用于制备免疫球蛋白单可变结构域的方法与流程
本发明涉及免疫球蛋白单可变结构域的制备方法。更具体地,本发明提供了制备免疫球蛋白单可变结构域的改进方法,其中获得了增加的产率。本发明进一步提供了用于本发明方法的核酸,遗传构建体和宿主细胞以及可通过本发明的方法获得的免疫球蛋白单可变结构域。
背景技术:
:对于治疗剂应用,抗体或抗体片段必须具有非常高的产品质量。这对生物治疗剂的生产过程提出了很高的要求。这些治疗化合物的生产成本受到生产过程中遇到的困难的强烈影响。低产量或缺乏同质性将影响生产过程的经济性,并因此整体影响治疗剂的成本。已经在广泛的表达系统中报道了获得常规免疫球蛋白及其片段的足够产量的功能性产物的限制,这些表达系统包括体外翻译,大肠杆菌,酵母如例如酿酒酵母(Saccharomycescerevisiae)和巴斯德毕赤酵母(Pichiapastoris),哺乳动物细胞如例如中国仓鼠卵巢细胞和昆虫细胞中的杆状病毒系统。除此之外,抗体表达的瓶颈似乎是轻链供应不足,内质网(ER)中的不适当加工和折叠以及重链片段的细胞内积累(Lange等.2001,J.Immunol.Methods255:103;Gasser等.2006,BiotechnolBioeng.94:353;Gach等.2007,J.Biotechnol.128:735;Jenkins等.2009,Biotechnol.Appl.Biochem.53:73)。已经将蛋白质折叠和分泌途径的干预描述为用于提高重组蛋白质(如单克隆抗体(MAb))的表达和质量的不同策略之一。然而,ER分泌机构的一种或多种组分的过表达在提高生产力方面产生了混合的结果。在生物制药生产中实现始终如一的高产量仍然存在许多挑战(Jenkins等.2009,Biotechnol.Appl.Biochem.53:73)。巴斯德毕赤酵母(Pichiapastoris)已被开发为异源蛋白质生产的宿主。虽然这种宿主被称为高效表达系统,但特别是复杂蛋白质的产生已经证明具有相当低的成功率。在巴斯德毕赤酵母中,免疫球蛋白结合蛋白(BiP)的共过表达导致scFv(A33scFv)的分泌水平增加约三倍。相反,蛋白质二硫键异构酶(PDI1)的共过表达对A33scFv的分泌没有明显影响。在巴斯德毕赤酵母中共过表达BiP和PDI1没有增加A33scFv分泌,并且蛋白质水平保持与对照菌株相同(Damasceno等.2007,Appl.Microbiol.Biotechnol.74:381)。与对照菌株相比,巴斯德毕赤酵母中2F5Fab片段的产率可以提高1.2倍(在与BFR2共过表达的情况下)至2.3倍(过表达SSE1或KIN2时)(Gasser等.2007,Appl.Environ.Microbiol.73:6499)。基础亮氨酸拉链(bZIP)转录因子HAC1的过表达对巴斯德毕赤酵母中Mab2F5Fab片段分泌具有轻微影响(1.3倍),而PDI1的过表达使Fab水平增加1.9倍(Gasser等.2006,BiotechnolBioeng.94:353)。作者得出结论,轻链的充足供应和链间二硫键的形成可被视为Fab组装和随后分泌的主要速率限制因素。与用常规四链抗体或其片段(包括Fab和scFv)观察到的这些困难相反,已知免疫球蛋白单可变结构域以足够的速率和水平容易地从宿主如大肠杆菌或巴斯德毕赤酵母中表达和分泌。免疫球蛋白单可变结构域不具有链间二硫键,其特征在于由单免疫球蛋白可变结构域形成抗原结合位点,其不需要与另一结构域(例如以VH/VL相互作用的形式)相互作用用于抗原识别。例如,已经广泛描述了在原核宿主例如大肠杆菌中产生纳米抗体(Nanobody,作为免疫球蛋白单可变结构域的一个具体实例)(参见例如Ghahroudi等.1997,FEBSLetters414:521-526;Muyldermans2001,J.Biotechnol.74:277-302;Vranken等.2002,Biochemistry41:8570-8579)。Frenken等.2000(J.Biotechnol.78:11-21),WO94/25591,WO2010/125187,WO2012/056000和WO2012/152823描述了在低等真核宿主例如巴斯德毕赤酵母中产生纳米抗体。在没有任何条件优化的情况下,当在振荡培养瓶中生长的大肠杆菌中表达时,常规以5-10mg/l的水平获得重组骆驼科单可变结构域(Ghahroudi等.1997)。使用其他表达系统,甚至可以获得更高的VHH表达产量。Frenken等.2000描述了在摇瓶中每升酵母培养物中生产水平为9.3mg/l/OD660或~250mg分泌蛋白。最近,已经描述了在巴斯德毕赤酵母中表达后纳米抗体超过1g/升的产量(WO2010/139808,WO2012/152823)。WO2010/125187描述了用于在酵母(如巴斯德毕赤酵母)中产生单可变结构域的方法。WO2010/125187的方法应用促进单可变结构域中二硫键形成的条件。提出的条件之一是增强硫醇异构酶(如例如PDI1)的表达。全功能免疫球蛋白单可变结构域易于在大肠杆菌或酵母中以足够的速率和水平生产,这个事实代表了这种免疫球蛋白形式相对于常规免疫球蛋白的重要优势。发明概述尽管可以使用本领域中描述的用于表达VHH和纳米抗体的表达系统来表达它们(WO1994/25591;Ghahroudi等.1997,FEBSLetters414:521-526;Frenken等.2000,J.Biotechnol.78:11-21;Muyldermans2001,J.Biotechnol.74:277-302;Vranken等.2002,Biochemistry41:8570-8579;WO2010/125187;WO2012/056000;WO2012/152823以及埃博灵克斯股份有限公司(AblynxN.V.)的其他专利申请),发明人发现在一些情况下(例如多价VHH和纳米抗体和/或VHH和纳米抗体具有多于一个二硫键),VHH和纳米抗体的表达更难,导致表达水平和/或产量远低于预期。例如,发明人出乎意料地观察到一些治疗性VHH1型免疫球蛋白单可变结构域的产生方面的问题。在巴斯德毕赤酵母中表达这些免疫球蛋白单可变结构域后,与通常获得的VHH2型和VHH3型免疫球蛋白单可变结构域的产量相比,发明人获得了这些免疫球蛋白单可变结构域的低得多的产率。与针对免疫球蛋白单可变结构域证实的相反,某些免疫球蛋白单可变结构域(如例如,VHH1型免疫球蛋白单可变结构域)在巴斯德毕赤酵母中的表达不产生大量功能性产物。发明人已经提供了该问题的解决方案,这将在本文中进一步阐述。此外,他们观察到提出的提高免疫球蛋白单可变结构域产率的解决方案是普遍适用的,即不仅可以提高VHH1型免疫球蛋白单可变结构域的产率,还可以提高其他免疫球蛋白单可变结构域(如例如VHH2和VHH3型免疫球蛋白单可变结构域)的产率。因此,在一个方面,本发明涉及观察到某些免疫球蛋白单可变结构域在巴斯德毕赤酵母中表达后的低产量。在本发明的另一方面,提供了用于制备免疫球蛋白单可变结构域的方法,其中所获得的产物的产率增加。这些方法在本文中也称为“本发明的方法”。因此,本发明还提供了制备免疫球蛋白单可变结构域的方法,该方法克服了这一意想不到的问题。更具体地,发明人已发现(通过筛选辅助蛋白文库)在毕赤酵母宿主中增强某些辅助蛋白(PDI1(SEQIDNO:5),Kar2p(SEQIDNO:4),RPP0(SEQIDNO:6)并且特别是经剪接的HAC1(SEQIDNO:14)的表达导致免疫球蛋白单可变结构域在毕赤酵母宿主中表达时产率增加。获得的产率增加超过2倍至超过10倍(或甚至更多)。因此,在一个方面,本发明涉及增加毕赤酵母(如巴斯德毕赤酵母)中免疫球蛋白单可变结构域的表达和/或产率的方法。在本发明的方法中,表达免疫球蛋白单可变结构域,同时增强经剪接的HAC1的表达。用于本发明方法的免疫球蛋白单可变结构域可以形成多肽的一部分(也称为“本发明的多肽”),其可以包含一个或多个(即至少一个)免疫球蛋白单可变结构域或由其组成,以及其可任选地进一步包含一个或多个其他氨基酸序列(全部任选地经由一个或多个合适的接头连接)。因此,本发明提供了在毕赤酵母宿主(如巴斯德毕赤酵母)中产生包含至少一个免疫球蛋白单可变结构域或由其组成的多肽(在本文中称为“本发明的多肽”)的方法,所述方法包括在毕赤酵母宿主中表达本发明多肽并在所述毕赤酵母宿主中增强经剪接的HAC1蛋白表达的步骤。本发明的方法还可包括分离和/或纯化本发明多肽的步骤。本发明的方法特别适用于以下免疫球蛋白单可变结构域和多肽,其在标准条件下(如在本文中进一步定义的)表达时在毕赤酵母(例如巴斯德毕赤酵母)中不容易表达或以非常低的产率表达;因此,当本发明的这些免疫球蛋白单可变结构域或多肽在毕赤酵母(如巴斯德毕赤酵母)中表达后,不能获得足够量的免疫球蛋白单可变结构域和/或多肽。因此,在一个方面,本发明的免疫球蛋白单可变结构域和/或多肽选自以下免疫球蛋白单可变结构域和/或多肽,其在标准毕赤酵母表达条件(如本文进一步定义)下在毕赤酵母宿主(如巴斯德毕赤酵母)中表达后,获得0.5g/l或更低的产率,例如0.4g/L,0.3g/L,0.2g/L,0.1g/L,0.05g/L,0.01g/L,或甚至更少。另一方面,本发明的免疫球蛋白单可变结构域和/或多肽选自以下免疫球蛋白单可变结构域和/或多肽,其在毕赤酵母表达条件(如本文进一步定义)下在毕赤酵母宿主(如巴斯德毕赤酵母)中表达后,所得产率显示与编码所述免疫球蛋白单可变结构域和/或多肽的核酸的拷贝数呈负相关(如本文进一步定义)。在本发明的具体方面,本发明的方法特别适用于在毕赤酵母(如巴斯德毕赤酵母)中易于表达的本发明的免疫球蛋白单可变结构域和多肽。因此,在一个方面,本发明的免疫球蛋白单可变结构域和/或多肽选自以下免疫球蛋白单可变结构域和/或多肽,其在标准毕赤酵母表达条件(如本文进一步定义)下在毕赤酵母宿主(如巴斯德毕赤酵母)中表达后,获得0.5g/l或更高的产率,例如1g/L,2g/L,3g/L,4g/L,5g/L,6g/L,或甚至更高。在上述方法中,本发明的多肽可包含两个或更多个免疫球蛋白单可变结构域或基本上由其组成。此类多肽也称为多价多肽。因此,在一个具体方面,通过本发明的方法表达的本发明的多肽是多价多肽。备选地,通过本发明的方法表达的多肽可包含一个免疫球蛋白单可变结构域或基本上由其组成。此类多肽在本文中也称为单价多肽。因此,在另一个具体方面,通过本发明的方法表达的本发明的多肽是单价多肽。在上述方法中,免疫球蛋白单可变结构域(可能存在于本发明的多肽中)可以是(但不限于)免疫球蛋白单可变结构域,其是轻链可变结构域或重链可变结构域;更具体地是免疫球蛋白单可变结构域,其是衍生自常规四链抗体的重链可变结构域或衍生自重链抗体的重链可变结构域;特别是结构域抗体(或适合用作结构域抗体的氨基酸序列),单结构域抗体(或适合用作单结构域抗体的氨基酸序列),“dAb”(或适合用作dAb的氨基酸序列)或纳米抗体(包括但不限于VHH序列),优选纳米抗体。在优选的方面,在本发明的方法中表达的纳米抗体是VHH序列(VHH),(部分)人源化VHH序列(人源化VHH),骆驼化重链可变结构域(骆驼化VH)或纳米抗体,其已经通过亲和力成熟等技术获得。在本发明的另一个具体方面,在本发明的方法中表达的免疫球蛋白单可变结构域(可能存在于本发明的多肽中)包含至少两个二硫键。在另一个具体方面,免疫球蛋白单可变结构域属于VHH1型免疫球蛋白单可变结构域的组。另一方面,免疫球蛋白单可变结构域(可能存在于本发明的多肽中)不属于VHH1型免疫球蛋白单可变结构域的组,但属于免疫球蛋白单可变结构域的另一个组,如VHH2,VHH3或任何其他类型的免疫球蛋白单可变结构域。在另一个具体方面,在本发明的方法中表达的免疫球蛋白单可变结构域(可能存在于本发明的多肽中)(如例如,WO2012/042026和WO2013/045707中描述的免疫球蛋白单可变结构域)特异性结合c-Met。因此,包含至少一个特异性结合c-Met的免疫球蛋白单可变结构域的本发明多肽(其中所述免疫球蛋白单可变结构域包含两个二硫键)形成本发明的特定但非限制性的方面。用于本发明方法的特定纳米抗体是SEQIDNO:49。在一个优选的方面,在本发明的方法中表达的一个或多个免疫球蛋白单可变结构域(可能存在于本发明的多肽中)(如例如埃博灵克斯股份有限公司的美国临时申请US62/254,375中描述的免疫球蛋白单可变结构域(也参见PCT/EP2016/077595))特异性结合TNF。因此,包含一个或多个(至少一个)特异性结合TNF的免疫球蛋白单可变结构域的本发明多肽形成本发明的特定和非限制性方面。在一个具体方面,在本发明的方法中表达的本发明的多肽包含一个免疫球蛋白单可变结构域或基本上由其组成。此类多肽在本文中也称为单价多肽。因此,在一个优选的方面,在本发明的方法中,多肽是单价多肽。这种包含一个免疫球蛋白单可变结构域或基本上由其组成的单价多肽可包含至少两个二硫键。因此,特异性结合TNF的免疫球蛋白单可变结构域属于VHH1型免疫球蛋白单可变结构域。备选地,免疫球蛋白单可变结构域包含一个二硫键。因此,在优选的方面,特异性结合TNF的免疫球蛋白单可变结构域不属于VHH1型免疫球蛋白单可变结构域的组,但属于免疫球蛋白单可变结构域的另一个组,如VHH2,VHH3或任何其他类型的免疫球蛋白单可变结构域。在优选的方面,在本发明的方法中,免疫球蛋白单可变结构域基本上由4个框架区(分别为FR1至FR4)和3个互补决定区(分别为CDR1至CDR3)组成,其中CDR1为SEQIDNO:58,CDR2是SEQIDNO:60,CDR3是SEQIDNO:62。