特异性靶向CD19抗原且高水平稳定表达PD-1抗体的CAR-T细胞及用途的制作方法
本发明涉及特异性靶向cd19抗原且高水平稳定表达抗pd-1抗体的cart细胞及其抗肿瘤效果。
背景技术:
嵌合抗原受体t细胞(chimericantigenreceptortcell,car-t)治疗技术无疑是肿瘤免疫细胞治疗领域中一颗冉冉升起的巨星。car-t技术是通过基因工程技术将识别某抗原分子的抗体可变区基因序列与t淋巴细胞免疫受体的胞内区序列拼接后,通过逆转录病毒或慢病毒载体、转座子或转座酶系统或直接mrna转导到淋巴细胞内,并表达融合蛋白于细胞表面,使t淋巴细胞能通过非mhc限制性的方式识别特定抗原,增强其识别和杀伤肿瘤的能力。
car的结构自1989年以色列eshhar研究小组首次提出后,经过近30年的发展,已经被证实经过car结构修饰的t细胞在肿瘤免疫治疗中具有较好的疗效。第一代car受体包含了胞外特异识别肿瘤抗原的片段(single-chainvariablefragment,scfv),胞内激活信号由cd3ζ信号链来传递。但是第一代car受体缺乏t细胞的共刺激信号,导致t细胞只能发挥瞬间效应,在体内存在时间短、细胞因子分泌少。第二代car受体增加了共刺激信号分子的胞内结构域,包括如cd28,cd134/ox40,cd137/4-1bb,lymphocyte-specificproteintyrosinekinase(lck),induciblet-cellco-stimulator(icos)以及dnax-activationprotein10(dap10)等结构域,增强了t细胞的增殖能力及细胞因子的分泌功能,il-2、ifn-γ以及gm-csf增加,从而突破肿瘤微环境的免疫抑制、延长aicd(activationinducedcelldeath,aicd)。第三代car受体则是在共刺激结构cd28和itam信号链的之间再融合一个二级共刺激分子如4-1bb,因而产生了一个三重信号的car受体,第三代car受体改造的t细胞具有更好的效应功能和体内存活时间。常用的经典car-t结构即为二代car受体,其结构具体可分为以下4部分:识别肿瘤抗原的抗体单链可变区(scfv)、铰链区、跨膜区、胞内刺激信号结构区域。car结构铰链区负责形成正确构象,形成二聚体。铰链区的长短及氨基酸序列特征,决定了car的空间构象,也决定了其与肿瘤细胞表面抗原结合的能力。
恶性淋巴瘤分为霍奇金淋巴瘤(hl)和非霍奇金淋巴瘤(nhl)两大类。霍奇金淋巴瘤占淋巴瘤的10%-15%,而非霍奇金淋巴瘤是发病患者增加最快的恶性肿瘤。据who统计,目前全球每年约有35万新发nhl,死亡人数超过20万。在霍奇金淋巴瘤和非霍奇金淋巴瘤两大类中均可见b细胞淋巴瘤。目前淋巴瘤临床治疗药物包括糖皮质激素和烷化剂等细胞毒药物及基于特定分子靶标的靶向药物(如利妥昔单抗等),其中基于靶向药物的联合化疗显著提高部分淋巴瘤患者临床缓解率和治愈率。但目前仍有相当一部分淋巴瘤患者对现有治疗方案不敏感或疗效欠佳而成为“真正”难治性患者。一些新型治疗手段(如细胞免疫治疗等)使部分复发或难治性淋巴瘤患者获得缓解并延长其生存期。目前正在开发的针对血液系统恶性肿瘤的car-t的种类很多,主要包括利用抗cd19、抗cd20、抗kappa轻链、抗cd22、抗cd23、抗cd30、抗cd70等抗体构建car修饰的t细胞进行抗肿瘤研究,其中以抗cd19、抗cd20单克隆抗体最为热门。
选择合适的肿瘤抗原作为靶标是设计安全有效car-t的关键。由于cd19仅表达于各分化阶段的正常及恶性b细胞而不表达于其他非b细胞(如造血干细胞等)表面,是治疗b系肿瘤很有潜力的靶点,也是car-t研究中的热点,因而cd19car-t被广泛用于急性b淋巴细胞白血病(b-all)、慢性b淋巴细胞白血病(b-cll)、套细胞淋巴瘤(mcl)、nhl和多发性骨髓瘤(mm)等恶性b细胞淋巴瘤的临床试验。
pd-1(programmeddeath1,重编程细胞死亡受体1)是调节性t细胞cd28家族的成员,属于免疫球蛋白超家族受体。pd-1和它的配体pd-l1/pd-l2在t细胞的共抑制和衰竭中起重要作用,他们的相互作用抑制了共刺激分子调节的t细胞的增殖和细胞因子的分泌,下调了抗凋亡分子bcl-xl的表达,削弱了肿瘤特异性t细胞的功能,导致一些肿瘤患者不能完全消除肿瘤。pd-1抗体通过与肿瘤特异性t细胞表面的pd-1分子结合,竞争了pd-1与其配体pd-l1/pd-l2的结合,从而缓解pd-1与pd-l1/pd-l2结合引起的免疫微环境抑制作用。目前商品化的pd-1抗体有nivolumab及pidilizumab。这2种单抗已经证实在黑色素瘤、结肠癌、前列腺癌、非小型细胞肺癌、肾细胞癌等实体瘤上取得了良好的临床疗效,近期的临床研究证实pd-1抗体在淋巴瘤治疗上也取得了很大的进步。但pd-1抗体在临床应用中仍然存在一些不可避免的问题。一方面由于pd-1单抗是静脉注射全身用药,大多数接受pd-1抗体阻断治疗的患者都会出现不同程度的药物毒副作用。另一方面pd-1单抗的生产涉及复杂的生产制备与纯化工艺,成本高,导致治疗费用昂贵。
综上所述,car-t细胞具有杀灭肿瘤细胞能力,且能有效进入肿瘤组织内部,但其活性容易在肿瘤微环境中被抑制;而pd-1抗体能重新激活t细胞的抗肿瘤活性,但大分子抗体对实体瘤的穿透力不足,全身用药具有较大的毒副作用,且药物成本很高。因而,如果在保持car-t细胞杀伤毒性的前提下,能由其高效表达pd-1抗体,通过car-t细胞的肿瘤趋向特性,在肿瘤局部高水平表达pd-1抗体,将同时克服car-t细胞治疗与pd-1抗体治疗各自的缺陷,发挥两者的协同作用而提高疗效,同时降低治疗成本。
技术实现要素:
基于此,本发明设计了靶向cd19抗原且能高水平稳定表达pd-1抗体的多能car-t细胞,从而达到良好的识别抗原和杀伤淋巴瘤作用。
具体而言,本发明的多能t细胞包含识别cd19抗原的嵌合抗原受体(car)及稳定表达pd-1抗体。
在一个或多个实施方案中,所述嵌合抗原受体包含识别cd19抗原的scfv、铰链区、跨膜区、胞内共刺激信号域和胞内信号域。
在一个或多个实施方案中,所述scfv的氨基酸序列如seqidno:2所示。
在一个或多个实施方案中,所述铰链区为cd8铰链区或igg4ch2ch3铰链区。
在一个或多个实施方案中,所述cd8铰链区的氨基酸序列如seqidno:3所示;所述igg4ch2ch3铰链区的氨基酸序列如seqidno:4所示。
在一个或多个实施方案中,所述跨膜区为cd28跨膜区、cd8跨膜区、cd3ζ跨膜区、cd134跨膜区、cd137跨膜区、icos跨膜区和dap10跨膜区中的一种;优选为cd8跨膜区,优选其氨基酸序列如seqidno:5所示。
在一个或多个实施方案中,所述胞内共刺激信号域包括共刺激信号分子的胞内结构域,包括cd28、cd134/ox40、cd137/4-1bb、淋巴细胞特异性蛋白酪氨酸激酶(lck)、诱导性t细胞共刺激因子(icos)和dnax激活蛋白10(dap10)的胞内结构域。
在一个或多个实施方案中,所述胞内共刺激信号域为cd137/4-1bb的胞内结构域;优选地,所述cd137/4-1bb的氨基酸序列如seqidno:7所示。
在一个或多个实施方案中,所述胞内信号域为cd3ζ胞内信号域或fcεriγ胞内信号域;优选为cd3ζ胞内信号域,优选地所述cd3ζ胞内信号域的氨基酸序列如seqidno:8所述。
在一个或多个实施方案中,所述嵌合抗原受体从n端到c端依次含有:scfv、cd8铰链区、cd8跨膜区、4-1bb和cd3ζ胞内信号域;优选地,所述嵌合抗原受体的氨基酸序列如seqidno:9第22-486位氨基酸残基所示。
在一个或多个实施方案中,所述嵌合抗原受体还含有信号肽,优选地,所述信号肽为分泌型信号肽和膜结合型信号肽,优选为cd8信号肽,更优选地,所述cd8信号肽的氨基酸序列如seqidno:1所示。
在一个或多个实施方案中,所述嵌合抗原受体的氨基酸序列如seqidno:9所示。
在一个或多个实施方案中,所述pd-1抗体的氨基酸序列如seqidno:11第21-495位氨基酸残基所示。
在一个或多个实施方案中,所述t细胞转入了含所述嵌合抗原受体编码序列的载体和含所述pd-1抗体的编码序列的载体。
在一个或多个实施方案中,所述载体为包含piggbac转座元件的真核表达载体。
在一个或多个实施方案中,所述嵌合抗原受体的编码序列及所述pd-1抗体的编码序列整合到所述t细胞的基因组中。
本发明还提供一种pd-1抗体,所述pd-1抗体的氨基酸序列如seqidno:11第21-495位氨基酸残基所示。
本发明还提供本发明pd-1抗体的编码序列及其互补序列,优选地,所述编码序列如seqidno:12第61-1488位氨基酸残基所示。
本发明还提供seqidno:11及其编码序列或互补序列。优选地,所示编码序列如seqidno:12所示。
本发明还提供一种组合物,所述组合物含有含本文所述嵌合抗原受体编码序列的载体和含本文所述pd-1抗体的编码序列的载体。
本发明还提供一种试剂盒,所述试剂盒含有含本文所述嵌合抗原受体编码序列的载体和含本文所述pd-1抗体的编码序列的载体,或者含有本文所述的组合物。
在一个或多个实施方案中,所述载体为包含piggbac转座元件的真核表达载体。
在一个或多个实施方案中,所述试剂盒还含有将所述载体转入细胞中的试剂。
本发明还提供一种药物组合物,所述药物组合物含有本文所述的t细胞。
本发明还提供本文所述的t细胞或其药物组合物在制备治疗治疗或预防恶性肿瘤的药物中的用途。
在一个或多个实施方案中,所述恶性肿瘤为恶性b细胞淋巴瘤,包括急性b淋巴细胞白血病(b-all)、慢性b淋巴细胞白血病(b-cll)、套细胞淋巴瘤(mcl)、nhl和多发性骨髓瘤(mm)。
附图说明
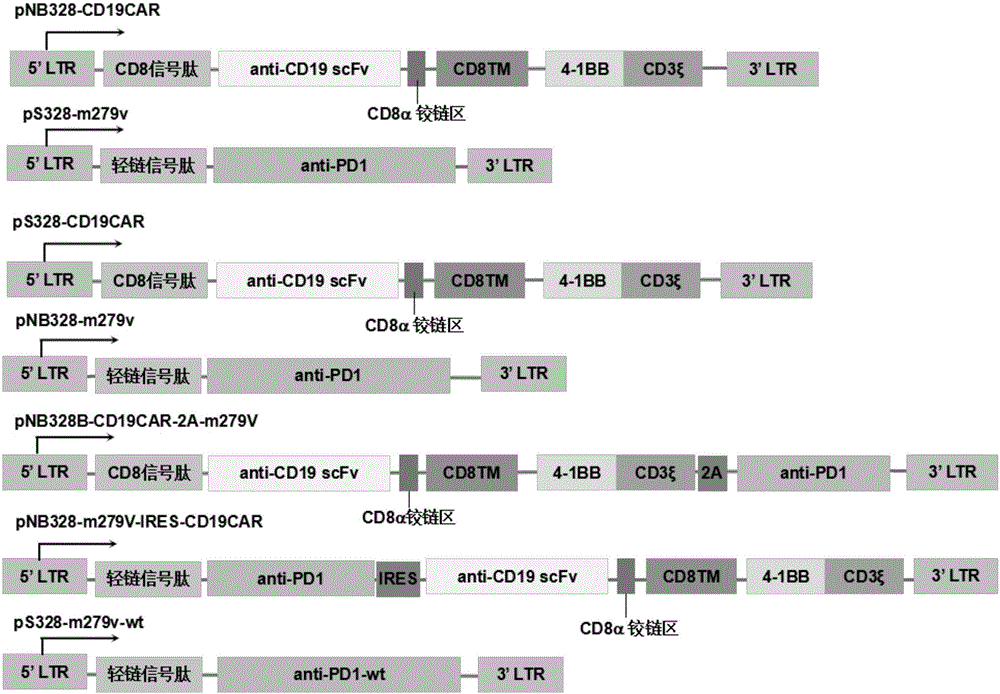
图1:pnb328-cd19car、ps328-m279v、ps328-cd19car、pnb328-m279v、pnb328-cd19car-2a-m279v、pnb328-m279v-ires-cd19car、ps328-m279v-wt的表达框模式。
图2:不同患者来源的pbmcs经cd19car基因及pd-1抗体基因修饰后检测cd19car基因及pd-1抗体基因表达的westernblotting检测图。
图3:不同患者来源的pbmcs经cd19car基因及pd-1抗体基因修饰活化后t细胞表达cd19car基因阳性率测定。
图4:不同患者来源的pbmcs经pd-1抗体基因修饰后pd-1抗体基因表达的elisa检测图。
图5:cd19car-antipd1多能t细胞的杀伤作用检测。
图6:cd19car-antipd1多能t细胞在体外可以增强t细胞的杀伤活性。
图7:cd19car-antipd1t细胞的体内杀伤功能检测。
具体实施方式
下面对本发明涉及的部分术语进行解释。
