神经系统肿瘤细胞自噬体的制备方法及其在制备疫苗中的用途与流程
本发明属生物技术领域,涉及神经系统肿瘤细胞自噬体及其用途,具体涉及神经系统肿瘤细胞自噬体的制备方法及其在制备疫苗中的用途,所述的疫苗是针对神经系统来源肿瘤细胞自噬体的树突状细胞肿瘤疫苗。
背景技术:
现有技术公开了脑胶质瘤为起源于神经胶质细胞的肿瘤,是最常见的原发性颅内肿瘤,其中,胶质母细胞瘤(whoiv级)的发病率最高,占46.1%,发病率约为3.20/10万,中青年病死率排名第二。临床实践显示,包括手术、辅助同步放化疗在内的治疗干预方案,仅能延缓其进程,所述的疾患均不可避免地复发。据有关统计显示,脑胶质瘤疾患其平均的进展时间为6.9个月,总生存期仅14.6个月,临床治疗干预效果很不理想。寻找更多的新技术性干预方案进而提升患者的生存状态,已成为本领域研究人员关注的重点问题。研究显示,中枢神经系统(cns)存在活跃且被严密调控的免疫系统,为脑胶质母细胞瘤的免疫治疗提供了理论依据,并且,从整个肿瘤的领域来看,随着免疫肿瘤学的逐渐明晰,本领域研究人员预测,在未来的几十年内,肿瘤的治疗将会发生以肿瘤为中心的治疗策略向以肿瘤患者免疫系统为中心的治疗策略的转变。
有研究报道,机体免疫系统对体内发生畸变的细胞有免疫监视的作用,而在肿瘤个体体内,肿瘤细胞则属逃避了机体的免疫监视的特例;肿瘤免疫逃逸的情形纷繁复杂,而其中比较重要的一个环节是肿瘤抗原不能有效的呈递给t细胞。还有研究报道,树突状细胞(dc)作为抗原呈递细胞的一个重要组成部分,在肿瘤个体内往往数量减少,功能低下甚至缺陷,导致肿瘤的免疫逃逸,使免疫系统不能有效的控制肿瘤的形成和发展。业内认为,dc细胞作为唯一能活化初次免疫应答的抗原呈递细胞,激活免疫应答的能力十分强大,所以目前,基于dc的肿瘤疫苗仍是一类十分重要而有效的肿瘤疫苗,并且已经有了大量基础研究和临床试验的数据。
有效的dc肿瘤疫苗的有效实现依赖于两个方面,一为dc自身具备有效的抗原呈递作用;二是负载特异性的有效抗原。然而在具体的研究试验中,还存在许多待解决的问题,包括dc的来源、诱导、制备过程,dc抗原的选择,抗原的用量、负载方式,最后dc肿瘤疫苗的使用途径、频率、剂量等等,而抗原的制备和负载方式是其中的重要的环节。
研究认为,肿瘤自噬体很适合成为dc肿瘤疫苗所负载的有效抗原;自噬(autophagy)作为细胞内两大蛋白降解途径之一(另一途径为泛素-蛋白酶体系统),是普遍存在的细胞生物学现象,即便在神经系统也是如此。自噬是指胞浆的组成成分以及细胞器,在一定的诱因下,被脂质囊泡包裹形成自噬小体(autophagosome),后与溶酶体融合形成自噬溶酶体复合物(autolysosome),在溶酶体的酸性环境下,内容物被降解,最终实现胞内蛋白的降解。而该过程不仅为生命活动提供再利用的原材料,也将过剩、甚至错误发生的蛋白进行清除。而肿瘤的恶变过程往往会伴随着大量错误折叠蛋白的产生,而自噬的固有功能恰好可以富集上述错误折叠蛋白,有利于所诱导肿瘤疫苗的特异性。鉴于此特点,肿瘤自噬体抗原相较常规含有大量自身抗原的肿瘤冻融裂解抗原,所负载的dc肿瘤疫苗的特异性会有大幅提升。
关于肿瘤自噬体负载dc的肿瘤疫苗,已有不少的研究结果加以证实,抑制自噬的方法如自噬抑制剂、sirna干扰等抑制肿瘤细胞自噬后,会明显降低dc交叉提呈肿瘤抗原的免疫效果;而促进自噬的方法如自噬体诱导剂、饥饿等诱导肿瘤细胞自噬后,则会明显增强dc交叉提呈的免疫效果。