用于本发明方法的特定纳米抗体选自由SEQIDNO:55和56组成的组。包含一个或多个免疫球蛋白单可变结构域或基本上由其组成的本发明多肽可以进一步包含任选地经由一个或多个肽接头连接的一个或多个其他残基或结合单元。在一个方面,所述一个或多个其他残基可有效预防或减少血清中已存在的抗体(所谓的“已存在抗体”)与本发明多肽的结合(如本文进一步描述的)。在另一个方面,所述一个或多个其他残基或结合单元还可以选自由以下各项组成的组:免疫球蛋白单可变结构域,结构域抗体,适合用作结构域抗体的氨基酸序列,单结构域抗体,适合用作单结构域抗体的氨基酸序列,“dAb”,适合用作dAb的氨基酸序列,或纳米抗体。包含两个或更多个结合单元或基本上由其组成的多肽也称为多价构建体。在本发明的具体方面,包含本发明方法中表达的一个或多个免疫球蛋白单可变结构域或基本上由其组成的多肽是多价构建体。在另一个具体方面,本发明的多肽是二价,三价或四价多肽。在本发明的特定方面,与不含所述一个或多个其他结合单元的多肽相比,所述一个或多个其他结合单元可以为本发明的多肽提供增加的半衰期。不受限制地,为多肽提供增加的半衰期的所述一个或多个其他结合单元可以选自由以下各项组成的组:可以结合血清白蛋白(如人血清白蛋白)或血清免疫球蛋白(如IgG)的结合单元。在本发明的方法中,可以通过将一个或多个编码经剪接的HAC1蛋白的核酸引入毕赤酵母宿主中来增强经剪接的HAC1的表达。另一方面,可以通过将一个或多个控制编码经剪接的HAC1蛋白的核酸表达的强启动子引入毕赤酵母宿主中来增强经剪接的HAC1蛋白的表达。包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽和经剪接的HAC1蛋白可以从相同的遗传构建体表达。在本发明的该特定方面,编码包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽的核酸的转录和编码经剪接的HAC1蛋白的核酸的转录可以由相同的启动子或不同的启动子控制。编码经剪接的HAC1蛋白的核酸可以位于遗传构建体上在编码包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽的核酸的下游。备选地,编码包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽的核酸可以位于遗传构建体上在编码经剪接的HAC1蛋白的核酸的下游。在本发明的另一方面,包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽和经剪接的HAC1蛋白可以从不同的遗传构建体表达。在本发明的该特定方面,编码包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽的核酸和编码经剪接的HAC1蛋白的核酸从不同遗传构建体的转录可以由可以是相同或不同的两个单独的启动子来控制。包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽和/或经剪接的HAC1蛋白还能够从染色体表达。在该特定方面,可以通过将强启动子引入毕赤酵母宿主的染色体来增强经剪接的HAC1蛋白的表达。备选地,可以将由强启动子控制的编码经剪接的HAC1蛋白的一个或多个核酸引入毕赤酵母宿主的染色体中。还可以将编码包含至少一个免疫球蛋白单可变结构域或基本上由其组成的多肽的一个或多个核酸引入毕赤酵母宿主的染色体中。在本发明的一个方面,编码包含免疫球蛋白单可变结构域或基本上由其组成的多肽的核酸的数目是一个。在本发明的另一方面,编码包含免疫球蛋白单可变结构域或基本上由其组成的多肽的核酸的数目是两个或更多个。另一方面,本发明还涉及编码本发明多肽(或其合适片段)并且编码经剪接的HAC1蛋白的核酸。这种核酸在本文中也称为“本发明的核酸”,并且可以例如是如本文进一步描述的遗传构建体的形式。因此,本发明还涉及作为遗传构建体形式的本发明的核酸。这种遗传构建体在本文中也称为“本发明的遗传构建体”,因此包含编码本发明多肽的核酸和编码经剪接的HAC1蛋白的核酸。在一个具体方面,在本发明的这种遗传构建体中,编码经剪接的HAC1蛋白的核酸位于编码本发明多肽的核酸的下游。在另一个具体方面,在本发明的这种遗传构建体中,编码本发明多肽的核酸位于编码经剪接的HAC1蛋白的核酸的下游。在本发明的遗传构建体中,编码本发明多肽的核酸的表达和编码经剪接的HAC1蛋白的核酸的表达可以由相同的启动子或不同的启动子控制。启动子可以是组成型或诱导型启动子。在本发明的一个方面,编码本发明多肽的核酸的拷贝数是一个。在本发明的另一个方面,编码本发明多肽的核酸的拷贝数是两个或更多个。在本发明的一个方面,辅助蛋白的数目是一个,其中辅助蛋白是经剪接的HAC1。在本发明的另一方面,一个或多个另外的辅助蛋白的表达被增强。在本发明的另一个方面,另外的辅助蛋白选自PDI1,Kar2p和RPPO。在本发明的另一个方面,辅助蛋白的数目是两个。在本发明的另一方面,辅助蛋白的数量多于两个,例如三个或更多个。在另一方面,两个或更多个辅助蛋白选自以下辅助蛋白的组合:-PDI1和经剪接的HAC1;-Kar2p和经剪接的HAC1;-RPP0和经剪接的HAC1;-PDI1,Kar2p和经剪接的HAC1;-PDI1,RPP0和经剪接的HAC1;-Kar2p,RPP0和经剪接的HAC1;和-PDI1,Kar2p,RPP0和经剪接的HAC1.在一个优选的方面,以下辅助蛋白的表达被增强:-PDI1,Kar2p和经剪接的HAC1;或-Kar2p,RPP0和经剪接的HAC1。在本发明的一个优选方面,辅助蛋白是经剪接的HAC1(SEQIDNO:14)。在另一个优选的方面,另外的辅助蛋白选自PDI1(SEQIDNO:5),Kar2p(SEQIDNO:4),RPP0(SEQIDNO:6)和经剪接的HAC1(SEQIDNO:14)。在另一优选方面,两个或更多个辅助蛋白选自以下辅助蛋白的组合:-PDI1(SEQIDNO:5)和经剪接的HAC1(SEQIDNO:14);-Kar2p(SEQIDNO:4)和经剪接的HAC1(SEQIDNO:14);-RPP0(SEQIDNO:6)和经剪接的HAC1(SEQIDNO:14);-PDI1(SEQIDNO:5),Kar2p(SEQIDNO:4)和经剪接的HAC1(SEQIDNO:14);-PDI1(SEQIDNO:5),RPP0(SEQIDNO:6)和经剪接的HAC1(SEQIDNO:14);-Kar2p(SEQIDNO:4),RPP0(SEQIDNO:6)和经剪接的HAC1(SEQIDNO:14);和-PDI1(SEQIDNO:5),Kar2p(SEQIDNO:4),RPP0(SEQIDNO:6)和经剪接的HAC1(SEQIDNO:14).另一方面,本发明涉及在毕赤酵母宿主(在本文中也称为“本发明的毕赤酵母宿主”)中引入本发明的核酸和遗传构建体。除了毕赤酵母宿主的质粒或载体转化外,毕赤酵母宿主的染色体转化也包括在本发明中。可以在毕赤酵母宿主的染色体上引入强(诱导型)启动子(而不是天然辅助蛋白的天然启动子);或者可以将另一个(强)启动子控制下的辅助蛋白基因序列的另一个拷贝引入染色体中。另一方面,本发明涉及表达(或在适当情况下能够表达)本发明多肽并且其中经剪接的HAC1的表达被增强的毕赤酵母宿主;和/或含有本发明的核酸和/或本发明的遗传构建体的毕赤酵母宿主。在一个优选的方面,毕赤酵母宿主是巴斯德毕赤酵母(Pichiapastoris)。在另一个优选的方面,巴斯德毕赤酵母菌株选自巴斯德毕赤酵母X33和巴斯德毕赤酵母NRRLY-11430。本发明进一步涉及制备本发明的核酸,本发明的遗传构建体和本发明的毕赤酵母宿主的方法,以及涉及本发明的所述核酸,本发明的遗传构建体和本发明的毕赤酵母宿主用于产生免疫球蛋白单可变结构域和包含其的多肽的用途。本发明进一步涉及通过本文所述的任何方法能够获得的多肽和/或免疫球蛋白单可变结构域,包含此类多肽和/或免疫球蛋白单可变结构域的药物组合物和其他组合物,以及所述多肽和/或免疫球蛋白单可变结构域的治疗用途或包括使用所述多肽和/或免疫球蛋白单可变结构域的治疗方法。附图说明图1:纳米抗体A在如实施例1.2中所述产生的两个不同的毕赤酵母克隆中表达。一个毕赤酵母克隆在基因组中含有1个拷贝的纳米抗体A表达盒。第二个克隆含有插入基因组中的一个以上拷贝的纳米抗体A表达盒。在SDS-PAGE凝胶上比较来自不同克隆的等体积的上清液。进行密度测定分析,用于对应于完整纳米抗体产物的条带的相对定量。使用Imagequant软件(GEHealthcare)进行条带量的定量。观察到拷贝数和产率之间的负相关。图2:测试用纳米抗体A和辅助蛋白文库转化的克隆的提高的纳米抗体A的表达水平。在SDS-PAGE凝胶上比较来自不同克隆的等体积的上清液。进行密度测定分析,用于对应于完整纳米抗体产物的条带的相对定量。使用Imagequant软件(GEHealthcare)进行条带量的定量。发现4个克隆(6H1,4C2,5A6,9C4)与没有辅助蛋白的相应参比克隆(RefCN=1和RefCN>1)相比,在摇瓶中分泌显著更高水平的纳米抗体A。Ref:具有插入基因组中的纳米抗体A表达盒的指定拷贝数的参比克隆。图3:用辅助蛋白Kar2p,RPP0,Hac1剪接变体或PDI1之一转化的具有多于1个拷贝的纳米抗体A表达盒的参比克隆的摇瓶表达。在SDS-PAGE上分析纳米抗体产率,并与参比克隆(不含辅助蛋白)的纳米抗体产率进行比较。进行密度测定分析,用于对应于完整纳米抗体产物的条带的相对定量。使用Imagequant软件(GEHealthcare)进行条带量的定量。发明详述定义除非另有说明或另外定义,否则所使用的所有术语具有其在本领域中的通常含义,这对于本领域技术人员来说是清楚的。例如,参考标准手册,例如Sambrook等.1989(MolecularCloning:ALaboratoryManual,2ndEd.,Vols.1-3,ColdSpringHarborLaboratoryPress),Ausubel等.1987(Currentprotocolsinmolecularbiology,GreenPublishingandWileyInterscience,NewYork),Lewin1985(GenesII,JohnWiley&Sons,NewYork,N.Y.),Old等.1981(PrinciplesofGeneManipulation:AnIntroductiontoGeneticEngineering,2ndEd.,UniversityofCaliforniaPress,Berkeley,CA),Roitt等.2001(Immunology,6thEd.,Mosby/Elsevier,Edinburgh),Roitt等.2001(Roitt’sEssentialImmunology,10thEd.,BlackwellPublishing,UK),以及Janeway等.2005(Immunobiology,6thEd.,GarlandSciencePublishing/ChurchillLivingstone,NewYork),以及这里引用的一般
背景技术:
。除非另外指出,否则本领域技术人员将会清楚,没有详细描述的所有方法、步骤、技术和操作都可以以本身已知的方式执行并且已经被执行。再次参考例如本文提及的标准手册和一般
背景技术:
