一种通过干扰黄萎病菌VdRGS1基因表达显著提高棉花对黄萎病抗性的方法与流程
本发明涉及一个黄萎病菌vdrgs1基因及通过干扰vdrgs1基因表达显著提高棉花对黄萎病抗性的方法。黄萎病菌vd8和v991是两种侵染棉花的强致病力菌株,导致棉花黄萎病表型。rgs是g蛋白信号通路中g蛋白α亚基的负调控因子,利用农杆菌介导的方法将棉花黄萎病菌vd8中的vdrgs1基因敲除和互补,获得vdrgs1敲除和回补突变体。功能和表型分析表明vdrgs1调控黄萎病菌致病性、微菌核形成、孢子及菌丝的生长等,在黄萎病菌侵染寄主时发挥重要作用。通过创制表达绿色荧光蛋白(gfp)的转基因v991黄萎病菌,验证了基于病毒诱导的基因沉默技术(vigs)能够实现宿主诱导基因沉默(higs)。将表达gfp的黄萎病菌接种棉花后,通过vigs技术可在棉花中高效沉默该转基因菌中的gfp基因,表明基于vigs技术能够成功地在棉花中沉默入侵的黄萎病菌基因。利用以vigs为基础的higs技术,在棉花中沉默黄萎病菌vdrgs1,显著提高了棉花对黄萎病菌的抗性。
背景技术:
棉花(gossypiumspp.)作为全球最重要的经济作物之一,其产量和纤维品质严重受到黄萎病菌(verticilliumdahliae)的危害。棉花一旦感染黄萎病,轻则减产重则绝收,因此黄萎病被认为是棉花的“癌症”。据联合国粮农组织不完全统计,由于病害而造成的棉花减产可高达24%。我国每年因黄萎病害造成棉花产量损失达几十亿(jianetal.,2003;xuetal.,2012)。黄萎病菌是一种世界性毁灭性的植物病害,可危害660多种植物,造成寄主植物的萎蔫进而死亡,因此防治黄萎病刻不容缓(bhatandsubbarao,1999)。
黄萎病菌是一种真菌病原菌,主要存在两类,一种是大丽轮枝菌verticilliumdahliaekleb),另一种是黑白轮枝菌(verticilliumalboatrum),两种都是半知菌亚门(deuteromycotina)中淡色孢科(moniliaceae),轮枝菌属(verticillium)。目前主要危害我国棉花的黄萎病菌一般被认为是大丽轮枝菌(张绪振等,1981)。黄萎病菌开始生长的菌丝体呈无色,之后会逐渐变为橄榄褐色,膨胀状,其中菌丝芽殖变为微菌核,逆境条件下微菌核的形成会提前。黄萎病之所以难以防治,除了其强致病力外,更重要的是病菌以微菌核形式可以在土壤或秸秆中存活数十年,只要适宜的温度和湿度,就会立即打破休眠,从病菌孢子萌发成菌丝体,执行循环侵染。由于黄萎病菌的存活周期长,具有较强的适应性和抗逆性,所以黄萎病菌的防御和治理工作进展缓慢,化学和物理防治措施均未取得较理想的效果。
g蛋白信号是真菌中最重要的信号通路之一,g蛋白通过感知外界信号将其整合为内在信号转导途径,包括camp信号途径的激活(malbon,2005)。在真菌中,g蛋白调控着营养生长和致病性等多种多样的细胞功能,包括孢子的产生、菌落的形态、侵染结构的差异性和致病性。g蛋白的负调控因子(regulatorofgproteinsignaling,rgs)作为gtpase加速器主要促进gtp水解,然后负调控g蛋白,并且迅速关闭g蛋白偶联信号转导通路(siderovskietal.,2005;devriesetal.,2000)。rgs蛋白也可以作为效应受体蛋白的拮抗剂,阻止g蛋白与自身的效应蛋白互作。另外,rgs蛋白可以通过增强在gtp水解过程中gα亚基和βγ亚基的亲和力来改变参与互作的g蛋白βγ亚基数量(zhangetal.,2011)。在酵母真菌中有4个rgs蛋白,存在着功能的多样性。sst2是信息素反应相关的负调控因子,sst2可以特异绑定g蛋白α亚基gpa1,加速gtp酶的活性(chanandotte1982;apanovitchetal.1998;yietal.2003;dixitetal.,2014)。rgs2是gpa2的抑制因子,影响细胞的耐热性,同时,加速gpa2对gtp的绑定和水解(verseleetal.,1999)。rax1参与细胞极性的建立和维持(fujitaetal.,2004)。mdm1在高温生长条件下,影响细胞核和线粒体的遗传(mcconnelletal.,1990;fisketal.,1997)。在稻瘟病菌中,morgs1和morgs4正向调控表面疏水力、孢子的产生、病菌的致病性和成熟。不同于morgs1,morgs4同时参与调控漆酶和过氧化物酶的活性(zhangetal.,2011)。综上所述,rgs作为高度多样化和多功能的家族,参与g蛋白信号调控,在真菌病原菌的致病性及生长发育中起着至关重要的作用。
宿主诱导基因沉默(higs)是一种rna干扰(rnai)过程,宿主植物中产生一系列小rna沉默病菌相应基因的转录。目前,higs已经成为防治病害提高植物抗病性的有效途径之一,它主要通过沉默病原菌的关键基因,抑制病原菌在宿主植物中的生长或致病性。例如,在宿主植物拟南芥和大麦中利用rna干扰技术,沉默镰刀菌中的细胞色素氧化酶cyp51,显著影响病菌对寄主的侵染能力(kochetal.,2013)。利用higs技术,揭示了侵染大麦的白粉病菌中,其多个效应蛋白的致病性(pliegoetal.,2013)。在小麦中,利用higs技术沉默镰刀菌属中几丁质合成酶基因的转录,显著提高了小麦抗赤霉病和苗枯萎病的能力(chengetal.,2015)。