在本发明中,术语“表达框”是指表达一个基因所需的完整元件,包括启动子、基因编码序列、polya加尾信号序列。
术语“编码序列”在文中定义为核酸序列中直接确定其蛋白产物(例如car,单链抗体,铰链区和跨膜区)的氨基酸序列的部分。编码序列的边界通常是由紧邻mrna5’端开放读码框上游的核糖体结合位点(对于原核细胞)和紧邻mrna3’端开放读码框下游的转录终止序列确定。编码序列可以包括,但不限于dna、cdna和重组核酸序列。
术语“fc”即抗体的可结晶段(fragmentcrystallizable,fc),是指位于抗体分子"y"结构的柄部末端,包含抗体重链恒定区ch2和ch3结构域的肽段,是抗体与效应分子或者细胞相互作用的部位。
术语“共刺激分子”是指存在于抗原提呈细胞表面,能与th细胞上的共刺激分子受体结合,产生协同刺激信号的分子。淋巴细胞的增殖不仅需要抗原的结合,还需要接受共刺激分子的信号。共刺激信号传递给t细胞主要是通过表达在抗原呈递细胞表面的共刺激分子cd80,cd86与t细胞表面的cd28分子结合。b细胞接受共刺激信号可以通过一般的病原体成分例如lps,或者通过补体成分,或者通过激活了的抗原特异性的th细胞表面的cd40l。
术语“接头”或铰链是连接不同蛋白或多肽之间的多肽片段,其目的是使所连接的蛋白或多肽保持各自的空间构象,以维持蛋白或多肽的功能或活性。示例性的接头包括含有g和/或s的接头,以及例如furin2a肽。
术语“特异性结合”是指抗体或者抗原结合片段与其所针对的抗原之间的反应。在某些实施方式中,特异性结合某抗原的抗体(或对某抗原具有特异性的抗体)是指,抗体以小于大约10-5m,例如小于大约10-6m、10-7m、10-8m、10-9m或10-10m或更小的亲和力(kd)结合该抗原。“特异性识别”具有类似的含义。
术语“药学上可接受的辅料”是指在药理学和/或生理学上与受试者和活性成分相容的载体和/或赋形剂,其是本领域公知的(参见例如remington'spharmaceuticalsciences.editedbygennaroar,19thed.pennsylvania:mackpublishingcompany,1995),并且包括但不限于:ph调节剂,表面活性剂,佐剂,离子强度增强剂。例如,ph调节剂包括但不限于磷酸盐缓冲液;表面活性剂包括但不限于阳离子,阴离子或者非离子型表面活性剂,例如tween-80;离子强度增强剂包括但不限于氯化钠。
术语“有效量”是指可在受试者中实现治疗、预防、减轻和/或缓解本发明所述疾病或病症的剂量。
术语“疾病和/或病症”是指所述受试者的一种身体状态,该身体状态与本发明所述疾病和/或病症有关。
术语“受试者”或者“患者”可以指患者或者其它接受本发明药物组合物以治疗、预防、减轻和/或缓解本发明所述疾病或病症的动物,特别是哺乳动物,例如人、狗、猴、牛、马等。
术语“嵌合抗原受体”(car)是人工改造受体,能够将识别肿瘤细胞表面抗原的特异性分子(如抗体)锚定在免疫细胞(如t细胞)上,使免疫细胞识别肿瘤抗原或病毒抗原和杀死肿瘤细胞或病毒感染的细胞。car通常依次包含任选的信号肽、结合肿瘤细胞膜抗原的多肽如单链抗体、铰链区、跨膜区和胞内信号区。通常,结合肿瘤细胞膜抗原的多肽能够以中等亲和力结合肿瘤细胞广泛表达的膜抗原。结合肿瘤细胞膜抗原的多肽可以是天然多肽或人工合成多肽;优选地,人工合成多肽为单链抗体或fab片段。
术语“单链抗体”(scfv)是指由抗体轻链可变区(vl区)氨基酸序列和重链可变区(vh区)氨基酸序列经铰链连接而成,具有结合抗原能力的抗体片段。在某些实施方案中,感兴趣单链抗体(scfv)来自感兴趣的抗体。感兴趣的抗体可以是人抗体,包括人鼠嵌合抗体和人源化抗体。抗体可以是分泌型或膜锚定型;优选地为膜锚定型。
有研究表明pd-1抗体的igg4fc片段容易被单核/巨噬细胞识别而被吞噬,本发明对pd-1抗体igg4fc片段进行碱基突变改造以满足t细胞自身表达的pd-1抗体既可以很好的发挥功能又不引起adcc反应。
因此,本发明提供一种pd-1抗体,所述pd-1抗体含有抗pd-1单链抗体和igg4fc。在某些实施方案中,所述igg4fc的氨基酸序列如seqidno:11第267-495位氨基酸残基所示;优选地,其编码序列如seqidno:12第799-1485位碱基序列所示。
在某些实施方案中,所述抗pd-1单链抗体(scfv)中抗体轻链可变区(vl区)氨基酸序列如seqidno:11第21-131位氨基酸残基所示;优选地,其编码序列如seqidno:12第61-393位碱基序列所示。在某些实施方案中,所述抗pd-1单链抗体中的重链可变区(vh区)氨基酸序列如seqidno:11第147-266位氨基酸序列所示;优选地,其编码序列如seqidno:12第439-798位碱基序列所示。在某些实施方案中,所述抗pd-1单链抗体的氨基酸序列如seqidno:11第21-266位氨基酸残基所示;优选地,其编码序列如seqidno:12第61-798位碱基序列所示。
在某些实施方案中,所述pd-1抗体还含有轻链信号肽。在某些实施方案中,所述pd-1抗体从n端到c端,依次含有轻链信号肽、抗pd-1单链抗体和igg4fc。在某些实施方案中,所述轻链信号肽的氨基酸序列如seqidno:11第1-20位氨基酸残基所示;优选地,所示轻链信号肽的编码序列如seqidno:12第1-60位碱基序列所示。
在某些实施方案中,所述pd-1抗体的氨基酸序列如seqidno:11第21-495位氨基酸序列所示,或者如seqidno:11所示。
本发明还包括所述pd-1抗体的编码序列或其互补序列,所述编码序列至少包括本文所述的igg4fc的编码序列或其互补序列。在某些实施方案中,所述pd-1抗体的编码序列含有seqidno:12第61-1495位碱基序列所示的序列,优选含有seqidno:12所示的序列。
本发明还包括一种核酸构建物,所述核酸构建物含有本发明所述的pd-1抗体的编码序列或其互补序列。优选地,所述核酸构建物是表达载体或用于将所述编码序列或其互补序列整合入宿主细胞的整合载体。
本发明还提供一种宿主细胞,所述宿主细胞含有本文所述的核酸构建物。
本发明还提供所述pd-1抗体、其编码序列或互补序列、核酸构建物以及宿主细胞在制备治疗或预防恶性肿瘤中的用途,所述肿瘤尤其是与pd-1相关的肿瘤,包括但不限于本文所述的各种恶性肿瘤。
有研究表明,有多种方式可以将2种不同的蛋白在同一细胞中表达,包括以2a或ires连接2种基因片段形成单质粒后,再进行体外修饰细胞,该种方法可以确保当一种基因在某一细胞中表达后另一种基因必定能在同一细胞中表达,但该种质粒所携带的基因片段如果太长就会导致转录翻译的蛋白表达较弱。或者,也可将2种基因片段分别构建在2种载体上,同时体外修饰细胞。该种方法可以使得转录翻译的蛋白表达增强,但不能保证2种蛋白在同一个细胞中表达。
为了实现cd19car基因与pd-1抗体均能在细胞高效且稳定表达,本发明将cd19car基因与pd-1抗体做了多种组合形式的测试,包括以2a连接cd19car基因、pd-1抗体以及pb基因形成单个质粒、以ires连接cd19car基因、与pd-1抗体以及pb基因形成单个质粒、携带pb基因的cd19car基因质粒与pd-1抗体质粒的双质粒组合以及携带pb基因的pd-1抗体质粒与cd19car基因质粒的双质粒组合。通过测试表明携带pb基因的cd19car基因质粒与pd-1单链抗体质粒的双质粒组合能取得稳定的cd19car基因及pd-1抗体表达。
因此,本发明还提供一种经cd19car基因修饰并能表达pd-1抗体的多能t细胞,该t细胞能高水平稳定的表达cd19car基因及pd-1抗体,外源表达的cd19car基因可以准确的靶向cd19抗原,增强t细胞的增殖能力及细胞因子的分泌,表达的pd-1抗体能够帮助car-t细胞突破肿瘤微环境的抑制,从而增强car-t细胞对肿瘤细胞的杀伤,并通过增强免疫反应,发挥抗肿瘤作用。同时外源car基因及pd-1抗体基因可经pb转座酶系统整合到t细胞的基因组中,从而在t细胞中稳定持续的表达。本发明获得的高水平稳定表达car基因及pd-1抗体基因的t细胞可用于多种cd19高表达的恶性淋巴瘤的治疗。
本发明的car通常含有任选的信号肽序列、识别cd19抗原的scfv、铰链区、跨膜区、胞内共刺激信号域和胞内信号域。
信号肽是引导新合成的蛋白质向分泌通路转移的短肽链(长度5-30个氨基酸),常指新合成多肽链中用于指导蛋白质的跨膜转移(定位)的n-末端的氨基酸序列(有时不一定在n端),它负责把蛋白质引导到细胞含不同膜结构的亚细胞器内。信号肽可以是分泌型信号肽或膜结合型信号肽。在某些实施方案中,信号肽为cd8信号肽、cd28信号肽或cd4信号肽;更优选地为cd8信号肽。cd8信号肽的氨基酸序列可如seqidno:1所示;在某些实施方案中,其编码序列如seqidno:10第1-63位碱基所示。
识别cd19抗原的scfv可以是本领域常用的识别cd19抗原的scfv。在某些实施方案中,所述scfv的氨基酸序列如seqidno:2所示;在某些实施方案中,其编码序列如seqidno:10第64-789位碱基所示。
本文中,铰链区指免疫球蛋白重链ch1和ch2功能区之间的区域,该区富含脯氨酸,不形成α螺旋,易发生伸展及一定程度扭曲,有利于抗体的抗原结合部位与抗原表位间的互补性结合。适用于本文的铰链区可选自cd8的胞外铰链区、igg1fcch2ch3铰链区、igd铰链区、cd28的胞外铰链区、igg4fcch2ch3铰链区和cd4的胞外铰链区的任意一种或多种。铰链区优选是长50个氨基酸残基以上、更优选长80个氨基酸以上的铰链区。在某些实施方案中,本文使用cd8铰链区或igg4fcch2ch3铰链区。在某些实施方案中,cd8铰链区的氨基酸序列如seqidno:3所示;igg4ch2ch3铰链区的氨基酸序列如seqidno:4所示。在某些实施方案中,cd8铰链区的编码序列如seqidno:10第790-924位碱基所示。
跨膜区可以是cd28跨膜区、cd8跨膜区、cd3ζ跨膜区、cd134跨膜区、cd137跨膜区、icos跨膜区和dap10跨膜区中的一种;优选为cd8跨膜区,优选其氨基酸序列如seqidno:5所示;在某些实施方案中,其编码序列如seqidno:10第925-996位碱基所示。
胞内共刺激信号域包括共刺激信号分子的胞内结构域可选自cd28、cd134/ox40、cd137/4-1bb、淋巴细胞特异性蛋白酪氨酸激酶(lck)、诱导性t细胞共刺激因子(icos)和dnax激活蛋白10(dap10)的胞内结构域。在某些实施方案中,所述共刺激信号分子的胞内结构域为cd28的胞内结构域,优选其氨基酸序列如seqidno:6所示。在某些实施方案中,所述共刺激信号分子的胞内结构域为cd137/4-1bb的胞内结构域;优选地,所述cd137/4-1bb的氨基酸序列如seqidno:7所示;在某些实施方案中,其编码序列如seqidno:10第997-1122位碱基所示。
