本申请是申请日为2011年12月30日的题为“岩藻糖基转移酶及其应用”的中国专利申请号201180065581.1的分案申请。本发明涉及一种新型岩藻糖基转移酶及其应用。
背景技术:
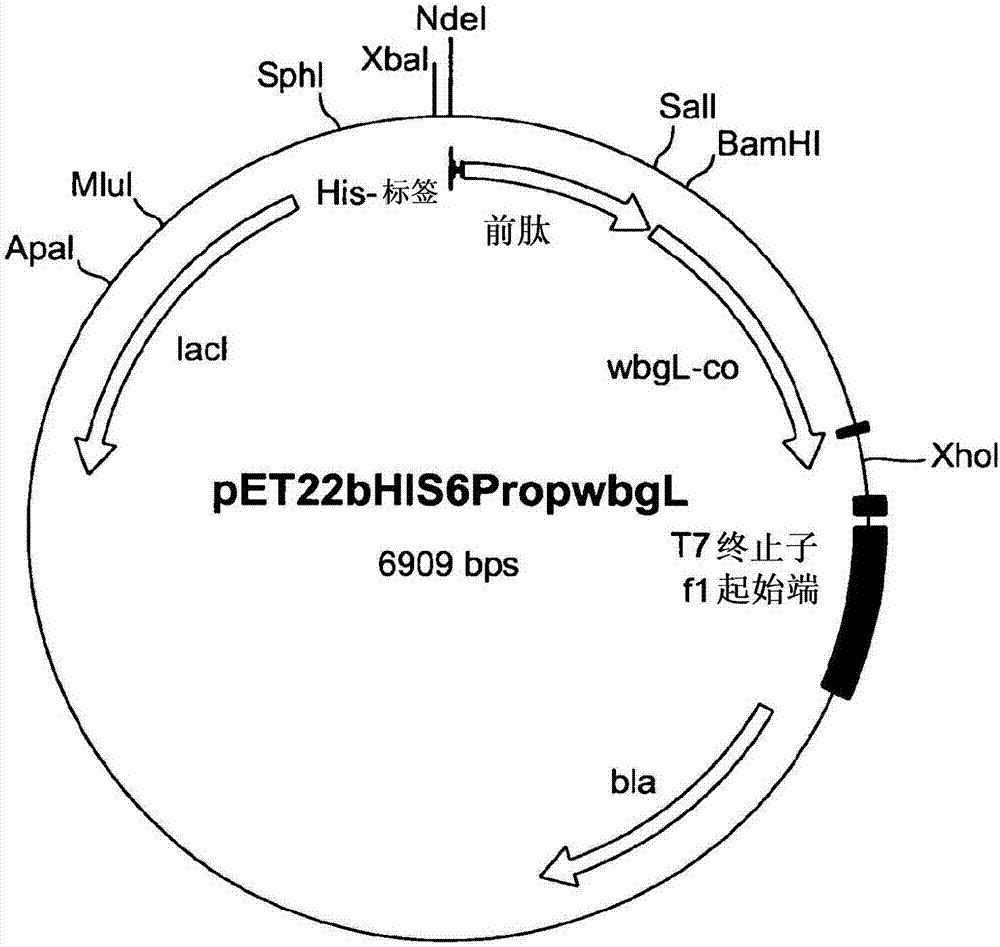
:许多(糖)蛋白、(糖)脂、或寡糖需要特定岩藻糖基化结构的存在,以呈现特定的生物活性。例如,许多细胞间的识别机制需要岩藻糖基化寡糖:例如,为了连接细胞粘附分子如l-选择蛋白,特定的细胞结构必须包括岩藻糖基化碳水化合物。具有生物功能的岩藻糖基化结构的另一个实例是形成abo血型系统的结构。此外,治疗性(糖)蛋白是发展最快的一类药物试剂,因此它们的药物代谢动力学性质以及稳定性归于它们的聚糖。由于它们的复杂性以及固有的化学性质,糖结合物(glycoconjugate)的化学合成是一项主要的挑战,并且具有大量困难。不同于针对自动合成仪的可商购的蛋白质和核酸,聚糖(还)不能使用一般的商业系统合成,更不必说糖结合物。除了对于控制立体化学的需求之外,特定连接键的形成仍然很困难。考虑到与糖结合物的化学合成或结合的酶/化学合成相关的复杂性,近年来的方法已使用糖基转移酶来酶促合成包括寡糖残基的(糖)蛋白以及(糖)脂。属于糖基转移酶的酶家族中的岩藻糖基转移酶,广泛地在脊椎动物、无脊椎动物、植物、以及细菌中表达。它们催化岩藻糖残基从供体(通常是二磷酸鸟苷岩藻糖(gdp-岩藻糖))转移到包括寡糖、(糖)蛋白、以及(糖)脂的受体。由此得到的岩藻糖基化受体底物涉及各种生物和病理过程。基于岩藻糖的添加部位,岩藻糖基转移酶被分类为:α-1,2-岩藻糖基转移酶、α-1,3/4-岩藻糖基转移酶、以及o-岩藻糖基转移酶。一些α-1,2-岩藻糖基转移酶已经被识别,例如,在幽门螺杆菌(helicobacterpylori)和大肠杆菌等细菌中、在哺乳动物、漂亮新小杆线虫(caenorhabditiselegans)和曼森血吸虫(schistosomamansoni)中、以及在植物中的α-1,2-岩藻糖基转移酶。这些酶的大多数或者不能在细菌系统中以活性形式表达、或者不能使用乳糖作为受体。在哺乳动物中,gdp-岩藻糖通过从头合成(denovosynthesis)和补救途径在细胞质中合成。随着从头合成,通过两种酶将gdp-甘露糖转化为gdp-岩藻糖,同时补救途径使用游离细胞质岩藻糖作为底物。在细胞中,gdp-岩藻糖浓缩成囊泡并被岩藻糖基转移酶识别为供体底物。然而,真核糖基转移酶的异源功能表达,尤其证明了岩藻糖基转移酶在原核表达系统中表达困难。哺乳动物,尤其是来源于原核来源的人类寡糖如hmo是未知的,因而发现了使得这些寡糖极其不同的糖基转移酶。由于许多商业上重要的寡糖、(糖)蛋白、以及(糖)脂的生物活性取决于特定岩藻糖残基的存在,因此现有技术需要有效地合成或产生具有期望的寡糖残基的糖结合物。技术实现要素:因此,本发明的目的是提供工具以及方法,通过所述工具和方法,岩藻糖基化底物可以以高效、省时、以及节省成本的方式进行生产,以获得高产量的期望底物。根据本发明,尤其是通过提供一种多核苷酸达到了该目的,所述多核苷酸可以是,例如,分离的、重组的或合成的编码具有α-1,2-岩藻糖基转移酶活性的多肽的多核苷酸,并且包括选自以下组中的序列或由选自以下组中的序列组成,该组由以下各项组成:a)随附序列表中的seqidno.1;b)与seqidno.1互补的核酸序列;c)在严格条件下与a)和b)中限定的核酸序列或它们的互补链杂交的核酸序列。根据本发明的多核苷酸(参见seqidno.1)是大肠杆菌菌种血清群o126的岩藻糖基转移酶。由于通过使用最新识别的岩藻糖基转移酶可以进行不能天然发生在源生物体中的反应,它们具有出乎意料的效果:在本发明的范围内,已经发现上述识别的α-1,2-岩藻糖基转移酶能够使用乳糖作为底物,并且能够产生岩藻糖基化寡糖,尤其是2’-岩藻糖基乳糖。至今,还没有已知的从细菌中分离的α-1,2-岩藻糖基转移酶显示使用乳糖作为天然底物用于产生岩藻糖基乳糖。因而,用于产生岩藻糖基化寡糖的最新识别的岩藻糖基转移酶的适用性极为出乎意料,因此,它的应用代表一种容易地、高效地、以及节省成本地生产诸如母乳寡糖(hmo)(如岩藻糖基乳糖)的优良工具。当今,已经表征了属于hmo的多于80种化合物的结构;它们代表一类具有益生元(prebiotic)作用的寡糖复合物。此外,hmo与上皮表位的同源结构是其对抗细菌病原体的保护性的原因。在婴儿胃肠道中,hmo选择性地提供选定的细菌菌种生长的养分,因而致使母乳喂养婴儿中的独特肠微生物群的生长。直到现在,因为这些寡糖的结构复杂性阻碍它们的合成制备,母乳仍然是hmo的主要来源。因此,需要容易地、轻松地得到hmo,这一需求可以通过使用本文中给出的出乎意料地适用的岩藻糖基转移酶提供。根据本发明,术语“多核苷酸”一般指的是任一种多核糖核苷酸或多脱氧核糖核苷酸,所述多核糖核苷酸或多脱氧核糖核苷酸可以是未改变的rna或dna、或改变的rna或dna。“多核苷酸”包括,但不限于,单链和双链dna,单链区和双链区混合的dna,或单链区、双链区以及三链区混合的dna,单链和双链rna,以及单链区和双链区混合的rna,包括dna和rna的杂交分子,所述杂交分子可以是单链的或更典型的是双链区、或三链区、或单链区和双链区的混合体。此外,本文中使用的“多核苷酸”指的是包括rna、或dna、或rna和dna的三链区。在此区域的链可以来自同一分子或来自不同的分子。该区域可以包括所有分子中的一个或多个,但更典型地只包括一些分子的一个区。三螺旋区的一个分子通常是寡核苷酸。如在本文中所使用的,术语“多核苷酸”还包括如上所述的包含一个或多个改变的碱基的dna或rna。因此,具有出于稳定性或其它原因改变的主链的dna或rna是本文中意指的术语“多核苷酸”。此外,仅举两个实例,包括稀有碱基如肌苷、或者修饰碱基如三苯甲基化碱基的dna或rna是本文中所使用的术语多核苷酸。将理解已经对本领域技术人员熟知的用于许多有用目的的dna和rna进行了大量改变。如在本文中使用的术语“多核苷酸”包括如化学地、酶促地、或代谢地改变的多核苷酸的形式,以及病毒和细胞(包括,例如简单和复杂细胞)特性的dna和rna的化学形式。另外,“多核苷酸”还包括短的多核苷酸,通常指的是寡核苷酸。“多肽”指的是包括两个或更多氨基酸通过肽键或改变的肽键彼此连接的任一种肽或蛋白质。“多肽”不仅指的是短链,一般指的是肽、寡肽以及寡聚物;还指的是较长的链,一般指的是蛋白质。多肽可以包括除20种基因编码的氨基酸之外的氨基酸。“多肽”包括通过自然过程如加工以及其它转录后修饰,以及通过化学修饰技术改变的那些。