黄萎病菌是一种寄主范围特别广泛的土传真菌,利用higs技术沉默黄萎病菌已经在棉花,拟南芥,番茄等多种植物上有成功报道(zhangetal.,2016;songandthomma,2016)。棉花黄萎病菌疏水蛋白基因vdh1,主要功能是促进正常的菌丝生长,微菌核及黑色素的形成。利用higs技术在棉花中沉默vdh1后,显著提高棉花对黄萎病菌的抗性(zhangetal.,2016)。在番茄和拟南芥上,验证和比较了vigs瞬时沉默和rna干扰稳定的转基因技术对黄萎病菌不同基因沉默的表型,结果发现两者都可以实现宿主诱导黄萎病菌基因的沉默,rna干扰转基因技术相对较稳定(songandthomma,2016)。因此,通过发掘棉花黄萎病菌关键致病基因,不仅可揭示黄萎病菌的致病机理,而且为通过转基因技术干扰病菌基因表达,提高棉花的抗病性奠定基础。
到目前为止,rgs在黄萎病菌中的功能分析及表型鉴定尚未报道,更没有被应用到提高棉花对黄萎病菌抗性的育种实践中。同时,以vigs为基础的higs技术在棉花和黄萎病互作方面仍没有被验证。本发明研究发现棉花黄萎病菌vdrgs1参与控制病菌的致病性、微菌核形成、孢子产生以及菌丝生长等功能,有望基于higs技术,沉默该基因,降低黄萎病菌致病力,实现增强棉花抗病性的目的。为了验证这一结果,通过转基因技术,创制了表达绿色荧光蛋白(gfp)的黄萎病菌,利用病毒诱导的基因沉默(vigs)技术,在棉花中高效沉默该黄萎病菌的gfp基因,表明基于vigs技术可实现宿主诱导病原菌基因沉默(higs),成功地在棉花中沉默入侵的黄萎病菌基因。结合vigs为基础的higs技术,在棉花中沉默黄萎病菌vdrgs1,显著提高了棉花对黄萎病菌的抗性。综上所述,利用higs技术在棉花中创制干扰vdrgs1表达的转基因材料,可显著提高棉花对黄萎病菌的抗性。
技术实现要素:
本发明的目的是提供一种黄萎病菌vdrgs1基因及该基因在黄萎病菌vd8的基因组序列,全长cdnaorf序列和编码的蛋白序列。
本发明的另一目的是提供vdrgs1基因的功能特征。利用农杆菌介导的方法将棉花黄萎病菌vd8中的vdrgs1基因敲除和互补,获得vdrgs1敲除和回补突变体。功能和表型分析表明vdrgs1调控黄萎病菌致病性、微菌核形成、孢子及菌丝的生长等,在黄萎病菌侵染寄主时发挥重要作用。
本发明的又一目的是提供一种以vigs为基础的higs技术实现宿主诱导基因沉默(higs),为棉花和黄萎病互作的功能研究提供一种方法。通过转基因技术,创制了表达绿色荧光蛋白(gfp)的黄萎病菌,利用病毒诱导的基因沉默(vigs)技术可在棉花中高效沉默该黄萎病菌的gfp基因,表明基于vigs技术实现了宿主诱导的病原菌基因沉默(higs),可成功地在棉花中沉默入侵的黄萎病菌基因。
本发明的又一目的是提供黄萎病菌vdrgs1基因在棉花抗病育种中的应用。以vdrgs1基因为靶基因,利用以vigs为基础的higs技术在棉花中沉默入侵黄萎病菌的vdrgs1基因,或利用higs技术在棉花中创制干扰vdrgs1表达的转基因材料,显著提高棉花对黄萎病菌的抗性。
本发明的目的通过以下技术方案实现
一种黄萎病菌vdrgs1基因,该基因在大丽轮枝菌vd8基因组中,其核苷酸序列如seqidno.1所示;该基因具有如seqidno.2所示的orf序列;该基因编码的蛋白具有如seqidno.3所示的氨基酸序列。
上述的vdrgs1基因在调控棉花黄萎病病菌致病性、微菌核形成、孢子及菌丝生长中的应用。
上述的应用中为明确vdrgs1基因的功能特征,利用农杆菌介导的方法将棉花黄萎病菌vd8中的vdrgs1基因敲除和互补,获得vdrgs1敲除和回补突变体。功能和表型分析表明vdrgs1基因调控黄萎病菌致病性、微菌核形成、孢子及菌丝的生长等,在黄萎病菌侵染寄主时发挥重要作用。
上述的vdrgs1基因在提高棉花抗病性及培育棉花新种质或新品种中的应用。
上述的应用是以vigs为基础的higs技术实现宿主诱导基因沉默(higs),为棉花和黄萎病互作的功能研究提供一种方法。通过转基因技术,创制了表达绿色荧光蛋白(gfp)的黄萎病菌,利用病毒诱导的基因沉默(vigs)技术可在棉花中高效沉默该黄萎病菌的gfp基因,表明基于vigs技术实现了宿主诱导的病原菌基因沉默(higs),可成功地在棉花中沉默入侵的黄萎病菌基因。
上述的应用,具体提供黄萎病菌vdrgs1基因在棉花抗病育种中的应用。以vdrgs1基因为靶基因,利用以vigs为基础的higs技术在棉花中沉默入侵黄萎病菌的vdrgs1基因,或利用higs技术在棉花中创制干扰vdrgs1表达的转基因材料,显著提高棉花对黄萎病菌的抗性。
含有上述vdrgs1基因或其部分基因片段的重组载体、表达盒、转基因细胞系或重组菌。
上述的重组载体、表达盒、转基因细胞系或重组菌在提高棉花抗黄萎病性和培育棉花抗病新种质/新品种中的应用。
一种提高棉花抗黄萎病的方法,以病毒诱导基因沉默为基础的宿主诱导基因沉默技术,实现宿主诱导的病原菌基因沉默,在棉花中降低上述的vdrgs1基因的表达剂量。
本发明的优点表现在:
(1)首次在棉花黄萎病菌中对vdrgs1基因功能分析
到目前为止,黄萎病菌中没有关于g蛋白信号相关基因的报道,我们在黄萎病菌中对rgs类蛋白进行分析,利用农杆菌介导法获得基因敲除和互补突变体,最终发现vdrgs1显著影响病菌的致病性、微菌核的形成、孢子的生长等,是应用于提高棉花黄萎病抗性的优选靶基因。
(2)以vigs为基础的higs技术在棉花上的验证
我们利用上述农杆菌介导的方法,在黄萎病中导入gfp基因,获得荧光信号强而不影响致病性的菌株,验证以vigs为基础的higs技术可有效用于在棉花上沉默入侵的黄萎病菌基因。以gfp为靶基因构建vigs载体,注射棉花幼苗,以注射空载和不注射的陆地棉品种军棉1号为对照。两周后,接种含gfp的黄萎病菌株。同时,在接病菌2周后,选择对照和注射含沉默gfp载体的多个不同植株,切取幼苗茎秆在土豆培养基上培养3天,观察孢子荧光信号。结果表明只有注射含gfp的vigs载体的幼苗中,具有荧光信号的黄萎病菌孢子数显著减少。表明利用该技术可以实现寄主棉花对入侵黄萎病菌基因的沉默。
