这有助于探究剪切产生不同的转录本的功能差异性研究。
实施例4:水稻转录因子osbzip86亚细胞定位
通过网站http://rapdb.dna.affrc.go.jp/viewer/gbrowse_details/irgsp1?name=os12g0233800预测osbzip86属于碱性亮氨酸拉链(basicleucinezipper,bzip)转录因子,定位于细胞核。目前的研究表明,植物bzip转录因子在细胞中的位置主要定位于细胞核中,因为在其紧靠亮氨酸拉链结构域的n末端,通过一个固定的n-x7-r/k结构与特异dna序列相结合,起核定位信号作用。
添加了kpni和bamhi酶切位点的引物进行pcr扩增osbzip86-2和osbzip86-3的cds,与经过改造的puc18载体连接,构建融合表达载体。以经过改造的puc18空载体作为对照和将osbzip86-2::sgfp、osbzip86-3::sgfp融合表达载体分别与核定位信号mcherry共转入水稻原生质体,获得高效的瞬时表达。结果表明,对照gfp蛋白在细胞内均有分布,且与mcherry的定位完全一致。osbzip86则与mcherry共定位于细胞核中(见图5),与网站预测结果一致。
将osbzip86-2::sgfp、osbzip86-3::sgfp片段融合载体pxu1301中,并通过农杆菌eha105介导法的遗传转化方法,获得osbzip86-2::sgfp、osbzip86-3::sgfp的稳定表达的转基因材料以及负对照zh11、正对照puc18-gfp。待根生长至约2cm。利用广东省能源所的激光扫描共聚焦显微镜zeiss-lsm750观察细胞。osbzip86-2和osbzip86-3稳定表达的转基因材料的亚细胞定位结果和瞬时表达的结果一致(见图6)。
实施例5:水稻转录因子osbzip86在干旱胁迫下被诱导表达
本发明中所用的水稻材料为本实验室萌发的水稻小苗,取水稻野生型材料zh11大约50粒左右的饱满种子,采用70%乙醇浸泡1min,清水冲洗2次,3%的次氯酸钠浸泡40min,之后用自来水清洗5遍,将种子置于清水中萌发约5天,后移至国际水稻所常规水稻营养液继续培养约2周(22℃,每天16小时光照/8小时黑暗)。将zh11幼苗倒掉水培液进行缺水胁迫,收集胁迫处理0、0.5、1、2、4h后的水稻幼叶各0.5g。其余幼苗继续复水处理,收集复水处理0.5、1、2、4、8、20h后的水稻幼叶各0.5g。用于提取总rna。trizol裂解法提取总rna。对提取的叶片总rna进行b-500biophotometer核酸蛋白检测仪检测浓度。采用两步法以总rna为模板进行逆转录反应获取cdna第一链。cdna单链的合成依照takaraclontech公司
通过primerpremier5设计osbzip86基因的real-timert-pcr引物。用于检测osbzip86基因表达模式。用于检测osbzip86基因表达模式的引物为seqidno.20和seqidno.21(osbzip86-qrt/f:5’-tgctccaactacaaacttgaacata-3’和osbzip86-qrt/r:5’-gggaagggctaatagaacaaacc-3’)。内参基因为水稻组成型表达基因eef-1a,引物为seqidno.22和seqidno.23(eef-1a-qrt/f:5’-gcacgctcttcttgctttc-3’和eef-1a-qrt/r:5’-agggaatcttgtcagggttg-3’)。
参考takaraclontech公司
所有检测均采用两个生物学样本重复,每个生物学样本进行三次重复检测反应。
分别选取经过干旱及复水处理的野生型水稻zh11幼苗的地上部分提取总rna,通过qrt-pcr分析水稻转录因子基因osbzip86在非生物胁迫下的相对表达量(见图6)。结果表明,受到干旱诱导后,osbzip86的表达显著提高,复水后其表达逐渐恢复到正常水平。osbzip86的表达显著的受到干旱诱导,表明该基因具有干旱胁迫应答的基本功能,可将该基因应用于改良植物对干旱的耐受性。
实施例6:水稻转录因子osbzip86抗逆性实验中的功能探究
1、osbzip86-ox、osbzip86-rnai转基因水稻植株对aba敏感性实验
将osbzip86-ox、osbzip86-rnai和zh11水稻种子各30粒无菌播种于aba浓度0、3、5、10μm的1/2ms中,28℃,16/24h光周期条件下萌发,于处理10天进行观察表型分析及拍照,统计萌根长以及芽长(见图7)。
在10μmaba处理10天时,osbzip86-ox根长和芽长与zh11相比差别最大,根长增长61%,芽长增长46%,且具有显著性差异;osbzip86-rnai的萌发速度均慢于zh11,osbzip86-rnai根长和芽长与zh11相比差别最大,芽长减少20%,根长减少18%。通统计结果分析表明osbzip86-ox受到aba的抑制较小。其对aba的耐受性显著高于对照zh11和osbzip86-rnai,其对aba敏感性较低。
2、osbzip86-ox、osbzip86-rnai转基因水稻植株抗旱性实验探究
将在国际水稻所常规水稻营养液中进行水培约15天的osbzip86-ox、osbzip86-rnai和zh11幼苗倒掉水培液8小时进行缺水胁迫。然后再进行复水实验,复水7天后拍照并统计成活率(见图8)。成活率分别为成活率分别为zh11:7.40%;osbzip86-rnai:0%;osbzip86-ox:22.20%。统计结果说明osbzip86-ox转基因植株幼苗对干旱的耐受性显著优于对照zh11和osbzip86-rnai,受到干旱胁迫较小。说明osbzip86对水稻对干旱的耐受性是必须的。