胞内信号域优选为免疫受体酪氨酸活化基序,可以是cd3ζ胞内信号域或fcεriγ胞内信号域;优选为cd3ζ胞内信号域,优选地所述cd3ζ胞内信号域的氨基酸序列如seqidno:8所述;在某些实施方案中,其编码序列如seqidno:10第1123-1458位碱基所示。
在某些实施方案中,所述嵌合抗原受体从n端到c端依次含有:scfv、cd8铰链区、cd8跨膜区、4-1bb和cd3ζ胞内信号域;优选地,所述嵌合抗原受体的氨基酸序列如seqidno:9第22-486位氨基酸残基所示。在某些实施方案中,所述嵌合抗原受体还含有信号肽,优选地,该嵌合抗原受体的氨基酸序列如seqidno:9所示。
应理解,本发明也包括本文所述的嵌合抗体受体及其编码序列。
形成本文嵌合抗原受体的上述各部分,如信号肽、抗muc1单链抗体的轻链可变区和重链可变区、铰链区、跨膜区、胞内共刺激信号域和胞内信号域等,相互之间可直接连接,或者可通过接头序列连接。接头序列可以是本领域周知的适用于抗体的接头序列,例如含g和s的接头序列。接头的长度可以是3~25个氨基酸残基,例如3~15、5~15、10~20个氨基酸残基。在某些实施方案中,接头序列是多甘氨酸接头序列。接头序列中甘氨酸的数量无特别限制,通常为2~20个,例如2~15、2~10、2~8个。除甘氨酸和丝氨酸来,接头中还可含有其它已知的氨基酸残基,例如丙氨酸(a)、亮氨酸(l)、苏氨酸(t)、谷氨酸(e)、苯丙氨酸(f)、精氨酸(r)、谷氨酰胺(q)等。
应理解,在基因克隆操作中,常常需要设计合适的酶切位点,这势必在所表达的氨基酸序列末端引入了一个或多个不相干的残基,而这并不影响目的序列的活性。为了构建融合蛋白、促进重组蛋白的表达、获得自动分泌到宿主细胞外的重组蛋白、或利于重组蛋白的纯化,常常需要将一些氨基酸添加至重组蛋白的n-末端、c-末端或该蛋白内的其它合适区域内,例如,包括但不限于,适合的接头肽、信号肽、前导肽、末端延伸等。因此,本文的car的氨基端或羧基端还可含有一个或多个多肽片段,作为蛋白标签。任何合适的标签都可以用于本文。例如,所述的标签可以是flag,ha,ha1,c-myc,poly-his,poly-arg,strep-tagii,au1,ee,t7,4a6,ε,b,ge以及ty1。这些标签可用于对蛋白进行纯化。
本文还包括编码所述嵌合抗原受体的多核苷酸序列。本文的多核苷酸序列可以是dna形式或rna形式。dna形式包括cdna、基因组dna或人工合成的dna。dna可以是单链的或是双链的。
本文所述的多核苷酸序列通常可以用pcr扩增法获得。具体而言,可根据本文所公开的核苷酸序列来设计引物,并用市售的cdna库或按本领域技术人员已知的常规方法所制备的cdna库作为模板,扩增得到有关序列。当序列较长时,常常需要进行两次或多次pcr扩增,然后再将各次扩增出的片段按正确次序拼接在一起。例如,在某些实施方案中,编码本文所述融合蛋白的多核苷酸序列如seqidno:14所示。
本文还包括核酸构建物,其含有本文所述的编码所述嵌合抗原受体的多核苷酸序列或编码所述pd-1抗体的多核苷酸序列,以及与这些序列操作性连接的一个或多个调控序列。在某些实施方案中,所述核酸构建物是表达框。
调控序列可以是合适的启动子序列。启动子序列通常与待表达蛋白的编码序列操作性连接。启动子可以是在所选择的宿主细胞中显示转录活性的任何核苷酸序列,包括突变的、截短的和杂合启动子,并且可以从编码与该宿主细胞同源或异源的胞外或胞内多肽的基因获得。
调控序列也可以是合适的转录终止子序列,由宿主细胞识别以终止转录的序列。终止子序列与编码该多肽的核苷酸序列的3’末端操作性连接。在选择的宿主细胞中有功能的任何终止子都可用于本文。
在某些实施方案中,所述核酸构建物是载体。具体而言,可将本文car的编码序列或pd-1抗体的编码序列克隆入许多类型的载体,例如这些类型的载体包括但不限于质粒、噬菌粒、噬菌体衍生物、动物病毒和粘粒。载体可以是表达载体。表达载体可以以病毒载体形式提供给细胞。可用作载体的病毒包括但不限于逆转录病毒、腺病毒、腺伴随病毒、疱疹病毒和慢病毒。
通常,合适的载体包含在至少一种有机体中起作用的复制起点、启动子序列、方便的限制酶位点和一个或多个可选择的标记。例如,在某些实施方案中,本发明使用逆转录病毒载体,该逆转录病毒载体含有复制起始位点,3’ltr,5’ltr,本文所述car的编码序列或pd-1抗体的编码序列,以及任选的可选择的标记。
合适的启动子包括但不限于即时早期巨细胞病毒(cmv)启动子序列。该启动子序列是能够驱动可操作地连接至其上的任何多核苷酸序列高水平表达的强组成型启动子序列。合适的启动子的另一个例子为延伸生长因子-1α(ef-1α)。然而,也可使用其他组成型启动子序列,包括但不限于类人猿病毒40(sv40)早期启动子、小鼠乳癌病毒(mmtv)、人免疫缺陷病毒(hiv)长末端重复(ltr)启动子、momulv启动子、鸟类白血病病毒启动子、eb病毒即时早期启动子、鲁斯氏肉瘤病毒启动子、以及人基因启动子,诸如但不限于肌动蛋白启动子、肌球蛋白启动子、血红素启动子和肌酸激酶启动子。进一步地,也可考虑使用诱导型启动子。诱导型启动子的使用提供了分子开关,其能够在期限表达时打开可操作地连接诱导型启动子的多核苷酸序列的表达,而在当表达是不期望的时关闭表达。诱导型启动子的例子包括但不限于金属硫蛋白启动子、糖皮质激素启动子、孕酮启动子和四环素启动子。
在某些实施方案中,可使用cn201510021408.1所公布的各种启动子序列,包括但不限于该申请seqidno:5所示的含mcmv增强子、hcmv增强子和ef1α启动子的ccef启动子;seqidno:7所示的含cd3e增强子、mcmv增强子、hcmv增强子和ef1α启动子的tcef启动子;seqidno:8所示的含mcmv增强子、hcmv增强子和含内含子的ef1α启动子的ccefi启动子;seqidno:3所示的含cd3e增强子和含内含子的ef1α启动子的tefi启动子;以及seqidno:3所示的含cd3e增强子、mcmv增强子、hcmv增强子和含内含子的ef1α启动子的tcefi启动子。本文将该申请的全部内容以引用的方式纳入本文。
可选择的标记包括可选择的标记基因或报道基因中的任一个或两者,以便于从被病毒载体感染的细胞群中鉴定和选择表达细胞。有用的可选择标记基因包括例如抗生素抗性基因,诸如neo等。合适的报道基因可包括编码荧光素酶、β-半乳糖苷酶、氯霉素乙酰转移酶、分泌型碱性磷酸酶或绿色萤光蛋白基因的基因。
在某些实施方案中,可将本文所述的嵌合抗原受体的编码序列和pd-1抗体的编码序列分别克隆到用于将目的核酸序列整合到宿主细胞的基因组中的载体(也称为整合载体)中,尤其是转座子载体。在某些实施方案中,该转座子载体是含有选自piggybac、sleepingbeauty、frogprince、tn5或ty的转座元件的真核表达载体。这类转座子载体含有相应转座子的5’反向末端重复序列(5’ltr)和相应转座子的3’反向末端重复序列(3’ltr)。转座酶可以是来自piggybac、sleepingbeauty、frogprince、tn5或ty转座系统的转座酶。当使用来自不同转座系统的转座酶时,所述载体中的5’ltr和3’ltr的序列也相应改变为与该转座系统适配的序列,这可由本领域技术人员容易地确定。在某些实施方案中,在5’ltr和3’ltr之间是本发明的car或抗体的表达框,包括相应的启动子序列、car或抗体的编码序列以及polya加尾信号序列。
在某些实施方案中,转座酶是来自piggybac转座系统的转座酶。因此,在这些实施方案中,转座子5’反向末端重复序列和3’反向末端重复序列分别为piggybac转座子的5’反向末端重复序列和3’反向末端重复序列。在某些实施方案中,转座子5’反向末端重复序列如cn201510638974.7(本文将其内容以引用的方式纳入本文)seqidno:1所示。在某些实施方案中,转座子3’反向末端重复序列如cn201510638974.7seqidno:4所示。在某些实施方案中,piggybac转座酶为含c-myc核定位信号编码序列的转座酶。在某些实施方案中,piggybac转座酶的编码序列如cn201510638974.7seqidno:5所示。
转座酶编码序列的启动子可以是本领域已知的用于控制转座酶编码序列表达的各种启动子。在某些实施方案中,使用cmv启动子控制转座酶编码序列的表达。cmv启动子的序列可如cn201510638974.7seqidno:6所示。
在某些实施方案中,本发明含嵌合抗原受体的编码序列的载体为cn201510638974.7所公开的pnb328载体。可采用本领域常规的方法制备本发明的嵌合抗原受体的编码序列,并将其克隆入合适的载体中。
在某些实施方案中,所述用于将目的基因整合到宿主细胞的基因组中的载体不含有转座酶编码序列。例如,可在pnb328载体的基础上除去转座酶编码序列即可获得这类载体。通常,用这类载体将pd-1抗体的编码序列及信号肽编码序列(如轻链信号肽的编码序列)整合到宿主细胞的基因组中。
在某些实施方案中,本文所述的经cd19car基因修饰并能表达pd-1抗体的t细胞可转入:
(1)用于在t细胞基因组中整合入嵌合抗原受体表达框的含转座酶编码序列的载体,和用于在t细胞基因组中整合入本文所述的pd-1抗体的表达框的含转座酶编码序列的载体;
(2)用于在t细胞基因组中整合入嵌合抗原受体表达框的不含转座酶编码序列的载体,和用于在t细胞基因组中整合入本文所述的pd-1抗体的表达框的不含转座酶编码序列的载体;
(3)用于在t细胞基因组中整合入嵌合抗原受体表达框的含转座酶编码序列的载体,和用于在t细胞基因组中整合入本文所述的pd-1抗体的表达框的不含转座酶编码序列的载体;或
(4)用于在t细胞基因组中整合入嵌合抗原受体表达框的不含转座酶编码序列的载体,和用于在t细胞基因组中整合入本文所述的pd-1抗体的表达框的含转座酶编码序列的载体。
优选地,所述t细胞转入了用于在t细胞基因组中整合入嵌合抗原受体表达框的含转座酶编码序列的载体,和用于在t细胞基因组中整合入本文所述的pd-1抗体的表达框的不含转座酶编码序列的载体。更优选地,所述t细胞转入了以pnb328载体为骨架载体构建的含嵌合抗原受体表达框的载体以及以ps328载体(与pnb328相比不含转座酶编码序列)为骨架载体构建的含pd-1抗体表达框的载体。在某些实施方案中,所述嵌合抗原受体的编码序列如seqidno:10所示;所述pd-1抗体的编码序列如seqidno:12第61-1488位碱基序列。在某些实施方案中,所述含pd-1抗体的编码序列的载体中,pd-1抗体的信号肽为轻链信号肽。示例性的轻链信号肽的氨基酸序列可如seqidno:13第1-20位氨基酸残基所示。更具体而言,在某些实施方案中,所述在t细胞基因组中整合入嵌合抗原受体编码序列的含转座酶编码序列的载体依次案由5’ltr、启动子、cd8信号肽编码序列、识别cd19抗原的scfv的编码序列、cd8铰链区的编码序列、cd8跨膜区的编码序列、4-1bb的编码序列、cd3ζ胞内信号域的编码序列、polya加尾信号序列、3’ltr和转座酶的编码序列及其启动子;所述在t细胞基因组中整合入本文所述的pd-1抗体的编码序列的不含转座酶编码序列的载体在5’ltr和3’ltr之间依次含有启动子、轻链信号肽的编码序列、pd-1抗体的编码序列和polya加尾信号序列。
