改进的分化方法与流程
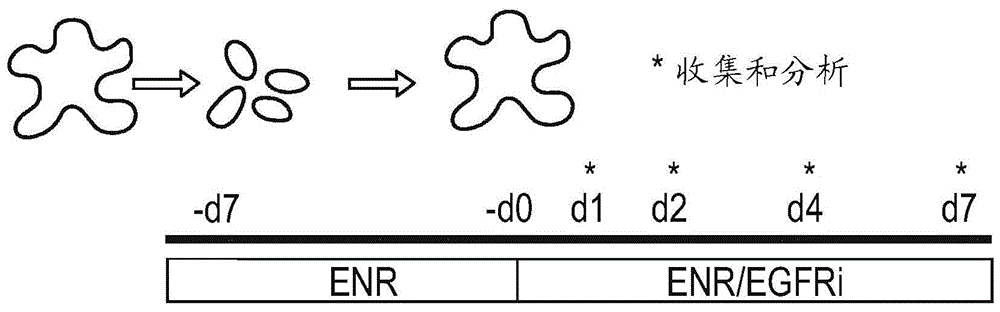
本发明属于细胞培养基和细胞培养方法领域,特别是用于分化祖细胞(例如人上皮干细胞)的培养基和培养方法领域。
背景技术:
:人们对用于分化祖细胞的培养基和培养方法非常感兴趣。祖细胞及其分化子代可用于细胞分析、药物筛选和毒性测定。祖细胞及其分化子代还显示出用于基于细胞的疗法的前景,如在用于治疗受损组织的再生医学方面。此外,有效的细胞培养基对于提供并维持细胞群以用于研究目的是重要的。肠内分泌细胞(eec)是罕见的激素分泌细胞,其可以从lgr5干细胞产生(koo和clevers(2014)gastroenterology147:289-302)。最常见的是,亚型基于其分泌的激素进行区分,并且包括生长激素抑制素+(sst)树突状细胞、胃抑制蛋白+(gip)k-细胞、分泌素+(sct)s-细胞、胆囊收缩素(cck)i-细胞、胰高血糖素样蛋白+(glp-1)l-细胞、神经降压素+(nts)n-细胞和产血清素肠嗜铬细胞(gunawardene等人(2011)internationaljournalofexperimentalpathology92:219-231)。然而,单个eec可以表达不同水平的多种激素,强调了高水平的异质性(egerod等人(2012)naturecellbiology14:1099-1104)。虽然eec被认为在控制肠功能和生物代谢的各个方面起着至关重要的作用,但它们的稀缺性已经成为他们深入研究和开发的障碍。已描述了用于分化来源于数种组织(例如胰腺、结肠、肠隐窝和胃)的祖细胞的方法(参见wo2010/090513、wo2012/014076、wo2012/168930和wo2015/173425)。用于诱导肠上皮细胞、杯状细胞和帕内特细胞分化的方法是已知的。需要改进的培养基和培养方法,其导致更高的祖细胞至eec命运的分化效率。发明概述本发明提供了用于分化祖细胞的方法,其中所述方法包括:在分化培养基中培养所述细胞,所述分化培养基包含基础培养基并且还包含一种或多种egfr通路抑制剂、notch抑制剂和一种或多种wnt抑制剂。本发明还提供了分化培养基,所述分化培养基包含基础培养基并且还包含一种或多种egfr通路抑制剂、notch抑制剂和一种或多种wnt抑制剂。本发明还提供了用于分化肠祖细胞以获得肠内分泌细胞中富含的肠细胞群的方法,其中所述方法包括:在本发明的分化培养基中培养所述肠祖细胞。本发明还提供了用于培养上皮干细胞、优选地以获得类器官的方法,其中所述方法包括:在扩增培养基存在下,培养与细胞外基质接触的一种或多种上皮干细胞;和在本发明的分化培养基中培养一种或多种扩增的上皮干细胞。本发明还提供了用于培养肠上皮干细胞优选以获得分化的肠类器官的方法,并且其中所述方法包括:在扩增培养基存在下,培养与细胞外基质接触的一种或多种肠上皮干细胞;优选地,其中所述扩增培养基包含基础培养基,并且还包含:受体酪氨酸激酶配体(例如egf)、bmp抑制剂(例如头蛋白(noggin))和wnt激动剂(例如rspondin),并且任选地,丙戊酸和gsk-3抑制剂(例如chir99021);以及随后在本发明的分化培养基存在下,培养与细胞外基质接触的一种或多种扩增的肠上皮干细胞。本发明还提供了通过本发明的方法可获得的或获得的类器官。本发明还提供了类器官,其中至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或至少99%的细胞表达肠内分泌细胞标志物。本发明还提供了本发明的类器官或来源于所述类器官的细胞,其用于医学中的用途。本发明还提供了本发明的类器官或来源于所述类器官的细胞在下述中的用途:药物发现筛选;毒性测定;诊断;组织胚胎学、细胞谱系和分化途径的研究;鉴别导致各自激素释放的化学和/或神经元信号的研究;包括重组基因表达的基因表达研究;组织损伤和修复中涉及的机制研究;炎性和传染性疾病的研究;发病机制研究;或者细胞转化和癌症病因机制的研究。本发明还提供了药物制剂,其包含一种或多种egfr通路抑制剂、notch抑制剂和一种或多种wnt抑制剂。用于筛选治疗性或预防性药物或化妆品的方法,其中所述方法包括:使本发明的分化的类器官与候选分子(或候选分子库)接触,评估所述类器官的任何影响(例如细胞的任何变化,如增殖的减少或丧失、形态变化和/或细胞死亡)或类器官的变化(例如器官大小或运动性);将导致所述影响的候选分子鉴定为潜在的药物或化妆品;并且任选地,将所述候选分子制备为药物或化妆品。本发明还提供了用于诱导lgr5+干细胞静止的方法,其中所述方法包括:用一种或多种egfr通路抑制剂处理细胞。本发明还提供了通过用于诱导本发明的lgr5+祖干细胞静止的方法获得的静止干细胞群,其中所述细胞表达lgr5和lef1并且不表达ki67和m期标志物磷酸化-组蛋白h4。本发明还提供了获得富含eec的细胞群的方法,其中所述方法包括在本发明的分化培养基中培养细胞群。本发明还提供了获得富含分泌glp1的eec的细胞群的方法,其中所述方法包括在本发明的分化培养基中培养细胞群,其中所述分化培养基包含bmp抑制剂。本发明还提供了获得富含分泌分泌素的eec的细胞群的方法,其中所述方法包括在本发明的分化培养基中培养细胞群,其中所述分化培养基包含bmp通路激活剂。本发明还提供了用于治疗或预防糖尿病或相关疾病或病症的方法的bmp抑制剂,其中所述方法包括向有需要的受试者施用治疗有效量的bmp抑制剂。本发明还提供了用于治疗胃酸过多或肥胖的方法的bmp激活剂,其中所述方法包括向有需要的受试者施用治疗有效量的bmp激活剂。发明详述先前已在wo2010/090513、wo2012/014076、wo2012/168930和wo2015/173425中描述了用于分化来自多种组织的祖细胞的方法。已经描述了用于促进肠祖细胞向肠细胞、杯状细胞或潘氏细胞命运分化的方法。这些总结在图11中。然而,先前未描述用于增强肠祖细胞分化为eec命运的方法和培养基。本发明人惊奇地发现,在分化培养基中wnt抑制剂、egfr通路抑制剂和notch抑制剂的组合增强了祖细胞分化为eec命运(参见实施例2)。eec占体内肠上皮细胞的不到1%。然而,本发明人的方法和分化培养基能够产生其中约50%的细胞是eec的类器官。本发明人还惊奇地发现,通过调节bmp信号传导通路,它们可以调节eec表型,特别是与表达水平和激素分泌有关(参见实施例5)。本发明人已经表明,某些eec表型可以通过激活或抑制bmp信号传导来获得。本发明人还表明,这些方法在体内可以是有效的,因此在疗法中为bmp激活剂和抑制剂提供了有希望的新用途。本发明人还惊奇地发现,egfr通路抑制剂在培养基中的存在促进了lgr5+干细胞的静止(参见实施例1)。特别地,本发明人发现了一种新的静止状态,其中维持了干细胞潜能但暂停了增殖。先前不知道可以分开干细胞潜能和增殖。wnt抑制剂本发明的分化培养基包含wnt抑制剂。可以使用任何合适的wnt抑制剂。当被激活时,wnt信号转导通路通常阻止β-连环蛋白降解并增强β-连环蛋白介导的信号转导。该通路由当包含卷曲蛋白受体和lrp5/6的细胞表面wnt受体复合物被激活时发生的一系列事件所限定,所述激活通常通过细胞外信号转导分子(如wnt家族成员)进行。这导致蓬乱蛋白家族蛋白的激活,其抑制降解细胞内β-连环蛋白的蛋白质的破坏复合物。破坏复合物由招募酪蛋白激酶ck1α、δ和ε以及gsk-3的结构组分(包括apc和轴蛋白)形成。破坏复合物被认为使β-连环蛋白磷酸化并将其暴露于泛素连接酶β-trcp。然后β-连环蛋白的泛素化导致其在蛋白酶体中降解。β-连环蛋白的主要效应功能在细胞核中,其中它通过与多种转录因子的相互作用调节转录,所述转录因子包括tcf/lef家族转录因子(例如tcf-1、tcf-3、tcf-4和lef1)。wnt通路受到高度调控。例如,当rspondin与其受体(lgr4、lgr5和/或lgr6)结合时,wnt信号转导被增强。然而,已经显示两种跨膜e3泛素连接酶rnf43和znrf3从细胞表面移除rspondin受体(例如lgr4、lgr5和/或lgr6)(参见例如delau等人,2016)。rspondin是脊椎动物特异性的wnt增强剂。此外,可以通过dapper家族蛋白(例如dapper1和dapper3)抑制蓬乱蛋白家族蛋白与卷曲蛋白受体的结合。此外,破坏复合物的活性被认为部分受apc、轴蛋白和gsk-3的磷酸化状态的调节。例如,磷酸酶(例如丝氨酸/苏氨酸磷酸酶,如pp1、pp2c或pp2a)对apc或轴蛋白的去磷酸化可以抑制β-连环蛋白的降解。此外,激酶(例如p38mapk、pka、pkb、pkc、p90rsk或p70s6k)对gsk-3的磷酸化可以抑制gsk-3的活性,并因此抑制β-连环蛋白的降解。破坏复合物的稳定性被认为部分受到两种parp,端锚聚合酶1和2的调节。这些端锚聚合酶对轴蛋白的聚(adp-核糖基)化和自身聚(adp-核糖基)化可以促进破坏复合物的去寡聚化。在细胞核中,蓬乱蛋白家族蛋白质可以与组蛋白脱乙酰酶sirt1形成复合物,所述组蛋白脱乙酰酶sirt1支持wnt靶基因的转录。被认为对wnt分泌关键的蛋白质是多通道膜蛋白porcupine(porc),其丧失导致wnt在内质网中积聚。可以在许多水平上抑制wnt信号转导通路,并且在voronkov和krauss(2013)currentpharmaceuticaldesign19:634664中详细综述了wnt抑制剂。wnt抑制剂被限定为抑制细胞或细胞群中tcf/lef介导的转录的试剂。因此,适用于本发明的wnt抑制剂包括:(1)wnt分泌抑制剂(例如porc的抑制剂,如lgk974、iwp-1或iwp-2),(2)wnt或rspondin与它们各自受体之间相互作用的竞争性和非竞争性抑制剂(例如omp-18r5、omp54f28),(3)促进wnt受体复合物的组分降解的因子如lrp(例如氯硝柳胺),和促进rspondin受体降解的因子如znrf3和/或rnf43,或者激活znrf3和/或rnf43的因子,(4)蓬乱蛋白家族蛋白抑制剂,如减少蓬乱蛋白家族蛋白与卷曲蛋白受体和/或破坏复合物组分的结合的抑制剂(例如dapper家族蛋白、fj9、舒林酸、3289-8625,j01-017a、nsc668036)或者下调蓬乱蛋白家族蛋白的表达的抑制剂(例如氯硝柳胺),(5)促进破坏复合物活性的因子,包括(a)使破坏复合物的组分(如轴蛋白和/或apc)去磷酸化的磷酸酶(例如pp1、pp2a和/或pp2c)的抑制剂(例如冈田酸或变构霉素(tautomycin))和(b)使gsk-3磷酸化的激酶(例如p38mapk、pka、pkb、pkc、p90rsk或p70s6k)的抑制剂(例如sb239063、sb203580或rp-8-br-camp),(6)破坏复合物去寡聚化的抑制剂,如端锚聚合酶1和/或2的抑制剂(例如xav939、iwr1、jw74、jw55、2-[4-(4-氟苯基)哌嗪-1-基]-6-甲基嘧啶-4(3h)-酮或pj34),以及(7)β-连环蛋白靶基因表达的抑制剂,包括β-连环蛋白:tcf/lef转录复合物的抑制剂,如破坏β-连环蛋白:tcf-4复合物的抑制剂(例如icrt3、cgp049090、pkf118310、pkf115-584、ztm000990、pnu-74654、bc21、icrt5、icrt14或fh535)和组蛋白脱乙酰酶sirt1的抑制剂(例如cambinol)。本发明的分化培养基包含wnt抑制剂。可以如以上(1)-(7)中所述使用任何合适的wnt抑制剂。例如,在一个优选的实施方案中,wnt抑制剂是wnt分泌抑制剂,如porc抑制剂(例如选自iwp-2、iwp-1和lgk974)。在另一个优选的实施方案中,wnt抑制剂是β-连环蛋白靶基因表达的抑制剂,例如β-连环蛋白:tcf/lef转录复合物的抑制剂或组蛋白脱乙酰酶sirt1的抑制剂(例如cambinol)。在一些实施方案中,β-连环蛋白:tcf/lef转录复合物的抑制剂是破坏β-连环蛋白:tcf-4复合物的抑制剂,例如选自以下的抑制剂:icrt3、cgp049090、pkf118310、pkf115-584、ztm000990、pnu-74654、bc21、icrt5、icrt14和fh535。在一些实施方案中,wnt抑制剂选自:iwp-2、omp-18r5、omp54f28、lgk974、3289-8625、fj9、nsc668036、iwr1和xav939。在一些实施方案中,wnt抑制剂选自:icrt3、pfk115-584、cgp049090、icrt5、icrt14和fh535。在一些实施方案中,wnt抑制剂是以下表1中列出的化合物之一。表1-wnt抑制剂在一些实施方案中,本发明的分化培养基包含表1中列出的任何wnt抑制剂中的一种或多种。优选向培养基中添加一定量的wnt抑制剂,如在相同细胞类型中所评估的,相对于所述分子不存在的情况下所述wnt活性的水平,所述量有效抑制细胞中至少10%,更优选至少20%,更优选至少30%,更优选至少50%,更优选至少70%,更优选至少90%,更优选100%的wnt活性。如技术人员已知的,可以通过测量wnt的转录活性来确定wnt活性,例如通过ptopflash和pfopflashtcf荧光素酶报告基因构建体(korinek等人,1997.science275:1784-1787)。因此,技术人员可以使用本领域已知的测定容易地鉴定新的wnt抑制剂。在一些实施方案中,本发明的分化培养基包含以下浓度的wnt抑制剂:0.01-150μm、0.1-150μm、0.5-100μm、0.1-100μm、0.5-50μm、1-100μm或10-80μm、1-20μm或者1-5μm。在一些实施方案中,本发明的分化培养基包含以下浓度的iwp-2:0.01-150μm、0.1-100μm、0.5-50μm、1-20μm或者1-5μm。例如,在一些实施方案中,本发明的分化培养基包含浓度为约1.5μm的iwp-2。在一些实施方案中,分化培养基不包含结合并激活wnt受体复合物的wnt激动剂,包括任何和所有wnt家族蛋白和rspondin。在其他实施方案中,分化培养基还包含wnt激动剂,如r-spondin1-4或其生物活性片段或变体。如上所述,r-spondin增强了细胞表面受体处的wnt信号传导。本发明人已经表明,从eec分化培养基中去除r-spondin降低了eec分化的效率(参见实施例5)。假设可能需要一些wnt信号传导将细胞导向分泌(而不是吸收)谱系。因此,在一些实施方案中,分化培养基包含wnt激动剂(特别是r-spondin)和wnt抑制剂。例如,在一些实施方案中,分化培养基包含r-spondin和porc抑制剂,如iwp-2。在一些实施方案中,r-spondin的使用终浓度为1至1000ng/ml、50至1000ng/ml或100至1000ng/ml。在一些实施方案中,r-spondin的使用终浓度为0.1至100μg/ml、0.1至50μg/ml、0.1至20μg/ml、0.1至10μg/ml、0.1至5μg/ml、0.5至100μg/ml、0.5至50μg/ml、0.5至20μg/ml、0.5至10μg/ml、0.5至5μg/ml、1至10μg/ml或1至5μg/ml。在一些实施方案中,r-spondin的使用终浓度为至少1ng/ml、至少50ng/ml、至少100ng/ml、至少500ng/ml或至少1μg/ml。在一些实施方案中,r-spondin的使用终浓度为约100ng/ml。在一些实施方案中,r-spondin的使用终浓度为约1μg/ml。egfr通路抑制剂本发明的分化培养基包含egfr通路抑制剂。可以使用如本文定义的任何合适的抑制剂。表皮生长因子受体(egfr),也称为erbb1或her1,是细胞外蛋白配体的表皮生长因子(egf)家族成员的细胞表面受体。egfr属于her受体家族,其包括四种相关蛋白(egfr(her1/erbb1)、erbb2(her2)、erbb3(her3)和erbb4(her4))。已知her受体通过与不同配体结合而被激活,所述配体包括egf、tgfa,结合肝素的egf样生长因子、双调蛋白、β细胞素和表皮调节素。在配体结合至受体的细胞外结构域后,受体形成功能活性二聚体(egfr-egfr(同二聚体)或egfr-her2、egfr-her3、egfr-her4(异二聚体))。二聚化诱导酪氨酸激酶结构域的激活,这导致受体在多个酪氨酸残基上的自身磷酸化。这导致募集一系列衔接子蛋白(如shc,grb2)并激活一系列细胞内信号传导级联以影响基因转录。已经很好地研究了介导egfr下游效应的通路,并且已经确定了三种主要的信号传导通路。第一通路涉及ras-raf-mapk通路,其中磷酸化的egfr通过grb2和shc衔接子蛋白募集鸟嘌呤-核苷酸交换因子,从而激活ras并随后刺激raf和map激酶通路以影响细胞增殖、肿瘤侵袭和转移。活化的ras激活raf激酶的蛋白激酶活性。raf激酶磷酸化并激活mek(也称为map2k或mapkk),所述mek磷酸化并激活map激酶(也称为erk,一种细胞外信号调控激酶)。第二通路涉及pi3k/akt通路,其通过激活核转录因子如nfkb来激活主要的细胞存活和抗凋亡信号。第三通路涉及jak/stat通路,所述jak/stat通路也参与激活与细胞存活相关的基因的转录。egfr激活还可能导致plcg磷酸化,随后磷脂酰肌醇4,5二磷酸(pip2)水解成肌醇1,4,5-三磷酸(ip3)和二酰基甘油(dag),导致蛋白激酶c(prkc)和camk激活。可以使用egfr抑制剂,如抗egfr单克隆抗体和小分子egfr酪氨酸激酶抑制剂。一些抗egfr抗体,如西妥昔单抗和帕尼单抗,结合至egfr单体的细胞外结构域,并通过内源配体竞争受体结合;以这种方式它们阻断配体诱导的受体激活。一些小分子egfr抑制剂,如厄洛替尼、吉非替尼和拉帕替尼,与atp竞争结合egfr的激酶结构域,这继而抑制egfr自身磷酸化和下游信号传导。可以使用一种或多种egfr通路抑制剂,例如2种、3种、4种或更多种。egfr通路抑制剂优选以如在相同细胞类型中评估,相对于在不存在所述分子的情况下所述egfr通路活性的水平,将细胞中egfr通路活性有效抑制至少10%、更优选至少20%、更优选至少30%、更优选至少50%、更优选至少70%、更优选至少90%、更优选100%的量加入培养基中。如技术人员所知,可以以多种方式测量egfr通路活性。例如,ghosh等人(2013)assayanddrugdevelopmenttechnologies11(1):44-51描述了用于监测egfr活性和抑制剂敏感性的测定。该特定测定涉及共价固定在磁珠上的肽底物。在激酶反应后,洗涤珠粒并使用针对磷酸化酪氨酸的hrp-缀合的一抗通过化学荧光检测肽的磷酸化。测量的荧光强度与底物磷酸化成正比,而底物磷酸化又与egfr激酶活性成比例。该测定还可用于筛选egfr通路中其他激酶(例如ras、raf、mek或erk)的抑制剂。用于测定激酶活性的可替代方法包括检测来自p32-标记的atp的末端磷酸根的掺入。因此,技术人员可以使用本领域已知的测定容易地鉴定新的egfr通路抑制剂。在一些实施方案中,egfr通路抑制剂是egfr抑制剂,其抑制egfr激酶活性至少10%、更优选至少20%、更优选至少30%、更优选至少50%、更优选至少70%、更优选至少90%、更优选100%。在一些实施方案中,egfr通路抑制剂是ras抑制剂,其抑制ras激酶活性至少10%、更优选至少20%、更优选至少30%、更优选至少50%、更优选至少70%、更优选至少90%、更优选100%。在一些实施方案中,egfr通路抑制剂是raf抑制剂,其抑制raf激酶活性至少10%、更优选至少20%、更优选至少30%、更优选至少50%、更优选至少70%、更优选至少90%、更优选100%。在一些实施方案中,egfr通路抑制剂是mek抑制剂,其抑制mek激酶活性至少10%、更优选至少20%、更优选至少30%、更优选至少50%、更优选至少70%、更优选至少90%、更优选100%。在一些实施方案中,egfr通路抑制剂是erk抑制剂,其抑制erk激酶活性至少10%、更优选至少20%、更优选至少30%、更优选至少50%、更优选至少70%、更优选至少90%、更优选100%。在一些实施方案中,egf以小于1mm的浓度存在于分化培养基中。在优选的实施方案中,egf不存在于分化培养基中。在一些实施方案中,egfr通路抑制剂是egfr抑制剂,如吉非替尼(santacruzbiotechnology)、ag-18、ag-490(酪氨酸磷酸化抑制剂b42)、ag-1478(酪氨酸磷酸化抑制剂ag-1478)、az5104、azd3759、布加替尼、厄洛替尼、西妥昔单抗、cl-387785(eki-785)、cnx-2006、埃克替尼、耐妥珠单抗、奥希替尼(azd9291)、osi-420、pd153035hcl、pd168393、培利替尼(ekb-569)、罗西替尼(co-1686、avl-301)、tak-285、酪氨酸磷酸化抑制剂9、凡德他尼、whi-p154、wz3146、wz4002、wz8040、帕尼单抗、札鲁目单抗、尼妥珠单抗或马妥珠单抗。在一些实施方案中,egfr抑制剂结合至egfr单体的细胞外结构域并竞争egf的受体结合。在一些实施方案中,egfr抑制剂与atp竞争结合egfr的激酶结构域。可以使用一种或多种egfr抑制剂,例如2种、3种、4种或更多种。