以及其中引用的其他参考文献;以及例如以下综述Presta2006(Adv.DrugDeliv.Rev.58:640),Levin和Weiss2006(Mol.Biosyst.2:49),Irving等.2001(J.Immunol.Methods248:31),Schmitz等.2000(Placenta21Suppl.A:S106),Gonzales等.2005(TumourBiol.26:31),其描述了用于蛋白质工程的技术,例如亲和力成熟和用于改善蛋白质如免疫球蛋白的特异性和其他所需性质的其他技术。当核苷酸序列或氨基酸序列分别被称为“包含”另一核苷酸序列或氨基酸序列或“基本上由另一核苷酸序列或氨基酸序列组成”时,这可意味着后提及的核苷酸序列或氨基酸序列序列分别已经被并入到先提及的核苷酸序列或氨基酸序列中,但更通常地这通常意味着先提及的核苷酸序列或氨基酸序列分别在其序列内包含一段具有与后提及的序列分别相同的核苷酸序列或氨基酸序列的核苷酸或氨基酸残基,而不管实际上如何生成或获得先提及的序列(其可以例如通过本文所述的任何合适的方法)。通过非限制性实例,当说本发明的多肽包含免疫球蛋白单可变结构域时,这可意味着所述免疫球蛋白单可变结构域序列已被并入本发明多肽的序列中,但更通常地这通常意味着本发明的多肽在其序列内包含免疫球蛋白单可变结构域的序列,而不管本发明的所述多肽是如何产生或获得的。而且,当核酸或核苷酸序列被认为包含另一核苷酸序列时,先提及的核酸或核苷酸序列优选是这样的,即当其表达产物(例如多肽)表达时,由后提及的核苷酸序列编码的氨基酸序列形成所述表达产物的一部分(换句话说,后提及的核苷酸序列与先提及的更大的核酸或核苷酸序列在相同的阅读框中)。“基本上由...组成”是指本发明方法中使用的免疫球蛋白单可变结构域与本发明的多肽完全相同或者对应于本发明的具有有限数量的氨基酸残基的多肽,如在免疫球蛋白单可变结构域的氨基端、羧基端或氨基端和羧基端两处添加的1-20个氨基酸残基,例如1-10个氨基酸残基,优选1-6个氨基酸残基,如1、2、3、4、5或6个氨基酸残基。例如与已经从中获得其的反应介质或培养介质相比,当核酸或氨基酸与至少一种与其通常在所述来源或介质中结合的其它组分(例如另一种核酸,另一种蛋白质/多肽,另一种生物组分或大分子或至少一种污染物、杂质或次要组分)已经分离时,所述核酸或氨基酸被认为是“(处于)(基本上)分离的(形式)”。具体而言,当核酸或氨基酸已经被纯化至少2倍,特别是至少10倍,更特别是至少100倍和多达1000倍或更多时,被认为是“(基本上)分离的”。“处于(基本上)分离的形式”的核酸或氨基酸优选基本上是均相的,如使用合适的技术如合适的色谱技术如聚丙烯酰胺凝胶电泳测定的。术语多肽“多肽(的)表达”或“表达多肽”(所述多肽是例如本发明的免疫球蛋白单可变结构域和/或多肽或辅助蛋白)是将来自基因的信息用于合成功能基因产物(即本发明的免疫球蛋白单可变结构域和/或多肽或辅助蛋白)的过程。当辅助蛋白或多肽如本发明的免疫球蛋白单可变结构域和/或多肽被称为从核酸,遗传构建体或染色体“表达”时,其通过将来自核酸,遗传构建体或染色体的信息用于合成功能基因产物(即本发明的免疫球蛋白单可变结构域和/或多肽或辅助蛋白)的过程合成。当蛋白质被称为“共表达”时,所述蛋白质同时表达。“增强基因的表达”意味着与不增强基因的表达的基因产物产生相比,基因产物(例如辅助蛋白)的产生增加。如进一步说明的,可以通过各种方法增强特定基因的表达,所述方法包括使用合适的控制序列,例如强启动子,和/或增加基因剂量,例如通过增加相应基因的拷贝数。本发明中使用的术语“产率”涉及在毕赤酵母宿主中表达后以功能形式产生的本发明的免疫球蛋白单可变结构域和/或多肽的量。产率表示为g(克)的本发明的免疫球蛋白单可变结构域和/或多肽/L(升)培养基。免疫球蛋白单可变结构域除非另有说明,否则术语“免疫球蛋白序列”-无论在本文中是指重链抗体还是常规的4链抗体-均用作通用术语,以包括全长抗体、其各个链以及其所有部分、结构域或片段(包括但不限于抗原结合结构域或片段,例如分别为VHH结构域或VH/VL结构域)。此外,如本文中使用的术语“序列”(例如在术语如“免疫球蛋白序列”,“抗体序列”,“可变结构域序列”,“VHH序列”或“蛋白质序列”中)通常应该被理解为包括相关氨基酸序列以及编码其的核酸或核苷酸序列,除非上下文需要更有限的解释。与“单可变结构域”可互换使用的术语“免疫球蛋白单可变结构域”定义了这样的分子,其中抗原结合位点存在于单个免疫球蛋白结构域上并由其形成。这使免疫球蛋白单可变结构域区别于“常规”免疫球蛋白或其片段(如Fab,scFv等)(其中两个免疫球蛋白结构域,特别是两个可变结构域相互作用形成抗原结合位点)。通常,在常规免疫球蛋白中,重链可变结构域(VH)和轻链可变结构域(VL)相互作用形成抗原结合位点。在这种情况下,VH和VL两者的互补决定区(CDR)都将有助于抗原结合位点,即总共6个CDR将参与抗原结合位点的形成。相反,免疫球蛋白单可变结构域的结合位点由单个VH或VL结构域形成。因此,免疫球蛋白单可变结构域的抗原结合位点由不超过三个CDR形成。因此,术语“免疫球蛋白单可变结构域”和“单可变结构域”不包含常规免疫球蛋白或其片段(其需要至少两个可变结构域的相互作用以形成抗原结合位点)。然而,这些术语确实包含以下常规免疫球蛋白的片段,其中抗原结合位点由单个可变结构域形成。通常,单可变结构域是基本上由4个框架区(分别为FR1至FR4)和3个互补决定区(分别为CDR1至CDR3)组成的氨基酸;或者是这类氨基酸的任何合适的片段(其通常含有形成这些CDR中的至少一个的氨基酸残基中的至少一些)。这种单可变结构域和片段最优选使得它们包含免疫球蛋白折叠或能够在合适的条件下形成免疫球蛋白折叠。如此,单可变结构域可以例如包含轻链可变结构域序列(例如VL序列)或其合适的片段;或重链可变结构域序列(例如,VH序列或VHH序列)或其合适的片段;只要其能够形成单抗原结合单元(即基本上由单可变结构域组成的功能性抗原结合单元,使得单抗原结合结构域不需要例如像以下可变结构域的情况那样与另一个可变结构域相互作用形成功能性抗原结合单元,所述可变结构域存在于例如常规抗体和scFv片段中,需要与另一可变结构域相互作用-例如通过VH/VL相互作用-以形成功能性抗原结合结构域)。在本发明的一个实施方案中,免疫球蛋白单可变结构域是轻链可变结构域序列(例如VL序列)或重链可变结构域序列(例如VH序列);更具体而言,免疫球蛋白单可变结构域可以是衍生自常规四链抗体的重链可变结构域序列或是衍生自重链抗体的重链可变结构域序列。例如,单可变结构域或免疫球蛋白单可变结构域(或适合用作免疫球蛋白单可变结构域的氨基酸)可以是(单)结构域抗体(或适合用作(单)结构域抗体的氨基酸),“dAb”或dAb(或适合用作dAb的氨基酸)或纳米抗体(如本文所定义,并且包括但不限于VHH);其他单个可变结构域或其任何一个的任何合适的片段。对于(单)域抗体的一般描述,还参考本文引用的现有技术,以及EP0368684。对于术语“dAb”,例如参考Ward等.1989(Nature341:544),参考Holt等.2003(TrendsBiotechnol.21:484);以及例如WO04/068820,WO06/030220,WO06/003388和DomantisLtd.的其他公开的专利申请。还应该注意的是,单可变结构域可以来自某些鲨鱼物种(例如所谓的“IgNAR结构域”,参见例如WO05/18629),尽管这在本发明的上下文中较不优选,因为它们不是哺乳动物来源的。特别地,免疫球蛋白单可变结构域可以是纳米抗体(如本文所定义的)或其合适的片段。[注意:纳米抗体(Nanobody,Nanobodies)和纳米克隆(Nanoclone)是埃博灵克斯股份有限公司的注册商标]对于纳米抗体的一般性描述,参考本文引用的现有技术,例如在WO08/020079(第16页)中描述的。免疫球蛋白序列特别是纳米抗体的氨基酸序列和结构可以被认为-但不限于-由在本领域和在下文中分别称为“框架区1”或“FR1”;“框架区2”或“FR2”;“框架区3”或“FR3”;“框架区4”或“FR4”的四个“框架区”或“FR”组成;这些框架区由在本领域和在下文中分别称为“互补决定区1”或“CDR1”;“互补决定区2”或“CDR2”;和“互补决定区3”或“CDR3”的三个“互补决定区”或“CDR”中断。纳米抗体中氨基酸残基的总数可以在110-120的范围内,优选112-115,最优选113。然而,应当注意,对纳米抗体的部分,片段,类似物或衍生物(如本文进一步描述的)的长度和/或大小没有特别限制,只要这些部分,片段,类似物或衍生物满足本文概述的进一步的要求并且还优选适用于本文所述的目的即可。关于VHH和纳米抗体的进一步描述,参考Muyldermans2001的综述文章(Rev.Mol.Biotechnol.74:277)以及以下作为一般
背景技术:
提及的专利申请:VrijeUniversiteitBrussel的WO94/04678、WO95/04079和WO96/34103;Unilever的WO94/25591、WO99/37681、WO00/40968、WO00/43507、WO00/65057、WO01/40310、WO01/44301、EP1134231和WO02/48193;VlaamsInstituutvoorBiotechnologie(VIB)的WO97/49805、WO01/21817、WO03/035694、WO03/054016和WO03/055527;AlgonomicsN.V.和埃博灵克斯股份有限公司的WO03/050531;加拿大国家研究委员会(NationalResearchCouncilofCanada)的WO01/90190;抗体研究所(InstituteofAntibodies)的WO03/025020(=EP1433793)以及埃博灵克斯股份有限公司的WO04/041867、WO04/041862、WO04/041865、WO04/041863、WO04/062551、WO05/044858、WO06/40153、WO06/079372、WO06/122786、WO06/122787和WO06/122825以及埃博灵克斯股份有限公司的进一步公开的专利申请。还参考了这些申请中提到的另外的现有技术,特别是国际申请WO06/040153的第41-43页上提及的参考文献列表,该列表和参考文献通过引用并入本文。如在这些参考文献中所描述的,纳米抗体(特别是VHH和部分人源化的纳米抗体)特别可以通过在一个或多个框架序列中存在一个或多个“标志残基”来表征。纳米抗体的进一步描述,包括纳米抗体的人源化和/或骆驼化,以及其他修饰、部分或片段、衍生物或“纳米抗体融合物”、多价构建体(包括接头序列的一些非限制性实例)和增加纳米抗体的半衰期的不同修饰以及它们的制备可以在WO08/101985和WO08/142164中找到。因此,在本发明的含义中,术语“免疫球蛋白单可变结构域”或“单可变结构域”包含衍生自非人来源的多肽,优选骆驼科,优选骆驼科重链抗体。如前所述,它们可以是人源化的。而且,该术语包括来源于非骆驼科动物来源(例如小鼠或人)的多肽,其已经被“骆驼化”,如描述于Davies和Riechmann1994(FEBS339:285),1995(Biotechonol.13:475)和1996(Prot.Eng.9:531)和Riechmann和Muyldermans1999(J.Immunol.Methods231:25)中的。术语“免疫球蛋白单可变结构域”包括不同来源的免疫球蛋白序列,包括小鼠,大鼠,兔,驴,人和骆驼科动物免疫球蛋白序列。它还包括完全人,人源化或嵌合免疫球蛋白序列。例如,它包括骆驼科动物免疫球蛋白序列和人源化骆驼科动物免疫球蛋白序列,或骆驼化免疫球蛋白单可变结构域,例如Ward等(参见例如WO94/04678和Davies和Riechmann1994,1995和1996)所述的骆驼化dAb。本发明可用于表达或产生本文所述的任何免疫球蛋白单可变结构域。本发明还可用于表达或产生本发明的多肽(即包含这种免疫球蛋白单可变结构域或基本上由其组成的多肽)。在特定方面,本发明可用于表达或产生包含两个二硫键的免疫球蛋白单可变结构域。本发明还可用于表达或产生具有两个二硫键的本发明的多肽(即包含这种免疫球蛋白单可变结构域或基本上由其组成的多肽)。本发明还可用于表达或产生本发明的多肽(即具有两个二硫键的包含这种免疫球蛋白单可变结构域或基本上由其组成的多肽)。已知所有VHH在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间含有至少一个二硫键(根据Kabat编号,参见埃博灵克斯股份有限公司的专利申请和Muyldermans和Lauwereys1999,J.Mol.Recognit.12:131)。虽然大多数VHH仅含有这种单一的二硫键,但也知道一些VHH含有总共两个(或在特殊情况下为三个)二硫键。例如,被称为“VHH-1型”,“VHH-1类”等(如本文进一步定义的)的一类VHH(和纳米抗体)通常在CDR2中的第50位半胱氨酸残基与CDR3中(或在特殊情况下在CDR1或CDR2中)存在的半胱氨酸残基之间具有第二个二硫键。此外,来自骆驼或单峰骆驼的一些VHH通常在CDR1中存在的半胱氨酸残基(或FR2中的第45位)与CDR3中存在的半胱氨酸残基之间具有二硫键(Vu等.1997,Mol.Immunol.34:1121;Muyldermans和Lauwereys1999)。来自美洲驼的一些VHH有时在CDR1中存在的半胱氨酸残基(例如在第33位)和CDR3中存在的半胱氨酸残基之间具有二硫键(Vu等,1997)。在一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在框架区之一中的半胱氨酸残基与CDR区之一中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在框架区之一中的半胱氨酸残基与CDR3中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在框架区二(FR2)中的半胱氨酸残基与CDR3中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在框架区二(FR2)中第45位的半胱氨酸残基与CDR3中的半胱氨酸残基之间形成的二硫键(如在一些衍生自骆驼和单峰骆驼的VHH中)。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在一个CDR中的半胱氨酸残基与另一个CDR中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR3中的半胱氨酸残基与另一个CDR(特别是CDR1)中的半胱氨酸残基之间形成的二硫键(如在一些衍生自骆驼,单峰骆驼和美洲驼的VHH中)。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR1中的半胱氨酸残基与另一个CDR中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR1中的半胱氨酸残基与CDR1中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR1中的半胱氨酸残基与CDR3中的半胱氨酸残基之间形成的二硫键(如在一些衍生自骆驼,单峰骆驼和美洲驼的VHH中)。