但神经系统的自噬的诱导一直为本领域视为较困难的问题,且认为经典的诱导剂组合“雷帕霉素+氯化铵”并不能在神经系统有效的诱导其自噬的产生;本申请的课题研究组经过长期摸索,发现上述原因在于经典的促进细胞自噬体释放的“超声裂解”法在提取神经系统来源细胞的自噬体时,极易裂碎神经系统自噬体膜,导致神经系统自噬体的提取效率低下。
dc交叉呈递的抗原是特异性免疫反应的物质基础、是效应淋巴细胞有效识别并杀伤肿瘤细胞的靶点。dc自噬体肿瘤疫苗所面临的难题,除了上述提取的困难外,还在于:一、肿瘤细胞表面呈递的主要为肿瘤细胞自身产生的短寿蛋白(半衰期约10分钟),而经自噬途径得到的抗原为肿瘤的长寿蛋白,两者不一定匹配,此为以“长寿蛋白”为基础的肿瘤疫苗的难题之一;二、dc在不同的状态下交叉呈递的能力不同,而交叉呈递效能的高低将直接影响dc肿瘤疫苗的效力,如何使得dc以最大的效力进行肿瘤抗原的交叉提呈,是以dc为基础的dc肿瘤疫苗的难题之二。
基于现有技术的现状,本申请的发明人拟提供神经系统肿瘤细胞自噬体的制备方法及其在制备疫苗中的用途,所述的疫苗是针对神经系统来源肿瘤细胞自噬体的树突状细胞肿瘤疫苗。
技术实现要素:
本发明的目的在于克服现有技术存在的缺陷与不足,提供一种神经系统来源的肿瘤细胞自噬体的制备方法及其在制备肿瘤疫苗中的用途,尤其是在制备针对神经系统来源肿瘤细胞自噬体的树突状细胞肿瘤疫苗中的用途。
为达到上述目的,本发明的具体技术方案如下:
一种神经系统来源的肿瘤细胞自噬体的制备方法,包括以下步骤:
步骤1,培养神经系统来源的肿瘤细胞;
步骤2,通过加入50-150ng/ml雷帕霉素、5-10mmol/l氯化铵,培养48±5小时诱导步骤1中培养细胞内产生自噬体。
或,
一种神经系统来源的肿瘤细胞自噬体的制备方法,包括以下步骤:
步骤1,培养神经系统来源的肿瘤细胞;
步骤2,诱导步骤1中培养的细胞自噬;
步骤3,用细胞刮子将步骤2诱导后的细胞刮起,用移液器直接反复吹打15-30次;
或,
一种神经系统来源的肿瘤细胞自噬体的制备方法,包括以下步骤:
步骤1,培养神经系统来源的肿瘤细胞;
步骤2,诱导步骤1中培养的细胞自噬;
步骤3,通过刮取和吹打,促进步骤2中肿瘤自噬体释放;
步骤4,将步骤3中吹打后的细胞及培养液进行第一步离心,取上清,弃沉淀;步骤5,将步骤4中离心后得到的上清液进行第二步离心,弃去上清,保留沉淀,收集备用。
上述制备的神经系统肿瘤细胞自噬体用于制备dc肿瘤疫苗,包括以下步骤:
步骤6,将步骤5中所获得自噬体抗原与未成熟dc细胞共培养,刺激成熟的dc细胞收集备用;
优选的,所述步骤2中的雷帕霉素、氯化铵的浓度分别为100±50ng/ml、10±2mmol/l。
优选的,所述步骤2中的加入雷帕霉素、氯化铵培养的时间为48±5小时。
优选的,所诉步骤3中的促自噬体释放方法为,用细胞刮子将将步骤2诱导后的细胞刮起,用移液器直接反复吹打15-30次;
优选的,所述步骤4中的第一步离心的离心力为400-550g。
优选的,所述步骤5中第二步离心的离心力为10000g-15000g。
优选的,所述肿瘤细胞自噬体负载树突状细胞制备肿瘤自噬体dc疫苗。
相对于现有技术,本发明的技术方案具有以下优点:
1,提高神经系统来源的肿瘤细胞自噬体的获得效率,显著促进了胞浆内自噬体的释放,获得自噬体快速而大量。
2,本发明的技术方案自噬体作为一种新的交叉提呈的抗原,负载dc后,可以有效的促进dc的成熟,激活t淋巴细胞,促进t淋巴细胞的增殖,并对肿瘤产生明显的杀伤作用,杀伤效力明显高于通常所用的反复冻融肿瘤细胞裂解液所制备的肿瘤抗原。
附图说明