在一些实施方案中,egfr通路抑制剂是egfr和erbb-2抑制剂,如阿法替尼(selleckchem)、二马来酸阿法替尼、ac480(bms-599626)、aee788(nvp-aee788)、ast-1306、卡奈替尼、cudc-101、达克替尼、拉帕替尼、来那替尼、波齐替尼(hm781-36b)、沙普替尼(azd8931)或瓦尼替尼。可以使用一种或多种egfr和erbb-2抑制剂,例如2种、3种、4种或更多种。在一些实施方案中,egfr通路抑制剂是ras-raf-mapk通路抑制剂。在一些实施方案中,egfr通路抑制剂是pi3k/akt通路抑制剂。在一些实施方案中,egfr通路抑制剂是jak/stat通路抑制剂。在一些实施方案中,egfr通路抑制剂是raf抑制剂,如gw5074、zm336372、nvp-bhg712、tak-632、达拉非尼(gsk2118436)、索拉非尼、甲苯磺酸索拉非尼、plx-4720、az628、cep-32496或维罗非尼(plx4032,rg7204)。在一些实施方案中,egfr通路抑制剂是mek抑制剂,如pd0325901(sigmaaldrich)。在一些实施方案中,egfr通路抑制剂是erk抑制剂,如sch772984(selleckchem)。在一些实施方案中,egfr通路抑制剂的使用浓度是0.01-200μm、0.01-100μm、0.1-50μm、0.1-20μm、1-100μm、1-50μm、1-30μm、5-100μm、5-50μm或5-20μm。例如,在一些实施方案中,分化培养基包含:(i)浓度为约5μm的吉非替尼,(ii)浓度为约10μm的阿法替尼,(iii)浓度为约5μm的pd0325901或(iv)浓度为约10μm的sch772984。notch抑制剂分化培养基包含notch抑制剂。可以使用任何合适的notch抑制剂。notch是跨膜表面受体,其可以通过多种蛋白水解切割而被激活,其中一种蛋白水解切割是通过被称为γ-分泌酶的具有蛋白酶活性的蛋白复合物进行的切割。γ-分泌酶是在膜内发挥其切割活性的蛋白酶。γ-分泌酶是多组分酶,并且由至少四种不同的蛋白(即早老素(早老素1或2)、呆蛋白(nicastrin)、pen-2和aph-1)组成。早老素是γ-分泌酶的催化中心。在配体结合时,notch受体经历允许胞外域通过是金属蛋白酶的adam蛋白酶的作用脱落的构象变化。紧接着是γ-分泌酶复合物的作用,其导致notch细胞内结构域(nicd)的释放。nicd易位至细胞核,在那它与csl(c启动子结合因子/重组信号序列结合蛋白jκ/无毛抑制因子/lagl(c-promoter-bindingfactor/recombinantsignal-sequencebindingproteinjκ/supressor-of-hairless/lagl))相互作用。nicd的结合将csl从转录抑制因子转变为激活因子,这导致notch靶基因的表达。在一些实施方案中,notch抑制剂是能够减少配体介导的notch激活的抑制剂(例如经由notch的显性阴性配体或者经由显性阴性notch或者经由能够至少部分阻断notch配体与notch之间的相互作用的抗体)或adam蛋白酶的抑制剂。在一些实施方案中,notch抑制剂是γ分泌酶抑制剂,例如dapt、二苯并氮杂(dbz)、苯并二氮杂(bz)或ly-411575。可以使用一种或多种notch抑制剂,例如2、3、4种或更多种。在一些实施方案中,以以下浓度使用notch抑制剂(例如dapt):0.001-200mm、0.01-100mm、0.1-50mm、0.1-20mm、0.5-10mm或0.5-5mm。例如,在一些实施方案中,分化培养基包含浓度为约1mm的dapt。基础培养基用于本发明的方法的分化培养基包含基础培养基。基础培养基是用于动物或人细胞的任何合适的基础培养基,其受本文提供的限制。用于动物或人细胞培养的基础培养基通常含有大量成分,这些成分是支持培养细胞的维持所必需的。考虑以下公开内容,技术人员可以容易地配制合适的成分组合。如文献中以及以上更加详细描述的,用于本发明的基础培养基将通常包含营养液,所述营养液包含标准细胞培养成分,如氨基酸、维生素、脂质补充剂、无机盐、碳能源和缓冲剂。在一些实施方案中,还将培养基补充有一种或多种标准细胞培养成分,例如选自氨基酸、维生素、脂质补充剂、无机盐、碳能源和缓冲剂的细胞培养成分。技术人员将从公知常识中理解培养基的类型,其可以在本发明的分化培养基中被用作基础培养基。潜在合适的细胞培养基可商购获得,包括但不限于杜尔贝科改良伊格尔培养基(dulbecco'smodifiedeaglemedia,dmem)、最小必需培养基(mem)、敲除-dmem(ko-dmem)、格拉斯哥最小必需培养基(glasgowminimalessentialmedium,g-mem)、伊格尔基础培养基(basalmediumeagle,bme)、dmem/ham'sf12、改良型dmem/ham'sf12、iscove改良杜尔贝科培养基和和最小必需培养基(mem)、ham'sf-10、ham'sf-12、培养基199以及rpmi1640培养基。例如,基础培养基可以选自补充有谷氨酰胺、胰岛素、青霉素/链霉素和转铁蛋白的dmem/f12和rpmi1640。在进一步优选的实施方案中,使用改良型dmem/f12或改良型rpmi,其针对无血清培养进行了优化,并且已经包含了胰岛素。在这种情况下,所述改良型dmem/f12或改良型rpmi培养基优选补充有谷氨酰胺和青霉素/链霉素。补充有n2和b27的addmem/f12(invitrogen)也是优选的。优选地,基础培养基是改良型dmem/f12。更优选地,基础培养基包含改良型dmem/f12、谷氨酰胺和b27。在一些实施方案中,基础培养基包含改良型dmem/f12、hepes、青霉素/链霉素、谷氨酰胺、n-乙酰半胱氨酸和b27。在一些实施方案中,基础培养基包含补充有青霉素/链霉素、10mmhepes、glutamax、b27(全部来自lifetechnologies,carlsbad,ca)和约1mmn-乙酰半胱氨酸(sigma)的改良型dmem/f12或者由其组成。另外优选地,所述基础培养基补充有纯化的、天然的、半合成的和/或合成的生长因子,并且不包含不确定的组分(如胎牛血清或小牛血清)。多种不同的血清替代物制剂可商购获得并且是技术人员已知的。当使用血清替代物时,根据常规技术,它可以以培养基体积的约1%至约30%被使用。如本文其它地方所述,本发明中使用的分化培养基可以包含血清,或者可以是无血清和/或无血清替代物的。培养基和细胞制备优选是符合fda对生物制剂产品所要求的标准并确保产品一致性的gmp过程。通常将在去离子蒸馏水中配制本发明的分化培养基。在使用前,通常将本发明的分化培养基进行灭菌以防止污染,例如通过紫外线、加热、辐射或过滤。可以冷冻分化培养基(例如在-20℃或-80℃下)以存储或运输。培养基可以含有一种或多种抗生素以防止污染。培养基可以具有低于0.1个内毒素单位/ml的内毒素含量,或者可以具有低于0.05个内毒素单位/ml的内毒素含量。用于确定培养基的内毒素含量的方法在本领域是已知的。优选的基础培养基是确定的合成培养基,其用基于碳酸盐的缓冲剂将ph缓冲为7.4(优选ph为7.2-7.6或者至少7.2且不高于7.6),同时在包含5%至10%co2或者至少5%且不超过10%co2,优选5%co2的环境中培养细胞。在一些实施方案中,提供的分化培养基具有准备用于分化方法的基础培养基。在其它实施方案中,提供的分化培养基不含基础培养基,例如作为培养基补充剂,并且可以在用于分化方法之前添加基础培养基(或其它组分)。因此,提供了本文所述的任何分化培养基,其中不存在基础培养基或者其中基础培养基仅是任选组分。另外的因子先前已经显示许多因子在某些情况下增强祖细胞的分化。在一些实施方案中,这些因子中的一种或多种包含在本发明的分化培养基中。这些因子包括但不限于p38抑制剂、tgf-β抑制剂、胃泌素、糖皮质激素、受体酪氨酸激酶配体、bmp通路激活剂、hedgehog激活剂、hedgehog抑制剂、mtor信号传导调节剂、gsk-3抑制剂、chir99021激动剂、ap-1刺激剂、毒蕈碱乙酰胆碱受体激动剂、卡巴胆碱激动剂和camp通路激活剂。在一些实施方案中,这些因子选自p38抑制剂、tgf-β抑制剂、胃泌素、糖皮质激素、受体酪氨酸激酶配体、bmp通路激活剂、bmp通路抑制剂、hedgehog激活剂、hedgehog抑制剂、mtor信号传导调节剂、gsk-3抑制剂、chir99021激动剂、ap-1刺激剂、毒蕈碱乙酰胆碱受体激动剂、卡巴胆碱激动剂和camp通路激活剂。在一些实施方案中,这些因子选自胃泌素、糖皮质激素、受体酪氨酸激酶配体、bmp通路激活剂、bmp通路抑制剂、hedgehog激活剂、hedgehog抑制剂、mtor信号传导调节剂、gsk-3抑制剂、chir99021激动剂、ap-1刺激剂、毒蕈碱乙酰胆碱受体激动剂、卡巴胆碱激动剂和camp通路激活剂。p38抑制剂在本发明的一些实施方案中,分化培养基还包含p38抑制剂,意指任何直接或间接负调控p38信号传导的抑制剂。在一些实施方案中,根据本发明的抑制剂结合至并降低p38(gi编号1432)的活性。p38蛋白激酶是丝裂原活化蛋白激酶(mapk)家族的一部分。mapk是丝氨酸/苏氨酸特异性蛋白激酶,其响应于细胞外刺激如环境应激和炎性细胞因子,并调控各种细胞活性,如基因表达、有丝分裂、分化、增殖和细胞存活/凋亡。p38mapk以α、β、β2、γ和δ同种型存在。p38抑制剂是结合至并降低至少一种p38同种型活性的试剂。用于确定物质是否是p38抑制剂的各种方法是已知的,并且可以与本发明结合使用。实例包括:磷酸-特异性抗体检测thr180/tyr182处的磷酸化,其提供了公认的细胞p38激活或抑制的量度;生化重组激酶测定;肿瘤坏死因子α(tnfα)分泌测定;和p38抑制剂的discoverrx高通量筛选平台(参见http://www.discoverx.com/kinases/literature/biochemical/collaterals/drx_poster_p38%20kba.pdf)。还存在几种p38活性测定试剂盒(例如millipore,sigma-aldrich)。各种p38抑制剂在本领域中是已知的。在一些实施方案中,直接或间接负调控p38信号传导的抑制剂选自由以下组成的组:sb-202190、sb-203580、vx-702、vx-745、pd-169316、ro-4402257和birb-796。在一个实施方案中,根据本发明的p38抑制剂结合至其靶标并如通过细胞测试评估与对照相比降低其靶标的活性大于10%;大于30%;大于60%;大于80%;大于90%;大于95%;或大于99%。用于测量靶抑制的细胞测定的实例是本领域熟知的,如上所述。sb-203580可以以50nm至100μm、或100nm至50μm、或1μm至50μm的浓度添加至分化培养基。例如,sb-203580可以以约30μm添加至培养基。tgf-β抑制剂在一些实施方案中,分化培养基还包含tgf-β抑制剂。tgf-β信号转导涉及许多细胞功能,包括细胞生长、细胞命运和细胞凋亡。信号转导通常始于tgf-β超家族配体与ii型受体的结合,所述ii型受体招募并磷酸化i型受体。然后,i型受体磷酸化smad,smad作为细胞核中的转录因子发挥作用并调节靶基因的表达。先前在促进祖细胞分化中已经涉及了tgf-β抑制剂信号转导通路。例如,向肝脏外植体中添加tgf-β促进了体外胆分化(clotman等人(2005)genesdev.19(16):1849-54)。此外,先前已经表明,在分化培养基中包含tgf-β抑制剂可以抑制胆管细胞命运并引发细胞向更多肝细胞表型分化(参见wo2012/168930)。具体地,发现在分化培养基中包含tgf-β抑制剂(如a83-01)增强成熟肝细胞标志物的表达并增加肝细胞样细胞的数量。tgf-β超家族配体包含骨形态发生蛋白(bmp)、生长和分化因子(gdf)、抗缪勒氏管激素(amh)、激活素、nodal和tgf-β。通常,smad2和smad3被tgf-β/激活素通路中的alk4、5和7受体磷酸化。相比之下,smad1、smad5和smad8被磷酸化作为骨形态发生蛋白(bmp)通路的一部分。尽管通路之间存在一些交叉,但在本发明的上下文中,“tgf-β抑制剂”或“tgf-β信号转导的抑制剂”优选是tgf-β通路抑制剂,其经由smad2和smad3和/或经由alk4、alk5或alk7起作用。因此,在一些实施方案中,tgf-β抑制剂不是bmp抑制剂,即tgf-β抑制剂不是头蛋白。在一些实施方案中,除tgf-β抑制剂之外,还向培养基中添加bmp抑制剂。因此,tgf-β抑制剂可以是任何试剂,其降低tgf-β信号转导通路,优选经由smad2和/或smad3起作用的信号转导通路,更优选经由alk4、alk5或alk7起作用的信号转导通路的活性。存在多种破坏tgf-β信号转导通路的方法,这些方法是本领域已知的并且可以与本发明联合使用。例如,可以通过以下方式破坏tgf-β信号转导:通过小干扰rna策略抑制tgf-β的表达;抑制弗林蛋白酶(tgf-β激活蛋白酶);通过生理抑制剂抑制通路;用单克隆抗体中和tgf-β;用tgf-β受体激酶1(也被称为激活素受体样激酶alk5)、alk4、alk6、alk7或其它tgf-β相关受体激酶的小分子抑制剂进行抑制;例如通过过表达它们的生理抑制剂smad7或者通过使用硫氧还蛋白作为smad锚使smad无法激活来抑制smad2和smad3信号转导(fuchs,o.inhibitionoftgf-signallingforthetreatmentoftumormetastasisandfibroticdiseases.currentsignaltransductiontherapy,第6卷,第1期,2011年1月,第29-43(15)页)。用于确定物质是否是tgf-β抑制剂的多种方法是已知的,并且可以与本发明联合使用。例如,可以使用细胞分析,其中用包含人pai-1启动子或smad结合位点的报告基因构建体稳定转染细胞,驱动荧光素酶报告基因。可以将相对于对照组的荧光素酶活性的抑制用作化合物活性的程度(degouville等人,(2005)brjpharmacol.145(2):166-177)。因此,本领域技术人员可以容易地鉴定新的tgf-β抑制剂。根据本发明的tgf-β抑制剂可以是蛋白质、肽、小分子、小干扰rna、反义寡核苷酸、适配子或抗体。抑制剂可以是天然存在的或合成的。在一个实施方案中,tgf-β抑制剂是alk4、alk5和/或alk7的抑制剂。例如,tgf-β抑制剂可以结合并直接抑制alk4、alk5和/或alk7。可以被用于本发明的上下文的优选的小分子tgf-β抑制剂的实例包括但不限于以下表2中列出的小分子抑制剂。表2:靶向受体激酶的小分子tgf-β抑制剂在一些实施方案中,tgf-β抑制剂是小分子抑制剂,其任选地选自:a83-01、sb-431542、sb-505124、sb-525334、ly364947、sd-208和sjn2511。在一些实施方案中,在分化培养基中存在不多于一种tgf-β抑制剂。在其它实施方案中,在分化培养基中存在多于一种的tgf-β抑制剂(例如,2、3、4种或更多种)。在一些实施方案中,本发明的分化培养基包含表2中列出的任何抑制剂中的一种或多种。分化培养基可以包含一种抑制剂与列出的另一种抑制剂的任何组合。例如,培养基可以包含sb-525334或sd-208或a83-01;或者sd-208和a83-01。技术人员将会理解存在许多其它小分子抑制剂,将其主要设计为靶向其它激酶,但是在高浓度下也可以抑制tgf-β受体激酶。例如,sb-203580是p38map激酶抑制剂,其在高浓度(例如大约10μm或更高)下被认为抑制alk5。也可以在本发明的上下文中使用抑制tgf-β信号转导通路的任何此类抑制剂。在一些实施方案中,tgf-β抑制剂(例如a83-01)以以下浓度存在于分化培养基中:至少1nm,例如至少5nm、至少50nm、至少100nm、至少300nm、至少450nm或至少475nm。例如,tgf-β抑制剂(例如a83-01)以以下浓度存在于分化培养基中:1nm-200μm、10nm-200μm、100nm-200μm、1μm-200μm、10nm-100μm、50nm-100μm、50nm-10μm、100nm-1μm、200nm-800nm、350-650nm或约500nm。因此,在一些实施方案中,分化培养基包含浓度为约500nm的a83-01。在一些实施方案中,分化培养基不包含tgf-β抑制剂。在一些实施方案中,分化培养基不包含a83-01。胃泌素在一些实施方案中,本发明的分化培养基还包含胃泌素。在一些实施方案中,本发明的分化培养基包含浓度为0.01-500nm、0.1-100nm、1-100nm、1-20nm或5-15nm的胃泌素。例如,在一些实施方案中,本发明的分化培养基包含浓度为约10nm的胃泌素。糖皮质激素在一些实施方案中,分化培养基还包含糖皮质激素。糖皮质激素是一类皮质类固醇,其是一类类固醇激素。糖皮质激素是与糖皮质激素受体结合的皮质类固醇。皮质醇是最重要的人糖皮质激素。氢化可的松是皮质醇的合成形式。存在许多具有相关结构的其它合成的糖皮质激素(例如泼尼松、泼尼松龙、甲基泼尼松龙、地塞米松、倍他米松、曲安西龙、倍氯米松和醋酸氟氢可的松)。糖皮质激素具有不同的激活糖皮质激素受体的效力。这被称为糖皮质激素效力,并且通常与皮质醇比较测量。多种糖皮质激素的生物化学、药理学和作用机理在例如ceciltextbookofmedicine(1988),第128-130页,和thescienceandpracticeofpharmacy第20版(2000),第1363-1370页中进行了综述。在一些实施方案中,分化培养基包含糖皮质激素。可以使用任何合适的糖皮质激素。在一些实施方案中,糖皮质激素选自以下中的一种或多种:皮质醇、可的松、醋酸氢化可的松、盐酸氢化可的松、戊酸氢化可的松、泼尼松、泼尼松龙、甲基泼尼松龙、地塞米松、倍他米松、二丙酸倍他米松、戊酸倍他米松、曲安西龙、曲安奈德、倍氯米松、二丙酸倍氯米松、氟氢可的松、醋酸氟氢可的松、氟替卡松、氟替卡松缩丙酮、丙酸氟替卡松、氟尼缩松、布地奈德、氯倍他索、丙酸氯倍他索、双氟拉松、二醋酸双氟拉松、卤倍他索、丙酸卤倍他索、安西奈德、去羟米松、氟轻松醋酸酯(fluocinonide)、氟轻松醋酸酯缩丙酮(fluocinonideacetonide)、哈西奈德、莫米松、糠酸莫米松、氟氢缩松、泼尼卡酯、阿氯米松、二丙酸阿氯米松、地奈德、氟新诺龙(flucinolone)、氟新诺龙缩丙酮(flucinoloneacetonide)、丙吗卡因以及盐酸丙吗卡因。地塞米松是最有效的糖皮质激素之一,并且是用于本发明的分化培养基的优选的糖皮质激素。在一些实施方案中,糖皮质激素是具有与地塞米松相同或更高的糖皮质激素效力的任何糖皮质激素。倍他米松和醋酸氟氢可的松也是高效的糖皮质激素。因此,倍他米松是用于本发明分化培养基的优选的糖皮质激素。在一些实施方案中,糖皮质激素是具有与倍他米松相同或更高的糖皮质激素效力的任何糖皮质激素。因此,醋酸氟氢可的松是用于本发明分化培养基的优选的糖皮质激素。在一些实施方案中,糖皮质激素是具有与醋酸氟氢可的松相同或更高的糖皮质激素效力的任何糖皮质激素。在一些实施方案中,糖皮质激素是具有与皮质醇相同或更高的糖皮质激素效力的任何糖皮质激素。下面提供了用于本发明的分化培养基的示例性糖皮质激素的列表。所呈现的效力是指口服给药。糖皮质激素糖皮质激素效力皮质醇(氢化可的松)1可的松0.8泼尼松3.5-5泼尼松龙4甲基泼尼松龙5-7.5地塞米松25-80倍他米松25-30曲安西龙5醋酸氟氢可的松15在一些实施方案中,以以下浓度使用糖皮质激素(例如地塞米松):0.01-150μm、0.1-15μm、0.5-10μm或1-5μm。在优选的实施方案中,糖皮质激素是地塞米松。例如,在一些实施方案中,分化培养基包含浓度为约3μm的地塞米松。在一些实施方案中,分化培养基不包含糖皮质激素。受体酪氨酸激酶配体在一些实施方案中,本发明的分化培养基还包含一种或多种受体酪氨酸激酶配体。受体酪氨酸激酶(rtk)是多肽生长因子、细胞因子和激素的高亲和力细胞表面受体。rtk是细胞维持、生长和发育的关键调节剂,并且在许多类型的癌症的发展和进展中发挥关键作用。在本发明的上下文中,受体酪氨酸激酶配体是激活rtk的任何配体。许多受体酪氨酸激酶配体是促有丝分裂生长因子。因此,在一些实施方案中,分化培养基中的一种或多种受体酪氨酸激酶配体包含一种或多种促有丝分裂生长因子。大约存在20种不同的已知类型的rtk、包括rtki类(egf受体家族)(erbb家族)、rtkii类(胰岛素受体家族)、rtkiii类(pdgf受体家族)、rtkiv类(fgf受体家族)、rtkv类(vegf受体家族)、rtkvi类(hgf受体家族)、rtkvii类(trk受体家族)、rtkviii类(eph受体家族)、rtkix类(axl受体家族)、rtkx类(ltk受体家族)、rtkxi类(tie受体家族)、rtkxii类(ror受体家族)、rtkxiii类(ddr受体家族)、rtkxiv类(ret受体家族)、rtkxv类(klg受体家族)、rtkxvi类(ryk受体家族)、rtkxvii类(musk受体家族)。在一些实施方案中,一种或多种受体酪氨酸激酶配体包含针对这20类rtk中的一种或多种或者全部的配体。