(3)基于higs技术沉默黄萎病菌中关键基因在棉花抗病中的应用
在黄萎病菌中发掘致病相关关键基因,结合黄萎病菌的防治难点,选择理想靶基因。研究表明vdrgs1参与黄萎病菌的致病性、微菌核的形成、孢子的生长等,利用以vigs为基础的higs技术在棉花中沉默入侵黄萎病菌的vdrgs1,或利用higs技术在棉花中创制干扰vdrgs1表达的转基因材料,显著提高棉花对黄萎病菌的抗性。
附图说明
图1黄萎病菌中rgs的鉴定与表达特征分析
a:大丽轮枝菌中8个rgs基因的进化分析;b:8个rgs蛋白结构域的验证,都包含rgs功能域。与其他rgs蛋白不同,仅vdrgs1含有2个dep结构域;c:8个rgs基因在棉花与黄萎病菌互作中表达特征分析。棉花幼苗处理黄萎病菌不同时间rgs表达水平变化,vdrgs1的表达和诱导表达水平表现最高,可能参与黄萎病菌致病性。
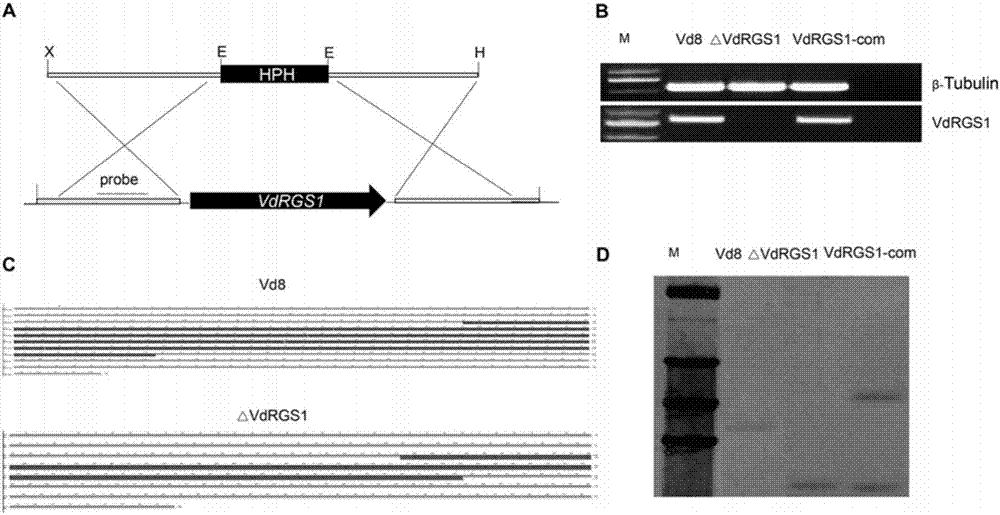
图2vdrgs1缺失和互补突变体获得与验证
a:vdrgs1缺失突变体同源置换的模式图;b:rt-pcr验证vdrgs1缺失和互补突变体(△vdrgs1和vdrgs1-com);c:双侧pcr验证△vdrgs1,序列分析表明目的基因vdrgs1被替换;d:以目的基因vdrgs1上游片段为探针,southernblot验证△vdrgs1和vdrgs1-com,发现vdrgs1被替换。与野生型相比,△vdrgs1中,目标片段分子量明显减小,且只有一个拷贝。在vdrgs1-com中,vdrgs1成功导入△vdrgs1突变体。
图3vdrgs1影响黄萎病菌孢子形态和菌丝生长
a:△vdrgs1菌株其孢子萌发提前,vdrgs1-com菌株的孢子则正常生长;b:vd8、△vdrgs1和vdrgs1-com单个孢子在完全培养基(cm)上生长24和36小时的表型,△vdrgs1呈哑铃状,vdrgs1-com表型正常。
图4vdrgs1影响菌体的形态及孢子的产生
a:△vdrgs1在完全培养基(cm)和马铃薯培养基(pda)上的形态,表现为微菌核缺失;b:vd8、△vdrgs1和vdrgs1-com的1×103个孢子在50毫升察氏培养基(cpk)培养7天,发现△vdrgs1的产孢量显著减少。
图5vdrgs1影响黄萎病菌微菌核的形成
a:vd8、△vdrgs1和vdrgs1-com的孢子在cm培养基上生长5天,发现△vdrgs1微菌核消失;b:vd8、△vdrgs1和vdrgs1-com以1×105个孢子/毫升喷施在0.22μm的硝酸膜上,膜下是基本培养基(bm)。5天后发现△vdrgs1微菌核消失;c:显微镜下观察硝酸膜上vd8、△vdrgs1和vdrgs1-com菌株表型,发现△vdrgs1黑色素与微菌核几乎消失(倍数:4×和10×);d:黑色素和微菌核合成相关的关键基因聚酮化合物合成酶(vdag_00190)、羟基萘还原酶(vdag_03665)、阿魏酸脱水酶(vdag_03393)和多色素还原酶(vdag_00183)在△vdrgs1中均显著下调。
图6vdrgs1影响黄萎病菌的致病力
a:水处理(ck)、vd8、△vdrgs1和vdrgs1-com接种感病的陆地棉品种军棉1号,接种11、15和20天后调查表型,发现△vdrgs1致病力丧失;b:接种15天后,劈杆发现,植株中△vdrgs1致病力和微菌核丧失;c:vd8、△vdrgs1和vdrgs1-com分别接种陆地棉军棉1号后,在不同时间的病叶率统计,△vdrgs1致病力显著降低。
图7vigs为基础的higs技术在棉花中的验证
黄萎病菌v991和vd8均为导致棉花黄萎病的强致病力菌株,通过转基因技术在黄萎病菌v991中整合gfp(v991-gfp),筛选获得具有绿色荧光信号而致病性不受影响的转gfp转基因菌株。棉花幼苗注射trv:gfp载体2周后,接种v991-gfp菌株。15天后,取ck、接种trv:00与trv:gfp的多个不同植株重复样品,劈杆培养3天后,利用激光共聚焦观察孢子的荧光信号,发现注射trv:gfp的棉花茎秆中约50%孢子荧光信号消失,其他对照孢子荧光信号无明显变化。
图8黄萎病菌不同生理型v991和vd8中vdrgs1的序列比较
分别在黄萎病菌不同生理型v991和vd8中克隆vdrgs1,序列比较发现该基因在上述两个菌株中基因序列无差异。
图9利用病毒诱导的基因沉默(vigs)技术在棉花中沉默入侵黄萎病菌vd8中的vdrgs1,显著提高棉花的抗病性
a:vigs沉默的棉花茎秆中,vd8中vdrgs1转录水平检测。接种15天后,取不同茎秆培养3天后,qrt-pcr检测注射不同载体的茎秆中vdrgs1的转录水平,以注射trv:00的茎秆中vd8为对照。数据表示三次独立实验的三个样本的标准偏差,“*”p<0.05;“**”p<0.01。b:选择vdrgs1基因的不同区段分别构建4个不同沉默载体。水处理、接种△vdrgs1与野生型vd8在陆地棉军棉1号及注射trv:00,trv:vdrgs1-1,trv:vdrgs1-2,trv:vdrgs1-3与trv:vdrgs1-4等不同载体的幼苗上,接种15和20天后植株表型。c:注射不同沉默载体和对照的军棉1号的平均病叶率,不同沉默载体每个处理至少30棵幼苗,三次生物学重复,用于统计学分析。以trv:00为对照进行数据分析。*p<0.05,**p<0.01。d:不同处理15天后的棉花幼苗微管颜色的变化。用体式显微镜mvx10观察,ck、trv:00、trv:vdrgs1-1与trv:vdrgs1-4的茎秆在接种vd8菌株15天后,微管微菌核和黑色素积累。水处理和△vdrgs1处理的茎秆,注射trv:vdrgs1-2与trv:vdrgs1-3的幼苗接种vd8菌株15天后,几乎不存在微管微菌核和黑色素的积累。与对照相比,注射trv:vdrgs1-2与trv:vdrgs1-3的幼苗其vdrgs1显著被沉默,表现出较好的抗病性,说明微管微菌核和黑色素的积累和vdrgs1沉默效率显著相关。