构成本发明的一部分的附图用来提供对本发明的进一步理解,本发明的示意性实施例及其说明用于解释本发明,并不构成对本发明的不当限定。在附图中:
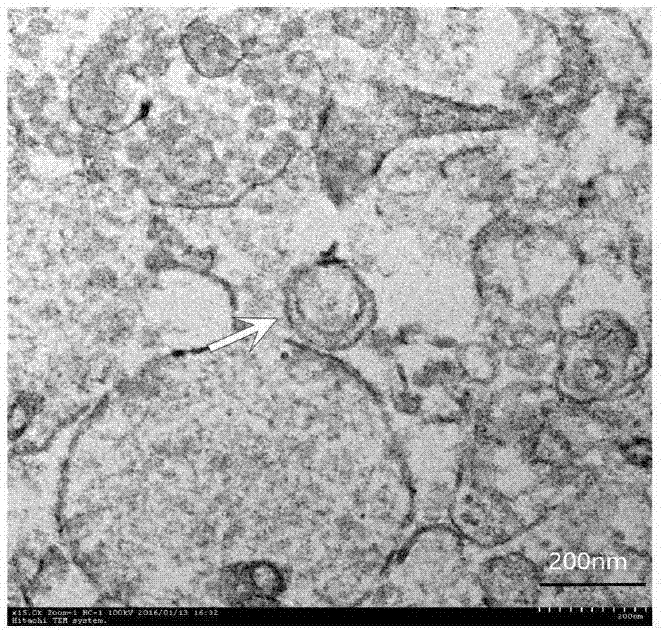
图1为电镜下观察提取物中自噬体形态。
图2为携带mcherry红色荧光lc3基因的慢病毒转染293细胞在本发明实施例的技术干预下出现明显自噬,在共聚焦显微镜下发出明显红色荧光。
图3为神经胶质瘤细胞系自噬体诱导试剂浓度的选择。
图4为神经胶质瘤细胞系自噬体诱导时间的选择。
图5为裂解胶质瘤细胞的方法,“超声裂解法”和“反复吹打”法提取自噬体效率的比较。
图6为所获得的未成熟dc细胞形态。
图7为空白抗原、自噬体抗原、肿瘤细胞冻融裂解液抗原促进dc成熟的能力的比较。
图8为自噬体抗原组、肿瘤细胞冻融裂解液抗原组dc促cd8+t细胞增殖能力的比较和最佳刺激比(成熟dc:cd8+t淋巴细胞)的选择。
图9为空白抗原组、肿瘤细胞冻融裂解液抗原组、自噬体抗原组效应t细胞对靶细胞杀伤效率的比较和杀伤比的确定(效应t细胞:靶细胞)。
具体实施方式
下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,均属于本发明保护的范围。
需要说明的是,在不冲突的情况下,本发明中的实施例及实施例中的特征可以相互组合。
以下将结合附图对本发明的实施例做具体阐释。
实施例1制备神经系统来源的肿瘤细胞自噬体,神经系统来源的肿瘤细胞采用c6/gl261胶质瘤细胞,
包括以下步骤:
(1)c6/gl261胶质瘤细胞系的培养:
复苏冻存的c6/gl261胶质瘤细胞,37℃水浴迅速融化,不时摇动,力求1分钟内达到室温以上,无菌下转移至5ml的含有10%胎牛血清的dmem的完全培养基内,1000r/min速度下离心5分钟,弃去上清液,加入适量上述完全培养基,调整接种密度约为1×10^6/ml,接种于t72培养瓶中,置37℃,5%二氧化碳的培养箱中静置培养,次日待细胞贴壁后换液一次,继续培养,视生长情况判断是否传代;若细胞密度尚可,弃去上清常规换液即可,若细胞密度过高,及时0.25%胰蛋白酶消化传代,以免细胞老化;
(2)c6/gl261胶质瘤细胞系的干预:
待步骤(1)中培养的细胞贴壁状态良好,密度适中,处于对数生长期时,加入0-250ng/ml雷帕霉素、0-10mmol/l氯化铵,优选量分别为100±50ng/ml、10±2mmol/l,并优选为100ng/ml、10mmol/l,诱导培养胶质瘤细胞系48±5小时后,提取胶质瘤细胞系自噬体;同时设置有对照组(只加入完全培养基,不含自噬诱导药物);
(3)提取c6/gl261胶质瘤细胞系自噬体:
将步骤(2)诱导后的细胞,使用经优选过的提取方法:直接用细胞刮子刮取,反复吹打,促进自噬体释放以及离心后,得到的沉淀即为自噬体;
步骤(3)里的具体自噬体提取步骤如下:
(3a)优选的,用细胞刮子将细胞从培养器具底部刮起,之后用移液器直接反复吹打15-30次,优选为15-20次,并优选为20次;
(3b)将步骤(3a)中的细胞及培养液混合液倒入50ml离心管中,低速离心400-550g,优选为400-500g,并优选为500g,10±2min,4℃,取上清,弃沉淀;
(3c)将步骤(3b)中离心后得到的上清液移入45ml圆底超高速离心管中,高速离心10000-15000g,优选为10000-12000g,并优选为12000g,20±5min,4℃,弃去上清,保留沉淀;
(3d)将步骤(3c)中得到的沉淀用预冷的pbs重悬后,定容分装,收集到1.5mlep管中,冻存于-20℃,备用。