在一些实施方案中,一种或多种受体酪氨酸激酶配体包含针对rtkiv类(fgf受体家族)的配体。在一些实施方案中,一种或多种受体酪氨酸激酶配体包含针对rtkvi类(hgf受体家族)的配体。在一些实施方案中,一种或多种受体酪氨酸激酶配体包含针对rtkiv类(fgf受体家族)的配体和针对rtkvi类(hgf受体家族)的配体。因此,在一些实施方案中,在分化培养基中的一种或多种受体酪氨酸激酶配体选自成纤维细胞生长因子(fgf)和肝细胞生长因子(hgf)。在一些实施方案中,一种或多种受体酪氨酸激酶配体包含fgf和hgf。在一些实施方案中,仅一种受体酪氨酸激酶配体被包含在分化培养基中,例如,其中受体酪氨酸激酶配体选自fgf和hgf。fgf优选的是能够结合至fgfr2或fgfr4的fgf,并且优选是fgf19。可在本发明的分化培养基中使用三种或更多种,例如3、4、5或更多种受体酪氨酸激酶配体。如上所述,本发明人发现抑制egfr通路有助于肠内分泌细胞分化。因此,优选地,一种或多种受体酪氨酸激酶配体不包含激活egfr的配体(例如egf)。在一些实施方案中,分化培养基包含小于1mm的egf。bmp通路激活剂本发明人惊奇地发现,在eec分化培养基中包含bmp通路激活剂增加了分化的eec群中分泌分泌素的细胞的存在(参见图13和14)。因此,在一些实施方案中,分化培养基还包含bmp通路激活剂。在一些实施方案中,分化培养基不包含bmp通路抑制剂(例如头蛋白)。在一些实施方案中,分化培养基还包含bmp通路激活剂,并且不包含bmp通路抑制剂(例如头蛋白)。用于鉴定合适的bmp激活剂的方法是本领域已知的。zilberberg等人,bmccellbiology20078:41中描述了用于测量bmp活性的合适测定。在一些实施方案中,分化培养基包含bmp通路激活剂。在一些实施方案中,bmp通路激活剂选自bmp7、bmp4和bmp2。bmp7是优选的。bmp7诱导smad1和smad5的磷酸化。因此,在一些实施方案中,bmp通路激活剂是能够诱导smad1和smad5的磷酸化的任何化合物。此外,当提及bmp7时,可以使用任何诱导smad1或smad5的磷酸化的化合物来代替bmp7。在一些实施方案中,bmp通路激活剂(如bmp4或bmp7)以以下浓度存在于分化培养基中:至少0.01ng.ml、至少0.1ng/ml、至少1ng/ml、至少10ng/ml、至少20ng/ml、至少25ng/ml、至少100ng/ml、至少500ng/ml、至少1μg/ml、至少10μg/ml或至少50μg/ml。在一些实施方案中,bmp通路激活剂(如bmp4或bmp7)以以下浓度存在于分化培养基中:约0.01ng/ml至约500ng/ml、约1ng/ml至约500ng/ml、约10ng/ml至约500ng/ml、约20ng/ml至约500ng/ml。在一些实施方案中,bmp通路激活剂(如bmp4或bmp7)以以下浓度存在于分化培养基中:约0.01ng/ml至约200ng/ml、约0.1ng/ml至约100ng/ml、约1ng/ml至约100ng/ml、约10ng/ml至约100ng/ml、约10ng/ml至约50ng/ml、约15ng/ml至约30ng/ml。在一些实施方案中,bmp通路激活剂(如bmp4或bmp7)以约25ng/ml存在于分化培养基中。在一些实施方案中,bmp4以约0.1μg/ml至约50μg/ml、约1至约50μg/ml、约5μg/ml至约25μg/ml或约5μg/ml至约15μg/ml存在于分化培养基中。在一些实施方案中,bmp4以约10μg/ml存在于分化培养基中。在一些实施方案中,分化培养基不包含bmp通路激活剂。还提供了用于增加包含eec的细胞群中分泌素分泌的方法,其中所述方法包括使细胞群与bmp激活剂接触。还提供了用于降低包含eec的细胞群中glp-1分泌的方法,其中所述方法包括使细胞群与bmp激活剂接触。还提供了用于增加包含eec的细胞群中pyy和/或nts表达的方法,其中所述方法包括使细胞群与bmp激活剂接触。bmp抑制剂本发明人还惊奇地发现,在eec分化培养基中包含bmp信号传导抑制剂促进分化的eec群体中的隐窝激素特征(hormonesignature)(参见实施例5)。隐窝激素特征的特征在于glp-1的高表达和缺乏分泌素表达。因此,在一些实施方案中,本发明的分化培养基还包含bmp抑制剂。在这些实施方案中,分化培养基优选不包含bmp激活剂。bmp是与两类细胞表面骨形态发生蛋白受体(bmpr-i和bmprii)结合的小信号传导分子。bmpr-1受体类别由三种受体类型组成:激活素受体样激酶-2(alk-2或actr-ia)、alk-3(bmpr-ia)和alk-6(bmpr-ib)。bmpr-ii受体类别由三种受体类型组成:bmpr-ii、actr-iia和actr-iib。bmp的结合导致形成含有两种i型和两种ii型受体的异四聚体复合物。除细胞外结合结构域外,每种bmp受体还含有细胞内丝氨酸/苏氨酸激酶结构域。结合bmp后,组成型活性ii型受体激酶磷酸化i型受体激酶结构域,进而磷酸化bmp-响应性smad1、5和8,其可进入细胞核并起转录因子的作用。这些特异性smad的磷酸化导致各种细胞效应,包括生长调控和分化。bmp抑制剂是任何通过这些通路导致信号传导显著减少的抑制剂。例如,bmp抑制剂可能能够破坏bmp与bmp受体的相互作用;结合至bmp受体并抑制下游信号传导的激活;抑制smad1、smad5或smad8的磷酸化;抑制smad1、smad5或smad8向细胞核的易位;抑制smad1、smad5或smad8介导的靶基因转录;或抑制bmp的表达、折叠或分泌。在一些实施方案中,bmp抑制剂通过bmpr-1受体类别减少信号传导。在一些实施方案中,bmp抑制剂通过bmpr-ii受体类别减少信号传导。在一些实施方案中,bmp抑制剂通过smad1/5/8减少信号传导。抑制可以是直接的或间接的。许多bmp抑制剂在本领域中是已知的,例如,如cuny,等人,(2008)structure-activityrelationshipstudyofbonemorphogeneticprotein(bmp)signalinginhibitors.bioorgmedchemlett18:4388-4392中所公开的。这些bmp抑制剂中的任一种都适用于本发明的方法。用于鉴定合适的bmp抑制剂的方法是本领域已知的。zilberberg等人,bmccellbiology20078:41中描述了合适的测定。适用于bmp抑制剂的另一种测定(特别是抑制通过alk2和alk3磷酸化smad1、5或8的bmp抑制剂)可以由本领域技术人员使用cuny,等人,(2008)structure-activityrelationshipstudyofbonemorphogeneticprotein(bmp)signalinginhibitors.bioorgmedchemlett18:4388-4392中描述的cytobot细胞elisa测定鉴定。在一些实施方案中,bmp抑制剂选自头蛋白、脊索发生素、卵泡抑素、gremlin、tsg(扭曲的原肠胚形成)、sog(短原肠胚形成)、dorsomorphin和ldn193189。在一些实施方案中,bmp抑制剂选自:a.dorsomorphin或ldn193189或其类似物或变体;和/或b.头蛋白、硬化蛋白、脊索发生素、ctgf、卵泡抑素、gremlin、tsg、sog或其类似物或变体。在一些优选的实施方案中,bmp抑制剂是头蛋白。头蛋白特别适用于体外培养方法。在其他优选的实施方案中,bmp抑制剂是ldn193189。ldn193189特别适用于体内治疗方法,因为它可口服并因此适用于口服施用(参见后面的治疗方法的进一步评论)。在一些实施方案中,头蛋白以以下终浓度包含在分化培养基中:1至1000ng/ml、10至1000ng/ml、100至1000ng/ml、1至500ng/ml、1至200ng/ml、1至100ng/ml、10至500ng/ml、20至500ng/ml、10至200ng/ml、20至200ng/ml、50至500ng/ml或between50至200ng/ml。在一些实施方案中,头蛋白以约100ng/ml的终浓度包含在分化培养基中。在一些实施方案中,ldn193189以以下终浓度包含在分化培养基中:1nm至10μm、5nm至10μm、10nm至10μm、100nm至10μm、1μm至10μm或1μm至5μm。在一些实施方案中,ldn193189以至少1nm、至少5nm、至少10nm、至少100nm、至少1μm或约10μm的终浓度包含在分化培养基中。还提供了用于增加包含eec的细胞群中glp-1分泌的方法,其中所述方法包括使细胞群与bmp抑制剂接触。还提供了用于降低包含eec的细胞群中分泌素分泌的方法,其中所述方法包括使细胞群与bmp抑制剂接触。还提供了用于增加包含eec的细胞群中tac1表达的方法,其中所述方法包括使细胞群与bmp抑制剂接触。还提供了用于增加包含eec的细胞群中gcg表达的方法,其中所述方法包括使细胞群与bmp抑制剂接触。hedgehog激活剂和抑制剂本发明人惊奇地发现,在eec分化培养基中包含hedgehog信号传导的抑制剂可以在一些情况下增加分化的eec群中分泌glp1和分泌cck的细胞的存在(参见图14)。因此,在一些实施方案中,本发明的分化培养基还包含一种或多种hedgehog抑制剂。一种或多种hedgehog抑制剂可以是降低细胞中hedgehog信号传导的任何合适的抑制剂。这种培养基适于获得富含分泌glp1和分泌cck的细胞的eec群。或者,使用还包含hedgehog激活剂的本发明的分化培养基可获得其中分泌glp1和分泌cck的细胞数已耗尽的eec群体。因此,在一些实施方案中,本发明的分化培养基还包含一种或多种hedgehog激活剂。一种或多种hedgehog激活剂可以是增加细胞中hedgehog信号传导的任何合适的激活剂。哺乳动物hedgehog(hh)配体包括sonichedgehog(shh)、indianhedgehog(ihh)和deserthedgehog(dhh)。hh配体通常作为前体蛋白合成,其在羧基末端进行自催化裂解和伴随的胆固醇修饰并且在氨基末端进行棕榈酰化,产生分泌的双重脂化蛋白。hh配体通过dispatched和scube2的联合作用从细胞表面释放,并随后通过与细胞表面蛋白lrp2和硫酸乙酰肝素蛋白聚糖的磷脂酰肌醇蛋白聚糖家族(gpc1-6)的相互作用在多个细胞上运输。hh蛋白通过与经典受体patched(ptch1)和共受体gas1、cdon和boc结合而启动信号传导。hh与ptch1的结合导致gpcr-氧蛋白smoothened(smo)的去阻遏,从而导致smo在纤毛中积累和其细胞质尾磷酸化。smo介导下游信号转导,包括gli蛋白(hh通路的转录效应子)与驱动蛋白家族蛋白kif7和关键细胞内hh通路调节子sufu的解离。在一些实施方案中,一种或多种hedgehog抑制剂包括smo抑制剂。在一些实施方案中,smo抑制剂是环巴胺或环巴胺竞争性抑制剂(例如,vismogenib、saridegib或环巴胺)。在其他实施方案中,smo抑制剂不是环巴胺竞争性抑制剂(例如伊曲康唑)。在一些实施方案中,一种或多种hedgehog抑制剂包括抑制hh与ptch1结合的抗ptch1抗体(参见,例如,nakamura等人(2007)anticancerresearch27:3743-3748)。在一些实施方案中,一种或多种hedgehog激活剂包括smo激活剂。在一些实施方案中,smo激活剂是sag(hh-ag1.3)或嘌吗啡胺。可以使用本领域已知的方法鉴定hedgehog抑制剂和hedgehog激活剂,例如使用rt-pcr方法测定gli1和ptch1的mrna水平(参见,例如,nakamura等人(2007)anticancerresearch27:3743-3748)。gli1和ptch1是gli1反式激活的靶基因,并因此可以用作hh信号传导通路活性的标志物。例如,gli1和ptch1的抑制表达指示hh信号传导通路活性降低。在一个实施方案中,如通过rt-pcr测定评估,根据本发明的hedgehog抑制剂将hedgehog信号传导通路的活性与对照相比降低大于10%;大于30%;大于60%;大于80%;大于90%;大于95%;或大于99%。用于测量抑制的rt-pcr测定的实例是如上所述的本领域熟知的。在一个实施方案中,如通过rt-pcr测定评估,根据本发明的hedgehog激活剂将hedgehog信号传导通路的活性与对照相比增加大于10%;大于30%;大于60%;大于80%;大于90%;大于95%;或大于99%。用于测量激活的rt-pcr测定的实例是如上所述的本领域熟知的。在一些实施方案中,hedgehog抑制剂以0.01-200μm、0.01-100μm、0.05-100μm、0.1-50μm、0.1-20μm、1-100μm、1-50μm、1-30μm、1-10μm、5-100μm、5-50μm或5-20μm的浓度使用。在一些实施方案中,hedgehog抑制剂(例如vismogenib)以约5μm的浓度存在。在一些实施方案中,hedgehog激活剂以0.01-200μm、0.01-100μm、0.05-100μm、0.1-50μm、0.1-20μm、1-100μm、1-50μm、1-30μm、1-10μm、5-100μm、5-50μm或5-20μm的浓度使用。在一些实施方案中,hedgehog激活剂(例如sag(hh-ag1.3)或嘌吗啡胺)以约5μm的浓度存在。在一些实施方案中,本发明的分化培养基包含两种、三种、四种或更多种hedgehog抑制剂。在一些实施方案中,本发明的分化培养基包含两种、三种、四种或更多种hedgehog激活剂。在一些实施方案中,分化培养基不包含hedgehog抑制剂。mtor信号传导的调节剂本发明人惊奇地发现,包含mtor信号传导的调节剂可以影响eec亚型的相对比例(例如mtor激活剂可以促进向表达glu的eec分化)。在一些实施方案中,本发明的分化培养基还包含mtor信号传导的一种或多种调节剂。在一些实施方案中,mtor信号传导的一种或多种调节剂包括mtor信号传导的抑制剂。一种或多种mtor抑制剂可以是降低细胞中mtor信号传导的任何合适的抑制剂。在一些实施方案中,mtor信号传导的一种或多种调节剂包括mtor信号传导的激活剂(例如mhy1485)。一种或多种mtor激活剂可以是增加细胞中mtor信号传导的任何合适的激活剂。雷帕霉素(mtor)的机制靶标是非典型的丝氨酸/苏氨酸激酶,其存在于两种不同的复合物中。第一种mtor复合物1(mtorc1)由mtor、raptor、gβl和deptor组成,并被雷帕霉素抑制。它是一种主要的生长调节剂,其感知和整合多种营养和环境因素,包括生长因子、能量水平、细胞应激和氨基酸。它通过磷酸化增强合成代谢过程(如mrna翻译和脂质合成)或限制分解代谢过程(如自噬)的底物,将这些信号与促进细胞生长偶联。小gtp酶rheb在其gtp结合状态下是mtorc1激酶活性的必需且有效的刺激物,其受其gap(结节性硬化异二聚体tsc1/2)的负调控。大多数上游输入通过akt和tsc1/2汇集,以调控rheb的核苷酸负载状态。相反,氨基酸独立于pi3k/akt轴向mtorc1发信号以促进mtorc1易位至溶酶体表面,其中它可在与rheb接触后被激活。该过程由多种复合物的协同作用介导,特别是v-atp酶、ragulator、raggtp酶和gator1/2。第二种复合物mtor复合物2(mtorc2)由mtor、rictor、gβl、sin1、prr5/protor-1和deptor组成。mtorc2通过激活akt促进细胞存活,通过激活pkcα调控细胞骨架动力学,并通过sgk1磷酸化控制离子转运和生长。在一些实施方案中,mtor信号传导的激活剂是mhy1485。在一些实施方案中,mtor信号传导的抑制剂是雷帕霉素或雷帕霉素类似物(例如雷帕霉素、地磷莫司(ap23573)、依维莫司(rad001)和替西罗莫司(cci-779))。在一些实施方案中,mtor信号传导的抑制剂是atp竞争性mtor激酶抑制剂(例如mln0128、pp242或azd8055)。可以使用本领域已知的方法鉴定mtor抑制剂和mtor激活剂,例如使用基于elisa的mtor激酶测定(例如使用k-lisatmmtor活性试剂盒,其是用于检测在atp存在下磷酸化p70s6k-gst融合蛋白(一种特异性mtor底物)的基于elisa的活性测定)。在一个实施方案中,如通过通过基于elisa的测定评估,根据本发明的mtor抑制剂与对照相比将mtor信号传导通路的活性降低大于10%;大于30%;大于60%;大于80%;大于90%;大于95%;或大于99%。用于测量抑制的基于elisa的测定的实例是如上所述的本领域熟知的。在一个实施方案中,如通过通过基于elisa的测定评估,根据本发明的mtor激活剂与对照相比将mtor信号传导通路的活性增加大于10%;大于30%;大于60%;大于80%;大于90%;大于95%;或大于99%。用于测量激活的基于elisa的测定的实例是如上所述的本领域熟知的。在一些实施方案中,mtor抑制剂以0.01-200μm、0.01-100μm、0.05-100μm、0.1-50μm、0.1-20μm、1-100μm、1-50μm、1-30μm、1-10μm、5-100μm、5-50μm或5-20μm的浓度使用。在一些实施方案中,mtor抑制剂(例如雷帕霉素、地磷莫司(ap23573)、依维莫司(rad001)或替西罗莫司(cci-779))以约5μm的浓度存在。在一些实施方案中,mtor激活剂以0.01-200μm、0.01-100μm、0.05-100μm、0.1-50μm、0.1-20μm、1-100μm、1-50μm、1-30μm、1-10μm、5-100μm、5-50μm或5-20μm的浓度使用。在一些实施方案中,mtor激活剂(例如mhy1485)以约5μm的浓度存在。在一些实施方案中,本发明的分化培养基包含两种、三种、四种或更多种mtor抑制剂。在一些实施方案中,本发明的分化培养基包含两种、三种、四种或更多种mtor激活剂。gsk-3抑制剂之前发现包含gsk-3抑制剂(例如,chir99021)也改善了上皮干细胞的分化(参见gb1603569.3)。如上所述,gsk-3是β-连环蛋白破坏复合物的组分。抑制gsk-3活性导致β-连环蛋白的破坏减少,并因此gsk-3抑制剂是wnt激动剂。通过添加抑制破坏复合物下游tcf/lef介导的转录的wnt抑制剂(icrt3),极大地增加了通过分化培养基中包含gsk-3抑制剂所引起的增强的分化。例如,当用于肝脏祖细胞的分化培养基中包括wnt激动剂(chir99021)和wnt抑制剂(icrt3)两者时,肝细胞标志物(白蛋白和cyp3a4)的表达显著提高。在不希望受任何理论束缚的情况下,发明人认为gsk-3抑制剂可刺激并抑制上皮干细胞的分化(其中刺激比抑制强)。通过gsk-3抑制剂抑制分化被认为经由促进β-连环蛋白的破坏而发生。相反,通过gsk-3抑制剂促进分化被认为是通过不同的效应机制发生的。因此,当gsk-3抑制剂被包含在分化培养基中时,wnt抑制剂应该是在包含gsk-3抑制剂的分化培养基中抑制tcf/lef介导的转录下游破坏复合物的wnt抑制剂,中和了gsk-3抑制剂的分化抑制作用。作用于破坏复合物下游的wnt抑制剂包括上述第(7)类中的wnt抑制剂,即β-连环蛋白靶基因表达的抑制剂,包括β-连环蛋白:tcf/lef转录复合物的抑制剂,如破坏β-连环蛋白:tcf-4复合物的抑制剂(例如icrt3、cgp049090、pkf118310、pkf115-584、ztm000990、pnu-74654、bc21、icrt5、icrt14或fh535)和组蛋白脱乙酰酶sirt1的抑制剂(例如cambinol)。因此,gsk-3抑制剂的分化促进作用保持不受影响,并且通过wnt抑制剂的分化促进作用而增加。因此,在一些实施方案中,本发明的分化培养基包含gsk-3抑制剂。可以使用任何合适的gsk-3抑制剂。gsk-3抑制剂被限定为降低gsk-3激酶活性的试剂。在人类中发现了两种不同的gsk-3同种型(gsk-3α和gsk-3β)。已知抑制剂抑制这些同种型中的一种或两种。因此,在一些实施方案中,gsk-3抑制剂抑制gsk-3α和gsk-3β。在一些实施方案中,gsk-3抑制剂抑制gsk-3α但不抑制gsk-3β。在一些实施方案中,gsk-3抑制剂抑制gsk-3β但不抑制gsk-3α。在一些实施方案中,分化培养基包含多于一种的gsk-3抑制剂(例如两种、三种、四种、五种或更多种gsk-3抑制剂)。在一些实施方案中,gsk-3抑制剂是chir99021。在一些实施方案中,向培养基中添加一定量的gsk-3抑制剂,如在相同细胞类型中所评估的,相对于所述分子不存在的情况下所述gsk-3活性的水平,所述量有效抑制细胞中至少10%,更优选至少20%,更优选至少30%,更优选至少50%,更优选至少70%,更优选至少90%,更优选100%的gsk-3活性。如技术人员已知的,可以通过监测特定gsk-3底物(例如β-连环蛋白)的磷酸化来测量gsk-3活性(参见例如,cole等人(2008)methodsmolbiol.468:45-65)。因此,技术人员可以使用本领域已知的分析容易地鉴定新的gsk-3抑制剂。