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在第33位的半胱氨酸与CDR3中的半胱氨酸残基之间形成的二硫键(如在一些衍生自骆驼,单峰骆驼和美洲驼的VHH中)。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR2中的半胱氨酸残基与另一个CDR中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR2中的半胱氨酸残基与CDR2中的半胱氨酸残基之间形成的二硫键。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在CDR2中的半胱氨酸残基与CDR3中的半胱氨酸残基之间形成的二硫键(如在一些衍生美洲驼的VHH中)。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在第50位的半胱氨酸残基与在另一个CDR(例如CDR1,CDR2或CDR3)中的半胱氨酸残基之间形成的二硫键(如在VHH和VHH1型的纳米抗体中)。在另一个具体但非限制性的方面,免疫球蛋白单可变结构域包含在第22位的半胱氨酸残基和第92位的半胱氨酸残基之间的二硫键,并且还包含在第50位的半胱氨酸残基与CDR3中的半胱氨酸残基之间形成的二硫键(如在VHH和VHH1型的纳米抗体中)。在优选但非限制性的实施方案中,免疫球蛋白单可变结构域可以是“VHH1型免疫球蛋白单可变结构域”。如果VHH1型免疫球蛋白单可变结构域或VHH1型序列与VHH1共有序列(SEQIDNO:46,QVQLVESGGGLVQPGGSLRLSCAASGFTLDYYAIGWFRQAPGKEREGVSCISSSDGSTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDTAVYYCAA)具有85%同一性(使用具有标准设置的blastp算法,即blosom62评分矩阵)并且强制性地在第50位具有半胱氨酸,即Cys50(使用Kabat编号),则氨基酸如例如本发明的免疫球蛋白单可变结构域或多肽被称为所述“VHH1型免疫球蛋白单可变结构域”或“VHH1型序列”。这些VHH1型免疫球蛋白单可变结构域通常具有(或能够形成)Cys50和CDR3(或在特殊情况下CDR1或CDR2)中的半胱氨酸残基之间的二硫键。如果VHH2型免疫球蛋白单可变结构域或VHH2型序列与VHH2共有序列(SEQIDNO:47,QVQLVESGGGLVQAGGSLRLSCAASGSIFSINAMGWYRQAPGKQRELVAAITSGGSTNYADSVKGRFTISRDNAKNTVYLQMNSLKPEDTAVYYCNA)具有85%同一性(使用具有标准设置的blastp算法,即blosom62评分矩阵),则氨基酸序列如例如本发明的免疫球蛋白单可变结构域或多肽被称为所述“VHH2型免疫球蛋白单可变结构域”或“VHH2型序列”。如果VHH3型免疫球蛋白单可变结构域或VHH3型序列与VHH3共有序列(SEQIDNO:48,QVQLVESGGGLVQAGGSLRLSCAASGRTFSSYAMGWFRQAPGKEREFVAAISWSGGSTYYADSVKGRFTISRDNAKNTVYLQMNSLKPEDTAVYYCAA)具有85%同一性(使用具有标准设置的blastp算法,即blosom62评分矩阵),则氨基酸序列如例如本发明的免疫球蛋白单可变结构域或多肽被称为所述“VHH3型免疫球蛋白单可变结构域”或“VHH3型序列”。尽管本发明特别适用于表达VHH1(其中存在两个二硫键非常常见),但应注意它也可用于表达VHH2或VHH3(其可能包含也可能不包含两个二硫键,虽然这不太常见)。对于针对c-Met并且可以使用本文描述的方法表达/产生的纳米抗体(和包含其的多肽)的一般描述和一些非限制性实例,参考WO2012/042026和WO2013/045707。对于针对TNF并且可以使用本文描述的方法表达/产生的纳米抗体(和包含其的多肽)的一般描述和一些非限制性实例,参考埃博灵克斯股份有限公司的美国临时申请US62/254,375(也参见PCT/EP2016/077595)。发明人还期望该教导不仅特别适用于具有两个或更多个二硫键的VHH和纳米抗体(例如VHH-1),而且还特别适用于包含两个或更多个二硫键的其他免疫球蛋白单可变结构域(如(单)结构域抗体,dAb,来自鲨鱼的IgNAR结构域等)。本发明的多肽在本发明方法中制备的免疫球蛋白单可变结构域可以形成蛋白质或多肽(也称为“本发明的多肽”)的一部分,其可以包含一个或多个(至少一个)免疫球蛋白单可变结构域或由其组成,以及其可任选地进一步包含一个或多个其他氨基酸序列(全部任选地经由一个或多个合适的接头连接)。术语“免疫球蛋白单可变结构域”也可包括本发明的此类多肽。一个或多个免疫球蛋白单可变结构域可用作这种蛋白质或多肽中的结合单元,所述蛋白质或多肽可任选地含有一个或多个可用作结合单元的其他氨基酸,以分别提供单价,多价或多特异性的本发明的多肽(对于含有一个或多个VHH结构域的多价和多特异性多肽及其制备,也参考Conrath等.2001(J.Biol.Chem.276:7346)以及例如WO96/34103,WO99/23221和WO2010/115998)。本发明的多肽可包含一个如上所述的免疫球蛋白单可变结构域或基本上由其组成。此类多肽在本文中也称为单价多肽。如上所述,本发明的多肽还可以包括含有两个或多个如上所述的单可变结构域形式的抗原结合单元的构建体。例如,可以连接具有相同或不同抗原特异性的两个(或更多个)免疫球蛋白单可变结构域以形成例如二价,三价或多价构建体。通过组合具有两种或更多种特异性的免疫球蛋白单可变结构域,可以形成双特异性、三特异性等构建体。例如,根据本发明的免疫球蛋白单可变结构域可包含针对相同靶标的两个或三个免疫球蛋白单可变结构域,或针对靶标A的一个或两个免疫球蛋白单可变结构域,和针对靶标B的一个免疫球蛋白单可变结构域。本领域技术人员可以容易想到的这样的构建体和其修饰都涵盖在在本文中使用的术语本发明的多肽中。此外,在本发明的方法中也制备了融合的免疫球蛋白序列,其包含标签或其他功能部分,例如毒素,标记,放射化学品等。在另一个方面,包含一个或多个免疫球蛋白单可变结构域(或其合适片段)或基本上由其组成的本发明多肽可以进一步包含一个或多个其他基团,残基,部分或结合单元。此类其他基团,残基,部分,结合单元或氨基酸序列可以为或可以不为免疫球蛋白单可变结构域(和/或其存在于其中的多肽)提供进一步的功能,并且可以改变或可以不改变免疫球蛋白单可变结构域的性质。例如,此类其他基团,残基,部分或结合单元可以是一个或多个另外的氨基酸,使得化合物,构建体或多肽是(融合)蛋白质或(融合)多肽。在一个优选但非限制性的方面,所述一个或多个其他基团、残基、部分或结合单元是免疫球蛋白。甚至更优选地,所述一个或多个其它基团、残基、部分或结合单元选自由以下各项组成的组:结构域抗体、适合用作结构域抗体的氨基酸、单结构域抗体、适合用作单结构域抗体的氨基酸、“dAb”、适合用作dAb的氨基酸,或纳米抗体。备选地,这种基团、残基、部分或结合单元可以例如是化学基团、残基、部分,其自身可以是或可以不是生物学上和/或药理学上活性的。例如但不限于,这种基团可以与一个或多个免疫球蛋白单可变结构域连接,以便提供免疫球蛋白单可变结构域的“衍生物”。在优选但非限制性的方面,所述其他残基可有效预防或减少所谓的“已存在的抗体”与本发明多肽的结合。为此目的,本发明的多肽和构建体可含有C端延伸(X)n(其中n为1至10,优选1至5,例如1,2,3,4或5(并且优选1或2,例如1);并且每个X是独立选择的(优选天然存在的)氨基酸残基,并且优选独立地选自由以下各项组成的组:丙氨酸(A),甘氨酸(G),缬氨酸(V),亮氨酸(L)或异亮氨酸(I),其参考以下共同未决的美国临时申请,名称皆为“改进的免疫球蛋白可变结构域(Improvedimmunoglobulinvariabledomains)”:2014年5月16日提交的US61/994552;2014年6月18日提交的US61/014,015;2014年8月21日提交的US62/040,167;和2014年9月8日提交的US62/047,560(均受让给埃博灵克斯股份有限公司)以及基于这些临时申请并于2015年11月19日发布的国际申请WO2015/173325。因此,在本发明的方法中,多肽可以进一步包含C端延伸(X)n,其中n是1至5,例如1,2,3,4或5,并且其中X是天然存在的氨基酸,优选不含半胱氨酸。在优选的方面,在本发明的方法中表达的本发明的多肽包含SEQIDNO:55或基本上由其组成。在特定方面,这种多肽由SEQIDNO:55组成。在上述多肽中,一个或更多个免疫球蛋白单可变结构域和一个或多个基团、残基、部分或结合单元可以彼此直接连接和/或经由一个或多个更适合的接头或间隔物连接。例如,当一个或多个基团、残基、部分或结合单元是氨基酸时,接头也可以是氨基酸,以使得到的多肽是融合蛋白或融合多肽。在本发明的一个具体方面,制备了与相应的免疫球蛋白单可变结构域相比具有增加的半衰期的本发明的多肽。包含这种延长半衰期的部分的本发明多肽包括但不限于以下多肽:多肽,其中免疫球蛋白单可变结构域合适地连接至一个或多个血清蛋白或其片段(如(人)血清白蛋白或其合适的片段)或连接至一个或多个可结合血清蛋白的结合单元(如例如能结合血清蛋白如血清白蛋白(如人血清白蛋白)、血清免疫球蛋白(如IgG)或转铁蛋白的结构域抗体,适合用作结构域抗体的氨基酸,单结构域抗体,适合用作单结构域抗体的氨基酸,“dAb”,适合用作dAb的氨基酸或纳米抗体);多肽,其中免疫球蛋白单可变结构域连接至Fc部分(如人Fc)或其合适的部分或片段;或多肽,其中一个或多个免疫球蛋白单可变结构域适当地连接至一个或多个能结合血清蛋白的小蛋白质或肽(如但不限于WO91/01743、WO01/45746或WO02/076489中描述的蛋白质和肽)。通常,具有增加的半衰期的本发明的多肽具有的半衰期优选是大于本发明的相应免疫球蛋白单可变结构域或多肽本身的半衰期至少1.5倍、优选至少2倍、如至少5倍、例如至少10倍或超过20倍。通常,在优选但非限制性的方面,与本发明的相应免疫球蛋白单可变结构域或多肽本身相比,本发明的这种多肽具有的血清半衰期增加多于1h、优选多于2h、更优选多于6h、如多于12h或甚至多于24、48或72h。在另一个优选的但非限制性的方面,本发明的这种多肽在人中显示至少约12h、优选至少24h、更优选至少48h、甚至更优选至少72h或者更长的血清半衰期。例如,本发明的多肽可具有至少5天(如约5至10天),优选至少9天(如约9至14天),更优选至少约10天(如约10至15天)或至少约11天(如约11至16天),更优选至少约12天(如约12至18天或更多)或多于14天(如约14至19天)的半衰期。本发明的方法特别适用于本发明的以下免疫球蛋白单可变结构域和多肽,其在适用于毕赤酵母(如巴斯德毕赤酵母)的表达条件下表达时,不容易在这些宿主中表达或以非常低的产率表达;因此,当本发明的这些免疫球蛋白单可变结构域或多肽在毕赤酵母(如巴斯德毕赤酵母)中表达后,不能获得足够的量。因此,本发明的方法特别适用于本发明的免疫球蛋白单可变结构域和/或多肽,其在标准毕赤酵母表达条件(如本文定义)下在毕赤酵母宿主(如巴斯德毕赤酵母)中表达后获得低产率。本发明中使用的“低产率”是指获得的本发明的免疫球蛋白单可变结构域和/或多肽的产率为0.5g/L或更低,如0.4g/L或更低,0.3g/L或更低,0.2g/L或更低,0.1g/L或更低,0.05g/L或更低,0.01g/L或更低,或甚至更低[表达为g(克)本发明免疫球蛋白单可变结构域或多肽/L(升)培养基]。本发明的方法还特别适用于本发明的免疫球蛋白单可变结构域和多肽,其在标准毕赤酵母表达条件(如本文定义)下在毕赤酵母宿主(如巴斯德毕赤酵母)中表达后,所得产率显示与编码所述免疫球蛋白单可变结构域和/或多肽的核酸的拷贝数呈负相关(如本文定义)。显示“负相关”的产率意指当在具有编码所述免疫球蛋白单可变结构域和/或多肽的核酸的一个拷贝的毕赤酵母中(在本文定义的标准毕赤酵母表达条件下)表达所述免疫球蛋白单可变结构域和/或多肽时获得的所述免疫球蛋白单可变结构域和/或多肽的产率高于当在具有编码所述免疫球蛋白单可变结构域和/或多肽的核酸的一个以上拷贝的毕赤酵母中(在本文定义的标准毕赤酵母表达条件下)表达所述免疫球蛋白单可变结构域和/或多肽时获得的所述免疫球蛋白单可变结构域和/或多肽的产率。用于本发明方法的优选多肽包括SEQIDNO:49至54。该序列也形成本发明的单独方面。因此,本发明还涉及具有SEQIDNO:49,50,51,52,53,54,55或56的多肽。用于本发明方法的其他特别优选的多肽包含以下免疫球蛋白单可变结构域(如例如具有SEQIDNO:55或56的氨基酸序列的免疫球蛋白单可变结构域)或基本上由其组成,所述免疫球蛋白单可变结构域基本上由4个框架区(分别为FR1至FR4)和3个互补决定区(分别为CDR1至CDR3)组成,其中CDR1是SEQIDNO:58,CDR2是SEQIDNO:60,以及CDR3是SEQIDNO:62。辅助蛋白如本文所用的术语“辅助蛋白”是指以下蛋白质,其有助于其他分子结构和/或蛋白质发挥其生物学功能,但当这些其他分子结构和/或蛋白质正在发挥其正常的生物学功能时,本身不在这些其他分子结构和/或蛋白质的结构中出现。