优选地,转染时,含嵌合抗原受体编码序列的载体与含pd-1抗体编码序列的载体的质量比为1:1~7,如1:1~5,优选1:1~3,更优选1:1~2,更优选1:1。
转染的方法为本领域常规的方法,包括但不限于:病毒转导、显微注射、粒子轰击、基因枪转化和电转等。在某些实施方案中,采用电转将所述载体转染感兴趣的细胞中。
感兴趣的细胞可以是本领域周知的各种t细胞,包括但不限于外周血t淋巴细胞、细胞毒杀伤t细胞(ctl)、辅助t细胞、抑制/调节性t细胞、γδt细胞以及细胞因子诱导的杀伤细胞(cik)、肿瘤浸润淋巴细胞(til)等混合细胞群体的t细胞。
本发明还提供一种组合物,所述组合物含有含本文所述嵌合抗原受体表达框的载体和含本文所述pd-1抗体的表达框的载体。该组合物中还可含有合适的试剂,包括但不限于转染用的试剂。
本发明还提供一种试剂盒,所述试剂盒含有含本文所述嵌合抗原受体表达框的载体和含本文所述pd-1抗体的表达框的载体,或者含有本文所述的组合物。试剂盒中还可配有将所述载体转入细胞中的试剂或仪器。
应理解,本文所述的表达框除含有本文所述的car或抗体的编码序列外,还至少含有合适的启动子和polya加尾信号序列。
本发明还提供一种药物组合物,所述药物组合物含有本文所述的t细胞或所述t细胞及其表达的pd-1抗体。药物组合物中可含有合适的药学上可接受的载体或辅料。药物组合物中含有治疗或预防有效量的t细胞。可根据患者的病情等因素确定t细胞的治疗或预防有效量。
本发明还提供本文所述的t细胞或所述t细胞及其表达的pd-1抗体或其药物组合物在制备治疗治疗或预防恶性肿瘤的药物中的用途。
本发明还提供恶性肿瘤的治疗或预防方法,所述方法包括给予需要的对象治疗或预防有效量的本发明所述的t细胞。恶性肿瘤为恶性b细胞淋巴瘤,包括急性b淋巴细胞白血病(b-all)、慢性b淋巴细胞白血病(b-cll)、套细胞淋巴瘤(mcl)、nhl和多发性骨髓瘤(mm)。
本发明特异性靶向cd19抗原且能高水平稳定表达pd-1抗体的多能t细胞能够促进t细胞的增殖及其在肿瘤组织的浸润,增强t细胞的杀伤功能,该种多能t细胞在肿瘤免疫细胞治疗或癌症治疗方面发挥细胞免疫和体液免疫的双重作用。这些细胞可以通过非mhc限制性途径特异性识别肿瘤表面抗原,发挥杀伤毒性,从而高效特异性杀灭肿瘤细胞。
下面将结合实施案例对本发明所涉及的实施方案进行详细描述。本领域技术人员将会理解,下面的实施案例仅用于说明本发明,而不应视为限定本发明的范围。实施案例中未注明具体技术或条件者,按照本领域内的文献所描述的技术或条件(例如参考j.萨姆布鲁克等著,黄培堂等译的《分子克隆实验指南》,第三版,科学出版社)或者按照产品说明书进行。
所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规产品。
实施例1:重组质粒pnb328-cd19car、ps328-m279v、ps328-cd19car、pnb328-m279v、pnb328-cd19car-2a-m279v、pnb328-m279v-ires-cd19car的构建和多种类型多能t细胞的获得。
1、委托上海捷瑞生物公司合成cd19car外源基因(含cd8信号肽,scfv,cd8跨膜区,cd8铰链区,4-1bb和cd3ζ),并在其上游引入多克隆酶切位点(bglii-xbai-ecori-bamhi),在其下游插入酶切位点(sali-nhei-hindiii-spei),将其装入用ecor1+sali双酶切的pnb328载体或ps328-ef1α载体中(pnb328的结构及序列参见cn201510638974.7,本文将其全部内容以引用的方式纳入本文;与pnb328相比,ps328缺少转座酶编码序列)构成重组质粒,分别命名为pnb328-cd19car和ps328-cd19car。
委托上海杰瑞生物公司合成突变型pd-1抗体的外源基因(该外源基因含有轻链信号肽的编码序列及突变型pd-1抗体的编码序列;轻链信号肽的氨基酸序列如seqidno:11第1-20位氨基酸残基所示,编码序列如seqidno:12第1-60位碱基所示;编码突变型pd-1抗体的核苷酸序列如seqidno:12第61-1488位碱基所示,氨基酸序列如seqidno:11第21-495位氨基酸残基所示),并在其上游引入多克隆酶切位点(bglii-xbai-ecori-bamhi),在其下游插入酶切位点(sali-nhei-hindiii-spei),将其装入用ecor1+sali双酶切的ps328载体或pnb328载体中,构成重组质粒,分别命名为ps328-m279v和pnb328-m279v。
将前述合成的cd19car外源基因和pd-1抗体的编码序列(seqidno:12)以2a(核苷酸序列如seqidno:15所示,氨基酸序列如seqidno:16所示)连接人工,并在其上游引入多克隆酶切位点(bglii-xbai-ecori-bamhi),在其下游插入酶切位点(sali-nhei-hindiii-spei),委托上海捷瑞生物公司合成,将其装入用ecor1+sali双酶切的pnb328载体中构成重组质粒,命名为pnb328-cd19car-2a-m279v。
将前述合成的pd-1抗体的外源基因与cd19car外源基因以ires(核苷酸序列如seqidno:17所示)连接,并在其上游引入多克隆酶切位点(bglii-xbai-ecori-bamhi),在其下游插入酶切位点(sali-nhei-hindiii-spei),委托上海捷瑞生物公司合成,将其装入用ecor1+sali双酶切的pnb328载体中构成重组质粒,命名为pnb328-m279v-ires-cd19car。
委托上海杰瑞生物公司合成野生型pd-1抗体的外源基因(该外源基因含有轻链信号肽的编码序列及野生型pd-1抗体的编码序列;轻链信号肽的氨基酸序列如seqidno:13第1-20位氨基酸残基所示,编码序列如seqidno:14第1-60位碱基所示;编码野生型pd-1抗体的核苷酸序列如seqidno:13第61-1488位碱基所示,氨基酸序列如seqidno:14第21-495位氨基酸残基所示),并在其上游引入多克隆酶切位点(bglii-xbai-ecori-bamhi),在其下游插入酶切位点(sali-nhei-hindiii-spei),将其装入用ecor1+sali双酶切的ps328载体中,构成重组质粒,命名为ps328-m279v-wt。
各载体结构分别如图1所示。各结构模式图中未示出启动子序列和polya加尾信号序列,其分别位于5’ltr和信号肽序列之间以及3’ltr之前。
2、获得cd19car-antipd1多能t细胞
利用filcoll分离法从患者血液中分离获得外周血单核细胞(pbmcs)。将pbmc贴壁培养2-4h,其中未贴壁的悬浮细胞即为初始t细胞,将悬浮细胞收集到15ml离心管中,1200rmp离心3min,弃上清,加入生理盐水,1200rmp离心3min,弃生理盐水,并重复此步骤。
取六个1.5ml离心管,每管加入5×106个细胞,编号a、b、c、d、e、f,1200rmp离心3min,弃上清,取电转试剂盒(来自lonza公司),a、b、c、d、e、f管按比例加入电转试剂共100ul,a管加入4ug重组质粒ps328-cd19car和4ug重组质粒pnb328-m279v;b管加入4ug重组质粒pnb328-cd19car和4ug重组质粒ps328-m279v;c管加入pnb328-cd19car-2a-m279v质粒;d管加入pnb328-m279v-ires-cd19car;e管中加入6ug重组质粒pnb328-cd19car;f管中加入4ug重组质粒pnb328-cd19car和4ug重组质粒ps328-m279v-wt。分别重悬混匀细胞;将混合液转移至电转杯中,放入电转仪,选取所需程序,进行电击;使用试剂盒中的微量吸管将电转好的细胞悬液转移到加好培养液的六孔板中(含2%fbs的aim-ⅴ培养液),混匀,置于37℃,5%co2培养箱培养,获得转入了不同载体的活化t细胞。
实施例2:pbmcs经cd19car基因及pd-1抗体基因不同组合形式修饰活化后表达cd19car基因阳性率及pd-1抗体表达测定。
将实施例1构建的活化t细胞分别按2×106细胞数目收集细胞并按2×106细胞/孔,铺在加有3mlaim-v培液的6孔板中,置于37℃,5%co2培养箱培养,并于培养24h后收集细胞上清,-20℃保存备用。
用双抗体夹心elisa方法(使用人源的pd-1重组蛋白包被酶标板,带hrp标记的鼠抗人igg4mab)检测,以商品化的pd-1抗体作为标准品,待测样品5倍稀释后定量检测基因修饰后的t细胞中pd-1抗体的表达量。同时收集1×106细胞沉淀,用pbs清洗细胞沉淀2遍,加入5ul的biotin-cd19抗体,4℃孵育30min,pbs清洗细胞沉淀2遍,加入2ulpe-链霉素二抗,pbs清洗细胞沉淀2遍,加入400ulpbs将细胞转移至流式管中,上机检测。
结果如下表1所示。
表1
结果显示,由pnb328-cd19car质粒和ps328-m279v质粒组合电转所获得的t细胞(本文命名为cd19car-antipd1t细胞)具有较高的cd19car基因及pd-1抗体分泌。
实施例3:pbmcs经pnb328-cd19car与ps328-m279v质粒不同质量比修饰后表达cd19car基因阳性率及pd-1抗体表达量测定。
将实施例1构建得到的重组质粒pnb328-cd19car和ps328-m279v以不同质量比电转pbmcs细胞(1:1,3:5,1:3,1:7),获得多种同时表达cd19car基因和pd-1抗体的t细胞。将这些t细胞按2×106细胞数目收集细胞并按2×106细胞/孔,铺在加有3mlaim-v培液的6孔板中,置于37℃,5%co2培养箱培养,并于培养24h后收集细胞上清,-20℃保存备用。
用双抗体夹心elisa方法(使用人源的pd-1重组蛋白包被酶标板,带hrp标记的鼠抗人igg4mab)检测,以商品化的pd-1抗体作为标准品,待测样品5倍稀释后定量检测基因修饰后的t细胞中pd-1抗体的表达量。同时收集1×106细胞沉淀,用pbs清洗细胞沉淀2遍,加入5ul的biotin-cd19抗体,4℃孵育30min,pbs清洗细胞沉淀2遍,加入2ulpe-链霉素二抗,pbs清洗细胞沉淀2遍,加入400ulpbs将细胞转移至流式管中,上机检测。
结果如下表2所示。
表2
结果显示,由pnb328-cd19car质粒和ps328-m279v质粒以1:1质量比进行电转所获得的t细胞具有较高的cd19car基因及pd-1抗体分泌。