在一些实施方案中,gsk-3抑制剂选自以下中的一种或多种:chir99201、6-bio、dibromocantharelline、海门地塞(hymenialdesine)、靛玉红(indirubins)、meridianins、ct98014、ct98023、ct99021、tws119、sb-216763、sb-41528、ar-a014418、azd-1080、阿特波龙(alsterpaullone)、cazpaullone、肯帕罗酮(kenpaullone)、aloisines、manzaminea、palinurine、tricantine、tdzd-8、np00111、np031115、tideglusib、hmk-32和l803-mts。在一些实施方案中,本发明的分化培养基包含以下浓度的gsk-3抑制剂:0.001-500μm、0.01-150μm、0.1-100μm、1-100μm、0.5-50μm、1-50μm、1-20μm或者1-5μm。在一些实施方案中,本发明的分化培养基包含以下浓度的chir99021:0.01-150μl、0.1-100μm、1-100μm、0.5-50μm、1-50μm、1-20μm或者1-5μm。例如,在一些实施方案中,本发明的分化培养基包含浓度为约3μm的chir99021。在一些实施方案中,本发明的分化培养基包含gsk-3抑制剂和作用于破坏复合物下游的wnt抑制剂(如如前所述的β-连环蛋白靶基因表达的抑制剂)。例如,在一些实施方案中,本发明的分化培养基包含gsk-3抑制剂和β-连环蛋白靶基因表达的抑制剂。β-连环蛋白靶基因表达的抑制剂包括β-连环蛋白:tcf/lef转录复合物的抑制剂,如破坏β-连环蛋白:tcf-4复合物的抑制剂(例如icrt3、cgp049090、pkf118310、pkf115-584、ztm000990、pnu-74654或bc21)和组蛋白脱乙酰酶sirt1的抑制剂(例如cambinol)。因此,在一些实施方案中,本发明的分化培养基包含gsk-3抑制剂(例如浓度为0.001-500μm、0.01-150μm、0.1-100μm、0.5-50μm、1-20μm或者1-5μm)和β-连环蛋白靶基因表达的抑制剂(例如浓度为0.001-500μm、0.01-150μm、0.1-100μm、0.5-50μm、1-500μm、1-150μm、1-100μm、1-50μm、120μm或者1-5μm)。在一些实施方案中,β-连环蛋白靶基因表达的抑制剂是β-连环蛋白:tcf/lef转录复合物的抑制剂。因此,在一些实施方案中,本发明的分化培养基包含chir99021(例如浓度为0.01-150μm、0.1-100μm、0.5-50μm、1-20μm或者1-5μm)和icrt3(例如浓度为0.05-150μm、0.1-150μm、1-150μm、0.5-100μm、1-100μm或者10-80μm)。例如,在一些实施方案中,本发明的分化培养基包含浓度为约3μm的chir99021和浓度为约50μm的icrt3。在一些实施方案中,分化培养基不包含gsk-3抑制剂。chir99021激动剂如上所述,之前发现在分化培养基中包含wnt激动剂chir99021促进了分化。因此,在一些实施方案中,本发明的分化培养基包含chir99021或chir99021激动剂。chir99021激动剂在本文中被限定为与chir99021共享一种或多种生物活性的试剂。ap-1刺激剂激活蛋白1(ap-1)复合物是转录因子,其是由属于fos蛋白家族、jun、atf和jdp家族的蛋白组成的异二聚体。该转录因子响应多种刺激调节基因表达,所述刺激包括细胞因子、生长因子、胁迫以及细菌和病毒感染。之前观察到,与原代肝细胞相比,人肝脏类器官具有降低的ap-1复合物组分的表达。基于这一观察,他们假设刺激ap-1复合物的形成将促进肝脏类器官中的上皮干细胞分化成肝细胞的命运。发明人发现,在分化培养基中包括ap-1刺激剂卡巴胆碱(氯化2-[(氨基羰基)氧基]-n,n,n-三甲基乙铵)增加了肝细胞标志物(包括cyp3a4和白蛋白)的表达。之前测试的ap-1刺激剂是毒蕈碱乙酰胆碱受体激动剂。存在五种毒蕈碱乙酰胆碱受体亚型(m1-m5)。这些是存在于多种细胞类型(例如神经元)的细胞膜中的g蛋白偶联受体。设想ap-1刺激剂或毒蕈碱乙酰胆碱受体激动剂还可以增强上皮干细胞从肝脏以外的组织(例如肠、胃、胰腺或肺)分化。因此,在一些实施方案中,本发明的分化培养基包含ap-1刺激剂。在一些实施方案中,ap-1刺激剂是毒蕈碱乙酰胆碱受体激动剂。可以使用任何合适的毒蕈碱乙酰胆碱受体激动剂。在一些实施方案中,毒蕈碱乙酰胆碱受体激动剂是m3毒蕈碱乙酰胆碱受体激动剂(例如乙酰胆碱、氨甲酰甲胆碱、卡巴胆碱、氧代震颤素或毛果芸香碱)。在一些实施方案中,毒蕈碱乙酰胆碱受体激动剂选自以下中的一种或多种:乙酰胆碱、氨甲酰甲胆碱、卡巴胆碱、氧代震颤素、l-689,660(1-氮杂双环[2.2.2]辛烷、3-(6-氯吡嗪基)马来酸盐)、毛果芸香碱、毒蕈碱、mcn-a343(4-[[[(3-氯苯基)氨基]羰基]氧基]-n,n,n-三甲基-2-丁炔-1-氯化铵)、77-lh-218-1(1-[3-(4-丁基-1-哌啶基)丙基]-3,4-二氢-2(1h)-喹啉酮)和乙酰甲胆碱。在一些实施方案中,毒蕈碱乙酰胆碱受体激动剂是卡巴胆碱。在一些实施方案中,ap-1刺激剂(例如毒蕈碱乙酰胆碱受体激动剂)以以下浓度存在于本发明的分化培养基中:0.001-500μm、0.01-500μm、0.01-250μm、0.01-150μm、0.1-500μm、0.1-100μm、0.5-500μm、0.5-100μm、0.5-50μm、1-500μm、1-300μm、1-200μm、1-20μm、1-5μm、10-300μm、50-300μm、50-200μm或50-150μm。在一些实施方案中,本发明的分化培养基包含以下浓度的卡巴胆碱:0.001-500μm、0.001-300μm、0.01-500μm、0.01-300μm、0.1-500μm、0.1-300μm、1-500μm、10-500μm、100-500μm、1-300μm、10-300μm、50-300μm、50-200μm或50-150μm。例如,在一些实施方案中,本发明的分化培养基包含浓度为100μm的卡巴胆碱。毒蕈碱乙酰胆碱受体激动剂如上所述,之前发现在分化培养基中包含毒蕈碱乙酰胆碱受体激动剂卡巴胆碱促进分化。因此,在一些实施方案中,本发明的分化培养基还包含毒蕈碱乙酰胆碱受体激动剂。可以使用任何合适的毒蕈碱乙酰胆碱受体激动剂。在一些实施方案中,毒蕈碱乙酰胆碱受体激动剂是m3毒蕈碱乙酰胆碱受体激动剂(例如乙酰胆碱、氨甲酰甲胆碱、卡巴胆碱、氧代震颤素或毛果芸香碱)。在一些实施方案中,毒蕈碱乙酰胆碱受体激动剂选自以下中的一种或多种:乙酰胆碱、氨甲酰甲胆碱、卡巴胆碱、氧代震颤素、l-689,660(1-氮杂双环[2.2.2]辛烷、3-(6-氯吡嗪基)马来酸盐)、毛果芸香碱、毒蕈碱、mcn-a343(4-[[[(3-氯苯基)氨基]羰基]氧基]-n,n,n-三甲基-2-丁炔-1-氯化铵)、77-lh-218-1(1-[3-(4-丁基-1-哌啶基)丙基]-3,4-二氢-2(1h)-喹啉酮)和乙酰甲胆碱。在一些实施方案中,毒蕈碱乙酰胆碱受体激动剂是卡巴胆碱。在一些实施方案中,ap-1刺激剂(例如毒蕈碱乙酰胆碱受体激动剂)以以下浓度存在于本发明的分化培养基中:0.001-500μm、0.01-500μm、0.01-250μm、0.01-150μm、0.1-500μm、0.1-100μm、0.5-500μm、0.5-100μm、0.5-50μm、1-500μm、1-300μm、1-200μm、1-20μm、1-5μm、10-300μm、50-300μm、50-200μm或50-150μm。在一些实施方案中,本发明的分化培养基包含以下浓度的卡巴胆碱:0.001-500μm、0.001-300μm、0.01-500μm、0.01-300μm、0.1-500μm、0.1-300μm、1-500μm、10-500μm、100-500μm、1-300μm、10-300μm、50-300μm、50-200μm或50-150μm。例如,在一些实施方案中,本发明的分化培养基包含浓度为100μm的卡巴胆碱。卡巴胆碱激动剂如上所述,之前发现在分化培养基中包含毒蕈碱乙酰胆碱受体激动剂卡巴胆碱促进分化。因此,在一些实施方案中,本发明的分化培养基包含还卡巴胆碱或卡巴胆碱激动剂。卡巴胆碱激动剂在本文中被限定为与卡巴胆碱共享一种或多种生物活性的试剂。camp通路激活剂在一些实施方案中,本发明的分化培养基还包含camp通路激活剂。camp通路激活剂可以是提高细胞中camp的水平的任何合适的激活剂。camp通路涉及激活许多类型的激素和神经递质g蛋白偶联受体。激素或神经递质与其膜结合受体的结合诱导受体的构象变化,这导致g蛋白的α-亚基的激活。激活的g亚基刺激,而未激活的g亚基抑制腺苷酸环化酶。腺苷酸环化酶的刺激催化细胞质atp向camp的转化,从而提高细胞中camp的水平。因此,camp通路激活剂可以是,例如,腺苷酸环化酶激活剂。合适的腺苷酸环化酶激活剂的实例包括毛喉素、毛喉素类似物和霍乱毒素。在一些实施方案中,camp通路激活剂是毛喉素。在一些实施方案中,camp通路激活剂不是霍乱毒素。在一些实施方案中,camp通路激活剂可以是camp类似物,例如8-溴-camp。8-溴-camp是一种细胞可渗透camp类似物,其对磷酸二酯酶水解比对camp水解具有更强的抗性。在一些实施方案中,camp通路激活剂是nkh477(例如目录号tocris1603)。camp通路激活剂可以使用本领域已知的方法、例如使用测量camp水平的竞争性免疫测定鉴定。cyclic-amp荧光测定试剂盒(moleculardevicesllc)是用于进行这种免疫测定的可商购获得的试剂盒的实例。样品或标准品中的camp与辣根过氧化物酶(hrp)-标记的camp缀合物竞争抗camp抗体上的结合位点。在不存在camp的情况下,大多数hrp-camp缀合物与抗体结合。增加浓度的camp竞争性地降低结合的缀合物的量,从而降低测量的hrp活性。与对照相比,camp通路激活剂将导致camp水平升高和测量的hrp活性降低。在一些实施方案中,camp通路激活剂的使用浓度为约10nm至约500μm、约10nm至约100μm、约1μm至约50μm、约1μm至约25μm、约5μm至约1000μm、约5μm至约500μm、约5μm至约100μm、约5μm至约50μm、约5μm至约25μm、约10μm至约1000μm、约10μm至约500μm、约10μm至约100μm、约10μm至约50μm、约10μm至约25μm或约20μm。在一些实施方案中,camp通路激活剂的使用浓度为至少10nm、20nm、50nm、100nm、200nm、500nm、1μm、至少2μm、至少5μm、至少10μm、至少20μm、至少30μm、至少50μm或至少100μm。选择的浓度可取决于所用的camp通路激活剂,并且可由本领域技术人员根据camp通路激活剂的效力确定。例如,nkh477通常比8-br-camp和毛喉素更有效。更有效的camp通路激活剂可以在较低浓度下使用以达到相同的效应。例如,在一些实施方案中,nkh477可以以约100nm至约10μm的浓度使用,或以约100nm、约1μm或约10μm的浓度使用。在一些实施方案中,8-br-camp或毛喉素可以以约1μm至约100μm的浓度使用,或以约1μm、约10μm或约100μm的浓度使用。在一些实施方案中,霍乱毒素可以以以下浓度使用:约1ng/ml至约500ng/ml、约10ng/ml至约100ng/ml、约50ng/ml至约100ng/ml、或约10ng/ml、约20ng/ml、约30ng/ml、约40ng/ml、约50ng/ml、约60ng/ml、约70ng/ml、约80ng/ml、约90ng/ml、约100ng/ml、约200ng/ml、约300ng/ml、约400ng/ml或约500ng/ml。补充剂本发明的分化培养基优选被补充有选自b27、n-乙酰半胱氨酸和n2的化合物中的一种或多种(例如1、2、3种或全部)。因此,在一些实施方案中,分化培养基还包含选自b27、n2和n-乙酰半胱氨酸的一种或多种组分。例如,在一些实施方案中,分化培养基还包含b27、n-乙酰半胱氨酸和n2。据信b27(invitrogen)、n-乙酰半胱氨酸(sigma)和n2(invitrogen)以及烟酰胺(sigma)控制细胞的增殖并有助于dna稳定性。在一些实施方案中,n-乙酰半胱氨酸以以下浓度存在于分化培养基中:0.1-200mm、0.1-100mm、0.1-50mm、0.1-10mm、0.1-5mm、0.5-200mm、0.5-100mm、0.5-50mm、0.5-10mm、0.5-5mm、1-100mm、1-50mm、1-10mm、1-5mm。在一些实施方案中,n-乙酰半胱氨酸以约1.25mm的浓度存在于分化培养基中。在一些实施方案中,b27补充剂是‘b27补充剂减去维生素a’(本文中也被称为“不含维生素a的b27”或“b27wovita”;可获自invitrogen,carlsbad,ca;www.invitrogen.com;当前目录号12587010;以及获自paalaboratoriesgmbh,pasching,奥地利;www.paa.com;目录号f01-002;brewer等人(1993)jneuroscires.35(5):567-76)。在一些实施方案中,可以用包含选自以下列出的一种或多种组分的通用制剂代替b27补充剂:生物素、胆固醇、亚油酸、亚麻酸、黄体酮、腐胺、视黄醇乙酸酯、亚硒酸钠、三碘甲腺原氨酸(t3)、dl-α生育酚(维生素e)、白蛋白、胰岛素和转铁蛋白。由paalaboratoriesgmbh提供的b27补充剂是液体的50x浓缩液,其中含有生物素、胆固醇、亚油酸、亚麻酸、黄体酮、腐胺、视黄醇、视黄醇乙酸酯、亚硒酸钠、三碘甲腺原氨酸(t3)、dl-α生育酚(维生素e)、白蛋白、胰岛素和转铁蛋白等其它成分。在这些成分中,至少亚麻酸、视黄醇、视黄醇乙酸酯和三碘甲腺原氨酸(t3)是核激素受体激动剂。可以将b27补充剂作为浓缩物添加到分化培养基中,或者在添加到分化培养基之前对其进行稀释。它可以以1x最终浓度或其它最终浓度(例如0.1x至4x的浓度、0.1x至2x的浓度、0.5x至2x的浓度、1x至4x的浓度或1x至2x的浓度)被使用。使用b27补充剂是向本发明的分化培养基中掺入生物素、胆固醇、亚油酸、亚麻酸、黄体酮、腐胺、视黄醇、视黄醇乙酸酯、亚硒酸钠、三碘甲腺原氨酸(t3)、dl-α生育酚(维生素e)、白蛋白、胰岛素和转铁蛋白的便捷方式。还设想可以向分化培养基中分别添加这些组分中的一些或全部,而非使用b27补充剂。因此,分化培养基可以包含这些组分中的一些或全部。在一些实施方案中,分化培养基中使用的b27补充剂中不存在视黄酸和/或分化培养基中不存在视黄酸。‘n2补充剂’(在本文中也被称为“n2”)可获自invitrogen,carlsbad,ca;www.invitrogen.com;目录号17502-048;以及获自paalaboratoriesgmbh,pasching,奥地利;www.paa.com;目录号f005-004;bottenstein&sato,pnas,76(1):514-517,1979。由paalaboratoriesgmbh提供的n2补充剂是100x液体浓缩,其含有500μg/ml人转铁蛋白、500μg/ml牛胰岛素、0.63μg/ml黄体酮、1611μg/ml腐胺和0.52μg/ml亚硒酸钠。可以将n2补充剂作为浓缩物添加到分化培养基中,或者在添加到分化培养基之前对其进行稀释。它可以以1x最终浓度或其它最终浓度(例如0.1x至4x的浓度、0.1x至2x的浓度、0.5x至2x的浓度、1x至4x的浓度或1x至2x的浓度)使用。使用n2补充剂是向本发明的分化培养基中掺入转铁蛋白、胰岛素、黄体酮、腐胺和亚硒酸钠的便捷方式。当然还设想可以向分化培养基中分别添加这些组分中的一些或全部,而非使用n2补充剂。因此,分化培养基可以包含这些组分中的一些或全部。在其中培养基包含b27的一些实施方案中,它也不包含n2。因此,如果需要,本发明的实施方案可以适于当存在b27时排除n2。在一些实施方案中,n2不存在于分化培养基中。在其中培养基包含n2的一些实施方案中,它也不包含b27。因此,如果需要,本发明的实施方案可以适于当存在n2时排除b27。在一些实施方案中,b27不存在于分化培养基中。在一些实施方案中,将分化培养基补充有b27和/或n2。在一些实施方案中,将基础培养基补充有150ng/ml至250ng/ml的n-乙酰半胱氨酸;优选地,将基础培养基补充有约200ng/ml的n-乙酰半胱氨酸。可以使用任何合适的ph。例如,培养基的ph可以在约7.0至7.8的范围内,在约7.2至7.6的范围内或者为约7.4。可以使用缓冲剂维持ph。本领域技术人员可以容易地选择合适的缓冲剂。可以使用的缓冲剂包括碳酸盐缓冲剂(例如nahco3)和磷酸盐(例如nah2po4)。通常以约50至约500mg/l使用这些缓冲剂。也可以使用其它缓冲剂如n-[2-羟乙基]-哌嗪-n’-[2-乙烷磺酸](hepes)和3-[n-吗啉代]丙磺酸(mops),通常为约1000至约10,000mg/l。在一些实施方案中,缓冲剂选自以下列出的一种或多种:磷酸盐缓冲剂(例如kh2po4、k2hpo4、na2hpo4、nacl、nah2po4)醋酸盐缓冲剂(例如hoac或naoac)、柠檬酸盐缓冲剂(例如柠檬酸或柠檬酸钠)或tris缓冲剂(例如tris、tris-hcl)或有机缓冲剂。在一些实施方案中,有机缓冲剂是两性离子缓冲剂,如good’s缓冲剂,例如选自hepes、mops、mes、ada、pipes、aces、mopso、氯化胆胺、bes、tes、dipso、乙酰氨基甘氨酸(acetamindoglycine)、tapso、popso、heppso、hepps、n-三(羟甲基)甲基甘氨酸(tricine)、甘氨酰胺、n-二羟乙基甘氨酸(bicine)、taps、ampso、cabs、ches、caps和capso。优选的缓冲剂是hepes,例如浓度为0.1-100mm、0.1-50mm、0.5-50mm、1-50mm、1-20mm或5-15mm的hepes。在一些实施方案中,以约10mm向培养基中添加hepes。分化培养基还可以包含ph指示剂(如酚红)以使得能够容易地监测培养基的ph状态(例如,以约5至约50mg/升)。用于本发明的分化培养基可以包含一种或多种氨基酸。技术人员理解用于分化培养基的氨基酸的适当类型和量。可以存在的氨基酸包括l-丙氨酸、l-精氨酸、l-天冬酰胺、l-天冬氨酸、l-半胱氨酸、l-胱氨酸、l-谷氨酸、l-谷氨酰胺、l-甘氨酸、l-组氨酸、l-异亮氨酸、l-亮氨酸、l-赖氨酸、l-甲硫氨酸、l-苯丙氨酸、l-脯氨酸、l-丝氨酸、l-苏氨酸、l-色氨酸、l-酪氨酸、l-缬氨酸及其组合。一些分化培养基将含有所有这些氨基酸。通常,当存在时,除了以约0.05至约1g/l(通常约0.1至约0.75g/l)存在的l-谷氨酰胺之外,每种氨基酸以约0.001至约1g/l培养基(通常为约0.01至约0.15g/l)存在。氨基酸可以是合成来源的。用于本发明的分化培养基可以包含一种或多种维生素。技术人员理解用于分化培养基的维生素的适当类型和量。可以存在的维生素包括硫胺素(维生素b1)、核黄素(维生素b2)、烟酸(维生素b3)、d-泛酸钙(维生素b5)、吡哆醛/吡哆胺/吡哆醇(维生素b6)、叶酸(维生素b9)、氰钴维生素(维生素b12)、抗坏血酸(维生素c)、钙化醇(维生素d2)、dl-α生育酚(维生素e)、生物素(维生素h)以及甲萘醌(维生素k)。用于本发明的分化培养基可以包含一种或多种无机盐。技术人员理解用于分化培养基的无机盐的适当类型和量。分化培养基中通常包括无机盐以帮助维持细胞的渗透平衡并帮助调节膜电势。可以存在的无机盐包括钙、铜、铁、镁、钾、钠、锌的盐。通常以氯化物、磷酸盐、硫酸盐、硝酸盐和碳酸氢盐的形式使用盐。可以使用的具体盐包括cacl2、cuso4-5h2o、fe(no3)-9h2o、feso4-7h2o、mgcl、mgso4、kcl、nahco3、nacl、na2hpo4、na2hpo4-h2o和znso4-7h2o。培养基的同渗容摩可以在约200至约400mosm/kg的范围内,在约290至约350mosm/kg的范围内或者在约280至约310mosm/kg的范围内。培养基的同渗容摩可以小于约300mosm/kg(例如约280mosm/kg)。用于本发明的分化培养基可以包含一种或多种糖形式的碳能源。技术人员理解用于分化培养基的糖的适当类型和量。可以存在的糖包括葡萄糖、半乳糖、麦芽糖和果糖。糖优选是葡萄糖,特别是d-葡萄糖(右旋糖)。碳能源通常将以约1至约10g/l存在。本发明的分化培养基可以包含血清。可以使用获自任何适当来源的血清,包括胎牛血清(fbs)、山羊血清或人血清。优选地,使用人血清。根据常规技术,可以以培养基体积的约1%至约30%使用血清。在其它实施方案中,本发明的分化培养基可以包含血清替代物。多种不同的血清替代物制剂可商购获得并且是技术人员已知的。当使用血清替代物时,根据常规技术,其可以以培养基体积的约1%至约30%被使用。在其它实施方案中,本发明的分化培养基可以是无血清和/或无血清替代物的。无血清培养基是不含任何类型动物血清的培养基。