图10利用病毒诱导的基因沉默(vigs)技术在棉花中沉默入侵黄萎病菌v991中的vdrgs1,显著提高棉花的抗病性
a:vigs沉默的棉花茎秆中,v991中vdrgs1和gfp转录水平检测,接种15天后,取不同茎秆培养3天,qrt-pcr检测注射不同载体的茎秆中vdrgs1和gfp转录水平,以注射trv:00的茎秆中v991为对照。数据表示三次独立实验的三个样本的标准偏差,“*”p<0.05;“**”p<0.01;b:水处理和v991-gfp处理的不同vigs幼苗15与20天的植株表型,注射trv:gfp,trv:00,trv:vdrgs1-2,trv:vdrgs1-3与ck表型差异不明显,注射trv:vdrgs1-2和trv:vdrgs1-3的植株vdrgs1转录水平未受显著影响。而注射trv:vdrgs1-1与trv:vdrgs1-4vdrgs1转录水平显著降低,同时影响了v991的致病性,明显增加棉花的抗病性。c:不同处理的棉株平均病叶率统计,发现注射trv:vdrgs1-1与trv:vdrgs1-4显著提高棉花的抗病能力。再次证明棉花抗病性与vdrgs1沉默效率显著相关。
具体实施方式
实施例1
(一)黄萎病菌vdrgs1的发现与功能分析
利用已报道的大丽轮枝菌基因组信息(http://genome.jgi.doe.gov/verda1/verda1.home.html),对rgs进行系统分析,验证后发现存在8个rgs基因。基于上述序列信息,设计引物进一步在棉花黄萎病菌vd8中进行同源克隆。用棉花幼苗根处理vd8菌液0、2、4、6、12和24小时,分析8个rgs基因在黄萎病菌接触棉花幼苗根组织不同时间的表达特征。发现只有vdrgs1基因表达量最高且诱导表达最明显,在接触棉花根部12和24小时后,表达水平上升20-30倍,暗示该基因在黄萎病菌致病方面起着重要的作用(图1)。研究所用引物列于表1。黄萎病菌vdrgs1基因在大丽轮枝菌vd8基因组中,测序结果显示,其核苷酸序列如seqidno.1所示;该基因具有如seqidno.2所示的orf序列;该基因编码的蛋白具有如seqidno.3所示的氨基酸序列。
为了明确vdrgs1基因在黄萎病菌中的功能特征,利用农杆菌介导的方法获得vdrgs1基因缺失和回补的vd8黄萎病菌突变体(△vdrgs1和vdrgs1-com),并在转录水平、dna水平以及通过southernblot验证该突变体真实性(图2)。我们观察突变体的孢子发现,△vdrgs1的孢子明显提前萌发,而vdrgs1-com与野生型vd8没有明显差异。另外,野生型vd8、△vdrgs1和vdrgs1-com在完全培养基(cm)上进行单孢培养,在生长24和36小时后分别观察不同菌株表型的差异,发现△vdrgs1的菌丝生长呈哑铃状,vdrgs1-com与野生型没有差异。表明vdrgs1的缺失显著影响孢子的萌发以及菌丝的生长(图3)。
将野生型vd8、△vdrgs1和vdrgs1-com的单个孢子分别在cm和马铃薯培养基(pda)生长,观察菌体形态,发现△vdrgs1微菌核消失。用野生型vd8、△vdrgs1和vdrgs1-com的1000个孢子在50毫升察氏培养基(cpk)中培养,每个菌株5次重复。结果发现△vdrgs1的产孢量显著减少,大约是野生型产孢量的41%,而vdrgs1-com与野生型vd8的产孢量没有显著差异。结果表明,vdrgs1参与黄萎病菌孢子的产生(图4)。
为了验证vdrgs1是否参与黄萎病菌微菌核形成,我们对野生型vd8、△vdrgs1和vdrgs1-com在cm培养基上进行单孢培养5天,发现vdrgs1基因的缺失突变体微菌核消失。另外,我们分别用野生型vd8、△vdrgs1和vdrgs1-com的1×105个/毫升孢子浓度,喷施在覆盖0.22μm的硝酸膜的bm培养基上,培养5天。发现只有野生型vd8和vdrgs1-com的微菌核与黑色素积累,而△vdrgs1没有发现上述表型。分别在4和10倍的显微镜下进一步验证了上述结果,只有野生型vd8和vdrgs1-com的微菌核与黑色素分布密集,而△vdrgs1几乎没有微菌核的分布。另外,目标基因表达分析发现,合成黑色素与微菌核的关键基因vdag_00190、vdag_03665、vdag_03393和vdag_00183,在△vdrgs1中其表达水平极显著下调(图5)。所涉及的扩增引物列于表2。
为了验证vdrgs1是否影响黄萎病菌vd8的致病性,我们将水对照、野生型vd8、△vdrgs1和vdrgs1-com等分别接种感病材料陆地棉军棉1号,每个处理30棵。观察发病情况,并分别统计不同接种时间(11、15、20、25和30天)的平均病叶率,结果表明△vdrgs1和h2o处理类似,致病性几乎丧失,而野生型vd8和vdrgs1-com致病力没有明显差异(图6)。综上所述,vdrgs1显著影响黄萎病菌致病性、孢子的产生和微菌核的形成。
表1:黄萎病菌vdrgs基因克隆和定量分析所用引物
表2本研究涉及的其他引物信息
(二)vigs为基础的higs技术在棉花与黄萎病菌互作中的验证及应用