辅助蛋白可以例如修饰其他分子结构和/或蛋白质的生物物理,药理学和/或表达特性。不受限制,辅助蛋白可以稳定其他分子结构和/或蛋白质(例如通过复合物形成),它们可以调节其他分子结构和/或蛋白质的活性,它们可以增加其他分子结构和/或蛋白质的(表面)表达,和/或它们可以帮助折叠和/或组装。在本发明的方法中表达被增强的辅助蛋白是功能性HAC1蛋白。HAC1蛋白可以源自任何物种,但优选来自酵母来源,最优选来自酵母菌目(Saccharomycetes)的酵母,如来自酵母属(Saccharomyces),Komagataella或毕赤酵母属(汉逊酵母属(Hansenula))的酵母,如酿酒酵母(Saccharomycescerevisiae)或巴斯德毕赤酵母(Pichiapastoris)。在具体方面,功能性HAC1蛋白是“经剪接的HAC1”或“经剪接的HAC1蛋白”(这两个术语在本文中可互换使用)。如Guerfal等.2010(MicrobialCellFactories9:49)所述,经剪接的HAC1是在HAC1mRNA的剪接事件(去除内含子)后获得的HAC1蛋白。在更具体的方面,经剪接的HAC1蛋白来自毕赤酵母属。在优选的方面,经剪接的HAC1蛋白具有Guerfal等.2010中描述的序列(MicrobialCellFactories9:49;SEQIDNO:14)。在本发明的方法中,除HAC1蛋白外,还任选地增强选自蛋白质二硫键异构酶(PDI1;EC5.3.4.1),Kar2p和保守核糖体蛋白P0(RPP0)的一种或多种另外的辅助蛋白的表达。这些辅助蛋白可以源自任何物种,只要它们在毕赤酵母宿主中的表达增强提供本发明的免疫球蛋白单可变和/或多肽的增加的产率。在优选的方面,辅助蛋白来源于真菌,如酵母;优选来源于酵母菌目的酵母,如来自酵母属,Komagataella或毕赤酵母属(汉逊酵母属)的酵母,如酿酒酵母或巴斯德毕赤酵母。在优选的方面,辅助蛋白选自以下:-巴斯德毕赤酵母经剪接的HAC1以及任选的以下的一个或多个:-巴斯德毕赤酵母蛋白质二硫键异构酶(PDI1):-巴斯德毕赤酵母Kar2p:和-巴斯德毕赤酵母60S酸性核糖体蛋白P0(RPP0):本发明的方法本发明涉及在毕赤酵母宿主中表达和/或产生免疫球蛋白单可变结构域和/或包含其的多肽的方法。在本发明的方法中,经剪接的HAC1蛋白在毕赤酵母宿主中的表达得到增强。本发明的方法包括以下步骤:a)在毕赤酵母宿主中表达编码本发明的免疫球蛋白单可变结构域和/或多肽的核酸;和b)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:c)分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽。因此,本发明的方法包括以下步骤:a)在毕赤酵母宿主繁殖的条件下培养所述毕赤酵母宿主;b)在所述毕赤酵母宿主表达和/或产生本发明的免疫球蛋白单可变结构域和/或多肽的条件下维持所述毕赤酵母宿主;和c)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:d)从培养基分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽。为了产生/获得本发明的免疫球蛋白单可变结构域和/或多肽的表达,通常可以在表达/产生本发明的(期望的)免疫球蛋白单可变结构域和/或多肽条件下保持,维持和/或培养转化的毕赤酵母宿主。合适的条件对于技术人员来说是清楚的,并且通常取决于所用的毕赤酵母宿主菌株,以及控制本发明(相关)核苷酸序列表达的调控元件。通常,合适的条件可包括使用合适的培养基,存在合适的食物来源和/或合适的营养素,使用合适的温度,以及任选地存在合适的诱导因子或化合物(例如当本发明的核苷酸序列在诱导型启动子的控制下时);所有这些都可以由技术人员选择。同样,在这样的条件下,本发明的免疫球蛋白单可变结构域和/或多肽可以以组成型方式,以瞬时方式表达,或仅在适当诱导时表达。在毕赤酵母中重组产生异源蛋白质的培养条件是例如由Higgins和Cregg1998(Eds,MethodsinMolecularBiology,Pichiaprotocols,Volume103,2ndEd.,HumanaPress)和由InvitrogenTM(InvitrogenTMPichiaExpressionKit;ForExpressionofRecombinantProteinsinPichiapastoris;Catalogno.K1710-01)描述。巴斯德毕赤酵母中免疫球蛋白单可变结构域的产生已在WO94/25591,WO2010/125187,WO2012/056000和WO2012/152823中广泛描述。这些参考文献的内容在涉及一般培养技术和方法(包括合适的培养基和条件)时明确提及。这些文件的内容通过引用并入本文。本发明还涉及本领域描述的具体条件,例如WO94/25591,Gasser等.2006(Biotechnol.Bioeng.94:535);Gasser等.2007(Appl.Environ.Microbiol.73:6499),或Damasceno等.2007(Microbiol.Biotechnol.74:381)中描述的一般培养方法。毕赤酵母,特别是巴斯德毕赤酵母,通常在30℃在使用甘油作为碳源的补料分批发酵中培养。这种培养基通常包含缓冲剂,甘油,微量元素和氢氧化铵。缓冲剂的实例包括(不限于)H3PO4,CaSO4.2H2O,K2SO4,MgSO4.7H2O,和KOH。常见的生长培养基(例如基础盐培养基)由以下组成:(每升)26.7mL85%H3PO4,0.93gCaSO4.2H2O,18.2gK2SO4,14.9gMgSO4.7H2O,4.13gKOH,40g甘油,2mL微量元素[由6g/L硫酸铜.5H2O;0.8g/L碘化钾;3g/L硫酸锰.H2O;0.2g/L钼酸钠.2H2O;0.2g/L硼酸;0.5g/L硫酸铜;20g/L氯化锌;65g/L硫酸亚铁.7H2O;0.2g/L生物素和5mL浓硫酸构成]。氢氧化铵(NH4OH)用于pH控制(例如pH5)和作为氮源。在补料分批阶段,甘油(例如50%v/v)以例如15mL/L/h的进料速率进料数小时。AOX1启动子用于驱动编码本发明所需免疫球蛋白单可变结构域和/或多肽的目的基因的表达。本发明的免疫球蛋白单可变结构域和/或多肽的表达在30℃进行,甲醇进料速率为4-10mL/L/h(如例如4mL/L/h)。这些条件在本文中也称为“标准毕赤酵母表达条件”。可以通过公知的方法在毕赤酵母宿主中增强辅助蛋白的表达,所述方法包括例如使用合适的控制序列,例如强启动子,和/或增加基因剂量,例如通过增加相应基因的拷贝数。拷贝数可以例如通过引入适合于表达辅助蛋白的遗传构建体(质粒或载体)来增加。质粒或载体的额外存在将增加总拷贝数。此外,可以使用能够独立于毕赤酵母宿主基因组繁殖并且在毕赤酵母宿主中以多拷贝存在的遗传构建体。例如,多拷贝质粒或载体可以在毕赤酵母宿主细胞中以5至50的拷贝数存在。除了毕赤酵母宿主的质粒或载体转化外,毕赤酵母宿主的染色体转化也包括在本发明中。可以在宿主的染色体上引入强(诱导型)启动子(而不是天然辅助蛋白的天然启动子);或者可以将另一个(强)启动子控制下的辅助蛋白基因序列的另一个拷贝引入染色体中。技术人员将知道增强辅助蛋白表达的多种可能性,所有这些都包括在本发明中。本发明的辅助蛋白和免疫球蛋白单可变结构域和/或多肽可以由相同或不同的核酸表达。两种或更多种蛋白质的共表达可以通过在相同的遗传构建体(质粒或载体或整合到宿主的染色体中)上表达这两种或更多种蛋白质来实现;或者通过在不同遗传构建体(质粒或载体或整合到宿主的染色体中)上表达这两种或更多种蛋白质来实现。当在相同的遗传构建体上表达时,编码所述两种或更多种蛋白质的核酸优选彼此相邻。编码所述两种或更多种蛋白质的核酸的转录可以由一种启动子控制(位于两种基因的前面);或者,编码这两种或更多种蛋白质之一的每种核酸可以由单独的启动子控制,所述启动子可以是相同或不同的启动子。当从不同的遗传构建体表达时,编码本发明多肽的核酸的转录和编码一种或多种辅助蛋白的核酸的转录可以由两个可以相同或不同的单独的启动子控制。启动子可以是组成型启动子或诱导型启动子。在一个优选的方面,启动子是诱导型启动子。在本发明的方法中表达增强的辅助蛋白的数量可以是一种(经剪接的HAC1蛋白质),或者可以是多于一种,例如两种,三种,四种,五种或更多种。在优选的方面,在本发明的方法中表达增强的辅助蛋白的数目是一种(经剪接的HAC1蛋白)。在另一个优选的方面,在本发明的方法中表达增强的辅助蛋白的数目是两种,三种,四种或更多种。在该优选方面,增强了经剪接的HAC1蛋白的表达,并且另外增强了一种或多种另外的辅助蛋白的表达。另外的辅助蛋白可以是本领域可获得和/或已知的任何辅助蛋白。优选地,另外的辅助蛋白选自PDI1,Kar2p和RPPO中的任何一种。因此,本发明的方法还包括在毕赤酵母宿主中表达和/或产生免疫球蛋白单可变结构域和/或包含其的多肽,其中经剪接的HAC1蛋白和一种或多种选自PDI1,Kar2p和RPP0的其他辅助蛋白的表达被增强。因此,本发明还涉及包括以下步骤的方法:a)在毕赤酵母宿主中表达编码本发明的免疫球蛋白单可变结构域和/或多肽的核苷酸序列;b)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达和一种,两种,三种(或更多种)编码选自PDI1,Kar2p和RPP0的辅助蛋白的核酸的表达;任选地继之以:c)分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽。在具体方面,本发明的方法包括以下步骤:a)在毕赤酵母宿主繁殖的条件下培养所述毕赤酵母宿主;b)在所述毕赤酵母宿主表达和/或产生本发明的免疫球蛋白单可变结构域和/或多肽的条件下维持所述毕赤酵母宿主;和c)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达和一种,两种,三种(或更多种)编码选自PDI1,Kar2p和RPP0的辅助蛋白的核酸的表达;任选地继之以:d)从培养基分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽。因此,在本发明方法的这个特定方面,可以增强以下辅助蛋白组合的表达:-PDI1和经剪接的HAC1;-Kar2p和经剪接的HAC1;-RPP0和经剪接的HAC1;-PDI1,Kar2p和经剪接的HAC1;-PDI1,RPP0和经剪接的HAC1;-Kar2p,RPP0和经剪接的HAC1;和-PDI1,Kar2p,RPP0和经剪接的HAC1.当辅助蛋白的数目是两种或更多种时,辅助蛋白可以通过相同遗传构建体的作用表达,例如在一个质粒或载体上表达两种或更多种辅助蛋白;或者通过在不同质粒或载体上表达两种或更多种辅助蛋白。除了毕赤酵母宿主的质粒或载体转化外,毕赤酵母宿主的染色体转化也包括在本发明中。可以在宿主的染色体上引入强(诱导型)启动子(而不是天然辅助蛋白的天然启动子);可以将另一个(强)启动子控制下的辅助蛋白基因序列的另一个拷贝引入染色体中。当在相同的遗传构建体上表达时,两种或更多种辅助蛋白可以由相同的启动子或不同的启动子控制。当从不同的遗传构建体表达时,两种或更多种辅助蛋白可以由两个可以相同或不同的单独启动子控制。启动子可以是组成型或诱导型启动子。通过使用上述方法,发明人能够增加有时在免疫球蛋白单可变结构域和/或包含其的多肽的情况下观察到的出乎意料的低产率。特别针对以下观察到低产率:免疫球蛋白单可变结构域(和/或包含其的多肽),其包含两个二硫键;免疫球蛋白单可变结构域(和/或包含其的多肽),其是VHH1型免疫球蛋白单可变结构域;和/或免疫球蛋白单可变结构域(和/或包含其的多肽),其在标准毕赤酵母表达条件下在毕赤酵母宿主中表达后,获得的产率显示与编码所述免疫球蛋白单可变结构域(和/或包含其的多肽)的核酸的拷贝数负相关。因此,本发明还提供了增加免疫球蛋白单可变结构域和/或包含其的多肽的(表达和/或产生)产率的方法,其包括以下步骤:a)在毕赤酵母宿主中表达编码本发明的免疫球蛋白单可变结构域和/或多肽的核酸;和b)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:c)分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽。因此,增加免疫球蛋白单可变结构域和/或多肽的(表达和/或产生)产率的方法包括以下步骤:a)在毕赤酵母宿主繁殖的条件下培养所述毕赤酵母宿主;b)在所述毕赤酵母宿主表达和/或产生本发明的免疫球蛋白单可变结构域和/或多肽的条件下维持所述毕赤酵母宿主;和c)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:d)从培养基分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽。在优选的方面,本发明的方法提供的本发明的免疫球蛋白单可变结构域和/或多肽的产率是未增强经剪接的HAC1蛋白表达的方法中获得的本发明的相同免疫球蛋白单可变结构域和/或多肽的产率的2倍或更多倍(优选3倍或4倍或更多倍,更优选5倍,7.5倍,10倍,15倍,20倍,25倍,30倍,40倍,50或甚至更多倍)。