无血清培养基可能是优选的,以避免干细胞可能的异种污染。无血清替代物培养基是尚未补充任何商业血清替代物制剂的培养基。在优选实施方案中,分化培养基补充有纯化的、天然的、半合成的和/或合成的生长因子,并且不包含不确定的组分,如胎牛血清或小牛血清。例如,诸如b27(invitrogen)、n-乙酰半胱氨酸(sigma)和n2(invitrogen)的补充剂刺激一些细胞的增殖。在一些实施方案中,分化培养基补充有这些补充剂中的一种或多种,例如这些补充剂中的一种、任意两种或全部三种。用于本发明的分化培养基可以包含一种或多种微量元素,如下述的离子:钡、溴、钴、碘、锰、铬、铜、镍、硒、钒、钛、锗、钼、硅、铁、氟、银、铷、锡、锆、镉、锌和/或铝。培养基可以包含还原剂,如浓度为约0.1mm的β-巯基乙醇。本发明的分化培养基可以包含一种或多种另外的试剂,如先前报道用于改善干细胞培养的营养物或生长因子,如胆固醇/转铁蛋白/白蛋白/胰岛素/黄体酮、腐胺、亚硒酸盐/其它因子。组合物和容器本发明还提供了组合物或细胞培养容器,其包含根据上述本发明任一方面所述的细胞和/或类器官以及根据上述本发明任一方面所述的分化培养基。例如,此类组合物或细胞培养容器可以包含在如上所述的分化培养基中根据本发明的方法培养的任何数量的细胞或类器官。根据本发明的另外的方面,提供了含有本发明的分化培养基的气密密封容器。气密密封容器可以优选用于运输或储存分化培养基,以防止污染。容器可以是任何合适的容器,如细颈瓶、平板、瓶子、罐子、小瓶或袋子。本发明的示例性分化培养基在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂和egfr抑制剂。在一些实施方案中,分化培养基还包含基础培养基。因此,在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂和egfr抑制剂(例如吉非替尼)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如,浓度为约1mm)和吉非替尼(例如,浓度为约5μm)。例如,在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂和egfr和erbb2抑制剂(例如阿法替尼)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如,浓度为约1mm)和阿法替尼(例如,浓度为约10μm)。在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂和ras-raf-mapk通路抑制剂(例如mek抑制剂)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如浓度为约1mm)和mek抑制剂,如pd0325901(例如,浓度为约1.5μm)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如浓度为约1mm)和erk抑制剂,如sch772984(例如,浓度约为10μm)。在一些实施方案中,本发明的分化培养基还包含bmp通路激活剂(例如bmp4)。如上所讨论,本发明人惊奇地发现,包含bmp通路激活剂可促进分泌素-产生细胞分化并抑制肠嗜铬细胞分化。在其他实施方案中,本发明的分化培养基还包含bmp抑制剂(例如头蛋白)。如上所讨论,本发明人惊奇地发现,包含bmp抑制剂可促进glp-1-产生细胞分化并抑制分泌素-产生细胞分化。因此,在一些实施方案中,本发明的分化培养基包含:wnt抑制剂(例如iwp2)、notch抑制剂(例如dapt)、egfr抑制剂(例如吉非替尼)和bmp通路激活剂(例如bmp4)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如,浓度为约1mm)、吉非替尼(例如,浓度为约5μm)和bmp4(例如浓度为约10μg/ml)。例如,在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂、egfr和erbb2抑制剂(例如阿法替尼)和bmp通路激活剂(例如bmp4)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如浓度为约1mm)、阿法替尼(例如浓度为约10μm)和bmp4(例如浓度为约10μg/ml)。在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂、ras-raf-mapk通路抑制剂(例如mek抑制剂)和bmp通路激活剂如bmp4(例如浓度约10μg/ml)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如浓度为约1.5μm)、dapt(例如浓度为约1mm)、mek抑制剂如pd0325901(例如浓度为约1.5μm)和bmp通路激活剂如bmp4(例如浓度为约10μg/ml)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如浓度为约1.5μm)、dapt(例如浓度为约1mm)、erk抑制剂如sch772984(例如浓度为约10μm)和bmp通路激活剂如bmp4(例如浓度为约10μg/ml)。在一些实施方案中,本发明的分化培养基包含:wnt抑制剂(例如iwp2)、notch抑制剂(例如dapt)、egfr抑制剂(例如吉非替尼)和bmp抑制剂(例如头蛋白)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如浓度为约1.5μm)、dapt(例如浓度为约1mm)、吉非替尼(例如浓度为约5μm)和头蛋白(例如浓度为约100ng/ml)。例如,在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂、egfr和erbb2抑制剂(例如阿法替尼)和bmp抑制剂(例如头蛋白)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如浓度为约1.5μm)、dapt(例如浓度为约1mm)、阿法替尼(例如浓度为约10μm)和头蛋白(例如浓度为约100ng/ml)。在一些实施方案中,本发明的分化培养基包含:wnt抑制剂、notch抑制剂、ras-raf-mapk通路抑制剂(例如mek抑制剂)和bmp抑制剂如头蛋白(例如浓度为约100ng/ml)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如浓度为约1.5μm)、dapt(例如浓度为约1mm)、mek抑制剂如pd0325901(例如浓度为约1.5μm)和bmp抑制剂如头蛋白(例如浓度为约100ng/ml)。例如,在一些实施方案中,本发明的分化培养基包含iwp-2(例如,浓度为约1.5μm)、dapt(例如浓度为约1mm)、erk抑制剂如sch772984(例如浓度为约10μm)和bmp抑制剂如头蛋白(例如浓度为约100ng/ml)。在一些实施方案中,本发明的分化培养基还包含mtor激活剂,如mhy1485(例如,浓度为约5μm)。在一些实施方案中,本发明的分化培养基还包含hedgehog抑制剂,如smo抑制剂(例如,浓度为约5μm)。在一些实施方案中,smo抑制剂是vismogenib。在优选的实施方案中,分化培养基还包含r-spondin(例如浓度为约1μg/ml)。在优选的实施方案中,培养基中egf的浓度小于1mm。细胞外基质在一些实施方案中,用于分化细胞的方法包括培养与细胞外基质(ecm)接触的细胞。可以使用任何合适的ecm。优选在微环境中培养细胞,所述微环境至少部分模拟所述细胞天然存在的细胞小生境。由细胞和所述小生境中的细胞分泌的ecm部分确定细胞小生境。可以通过在提供与细胞膜蛋白(如整合蛋白)的相互作用的生物材料或合成材料的存在下培养所述细胞来模拟细胞小生境。因此,如本文所述的ecm是例如通过与细胞膜蛋白(如整合蛋白)相互作用,模拟体内细胞小生境的任何生物材料或合成材料或其组合。在本发明优选的方法中,将细胞与ecm接触培养。“接触”意为物理或机械或化学接触,这意味着为了将所述得到的类器官或上皮细胞群与所述细胞外基质分离,需要使用力。在一些实施方案中,ecm是三维基质。在一些实施方案中,将细胞嵌入ecm中。在一些实施方案中,细胞附着于ecm。可以将本发明的培养基扩散进三维ecm中。在另一实施方案中,ecm处于悬浮状态,即细胞在悬浮系统中与ecm接触。在一些实施方案中,ecm以至少1%、至少2%或至少3%的浓度存在于悬浮液中。在一些实施方案中,ecm以1%至约10%或1%至约5%的浓度存在于悬浮液中。对于规模扩大的方法,悬浮方法可能具有优势。一种类型的ecm由上皮细胞、内皮细胞、腔壁内胚层样细胞(例如hayashi等人(2004)matrixbiology23:47-62中描述的englebreth-holm-swarm腔壁内胚层样细胞)和结缔组织细胞分泌。该ecm包含多种多糖、水、弹性蛋白和糖蛋白,其中糖蛋白包含胶原蛋白、内联蛋白(entactin)(巢蛋白(nidogen))、纤连蛋白和层粘连蛋白。因此,在一些实施方案中,用于本发明的方法的ecm包含选自以下列出的一种或多种组分:多糖、弹性蛋白和糖蛋白(例如其中所述糖蛋白包含胶原蛋白、内联蛋白(entactin)(巢蛋白(nidogen))、纤连蛋白和/或层粘连蛋白)。例如,在一些实施方案中,将胶原蛋白用作ecm。不同类型的ecm是已知的,其包含不同的组成,包括不同类型的糖蛋白和/或糖蛋白的不同组合。可以在移除这些细胞和添加分离的组织碎片或分离的上皮细胞之前通过在容器中培养产生ecm的细胞(如例如上皮细胞、内皮细胞、腔壁内胚层样细胞或成纤维细胞)来提供ecm。产生细胞外基质的细胞的实例是软骨细胞(主要产生胶原蛋白和蛋白聚糖)、成纤维细胞(主要产生iv型胶原、层粘连蛋白、间质前胶原和纤连蛋白)和结肠肌成纤维细胞(主要产生胶原蛋白(i型、iii型和v型)、硫酸软骨素蛋白聚糖、透明质酸、纤连蛋白和肌腱蛋白-c。这些是“天然产生的ecm”。可以商业提供天然产生的ecm。可商购获得的细胞外基质的实例包括:细胞外基质蛋白(invitrogen)和来自engelbreth-holm-swarm(ehs)小鼠肉瘤细胞的基底膜制剂(例如基底膜提取物(trevigen,inc.)或matrigeltm(bdbiosciences))。因此,在一些实施方案中,是天然产生的ecm。在一些实施方案中,ecm是含有层粘连蛋白的ecm,如matrigeltm(bdbiosciences)。在一些实施方案中,ecm是matrigeltm(bdbiosciences),其包含层粘连蛋白、内联蛋白和胶原蛋白iv。在一些实施方案中,ecm包含层粘连蛋白、内联蛋白、胶原蛋白iv和硫酸肝素蛋白聚糖(例如2型基底膜提取物(trevigen,inc.))。在一些实施方案中,ecm包含至少一种糖蛋白(如胶原蛋白和/或层粘连蛋白)。用于本发明方法的优选的ecm包含胶原蛋白和层粘连蛋白。进一步优选的ecm包含层粘连蛋白、内联蛋白和胶原蛋白iv。如果需要,可以使用天然产生的或合成的ecm材料的混合物。在另一实施方案中,ecm可以是合成的ecm。例如,可以使用合成的ecm,如pronectin(sigmaz378666)。在另一实例中,ecm可以是塑料(例如聚酯)或水凝胶。在一些实施方案中,可以用生物材料(例如一种或多种糖蛋白,如胶原蛋白或层粘连蛋白)包被合成基质三维ecm支持培养三维上皮类器官。细胞外基质材料通常将是平皿底部的液滴,其中细胞悬浮在其中。通常,当基质在37℃凝固时,添加培养基并扩散进ecm中。培养基中的细胞通过与其表面结构的相互作用(例如与整合蛋白的相互作用)而粘附于ecm。可以将培养基和/或细胞可以置于ecm上,嵌入ecm中或与ecm混合。例如,在一些实施方案中,将单个细胞、细胞群或组织碎片嵌入matrigeltm(其是任选地生长因子减少和/或不含酚红)中。在一些实施方案中,将培养基置于ecm的顶部。然后可以在需要时移除和补充培养基。在一些实施方案中,每1、2、3、4、5、6或7天补充培养基。如果向培养基“添加”或从培养基“移除”组分,则在一些实施方案中,这可以意味着从ecm移除培养基本身,和然后将含有“添加的”组分或具有排除的“移除的”组分的新培养基置于ecm上。用于培养的祖细胞和干细胞和获得所述细胞分化方法可用于祖细胞。祖细胞在本文被限定为具有分化潜能的任何细胞。因此,术语“祖细胞”涵盖干细胞,其包括但不限于成体干细胞、胚胎干细胞和ips细胞。祖细胞可以是例如原代干细胞或扩增的干细胞或部分分化的干细胞。在一些实施方案中,祖细胞是原代细胞,意为直接从活组织获得。在其它实施方案中,祖细胞是次级细胞,即已经培养和/或传代的细胞。在一些实施方案中,祖细胞是扩增的细胞。术语“扩增的”意为在分化之前细胞已在促进细胞扩增(例如增殖)的培养基中体外培养细胞。在优选实施方案中,祖细胞是成体祖细胞,即来源于成体组织。在一些实施方案中,成体祖细胞是成体干细胞(例如成体上皮干细胞)。在该情境下,“成体”包括新生儿或儿童,但排除胚胎或胎儿。在一些实施方案中,祖细胞不来源于胚胎干细胞或胚胎干细胞系(例如人胚胎干细胞或人胚胎干细胞系)。在优选的实施方案中,祖细胞是上皮祖细胞。例如,在优选的实施方案中,祖细胞来源于上皮组织,更优选成体上皮组织。上皮组织包括肝脏、胰腺、肠、胃、前列腺、肺、乳腺、卵巢、唾液腺、毛囊、皮肤、食道、耳、膀胱或甲状腺。因此,在一些实施方案中,祖细胞获自肝脏、胰腺、肠、胃、前列腺、肺、乳腺、卵巢、唾液腺、毛囊、皮肤、食道、耳、膀胱或甲状腺。在一些实施方案中,祖细胞获自胰腺、胃、肺或肠。在优选的实施方案中,祖细胞获自肠。在优选的实施方案中,祖细胞是哺乳动物细胞。例如,在优选的实施方案中,细胞来源于哺乳动物组织。例如,在一些实施方案中,祖细胞是人细胞。在一些实施方案中,祖细胞来自实验动物(例如小鼠、兔、大鼠、豚鼠)、伴侣动物(例如狗、猫、马)或农场动物(例如牛、猪、绵羊、山羊)。原代细胞代表了体内情况的最佳实验模型。在本发明的优选的实施方案中,祖细胞是(或在细胞培养物中来源于)原代祖细胞(例如原代上皮干细胞)。可以将原代细胞培养物传代以形成次级细胞培养物。除癌细胞外,传统的次级上皮细胞培养物具有有限的寿命。在一定数量的群体倍增(例如50-100代)后,细胞经历衰老过程并停止分裂。可以将来自次级培养物的细胞变成永生化以成为连续细胞系。永生化可以自发发生,或者可以是病毒或化学诱导的。永生化细胞系也被称为转化的细胞。在本发明优选的实施方案中,细胞获自扩增的上皮干细胞培养物,优选扩增的类器官,其已经被扩增和/或传代而没有永生化或转化。在一些实施方案中,这些扩增的上皮培养物或类器官可以是遗传异质性的(与传统细胞系不同)。因此,在一些实施方案中,祖细胞不是永生化的细胞或转化的细胞或者不来源于永生化的细胞系或转化的细胞系。可以通过任何合适的方法获得祖细胞,例如如wo2010/090513、wo2012/014076、wo2012/168930或wo2015/173425中所述。在一些实施方案中,例如dorell等人,2008(hepatology.2008oct;48(4):1282-91.surfacemarkersforthemurineovalcellresponse.dorrellc,erkerl,lanxon-cooksonkm,abrahamsl,victorofft,ros,canadayps,streeterpr,grompem)中所述,通过胶原酶消化来分离细胞。在一些实施方案中,在组织活检切片上进行胶原酶消化。在一些实施方案中,使用胶原酶和细胞消化液(accutase)消化来获得用于本发明的祖细胞。在一些实施方案中,祖细胞是表达lgr5的上皮干细胞。优选使用来自成体组织的细胞,优选来自成体组织的上皮干细胞,更优选表达lgr5的上皮干细胞获得类器官。在最优选的实施方案中,祖细胞是或包含表达lgr5的成体上皮干细胞。在一些实施方案中,祖细胞是正常细胞,意为细胞具有正常核型、基因型和/或表型。在可替代的实施方案中,祖细胞是疾病细胞,意为它们具有疾病核型、基因型和/或表型。例如,在一些实施方案中,祖细胞是癌细胞。因此,设想例如上皮干细胞可以是lgr5阳性癌症干细胞。因此,如果需要可以从肿瘤获得细胞。在可替代的实施方案中,祖细胞是患病祖细胞,例如用细胞内病原体(例如细菌、病毒或寄生虫)感染的祖细胞。示例性方法本发明提供了用于分化祖细胞的方法,其中所述方法包括在本文所述的分化培养基中培养细胞。在优选的实施方案中,如本文所述将细胞与ecm接触培养。本发明还提供了用于培养祖细胞的方法,其中所述方法包括在扩增培养基中培养细胞,并随后在本文所述的分化培养基中培养细胞。在一些实施方案中,在扩增和/或分化步骤期间,如本文所述将细胞与ecm接触培养。在一些实施方案中,在分化培养基中培养细胞之前,移除扩增培养基,例如通过反复洗涤或分离细胞培养物。本发明提供了用于培养单个上皮干细胞、上皮干细胞群或分离的组织碎片的方法,优选以获得类器官,其中所述方法包括:在扩增培养基中培养上皮干细胞、上皮干细胞群或分离的组织碎片以提供扩增的细胞群;任选地诱导扩增的细胞群的静止(例如通过用一种或多种egfr通路抑制剂处理);和在分化培养基中培养扩增的细胞群。本发明还提供了用于分化单个祖细胞或祖细胞群的方法,其中所述方法包括:在分化培养基中培养祖细胞或祖细胞群。分化培养基可以是本文描述的任何分化培养基。例如,在一些实施方案中,分化培养基包含wnt抑制剂(例如iwp-2)、notch抑制剂(例如dapt)和egfr通路抑制剂(例如,吉非替尼、阿法替尼、pd0325901和sch772984中的一种或多种)。该分化培养基特别适用于获得富含eec的细胞群的方法。eec特征性标志物包括chga、chgb、tac1、tph1、gip、fabp5、ghrl、pyy、nts、neurod1、sst、sct、胆囊收缩素、胰高血糖素和/或胰高血糖素原。grun等人(2015)nature525:251-255描述了另外的eec细胞类型及其特征。稍后参见表2。因此,在一些实施方案中,本发明的分化方法产生表达一种或多种选自以下的标志物的细胞群:chga、chgb、tac1、tph1、gip、fabp5、ghrl、pyy、nts、neurod1、sst、sct、胆囊收缩素、胰高血糖素和/或胰高血糖素原。在一些实施方案中,通过本发明的方法获得的eec表达血清素。在一些实施方案中,分化培养基包含:wnt抑制剂(例如iwp-2)、notch抑制剂(例如dapt)和egfr通路抑制剂(例如吉非替尼、阿法替尼、pd0325901和sch772984中的一种或多种)以及bmp抑制剂(例如头蛋白或ldn193189)。该分化培养基特别适用于获得富含分泌glp1的eec的细胞群的方法。在一些实施方案中,该方法产生对于选自tac1、glp1和chg的一种或多种标志物呈阳性的细胞群;对于选自分泌素、pyy和nts的一种或多种标记物呈阴性。在一些实施方案中,细胞群表达血清素。在一些实施方案中,分化培养基包含:wnt抑制剂(例如iwp-2)、notch抑制剂(例如dapt)和egfr通路抑制剂(例如吉非替尼、阿法替尼、pd0325901和sch772984中的一种或多种)以及bmp激活剂(例如bmp4)。该分化培养基特别适用于获得富含分泌分泌素的eec的细胞群的方法。在一些实施方案中,该方法产生对于选自tac1、glp1和chg的一种或多种标记物呈阴性的细胞群;对于选自分泌素、pyy和nts的一种或多种标记物呈阳性。在一些实施方案中,细胞群表达血清素。扩增培养基可以是用于上皮干细胞或祖细胞的任何合适的扩增培养基,优选用于上皮干细胞的合适的扩增培养基(例如wo2010/090513、wo2012/014076、wo2012/168930或wo2015/173425中所述)。在一些实施方案中,扩增培养基包含一种或多种受体酪氨酸激酶配体(例如egf、hgf和/或fgf10)、烟酰胺和一种或多种wnt激动剂(例如rspondin条件化培养基和/或wnt条件化培养基)。在一些实施方案中,扩增培养基包含一种或多种受体酪氨酸激酶配体(例如egf、hgf和/或fgf10)、一种或多种wnt激动剂(例如rspondin条件化培养基和/或wnt条件化培养基)和一种或多种tgf-β抑制剂(例如a83-01)。在一些实施方案中,扩增培养基包含一种或多种受体酪氨酸激酶配体(例如egf、hgf和/或fgf10)、烟酰胺、一种或多种wnt激动剂(例如rspondin条件化培养基和/或wnt条件化培养基)和一种或多种tgf-β抑制剂(例如a83-01)。在这些实施方案的任何实施方案中,扩增培养基还可以包含camp通路激活剂(例如毛喉素)、胃泌素和/或bmp抑制剂(例如头蛋白)。例如,在一些实施方案中(例如在用于肠的优选的实施方案中),扩增培养基包含(i)egf(例如约10至50ng/ml);(ii)头蛋白条件培养基(例如约50至100ng/ml或约5%终体积);(iii)rspondin条件培养基(例如约1μg/ml或约5%终体积)。在一些实施方案中,扩增培养基还包含n-乙酰半胱氨酸(例如约1mm)和1xb27。在一些实施方案中,扩增培养基还包含丙戊酸(例如约1mm)和gsk-3抑制剂(例如约3μm,如约3μm的chir99021)。