本实施例在雷帕霉素、氯化铵两种药物联合应用处理c6/gl261两种胶质瘤细胞系,诱导自噬体产生后,通过细胞刮子的直接刮取和对所得细胞的反复吹打20次,明显促进了胞浆内自噬体的释放,最后再通过分步高速离心的方法提取到c6/gl261胶质瘤细胞系的自噬体,获得自噬体快速而大量,提高了神经系统来源的肿瘤细胞自噬体的获得效率;
本实施例所制备的胶质瘤自噬体作为一种新的交叉提呈的抗原,经后续证实,负载dc后,可以有效的促进dc的成熟,激活t淋巴细胞,促进t淋巴细胞的增殖,并对肿瘤产生明显的杀伤作用,杀伤效力明显高于通常所用的反复冻融肿瘤细胞裂解液所制备的肿瘤抗原。
将本实施例提取得到的自噬体超高速离心,使其压缩紧密,弃上清液后,加入2.5%戊二醛固定沉淀,送电镜室进行后续处理;上镜观察自噬体形态,如图1中所示:电镜下可见双层膜结构的小体,直径在200nm左右(白色箭头所指),证实本发明的实施例有效的诱导和提取了c6/gl261胶质瘤细胞系自噬体。
本实施例进行了自噬体的鉴定,并对其相关生物学活性进行检测,通过电子显微镜可观察到典型的双层膜结构,直径在200nm左右,符合自噬体的特点,提示自噬体被有效的诱导以及提取。
本发明的实施例还通过慢病毒转染而携带mcherry红色荧光-lc3(microtubule-associatedprotein1lightchain3)基因的293工具细胞的建立:
1)第一天用dmem+10%胎牛血清(fbs)培养基铺板293细胞。确保第二天细胞有80-90%融合度;
2)用500ulopti-mem(减血清)培养基稀释2ugpwpxld-mcherry红色荧光lc3基因+1.5ugpspax2慢病毒载体+1.5ugpmd2g慢病毒载体;
3)用500ulopti-mem稀释15ullipofectamine2000转染试剂;
4)5min后,混合2)、3),室温静置30分钟;
5)从6孔板中吸出1ml培养基后,滴加1ml4)中所得质粒和脂质体混合物;
6)6-10h后,移除含有dna-脂质体复合物的培养基,代之以正常培养液dmed+10%fbs,继续再培养24h;
7)荧光显微镜下观察6)中所得细胞,可见红色荧光,阴性对照无此荧光;
8)为提升转染效果,可每隔48h收集含病毒的上清,0.45um滤膜过滤后,给目的细胞换液时按一份病毒液配两份完全培养基的比例反复感染目的细胞;
9)所获得的携带mcherry红色荧光lc3基因的293工具细胞直观的反映了细胞内自噬的强弱,侧面验证本技术即便是在不同的细胞系,也具有相当的有效性。
如图2所示:药物处理后的细胞在荧光下明显出现红色荧光,而红色荧光与lc3在同一转染基因,表明在干预的作用下,促进了自噬体的产生:一方面自噬诱导剂雷帕霉素诱导了自噬体的产生,另一方面,氯化铵则抑制了溶酶体的酸化,阻止了溶酶体和自噬体的融合;故而雷帕霉素和氯化铵共同协作,有效的促进了自噬体的产生。
本发明的实施例利用携带mcherry红色荧光lc3基因的慢病毒转染293工具细胞对流程进行优化,配合westernblot和电子显微镜验证,极大的优化了胶质瘤细胞系自噬体的提取线路。
本发明的实施例还对自噬体总蛋白含量测定(bca法):
1)取0.8ml蛋白标准配制液加入到一管蛋白标准(20mgbsa牛血清白蛋白)中,充分溶解后配制成25mg/ml的标准蛋白溶液,取适量25mg/ml标准蛋白溶液,稀释至终浓度为0.5mg/ml的标准品;