因此,在一个优选的方面,本发明还提供了增加免疫球蛋白单可变结构域和/或包含其的多肽的(表达和/或产生)产率的方法,其包括以下步骤:a)在毕赤酵母宿主中表达编码本发明的免疫球蛋白单可变结构域和/或多肽的核酸;和b)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:c)分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽,其中所述方法提供的本发明的免疫球蛋白单可变结构域和/或多肽的产率是未增强经剪接的HAC1蛋白表达的方法中获得的本发明的相同免疫球蛋白单可变结构域和/或多肽的产率的2倍或更多倍(优选3倍或4倍或更多倍,更优选5倍,7.5倍,10倍,15倍,20倍,25倍,30倍,40倍,50或甚至更多倍)。因此,增加免疫球蛋白单可变结构域和/或多肽的(表达和/或产生)产率的方法包括以下步骤:a)在毕赤酵母宿主繁殖的条件下培养所述毕赤酵母宿主;b)在所述毕赤酵母宿主表达和/或产生本发明的免疫球蛋白单可变结构域和/或多肽的条件下维持所述毕赤酵母宿主;和c)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:d)从培养基分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽,其中所述方法提供的本发明的免疫球蛋白单可变结构域和/或多肽的产率是未增强经剪接的HAC1蛋白表达的方法中获得的本发明的相同免疫球蛋白单可变结构域和/或多肽的产率的2倍或更多倍(优选3倍或4倍或更多倍,更优选5倍,7.5倍,10倍,15倍,20倍,25倍,30倍,40倍,50或甚至更多倍)。在另一个优选的方面,本发明的方法提供本发明的免疫球蛋白单可变结构域和/或多肽的产率为1g/L或更多,更优选为1.5g/L或更多,2g/L或更多,或甚至2.5g/L或更多。因此,在另一个优选的方面,本发明还提供了增加免疫球蛋白单可变结构域和/或包含其的多肽的(表达和/或产生)产率的方法,其包括以下步骤:a)在毕赤酵母宿主中表达编码本发明的免疫球蛋白单可变结构域和/或多肽的核酸;和b)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:c)分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽,其中所述方法提供本发明的免疫球蛋白单可变结构域和/或多肽的产率为1g/L或更多,更优选为1.5g/L或更多,2g/L或更多,或甚至2.5g/L或更多。因此,增加免疫球蛋白单可变结构域和/或多肽的(表达和/或产生)产率的方法包括以下步骤:a)在毕赤酵母宿主繁殖的条件下培养所述毕赤酵母宿主;b)在所述毕赤酵母宿主表达和/或产生本发明的免疫球蛋白单可变结构域和/或多肽的条件下维持所述毕赤酵母宿主;和c)在所述毕赤酵母宿主中增强编码经剪接的HAC1蛋白的核酸的表达;任选地继之以:d)从培养基分离和/或纯化由此获得的本发明的免疫球蛋白单可变结构域和/或多肽,其中所述方法提供本发明的免疫球蛋白单可变结构域和/或多肽的产率为1g/L或更多,更优选为1.5g/L或更多,2g/L或更多,或甚至2.5g/L或更多。本发明的免疫球蛋白单可变结构域和/或多肽在细胞外产生,并从培养毕赤酵母宿主细胞的培养基中分离。通常但非必要地,本发明的免疫球蛋白单可变结构域和/或多肽将至少具有将本发明的免疫球蛋白单可变结构域和/或多肽引向周质的转运信号。在本发明中,可以通过常规方法从培养基中除去毕赤酵母宿主。例如,可以通过离心或过滤除去毕赤酵母宿主。通过从培养基中除去毕赤酵母宿主获得的溶液也称为培养上清液或澄清的培养上清液。本领域技术人员还将清楚,本发明的免疫球蛋白单可变结构域和/或多肽可以(首先)以未成熟形式(如上所述)产生,然后可以进行翻译后修饰,这取决于使用的毕赤酵母宿主。而且,本发明的免疫球蛋白单可变结构域和/或多肽可以是糖基化的,这同样取决于使用的毕赤酵母宿主。随后可以从毕赤酵母宿主和/或从通过标准方法培养所述毕赤酵母宿主的培养基中分离本发明的免疫球蛋白单可变结构域和/或多肽。标准方法包括但不限于色谱方法,包括尺寸排阻色谱,疏水色谱,离子交换色谱和亲和色谱。这些方法可以单独进行或与其他纯化方法组合进行,例如,差异沉淀技术,凝胶电泳,亲和技术(例如使用与本发明的免疫球蛋白单可变结构域和/或多肽融合的特异性可切割氨基酸序列)和/或制备型免疫学技术(即使用针对待分离的本发明的免疫球蛋白单可变结构域和/或多肽的抗体)。技术人员可以基于公知常识设计用于免疫球蛋白单可变结构域的纯化方法的合适组合。对于具体实例,参考本文引用的技术。免疫球蛋白单可变结构域和/或包含其的多肽可以通过蛋白A亲和色谱,离子交换色谱和尺寸排阻色谱从培养上清液中纯化。提及任何“纯化步骤”,包括但不限于这些特定方法。更具体地,可以使用以下方法来从培养上清液纯化免疫球蛋白单可变结构域和/或包含其的多肽:其中将(通过离心获得的)澄清的上清液捕获在蛋白A树脂上,然后是SOURCE15S(GEHealthcare)阳离子交换色谱步骤和Superdex75(GEHealthcare)SEC步骤。除去毕赤酵母宿主后,本发明的免疫球蛋白单可变结构域和/或多肽可以存在于多种合适的缓冲液中。实例包括但不限于PBS,Tris-HCl,组氨酸或磷酸盐缓冲液。本发明的免疫球蛋白单可变结构域和/或多肽也可以存在于生理盐水中。通常,对于药物用途,本发明的免疫球蛋白单可变结构域和/或多肽可以配制成药物制剂或组合物,其包含至少一种本发明的免疫球蛋白单可变结构域和/或多肽和至少一种药学上可接受的载体,稀释剂或赋形剂和/或佐剂,和任选的一种或多种其他药学活性多肽和/或化合物。通过非限制性实例,这种配制品可以是适于口服施用,胃肠外施用(例如通过静脉内施用,肌内施用或皮下注射或静脉内输注施用),局部施用,吸入施用,通过皮肤贴剂施用,通过植入物施用,通过栓剂施用等的形式。这种合适的给药形式-可以是固体,半固体或液体,取决于施用方式-以及用于其制备的方法和载体对于本领域技术人员将是清楚的。本发明的核酸和遗传构建体本发明还涉及编码本发明的免疫球蛋白单可变结构域和/或多肽和经剪接的HAC1的核酸。这些核酸在本文中也称为“本发明的核酸”。如本领域技术人员将清楚的,本发明的核酸可以是遗传构建体的形式、存在于其中和/或作为其一部分。此类遗传构建体通常包含至少一种本发明的核酸,其任选地连接至本身已知的遗传构建体的一个或多个元件,如例如一个或多个合适的调控元件(如合适的启动子、增强子、终止子等)以及本文涉及的遗传构建体的其他元件。包含本发明的至少一种核酸的此类遗传构建体在本文中也将被称为“本发明的遗传构建体”。因此,本发明的遗传构建体至少编码本发明的免疫球蛋白单可变结构域和/或多肽以及经剪接的HAC1。由本发明的核酸和遗传构建体编码的辅助蛋白的数量可以是一种(即经剪接的HAC1蛋白)或可以多于一种,例如两种,三种,四种,五种或更多种。在优选的方面,由本发明的核酸和遗传构建体编码的辅助蛋白的数目是一种(即经剪接的HAC1蛋白)。在另一个优选的方面,由本发明的核酸和遗传构建体编码的辅助蛋白的数目是两种或更多种。因此,本发明还包括编码本发明的免疫球蛋白单可变结构域和/或多肽和两种或更多种辅助蛋白(包括经剪接的HAC1蛋白)的核酸或遗传构建体。如上所述,本发明的免疫球蛋白单可变结构域和/或多肽和辅助蛋白可以从单一核酸和/或遗传构建体共表达;或者从不同的(单独的)核酸和/或遗传构建体共表达(可能包括自宿主染色体表达)。编码本发明的免疫球蛋白单可变结构域和/或多肽和/或编码辅助蛋白的所有这些核酸和/或遗传构建体(作为一个构建体或作为单独的构建体)包括在术语“本发明的核酸”和“本发明的遗传构建体”中。本发明的核酸可以是单链或双链DNA或RNA的形式,并且优选为双链DNA的形式。例如,本发明的核酸可以是基因组DNA、cDNA或合成DNA(如具有专门适用于在毕赤酵母宿主细胞中表达的密码子使用的DNA)。根据本发明的一个实施方案,本发明的核酸处于如本文所定义的基本上分离的形式。本发明的核酸也可以是载体(如例如质粒,粘粒或YAC)的形式,可以存在于其中和/或是其一部分,所述载体也可以是基本上分离的形式。基于待表达的本发明的免疫球蛋白单可变结构域和/或多肽的以及用于共表达的辅助蛋白的信息,可以以本身已知的方式制备或获得本发明的核酸。此外,如本领域技术人员所清楚的,为了制备本发明的核酸,可以以合适的方式将若干个核苷酸序列,例如至少一个编码本发明的免疫球蛋白单可变结构域和/或多肽的核苷酸序列和至少一个编码辅助蛋白的核苷酸序列连接在一起。用于产生本发明的核酸的技术对于本领域技术人员将是清楚的,并且可以例如包括但不限于自动化DNA合成;定点诱变;组合两个或更多个天然存在的和/或合成的序列(或其两个或更多个部分),引入导致截短表达产物表达的突变;引入一个或多个限制性位点(例如以产生可使用合适的限制性酶容易地消化和/或连接的盒和/或区域)和/或通过PCR反应使用一个或多个“错配“引物引入突变。这些和其他技术对于技术人员将是清楚的,并且再次参考标准手册,例如本文提到的Sambrook等人和Ausubel等人,以及下面的实施例。本发明的遗传构建体可以是DNA或RNA,并且优选是双链DNA。本发明的遗传构建体也可以是适于转化毕赤酵母宿主的形式,适于整合到毕赤酵母宿主细胞的基因组DNA中的形式,或适于在毕赤酵母宿主中独立复制、维持和/或遗传的形式。例如,本发明的遗传构建体可以是载体的形式,例如质粒、YAC、病毒载体或转座子。特别地,载体可以是表达载体,即可以在毕赤酵母宿主中提供表达的载体。在优选但非限制性的实施方案中,本发明的遗传构建体包含:a)至少一个本发明的核酸;其可操作地连接至b)一个或多个调控元件,例如启动子和任选的合适的终止子;并且还任选c)本身已知的遗传构建体的一个或多个其他元件;其中术语“调控元件”、“启动子”、“终止子”和“可操作地连接”具有它们在本领域中的通常含义(如本文进一步所述);并且其中存在于遗传构建体中的所述“其他元件”可以例如是3'-或5'-UTR序列、前导序列、选择标记、表达标记/报道基因和/或可以促进或增加转化或整合(的效率)的元件。这些遗传构建体的这些和其他合适的元件对技术人员来说是清楚的,并且可以例如取决于所使用的构建体的类型;毕赤酵母宿主菌株;感兴趣的发明的核苷酸序列的表达方式(例如通过组成型、瞬时或诱导型表达);和/或要使用的转化技术。例如,本身已知用于抗体和抗体片段(包括但不限于(单)结构域抗体和ScFv片段)的表达和产生的调控序列、启动子和终止子可以以基本类似的方式使用。优选地,在本发明的遗传构建体中,本发明的所述至少一个核酸和所述调控元件,以及任选地所述一个或多个其他元件彼此“可操作地连接”,这通常意味着它们是彼此处于功能关系中。例如,如果启动子能够启动或以其他方式控制/调控编码序列的转录和/或表达(其中所述编码序列应被理解为在所述启动子“的控制下”),则认为所述启动子与所述编码序列“可操作地连接”。通常,当两个核苷酸序列可操作地连接时,它们将处于相同的取向并且通常也处于相同的阅读框中。它们通常也基本上是连续的,尽管这也可能不是必需的。在本发明的一个方面,编码本发明的免疫球蛋白单可变和/或多肽的核苷酸序列与编码辅助蛋白的核苷酸序列可操作地连接。它们可以在相同启动子的控制下,或者它们各自可以在单独的(相同或不同的)启动子的控制下。设计,创建或获得用于表达的核酸序列的方法,构建合适载体的方法,将核酸序列插入载体中的方法,选择合适的毕赤酵母宿主菌株的方法,将载体引入毕赤酵母宿主菌株的方法,引起或允许表达多肽或蛋白质的方法,从毕赤酵母宿主菌株分离核酸的方法或鉴定核酸序列和相应的蛋白质序列的方法是标准方法(Sambrook等1989),这是本领域普通技术人员所熟知的。本领域技术人员还可以基于常识,设计合适的遗传构建体用于在毕赤酵母宿主中表达本发明的免疫球蛋白单可变结构域和/或多肽。本发明还涉及本领域描述的遗传构建体,例如WO94/25591,Cereghino和Cregg2000(Curr.OpinionBiotechnol.10:422),Gasser等.2006(Biotechnol.Bioeng.94:535),Gasser等.2007(Appl.Environ.Microbiol.73:6499)或Damasceno等.2007(Microbiol.Biotechnol.74:381)中描述的质粒、启动子和前导序列。优选地,本发明的遗传构建体的调控和其他元件使得它们能够在毕赤酵母宿主中提供其预期的生物学功能。例如,启动子,增强子或终止子应该在毕赤酵母宿主中是“可操作的”,这意味着(例如)所述启动子应该能够启动或控制/调控与其可操作地连接(如本文所定义)的核苷酸序列例如编码序列的转录和/或表达。合适的启动子,终止子和其他元件的一些优选但非限制性的实例包括可用于在毕赤酵母宿主中表达的那些;特别是在此提及的那些和/或在下面的实施例中使用的那些。一些特别优选的启动子包括但不限于本身已知在毕赤酵母宿主中表达的启动子;特别是在此提及的那些和/或在实施例中使用的那些。启动子的具体序列决定了启动子的强度(“强启动子”导致高转录起始速率)。当编码辅助蛋白的核酸的表达被称为受“强启动子”控制时,意味着编码所述辅助蛋白的核酸的表达受以下启动子的控制,其比控制天然辅助蛋白转录起始的天然启动子导致更高的转录起始速率。除了“启动”转录的序列之外,启动子可以包括称为操纵子的其他序列,其控制启动子的强度。