3、干旱胁迫下osbzip86-ox、osbzip86-rnai转基因水稻中osnced3基因表达差异
本发明中所用的水稻材料为本实验室萌发的水稻小苗,取水稻转基因材料osbzip86-ox、osbzip86-rnai和zh11各50粒左右的饱满种子,采用70%乙醇浸泡1min,清水冲洗2次,3%的次氯酸钠浸泡40min,之后用自来水清洗5遍,将种子置于清水中萌发约5天,后移至国际水稻所常规水稻营养液继续培养约2周(28℃,每天16小时光照/8小时黑暗)。osbzip86-ox、osbzip86-rnai和zh11幼苗倒掉水培液进行缺水胁迫。收集胁迫处理0、1、2、3h后的水稻幼叶各0.5g,用于提取总rna。trizol裂解法提取总rna。对提取的叶片总rna进行b-500biophotometer核酸蛋白检测仪检测浓度。采用两步法以总rna为模板进行逆转录反应获取cdna第一链。cdna单链的合成依照takaraclontech公司
通过文献获得的osnced3基因的real-timert-pcr引物。用于检测osnced3基因表达模式。用于检测osnced3基因表达模式的引物为seqidno.24和seqidno.25(osnced3-qrt/f:5’-cccctcccaaaccatccaaaccga-3’和osnced3-qrt/r:5’-tgtgagcatatcctggcgtcgtga-3’)。内参基因为水稻组成型表达基因eef-1a,引物为seqidno.22和seqidno.23。
realtimert-pcr体系及程序参考实施例5。
如图9所示,在干旱胁迫处理下,osbzip86-ox中aba合成途径基因osnced3基因的表达受到干旱诱导,其最大诱导量达到4倍。osbzip86-rnai中osnced3基因的表达,在干旱处理2h时表现出显著性降低。以上公开的内容表明了osbzip86基因是一个可促进aba合成途径基因osnced3在干旱胁迫下高表达,从而增强水稻抗旱性。osbzip86还可能直接参与osnced3基因的转录表达。该基因具备应用于针对水稻耐旱分子育种工程改良的潜力。
4、干旱胁迫下osbzip86-ox、osbzip86-rnai转基因水稻中aba含量测定
本发明中所用的水稻材料为本实验室萌发的水稻小苗,取水稻转基因材料osbzip86-ox、osbzip86-rnai和zh11各50粒左右的饱满种子,采用70%乙醇浸泡1min,清水冲洗2次,3%的次氯酸钠浸泡40min,之后用自来水清洗5遍,将种子置于清水中萌发约5天,后移至国际水稻所常规水稻营养液继续培养约2周(28℃,每天16小时光照/8小时黑暗)。osbzip86-ox、osbzip86-rnai和zh11幼苗倒掉水培液进行缺水胁迫。收集胁迫处理4小时后的水稻幼叶各0.5g,干冰保存送至广州菲博生物公司采用植物激素脱落酸(aba)elisakit进行植物aba含量测定。osbzip86-ox中aba含量受到干旱诱导积累增加,增加量与zh11比较呈显著性差异,osbzip86-rnai中aba含量与zh11比较表现出降低趋势(见图10)。以上公开的内容表明了osbzip86基因是一个通过aba途径响应干旱胁迫的应答基因,该基因可作为丰富bzip家族在aba合成信号途径中的作用做进一步研究,有为水稻耐旱分子育种提供更为丰富的基因遗传资源的潜力。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
sequencelisting
<110>中国科学院华南植物园
<120>水稻osbzip86基因及其编码蛋白在干旱胁迫中的应用
<160>25
<170>patentinversion3.3
<210>1
<211>390
<212>prt
<213>osbzip86-2protein
<400>1
metglyserserglyalaaspalaprothrlysthrserlysalaser
151015
alaproglngluglnglnproproalaserserserthralathrpro
202530
alavaltyrproasptrpalaasnpheglnglytyrproproilepro
354045
prohisglyphepheproserprovalalaserserproglnglyhis
505560
protyrmettrpglyalaglnprometileproprotyrglythrpro
65707580
proproprotyrvalmettyrproproglyvaltyralahisproser
859095
metproproglyalahisprophethrprotyralametalaserpro
100105110
asnglyasnalaaspprothrglythrthrthrthralaalaalaala
115120125
alaalaglygluthraspglylyssersergluglylysglulysser
130135140
proilelysargserlysglyserleuglyserleuasnmetilethr
145150155160
glylysasnserthrgluhisglylysthrserglyalaseralaasn
165170175