黄萎病菌不同生理型vd8和v991均为棉花强致病力菌株,导致棉花落叶型黄萎病表型。分别以vd8和v991为受体,通过农杆菌介导法转化外源gfp基因,对转化体进行鉴定,最终获得荧光信号强且不影响致病力的v991转基因菌株(v991-gfp)。我们把gfp的一段编码区序列构建到trv载体上,注射军棉1号幼苗的子叶,两周后接种v991-gfp。接种15天后,我们取ck、trv:00和trv:gfp处理的不同植株,切取茎杆培养在cm培养基,3天后检测菌株中gfp的转录水平,同时在激光共聚焦显微镜下检测孢子的荧光信号。结果表明,注射trv:gfp幼苗的茎秆,其v991-gfp菌株中gfp的转录水平显著降低,同时所含的孢子荧光信号也明显消失。进一步统计发现,注射trv:gfp幼苗的茎秆中仅50%的孢子检测到绿色荧光,而在ck和trv:00茎秆中近100%的孢子均具有明显的绿色荧光信号(图7)。另外,经过对黄萎病菌不同生理型vd8和v991中vdrgs1的克隆和序列比对,发现两个生理型中的vdrgs1序列完全一致(图8)。可利用vigs技术分别对不同生理型vd8和v991中的vdrgs1基因实现沉默。
进一步开展了利用vigs为基础higs技术沉默棉花中入侵病菌的vdrgs1。分别选择vdrgs1的4个不同基因片段,构建了trv:vdrgs1-1,trv:vdrgs1-2,trv:vdrgs1-3和trv:vdrgs1-4等四个不同的vdrgs1沉默载体(vigs载体构建所用引物见表3),连同trv:00和trv:ghcla1一起注射棉花。ghcla1基因(genbank登录号:kj123647)编码1-deoxyxylulose5-phosphatesynthase,从陆地棉遗传标准系tm-1中克隆,参与叶绿体发育过程,该基因突变有明显的白化表型,是易于识别的标记性状(王心宇等,作物学报,2014,40(8):1356-1363)。两周后,注射trv:ghcla1的幼苗出现明显的白化表型,证明vigs技术的可行性。将所有vigs幼苗和对照植株接种野生型vd8菌株,同时对照幼苗接种△vdrgs1和h2o。15天后,取不同处理样品的植株,切取茎段在cm培养基上培养,分析vdrgs1转录水平。相对于trv:00内vd8中的vdrgs1表达水平,注射trv:vdrgs1-2和trv:vdrgs1-3载体的幼苗茎秆中,vd8菌株中vdrgs1表达水平显著下调,表明vdrgs1被沉默。调查接种15和20天的植株表型以及不同时间不同处理的植株发病率,发现h2o处理和接种△vdrgs1幼苗表型类似,再次证明△vdrgs1致病力丧失。注射trv:vdrgs1-2和trv:vdrgs1-3载体的幼苗发病率明显减少。另外,不同处理的幼苗接种15天后,发现trv:vdrgs1-2和trv:vdrgs1-3处理的植株与h2o和△vdrgs1接种的植株表型无显著差异,微菌核和黑色素几乎不存在。而vdrgs1没有被沉默的幼苗茎秆,例如接种vd8的ck、trv:00、trv:vdrgs1-1和trv:vdrgs1-4等植株,其微管中黑色素和微菌核积累明显增加。结果表明,沉默黄萎病菌中vdrgs1可以有效增加棉花对黄萎病菌的抗病性(图9)。
我们进一步使用v991-gfp黄萎病菌接种棉花,在棉花中沉默v991中的vdrgs1,观察是否增加棉花的抗病性。将trv:vdrgs1-1、trv:vdrgs1-2、trv:vdrgs1-3和trv:vdrgs1-4,连同trv:00和trv:gfp分别注射棉花幼苗,两周后取样,通过棉花切杆培养检测v991中vdrgs1或gfp的转录水平。发现只有trv:gfp茎秆内的v991中,其gfp被沉默,大部分孢子绿色荧光信号消失(图7)。我们检测到trv:vdrgs1-1和trv:vdrgs1-4幼苗茎秆中v991的vdrgs1被不同程度的沉默。接种黄萎病菌v991后,随着vdrgs1的沉默,相对应的vigs幼苗抗病性显著增加。通过不同时间,不同处理的发病率调查,进一步证明,trv:vdrgs1-1和trv:vdrgs1-4幼苗的发病率明显减少。因此,棉花中沉默v991的vdrgs1能有效提高棉花的抗病能力(图10)。
表3vigs载体构建所用引物
sequencelisting
<110>南京农业大学
<120>一种通过干扰黄萎病菌vdrgs1基因表达显著提高棉花对黄萎病抗性的方法
<130>2017
<160>3
<170>patentinversion3.3
<210>1
<211>2384
<212>dna
<213>大丽轮枝菌vd8(verticilliumdahliaeklebvd8)
<221>vdrgs1基因的核苷酸序列
<400>1
atggccgccatctcctattccaaccgctcctcaatccgagatcaactaccgcagtcgaca60
tcgacctcacgatcttccaccagagttgccagtcgccgctcttcactcgcctcgccttcg120
gctccctcatcacccgccgttgaccctttctcccacgagggccaagccgcctccacctcg180
acgggctaccgccagatcggccacttcagctctccttccacgagcaagatccagcgatcc240
ggaggcttgctcggccttgcagctgctgcaatcgacaagacgcagtctgcgctgtccacc300
ctgcagtcacagtcacagtcacaacccgactactccatacgtcagcgttcctccaattcc360
gctttgggcagagtcgcctccagtgccgattccgctttcagccccaccgacttggacaag420