例如,启动子可包括吸引或阻碍RNA与启动子的结合的蛋白质结合位点。该蛋白质的存在与否会影响启动子的强度。这种启动子被称为受调控的启动子。巴斯德毕赤酵母醇氧化酶I(AOX1)启动子是已知的最强的,调控性最好的启动子之一。相反,巴斯德毕赤酵母第二醇氧化酶(AOX2)由弱得多的启动子控制。巴斯德毕赤酵母甘油醛-3-磷酸脱氢酶(GAP)启动子在葡萄糖,甘油和甲醇培养基上提供组成型高水平的表达(Waterham等.1997,Gene186:37)。巴斯德毕赤酵母甲醛脱氢酶(FLD1)启动子可以通过甲醇或甲胺诱导,其表达水平与甲醇中AOX1启动子获得的表达水平相当(Shen等.1998,Gene216:93)。Peroxin8(PEX8)启动子在葡萄糖上产生低表达,并且当细胞转移到甲醇时被适度诱导(约10倍)(Johnson等.1999,Genetics151:1379)。多形汉逊酵母(H.polymorpha)中的强启动子包括衍生自甲醇氧化酶(MOX),甲酸脱氢酶(FMD)和二羟基丙酮合酶(DHAS)基因的元件(Song等.2006,Biotechnol.Lett.25:1999)。多形汉逊酵母中的甘油醛-3-磷酸脱氢酶(GAP1)启动子(Sohn等.1999,Appl.Microbiol.Biotechnol.51:800)和PMA1启动子(Cox等.2000,Yeast16:1191)是组成型元件。就高表达水平而言,PMA1启动子与MOX启动子竞争。甲醇毕赤酵母(P.methanolica)醇氧化酶(AUG1)启动子P(MOD1)和P(MOD2)受甲醇(P(MOD1)和P(MOD2))和甘油(仅P(MOD1))的强烈和严格调节(Nakagawa等.2006,Yeast23:15)。来自博伊丁假丝酵母(C.boidinii)的强启动子包括醇氧化酶(AOD1)启动子和二羟基丙酮合酶(DAS1)启动子(Yurimoto等.2000,Biochim.Biophys.Acta1493:56)。DAS1和甲酸脱氢酶(FMD)启动子均可获得于博伊丁假丝酵母(Sakai等.1995,Appl.Microbiol.Biotechnol.42:860;1996,Biochim.Biophys.Acta1308:81)和多形汉逊酵母(Hollenberg和Gellissen1997,Curr.Opin.Biotechnol.8:554)。选择标记应当是这样的,其允许-即在适当的选择条件下-已经(成功)用本发明的核酸转化的毕赤酵母宿主细胞区别于尚未(成功)转化的毕赤酵母宿主细胞。这些标记的一些优选但非限制性的实例是提供抗生素(如例如博莱霉素(Zeocin),杀稻瘟素,遗传霉素(G418),腐草霉素,卡那霉素或氨苄青霉素)抗性的基因,提供耐温性的基因,或允许毕赤酵母宿主在不存在对于非转化细胞存活必需的某些因子,化合物和/或(食物)成分的情况下在培养基中维持的基因。前导序列应该是使得-在毕赤酵母宿主中-它允许所需的翻译后修饰和/或从所述细胞分泌表达产物。因此,前导序列可以是在毕赤酵母宿主中有效的任何原(pro)序列,前(pre)序列或前原(prepro)序列。然而,通常但非必要地,本发明的免疫球蛋白单可变结构域和/或多肽将至少具有将本发明的免疫球蛋白单可变结构域和/或蛋白质引向周质的转运信号。例如,本身已知用于抗体和抗体片段(包括但不限于(单)结构域抗体和ScFv片段)的表达和产生的前导序列可以以基本类似的方式使用。一些优选但非限制性的分泌序列包括酿酒酵母衍生的α-交配因子信号序列,巴斯德毕赤酵母衍生的酸性磷酸酶(PHO1)信号序列,巴斯德毕赤酵母衍生的磷酸酶(pho1)前导序列,酵母转化酶(Suc)的分泌信号,人血清白蛋白信号肽,S.occidentalis衍生的GAM1信号序列,以及普通滨蟹(Carcinusmaenas)衍生的高血糖激素(CHH)序列等。技术人员还可以设想使用衍生自基因组测序实验的预测信号肽。这些预测的信号肽序列可以源自任何物种,但优选来自酵母来源,最优选来自酵母菌目的酵母,例如来自酵母属,Komagataella或毕赤酵母属(汉逊酵母属)的酵母,例如酿酒酵母或巴斯德毕赤酵母。DeSchutter等.2009(NatureBiotech27(6):561-566)描述了一些优选但非限制性的源自巴斯德毕赤酵母的预测信号肽。对于将这种预测的信号肽用于产生免疫球蛋白单可变结构域,另外参考WO2012/152823。还可以修饰已知或预测的分泌序列以改善所产生的本发明的免疫球蛋白单可变结构域和/或多肽的性质。这些修饰可以例如通过提高加工效率来提高本发明的免疫球蛋白单可变结构域和/或多肽的纯度。例如在WO2012/152823中描述了用于提高处理效率的α-交配因子信号序列的修饰。表达标记或报告基因应该是使得-在毕赤酵母宿主中-它允许检测遗传构建体(上存在的基因或核苷酸序列)的表达。此类报告基因也可以表达为与本发明的免疫球蛋白单可变结构域和/或多肽融合的蛋白质。一些优选但非限制性的实例包括荧光蛋白,例如GFP和荧光素酶(LUC)。本发明的遗传构建体通常可以通过以下来提供:例如使用在一般手册例如上文提到的Sambrook等人和Ausubel等人中所描述的技术,适当地将本发明的核酸和/或核苷酸序列与上述一个或多个其他元件连接。通常,本发明的遗传构建体将通过将本发明的核酸和/或核苷酸序列插入本身已知的合适(表达)载体中获得。合适的表达载体的一些优选的但非限制性的例子是在以下实施例中使用的那些,以及本文提及的那些。用于本发明遗传构建体的一些优选但非限制性载体包括用于在酵母或其他真菌细胞中表达的载体,例如pYES2(Invitrogen),pUR3515和pUR3501(Sierkstra等.1991,Curr.Genet.19:81)和毕赤酵母表达载体,如例如(不限于)由Invitrogen提供的用于巴斯德毕赤酵母表达的pPICZ载体,pPIC3.5,pPIC3.5K,pPIC6a,pPIC9,pPIC9K,pHIL-D2,pHIL-S1,用于甲醇毕赤酵母表达的pMET,pMETα。对于毕赤酵母表达载体的非详尽列表,还参考Daly和Hearn2004(J.Mol.Recognition18:119),PichiaProtocols2007(Ed.Cregg,2ndEd.,HumanaPress,NJ)和Gelissen2000(Appl.Microbiol.Biotechnol.54:741)。本发明还包括制备本发明核酸和遗传构建体的方法,包括在合适的载体中克隆辅助蛋白基因和/或编码本发明的免疫球蛋白单可变结构域和/或多肽的核苷酸序列的步骤。本发明的核酸和/或本发明的遗传构建体可用于转化毕赤酵母宿主,即用于表达和/或产生本发明的免疫球蛋白单可变结构域和/或多肽。用于转化毕赤酵母宿主的合适技术对于技术人员来说是清楚的。再次参考上述手册和专利申请。关于转化方法的更多细节,参考Invitrogen的EasySelectTM毕赤酵母表达手册(PichiaExpressionmanual),参考Wu和Letchworth2004(BioTechniques36:152),参考PichiaProtocols2007(Ed.Cregg,2ndEd.,HumanaPress,NJ)和参考Faber1994(Curr.Genet.25:305)。转化后,可以进行检测和选择已经用本发明的核苷酸序列/遗传构建体成功转化的毕赤酵母宿主细胞的步骤。例如,这可以是基于本发明的遗传构建体中存在的选择标记的选择步骤或涉及例如使用特异性抗体检测本发明的氨基酸序列的步骤。因此,本发明还涉及包含本发明的遗传构建体或核酸的毕赤酵母宿主细胞的制备。技术人员可以通过常规措施例如转化将本发明的核酸或遗传构建体引入毕赤酵母宿主中。然后,技术人员可以例如通过监测辅助蛋白在核酸和/或蛋白质水平上的表达选择包含核酸或遗传构建体的合适的毕赤酵母宿主细胞。将选择具有令人满意的表达水平的菌株。期望辅助蛋白的高表达,但是,其不应高到导致与本发明的免疫球蛋白单可变结构域和/或多肽的表达竞争。这可以通过常规方法确定。转化的毕赤酵母宿主细胞(其可以是稳定细胞系的形式)形成本发明的另一方面。毕赤酵母宿主因此,本发明还涉及包含如上所述的这种遗传构建体或核酸的毕赤酵母宿主。术语“毕赤酵母宿主”和“毕赤酵母宿主细胞”可互换使用,并且是指具有球形,椭圆形或长方渐尖形细胞的酵母科(Saccharomycetaceae)中的毕赤酵母属(汉逊酵母属和生丝毕赤酵母属(Hyphopichia)是废弃的同义词)。毕赤酵母是一种有性型,在有性繁殖期间形成帽状,半球形或圆形子囊孢子。一些毕赤酵母物种的无性型是假丝酵母(Candida)物种。无性繁殖是通过多边出芽。本发明无限制地涉及毕赤酵母宿主,条件是它们适用于产生免疫球蛋白单可变结构域和/或包含其的多肽。出于本发明的目的,术语“毕赤酵母宿主”还包括汉逊酵母和假丝酵母物种。本发明的毕赤酵母宿主能够产生本发明的免疫球蛋白单可变结构域和/或多肽和经剪接的HAC1蛋白,以及任选的一种或多种另外的辅助蛋白,如例如蛋白二硫键异构酶(PDI1),Kar2p或保守核糖体蛋白P0(RPP0)。通常对其进行遗传修饰,使其包含一个或多个编码本发明的免疫球蛋白单可变结构域和/或多肽的核酸,并增强经剪接的HAC1蛋白的表达。遗传修饰的非限制性实例包括例如用质粒或载体转化,或用病毒载体转导。一些宿主可以通过融合技术进行遗传修饰。遗传修饰包括将分开的核酸分子例如质粒或载体引入宿主,以及例如通过同源重组例如通过整合到宿主的染色体中对宿主遗传物质的直接修饰。通常将两者组合,例如,用质粒转化宿主,该质粒在同源重组后(至少部分地)整合到宿主染色体中。技术人员知道对宿主的遗传修饰的合适方法,以使宿主能够产生本发明的免疫球蛋白单可变结构域和/或多肽。合适的毕赤酵母宿主对于技术人员来说是清楚的,并且可以是例如酵母,包括但不限于毕赤酵母,汉逊酵母或假丝酵母,例如甲基营养型酵母,包括巴斯德毕赤酵母,甲醇毕赤酵母,多形汉逊酵母(Hansenulapolymorpha,Pichiaangusta)和博伊丁假丝酵母。对于非详尽的毕赤酵母菌株列表,参考例如Gelissen2000(Appl.Microbiol.Biotechno.54:741)。不受限制,巴斯德毕赤酵母菌株列于Daly和Hearn2004(J.Mol.Recognition,18:119)和PichiaProtocols2007(Ed.Cregg,2ndEd.,HumanaPress,NJ)。巴斯德毕赤酵母菌株的实例包括(不限于)由Invitrogen提供的由Cereghino和Cregg2004(FEMSMicrobiol.Rev.24:45)、Macauley-Patrick等.2005(Yeast22:249)和/或Damasceno等.2007(Appl.Microbiol.Biotechno.74:381)描述的X33,GS115,KM71,KM71H,SMD1163,SMD1165,SMD1168,SMD1168H,NRRL-Y11430,GS200。多形汉逊酵母菌株的实例包括(不限于)A16(Veale等.1992,Yeast8:361),GF16(Faber1994,Proc.Natl.Acad.Sci.USA91:12985),CBS4732(CCY38-22-2;ATCC34438,NRRL-Y-5445),DL-1(NRRL-Y-7560;ATCC26012),以及菌株NCYC495(CBS1976;ATAA14754,NRLL-Y-1798)。甲醇毕赤酵母菌株的实例包括(不限于)Invitrogen提供的PMAD11和PMAD16。Nakagawa等.1996(J.FermentationBioeng.81:498)描述了甲醇毕赤酵母菌株(IAM12901和IAM12481)和博伊丁假丝酵母菌株(IAM12875)。还参考上文引用的一般
背景技术:
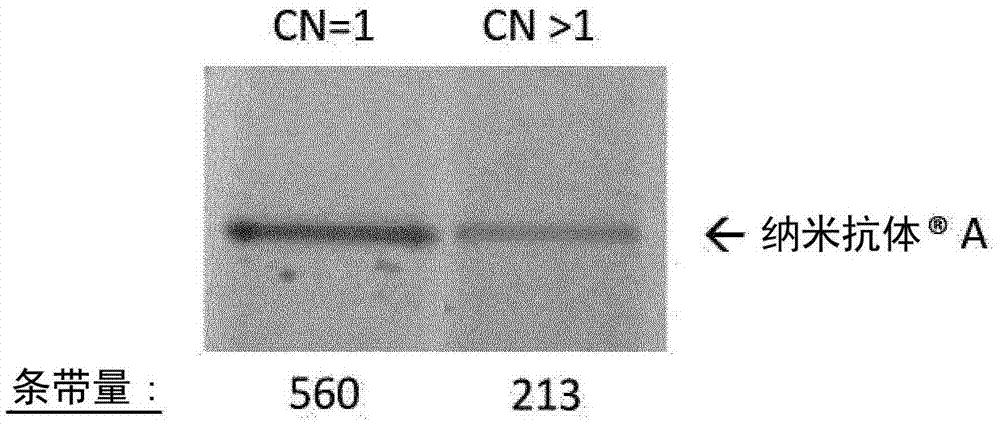