有利地,发现包含丙戊酸和gsk-3抑制剂产生富含干细胞的细胞群。人类器官的优选扩增培养基包含(i)egf(例如约10至50ng/ml);(ii)头蛋白条件培养基(例如约50至100ng/ml或约5%终体积);(iii)rspondin条件培养基(例如约1μg/ml或约5%终体积);(iv)p38抑制剂(例如浓度约30μm的sb-203580);(v)tgf-β抑制剂(例如浓度约500nm的a83-01);(vi)烟酰胺(例如浓度约为10mm)。优选地,在分化之前扩增细胞以产生一种或多种(例如至少2、3、4、5、6、10、15、20或多于20种)类器官。在一些实施方案中,最初在本文所述的扩增培养基中培养细胞,一旦建立成功的类器官,则用本文所述的分化培养基替换扩增培养基。因此,在一些实施方案中,在一次或多次(例如在两次、三次、四次、五次、六次、七次、八次、九次、十次或更多次之后)传代之后,用分化培养基替换扩增培养基。在一些实施方案中,传代每周进行。在一些实施方案中,在2、3、4、5、6、7、8、9、10或更多天后,用分化培养基替换扩增培养基。在优选的实施方案中,在五天或更多天后用分化培养基替换扩增培养基。在一些实施方案中,在分化之前诱导细胞静止。在一些实施方案中,在分化之前,洗涤上皮干细胞、上皮干细胞群或分离的组织片段并涂铺在细胞外基质(例如matrigel)中。在一些实施方案中,用基础培养基或pbs进行洗涤。不希望受任何理论束缚,本发明人认为该洗涤步骤有利于分化,因为其从上皮干细胞、上皮干细胞群或分离的组织片段中去除干细胞因子。在一些实施方案中,上皮干细胞群或上皮干细胞或分离的组织片段在包含bmp抑制剂(例如头蛋白)和rspondin并且不包含egf、烟酰胺、tgfβ抑制剂或wnt条件培养基的分化培养基中,之后在本发明的分化培养基中分化。在一些实施方案中,在第一分化培养基中培养约一天。对于本领域技术人员显而易见的是,本发明优选的培养方法是有利的,因为不需要饲养细胞。通常使用饲养细胞层以支持干细胞的培养,并抑制它们的分化。饲养细胞层通常是与目标细胞共培养的单层细胞,其提供适于目标细胞生长的表面。饲养细胞层提供了其中目标细胞可以生长的环境。饲养细胞通常是有丝分裂失活的(例如通过辐射或用丝裂霉素c处理)以防止它们的增殖。饲养细胞的使用是非期望的,因为它使细胞的传代变得复杂(在每次传代时,必须将细胞与饲养细胞分离,并且在每次传代时需要新的饲养细胞)。饲养细胞的使用还可导致饲养细胞污染所需细胞。这对于任何医学应用显然是有问题的,并且甚至在研究情境下,这使对细胞进行的任何实验的结果分析复杂化。如本文其它地方所指明的,本发明的培养基是特别有利的,因为可以将它们用于在没有饲养细胞接触的情况下培养细胞,即本发明的方法不需要饲养细胞层来支持其生长受到帮助的细胞。因此,本发明的组合物可以是不含饲养细胞的组合物。如果组合物中的细胞在不存在饲养细胞层的情况下已经培养了至少一代,则通常认为组合物不含饲养细胞。本发明的不含饲养细胞的组合物将通常含有小于约5%,小于约4%,小于约3%,小于约2%,小于约1%的饲养细胞(表示为组合物中的细胞总数的%)或者优选根本不含饲养细胞。在另一方面,提供了获得分化细胞群或类器官的方法,其中所述方法包括在本发明的分化培养基中培养祖细胞。优选地,该方法包括使用如本文所述的分化方法在本发明的分化培养基中培养祖细胞。在一些实施方案中,该方法包括从单个上皮干细胞获得类器官/分化细胞群。在另一实施方案中,该方法包括从上皮干细胞群或从上皮组织碎片获得类器官/分化细胞群。在具体的实施方案中,提供了获得分化的类器官的方法,其中所述方法包括在本发明的分化培养基中培养上皮祖细胞(例如上皮干细胞,任选地表达lgr5的上皮干细胞),优选地其中上皮祖细胞与ecm(优选三维ecm)接触。在一些实施方案中,该方法包括在扩增培养基中培养祖细胞一段时间,例如3天至10周,1至10周,1至4周或10天至3周,然后将细胞传代(例如将细胞分离成单细胞密度,以每个容器(例如每个孔)1个细胞的比例接种一个或多个细胞),使用扩增培养基继续扩增细胞一段时间,例如3天至10周,1至10周,1至4周或10天至3周,并在分化细胞之前重复传代和扩增步骤至少一次、至少两次、至少三次、至少四次、至少五次、至少六次、至少七次、至少八次、至少九次、至少十次、至少十一次、至少十二次、至少十三次或至少十四次。在一些实施方案中,该方法包括在分化培养基中培养祖细胞至少1天、至少2天、至少3天、至少4天、至少5天、至少6天、至少7天、至少8天、至少9天、至少10天、至少11天、至少12天、至少13天、至少14天。在一些实施方案中,该方法包括在分化培养基中培养祖细胞约1至约20天、约1至约10天或约1至约5天。分化之后,该方法还可以包括获得和/或分离一种或多种分化的细胞或分化的类器官。例如,在培养祖细胞后,从培养基中移出培养基中培养的一种或多种细胞和/或一种或多种类器官用于后续应用可能有用。可以使用本领域已知的许多物理分离方法中的任何一种来选择本发明的细胞并将它们与其它细胞类型区分开。此类物理方法可以包括基于由本发明的细胞特异性表达的标志物的facs和多种免疫亲和方法。在一个实施方案中,可以利用抗体(例如针对这些标志物之一的抗体)通过facs分离细胞,。对本领域技术人员显而易见的是,这可以通过荧光标记的抗体或者通过对一级抗体具有结合特异性的荧光标记的二级抗体来实现。合适的荧光标记的实例包括但不限于fitc、alexa488、gfp、cfse、cfda-se、dylight488、pe、percp、pe-alexa700、pe-cy5(tri-))、pe-cy5.5、pi、pe-750和pe-cy7。该列表仅作为示例提供,并非旨在进行限制。可替代地,可以通过免疫亲和纯化分离细胞,免疫亲和纯化是本领域众所周知的分离方法。该方法依赖于抗体在纯化柱上的固定。然后将细胞样品加载到柱上,允许适当的细胞与抗体结合,并因此与柱结合。洗涤步骤之后,使用优先与固定化抗体结合的竞争剂从柱上洗脱细胞,并允许细胞从柱中被释放出来。对本领域技术人员显而易见的是,使用固定化抗体的免疫亲和纯化将提供纯化的细胞群。然而,在一些实施方案中,可以优选通过使用一种或多种其它可鉴定的标志物进行又一轮的免疫亲和纯化来进一步纯化细胞群,并使用等分试样的分离的克隆来确定其它相关的细胞内标志物的表达。对本领域技术人员显而易见的是,不一定需要顺序纯化步骤涉及相同的物理分离方法。分化的细胞和类器官本发明还提供了分化的类器官或一种或多种分化细胞群。分化的类器官是包含分化的上皮细胞类型的三维结构。分化的类器官通常是自组织的,这意味着当细胞分化时,类器官中细胞的三维排列自发地发生。在一些实施方案中,分化的类器官来源于上皮干细胞,任选地表达lgr5的上皮干细胞。在一个实施方案中,本发明提供了通过本发明的方法可获得的或获得的分化的类器官或一种或多种分化细胞群,例如所述本发明的方法包括在本发明的分化培养基中培养祖细胞,优选地其中祖细胞是与三维ecm接触的。细胞“群”是大于1的任何数目的细胞,但优选至少10个细胞、至少50个细胞、至少100个细胞、至少500个细胞、至少1x103个细胞、至少1x104个细胞、至少1x105个细胞、至少1x106个细胞、至少1x107个细胞、至少1x108个细胞或者至少1x109个细胞。根据本发明的分化的类器官可以包含具有以下细胞数的细胞群:至少10个细胞,至少50个细胞,至少100个细胞,至少500个细胞,至少1x103个细胞,至少1x104个细胞,至少1x105细胞,至少1×106个细胞,至少1×107个细胞或者更多。在一些实施方案中,每个类器官包含大约1×103个细胞至5×103个细胞;通常,一个孔(例如24孔板的一个孔)中可以一起生长10-20个类器官。技术人员清楚的是,本发明的类器官不是天然存在的组织碎片和/或不包含血管。例如,本发明的类器官与天然存在的组织不同,因为它们仅包含上皮细胞类型(并且不包含间充质细胞或其它结构细胞类型)。因此,在一些实施方案中,本发明的分化的类器官仅包含上皮细胞。在一些实施方案中,分化的类器官不包含非上皮细胞。例如,在具体的实施方案中,分化的类器官不包含间充质细胞。本文所述的分化培养基优选在培养至少五天期间诱导或促进细胞的特异性分化。如本文所限定的,可以通过检测与特定组织谱系(例如肠内分泌谱系)相关的特定标志物的存在来判断分化。如本文所限定的,可以通过检测与组织谱系(例如肠内分泌谱系)相关的特定标志物的存在来判断分化。根据标志物的特性,可以在如本文所限定的分化培养基中培养至少5、7、8、9、10、11、12、13、14、15、16或更多天之后通过rtpcr或免疫组织化学来评估所述标志物的表达。术语“表达”被用于描述细胞内标志物的存在。为了被认为是表达,标志物必须以可检测的水平存在。“可检测的水平”意为可以使用标准实验室方法之一(如pcr、印迹或facs分析)来检测标志物。如果在30个pcr循环(这对应于每个细胞至少约100个拷贝的细胞中的表达水平)后可以合理地检测到表达,则认为基因由本发明群体的细胞表达。术语“表达(express)”和“表达(expression)”具有一致的含义。在低于该阈值的表达水平,认为标志物不被表达。可以优选通过比较已经从相同物种中分离出来的两种细胞类型来进行本发明的细胞中的标志物的表达水平与另一细胞(如例如胚胎干细胞)中相同标志物的表达水平之间的比较。优选地,该物种是哺乳动物,并且更优选地该物种是人。可以使用逆转录酶聚合酶链式反应(rt-pcr)实验方便地进行此类比较。本发明的分化的类器官或本发明的分化的细胞群优选包含至少50%的活细胞,更优选至少60%的活细胞,更优选至少70%的活细胞,更优选至少80%的活细胞,更优选至少90%的活细胞。可以在facs中使用hoechst染色或中的碘化丙啶染色来评估细胞的活力。活细胞优选具有相应的体内功能或特性。例如,活的肠内分泌细胞优选具有肠内分泌功能或肠内分泌细胞的特征。在优选的实施方案中,类器官是肠类器官。这意味着类器官源自肠细胞。在优选的实施方案中,本发明的分化细胞群源自肠细胞。发明人已经表明,通过本发明的方法获得的肠类器官得到了改善,因为相较于先前在grün等人(2015)nature525(7568):251-5中所述的分化的肠类器官中,这些类器官中更大比例的细胞是肠内分泌细胞。在哺乳动物胃肠道中发现的一些肠内分泌细胞类型总结在下表2中。表2:哺乳动物胃肠道的肠内分泌细胞eec特征性标志物包括chga、chgb、tac1、tph1、gip、fabp5、ghrl、pyy、nts、neurod1、sst、sct、胆囊收缩素、胰高血糖素和/或胰高血糖素原。grun等人(2015)nature525:251-255描述了其他eec细胞类型及其特征。在一些实施方案中,本发明的分化的类器官是分化的肠类器官,其中至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或至少99%的细胞表达肠内分泌细胞标志物(例如chga、chgb、tac1、tph1、gip、fabp5、ghrl、pyy、nts、neurod1、sst、sct、胆囊收缩素、胰高血糖素和/或胰高血糖素原)。在一些实施方案中,通过单细胞rna测序分析测量mrna表达。在一些实施方案中,本发明的分化的类器官是分化的肠类器官,其中至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或至少99%的细胞表达特定肠内分泌细胞类型、特别是表2中所述的细胞类型的特征性肠内分泌细胞标记物。例如,在一些实施方案中,特定肠内分泌细胞类型的特征性肠内分泌细胞标记物是gcg和/或glp-1。gcg是表达glp-1来源的前胰高血糖素原的基因。在其他实施方案中,特定肠内分泌细胞类型特征性肠内分泌细胞标记物是sct。在一些实施方案中,本发明的分化的类器官是分化的肠类器官,其中至少10%、至少20%、至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%或至少99%的细胞表达表2中列出的一种或多种(例如2、3、4或更多种)产物。在一些实施方案中,本发明的分化的类器官是分化的肠类器官,其中小于50%、小于40%、小于30%、小于20%、小于10%或小于1%的细胞表达goblet或帕内特细胞标志物(例如lyz1、defa6、agr2、gob5、muc2、ttf3和/或defa24)。在一些实施方案中,通过单细胞rna测序分析测量mrna表达。在一些实施方案中,本发明的分化的类器官是分化的肠类器官,其中小于50%、小于40%、小于30%、小于20%、小于10%或小于1%的细胞表达肠上皮细胞标志物(例如aldob、apoa1和/或alpi)。在一些实施方案中,通过单细胞rna测序分析测量mrna表达。在一些实施方案中,本发明的分化的类器官是分化的肠类器官,其中小于50%、小于40%、小于30%、小于20%、小于10%或小于1%的细胞表达tuft细胞标志物(例如dclk1和/或trpm5)。在一些实施方案中,通过单细胞rna测序分析测量mrna表达。在一些实施方案中,分化的类器官具有带有中央腔的囊性结构。在一些实施方案中,中央腔被上皮单层包围。还提供了在本发明的分化培养基中的本发明的分化的类器官或分化的肠内分泌细胞群。在一个实施方案中,提供了在分化培养基中的类器官(例如,如本文所述)。在实施方案中,分化的类器官是仍然使用本发明的方法培养并因此与细胞外基质接触的类器官。优选地,将分化的类器官嵌入非间充质细胞外基质中。类器官或祖细胞群可以来自任何哺乳动物组织,但优选来自人。在一些实施方案中,它来自小鼠、兔、大鼠、豚鼠或其它非人哺乳动物。表3-分化的肠类器官与原代肠组织之间的差异分化的类器官原代组织细胞组成约30-80%eec约1%eec分化的类器官的用途同样提供了本文所述的类器官和来源于所述类器官的细胞的用途。当在本部分提及类器官时,这些是根据本发明的分化的类器官。本领域技术人员将会理解,其用途也可适用于通过本发明的方法获得的和/或可获得的分化细胞群。还提供了通过本发明的方法获得的和/或可获得的分化的细胞群的此类用途。例如,本发明提供了分化的类器官或来源于所述类器官的细胞在下述中的用途:药物发现筛选;毒性测定;组织胚胎学、细胞谱系和分化途径的研究;鉴别导致各自激素释放的化学和/或神经元信号的研究;包括重组基因表达的基因表达研究;组织损伤和修复中涉及的机制研究;炎性和传染性疾病研究;发病机制研究;或者细胞转化和癌症病因机制的研究。本发明还提供了本发明的类器官或来源于所述类器官的细胞,其用于医学中的用途。本发明还提供了本发明的类器官或来源于所述类器官的细胞,其用于治疗病症、病况或疾病中的用途。本发明还提供了本发明的类器官或来源于所述类器官的细胞,其用于再生医学中的用途,例如其中所述用途涉及将类器官或细胞移植到患者体内。本发明提供了本发明的类器官或来源于所述类器官的细胞在药物筛选、(药物)靶标验证、(药物)靶标发现、毒理学和毒理学筛选、个性化医学、再生医学和/或作为离体细胞/器官模型(如疾病模型)中的用途。认为根据本发明的培养基和方法培养的细胞和类器官忠实地代表了体内情况。这对于生长自正常组织的分化的细胞群和类器官以及对于生长自患病组织的分化的细胞群和类器官都是如此。因此,除了提供正常的离体细胞/器官模型外,可以将本发明的类器官用作离体疾病模型。本发明的类器官也可以被用于培养病原体,并因此可以被用作离体感染模型。可以使用本发明的类器官进行培养的病原体的实例包括在其动物宿主中引起疾病的病毒、细菌、朊病毒或真菌。因此,本发明的类器官可以被用作代表感染状态的疾病模型。在本发明的一些实施方案中,可以将类器官用于疫苗开发和/或生产。因此,可以通过本发明的类器官研究的疾病包括遗传性疾病、代谢性疾病、病原性疾病、炎性疾病等,例如包括但不限于:糖尿病(如i型或ii型)、囊性纤维化、癌、腺癌、腺瘤、胃肠胰腺神经内分泌肿瘤、炎症性肠病(如克罗恩氏病(crohn’sdisease))。传统上,细胞系和最近的ips细胞已被用作离体细胞/器官和/或疾病模型(例如参见robinton等人,nature481,295,2012)。然而,这些方法存在许多挑战和缺点。例如,不能从所有患者获得细胞系(仅某些活组织检查产生成功的细胞系),并因此细胞系不能用于个性化诊断和医学。ips细胞通常要求一定水平的遗传操作以将细胞重编程进特定的细胞命运。可替代地,使它们经受影响核型完整性的培养条件,并且因此培养时间必须保持最短(人胚胎干细胞也是如此)。这意味着ips细胞不能准确地代表体内情况,而是试图模拟体内细胞的行为。细胞系和ips细胞也经受遗传不稳定性。相比之下,本发明的类器官提供了忠实地代表体内情况的遗传稳定的平台。在一些实施方案中,本发明的类器官包含存在于相应体内环境中的所有分化细胞类型。在其它实施方案中,可以进一步分化本发明的类器官以提供体内存在的所有分化的细胞类型。因此,可以将本发明的类器官用于获得对多种疾病和治疗的机理洞察,以进行体外药物筛选,评估潜在的治疗,鉴定用于未来新型(药物)疗法开发的可能靶标(例如蛋白质)和/或探索与细胞替代疗法相结合的基因修复。可以冷冻和解冻本发明的类器官并投入培养而不丧失其遗传完整性或表型特性并且不丧失增殖能力。因此,可以容易地储存和运输类器官。因此,在一些实施方案中,本发明提供冷冻的类器官。由于这些原因,本发明的类器官或分化的细胞群可以是用于药物筛选、靶标验证、靶标发现、毒理学和毒理学筛选以及个性化医学的工具。因此,在另一方面,本发明提供了根据本发明的类器官或来源于所述类器官的细胞在药物发现筛选、毒性测定或在医学(如再生医学)中的用途。例如,可将肠类器官中的任何一种用于药物发现筛选、毒性测定或用于医学(如再生医学)中。粘膜疫苗类器官的另一个重要用途是开发粘膜接种疫苗。粘膜疫苗是经粘膜施用的疫苗。这可以是任何粘膜表面,如经鼻、口或直肠。它们可以经吸入器、喷雾器或其它外部辅助施用。与注射相比,这具有几个明显的益处,如不需要医务人员来施用疫苗,例如在发展中国家,这可能是重要的。在肠中,m细胞(或“微皱褶细胞”)是在回肠聚集的淋巴结的滤泡相关上皮中发现的细胞。它们将来自肠腔的生物体和颗粒穿过上皮屏障运输到免疫细胞,并因此在刺激粘膜免疫中是重要的。它们具有经由内吞作用或吞噬作用从小肠腔摄取抗原的独特能力,然后经由转胞吞作用将其递送到位于它们基底侧的独特口袋样结构的树突状细胞(抗原递呈细胞)和淋巴细胞(即t细胞)。m细胞是一类肠内分泌细胞。如上所述,本发明人发现了一种用于将祖细胞分化为肠内分泌细胞命运的改进方法。因此,本发明的分化的类器官富含肠内分泌细胞,例如m细胞。在一些情况下,当用rank配体刺激时,类器官可以发育成m细胞(例如参见wo2012/169830的图49)。因此,在一些实施方案中,分化的培养基还包含rank配体。当粘膜疫苗靶向m细胞时,它们的效率可以被显著提高。因此,可以将本发明的分化的细胞群或类器官用于测试m细胞摄取病原体或抗原并将其呈递给免疫系统的能力。因此,在一些实施方案中,本发明提供了本发明的类器官在药物筛选(例如在疫苗开发和/或疫苗生产)中的用途。例如,在一些实施方案中,可以将类器官用于开发或生产针对病毒、细菌、真菌或其它寄生虫感染(例如(但不限于)霍乱、呼吸道合胞体病毒(rsv)、轮状病毒和hiv)的疫苗。在具体的实施方案中,本发明提供了在本发明的培养基中已经分化的类器官在粘膜疫苗开发中的用途。药物筛选为了优选地高通量目的,将本发明所述类器官在多孔板(例如96孔板或384孔板)中培养。将分子文库用于鉴定影响所述类器官的分子。优选的文库包括抗体片段文库、肽噬菌体展示文库、肽文库(例如lopaptm,sigmaaldrich)、脂质文库(biomol)、合成化合物文库(例如lopactm,sigmaaldrich)或天然化合物文库(specs,timtec)。此外,可以使用遗传文库来诱导或抑制干细胞子代中一种或多种基因的表达。这些遗传文库包含cdna文库、反义文库和sirna或其它非编码rna文库。优选将细胞暴露于多种浓度的测试试剂一段时间。在暴露期结束时,评估培养物。术语“影响”用于覆盖细胞的任何变化,包括但不限于增殖的减少或丧失、形态变化和细胞死亡。还可以将本发明的所述类器官用于鉴定特异性靶向上皮癌细胞而非本发明所述的类器官的药物。在短时间(天)内获得本发明的可用类器官的能力表明,类器官对于测试个体患者对特定药物的响应并根据响应性定制治疗非常有用。在一些实施方案中,其中类器官获自患者的活组织检查,将所述类器官培养小于21天、例如小于14天、小于13天、小于12天、小于11天、小于10天、小于9天、小于8天、小于7天(等)。类器官也可用于更广泛的药物发现目的(例如参见wo2013/093812,其描述了筛选用于囊性纤维化或霍乱的药物)。因此,在一些实施方案中,可以将本发明的类器官用于筛选囊性纤维化药物。然而,本领域技术人员将会理解,本发明的类器官将广泛适用于下述的药物筛选工具:人胃肠道的感染性、炎症性和肿瘤病理学和胃肠道的其它疾病以及本文所述的其它组织(如胰腺、胃或肺)的感染性、炎症性和肿瘤病理学以及其它疾病。在一些实施方案中,可以将本发明的类器官用于筛选癌症药物。在一些实施方案中,可以将本发明的类器官用于测试化学品、抗体、天然产物(植物提取物)等的文库用作药物、化妆品和/或预防性药物的适用性。例如,在一些实施方案中,可以使用本发明的培养基和方法培养来自目标患者的细胞活组织检查(如来自癌症患者的肿瘤细胞),然后用化学化合物或化学文库对其进行处理。然后可以确定哪些化合物有效地修饰、杀死和/或处理患者的细胞。这允许测试具体患者对特定药物的响应性,从而允许针对具体患者定制治疗。