在基础用书(basictext)和在详细的专著中、以及在长篇研究文献中很好地描述了这些改变,并且本领域的技术人员已经熟知。将理解相同类型的改变可以以相同或不同的程度出现在给定的多肽的一些位点。另外,给定的多肽可以包括许多类型的修饰。修饰可以发生在多肽的任何部位,包括肽主链、氨基酸侧链、以及氨基端或羧基端。修饰包括,例如,乙酰化、酰化、adp-核糖基化、酰胺化、黄素的共价连接、血红素部分的共价连接、核苷酸或核苷酸衍生物的共价连接、脂质或脂质衍生物的共价连接、磷脂酰肌醇的共价连接、交联、环化、二硫键形成、脱甲基化、共价交联的形成、焦谷氨酸的形成、甲酰化、γ-羧基化、糖基化、gpi锚定点(anchor)的形成、羟基化、碘化、甲基化、豆蔻酰化、氧化、蛋白水解处理、磷酰化、异戊烯化、外消旋化、脂质连接、硫酸化、谷氨酸残基的γ-羧基化、羟基化和adp-核糖基化、硒酰化、转运rna介导的氨基酸至蛋白质的添加(如精氨酰化、遍在蛋白化)。多肽可以是支链、或带有或不带有支链的环。环、支链、以及支链环状的多肽可以由转录后的天然过程产生,并且还可以完全通过合成法制备。“分离的”意味着从它的天然状态“通过人工”改变,即,如果天然发生,则已经从它的原始环境改变或转移、或即改变又转移。例如,在活有机体中天然存在的多核苷酸或多肽不是“分离的”,但是从其自然状态的共存物质中分离得到的相同多核苷酸或多肽是“分离的”,如在本文中使用的术语。类似地,“合成”序列,如在本文中使用的术语,意味着任一种通过合成产生而不是直接从天然来源分离的序列。“重组”意味着通过从一种物种中移植或剪接基因至不同种类的宿主有机体细胞中而制备的基因工程dna。这种dna成为宿主基因构成(makeup)的部分并被复制。本文中使用的术语“编码多肽的多核苷酸”包括以下多核苷酸,所述多核苷酸包括编码本发明的多肽,尤其是具有seqidno.2给出的氨基酸序列α-1,2-岩藻糖基转移酶的序列。术语还包括以下多核苷酸,所述多核苷酸包括编码多肽(例如,通过整合噬菌体、或插入序列、或编译而间断的)的单个连续区或不连续区、以及还可以包括编码序列和/或非编码序列的附加区域。本文中使用的术语“变体”是分别不同于参照多核苷酸或多肽的多核苷酸或多肽,但是保留基本性质。多核苷酸的典型变体的核苷酸序列不同于参照多核苷酸。变体的核苷酸序列的变化可以改变或者可以不改变由参照多核苷酸编码的氨基酸序列。核苷酸变化可以产生如下所述的由参照序列编码的多肽中氨基酸的取代、添加、缺失、融合、以及截短。多肽的典型变体的氨基酸序列不同于参照多肽。一般地,不同是有限的,使得参照多肽与变体的序列接近全部相似,并且在许多区域完全相同。变体和参照多肽可以通过以任意组合的一个或多个取代、添加、缺失而具有不同的氨基酸序列。取代或插入氨基酸残基可以是或可以不是由基因密码编码的氨基酸残基。多核苷酸或多肽的变体可以是天然产生的如等位基因变体,或者可以是未被认为是天然产生的变体。多核苷酸和多肽的非天然产生的变体可以通过诱变技术、通过直接合成、以及通过本领域技术人员已知的其他的重组方法制备。术语“α-1,2-岩藻糖基转移酶”、或“岩藻糖基转移酶”、或编码“α-1,2-岩藻糖基转移酶”或“岩藻糖基转移酶”的核酸/多核苷酸指的是催化岩藻糖从供体底物,例如gdp-岩藻糖,以α-1,2连接键转移至受体分子的糖基转移酶。受体分子可以是碳水化合物、寡糖、蛋白或糖蛋白、或脂质或糖脂;并且可以是,例如,n-乙酰葡糖胺、n-乙酰乳糖胺、半乳糖、岩藻糖、唾液酸、葡萄糖、乳糖、或它们的任意组合。在本发明范围内,那些术语中包括的核酸/多核苷酸、以及多肽的多态变体、等位基因、突变体、以及异种同源体,具有与由seqidno.1的核酸编码的多肽或seqidno.2的氨基酸序列大于约60%的氨基酸序列同一性,65%、70%、75%、80%、85%、90%,优选91%、92%、93%、94%、95%、96%、97%、98%、或99%、或更高的氨基酸序列同一性的氨基酸序列,优选在至少约25、50、100、200、500、1000、或更多氨基酸的区域内。此外,可以通过肽序列的添加或缺失来改变α-1,2-岩藻糖基转移酶多肽,以改变它的活性。例如,可以将多肽序列融合到α-1,2-岩藻糖基转移酶多肽中,以得到增加的酶活性。可替换地,可以删除氨基酸以去除或改变蛋白活性。可以将蛋白质改变为缺乏α-1,2-岩藻糖基转移酶的酶促活性,但是仍然保留它的三维结构。这种改变将有利于针对α-1,2-岩藻糖基转移酶多肽的抗体的发展。此外,α-1,2-岩藻糖基转移酶基因产物可以包括代表功能等价基因产物的蛋白或多肽。这种等价α-1,2-岩藻糖基转移酶基因产物可以包括在由上述α-1,2-岩藻糖基转移酶基因序列编码的氨基酸序列中的氨基酸残基的缺失、添加或取代,但是这会导致沉默(silent)的变化,由此产生功能等价α-1,2-岩藻糖基转移酶基因产物。氨基酸取代可以基于相关残基的极性、电荷、溶解性、疏水性、亲水性和/或两亲性的相似性进行。例如,非极性(疏水)氨基酸包括丙胺酸、亮氨酸、异亮氨酸、缬氨酸、脯氨酸、苯丙氨酸、色氨酸、以及甲硫氨酸;平面(planar)中性氨基酸包括甘氨酸、丝氨酸、苏氨酸、半胱氨酸、酪氨酸、天冬酰胺、以及谷氨酰胺;正电荷(碱性)氨基酸包括精氨酸、赖氨酸、以及组氨酸;以及负电荷(酸性)氨基酸包括天冬氨酸和谷氨酸。在本发明的上下文中,这里使用的“功能等价”指的是能够呈现与由上述α-1,2-岩藻糖基转移酶基因序列编码的内源α-1,2-岩藻糖基转移酶基因产物基本相似的体内活性的多肽,通过许多标准中的任一标准进行判断,所述标准包括,但不限于,抗原性,即结合至抗-α-1,2-岩藻糖基转移酶抗体的能力;免疫原性,即产生能够结合α-1,2-岩藻糖基转移酶蛋白或多肽的抗体的能力;以及酶促活性。包含在本发明的范围内的是在翻译期间或翻译后不同地改变的α-1,2-岩藻糖基转移酶蛋白、多肽、及衍生物(包括片段)。此外,非典型的氨基酸或化学氨基酸类似物可以作为取代或添加被引入α-1,2-岩藻糖基转移酶多肽序列中。α-1,2-岩藻糖基转移酶多肽可以通过使用本领域已知的重组dna技术产生。本领域技术人员熟知的方法可以用于构建包含α-1,2-岩藻糖基转移酶编码序列的表达载体,以及合适的转录翻译控制信号。这些方法包括,例如,体外重组dna技术、合成技术、以及体内遗传重组。参见,例如,sambrooketal.molecularcloning:alaboratorymanual,2nded.,coldspringharborlaboratorypress,coldspringharbor,n.y.(1989)中描述的技术.本文中使用的、以及如在现有技术中通常理解的术语“寡糖”指的是包含小数量的,典型的是3至10个简单糖类(即单糖)的糖类聚合物。