2)将标准品按0、1、2、4、8、12、16、20μl加到96孔板的标准品孔中,加标准品稀释液补足到20ul,相当于标准品浓度分别为0、0.025、0.05、0.1、0.2、0.3、0.4、0.5mg/ml;
3)将待测自噬体的pbs(磷酸缓冲盐)溶液,取适量体积,加入96孔板中,加pbs补足到20ul;
4)根据样品数量,按50体积bca试剂a加1体积bca试剂b(50:1)配制适量bca工作液,充分混匀;
5)各孔加入200μlbca工作液,室温放置2小时;
6)用酶标仪测定a546的波长的吸光度;
7)根据标准曲线和使用的样品体积计算出样品的蛋白浓度。结果大约为3ug/ul。
本发明的实施例还通过westernblot检测所提取自噬体总蛋白中标记蛋白lc3的表达:
lc3按所在部位的区别分为胞浆型即lc3-i,参与了自噬体的形成;在自噬发生时,lc3-i与磷脂酰乙醇胺共价结合后变为lc3-ii即膜型,锚定于自噬体的双层膜上,也因此,lc3-ii被公认为自噬体的特征性标记物;
1)配制分离胶和浓缩胶:配方如下
a.分离胶(15%)配制(总体积5ml):
b.浓缩胶(5%)的制备(总体积2ml):
2)sds-page电泳
待浓缩胶凝固,在电泳液中拔去梳子。根据蛋白定量的浓度调整蛋白浓度至均一浓度,调整每孔上样量。取样品蛋白,每孔约8ul,加入5*sds样品加样缓冲液2ul混匀,100℃煮沸3min,加入蛋白分子量标准,靠外的两槽再加入等体积的工1*sds上样缓冲液,加入电泳液,开始电泳,初始电压为45v进行稳流电泳,当电压达到65v时改为稳压电泳。在带蓝色条带移动至胶板下缘1cm以上结束电泳;
3)转膜
转膜开始前开始预冷转膜所需装置和转膜液。电泳结束后,将胶取下,根据目的蛋白分子量切下相应位置的胶,浸泡与转膜液中,根据胶的大小剪出同样大小的pvdf膜,用pvdf膜进行活化,待膜浸润透之后,移至转膜液中继续浸泡约30分钟,按:负极/海绵/3层滤纸/胶/膜/3层滤纸/海绵/正极的顺序制备“转移三明治”,每层放好后,用试管赶去气泡。恒流100ma电转移60min;
4)封闭
用1*tbst浸泡洗涤pvdf膜5min后,用含5%脱脂高蛋白牛奶的tbst在室温下封闭,慢摇1小时;
5)一抗孵育
tbst1:1000稀释兔抗鼠lc3单克隆抗体,4℃过夜。次日tbst洗涤3次,每次10min;
6)二抗孵育和ecl显影
tbst1:2000稀释辣根过氧化物酶(hrp)标记山羊抗兔igg(h+l),室温孵育1小时,tbst洗涤3次,每次10min。滴加ecl超敏发光底物曝光显影。
如图3所示:不同浓度的雷帕霉素(rapa)和氯化铵(nh4cl)干预后,lc3表达情况的变化,其中100ng/ml雷帕霉素+10mmol氯化铵的药物处理后,lc3ii/lc3i比值最高,预示着此时自噬体产生的量最高,表明100ng/ml雷帕霉素+10mmol氯化铵的药物处理浓度为小鼠胶质瘤细胞系自噬体的最佳诱导浓度。
如图4所示:不同诱导时间下,lc3表达情况的变化,其中48小时时,lc3ii/lc3i比值最高,预示着此时自噬体产生的量最高。表明48小时为提取自小鼠胶质瘤细胞系噬体的最佳提取时间。
如图5所示超声裂解细胞(c)和反复吹打(f)两种方法提取自噬体效率的比较,其中“反复吹打”法的lc3ii/lc3i比值始终明显高于超声裂解法各裂解时间段的处理组,预示着此时促进自噬体释放的程度最高。表明本技术中的“反复吹打”比使用超声裂解更适合提取自噬体。
实施例2
在制备肿瘤疫苗的应用中,本发明的实施例通过获取小鼠dc和dc负载自噬体抗原:
1)小鼠dc的获取