glyalaileserglnserglygluserglysergluserserserglu
180185190
glyserglualaasnserglnasnaspserhishislysglusergly
195200205
glngluglnaspglygluvalargserserglnasnglyvalserarg
210215220
serproserglnalalysleuasnglnthrmetalailemetpromet
225230235240
thrserserglyprovalproalaprothrthrasnleuasnilegly
245250255
metasptyrtrpalaasnthralaserserthrproalailehisgly
260265270
lysalathrprothralaalaproglysermetvalproglyglugln
275280285
trpvalglnaspgluarggluleulysargglnargarglysglnser
290295300
asnarggluseralaargargserargleuarglysglnalaglucys
305310315320
glugluleualaglnargalagluvalleulysglngluasnthrser
325330335
leuargaspgluvalasnargilearglysglutyraspgluleuleu
340345350
serlysasnserserleulysglulysleugluasplysglnhislys
355360365
thraspglualaglyvalaspasnlysleuglnhisserglyaspasp
370375380
serglnlyslysglyasn
385390
<210>2
<211>312
<212>prt
<213>osbzip86-3protein
<400>2
metglyserserglyalaaspalaprothrlysthrserlysalaser
151015
alaproglngluglnglnproproalaserserserthralathrpro
202530
alavaltyrproasptrpalaasnpheglnglnglytyrproproile
354045
proprohisglyphepheproserprovalalaserserproglngly
505560
hisprotyrmettrpglyalaglnprometileproprotyrglythr
65707580
proproproprotyrvalmettyrproproglyvaltyralahispro
859095
sermetproproglyalahisprophethrprotyralametalaser
100105110
proasnglyasnalaaspprothrglythrthrthrthralaalaala
115120125
alaalaalaglygluthraspglylyssersergluglylysglulys
130135140
serproilelysargserlysglyserleuglyserleuasnmetile
145150155160
thrglylysasnserthrgluhisglylysthrserglyalaserala
165170175
asnglyalaileserglnserglygluserglysergluserserser
180185190
gluglyserglualaasnserglnasnaspserhishislysgluser
195200205
glyglngluglnaspglygluvalargserserglnasnglyvalser
210215220
argserproserglnalalysleuasnglnthrmetalailemetpro
225230235240
metthrserserglyprovalproalaprothrthrasnleuasnile
245250255
glymetasptyrtrpalaasnthralaserserthrproalailehis
260265270
glylysalathrprothralaalaproglysermetvalproglyglu
275280285
glntrpvalglnvalcysserileserproserleuglngluprocys
290295300
thrcysasnilephepheproleu
305310
<210>3
<211>935
<212>dna
<213>osbzip86-3
<400>3
atgggtagcagtggcgcagacgcaccgactaagacaagcaaggcatctgcacctcaggag60
caacagccacctgctagttcaagcactgcaacaccggctgtttacccagattgggccaac120
tttcagggatatcctccaattccaccacatggcttctttccatcacctgtggcgtcaagc180
ccacagggtcatccttacatgtggggagctcagcctatgataccaccatatggaacccca240
ccaccaccatatgtcatgtatcctccaggagtatatgctcacccatctatgcctccggcg300