根据本发明的另一方面,提供的载体包括以上给出的编码具有α-1,2-岩藻糖基转移酶活性的多肽的核酸序列,其中,所述核酸序列可操作地连接至由载体转化的宿主细胞识别的控制序列。在特别优选的实施方式中,载体是一种表达载体,并且根据本发明的另一方面,载体可以以质粒、粘粒、噬菌体、脂质体、或病毒的形式存在。另外,本发明涉及包含由如上所述的、根据本发明的多核苷酸组成的序列的宿主细胞,其中所述序列是宿主细胞的外源序列,并且其中,将所述序列整合在宿主细胞的基因组中。因此,“宿主细胞的外源”意味着在所述宿主细胞中序列不能天然产生,即,序列相对于宿主细胞是异源的。异源序列可以稳定地引入宿主细胞的基因组中,例如,通过转染、转化、或转导,其中可以根据将引入宿主细胞中的序列施用技术。本领域的技术人员已知各种技术,并且,例如公开在sambrooketal.,1989(同上)中。因此,已引入异源序列的宿主细胞将产生根据本发明的多核苷酸编码的异源蛋白。对于重组生产,宿主细胞可以并入遗传工程的表达系统、或其部分、或本发明的多核苷酸。将多核苷酸引入宿主细胞可以通过许多标准实验室手册如davisetal.basicmethodsinmolecularbiology,(1986)和sambrooketal.,1989(同上)中描述的方法实现。因此,根据本发明的多核苷酸可以,例如,包括在将稳定地转化/转染至宿主细胞中的载体中。在该载体中,本发明的多核苷酸在例如诱导型启动子的控制下,以使基因/多核苷酸的表达可以特异性地靶向,并且,如果期望,基因可以以该方式过表达。大量的表达系统可以用于生产本发明的多肽。这种载体,除了其它,还包括染色体的、游离型以及源于病毒的载体,例如源于细菌质粒的载体、源于噬菌体的载体、源于转座子的载体、源于酵母附加体的载体、源于插入元件的载体、源于酵母染色体元件的载体、源于病毒的载体、以及源于它们的组合的载体(如源于质粒和噬菌体基因元件的载体、如源于粘粒和噬菌粒的载体)。表达系统构建体可以包括调节以及引起表达的控制区。一般地,在这一方面,适合保持、增殖、或表达多核苷酸和/或在宿主中表达多肽的任何系统或载体可以用于表达。合适的dna序列可以通过多种已知的和常规技术中的任一种插入表达系统中,如,例如,参见以上sambrooketal给出的那些技术。因此,本发明还涉及由选自以下组中的氨基酸序列组成的具有α-1,2-岩藻糖基转移酶活性的分离多肽,该组由以下各项组成:a)在seqidno.2中所示的氨基酸序列;b)在seqidno.2中所示的氨基酸序列等位基因变体的氨基酸序列,其中所述等位基因变体由在严格条件下杂交至在seqidno.1中所示的核酸分子的相反链的核酸分子编码;c)在seqidno.2中所示的氨基酸序列的同源体的氨基酸序列,其中所述同源体由在严格条件下杂交至在seqidno.1中所示的核酸分子的相反链的核酸分子编码;以及d)在seqidno.2中所示的氨基酸序列的片段,其中所述片段包括至少10个连续的氨基酸,并且其中所述片段具有α-1,2-岩藻糖基转移酶活性。术语“严格条件”指的是探针将杂交至其靶序列而不是其它序列的条件。严格条件取决于序列,并且在不同的环境下将不同。较长的序列具体地在较高的温度下杂交。一般地,在限定的离子强度和ph下,对于特定序列的严格条件选择为比热溶解温度(tm)约低15℃。tm是在平衡状态下将50%的互补于靶序列的探针杂交至靶序列的温度(在限定的离子强度、ph、以及核酸浓度下)。示例性的严格杂交条件如下:50%的甲酰胺、5xssc、以及1%的sds,在42℃下温育;或5xssc、1%的sds,在65℃下温育;并用0.2xssc以及0.1%的sds,在65℃下冲洗。另外,本发明涉及包含如上所定义的载体,或者包含根据本发明的多核苷酸作为引入宿主细胞基因组中的异源序列的宿主细胞,尤其是选自由以下各项组成的组中的宿主细胞:包括酵母菌的真菌、细菌、昆虫、动物细胞和植物细胞。特别地优选宿主细胞为大肠杆菌细胞。如在本文中所使用的,术语“宿主细胞”目前被定义为已转化或转染的细胞,或者能够被外源多核苷酸序列转化或转染的细胞。各种宿主表达载体系统可以用于表达本发明的α-1,2-岩藻糖基转移酶基因密码序列。这种宿主表达系统代表运载体(vehicle),通过该运载体可以生成感兴趣的编码序列,随后纯化,还代表当通过适当的核苷酸编码序列转化或转染时,在原位产生本发明的α-1,2-岩藻糖基转移酶基因产物的细胞。许多合适的表达系统以及宿主可以在,例如,涉及来自幽门螺杆菌的α-1,2-岩藻糖基转移酶的wo/0026383和ep1243647中找到,在此明确地引用了此出版物。根据本发明的另一方面,编码具有α-1,2-岩藻糖基转移酶活性的多肽的核酸适合各细胞的密码子使用。本发明进一步涉及根据本发明的多核苷酸、载体、或多肽分别用于生产岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂的应用。因此,根据应用的一个方面,所述岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂的生产通过编码α-1,2-岩藻糖基转移酶的多核苷酸的异源或同源表达进行。根据应用的另一方面,岩藻糖基化寡糖是已知来自于母乳中的寡糖,如2’-岩藻糖基乳糖。本发明还涉及用于生产岩藻糖基化寡糖、(糖)蛋白、以及(糖)脂的方法,该方法包括以下步骤:a.提供根据本发明的具有α-1,2-岩藻糖基转移酶活性的多肽,b.在多肽催化岩藻糖残基从供体底物转移至受体底物的条件下,使步骤a中的具有α-1,2-岩藻糖基转移酶活性的多肽与包括供体底物的混合物接触,该供体底物包括岩藻糖残基,并且受体底物包括单糖或寡糖、(糖)蛋白、或(糖)脂,由此生产岩藻糖基化寡糖、(糖)蛋白、或(糖)脂。根据本发明,用于生产岩藻糖基化寡糖的方法可以在无细胞系统或含细胞系统中进行。在充足条件下使底物与α-1,2-岩藻糖基转移酶多肽反应足够长的时间,以使得酶促产物能够形成。应理解,无论是无细胞系统还是基于细胞的系统,这些条件将随着底物和酶的量和纯度而改变。这些变量将易于由本领域的技术人员调整。在无细胞系统中,根据本发明的多肽、受体底物、供体底物在这种情况下可以是其它反应混合成分,包括其它糖基转移酶,并且辅酶(accessoryenzyme)通过在含水反应介质中与混合物结合。在溶液中可以使用游离酶,或者它们可以结合或固定到载体如聚合物上,并且底物可以加入载体中。载体可以,例如,堆积在柱中。用于合成岩藻糖基化寡糖的含细胞系统可以包括重组改变的宿主细胞。因此,本发明还涉及用于生产岩藻糖基化寡糖、(糖)蛋白、以及(糖)脂的方法,包括以下步骤:a.