1、断颈处死c57小鼠,浸泡于75%酒精当中,浸泡约10min后,使用无菌手术台固定小鼠;
2、无菌取出小鼠胫骨、股骨,浸泡于pbs;
3、用1ml注射器抽取pbs冲洗骨髓腔,至骨质变白,吹打混匀后,200目滤网过滤后加入50ml离心管;
4、1000rpm,5分钟,室温离心,去上清;
5、加入2ml红细胞裂解液,室温放置3分钟;
6、加入10mlpbs混匀,终止裂解。1000rpm,5分钟,室温离心,弃上清;
7、加入3ml含40ng/ml的il-4、40ng/ml的gm-csf和10%胎牛血清的1640培养基(dc诱导培养基)重悬;
8、调整细胞浓度到1×10^6个/ml后接种培养;
9、48小时后换液,去除悬浮细胞,保留贴壁细胞;
10、120小时后换液,吹打起悬浮和疏松贴壁的细胞,收集到50ml离心管中,1000rpm,5分钟,室温离心,去上清;
11、用3ml含40ng/ml的il-4、40ng/ml的gm-csf和10%胎牛血清的1640培养基(dc诱导培养基)重悬;
12、调整细胞浓度到1×10^6个/ml后接种到一个新的培养板中培养;
13、48小时,吹打起悬浮和疏松贴壁的细胞,收集到50ml离心管中,1000rpm,5分钟,室温离心,去上清。
即获得未成熟dc细胞,用dc诱导培养基重悬后调整浓度至1×10^6个/ml后接种。
如图6所示,未成熟的dc细胞,胞体变粗糙,出现不规则形态,有毛刺状突起,随培养时间的延长逐渐呈现出典型的树突状形态。
2)小鼠dc负载自噬体抗原和dc成熟情况鉴定:
1、往dc培养基内分三组分别加入自噬体抗原10±2ug/ml、肿瘤细胞冻融裂解液和空白对照组,培养48h后,收获成熟dc细胞;
2、取获得的dc细胞用1%bsa-pbs调整细胞密度为1×10^6个/ml,分别移入各自的enpendoff管;
3、加入流式所用的cd11c和cd86、cd80、mhc-i、mhc-ii的荧光标记抗体各1ul,4℃,避光30min孵育;
4、用洗涤液洗涤2遍,去除多余抗体,加入适量pbs重悬细胞,上机进行流式细胞检测,用cellquestplot软件分析所得结果。
如图7所示:加入自噬体、肿瘤细胞冻融裂解液抗原刺激后,dc细胞较空白对照组(kb)明显的上调表达了与dc细胞成熟度相关的cd86、cd80、mhc-i、mhc-ii分子和cd11c分子,表面抗原有效的刺激了dc细胞的成熟,而自噬体抗原处理组(zs)较肿瘤细胞冻融裂解液处理组(lj)的dc细胞的成熟度要更高(流式图中,处于右上格双阳性的细胞比例有比较明显增加),表明本发明实施例所获得的自噬体抗原较肿瘤细胞冻融裂解液能更有效的促进dc细胞的成熟。
实施例3自噬体疫苗诱导的特异性免疫应答实验
1)小鼠外周血源性淋巴细胞的制备
1、将小鼠固定后,通过眼眶采血采集小鼠外周血约1ml,放入预先涂有100uledta抗凝剂的5ml离心管中;
2、取一支15ml离心管,室温下加入3ml小鼠淋巴细胞分离液后,将所获得的抗凝血样本加于分离液液面上,400g,20min,室温离心;
3、用1ml枪头小心吸出分离液上层以上5mm的上清,弃去;再用1ml枪头小心吸取乳白色的淋巴细胞层至新离心管内;
4、离心管内加入适量的细胞洗涤液混匀,300g,10min,室温离心;
5、重复上述步骤洗涤后,即可得到分离好的外周血源性淋巴细胞;
2)小鼠外周血源性t淋巴细胞的诱导和维持培养
1、将分离得到的外周血源性淋巴细胞用含40ng/mlil-2和10%胎牛血清的1640培养基(t淋巴细胞培养液)重悬,置于37℃,5%二氧化碳的培养箱中静置培养;