aacgcacacgcgcgatacagaaggtccaacaaccattcccccgcctcttcttcgacaggc480
actttcgccgccgagggcaaacacccgagtcaggtgtcgctggccaacgaagtgccctcc540
tctccctattccgacactgaccccaaccgccctcttccaatccgcatcccagccgtcgag600
aacaagatgcaccaaacttcttcgcggttgctccgcatgacggacgacgacaagccattt660
acaaaggtaggcatcgccatctgttgcccttgttcaattgatgtgttgtaccattgtgcg720
cgggaacgcatgtctcgaagggcaatcgatctcgcctcgtgttgagcattccgccgcact780
caaccccgccatccccatgcccccctcttccctgccctcttccacccaggcctctcttgt840
caaaaagctcacacaccacacaggatttcaaagatctctttgcaaccctcgtcgtcagcc900
tattgcccctatctgcgcatcgcgtacgcctcactcgggttgagaacactttcctctcag960
aagatgctatcaacaatctcggttccctgaaattctcccagtcgaaccgcatgccggacc1020
cgaaagacccctcccgcatcgtaacaacgacaacgacgacaaccttctccatggctaaag1080
acatggctcgttctatctgccaaaggtttctcgaggcacggttcattgaatccgccgacg1140
gaaaataccagcaggtctacaacatgaagggctctgtctggcaacttacgccaaaaggcg1200
tcactattcttgatcgattctgctctcgtaacggcattcagcagaagcaaacatccgacc1260
ttatcggatctaccctttcgcagctcgttatcctggagcgtgatggccaaaccgacaagc1320
tcattaccgaccgtggcactgtcgaggttgtcttccgtcgattcgtcggcacacacggtc1380
ccaacatcaagtccagcgtcacatcggcagattccgattcgctcagtgactaccgggatg1440
gcctcacgggtgtcaagatggctagcgagcgtaagatcaatggcaagacgtaccgcgaca1500
ccttcacaggcaagggtgcgacggactggctcatggactgctgtaccaccgtggacagga1560
gggaggctgtggacattgcaacgctctttgtgcagtttgacctgatggaggcaatcgtgc1620
aagaccgagcacacatgtcgcagtatgccggccacaacgtcttccaacccaccaagaatg1680
ctatttaccagctcaccgccaagggcaaggacctgatcaacaatgtgggctctagggggc1740
gagcttctgaaagcgaaggtacaaacccaagccgcaatgcgatcgcgcgcgactcaaaca1800
cgcagaggctggacaagattcttaacgatgccgcactccgcttgctgttccgcgagaacc1860
tcagggaaactcattgtgaagaaaacctgtccttctacctcgacgtcgacgacttcgtca1920
agagctgcaaggtcgccatccgttccgctcagaagaacccgaactctacttctatggacg1980
ggattaaagaaatcatggcacaagcttatggcatctacaacgccttcctagcccccggat2040
ccccttgcgagttgaacattgaccatcagctgcgcaacaacctggcaactcgcatgacga2100
aggctgttggccaagacgtcgccatgatagacacgctgcaagaggtgacggctctgtttg2160
aagacgcccagaacgccgtcttcaagcttatggctagtgtgagtctcaccgcctctttaa2220
tcgcaacggagccgtgtactaacaatataacccaggattctgtgccaaagtttcttcgca2280
gtccaaagtacgagcacaccttgaagcactacgactttgactccctcaccaacgttcgcc2340
ctggcggcttggagcgaagccagagtcggtcgaaccgaccatga2384
<210>2
<211>2130
<212>dna
<213>人工序列(artificialsequence)
<221>vdrgs1基因的cdnaorf序列
<400>2
atggccgccatctcctattccaaccgctcctcaatccgagatcaactaccgcagtcgaca60
tcgacctcacgatcttccaccagagttgccagtcgccgctcttcactcgcctcgccttcg120
gctccctcatcacccgccgttgaccctttctcccacgagggccaagccgcctccacctcg180