因此,这允许个性化医学方法。以这种方式使用类器官鉴定药物的附加优点是它还可以筛选正常的类器官(来源于健康组织的类器官)以检查哪些药物和化合物对健康组织的影响最小。这允许筛选具有最小脱靶活性或非期望的副作用的药物。可以以这种方式筛选用于许多疾病的药物。例如,可以将本发明的类器官用于筛选用于糖尿病、囊性纤维化、癌、腺癌、腺瘤、胃肠胰腺神经内分泌肿瘤、炎症性肠病(如克罗恩氏病)等的药物。测试参数取决于目标疾病。例如,当筛选用于癌症的药物时,癌细胞死亡通常是最终目标。对于囊性纤维化,测量在针对药物和cftr的刺激的响应中类器官的扩增是目的。在其它实施方案中,可以评估代谢物或基因表达以研究筛选的化合物和药物对目标细胞或类器官的影响。因此,本发明提供了筛选治疗性或预防性药物或化妆品的方法,其中所述方法包括:使分化的细胞群或类器官与候选分子(或候选分子库)接触,评估所述分化的细胞群或类器官的任何影响(例如细胞的任何变化,如增殖的减少或丧失、形态变化和/或细胞死亡)或类器官的变化(例如类器官大小或运动性);将导致所述影响的候选分子鉴定为潜在的药物或化妆品;以及任选地将所述候选分子制备为药物或化妆品。在一些实施方案中,本发明提供了制备药物或化妆品的方法,其中所述方法包括:使分化的细胞群或类器官与候选分子(或候选分子库)接触,评估所述分化的细胞群或类器官的任何影响(例如细胞的任何变化,如增殖的减少或丧失、形态变化和/或细胞死亡)或类器官的变化(例如类器官大小或运动性);将导致所述影响的候选分子鉴定为潜在的药物或化妆品;以及任选地将所述候选分子制备为药物或化妆品。在一些实施方案中,采用计算机或机器人辅助的培养和数据收集方法来增加筛选通量。在一些实施方案中,类器官来源于患者活组织检查。在一些实施方案中,向所述患者施用引起对培养的分化细胞群(例如类器官)的期望影响的候选分子。因此,在一个方面,提供了治疗患者的方法,其包括:(a)从患者的目标患病组织获得活组织检查切片;(b)培养活组织检查切片以获得类器官;(c)使用本发明的筛选方法筛选合适的药物;以及(d)用步骤(c)中获得的药物治疗所述患者。在一些实施方案中,将药物或化妆品用于治疗、预防或改善遗传性疾病、代谢性疾病、病原性疾病、炎性疾病等的症状,例如包括但不限于:囊性纤维化、炎症性肠病(如克罗恩氏病)、癌、腺瘤、腺癌、结肠癌、糖尿病(如i型或ii型)、胃肠胰腺神经内分泌肿瘤等。在一些实施方案中,本发明提供了用于筛选用于再生医学的药物的方法。靶标发现在一些实施方案中,可以将本发明的类器官用于靶标发现。可以将来源于健康或患病组织的类器官细胞用于靶标识别。可以将本发明的类器官用于发现以下疾病的药物靶标:囊性纤维化、炎症性肠病(如克罗恩氏病)、癌、腺瘤、腺癌、结肠癌、糖尿病(如i型或ii型)、胃肠胰腺神经内分泌肿瘤等。根据本发明的培养基和方法培养的类器官被认为忠实地代表了体内情况。因此,它们可以成为发现特定疾病的新的(分子)靶标的工具。为了寻找新的药物靶标,可以将化合物文库(如sirna)用于转导细胞并使特定基因失活。在一些实施方案中,用sirna转导细胞以抑制(大)基因组的功能。可以使用基因组的任何功能性读出或特定细胞功能来确定靶标是否与研究相关。可以使用本领域众所周知的分析确定疾病特异性读出。例如,分析细胞增殖以测试癌症中涉及的基因。例如,可以将如本文所述的topflash分析用于检测由sirna抑制引起的wnt活性变化。当生长减少或细胞死亡发生时,可以通过本领域已知的方法鉴定相应的sirna相关基因。这些基因是抑制这些细胞生长的可能靶标。在鉴定时,将需要通过本领域众所周知的方法确定所鉴定靶标对研究的细胞过程的特异性。使用这些方法,可以将新分子鉴定为疗法的可能药物靶标。靶标和药物验证筛选可以将从患病和/或正常组织获得的患者特异性类器官用于高通量筛选中鉴定的分子的靶标验证。对于在高通量筛选中被鉴定为可能的治疗药物的化合物的验证也是如此。在类器官培养系统中分化的原代患者材料的使用可用于测试来自高通量药物发现细胞系研究的假阳性等。在一些实施方案中,可以将本发明的类器官用于验证在高通量筛选中已被鉴定为可能的药物或化妆品的化合物。培养病原体此外,可以将本发明的类器官用于培养病原体,如目前缺乏合适的组织培养物或动物模型的诺如病毒。再生医学和移植本发明提供了类器官在再生医学和/或移植中的用途。本发明还提供了治疗方法,其中该方法包括将类器官移植到动物或人体内。本发明的类器官(如胃类器官、肠类器官或胰腺类器官)可用于再生医学,例如用于肠上皮的放射后和/或手术后修复、罹患炎症性肠病(如克罗恩氏病和溃疡性结肠炎)的患者中肠上皮的修复,以及罹患短肠综合征的患者中肠上皮的修复。另外的用途存在于在患有小肠/结肠遗传性疾病患者的肠上皮修复中。包含胰腺类器官的培养物也可用于再生医学,例如作为胰腺或其部分切除之后的移植物,以及用于治疗糖尿病(如i型糖尿病和ii型糖尿病)。在可替代的实施方案中,将类器官或从类器官中分离的细胞重编程为相关的组织命运,例如包括胰腺β-细胞在内的胰腺细胞。本发明的培养方法将使得能够分析将密切相关的祖细胞转分化为胰腺细胞(包括胰腺β-细胞或肝细胞)的因子。技术人员清楚的是,可以将基因疗法另外用于旨在修复受损或疾病组织的方法中。例如,可以使用腺病毒或逆转录病毒基因递送载体来向干细胞递送遗传信息(如dna和/或rna)。技术人员可以替换或修复基因疗法中靶向的特定基因。例如,可以将正常基因插入基因组内的非特异性位置以替换非功能基因。在另一实例中,可以通过同源重组将异常基因序列替换为正常基因序列。可替代地,选择性回复突变可使基因恢复其正常功能。另一实例是改变具体基因的调控(基因打开或关闭的程度)。优选地,通过基因治疗方法离体处理类器官细胞或来源于类器官的细胞,并随后转移至哺乳动物,优选需要治疗的人。由于可以在没有任何明显限制或遗传损害的情况下扩增从成年捐赠者获取的小活组织检查切片,因此该技术可用于产生用于再生目的的可移植上皮。可以将类器官冷冻和解冻并投入培养而不丧失它们的3d结构和完整性并且没有显著的细胞死亡的事实进一步增加了类器官用于移植目的的适用性。此外,在一些实施方案中,可将嵌入ecm中或与ecm接触的类器官移植进哺乳动物中,优选移植进人体内。在另一实施方案中,可以将类器官和ecm同时移植进哺乳动物中,优选移植进人体内。技术人员将会理解,可以将ecm用作3d支架以获得包含根据本发明的扩增的细胞群或类器官的组织样结构。然后可以通过本领域众所周知的方法将此类结构移植进患者体内。可以使用ecm蛋白(如胶原蛋白和/或层粘连蛋白)合成制备ecm支架,或者可替代地可以通过“脱细胞化(decellularising)”分离的器官或组织碎片以留下由ecm组成的支架而获得ecm支架(例如参见macchiarini等人,thelancet,372卷,9655期,第2023-2030页,2008)。在一些实施方案中,可以通过使器官或组织碎片脱细胞化来获得ecm支架,其中任选地所述器官或组织碎片来自肠、胰腺、肝脏或胃。本发明提供了本发明的类器官或来源于所述类器官的细胞在移植进哺乳动物,优选移植进人体内的用途。还提供了治疗需要移植的患者的方法,其包括将本发明的类器官或来源于所述类器官的细胞移植进所述患者体内,其中所述患者是哺乳动物,优选人。在一些实施方案中,在移植进所述患者之前,进一步分化类器官。例如,从成年捐赠者获取小活组织检查切片并通过扩增方法扩增,并随后根据本发明进行分化。因此,可以将本文提供的技术用于产生用于再生目的的可移植上皮。本发明提供了治疗患者的胰岛素缺乏病症(如糖尿病)或者治疗患有胰腺功能障碍的患者的方法,其包括将本发明的胰腺类器官或来自本发明的胰腺类器官的细胞移植进患者体内。在一些实施方案中,细胞或类器官在移植进患者中时不表达或分泌胰岛素,但在患者体内分化以使它们分泌胰岛素。例如,分泌胰岛素的能力可能在移植之后不能立即检测到,但可能在移植之后约一个月存在,例如移植之后6周、2个月或3个月。患者优选是人,但也可以是非人哺乳动物(如猫、狗、马、牛、猪、绵羊、兔或小鼠)。因此,在本发明的范围包括通过细胞疗法治疗人或非人动物患者的方法。此类细胞疗法涵盖通过任何适当的方法将本发明的干细胞或类器官应用于患者。具体地,此类治疗方法涉及受损组织的再生。根据本发明,可以用同种异体或自体干细胞或类器官治疗患者。“自体”细胞是来源于相同生物体的细胞,它们被重新引入其中用于细胞疗法,例如为了允许组织再生。然而,细胞不一定分离自与它们被引入的组织相同的组织。自体细胞不需要与患者匹配以克服排斥问题。“同种异体”细胞是来源于不同于细胞被引入以用于细胞疗法(例如为了允许组织再生)的个体的个体的细胞,尽管是相同的物种。仍可能需要某种程度的患者匹配以防止排斥问题。因此,在一些实施方案中,移植涉及自体细胞。在一些实施方案中,移植涉及同种异体细胞。通常,通过注射或植入将本发明的细胞或类器官引入患者体内。通常将细胞直接注射进其中它们要作用的组织中。可替代地,将通过门静脉注射细胞。含有本发明的细胞和药学上可接受的载体的注射器被包括在本发明的范围内。与含有本发明的细胞和药学上可接受的载体的注射器相连的导管被包括在本发明的范围内。技术人员将能够根据待移植的材料(即细胞群、细胞悬液中的单个细胞、类器官或类器官碎片)以及待治疗的器官来选择合适的施用方法和通路。如上所讨论的,可以将本发明的类器官或细胞用于组织再生。为了实现该功能,可以根据细胞在体内的位置将细胞直接注射或植入它们可以繁殖并最终分化成所需的细胞类型处的受损组织中。可替代地,可以将类器官直接注射或植入受损组织中。易于治疗的组织包括所有受损组织,特别包括那些可能因疾病、损伤、创伤、自身免疫反应或者通过病毒或细菌感染而受损的那些组织。在本发明的一些实施方案中,本发明的细胞或类器官被用于再生结肠、小肠、肺、胰腺或胃系统。例如,在一个实施方案中,使用汉密尔顿注射器将本发明的细胞或类器官注射进患者体内。技术人员将会意识到对于待治疗的具体病况本发明的细胞或类器官的适当剂量是多少。在一个实施方案中,本发明的类器官或细胞,无论是在溶液中,微球中还是多种组合物的微粒中,都将被施用进动脉灌注需要再生的组织或受损器官部分。通常,将使用导管进行此类施用。导管可以是用于血管成形术和/或细胞递送的多种球状导管之一,或者是被设计用于向身体特定部位递送细胞的特定目的的导管。对于某些用途,可以将细胞或类器官包封到由许多不同的可生物降解的化合物制成的并且直径为约15μm的微球中。该方法可以允许血管内施用的细胞或类器官以保留在损伤部位,而不是通过毛细血管网络并进入第一阶段的体循环。毛细血管网络的动脉侧的保留也可以促进它们易位到血管外的空间。在另一实施方案中,可以将类器官或细胞逆行注射到血管树中,或者通过静脉将它们递送到整个身体,或者局部注射到流入细胞或类器官所针对的组织或身体部位的特定静脉中。对于该实施方案,可以使用许多上述制剂。在另一实施方案中,可以将本发明的细胞或类器官植入粘附于生物相容性植入物的受损组织中。在该实施方案中,细胞可以在植入患者体内之前体外粘附于生物相容性植入物上。本领域技术人员将会清楚,在植入之前,可以使用多种粘附物中的任何一种将细胞粘附到植入物上。仅举例而言,此类粘附物可以包括纤维蛋白、整合蛋白家族的一个或多个成员、钙粘蛋白家族的一个或多个成员、选择蛋白家族的一个或多个成员、一种或多种细胞粘附分子(cam)、免疫球蛋白家族中的一种或多种以及一种或多种人工粘附物。该列表仅以说明的方式提供,并且不旨在具有限制性。本领域技术人员将会清楚,可以使用一种或多种粘附物的任何组合。在另一实施方案中,在将基质植入患者体内之前,可以将本发明的类器官或细胞嵌入基质中。通常,将基质植入患者的受损组织中。基质的实例包括基于胶原蛋白的基质、基于纤维蛋白的基质、基于层粘连蛋白的基质、基于纤连蛋白的基质和人造基质。该列表仅以说明的方式提供,并且不旨在具有限制性。在又一实施方案中,可以将本发明的类器官或细胞与基质形成组分一起植入或注射到患者体内。这可以允许细胞在注射或植入之后形成基质,确保细胞或类器官保留在患者体内的适当位置。基质形成组分的实例包括纤维蛋白胶液体烷基、氰基丙烯酸酯单体、增塑剂、多糖(如葡聚糖)、含环氧乙烷的寡聚物、嵌段共聚物(如泊洛沙姆和普朗尼克类)、非离子表面活性剂(如吐温和triton'8'),以及人造基质形成组分。该列表仅以说明的方式提供,并且旨在具有限制性。本领域技术人员将会清楚,可以使用一种或多种基质形成组分的任何组合。在又一实施方案中,可以将本发明的类器官或细胞包含在微球内。在该实施方案中,可以将细胞包封在微球的中心内。同样在该实施方案中,可以将细胞嵌入微球的基质材料中。基质材料可包括任何合适的生物可降解聚合物,包括但不限于藻酸盐、聚乙二醇(plga)和聚氨酯。该列表仅作为示例提供,并且不旨在具有限制性。在又一实施方案中,可以将本发明的细胞或类器官粘附于用于植入的医疗装置上。此类医疗装置的实例包括支架、针、缝线、分离器(splits)、起搏器、假关节、人造皮肤和杆。该列表仅以说明的方式提供,并且不旨在具有限制性。本领域技术人员将会清楚,可以通过多种方法使细胞粘附到医疗装置上。例如,可以使用纤维蛋白、整合蛋白家族的一个或多个成员、钙粘蛋白家族的一个或多个成员、选择蛋白家族的一个或多个成员、一种或多种细胞粘附分子(cam)、一种或多种免疫球蛋白家族以及一种或多种人工粘附物使细胞或类器官粘附到医疗装置上。该列表仅以说明的方式提供,并且不旨在具有限制性。本领域技术人员将会清楚,可以使用一种或多种粘附物的任何组合。使用本发明的方法获得的类器官或分化的细胞群具有多种用途。例如,本发明提供了如本文所述的类器官或分化的细胞群在下述中的用途:药物发现筛选;毒性测定;胚胎学、细胞谱系和分化途径的研究;鉴别导致各自激素释放的化学和/或神经元信号的研究;包括重组基因表达的基因表达研究;损伤和修复中涉及的机制研究;炎性和传染性疾病研究;发病机制研究;或者细胞转化和癌症病因机制的研究。在一个方面,本发明提供了如本文所述的类器官或分化的细胞群在药物发现筛选、毒性测定或再生医学中的用途。类似地,本发明提供了本发明的类器官子代在这些用途中的用途。毒性测定可以是使用类器官或其部分或来源于类器官的细胞的体外分析。这些子代和类器官易于培养并且比例如当前用于毒性测定的上皮细胞系(如caco-2(atcchtb-37)、i-407(atccccl6)和xbf(atcccrl8808))更类似于原代上皮细胞。预期用类器官获得的毒性结果更类似患者中获得的结果。使用基于细胞的毒性测试来确定器官特异性细胞毒性。在所述测试中测试的化合物包含癌症化学预防剂、环境化学品、食品补充剂和潜在毒物。将细胞暴露于多种浓度的测试试剂一段时间。在初步分析中使用5天的暴露和来自最高可溶性浓度的对数稀释来确定分析中测试试剂的浓度范围。在暴露期结束时,评估培养物对生长的抑制。分析数据以确定抑制50%终点的浓度(tc50)。例如,根据本发明的该方面,可以使候选化合物与如本文所述的细胞或类器官接触,并且可以监测细胞或细胞活性的任何变化。出于高通量的目的,在多孔板(如例如96孔板或384孔板)中培养所述类器官。将分子文库用于鉴定影响所述类器官的分子。优选的文库包含抗体片段文库、肽噬菌体展示文库、肽文库(例如lopaptm,sigmaaldrich)、脂质文库(biomol)、合成化合物文库(例如lopactm,sigmaaldrich)或天然化合物文库(specs,timtec)。此外,可以使用遗传文库来诱导或抑制腺瘤细胞后代中一种或多种基因的表达。这些遗传文库包括cdna文库、反义文库和sirna或其它非编码rna文库。优选将细胞暴露于多种浓度的测试剂一段时间。在暴露期结束时,评估培养物。术语“影响”用于覆盖细胞的任何变化,包括但不限于增殖的减少或丧失、形态变化和细胞死亡。还可以将所述类器官用于鉴定特异性靶向上皮癌细胞而非所述类器官的药物。根据本发明的类器官还可以在药物发现筛选和潜在新药或已知药物或者已知或新型食品补充剂的毒性测定中替代细胞系(如caco-2细胞)的使用。此外,可以将此类类器官用于培养病原体。本发明还提供了本发明的分化的类器官或本发明的分化的细胞群在治疗中的用途。还提供了本发明的分化类器官或来源于所述类器官的细胞在治疗如本文所述的疾病或病况中的用途。类似地,提供了治疗如本文所述的疾病或病况的方法,其包括施用一种或多种本发明的类器官或来源于所述类器官的细胞。发明人还证明了将类器官成功移植进免疫缺陷小鼠中(参见wo2012/014076的实施例7),其中移植的肝脏类器官源性细胞在体内产生胆管上皮细胞和肝细胞。因此,在一个实施方案中,本发明提供了用于移植进人或动物的本发明的类器官或类器官源性细胞。本发明的类器官的优点是它们可以被冷冻并随后解冻而不丧失功能。这使得能够进行细胞存储、易于存储和急性用途的快速可用性。例如,这可用于制备“现成的”产品,例如,在肝脏的情况下,可以被用于治疗急性肝脏毒性的“现成的”产品。类器官也可以生长自作为取自活体捐赠者的小活组织检查切片的细胞或组织碎片,从而使对治疗的任何伦理反对最小化。捐赠者甚至可以来自待治疗的患者,这可以减少与外来细胞和器官移植相关的任何负面副作用,并减少对免疫抑制药物的需求。药物制剂在一些实施方案中,本发明还提供了药物制剂,其包含本文所述的分化培养基的组分和药学上可接受的稀释剂和/或赋形剂。例如,提供了包含一种或多种wnt抑制剂(例如iwp-2)、一种或多种egfr通路抑制剂(例如,吉非替尼、阿法替尼、pd0325901和sch772984中的一种或多种)、一种或多种notch抑制剂(例如dapt)和药学上可接受的稀释剂和/或赋形剂的药物制剂。在优选的实施方案中,药物制剂不包含基础培养基。在一些实施方案中,药物制剂不包含细胞外基质。设想这样的制剂可以适合于促进体内干细胞的分化,例如用于再生疗法。此类制剂可原位施用(例如在组织损伤部位处)或全身施用。或者,可以配制制剂,使其适于通过本领域已知的任何施用途径施用,例如静脉内、皮下、肌内施用、粘膜、真皮内、皮内、口服和经眼。因此,药物制剂可以是适于这种施用的任何形式,例如片剂、输液、胶囊剂、糖浆剂等。在一些实施方案中,提供了药物制剂,其包含一种或多种wnt抑制剂(例如iwp-2)、一种或多种egfr通路抑制剂(例如,吉非替尼、阿法替尼、pd0325901和sch772984中的一种或多种)、一种或多种notch抑制剂(例如dapt)、一种或多种bmp抑制剂(例如dorsomorphin或ldn193189)和药学上可接受的稀释剂和/或赋形剂。在一些实施方案中,提供了药物制剂,其包含一种或多种wnt抑制剂(例如iwp-2)、一种或多种egfr通路抑制剂(例如,吉非替尼、阿法替尼、pd0325901和sch772984中的一种或多种)、一种或多种notch抑制剂(例如dapt)、一种或多种bmp激活剂(例如bmp4、bmp7或bmp2)和药学上可接受的稀释剂和/或赋形剂。治疗方法调节体内激素水平还提供了涉及分化培养基的一种或多种组分和/或本发明的药物组合物的治疗方法。特别地,设想体内eec可以针对表达特定激素的特定eec表型,并因此在一些实施方案中,本发明的方法可以用于调节体内激素水平。例如,本发明人已经表明bmp激活剂促进eec中的分泌素分泌(和抑制glp-1分泌)(参见实施例5)。因此设想bmp激活剂或包含bmp激活剂的药物组合物(具有或不具有本文所述的分化培养基的其他组分)可用于与升高的分泌素水平或受抑制的glp-1水平相关的医学用途的背景下。分泌素与通过抑制胃酸分泌来中和胃ph(afroze等人,anntranslmed.2013年10月;1(3):29)和食欲抑制(cheng等人,neuropsychopharmacology.2011年1月;36(2):459-471)有关。因此,体内分泌素水平的提高可能是用于治疗胃酸过多(过量胃酸)或肥胖的有用机制。因此,提供了治疗胃酸过多或肥胖的方法,其中该方法包括向有此需要的受试者施用bmp激活剂。还提供了用于治疗胃酸过多或肥胖的方法中的bmp激活剂,其中该方法包括向有需要的受试者施用治疗有效量的bmp激活剂。还提供了bmp激活剂在制造用于治疗胃酸过多或肥胖的药物中的用途,其中该方法包括向有需要的受试者施用治疗有效量的bmp激活剂。合适的bmp激活剂的实例是本领域已知的并且早期在本申请中公开。本发明人还表明,bmp抑制剂促进eec中glp-1分泌(并抑制分泌素分泌)(参见实施例5)。因此设想bmp抑制剂或包含bmp抑制剂的药物组合物(具有或不具有本文所述的分化培养基的其他组分)可用于涉及升高的glp-1水平或抑制的分泌素水平的医学用途的背景下。例如,glp-1(胰高血糖素样肽-1)是内源性肠降血糖素并且在葡萄糖稳态中起重要作用(manandhar&ahnjmedchem.2015年2月12日;58(3):1020-1037)。它结合至并激活属于g蛋白偶联受体(gpcr)b类家族的glp-1受体(glp-1r),以发挥其调控功能。β-细胞上受体的激活导致camp和细胞内钙水平的快速升高,然后以葡萄糖依赖性方式进行胰岛素胞吐作用。尽管α-细胞中的glp-1r是β-细胞中的glp-1r的<0.2%,但glp-1通过调节钙通道活性抑制50%的胰高血糖素分泌。已经表明,glp-1疗法增强了健康和糖尿病患者的胰岛素分泌。与其他糖尿病药物不同,glp-1的促胰岛素作用是自限性的,因为一旦血浆葡萄糖水平降低到正常范围,它就会消退,从而降低低血糖的风险。此外,glp-1通过几种其他机制调控餐后血糖升高,包括促进胰岛素基因转录、刺激胰腺β-细胞增殖和新生、抑制β-细胞凋亡和阻断胰高血糖素释放。它还可以防止胃排空并引起饱腹感,导致体重下降。