cacacccatttactccatatgccatggcctctccaaatggcaatgctgatcctactggaa360
ctacaactactgctgctgctgctgctgctggtgagacggatggcaaatcctctgaaggaa420
aagaaaaaagtcccattaaaagatctaaaggaagtttaggtagcttgaatatgattacag480
ggaagaactctactgaacatggtaaaacctctggggcatcagctaatggagccatttctc540
aaagtggggaaagtggaagtgaaagttctagtgaaggaagtgaagcaaattctcagaatg600
attcacatcacaaggaaagtggacaagagcaagatggagaggttcgaagttcccagaatg660
gtgtatcacgttcaccatcccaggcaaagttgaatcaaactatggcaatcatgccaatga720
catcaagtggtccagtacctgctccaactacaaacttgaacataggaatggactactggg780
ccaacacagctagctccactccagcaatacatggtaaagcaaccccaactgcagctccag840
ggtctatggttccaggagagcagtgggtgcaggtttgttctattagcccttcccttcaag900
aaccgtgcacttgtaatattttcttcccgttgtag935
<210>4
<211>1173
<212>dna
<213>osbzip86-2
<400>4
atgggtagcagtggcgcagacgcaccgactaagacaagcaaggcatctgcacctcaggag60
caacagccacctgctagttcaagcactgcaacaccggctgtttacccagattgggccaac120
tttcagggatatcctccaattccaccacatggcttctttccatcacctgtggcgtcaagc180
ccacagggtcatccttacatgtggggagctcagcctatgataccaccatatggaacccca240
ccaccaccatatgtcatgtatcctccaggagtatatgctcacccatctatgcctccgggc300
gcacacccatttactccatatgccatggcctctccaaatggcaatgctgatcctactgga360
actacaactactgctgctgctgctgctgctggtgagacggatggcaaatcctctgaagga420
aaagaaaaaagtcccattaaaagatctaaaggaagtttaggtagcttgaatatgattaca480
gggaagaactctactgaacatggtaaaacctctggggcatcagctaatggagccatttct540
caaagtggggaaagtggaagtgaaagttctagtgaaggaagtgaagcaaattctcagaat600
gattcacatcacaaggaaagtggacaagagcaagatggagaggttcgaagttcccagaat660
ggtgtatcacgttcaccatcccaggcaaagttgaatcaaactatggcaatcatgccaatg720
acatcaagtggtccagtacctgctccaactacaaacttgaacataggaatggactactgg780
gccaacacagctagctccactccagcaatacatggtaaagcaaccccaactgcagctcca840
gggtctatggttccaggagagcagtgggtgcaggatgaacgggaactcaaaaggcagaga900
agaaaacaatccaacagggagtctgctcgcagatctaggttgcgtaagcaggctgaatgt960
gaggagttggctcaacgtgctgaagttttaaagcaagaaaatacttcacttagagatgaa1020
gtgaaccggatcagaaaggagtatgatgagcttctatcaaagaatagctcattaaaggaa1080
aaacttgaggacaaacaacacaaaactgatgaggcaggagttgacaataagctgcaacat1140
tctggtgatgacagccagaaaaaaggaaactaa1173
<210>5
<211>369
<212>dna
<213>osbzip86-3△c
<400>5
tctagtgaaggaagtgaagcaaattctcagaatgattcacatcacaaggaaagtggacaa60
gagcaagatggagaggttcgaagttcccagaatggtgtatcacgttcaccatcccaggca120
aagttgaatcaaactatggcaatcatgccaatgacatcaagtggtccagtacctgctcca180
actacaaacttgaacataggaatggactactgggccaacacagctagctccactccagca240
atacatggtaaagcaaccccaactgcagctccagggtctatggttccaggagagcagtgg300