在允许岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂生产,以及允许具有α-1,2-岩藻糖基转移酶活性的多肽表达的合适营养条件下,培养上述宿主细胞;b.与步骤a同时或在步骤a之后,提供包括岩藻糖残基的供体底物以及包括寡糖、(糖)蛋白、或(糖)脂的受体底物,使得在所述宿主细胞中表达的α-1,2-岩藻糖基转移酶催化岩藻糖残基从供体底物转移到受体底物,由此生产岩藻糖基化寡糖、(糖)蛋白、或(糖)脂;以及c.从宿主细胞或它的生长介质中分离所述岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂。根据本发明的方法,供体底物可以是gdp-岩藻糖。特别优选的供体底物为gdp-岩藻糖。根据本发明的一个方面,受体底物选自n-乙酰葡糖胺、n-乙酰乳糖胺、半乳糖、岩藻糖、唾液酸、葡萄糖、乳糖、或它们的任意组合。特别地,优选乳糖为受体底物。本文中使用的术语“底物”意味着可以通过根据本发明的多肽作用以生产岩藻糖基化寡糖、(糖)蛋白、或(糖)脂的任何物质或不同物质的组合。在充足条件下使底物与α-1,2-岩藻糖基转移酶多肽反应足够长的时间,以使得酶产物能够生成。无论系统是无细胞系统还是基于细胞的系统,这些条件将随底物和酶的量以及纯度而变化。这些变量将易于由本领域的技术人员调整。根据本发明的方法的一个方面,供体底物由通过宿主细胞产生的步骤b提供。这样做,例如,使得被加入到宿主细胞中的岩藻糖转化成gdp-岩藻糖的酶同时在宿主细胞内表达。这种酶可以是,例如,双功能的岩藻糖激酶/岩藻糖-1-磷酸鸟苷酰基转移酶(如脆弱拟杆菌(bacteroidesfragilis)中的fkp)、或单独的岩藻糖激酶中的一种以及单独的岩藻糖-1-磷酸鸟苷酰基转移酶中的一种(如,来自包括智人(homosapiens)、猪(susscrofa)、以及褐家鼠(rattusnorvegicus)等一些物种中的那些已知)的组合。可替换地,在步骤b中,供体底物可以添加到培养基/宿主细胞中,或者通过细胞自身的代谢产生。在还进一步的实施方式中,本发明还涉及包括以下步骤的方法:a.在合适的营养条件下,培养转化或转染的宿主细胞以包括选自以下各项中的核酸序列:i)随附序列表中seqidno.1的核酸序列;ii)与seqidno.1互补的核酸序列;以及iii)在严格条件下与在i)和ii)中限定的核酸序列或它们的互补链杂交的核酸序列,使得选自i)、ii)、和iii)中的核酸序列被表达为具有α-1,2-岩藻糖基转移酶活性的多肽。b.与步骤a同时或在步骤a之后,提供包括岩藻糖残基的供体底物以及包括寡糖、(糖)蛋白、或(糖)脂的受体底物,使得在所述宿主细胞中表达的α-1,2-岩藻糖基转移酶催化岩藻糖残基从供体底物转移到受体底物,由此生产岩藻糖基化寡糖、(糖)蛋白、或(糖)脂;以及c.从宿主细胞或它的生长培养基中分离所述岩藻糖及化寡糖、(糖)蛋白和/或(糖)脂。在根据本发明的方法中,在宿主细胞中表达的肽显示出α-1,2-岩藻糖基转移酶活性,因此,将岩藻糖残基从供体,例如,二磷酸鸟苷岩藻糖(gdp-岩藻糖)转移到包括寡糖、(糖)蛋白、以及(糖)脂的受体。以这种方式,得到的岩藻糖基化受体底物可以用作食品添加剂,用于婴儿食品的补充物,或者用作治疗或药物活性物质。通过该新方法,可以容易、有效地提供岩藻糖基化产物,而不需要复杂、耗时和消耗成本的合成过程。如本文中使用的术语“分离”意味着从基因表达系统中获取、收集、或分离通过根据本发明的α-1,2-岩藻糖基转移酶产生的产物。因此,本发明还涉及通过根据本发明的方法获得的岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂,还涉及上述用于生产岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂的多核苷酸、载体、或多肽的应用。根据又一个实施方式,所述岩藻糖基化寡糖、(糖)蛋白和/或(糖)脂的生产利用编码α-1,2-岩藻糖基转移酶的多核苷酸的异源或同源(过)表达进行。除非另有定义,本文中使用的所有技术和科学术语与本发明所属领域的任何普通技术人员通常理解的具有相同的意义。一般地,本文中使用的术语以及在细胞培养、分子遗传学、有机化学、核酸化学以及上述和下述杂交中的实验室规程是本领域熟知的,和通常使用的。对于核酸和肽合成使用标准方法。一般地,酶促反应和纯化步骤根据生产商的说明书进行。本发明还涵盖了本文中公开的多核苷酸序列的片段。实施方式和附图的描述给出进一步的优势。上述特征以及以下将要说明的特征不仅可以以各自特定组合使用,还可以以其它组合或单独使用,而没有背离本发明的范围。附图说明本发明的一些实施方式描述在图形中,并且借助以下描述进行更加详细的说明。在附图中:图1a示出了编码来自大肠杆菌o126的α-1,2-岩藻糖基转移酶wbgl的基因的dna和氨基酸序列;图1b示出了pet22bhis6propwbgl的载体图谱,即,将来自大肠杆菌o126的编码新型α-1,2-岩藻糖基转移酶wbgl的密码子优化基因wbgl克隆至pet22b(+)(novagen,darmstadt,germany,viabamhi/xhoi)中,在经由ndei/bamhi酶切之前,向其中加入编码n-端his-标签(his-tag)的18bp以及来自猪葡萄球菌脂肪酶的前肽的621bp(参见以下描述的);图1c示出了pacyc-wbgl的载体图谱,即,将来自大肠杆菌o126的编码新型α-1,2-岩藻糖基转移酶wbgl的密码子优化基因wbgl克隆至pacycduet-1(novagenviancoi/bamhi)中;图2示出了由imacni2+-ntasepharose柱纯化wbgl的色谱图(蓝色:在280nm下的吸收;红色:电导率;包含从柱中洗脱的his6-前肽-wbgl的部分被称为imac池);图3示出了在大肠杆菌jm109(de3)pet22bhis6propwbgl的粗提物和不溶部分中、以及在his6-前肽-wbgl的纯化物的洗涤部分中his6-前肽-wbgl表达体的sds-page分析,以及纯化的his6-前肽-wbgl的sds-page分析;图4示出了用dab底物(rochediagnostics,mannheim,germany)温育之后,使用结合到辣根过氧化物酶(hrp)(rochediagnostics,mannheim,germany)上的单克隆抗-his6抗体检测免疫印迹上的his6-前肽-wbgl;图5示出了用0.4mm的gdp-β-l-岩藻糖和5mm的乳糖纯化的his6-前肽-wbgl的连续分光光度测定,其中活性可以通过由nadh的氧化引起的在340nm处吸收的减弱测定;图6示出了由通过hpaec-pad的wbgl反应制备2’-岩藻糖基乳糖的测定,在反应初期不存在2’-岩藻糖基乳糖(a),当wbgl与2mm的gdp-β-l-岩藻糖和5mm乳糖反应1小时后产生2’-岩藻糖基乳糖(b);图7示出了通过毛细管电泳法对起始于2mm的gdp-岩藻糖、5mm乳糖、以及wbgl反应混合物的分析,其中在反应初期,仅能检测到gdp-β-l-岩藻糖(a);反应16小时之后,仅能检测到由wbgl活性释放的gdp的降解产物鸟苷(b);图8示出了在50mm的tris-hcl缓冲液中在不同ph值(ph6.8至8.4)下wbgl的相对活性;图9示出了在起始于2mm的gdp-β-l-岩藻糖以及5mm乳糖的wbgl反应的16小时反应过程中,在不同时间点测定的2’-岩藻糖基乳糖的浓度增加;图10示出了通过以水作为洗脱液的phenomenexrezexrcmca2+柱(在80℃下,以0.6ml/min洗脱30分钟)分离,并通过折光率检测器(shimadzu,germany)进行检测得到的hplc色谱图(a);以及通过以乙腈/水(68:32)作为洗脱液的reprosilcarbohydrate柱(5μm250x4.6mm)(在35℃下,以1.4ml/min洗脱20分钟)分离,并通过折光率检测器(shimadzu,germany)进行检测得到的hplc色谱图(b);图11示出了由wbgl产生的2’-岩藻糖基乳糖的nmr核磁共振光谱。具体实施方式实施例基因克隆为了推定的α-1,2-岩藻糖基转移酶wbgl的细胞质表达,对表达载体pet22b(+)(novagen,darmstadt,germany)进行改性。因此,使用限制性内切酶ndei和bamhi切割诱导大肠杆菌中周质表达的pelb前导序列。猪葡萄球菌脂肪酶的前肽基因序列(seqidno.3)(sauerzapfe,b.,d.j.namdjou,etal.(2008):"characterizationofrecombinantfusionconstructsofhumanbeta-1,4-galactosyltransferase1andthelipasepre-propeptidefromstaphylococcushyicus."journalofmolecularcatalysisb:enzymatic50(2-4):128-140)通过使用引物5`-catatgcaccaccaccaccaccacaatgattcgacaacacaaacaacgac-3`(seqidno.5)以及3′-ggatccgtatggttttttgtcgctcgcttg-5`(seqidno.6)从质粒plgaltδ38扩增,并且融合至产生载体pet22bhis6prop的改性pet22b载体。产生的载体被bamhi和xhoi消化。包括bamhi和xhoi限制位点的基因编码由geneart(geneart)合成,该基因编码用于推定的大肠杆菌o126中的α-1,2-岩藻糖基转移酶wbgl。将α-1,2-岩藻糖基转移酶wbgl连接到载体pet22bhis6prop上产生表达载体pethis6propwbgl(参见图1a),用异丙基硫代半乳糖苷(iptg)诱导pethis6propwbgl后,在大肠杆菌的细胞质中产生his6-前肽-wbgl(513个氨基酸)。可替换地,使用引物5’-gatcaccatgggcagcattattcgtctgcagggtggtc-3’(seqidno.7)和5’-gatcaggatccttagcagctgctatgtttatcaacgttgatcc-3’(seqidno.8)扩增之后,基因wbgl通过ncoi/bamhi克隆到载体pacycduet-1(novagen,darmstadt,germany)以产生载体pacyc-wbgl(见图1b)。使用用于大肠杆菌novablue(novagen,darmstadt,germany)或大肠杆菌top10(invitrogen,darmstadt,germany)进行质粒增殖,以及使用大肠杆菌jm109(de3)(madison,usa)进行his6-前肽-wbgl表达。岩藻糖基转移酶的培养和表达在包含20ml的lb培养基以及100μg/ml的氨苄青霉素的100ml爱伦美氏烧瓶中培养转化株,并且在37℃以及130rpm下温育过夜。在含有1000ml的tb培养基的5l爱伦美氏烧瓶中,在37℃以及80rpm下培养蛋白生产细胞。lacz启动子的诱导通过向培养物中添加iptg至0.1mm的最终浓度进行(od600=0.6-0.8)。温育在25℃下持续20h,并且最终通过离心收获细胞。酶纯化在50mmph7.6的tris-hcl缓冲液中,大肠杆菌细胞的40%(w/w)细胞悬液通过超声破碎(4x15s)。离心(15000rpm,30min)后,保留颗粒(pellet),通过sds-page分析内含体(inclusion)的产生。通过使用先前由100ml50mmph7.6的tris-hcl(缓冲液a)平衡的ni2+-ntasepharose(hilden,germany)将粗提物(15ml)上样到imac柱(0.8cm2×10cm,1.5ml/min)中。经包含0.3mnacl和20mm咪唑的缓冲液b的洗涤步骤后,利用在缓冲液c(50mmtris-hclph7.6,0.3mnacl以及300mm咪唑)中的浓度为300mm的咪唑洗脱蛋白质。所有部分进行蛋白浓度分析以及酶活性检测。将用缓冲液c洗脱的包含活性可溶wbgl的所有部分合并,并将得到的溶液称为imac池(参见图2)。sds-page和免疫印迹检测his6-前肽-wbgl的表达通过sds-page和免疫印迹监测。因此使用根据(paisley,uk)的凝胶灌注(gelcasting)说明书灌注的10%丙烯酰胺凝胶进行sds-page分析。将蛋白样品(40μg)上样在每一个凝胶槽(slot)上。预染蛋白ladderpagerulertm(vilnius,lithuania)用于检测分子质量。用考马斯亮蓝对蛋白凝胶染色(参见图3)。通过使用特定的抗-his6抗体进行免疫印迹,以检测his6propwbgl。因此,大肠杆菌jm109(de3)细胞碎片的样品、粗提物、以及his6-前肽-wbgl的imac部分在sds-page凝胶上分离,并通过nupage印迹转移方案(biorad,münchen,germany)转移到pvdf膜上。用在tbs缓冲液(10mmtris-hcl,150mmnacl,ph7.2)中的3%bsa封闭(block)膜。结合到辣根过氧化酶(hrp)(rochediagnostics,mannheim,germany)的单克隆抗-his6抗体用于特异性结合。