2、每3-5天换一次液,加入新鲜的t淋巴细胞培养液,即可完成对t淋巴细胞的维持培养;
3)磁珠分选小鼠cd8+t淋巴细胞
1、取生长良好的t淋巴细胞(以1×10^7个为例),300g,10min,室温离心,弃上清;
2、加入2mlbuffer重悬,洗涤,300g,10min,室温离心,弃上清;
3、加入约40ulbuffer重悬pbmc;
4、加入10ulcd8+t细胞bilton-抗体,充分混匀,4℃,孵育15min;
5、加入30ulbuffer,再加入20ulcd8+t细胞microbeadcocktail,充分混匀,4摄氏度,孵育15min;
6、加入2mlbuffer洗涤细胞,300g,10分钟,弃上清;
7、用500ulbuffer重悬细胞,过磁力柱分选,即可得到小鼠cd8+t淋巴细胞;
4)成熟dc促cd8+t细胞增殖(cck8法)
1、将前所获得的自噬体负载和肿瘤细胞冻融裂解液负载所得的成熟dc,调整dc细胞密度,使得每孔有1×10^3个成熟的dc细胞,接种到96孔板中,每个样本设置3个复孔;
2、分别按成熟dc:cd8+t淋巴细胞比例为1:50;1:20;1:10的比例添加cd8+t淋巴细胞,混匀;调整每孔细胞悬液体积为100ul;
3、按上述细胞数量和浓度设置各自对应的只有成熟dc细胞、只有t淋巴细胞和只有培养基的孔,同样设置3个复孔,调整悬液体积为100ul。每孔加入10ulcck8后轻轻敲击培养板帮助混匀,于37℃培养箱中进行共培养;
4、于共培养培养72h时测定各孔在450nm处的吸光度;
5、将上述所得到的各组3复孔吸光度值取平均数后,按增值率计算公式:增殖率=(实验组od-dc对照孔od)/(t对照组od-培养基对照组od)×100%,即可得到增殖倍数结果。
如图8所示,自噬体负载的成熟dc细胞在不同的激活比下(dc比cd8+t淋巴细胞)均能较肿瘤细胞冻融裂解液负载的成熟dc细胞能更好的促进cd8+t淋巴细胞的增殖;且在激活比为1:10时的增殖倍数为最高,为后续的激活比的设定提供了依据,表明本发明实施例所获得的肿瘤自噬体抗原较传统肿瘤细胞冻融裂解液抗原在同等条件下,更能有效的促进cd8+t淋巴细胞的增殖。
5)肿瘤细胞的免疫杀伤
1、计数合适数量的靶细胞,加入2umol/l的cfse,37摄氏度,孵育30min;
2、1000rpm,5min离心,用pbs清洗三遍;
3、将分别用:pbs+dc、肿瘤细胞冻融裂解液抗原+dc、肿瘤自噬体抗原+dc,激活的cd8+效应t细胞,与靶细胞混合,于37℃培养箱中进行共培养,5h;
4、取出细胞,用pbs洗三次;
5、添加pi工作液5ul,避光孵育30min;
6、流式检测;
7、将上述流式检测所获得的数据,按杀伤效率计算公式:杀伤效率=pi阳性cfse阳性细胞数/(pi阴性cfse阳性+pi阳性cfse阳性细胞数)×100%,得到杀伤率的结果。
如图9所示:较阴性对照而言,自噬体抗原组和肿瘤细胞冻融裂解液抗原组,均能有对靶细胞的有效的杀伤作用,而比较肿瘤细胞冻融裂解液抗原组,自噬体抗原组诱导的效应t细胞的杀伤效率要更胜一筹,表明本发明的方法获得的肿瘤细胞较传统肿瘤细胞冻融裂解液抗原更能有效的促进效应t细胞杀伤靶细胞。
以上对本发明的具体实施例进行了详细描述,但其只是作为范例,本发明并不限制于以上描述的具体实施例。对于本领域技术人员而言,任何对本发明进行的等同修改和替代也都在本发明的范畴之中。因此,在不脱离本发明的精神和范围下所作的均等变换和修改,都应涵盖在本发明的范围内。
- 还没有人留言评论。精彩留言会获得点赞!