实施例4:不同患者来源的pbmcs经cd19car基因及pd-1抗体基因修饰后检测cd19car基因及pd-1抗体基因表达的westernblotting检测图。
将实施例1构建得到的重组质粒pnb328-cd19car和ps328-m279v以质量比为1:1的比例电转不同患者的pbmc,获得的活化t细胞按2×106细胞数目收集细胞沉淀,pbs清洗细胞沉淀1遍,加入80ul的2xsds-page加样缓冲液100℃煮沸10min,-20℃保存备用。
westernblotting(使用cd3抗体以及igg4fc抗体作为一抗,hrp-羊抗人为二抗)实验检测cd19car及pd-1抗体的表达。结果发现如图2所示,t细胞可以稳定表达cd19car重组蛋白(上图)及pd-1抗体(下图)。
实施例5:pbmcs经cd19car基因及pd-1抗体基因修饰活化后t细胞表达cd19car基因阳性率测定。
将实施例1构建得到的重组质粒pnb328-cd19car和ps328-m279v以质量比为1:1的比例电转pbmc,获得的活化t细胞按1×106细胞数目收集细胞沉淀,pbs清洗细胞沉淀2遍,加入5ul的biotin-cd19抗体,4℃孵育30min,pbs清洗细胞沉淀2遍,加入2ulpe-链霉素二抗,pbs清洗细胞沉淀2遍,加入400ulpbs将细胞转移至流式管中,上机检测。
结果如图3所示,cd19car重组蛋白可以在t细胞表面稳定表达。
实施例6:不同患者来源的pbmcs经cd19car基因及pd-1抗体基因修饰活化后t细胞表达pd-1抗体表达量的定量检测。
将实施例1构建得到的重组质粒pnb328-cd19car和ps328-m279v以质量比为1:1的比例电转不同患者的pbmc,获得的活化t细胞按2×106细胞数目收集细胞并按2×106细胞/孔,铺在加有3mlaim-v培液的6孔板中,置于37℃,5%co2培养箱培养,并于培养24h后收集细胞上清,-20℃保存备用。
用双抗体夹心elisa方法(使用人源的pd-1重组蛋白包被酶标板,带hrp标记的鼠抗人igg4mab)检测,以商品化的抗pd-1抗体作为标准品,待测样品50倍稀释后定量检测经cd19car基因及pd-1抗体基因修饰后的t细胞中pd-1抗体的表达量。
结果如图4所示,经pd-1抗体基因修饰的cd19cart细胞能够高水平稳定表达pd-1抗体(下图;上图为标准品的抗体浓度曲线)。
实施例7:不同患者来源的pbmcs细胞经cd19car基因及pd-1抗体基因修饰后t细胞基因组中cd19car基因组表达水平的检测。
提取mockt细胞,实施例1制备得到的仅转入了pnb328-cd19car的cd19cart细胞及实施例1制备得到的转入了pnb328-cd19car和ps328-m279v的cd19car-antipd1t细胞的基因组dna(试剂盒法),实验步骤参照试剂盒内附带的说明书,测定mockt细胞、cd19cart细胞及cd19car-antipd1t细胞的dna浓度,采用实时荧光定量pcr的方法检测cd19car基因组的表达水平,反应程序为:50℃,2min→95℃,10min→95℃,15s→60℃,1min,40个循环。得到的cd19car基因组的ct值及actin的ct值根据相应的公式计算出绝对拷贝数含量。
结果如下表3所示。
表3
结果显示,经pb转座酶系统,cd19car基因组被整合到t细胞基因组中。
实施例8:mockt细胞、cd19cart细胞及cd19car-antipd1t细胞在体外对培养的肿瘤细胞的杀伤实验。
选取mhcclassi分型匹配的效应细胞与靶细胞,应用delfiaeutda细胞毒性实验检测cart细胞的体外杀伤活性,具体步骤如下:
(1)离心收集raji细胞,并用pbs清洗1次;
(2)离心,收集细胞沉淀并用1640培养基重悬细胞并计数,调整细胞密度为1×106/ml;
(3)取2-4ml上述细胞,加入5ul荧光增强配体,置于37℃,5%co2细胞培养箱中20min;
(4)pbs清洗细胞3-5次;
(5)离心,收集细胞沉淀并用1640培养基重悬细胞并计数,调整细胞密度为5×104/ml,取100ul细胞悬液加入96孔培养板中。
(6)将mockt细胞、实施例1制备得到的仅转入了pnb328-cd19car的cd19cart细胞及实施例1制备得到的转入了pnb328-cd19car和ps328-m279v的cd19car-antipd1t细胞进行细胞计数,按照不同的效靶比4:1取100ul细胞悬液对应加入上述raji细胞中,并设置高对照组(肿瘤细胞加裂解液裂解),低对照组(仅含有肿瘤细胞),空白对照组(仅含有培养基);
(7)置于37℃,5%co2细胞培养箱中20min共培养3h;
(8)转移20ul培养上清至96孔白板中;
(9)加入200uleuropium溶液;
(10)室温振荡混匀15min;
(11)在酶标仪中使用时间分辨荧光检测,读值;
结果如图5所示,cd19cart细胞及cd19car-antipd1t具有较强的对肿瘤细胞的杀伤作用且杀伤作用相当。
实施例9:流式检测mockt细胞、cd19cart细胞和cd19car-antipd1t细胞活化表型和细胞因子分泌的区别。
1、收集悬浮的mockt细胞、实施例1制备得到的仅转入了pnb328-cd19car的cd19cart细胞和实施例1制备得到的转入了pnb328-cd19car和ps328-m279v的cd19car-antipd1t细胞,pbs清洗两遍,1200rpm离心5min,分别加入2ul的同型对照抗体igg1-pe,荧光流式抗体anti-cd69-pe、anti-klrg1-pe,anti-pd1-pe;同型对照抗体igg1-pc5,荧光流式抗体anti-cd107-pc5;同型对照抗体igg1fitc,荧光流式抗体anti-cd62l-fitc;同型对照抗体igg1-pc5,荧光流式抗体anti-cd45ro-pc5;同型对照抗体igg1-pe,荧光流式抗体anti-ccr7-pe。轻弹沉淀使其混合均匀,室温避光孵育30min,pbs清洗一遍,加入400ulpbs将细胞转移至流式管中,上机检测。
结果如图6(第1和2张图)所示,cd19car-antipd1t细胞分泌的pd-1单链抗体可以很好的封闭t细胞表面的pd-1蛋白,且cd19cart细胞和cd19car-antipd1t细胞在体外均有明显的杀伤活性,同时也能促进记忆t的形成,而活化标记cd69明显高于mockt细胞而耗竭标记lag3要明显低于mockt细胞。
2、用5ug/ml的cd19抗原包被24孔板,4℃包被过夜,pbs清洗3遍,加入3×105的mockt细胞、cd19cart细胞或cd19car-antipd1t细胞,培养24h后收集细胞上清。用bdtmcbahumanth1/th2cytokinekitii检测cd19cart细胞和cd19car-antipd1t细胞受cd19抗原刺激后细胞因子的分泌情况:
(1)混合人的il-2、il-4、il-6、il-10、tnf-α、ifn-γ捕获磁珠,涡旋振荡混匀捕获磁珠,每管加入50ul混匀后的捕获磁珠;
(2)加入50ul人的th1/th2细胞因子标准品(倍比稀释5000pg/ml、2500pg/ml、1250pg/ml、625pg/ml、312.5pg/ml、156pg/ml、80pg/ml、40pg/ml、20pg/ml、0pg/ml)和50ul的待测样品(经稀释液2倍稀释);
(3)每管加入50ul的人的th1/th2-ii-pe的检测抗体;
(4)室温避光孵育3h;
(5)每管加入1ml的洗涤缓冲液,200离心5min,弃上清;
(6)每管加入300ul的洗涤缓冲液重悬细胞,并转移至流式管中,用流式细胞仪检测荧光值;
结果如图6(第3张图)所示,cd19cart细胞及cd19car-antipd1t细胞分泌的il-2,tnf-α和ifn-γ相比于mockt细胞而言有了很大的提高,而三种细胞分泌的il-4、il-6和il-10并没有实质差异。
3、收集1×106个的cd19cart细胞和cd19car-antipd1t细胞,分别加入1.5ml的ep管中,pbs清洗两遍,1200rpm离心5min,加入2ul的-cd3cd4cd8抗体,室温避光孵育30min,pbs清洗一遍,加入400ulpbs将细胞转移至流式管中,上机检测。
结果如图6(第4张图)所示,cd19cart细胞、cd19car-antipd1t细胞和mockt中cd3+cd4+、cd3+cd8+细胞所占的百分比并没有很大的差异。
实施例10:cd19cart细胞、cd19car-antipd1t细胞以及cd19car-antipd1-wtt细胞的体内功能实验。
实验使用4~6周龄nsg完全免疫缺陷小鼠12只,平均重量22~27g,由北京百奥赛图生物技术有限公司提供,spf级动物实验室饲养。
体外培育人b细胞淋巴瘤raji-luc细胞,取对数生长期生长细胞,离心、收集细胞后用pbs液重悬,3000g室温离心2分钟,弃上清,再用pbs液重悬后离心收集细胞,调整细胞悬液浓度至5×107个/ml。于小鼠右肋背部皮下接种所述raji-luc细胞,0.1ml/只。接种10天左右后,可通过活体成像仪观察肿瘤大小,将nsg免疫缺陷小鼠随机分分为4组。分别为pbs组、mockt组、cd19cart组、cd19car-antipd1t组和cd19car-antipd1-wtt组(实施例1制备得到的转入了pnb328-cd19car和ps328-m279v-wt的t细胞),每组注射相应的t细胞为1×107个/100ul,pbs组给予100ul的pbs,给药途径为尾静脉注射。每日观察小鼠的生活状态并每隔7-8天通过活体成像仪观察小鼠肿瘤变化。
结果如图7所示。
尽管本发明的具体实施方式已经得到详细的描述。本领域技术人员将会理解。根据已经公开的所有教导,可以对那些细节进行各种修改和替换,这些改变均在本发明的保护范围之内。本发明的全部范围由所附权利要求及其任何等同物给出。
序列表
<110>上海细胞治疗研究院
上海细胞治疗工程技术研究中心集团有限公司
<120>特异性靶向cd19抗原且高水平稳定表达pd-1抗体的car-t细胞及用途
<130>179825
<160>17
<170>siposequencelisting1.0
<210>1
<211>21
<212>prt
<213>人工序列(artificialsequence)
<400>1
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargpro
20
<210>2
<211>242
<212>prt
<213>人工序列(artificialsequence)
<400>2
aspileglnmetthrglnthrthrserserleuseralaserleugly
151015
aspargvalthrilesercysargalaserglnaspileserlystyr
202530