acgggctaccgccagatcggccacttcagctctccttccacgagcaagatccagcgatcc240
ggaggcttgctcggccttgcagctgctgcaatcgacaagacgcagtctgcgctgtccacc300
ctgcagtcacagtcacagtcacaacccgactactccatacgtcagcgttcctccaattcc360
gctttgggcagagtcgcctccagtgccgattccgctttcagccccaccgacttggacaag420
aacgcacacgcgcgatacagaaggtccaacaaccattcccccgcctcttcttcgacaggc480
actttcgccgccgagggcaaacacccgagtcaggtgtcgctggccaacgaagtgccctcc540
tctccctattccgacactgaccccaaccgccctcttccaatccgcatcccagccgtcgag600
aacaagatgcaccaaacttcttcgcggttgctccgcatgacggacgacgacaagccattt660
acaaaggatttcaaagatctctttgcaaccctcgtcgtcagcctattgcccctatctgcg720
catcgcgtacgcctcactcgggttgagaacactttcctctcagaagatgctatcaacaat780
ctcggttccctgaaattctcccagtcgaaccgcatgccggacccgaaagacccctcccgc840
atcgtaacaacgacaacgacgacaaccttctccatggctaaagacatggctcgttctatc900
tgccaaaggtttctcgaggcacggttcattgaatccgccgacggaaaataccagcaggtc960
tacaacatgaagggctctgtctggcaacttacgccaaaaggcgtcactattcttgatcga1020
ttctgctctcgtaacggcattcagcagaagcaaacatccgaccttatcggatctaccctt1080
tcgcagctcgttatcctggagcgtgatggccaaaccgacaagctcattaccgaccgtggc1140
actgtcgaggttgtcttccgtcgattcgtcggcacacacggtcccaacatcaagtccagc1200
gtcacatcggcagattccgattcgctcagtgactaccgggatggcctcacgggtgtcaag1260
atggctagcgagcgtaagatcaatggcaagacgtaccgcgacaccttcacaggcaagggt1320
gcgacggactggctcatggactgctgtaccaccgtggacaggagggaggctgtggacatt1380
gcaacgctctttgtgcagtttgacctgatggaggcaatcgtgcaagaccgagcacacatg1440
tcgcagtatgccggccacaacgtcttccaacccaccaagaatgctatttaccagctcacc1500
gccaagggcaaggacctgatcaacaatgtgggctctagggggcgagcttctgaaagcgaa1560
ggtacaaacccaagccgcaatgcgatcgcgcgcgactcaaacacgcagaggctggacaag1620
attcttaacgatgccgcactccgcttgctgttccgcgagaacctcagggaaactcattgt1680
gaagaaaacctgtccttctacctcgacgtcgacgacttcgtcaagagctgcaaggtcgcc1740
atccgttccgctcagaagaacccgaactctacttctatggacgggattaaagaaatcatg1800
gcacaagcttatggcatctacaacgccttcctagcccccggatccccttgcgagttgaac1860
attgaccatcagctgcgcaacaacctggcaactcgcatgacgaaggctgttggccaagac1920
gtcgccatgatagacacgctgcaagaggtgacggctctgtttgaagacgcccagaacgcc1980
gtcttcaagcttatggctagtgattctgtgccaaagtttcttcgcagtccaaagtacgag2040
cacaccttgaagcactacgactttgactccctcaccaacgttcgccctggcggcttggag2100
cgaagccagagtcggtcgaaccgaccatga2130
<210>3
<211>709
<212>prt
<213>人工序列(artificialsequence)
<221>
<400>3
metalaalailesertyrserasnargserserileargaspglnleu
151015
proglnserthrserthrserargserserthrargvalalaserarg
202530
argserserleualaserproseralaproserserproalavalasp
354045
propheserhisgluglyglnalaalaserthrserthrglytyrarg
505560