经包含0.1%吐温20的tbs缓冲液的洗涤步骤后,用dab底物(rochediagnostics,mannheim,germany)温育印迹,并且通过除去溶液以及添加蒸馏水使hrp反应停止(参见图4)。活性测定通过2’-岩藻糖基乳糖的分光光度测定以及hpaec-pad分析,测定粗提物和imac部分中his6-前肽-wbgl的酶活性。分光光度测定通过使用用于检测释放的gdp(barratt,d.h.,l.barber,etal.(2001)."multiple,distinctisoformsofsucrosesynthaseinpea."plantphysiology127(2):655-664.)的丙酮酸激酶/乳酸脱氢酶系统进行。用于微量滴定板检测的反应混合物包括50mmph7.6tris-hcl、2mmgdp-β-l-岩藻糖、5mm乳糖、1mm磷酸烯醇丙酮酸、1mmdtt、0.25mmnadh、2mmmncl2、5u丙酮酸激酶、以及5u乳酸脱氢酶,其总体积为250μl。通过添加100μl酶溶液开始反应,然后在30℃下置于340nm处。副活性(sideactivity)检测的对照试验在没有受体底物乳糖的情况下进行(参见图5)。酶活性还通过用于2′-岩藻糖基乳糖检测的hpaec-pad分析测定。检测溶液包括2mmgdp-β-l-岩藻糖、5mm乳糖、含2mmmncl2的50mmph7.6tris-hcl、1mmdtt、1u碱性磷酸酶,其总体积为175μl,然后加入175μl粗提物之后在30℃下温育,并纯化酶溶液。通过在95℃下加热5分钟停止反应,在不同的时间点转化率是线性的。使用软件chromeleontm6.40,通过在carbopacpa1柱上的dionexhpaec-pad(dionexcorporation,sunnyvale,ca,usa)分析离心的样品。在30℃下用50mmnaoh进行洗脱(流速:1ml/min,注射量:50μl)。通过商用2`-岩藻糖基乳糖标准曲线测定产生的三糖2`-岩藻糖基乳糖的浓度,并且将浓度用于随后的酶活性的计算(参见图6)。对于hpaec-pad和分光光度测定两者,1u的his6-前肽-wbgl是在标准检测条件下每分钟产生1μmol产物(gdp或2′-岩藻糖基乳糖)的酶量。通过在配有uv检测器的来自beckmancoulter(krefeld,germany)的p/acemdq装置上进行毛细管电泳来分析gdp-β-l-岩藻糖及其消耗量。用于测定活化供体底物gdp-β-l-岩藻糖的样品通过加热(95℃)5分钟停止,并且在15000rpm(rotina35r,hettich,tuttlingen,germany)下离心10分钟。检测通过使用50mm的na2b4o7x10h2o/64mm的硼酸缓冲液(ph8.9)在未经处理的弹性石英毛细管(i.d.75mm,总毛细管长度:57cm,距检测器50cm)上完成。迁移和检测条件分别是:25.8℃下25kv(23ma)以及uv检测波长为254nm。样品通过压力(在0.5psi条件下正向推进5.0秒)注射(参见图7)。gdp-β-l-岩藻糖和产生的鸟苷的识别由商用底物确定。最适ph以及金属离子依赖性为了研究重组his6-前肽-wbgl活性的最适ph值,检测在50mmtris-hcl缓冲液不同的ph值下进行,ph值范围为6.8至8.4。最适ph值为7.6(参见图8)。向标准检测中加入金属离子mn2+以研究金属离子依赖性。所有样品通过上述hapaec-pad进行分析。结果表明,酶不依赖于mn2+离子。动力学分析用于受体底物乳糖和供体底物gdp-β-l-岩藻糖的his6-前肽-wbgl的动力学常数来自于使用上述分光光度测定在变化的底物浓度下的初始速率分析。gdp-β-l-岩藻糖在恒定浓度为10mm的乳糖中的变化范围为0.02mm至4mm,并且在恒定浓度为2mmgdp-β-l-岩藻糖中乳糖的变化范围为0.05mm至40mm。所有数据通过使用sigmaplot10软件(spsssciencesoftwaregmbh,erkrath,germany)根据michaelis-menten方程非线性回归分析测定。表1.重组α-1,2-岩藻糖基转移酶his6-前肽-wbgl的动力学常数底物km值[mm]vmax[mu/mg]vmax/km[mu*mg-1mm-1]r2乳糖5.3170320.9935gdp-岩藻糖0.271676180.9907受体底物光谱研究通过根据上述活性测定的hpaec-pad分析重组his6-前肽-wbgl的底物光谱。测试替代5mm乳糖的不同受体底物以测定相对于乳糖的相对活性。表2.α-1,2-岩藻糖基转移酶his6-前肽-wbgl的受体光谱底物相对活性[%]乳糖100乳果糖124lacnactypⅰ28lacnactypⅱ95d-半乳糖713-岩藻糖基乳糖0β-苄基乳糖50d-galnac0d-galnh20lacdinac0岩藻糖基化化合物的产生用载有合适的岩藻糖基转移酶基因的pacycduet-1转化大肠杆菌细胞bl21(de3)δlaczpdest14-fkppcola-lacy-fucp。菌落在含有合适的抗生素的2yt平板上生长。用5ml过夜培养物(含有抗生素的2yt)培养各菌株,并且将该培养物的各15ml矿物培养基接种至1%。细胞通过使用甘油作为碳源生长,然后使用0.1mmiptg、40mm乳糖在od600=0.5下诱导,再加入30mm乳糖。培养物在30℃以及120rpm下温育。hplc分析监测2’-岩藻糖基乳糖的产生。通过来自幽门螺杆菌的fuct2表达产生的2’-岩藻糖基乳糖(2’-fl)量相对于来自大肠杆菌o126的wbgl表达产生的2’-岩藻糖基乳糖(2’-fl)量的比较显示于以下表1中:表3.使用来自幽门螺杆菌的α-1,2-岩藻糖基转移酶fuct2以及来自大肠杆菌o126的wbgl产生的2’-岩藻糖基乳糖量的比较岩藻糖基转移酶2’-fl产量[mm]无(阴性对照)0.00fuct2(幽门螺杆菌)2.01wbgl(大肠杆菌o126)4.05从表1中可以看出,当使用根据本发明的α-1,2-岩藻糖基转移酶,即来自大肠杆菌o126的wbgl时,岩藻糖基化产物2’-岩藻糖基乳糖的量明显高于现有技术中使用来自幽门螺杆菌的α-1,2-岩藻糖基转移酶fuct2。岩藻糖基化产物的纯化如上所述产生的2’-岩藻糖基乳糖以以下几个步骤进行纯化。第一步是通过活性炭吸附进行纯化。将来自生产步骤的培养上清液应用于活性炭床。收集和分析通流(flow-through),但没有检测到残余的2’-岩藻糖基乳糖。为了除去不确定地结合的培养基化合物,如,例如,盐类和氨基酸,用蒸馏水洗涤活性炭床(在通流中没有2’-fl)。然后,用96%的乙醇洗脱2’-fl、残留的乳糖、以及岩藻糖。