leuasntrptyrglnglnlysproaspglythrvallysleuleuile
354045
tyrhisthrserargleuhisserglyvalproserargphesergly
505560
serglyserglythrasptyrserleuthrileserasnleuglugln
65707580
gluaspilealathrtyrphecysglnglnglyasnthrleuprotyr
859095
thrpheglyglyglythrlysleugluilethrglyglyglyglyser
100105110
glyglyglyglyserglyglyglyglysergluvallysleuglnglu
115120125
serglyproglyleuvalalaproserglnserleuservalthrcys
130135140
thrvalserglyvalserleuproasptyrglyvalsertrpilearg
145150155160
glnproproarglysglyleuglutrpleuglyvaliletrpglyser
165170175
gluthrthrtyrtyrasnseralaleulysserargleuthrileile
180185190
lysaspasnserlysserglnvalpheleulysmetasnserleugln
195200205
thraspaspthralailetyrtyrcysalalyshistyrtyrtyrgly
210215220
glysertyralametasptyrtrpglyglnglythrservalthrval
225230235240
serser
<210>3
<211>45
<212>prt
<213>人工序列(artificialsequence)
<400>3
thrthrthrproalaproargproprothrproalaprothrileala
151015
serglnproleuserleuargproglualacysargproalaalagly
202530
glyalavalhisthrargglyleuaspphealacysasp
354045
<210>4
<211>228
<212>prt
<213>人工序列(artificialsequence)
<400>4
gluserlystyrglyproprocysproprocysproalaproproval
151015
alaglyproservalpheleupheproprolysprolysaspthrleu
202530
metileserargthrprogluvalthrcysvalvalvalaspvalser
354045
glngluaspprogluvalglnpheasntrptyrvalaspglyvalglu
505560
valhisasnalalysthrlysproargglugluglnpheglnserthr
65707580
tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn
859095
glylysglutyrlyscyslysvalserasnlysglyleuproserser
100105110
ileglulysthrileserlysalalysglyglnproarggluprogln
115120125
valtyrthrleuproproserglngluglumetthrlysasnglnval
130135140
serleuthrcysleuvallysglyphetyrproseraspilealaval
145150155160
glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro
165170175
provalleuaspseraspglyserphepheleutyrserargleuthr
180185190
valasplysserargtrpglngluglyasnvalphesercysserval
195200205
methisglualaleuhisasnhistyrthrglnlysserleuserleu
210215220
serleuglylys
225
<210>5
<211>24
<212>prt
<213>人工序列(artificialsequence)
<400>5
iletyriletrpalaproleualaglythrcysglyvalleuleuleu
151015
serleuvalilethrleutyrcys
20
<210>6
<211>28
<212>prt
<213>人工序列(artificialsequence)
<400>6
prophetrpvalleuvalvalvalglyglyvalleualacystyrser
151015
leuleuvalthrvalalapheileilephetrpval
2025
<210>7
<211>42
<212>prt
<213>人工序列(artificialsequence)
<400>7
lysargglyarglyslysleuleutyrilephelysglnprophemet
151015
argprovalglnthrthrglnglugluaspglycyssercysargphe
202530
proglugluglugluglyglycysgluleu
3540
<210>8
<211>112
<212>prt
<213>人工序列(artificialsequence)
<400>8
argvallyspheserargseralaaspalaproalatyrglnglngly
151015
glnasnglnleutyrasngluleuasnleuglyargarggluglutyr
202530
aspvalleuasplysargargglyargaspproglumetglyglylys
354045
proargarglysasnproglngluglyleutyrasngluleuglnlys
505560
asplysmetalaglualatyrsergluileglymetlysglygluarg
65707580
argargglylysglyhisaspglyleutyrglnglyleuserthrala
859095
thrlysaspthrtyraspalaleuhismetglnalaleuproproarg
100105110
<210>9
<211>486
<212>prt
<213>人工序列(artificialsequence)
<400>9
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargproaspileglnmetthrglnthrthrserserleu
202530
seralaserleuglyaspargvalthrilesercysargalasergln
354045
aspileserlystyrleuasntrptyrglnglnlysproaspglythr
505560
vallysleuleuiletyrhisthrserargleuhisserglyvalpro
65707580
serargpheserglyserglyserglythrasptyrserleuthrile
859095
serasnleugluglngluaspilealathrtyrphecysglnglngly
100105110
asnthrleuprotyrthrpheglyglyglythrlysleugluilethr
115120125
glyglyglyglyserglyglyglyglyserglyglyglyglyserglu
130135140
vallysleuglngluserglyproglyleuvalalaproserglnser
145150155160
leuservalthrcysthrvalserglyvalserleuproasptyrgly
165170175
valsertrpileargglnproproarglysglyleuglutrpleugly
180185190
valiletrpglysergluthrthrtyrtyrasnseralaleulysser
195200205
argleuthrileilelysaspasnserlysserglnvalpheleulys
210215220
metasnserleuglnthraspaspthralailetyrtyrcysalalys
225230235240
histyrtyrtyrglyglysertyralametasptyrtrpglyglngly
245250255
thrservalthrvalserserthrthrthrproalaproargpropro
260265270
thrproalaprothrilealaserglnproleuserleuargproglu
275280285
alacysargproalaalaglyglyalavalhisthrargglyleuasp
290295300
phealacysaspiletyriletrpalaproleualaglythrcysgly
305310315320
valleuleuleuserleuvalilethrleutyrcyslysargglyarg
325330335
lyslysleuleutyrilephelysglnprophemetargprovalgln
340345350
thrthrglnglugluaspglycyssercysargpheproglugluglu
355360365
gluglyglycysgluleuargvallyspheserargseralaaspala
370375380
proalatyrglnglnglyglnasnglnleutyrasngluleuasnleu
385390395400
glyargarggluglutyraspvalleuasplysargargglyargasp
405410415
proglumetglyglylysproargarglysasnproglngluglyleu
420425430
tyrasngluleuglnlysasplysmetalaglualatyrsergluile
435440445
glymetlysglygluargargargglylysglyhisaspglyleutyr
450455460
glnglyleuserthralathrlysaspthrtyraspalaleuhismet
465470475480
glnalaleuproproarg