glnileglyhispheserserproserthrserlysileglnargser
65707580
glyglyleuleuglyleualaalaalaalaileasplysthrglnser
859095
alaleuserthrleuglnserglnserglnserglnproasptyrser
100105110
ileargglnargserserasnseralaleuglyargvalalaserser
115120125
alaaspseralapheserprothraspleuasplysasnalahisala
130135140
argtyrargargserasnasnhisserproalaserserserthrgly
145150155160
thrphealaalagluglylyshisproserglnvalserleualaasn
165170175
gluvalproserserprotyrseraspthraspproasnargproleu
180185190
proileargileproalavalgluasnlysmethisglnthrserser
195200205
argleuleuargmetthraspaspasplysprophethrlysaspphe
210215220
lysaspleuphealathrleuvalvalserleuleuproleuserala
225230235240
hisargvalargleuthrargvalgluasnthrpheleusergluasp
245250255
alaileasnasnleuglyserleulyspheserglnserasnargmet
260265270
proaspprolysaspproserargilevalthrthrthrthrthrthr
275280285
thrphesermetalalysaspmetalaargserilecysglnargphe
290295300
leuglualaargpheilegluseralaaspglylystyrglnglnval
305310315320
tyrasnmetlysglyservaltrpglnleuthrprolysglyvalthr
325330335
ileleuaspargphecysserargasnglyileglnglnlysglnthr
340345350
seraspleuileglyserthrleuserglnleuvalileleugluarg
355360365
aspglyglnthrasplysleuilethraspargglythrvalgluval
370375380
valpheargargphevalglythrhisglyproasnilelysserser
385390395400
valthrseralaaspseraspserleuserasptyrargaspglyleu
405410415
thrglyvallysmetalasergluarglysileasnglylysthrtyr
420425430
argaspthrphethrglylysglyalathrasptrpleumetaspcys
435440445
cysthrthrvalaspargargglualavalaspilealathrleuphe
450455460
valglnpheaspleumetglualailevalglnaspargalahismet
465470475480
serglntyralaglyhisasnvalpheglnprothrlysasnalaile
485490495
tyrglnleuthralalysglylysaspleuileasnasnvalglyser
500505510
argglyargalaserglusergluglythrasnproserargasnala
515520525
ilealaargaspserasnthrglnargleuasplysileleuasnasp
530535540
alaalaleuargleuleuphearggluasnleuarggluthrhiscys
545550555560
glugluasnleuserphetyrleuaspvalaspaspphevallysser
565570575
cyslysvalalaileargseralaglnlysasnproasnserthrser
580585590
metaspglyilelysgluilemetalaglnalatyrglyiletyrasn
595600605
alapheleualaproglyserprocysgluleuasnileasphisgln
610615620
leuargasnasnleualathrargmetthrlysalavalglyglnasp
625630635640
valalametileaspthrleuglngluvalthralaleuphegluasp
645650655
alaglnasnalavalphelysleumetalaseraspservalprolys
660665670
pheleuargserprolystyrgluhisthrleulyshistyraspphe
675680685
aspserleuthrasnvalargproglyglyleugluargserglnser
690695700
argserasnargpro
705709
- 还没有人留言评论。精彩留言会获得点赞!