,以及例如WO94/29457,Frenken等.1998(Res.Immunol.149:589),vanderLinden2000(J.Biotechnol.80:261),Joosten等.2003(Microb.CellFact.2:1),以及本文引用的其他参考文献。对于工业规模的生产,用于(工业)生产含有免疫球蛋白单可变结构域的蛋白质治疗剂的优选异源宿主包括适合于大规模表达/生产/发酵特别是大规模药物表达/生产/发酵的巴斯德毕赤酵母菌株。这种菌株的合适实例对于技术人员来说是清楚的。这种菌株和生产/表达系统也可由例如AveciaBiologics(Billingham,NorthEastEngland,UK),BIOMEVAGmbH(Heidelberg,Germany),PharmedArtisGmbH(Aachen,Germany),Richter-Helm(Hamburg,Germany),和CMCBiologics(Copenhagen,Denmark)等公司提供。本发明还包括本发明的毕赤酵母宿主细胞的其他世代,后代和/或子代,其可以例如通过细胞分裂获得。药物制剂本发明还涉及通过本文所述的本发明方法能够获得的本发明的免疫球蛋白单可变结构域和/或多肽。因此,本发明还涉及药物制剂和其他组合物,其包含通过本发明的方法能够获得的本发明的免疫球蛋白单可变结构域和/或多肽。本发明还涉及通过本发明的方法能够获得的本发明的免疫球蛋白单可变结构域和/或多肽的医学用途。技术人员可以基于共同的常识容易地配制药学上合适的配制品。此外,明确提及了本文引用的具体涉及免疫球蛋白单可变结构域和/或纳米抗体的参考文献。非限制性地,可以制备用于标准施用途径的配制品,包括用于鼻,口,静脉内,皮下,肌内,腹膜内,阴道内,直肠施用,局部施用或通过吸入施用的配制品。基于本发明,技术人员还可以容易地设计合适的治疗方法,其特征在于使用通过本发明的方法能够获得的治疗有效量的本发明的免疫球蛋白单可变结构域和/或多肽。通过以下实施例进一步说明本发明,这些实施例决不应被解释为进一步的限制。本申请通篇引用的所有参考文献(包括参考文献、授权专利、公开的专利申请和共同未决的专利申请)的全部内容在此明确地通过引用并入,特别是对于上文提到的教导。实施例实验部分描述了在巴斯德毕赤酵母中单价和多价免疫球蛋白单可变结构域表达后的低产率的出人意料的观察结果。我们还观察到,当巴斯德毕赤酵母的基因组中存在多于1个拷贝的表达盒时,产率进一步降低。还描述了通过增强经剪接的HAC1的表达来增加所述免疫球蛋白单可变结构域的表达产率的方法。实施例1:鉴定增加巴斯德毕赤酵母中纳米抗体表达的辅助蛋白1.1表达载体的构建先前在WO2013/045707中描述为SEQIDNO:7的纳米抗体A是由重链美洲驼抗体的两个序列优化的可变结构域组成的二价纳米抗体。纳米抗体A中的N端亚基是VHH1型免疫球蛋白单可变结构域,并且特异结合c-Met,而C端亚基结合人血清白蛋白(HSA)。使用9G/S接头使亚基头尾融合。纳米抗体A(SEQIDNO:49)的序列描述于表A-1中。先前显示纳米抗体A在巴斯德毕赤酵母中发酵后给出非常低的产率(0.2g/L或更低)。将含有纳米抗体A的编码信息的DNA片段克隆到含有杀稻瘟素TM抗性基因标记的毕赤酵母表达载体(pPIC6a的衍生物,Invitrogen)的多克隆位点中,使得纳米抗体序列位于aMF信号肽序列下游并与之同框。为了产生基因组中具有多于1个拷贝数的表达盒的毕赤酵母克隆,使用毕赤酵母表达载体中的独特BglII位点来引入纳米抗体A的第二表达盒。将表A-2中描述的辅助蛋白的编码序列克隆(使用限制性酶BstBI和NotI)到含有博莱霉素TM抗性基因标记的毕赤酵母表达载体(pPICZa的衍生物,Invitrogen)中。使用限制性酶AfeI和NotI克隆在其编码序列中含有BstBI位点的辅助蛋白。pPIC6a和pPICZa载体中的纳米抗体和辅助蛋白均在AOX1甲醇诱导型启动子的控制下。1.2纳米抗体编码序列的转化以及纳米抗体在巴斯德毕赤酵母中的表达和分泌野生型毕赤酵母X33的转化和表达研究通过标准技术并根据‘UsermanualforpPicZalphaA,BandC’(版本D,110801,手册部分号25-0148;Invitrogen)和MethodsinMolecularBiology2007(HumanaPressInc.)进行。首先,用具有纳米抗体A的单或双表达盒的合适的表达载体转化巴斯德毕赤酵母菌株。转化体在含有杀稻瘟素TM的选择性培养基上生长。通过qPCR表征一些个体菌落以选择具有整合到基因组中的1个拷贝的表达载体的克隆和具有整合到基因组中的一个以上拷贝的表达盒的克隆。验证了纳米抗体在培养基中的表达和分泌(图1)。1.3辅助蛋白编码序列的转化以及纳米抗体在巴斯德毕赤酵母中的表达和分泌一旦鉴定出合适的表达纳米抗体的菌落,就将其接种物繁殖并制备成感受态细胞。然后用含有表A-2中所示的22种辅助蛋白的表达载体文库转化这些细胞。转化体在含有不同选择标记(博莱霉素TM)的选择培养基上生长,这样,获得含有目标纳米抗体和表A-2的一种或多种辅助蛋白的共转化体。摇瓶表达在BMCM培养基中的5mL培养物中进行,并如毕赤酵母方案中所述(参见例如MethodsinMolecularBiology2007,HumanaPressInc.)通过加入甲醇进行诱导。在每个设置中针对提高的表达筛选1128个克隆,并与它们相应的参比克隆进行比较,所述参比克隆仅含有整合到基因组中的1个或一个以上拷贝的纳米抗体表达盒但没有编码一种或多种辅助蛋白的表达载体。对于每个设置,我们发现2个克隆的产量显著高于其参比克隆。克隆6H1和4C2具有整合在基因组中的一个拷贝的纳米抗体编码序列(拷贝数=1),克隆5A6和9C4具有整合在基因组中的一个以上拷贝的纳米抗体编码序列(拷贝数>1)(图2)。1.4鉴定对表达产率具有积极作用的辅助蛋白通过使用序列特异性PCR引物的基因组DNAPCR,鉴定对巴斯德毕赤酵母中的纳米抗体的表达产率具有积极作用的辅助蛋白。使用的引物列表如表A-3所示。鉴定的辅助蛋白如表1所示。表1:用特异性PCR引物鉴定存在于4个显示纳米抗体A表达水平增加的克隆(6H1,4C2,5A6,9C4)中的辅助蛋白。黑框表示相应克隆中具体辅助蛋白的存在。存在于克隆4C2和6H1两者中或5A6和9C4两者中的辅助蛋白用箭头指示。1.5确定不同克隆的表达产率在表达实验中通过对培养基中表达和分泌的纳米抗体产率进行定量,将纳米抗体/辅助蛋白共转化体的表达产率与对照(不增强一种或多种辅助蛋白的表达的纳米抗体转化体)的表达产率进行比较。使用标准的补料分批发酵条件。进行甘油补料分批并通过加入甲醇引发诱导。在pH6,30℃以2L规模,在复合培养基中进行生产,甲醇进料速率为4ml/L*h。对样品进行SDS-PAGE分析。通过考马斯染色的SDS-PAGE光密度测定扫描测量进行蛋白质的相对定量(表2)。表2:具有和不具有增强的辅助蛋白表达的纳米抗体A的表达产率。使用SDS-PAGE/考马斯染色评估产率,并使用Imagequant软件(GEHealthcare)进行条带量的定量。与其参比克隆(1个表达盒拷贝整合到基因组中)相比,克隆4C2和6H1表现出显著的表达增加。表达的这种增加可能是两个克隆中存在的PDI1的共表达的结果。类似地,与其参比克隆相比,克隆5A6和9C4显示出大量的产率增加。在克隆5A6和9C4中均表达的辅助蛋白是Kar2p,RPP0和经剪接的HAC1。最有可能的是,这些辅助蛋白参与提高纳米抗体A的表达。有趣的是,克隆4C2的表达水平显著高于克隆6H1,这可能是Kar2p和经剪接的HAC1的共表达的结果,其也存在于最高表达克隆5A6和9C4中。具有最高产率的克隆全部共表达经剪接的HAC1(表1和2)。实施例2:当增强单个辅助蛋白的表达时,评估纳米抗体A产率将单个辅助蛋白PDI1,Kar2p,RPP0和经剪接的HAC1转化到参比克隆中,如实施例1.3中所述,所述参比克隆在基因组中具有一个以上拷贝的纳米抗体表达盒。转化体在含有博莱霉素TM的选择性培养基上生长。获得含有纳米抗体A和具体辅助蛋白的共转化体。摇瓶表达在BMCM培养基中的5mL培养物中进行并通过加入甲醇进行诱导,如毕赤酵母方案中所述(参见例如MethodsinMolecularBiology2007,HumanaPressInc.)。通过考马斯染色的SDS-PAGE光密度测定扫描测量进行蛋白质的相对定量(图3)。共表达辅助蛋白之一的所有克隆显示出纳米抗体A的产率显著增加。再次,我们观察到含有经剪接的HAC1的克隆显示出最大的产率提高。实施例3:当增强单个辅助蛋白的表达时,评估纳米抗体B产率3.1表达载体的构建纳米抗体B是由重链美洲驼抗体的三个序列优化的可变结构域组成的三价纳米抗体。纳米抗体B中的亚基不是VHH1型免疫球蛋白单可变结构域,并且不与人血清白蛋白(HSA)结合。使用35G/S接头使亚基头尾融合。先前显示纳米抗体B在巴斯德毕赤酵母中发酵后给出非常低的产率(0.29g/L或更低)。将含有纳米抗体B的编码信息的DNA片段克隆到含有博莱霉素TM抗性基因标记的毕赤酵母表达载体(pPpT4_α_S的衍生物;等.2012,PLoSOne7:e39720)的多克隆位点中,使得纳米抗体序列位于aMF信号肽序列下游并与之同框。将辅助蛋白经剪接的HAC1,Kar2p,PDI1和RPP0的编码序列克隆到含有杀稻瘟素TM抗性基因标记的毕赤酵母表达载体中。纳米抗体和辅助蛋白均在AOX1甲醇诱导型启动子的控制下。3.2转化将单独的辅助蛋白PDI1,Kar2p,RPP0和经剪接的HAC1转化到巴斯德毕赤酵母菌株NRRLY-11430(ATCC编号76273)中。转化体在含有杀稻瘟素TM的选择性培养基上生长。分离单个克隆,随后用纳米抗体B转化。3.3确定单个克隆的表达产率如实施例1.5中所述进行表达分析。通过考马斯染色的SDS-PAGE光密度测定扫描测量进行蛋白质的相对定量(表3)。只有共表达经剪接的HAC1辅助蛋白的克隆显示纳米抗体B的产率大大增加,这再次表明辅助蛋白经剪接的HAC1的增强表达最有效地提高了产率。表3:具有和不具有增强的辅助蛋白表达的纳米抗体B的表达产率。使用SDS-PAGE/考马斯染色评估产率,并使用Imagequant软件(GEHealthcare)进行条带量的定量。实施例4:当增强单独辅助蛋白的表达时,评估良好表达的纳米抗体C的产率纳米抗体C是由重链美洲驼抗体的两个序列优化的可变结构域组成的二价纳米抗体。纳米抗体C中的亚基不是VHH1型免疫球蛋白单可变结构域。C端亚基与人血清白蛋白(HSA)结合。使用35G/S接头使亚基头尾融合。如实施例3中所述克隆单独的辅助蛋白PDI1,Kar2p,RPP0和经剪接的HAC1,并转化到巴斯德毕赤酵母菌株NRRLY-11430中。转化体在含有杀稻瘟素TM的选择性培养基上生长。分离单个克隆,随后用纳米抗体C转化。如实施例1.5中所述进行表达分析。通过考马斯染色的SDS-PAGE光密度测定扫描测量进行蛋白质的相对定量(表4)。只有共表达经剪接的HAC1辅助蛋白的克隆显示出纳米抗体C的产率显著增加。这再次表明辅助蛋白经剪接的HAC1的增强表达对于提高纳米抗体产率是最有效的。这说明辅助蛋白经剪接的HAC1的增强表达还可以进一步增加良好表达的纳米抗体的产率。表4:具有和不具有增强的辅助蛋白表达的纳米抗体C的表达产率。使用SDS-PAGE/考马斯染色评估产率,并使用Imagequant软件(GEHealthcare)进行条带量的定量。实施例5:当增强辅助蛋白经剪接的HAC1的表达时,评估不同纳米抗体(纳米抗体D,纳米抗体E,纳米抗体F,纳米抗体G和纳米抗体H)的表达产量纳米抗体D是由重链美洲驼抗体的两个序列优化的可变结构域组成的二价纳米抗体。纳米抗体D中的N端亚基是VHH1型免疫球蛋白单可变结构域,并且特异结合c-Met,而C端亚基结合人血清白蛋白(HSA)。亚基使用9G/S接头头尾融合并含有C端Flag3-His6表位标签。纳米抗体D的序列(SEQIDNO:50)描述于表A-1中。纳米抗体E是由重链美洲驼抗体的三个序列优化的可变结构域组成的三价纳米抗体。纳米抗体E中的C端亚基是VHH1型免疫球蛋白单可变结构域,而中心亚基结合人血清白蛋白(HSA)。亚基使用G/S接头头尾融合并含有C端Flag3-His6表位标签。纳米抗体E的序列(SEQIDNO:51)描述于表A-1中。纳米抗体F是由重链美洲驼抗体的三个序列优化的可变结构域组成的三价纳米抗体。纳米抗体F中的C端亚基是VHH1型免疫球蛋白单可变结构域。纳米抗体F中的亚基不与人血清白蛋白(HSA)结合。亚基使用35G/S接头头尾融合并含有C端Flag3-His6表位标签。纳米抗体F的序列(SEQIDNO:52)描述于表A-1中。纳米抗体G和H是由重链美洲驼抗体的四个序列优化的可变结构域组成的四价纳米抗体。纳米抗体G和H中的C端亚基是VHH1型免疫球蛋白单可变结构域,而中心亚基之一结合人血清白蛋白(HSA)。使用35G/S接头使亚基头尾融合。纳米抗体G和H的序列(分别为SEQIDNO:53和SEQIDNO:54)描述于表A-1中。纳米抗体I是特异性结合TNF的单价纳米抗体。纳米抗体I基本上由一个序列优化的可变结构域组成,其进一步包含一个丙氨酸残基作为C端延伸。纳米抗体I不是VHH1型免疫球蛋白单可变结构域。纳米抗体I的序列(SEQIDNO:55)描述于表A-1中。如3.1中所述克隆单独的辅助蛋白经剪接的HAC1,并转化到巴斯德毕赤酵母菌株NRRLY-11430中。转化体在含有杀稻瘟素TM的选择性培养基上生长。分离单个克隆,随后用纳米抗体D,E,F,G或H转化。如实施例1.5中所述进行表达分析。通过考马斯染色的SDS-PAGE光密度测定扫描测量进行蛋白质的相对定量(表5)。增强经剪接的HAC1辅助蛋白的表达提高了所有纳米抗体的产率。这再次表明增强辅助蛋白经剪接的HAC1的表达有效地提高了纳米抗体的产率。表5:具有和不具有增强的经剪接的HAC1辅助蛋白表达的纳米抗体D,E,F,G或H的表达产率。使用SDS-PAGE/考马斯染色评估产率,并使用Imagequant软件(GEHealthcare)进行条带量的定量。表表A-1.免疫球蛋白单可变结构域序列表A-2.筛选的用于提高纳米抗体A表达的辅助蛋白表A-3:在基因组DNAPCR中使用的引物,用于鉴定对巴斯德毕赤酵母中纳米抗体A的表达产率具有积极作用的辅助蛋白质表A-4:结合TNF的纳米抗体I的CDR和框架序列。CDR1,CDR2和CDR3根据Kontermann,2010确定。SEQIDNOFR157DVQLVESGGGVVQPGGSLRLSCTASCDR158GFTFSTADMGFR259WFRQAPGKGREFVACDR260RISGIDGTTYFR361YDEPVKGRFTISRDNSKNTVYLQMNSLRPEDTALYYCRSCDR362PRYADQWSAYDYFR463WGQGTLVTVSS当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1