485
<210>10
<211>1461
<212>dna
<213>人工序列(artificialsequence)
<400>10
atggccttaccagtgaccgccttgctcctgccgctggccttgctgctccacgccgccagg60
ccggacatccagatgacacagactacatcctccctgtctgcctctctgggagacagagtc120
accatcagttgcagggcaagtcaggacattagtaaatatttaaattggtatcagcagaaa180
ccagatggaactgttaaactcctgatctaccatacatcaagattacactcaggagtccca240
tcaaggttcagtggcagtgggtctggaacagattattctctcaccattagcaacctggag300
caagaagatattgccacttacttttgccaacagggtaatacgcttccgtacacgttcgga360
ggggggactaagttggaaataacaggtggaggcggttcaggcggaggtggcagcggcggt420
ggcgggtcggaggtgaaactgcaggagtcaggacctggcctggtggcgccctcacagagc480
ctgtccgtcacatgcactgtctcaggggtctcattacccgactatggtgtaagctggatt540
cgccagcctccacgaaagggtctggagtggctgggagtaatatggggtagtgaaaccaca600
tactataattcagctctcaaatccagactgaccatcatcaaggacaactccaagagccaa660
gttttcttaaaaatgaacagtctgcaaactgatgacacagccatttactactgtgccaaa720
cattattactacggtggtagctatgctatggactactggggtcaaggaacctcagtcacc780
gtctcctcaaccacgacgccagcgccgcgaccaccaacaccggcgcccaccatcgcgtcg840
cagcccctgtccctgcgcccagaggcgtgccggccagcggcggggggcgcagtgcacacg900
agggggctggacttcgcctgtgatatctacatctgggcgcccctggccgggacttgtggg960
gtccttctcctgtcactggttatcaccctttactgcaaacggggcagaaagaagctcctg1020
tatatattcaaacaaccatttatgagaccagtacaaactactcaagaggaagatggctgt1080
agctgccgatttccagaagaagaagaaggaggatgtgaactgagagtgaagttcagcagg1140
agcgcagacgcccccgcgtaccagcagggccagaaccagctctataacgagctcaatcta1200
ggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatgggg1260
ggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataag1320
atggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcac1380
gatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatg1440
caggccctgccccctcgctga1461
<210>11
<211>495
<212>prt
<213>人工序列(artificialsequence)
<400>11
metglualaproalaglnleuleupheleuleuleuleutrpleupro
151015
aspthrthrglygluilevalleuthrglnserproalathrleuser
202530
leuserproglygluargalathrleusercysargalaserlysgly
354045
valserthrserglytyrsertyrleuhistrptyrglnglnlyspro
505560
glyglnalaproargleuleuiletyrleualasertyrleugluser
65707580
glyvalproalaargpheserglyserglyserglythraspphethr
859095
leuthrileserserleugluprogluaspphealavaltyrtyrcys
100105110
glnhisserargaspleuproleuthrpheglyglyglythrlysval
115120125
gluilelysglyglyglyglyserglyglyglyglyserglyglygly
130135140
glyserglnvalglnleuvalglnserglyvalgluvallyslyspro
145150155160
glyalaservallysvalsercyslysalaserglytyrthrphethr
165170175
asntyrtyrmettyrtrpvalargglnalaproglyglnglyleuglu
180185190
trpmetglyglyileasnproserasnglyglythrasnpheasnglu
195200205
lysphelysasnargvalthrleuthrthraspserserthrthrthr
210215220
alatyrmetgluleulysserleuglnpheaspaspthralavaltyr
225230235240
tyrcysalaargargasptyrargpheaspmetglypheasptyrtrp
245250255
glyglnglythrthrvalthrvalsersergluserlystyrglypro
260265270
procysproprocysproalaprogluphegluglyglyproserval
275280285
pheleupheproprolysprolysaspthrleumetileserargthr
290295300
progluvalthrcysvalvalvalaspvalserglngluaspproglu
305310315320
valglnpheasntrptyrvalaspglyvalgluvalhisasnalalys
325330335
thrlysproargglugluglnpheglnserthrtyrargvalvalser
340345350
valleuthrvalleuhisglnasptrpleuasnglylysglutyrlys
355360365
cyslysvalserasnlysglyleuproserserileglulysthrile
370375380
serlysalalysglyglnproarggluproglnvaltyrthrleupro
385390395400
proserglngluglumetthrlysasnglnvalserleuthrcysleu
405410415
vallysglyphetyrproseraspilealavalglutrpgluserasn
420425430
glyglnprogluasnasntyrlysthrthrproprovalleuaspser
435440445
aspglyserphepheleutyrserargleuthrvalasplysserarg
450455460
trpglngluglyasnvalphesercysservalmethisglualaleu
465470475480
hisasnhistyrthrglnlysserleuserleuserleuglylys
485490495
<210>12
<211>1488
<212>dna
<213>人工序列(artificialsequence)
<400>12
atggaagccccagctcagcttctcttcctcctgctactctggctcccagataccaccgga60
gaaattgtgttgacacagtctccagccaccctgtctttgtctccaggggaaagagccacc120
ctctcctgcagggccagcaaaggtgtcagtacatctggctatagttatttgcactggtat180
caacagaaacctggccaggctcccaggctcctcatctatcttgcatcctacctagaatct240
ggcgtcccagccaggttcagtggtagtgggtctgggacagacttcactctcaccatcagc300
agcctagagcctgaagattttgcagtttattactgtcagcacagcagggaccttccgctc360
acgttcggcggagggaccaaagtggagatcaaaggtggaggcggttcaggcggaggtggc420
agcggcggtggcgggtcgcaggtgcagctggtgcagtccggcgtggaggtgaagaagcct480
ggcgcctccgtcaaggtgtcctgtaaggcctccggctacaccttcaccaactactacatg540
tactgggtgcggcaggccccaggccagggactggagtggatgggcggcatcaacccttcc600
aacggcggcaccaacttcaacgagaagttcaagaaccgggtgaccctgaccaccgactcc660
tccaccacaaccgcctacatggaactgaagtccctgcagttcgacgacaccgccgtgtac720
tactgcgccaggcgggactaccggttcgacatgggcttcgactactggggccagggcacc780
accgtgaccgtgtcctccgagtccaaatatggtcccccatgcccaccatgcccagcacct840
gagttcgaggggggaccatcagtcttcctgttccccccaaaacccaaggacactctcatg900
atctcccggacccctgaggtcacgtgcgtggtggtggacgtgagccaggaagaccccgag960
gtccagttcaactggtacgtggatggcgtggaggtgcataatgccaagacaaagccgcgg1020
gaggagcagttccagagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggac1080
tggctgaacggcaaggagtacaagtgcaaggtctccaacaaaggcctcccgtcctccatc1140
gagaaaaccatctccaaagccaaagggcagccccgagagccacaggtgtacaccctgccc1200
ccatcccaggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttc1260
taccccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaag1320
accacgcctcccgtgctggactccgacggctccttcttcctctacagcaggctaaccgtg1380
gacaagagcaggtggcaggaggggaatgtcttctcatgctccgtgatgcatgaggctctg1440
cacaaccactacacacagaagagcctctccctgtctctgggtaaatga1488
<210>13
<211>495
<212>prt
<213>人工序列(artificialsequence)
<400>13
metglualaproalaglnleuleupheleuleuleuleutrpleupro
151015
aspthrthrglygluilevalleuthrglnserproalathrleuser
202530
leuserproglygluargalathrleusercysargalaserlysgly
354045
valserthrserglytyrsertyrleuhistrptyrglnglnlyspro
505560
glyglnalaproargleuleuiletyrleualasertyrleugluser
65707580
glyvalproalaargpheserglyserglyserglythraspphethr
859095
leuthrileserserleugluprogluaspphealavaltyrtyrcys
100105110
glnhisserargaspleuproleuthrpheglyglyglythrlysval
115120125
gluilelysglyglyglyglyserglyglyglyglyserglyglygly
130135140
glyserglnvalglnleuvalglnserglyvalgluvallyslyspro
145150155160
glyalaservallysvalsercyslysalaserglytyrthrphethr
165170175
asntyrtyrmettyrtrpvalargglnalaproglyglnglyleuglu
180185190
trpmetglyglyileasnproserasnglyglythrasnpheasnglu
195200205
lysphelysasnargvalthrleuthrthraspserserthrthrthr
210215220
alatyrmetgluleulysserleuglnpheaspaspthralavaltyr
225230235240
tyrcysalaargargasptyrargpheaspmetglypheasptyrtrp
245250255
glyglnglythrthrvalthrvalsersergluserlystyrglypro
260265270
procysproprocysproalaproglupheleuglyglyproserval
275280285
pheleupheproprolysprolysaspthrleumetileserargthr
290295300
progluvalthrcysvalvalvalaspvalserglngluaspproglu
305310315320
valglnpheasntrptyrvalaspglyvalgluvalhisasnalalys
325330335
thrlysproargglugluglnpheasnserthrtyrargvalvalser
340345350
valleuthrvalleuhisglnasptrpleuasnglylysglutyrlys
355360365
cyslysvalserasnlysglyleuproserserileglulysthrile
370375380
serlysalalysglyglnproarggluproglnvaltyrthrleupro
385390395400
proserglngluglumetthrlysasnglnvalserleuthrcysleu
405410415
vallysglyphetyrproseraspilealavalglutrpgluserasn
420425430
glyglnprogluasnasntyrlysthrthrproprovalleuaspser
435440445
aspglyserphepheleutyrserargleuthrvalasplysserarg
450455460
trpglngluglyasnvalphesercysservalmethisglualaleu
465470475480
hisasnhistyrthrglnlysserleuserleuserleuglylys
485490495
<210>14
<211>1488
<212>dna
<213>人工序列(artificialsequence)
<400>14
atggaagccccagctcagcttctcttcctcctgctactctggctcccagataccaccgga60
gaaattgtgttgacacagtctccagccaccctgtctttgtctccaggggaaagagccacc120
ctctcctgcagggccagcaaaggtgtcagtacatctggctatagttatttgcactggtat180
caacagaaacctggccaggctcccaggctcctcatctatcttgcatcctacctagaatct240
ggcgtcccagccaggttcagtggtagtgggtctgggacagacttcactctcaccatcagc300
agcctagagcctgaagattttgcagtttattactgtcagcacagcagggaccttccgctc360
acgttcggcggagggaccaaagtggagatcaaaggtggaggcggttcaggcggaggtggc420
agcggcggtggcgggtcgcaggtgcagctggtgcagtccggcgtggaggtgaagaagcct480
ggcgcctccgtcaaggtgtcctgtaaggcctccggctacaccttcaccaactactacatg540
tactgggtgcggcaggccccaggccagggactggagtggatgggcggcatcaacccttcc600
aacggcggcaccaacttcaacgagaagttcaagaaccgggtgaccctgaccaccgactcc660
tccaccacaaccgcctacatggaactgaagtccctgcagttcgacgacaccgccgtgtac720
tactgcgccaggcgggactaccggttcgacatgggcttcgactactggggccagggcacc780
accgtgaccgtgtcctccgagtccaaatatggtcccccatgcccaccatgcccagcacct840
gagttcctggggggaccatcagtcttcctgttccccccaaaacccaaggacactctcatg900
atctcccggacccctgaggtcacgtgcgtggtggtggacgtgagccaggaagaccccgag960
gtccagttcaactggtacgtggatggcgtggaggtgcataatgccaagacaaagccgcgg1020
gaggagcagttcaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggac1080
tggctgaacggcaaggagtacaagtgcaaggtctccaacaaaggcctcccgtcctccatc1140
gagaaaaccatctccaaagccaaagggcagccccgagagccacaggtgtacaccctgccc1200
ccatcccaggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttc1260
taccccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaag1320
accacgcctcccgtgctggactccgacggctccttcttcctctacagcaggctaaccgtg1380
gacaagagcaggtggcaggaggggaatgtcttctcatgctccgtgatgcatgaggctctg1440
cacaaccactacacacagaagagcctctccctgtctctgggtaaatga1488
<210>15
<211>78
<212>dna
<213>人工序列(artificialsequence)
<400>15
cgtaggaaacgaggcagcggcgccacaaacttctctctgctaaagcaagcaggtgatgtt60
gaagaaaaccccgggcct78
<210>16
<211>26
<212>prt
<213>人工序列(artificialsequence)
<400>16
argarglysargglyserglyalathrasnpheserleuleulysgln
151015
alaglyaspvalglugluasnproglypro
2025
<210>17
<211>197
<212>dna
<213>人工序列(artificialsequence)
<400>17
ccggcgggtttctgacatccggcgggtttctgacatccggcgggtttctgacatccggcg60
ggtttctgacatccggcgggtttctgacatccggcgggtttctgacatccggcgggtttc120
tgacatccggcgggtttctgacatccggcgggtttctgacatccggcgggtgactcacaa180
ccccagaaacagacata197
- 还没有人留言评论。精彩留言会获得点赞!