gtgcaggtttgttctattagcccttcccttcaagaaccgtgcacttgtaatattttcttc360
ccgttgtag369
<210>6
<211>13
<212>dna
<213>人工序列
<400>6
atgggtagcagtg13
<210>7
<211>20
<212>dna
<213>人工序列
<400>7
ttagtttccttttttctggc20
<210>8
<211>13
<212>dna
<213>人工序列
<400>8
atgggtagcagtg13
<210>9
<211>25
<212>dna
<213>人工序列
<400>9
ctacaacgggaagaaaatattacaa25
<210>10
<211>22
<212>dna
<213>人工序列
<400>10
acggaattcatgggtagcagtg22
<210>11
<211>28
<212>dna
<213>人工序列
<400>11
cgggatccttagtttccttttttctggc28
<210>12
<211>22
<212>dna
<213>人工序列
<400>12
acggaattcatgggtagcagtg22
<210>13
<211>33
<212>dna
<213>人工序列
<400>13
cgggatccctacaacgggaagaaaatattacaa33
<210>14
<211>22
<212>dna
<213>人工序列
<400>14
acggaattcatgggtagcagtg22
<210>15
<211>31
<212>dna
<213>人工序列
<400>15
cgggatccagaatttgcttcacttccttcac31
<210>16
<211>29
<212>dna
<213>人工序列
<400>16
cggaattctctagtgaaggaagtgaagca29
<210>17
<211>28
<212>dna
<213>人工序列
<400>17
cgggatccttagtttccttttttctggc28
<210>18
<211>29
<212>dna
<213>人工序列
<400>18
cggaattctctagtgaaggaagtgaagca29
<210>19
<211>33
<212>dna
<213>人工序列
<400>19
cgggatccctacaacgggaagaaaatattacaa33
<210>20
<211>25
<212>dna
<213>人工序列
<400>20
tgctccaactacaaacttgaacata25
<210>21
<211>23
<212>dna
<213>人工序列
<400>21
gggaagggctaatagaacaaacc23
<210>22
<211>19
<212>dna
<213>人工序列
<400>22
gcacgctcttcttgctttc19
<210>23
<211>20
<212>dna
<213>人工序列
<400>23
agggaatcttgtcagggttg20
<210>24
<211>24
<212>dna
<213>人工序列
<400>24
cccctcccaaaccatccaaaccga24
<210>25
<211>24
<212>dna
<213>人工序列
<400>25
tgtgagcatatcctggcgtcgtga24
- 还没有人留言评论。精彩留言会获得点赞!
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 降低种子重量的白桦AP2基因及其编码蛋白的制造方法与工艺
- 可编码ArtRD蛋白基因及其蛋白和应用的制造方法与工艺
- 蝎神经毒素AaIT及其编码基因与应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 腈水解酶突变体及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白AsTMK1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种花狭口蛙的基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种泽陆蛙基因及其编码的多肽和该多肽的用途的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 甘薯耐逆相关蛋白IbFBA2、编码基因、重组载体与应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺
- 植物抗病毒相关蛋白GmSN1及其编码基因和应用的制造方法与工艺
- 植物抗旱相关蛋白EtSnRK2.2及其编码基因和应用的制造方法与工艺
- OsSAPK7蛋白及其编码基因在提高水稻白叶枯病抗性中的应用的制造方法与工艺
- 一种抗除草剂抗虫融合基因、编码蛋白及应用的制造方法与工艺
- 毛果杨PtrZFP103基因及其编码蛋白和应用的制造方法与工艺
- 大豆bHLH转录因子基因GmFER及其编码蛋白和应用的制造方法与工艺
- 一种水稻MIT1基因、其编码蛋白及应用的制造方法与工艺
- 蛋白StTrxF、编码基因及其在提高植物淀粉含量中的应用的制造方法与工艺
- 蛋白质及其编码基因在调控植物抗黄萎病性中的应用的制造方法与工艺
- 植物抗旱相关蛋白ZmNAC111及其编码基因与应用的制造方法与工艺
- 核盘菌凝集素SSL‑6蛋白及其编码基因与应用的制造方法与工艺