随后在旋转蒸发器中蒸发乙醇,并且通过10kda的横流模块(crossflowmodule)(microdynnadir,germany)过滤残余物。通过电渗析(electrodialysation)除去残留的盐类,其后通过使用横流模块(pall,germany)过滤除去内毒素。然后,通过利用在520mmx428mm的玻璃柱中堆积玻璃料的凝胶渗透色谱法材料biogelp-2(biorad,germany),将2’-fl以克级规模从乳糖和岩藻糖中分离。通过薄层色谱法监测2’-fl的纯化。合并仅包含2’-岩藻糖基乳糖的部分,并将其冷冻干燥。产物身份(identity)的确认通过1h-nmr分析利用本发明给出的岩藻糖基转移酶产生的纯化2’-岩藻糖基乳糖(参见图11)。得到的谱图与2’-fl的标准谱图(dextra,reading,uk)一致。除此之外,应用不同的hplc方法确认得到的2’-fl的身份。使用如上所述的hpaec-pad。其它方法是通过使用以水作为洗脱液的phenomenexrezexrcmca2+柱(在80℃下,以0.6ml/min洗脱30分钟;通过折光率检测器(shimadzu,germany)检测)分离(参见图10a),以及通过使用以乙腈/水(68:32)作为洗脱液的5μm250x4.6mmreprosil碳水化合物(在35℃下,以1.4ml/min洗脱20分钟;通过折光率检测器(shimadzu,germany)检测)分离(参见图10b)。序列表<110>詹尼温生物技术有限责任公司(jenneweinbiotechnoloiegmbh)<120>新型岩藻糖基转移酶及其应用<130>2827p104wo<160>8<170>siposequencelisting1.0<210>1<211>894<212>dna<213>大肠杆菌(escherichiacoli)<400>1atgagcattattcgtctgcagggtggtctgggtaatcagctgtttcagtttagctttggt60tatgccctgagcaaaattaatggtacaccgctgtatttcgacattagccattatgccgaa120aacgatgatcatggtggttatcgtctgaataatctgcagattccggaagaatatctgcag180tattataccccgaaaattaataatatttataaactgctggtgcgtggcagccgtctgtat240ccggatatttttctgtttctgggcttttgcaacgaatttcatgcctatggctacgatttt300gaatatattgcccagaaatggaaaagcaaaaaatacattggctactggcagagcgaacac360ttttttcataaacatattctggacctgaaagaattttttattccgaaaaatgtgagcgaa420caggcaaatctgctggcagcaaaaattctggaaagccagagcagcctgagcattcatatt480cgtcgtggcgattatattaaaaacaaaaccgcaaccctgacacatggtgtttgtagcctg540gaatattataaaaaagccctgaacaaaatccgcgatctggcaatgattcgtgatgtgttt600atctttagcgacgatatcttctggtgcaaagaaaatattgaaaccctgctgagcaaaaaa660tataatatttattatagcgaagatctgagccaagaagaggatctgtggctgatgagcctg720gcaaatcatcatattattgccaatagcagctttagttggtggggtgcatatctgggtagc780agcgcaagccagattgttatttatccgaccccgtggtatgatattaccccgaaaaacacc840tatatcccgattgtgaaccattggatcaacgttgataaacatagcagctgctaa894<210>2<211>297<212>prt<213>大肠杆菌(escherichiacoli)<400>2metserileileargleuglnglyglyleuglyasnglnleuphegln151015pheserpheglytyralaleuserlysileasnglythrproleutyr202530pheaspileserhistyralagluasnaspasphisglyglytyrarg354045leuasnasnleuglnileprogluglutyrleuglntyrtyrthrpro505560lysileasnasniletyrlysleuleuvalargglyserargleutyr65707580proaspilepheleupheleuglyphecysasngluphehisalatyr859095glytyrasppheglutyrilealaglnlystrplysserlyslystyr100105110ileglytyrtrpglnsergluhisphephehislyshisileleuasp115120125leulysgluphepheileprolysasnvalsergluglnalaasnleu130135140leualaalalysileleugluserglnserserleuserilehisile145150155160argargglyasptyrilelysasnlysthralathrleuthrhisgly165170175valcysserleuglutyrtyrlyslysalaleuasnlysileargasp180185190leualametileargaspvalpheilepheseraspaspilephetrp195200205cyslysgluasnilegluthrleuleuserlyslystyrasniletyr210215220tyrsergluaspleuserglnglugluaspleutrpleumetserleu225230235240alaasnhishisileilealaasnserserphesertrptrpglyala245250255tyrleuglyserseralaserglnilevaliletyrprothrprotrp260265270tyraspilethrprolysasnthrtyrileproilevalasnhistrp275280285ileasnvalasplyshissersercys290295<210>3<211>642<212>dna<213>猪葡萄球菌(staphylococcushyicus)<400>3atgcaccaccaccaccaccacaatgattcgacaacacaaacaacgacaccactggaagtc60gctcaaacgtcgcagcaagaaacacatacacatcaaacacctgttacatcattacatact120gcaacacctgaacatgttgatgactctaaagaagcaacacctttacctgaaaaagcagag180tcaccaaaaaccgaagtgacagttcaaccttcatcgcatacacaggaagtacctgcgtta240cataaaaaaacacagcaacaaccggcgtataaggataaaacggtaccagagtcaacgata300gcatcaaagtcggttgaatcaaataaagcaacagaaaatgagatgtcacctgttgaacat360catgcttcaaatgtggaaaaacgtgaagatagattggagactaatgagacaacaccgcca420tcagtggaccgtgaatttagccataaaatcatcaataatgcgcacgtaaatccaaaaacg480gatggacaaacaaacgttaatgttgatacgaaaacgatagacaccgtttcaccgaaagat540gacagaatagatacggcgcaaccgaaacaagtcgacgctcctaaagaaaatacaacggca600caaaataaatttacatcacaagcgagcgacaaaaaaccatac642<210>4<211>214<212>prt<213>猪葡萄球菌(staphylococcushyicus)<220><221>mutagen<222>(2)..(7)<223>his-标签<400>4methishishishishishisasnaspserthrthrglnthrthrthr151015proleugluvalalaglnthrserglnglngluthrhisthrhisgln202530thrprovalthrserleuhisthralathrprogluhisvalaspasp354045serlysglualathrproleuproglulysalagluserprolysthr505560gluvalthrvalglnproserserhisthrglngluvalproalaleu65707580hislyslysthrglnglnglnproalatyrlysasplysthrvalpro859095gluserthrilealaserlysservalgluserasnlysalathrglu100105110asnglumetserprovalgluhishisalaserasnvalglulysarg115120125gluaspargleugluthrasngluthrthrproproservalasparg130135140glupheserhislysileileasnasnalahisvalasnprolysthr145150155160aspglyglnthrasnvalasnvalaspthrlysthrileaspthrval165170175serprolysaspaspargileaspthralaglnprolysglnvalasp180185190alaprolysgluasnthrthralaglnasnlysphethrserglnala195200205serasplyslysprotyr210<210>5<211>50<212>dna<213>人工序列(artificialsequence)<220><221>misc_binding<223>来自猪葡萄球菌(staphylococcushyicus)的207个氨基酸his-标记多肽的正向引物<400>5catatgcaccaccaccaccaccacaatgattcgacaacacaaacaacgac50<210>6<211>30<212>dna<213>人工序列(artificialsequence)<220><221>misc_binding<223>来自猪葡萄球菌(staphylococcushyicus)的207个氨基酸his-标记多肽的反向引物<400>6ggatccgtatggttttttgtcgctcgcttg30<210>7<211>38<212>dna<213>人工序列(artificialsequence)<220><221>misc_binding<223>经由ncoi将密码子优化基因wbgl克隆至pacycduet-1中的正向引物<400>7gatcaccatgggcagcattattcgtctgcagggtggtc38<210>8<211>43<212>dna<213>人工序列(artificialsequence)<220><221>misc_binding<223>经由bamhi将密码子优化基因wbgl克隆至pacycduet-1中的反向引物<400>8gatcaggatccttagcagctgctatgtttatcaacgttgatcc43当前第1页12
- 该技术已申请专利。仅供学习研究,如用于商业用途,请联系技术所有人。
- 技术研发人员:尤利娅·帕尔科特;埃里克·许夫纳;斯特凡·延内魏因;洛塔尔·埃林;勒奥尼·恩格斯
- 技术所有人:詹尼温生物技术有限责任公司
- 我是此专利的发明人
- 该领域下的技术专家
- 如您需求助技术专家,请点此查看客服电话进行咨询。
- 1、薛老师:1.CRISPR-Cas系统 2.基因编辑 3.基因修复 4.天然产物合成 5.单分子技术开发与应用
- 2、张老师:1.探索新型氧化还原酶结构-功能关系,电催化反应机制 2.酶电催化导向的酶分子改造 3.纳米材料、生物功能多肽对酶-电极体系的影响4. 生物电化学传感和生物电合成体系的设计与应用。
- 3、豆老师:1.环境纳米材料及挥发性有机化合物(VOCs) 2.CO污染物的催化氧化 3.低温等离子体 4.吸脱附等控制技术
- 4、赵老师:1.高分子材料改性及加工技术 2.微孔及过滤材料 3.环境友好高分子材料
- 5、邬老师:1.高分子材料的共混与复合 2.涉及材料功能化及结构与性能的研究; 高分子热稳定剂的研发
- 如您是高校老师,可以点此联系我们加入专家库。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
精彩留言,会给你点赞!