glp-1疗法似乎也提供心脏保护作用。然而,内源性glp-1具有非常短的半衰期,这是由于蛋白酶如二肽基肽酶iv(dpp-iv)和中性内肽酶24.11(nep24.11)的快速代谢降解导致的。这限制了其作为治疗剂的用途。存在glp激动剂(例如dpp-iv抑制剂),其被认为稳定glp-1。dpp-iv抑制剂的一个可疑缺点是glp1刚刚稳定,并且不受内源性食物摄入的控制。因此,需要用于治疗糖尿病或与其相关的疾病和病症的替代和改进疗法的替代方法。本发明人假设bmp抑制剂的体内施用可以增加eec中的glp-1分泌,并因此用作用于治疗糖尿病和相关疾病和病症的疗法。本发明人显示向小鼠施用bmp抑制剂增加了glp-1的分泌(参见实施例6)。增加glp1细胞的数量对于疗法是有利的,因为这些细胞仍然需要食物摄入以释放其glp1。因此,当受试者需要增加胰岛素水平时,glp1峰值将更高。对于具有低数量glp1细胞的患者也是特别有利的。因此,提供了用于治疗糖尿病或与其相关的疾病或病症的方法,其中该方法包括向有需要的受试者施用治疗有效量的bmp抑制剂。还提供了bmp抑制剂,其用于治疗糖尿病或与其相关的疾病或病症的方法,其中该方法包括向有需要的受试者施用治疗有效量的bmp抑制剂。还提供了bmp抑制剂在制造用于治疗糖尿病或与其相关的疾病或病症的药物中的用途,其中该方法包括向有需要的受试者施用治疗有效量的bmp抑制剂。合适的bmp抑制剂的实例是本领域已知的并且早期在本申请中公开。“受试者”可以指人或任何非人动物(例如任何小鼠、大鼠、兔、狗、猫、牛、猪、绵羊、马或灵长类动物)。在优选的实施方案中,受试者是哺乳动物,更优选是人。受试者可以是患者,其是指呈现给医疗提供者以诊断或治疗疾病的人。受试者可能患有或易患疾病或病症,但可能会或可能不会表现出疾病或病症的症状。“治疗有效量”是指当施用于患有或易患疾病、病症和/或病状的受试者时,足以治疗、诊断、预防所述疾病、病症和/或病状的症状和/或延迟所述疾病、病症和/或病状的症状发作的治疗剂的量。本领域技术人员将理解,治疗有效量通常通过包含至少一个单位剂量的给药方案施用。贯穿本公开使用的“治疗(treating)”、“治疗(treat)”、“治疗(treatment)”是指用于部分或完全缓解、改善、缓解、抑制、预防特定疾病、病症和/或病状的一种或多种更多症状或特征、延迟其发作、降低其严重性和/或降低其发病率的任何方法。治疗可以施用于没有表现出疾病表征和/或仅表现出疾病早期表征的受试者,目的是降低发生与疾病相关的病理的风险。“糖尿病”可以是i型糖尿病和ii型糖尿病。或者它可以是妊娠糖尿病。“糖尿病”还包括对胰岛素不敏感但仍可能是糖尿病前期的患者。“相关疾病和病症”包括但不限于高血糖症、肥胖、乳糜泻、甲状腺疾病、多囊卵巢综合征、尿崩症、糖尿病脂性渐进性坏死、乳腺病、肌肉病和牙齿问题。例如,本发明提供以下编号的实施方案。1.用于治疗或预防糖尿病或相关疾病或病症的方法的bmp抑制剂,其中所述方法包括将治疗有效量的bmp抑制剂施用给有需要的受试者。2.bmp抑制剂,其用于通过增加肠内分泌细胞的glp-1分泌(以提高胰岛素水平并从而降低血浆葡萄糖水平)来治疗或预防糖尿病或相关疾病或病症的方法,其中所述方法包括施用治疗有效量的bmp抑制剂给有需要的受试者。3.用于如实施方案1或实施方案2所述的用途的bmp抑制剂,其中所述bmp抑制剂能够a.破坏bmp与bmp受体的相互作用;b.结合至bmp受体并抑制下游信号传导的激活;c.抑制smad1、smad5或smad8的磷酸化;d.抑制smad1、smad5或smad8向细胞核的易位;e.抑制smad1、smad5或smad8介导的靶基因转录;或f.抑制bmp的表达、折叠或分泌。4.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述bmp抑制剂抑制smad1、smad5或smad8的磷酸化,并且其是经取代的吡唑并[1,5-a]嘧啶衍生物,例如,根据式i:其中x和y独立地选自cr15和n;z选自cr3和n;ar选自经取代或未取代的芳基和杂芳基;l1不存在或选自经取代或未取代的烷基和杂烷基;a和b在每次出现时都独立地选自cr16和n;e和f均是cr5并且两次出现的r5与e和f一起形成经取代或未取代的5-或6-元环烷基环、杂环烷基环、芳基环或杂芳基环;r3选自h和经取代或未取代的烷基、环烷基、卤素、酰氨基、氨基甲酸酯、氰基、磺酰基、亚磺酰基(sulfoxido)、氨磺酰基或磺酰氨基;r4选自h和经取代或未取代的烯基、炔基、环烷基、杂环基、芳基、杂芳基、酰基、羧基、酯、羟基、烷氧基、烷基硫基、酰氧基、氨基、酰氨基、氨基甲酸酯、酰氨基、脒基、磺酰基、亚磺酰基、氨磺酰基或磺酰氨基;r15在每次出现时独立地选自h和经取代或未取代的烷基、环烷基、杂环基、环烷基烷基、杂环基烷基、卤素、酰氨基、氨基甲酸酯、氰基、磺酰基、亚磺酰基、氨磺酰基或磺酰氨基;r16在每次出现时独立地不存在或选自h和经取代或未取代的烷基、烯基、炔基、芳烷基、环烷基、杂环基、芳基、杂芳基、杂芳烷基、环烷基烷基、杂环基烷基、卤素、酰基、羧基、酯、羟基、烷氧基、烷基硫基、酰氧基、氨基、酰氨基、氨基甲酸酯、酰氨基、脒基、氰基、磺酰基、亚磺酰基、氨磺酰基或磺酰氨基,或其药学上可接受的盐或酯。5.用于如实施方案4所述的用途的bmp抑制剂,其中:a.a和b各自为ch;b.e和f各自为cr5,并且与r5的两个实例所附接的原子一起形成6元环;c.e和f一起表示以下基团:其中r40不存在或代表选自以下的1-4个取代基:经取代或未取代的烷基、环烷基、卤素、酰氨基、氨基甲酸酯、氰基、磺酰基、亚磺酰基、氨磺酰基或磺酰氨基;d.l1具有结构其中q选自cr10r11、nr12、o、s、s(o)和so2;和r10和r11在每次出现时独立地选自h和经取代或未取代的烷基、环烷基、杂环基、环烷基烷基、杂环基烷基、氨基、酰氨基、氨基甲酸酯、酰氨基、脒基、氰基、磺酰基、亚磺酰基、氨磺酰基或磺酰氨基;r12选自h和经取代或未取代的烷基、环烷基、杂环基、杂环基烷基、氨基、酰氨基、氨基甲酸酯、酰氨基、脒基、磺酰基、氨磺酰基或磺酰氨基;和n是0-4的整数;e.r4选自:其中w不存在或是c(r21)2、o或nr21;r20不存在或选自经取代或未取代的烷基、芳烷基、环烷基、杂环基、芳基、杂芳基、杂芳烷基、环烷基烷基、杂环基烷基、酰基、磺酰基、亚磺酰基、氨磺酰基和磺酰氨基;和r21在每次出现时独立地选自h和经取代或未取代的烷基、芳烷基、环烷基、杂环基、芳基、杂芳基、杂芳烷基、环烷基烷基、杂环基烷基、酰基、磺酰基、氨磺酰基或磺酰氨基;和/或f.ar是6元芳基环或杂芳基环,任选地,其中l1相对于双环核位于ar的对位上。6.用于如实施方案4或5所述的用途的bmp抑制剂,其中所述治疗有效量是a.至少0.1mg/kg、至少0.2mg/kg、至少0.5mg/kg、至少1.0mg/kg、至少2mg/kg、至少5mg/kg、至少10mg/kg、至少20mg/kg、至少30mg/kg或约35mg/kg;b.0.1mg/kg至50mg/kg、0.1mg/kg至30mg/kg、0.1mg/kg至10mg/kg、0.1mg/kg至1mg/kg、1mg/kg至50mg/kg、1mg/kg至30mg/kg、1mg/kg至10mg/kg;和/或c.其中所述治疗有效量每日施用一次、两次或三次。7.用于如实施方案1至3中任一项所述的用途的bmp抑制剂,其中所述bmp抑制剂选自:a.dorsomorphin或ldn193189或其类似物或变体;和/或b.头蛋白、硬化蛋白、脊索发生素、ctgf、卵泡抑素、gremlin、tsg、sog或其类似物或变体。8.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中bmp抑制剂呈药学上可接受的盐的形式。9.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述受试者是哺乳动物,优选人、猫或狗。10.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述受试者是人。11.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述bmp抑制剂以如下方式施用a.口服、局部或通过注射,优选口服,和/或b.全身或局部。12.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述bmp抑制剂与一种或多种另外的糖尿病治疗剂组合施用,例如磺酰脲、双胍、二甲双胍、α-葡糖苷酶抑制剂、噻唑烷二酮、美各里替尼、二肽基肽酶-4抑制剂或其他肠降血糖素模拟物、胰淀素类似物或糖尿剂。13.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述bmp抑制剂与例如选自以下的glp-1受体激动剂组合施用:艾塞那肽、利拉鲁肽、他司鲁泰、利西拉肽。14.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述bmp抑制剂与胰岛素或其生物活性类似物组合施用。15.用于如实施方案12至14中任一项所述的用途的bmp抑制剂,其中所述组合作为单一组合物或作为两种单独的组合物施用。16.用于如实施方案12至15中任一项所述的用途的bmp抑制剂,其中所述组合同时或依序施用。17.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中糖尿病是1型糖尿病、2型糖尿病、妊娠糖尿病或胰岛素不敏感,优选2型糖尿病。18.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述受试者具有异常低水平的glp-1。19.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述方法通过增加表达隐窝特征性激素的肠内分泌细胞的数量来治疗糖尿病,其中所述隐窝特征性激素包括glp-1、神经激肽a和p物质以及胰高血糖素。20.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述方法导致循环/肠/胰腺glp-1激素水平相对于在同一患者中施用bmp抑制剂之前增加至少10%、20%、30%、40%、50%、60%、70%、80%、90%或100%。21.用于如前述实施方案中任一项所述的用途的bmp抑制剂,其中所述方法导致所述受试者的空腹血浆葡萄糖水平小于10.0mmol/l、小于9.0mmol/l、小于8.0mmol/l、小于7.0mmol/l、小于6.9mmol/l、小于6.8mmol/l、小于6.7mmol/l、小于6.5mmol/l、小于6.4mmol/l、小于6.3mmol/l、小于6.2mmol/l、小于6.1mmol/l或小于6.0mmol/l。细胞疗法本发明范围内还包括通过细胞疗法治疗人或非人动物患者的方法。这里的术语"动物"表示所有哺乳动物。患者可能处于任何发育阶段,包括胚胎和胎儿阶段。例如,患者可以是成年人,或者疗法可以用于儿科用途(例如,新生儿、儿童或青少年)。这种细胞疗法包括通过任何适当的方式将根据本发明产生的细胞或类器官施用给患者。具体地,这种治疗方法涉及受损组织的再生。如本文所用的术语"施用"是指公认的施用形式,例如静脉内或注射,以及通过移植施用,例如通过手术移植、接枝或移植源自根据本发明的细胞或类器官的组织工程化细胞群。在细胞的情况下,可以对个体进行全身施用,例如,通过经由胸导管输注到肠系膜上动脉、腹腔动脉、锁骨下静脉,经由上腔静脉输注到心脏,或输注到腹膜腔、随后经由膈下淋巴管迁移细胞,或经由输注到肠动脉血液供应(例如进入上肠系膜动脉或下肠系膜动脉)直接进入肠道位点。在一些实施方案中,每次输注施用104至1013个细胞/100kg人。优选地,可以每100kg人静脉内输注约1-5x104至1-5x107个细胞。更优选地,可以每100kg人静脉内输注约1x104至10x106个细胞。在一些实施方案中,提供了单次施用细胞或类器官。在其他实施方案中,使用多次施用。可以在初始治疗方案中提供多次施用,例如连续3-7天,然后在其他时间重复施用。如本文所解释的,还可以从表达lgr5的单个细胞中获得类器官。可通过引入如本文所定义的核酸构建体来修饰该单细胞,例如以校正遗传缺陷或突变。还可以根据需要特异性地消除表达,例如使用sirna。待表达的潜在多肽可以是代谢疾病中所缺乏的那些多肽种的任一种,包括例如代谢性肝病中的多肽缺乏,例如aat(α抗胰蛋白酶)。为了阐明生理学,我们还可以表达或灭活wnt、egf、fgf、bmp或notch通路中涉及的的基因。本领域技术人员清楚的是,基因疗法可另外用于旨在修复受损或患病组织的方法中。例如,可以使用腺病毒或逆转录病毒基因递送媒介物来向干细胞递送遗传信息,如dna和/或rna。技术人员可以替换或修复基因疗法中靶向的特定基因。例如,可以将正常基因插入基因组内的非特异性位置以替换非功能性基因。在另一个实例中,异常基因序列可以通过同源重组替换为正常基因序列。或者,选择性反向突变可使基因恢复其正常功能。另一个例子是改变特定基因的调控(基因打开或关闭的程度)。优选地,干细胞通过基因疗法方法离体治疗,随后转移至哺乳动物,优选需要治疗的人。例如,在移植入患者之前,类器官-来源的细胞可以在培养基中进行遗传修饰。因此,在一些实施方案中,上皮干细胞的类器官或群用于医学,例如,用于治疗病症、病状或疾病和/或用于再生医学。在一个优选的实施方案中,例如,如果将类器官用于再生医学,该方法可以从上皮细胞或从其中细胞或组织片段是自体的或同种异体的组织片段开始。仍可能需要某种程度的患者匹配来防止排斥问题。用于使组织排斥最小化的技术对于本领域技术人员来说是已知的。在将类器官和/或细胞移植到患者体内的实施方案中,将细胞施用于支架中可能是有利的。因此,提供了一种支架,其包含一种或多种本发明的类器官或源自所述类器官的细胞。支架提供二维或三维网络。用于这种支架的合适的合成材料包括选自多孔固体、纳米纤维和水凝胶的聚合物,例如如包括自组装肽的肽,由聚乙二醇磷酸酯、聚乙二醇富马酸酯、聚丙烯酰胺、聚羟乙基甲基丙烯酸酯、聚醋酸纤维素组成的水凝胶,和/或其共聚物(参见,例如,saha等人.(2007)curropinchembiol.11(4):381-387;saha等人(2008)biophysicaljournal95:4426-4438;little等人(2008)chem.rev108:1787-1796)。如本领域技术人员已知的,机械性质,例如如支架的弹性影响干细胞的增殖、分化和迁移。优选的支架包含可生物降解的(共)聚合物,其在受试者中移植后被天然存在的组分替代,例如以促进组织再生和/或伤口愈合。进一步优选所述支架在受试者中移植后基本上不诱导免疫原性响应。所述支架补充有天然、半合成或合成配体,其提供干细胞增殖和/或分化和/或迁移所需的信号。在优选的实施方案中,所述配体包含确定的氨基酸片段。所述合成聚合物的实例包括f127嵌段共聚物表面活性剂(basf)和(johnson和johnson)。在一些实施方案中,细胞在支架中培养。在其他实施方案中,将它们培养,然后加入到支架中。本发明的用途可以使用单一的类器官,或者它们可以使用一种以上的类器官,例如,2、3、4、5、10、15、20、30、50、100、200或更多种类器官。有利地,本发明的方法允许在短时间内产生大量的类器官和上皮干细胞,因为它们导致指数生长,从而确保足够的细胞可用于目标应用。在本文中提及“治疗方法”或“用于治疗的方法”的任何地方,例如涉及类器官或从本发明的类器官获得的细胞,这同样也指“用于治疗用途”的类器官或细胞”和“用于制造药物用途”的类器官或细胞。人造器官在一些实施方案中,提供了分化的类器官或来源于分化的类器官的细胞在人造器官中的用途。可以通过其它地方解释的方法体内移植人造器官。可替代地,人造器官可以是离体的。在一些实施方案中,离体人造器官可以例如经由血液供应与患者相连。例如,可以将包含分化的类器官的人造器官用作血液透析机的一部分。因此,可以将分化的类器官用于支持具有患病的或损伤的上皮组织的患者。肠类器官和细胞群的用途如本实施例所示,本发明的分化培养基和方法增强了从胃获得的祖细胞向肠内分泌细胞命运的分化。因此,在一些实施方案中,本发明提供了肠类器官或从肠类器官获得的细胞,用于医学例如用于治疗胃病症、病状或疾病,或用于再生医学。本发明还提供了用于治疗糖尿病(如i型或ii型)、囊性纤维化和炎性肠病(如克罗恩病)的肠类器官或从肠类器官获得的细胞,其中所述治疗任选地包括将类器官或从胃类器官获得的细胞移植到有需要的患者体内。胃类器官和细胞群的用途用于培养胃细胞的方法描述于wo2010/090513中。设想本发明的分化培养基和方法将增强从胃获得的祖细胞向肠内分泌细胞命运的分化。因此,在一些实施方案中,本发明提供了胃类器官或从胃类器官获得的细胞,用于医学例如用于治疗胃病症、病状或疾病,或用于再生医学。本发明还提供了胃类器官或从胃类器官获得的细胞,用于治疗胃炎、萎缩性胃炎、幽门狭窄、胃癌或消化性溃疡,其中所述治疗任选地包括将类器官或从胃类器官获得的细胞移植到有需要的患者体内。胰腺类器官和细胞群的用途培养胰腺细胞的方法描述于wo2010/090513中。设想本发明的分化培养基和方法将增强获自胰腺的上皮干细胞的分化。因此,在一些实施方案中,本发明提供了胰腺类器官或获自胰腺类器官的细胞在医学中的用途,例如在治疗胰腺病症、病况或疾病中的用途或者在再生医学中的用途。本发明还提供了胰腺类器官或获自胰腺类器官的细胞在治疗糖尿病(例如i型或ii型糖尿病)、胰腺炎、胰腺癌或囊性纤维化中的用途,其中治疗任选地包括将类器官或获自胰腺类器官的细胞移植进有需要的患者中。在一些实施方案中,移植的细胞是胰岛素分泌细胞。在其它实施方案中,细胞是在移植进胰岛素分泌细胞后进一步成熟的祖细胞。肺类器官和细胞群的用途用于培养肺细胞的方法描述于wo2016/083613中。设想本发明的分化培养基和方法将增强从肺获得的上皮干细胞向肠内分泌细胞命运的分化。因此,在一些实施方案中,本发明提供了肺类器官或从肺类器官获得的细胞,用于医学例如用于治疗肺病症、病状或疾病,或用于再生医学。本发明还提供了肺类器官或从肺类器官获得的细胞,用于治疗小细胞肺癌或非小细胞肺癌(例如腺癌、鳞状细胞癌或大细胞癌)、间质性肺病、肺炎(例如组织性肺炎)、肺结核、囊性纤维化、支气管炎、肺纤维化、结节病、ii型增生、慢性阻塞性肺病、肺气肿、哮喘、肺水肿、急性呼吸窘迫综合征、喘息、支气管扩张、汉坦病毒肺综合征、中东呼吸综合征(mers)、严重急性呼吸系统综合症(sars)或尘肺病。本发明还提供了肺类器官或从肺类器官获得的细胞用于治疗由病原体如腺病毒、冠状病毒(例如sars-cov或mers-cov)、人偏肺病毒、流感病毒、副流感病毒、呼吸道合胞病毒、鼻病毒、汉坦病毒、肠道病毒(例如肠道病毒d68(ev-d68))、百日咳博德特氏菌(bordetellapertussis)、肺炎衣原体(chlamydophilapneumoniae)、白喉棒状杆菌(corynebacteriumdiphtheria)、伯氏考克斯体(coxiellaburnetii)、流感嗜血杆菌(haemophilusinfluenzae)、嗜肺军团菌(legionellapneumophila)、卡他莫拉菌(moraxellacatarrhalis)、结核分枝杆菌(mycobacteriumtuberculosis)、肺炎支原体(mycoplasmapneumoniae)、金黄色葡萄球菌(staphylococcusaureus)、链球菌肺炎链球菌(streptococcuspneumoniae)或化脓性链球菌(streptococcuspyogenes)引起的病原性疾病。静止lgr5+干细胞静止lgr5+干细胞存在于体内,但之前未在体外产生。本发明人惊奇地发现egfr通路的抑制可以在体外诱导lgr5+干细胞的静止。因此,本发明提供了用于诱导lgr5+干细胞中的静止的方法,其中所述方法包括:用一种或多种egfr通路抑制剂处理细胞。在一些实施方案中,该方法是体外方法。本发明还提供了通过诱导本发明的lgr5+祖干细胞静止的方法获得的静止干细胞群,其中所述细胞表达lgr5和lef1并且不表达ki67和m期标志物磷酸化组蛋白h4。本发明还提供了包含本发明的静止干细胞群的体外培养物。在一些实施方案中,静止状态维持至少5天、至少6天、至少7天、至少8天、至少9天或至少10天。因此,在一些实施方案中,静止状态维持至少7-10天。本发明的静止干细胞群具有多种优点和应用。例如,静止干细胞群可以存储(例如在冰箱中)并且它将比非静止干细胞群更有效地再生。在一些实施方案中,本发明的静止干细胞群用于研究干细胞的细胞周期或鉴定诱导干细胞进入细胞周期的分子。缩写词β-trcp:含有β-转导蛋白重复的蛋白bme:基底膜提取物cck:胆囊收缩素chga:嗜铬粒蛋白adapi:4',6-二脒基-2-苯基吲哚edu:5-乙炔基-2’-脱氧尿苷eec:肠内分泌细胞gip:胃抑制蛋白glp-1:胰高血糖素-样蛋白1gsk-3:糖原合成酶激酶3idmi:iwp2、dapt和mek抑制剂lgr:含有富亮氨酸重复的g蛋白偶联受体lrp:低密度脂蛋白受体相关蛋白nts:神经降压素pp1:蛋白磷酸酶1pp2a:蛋白磷酸酶2app2c:蛋白磷酸酶2csct:分泌素sst:生长激素抑制素定义如本文所用的动词“包含”及其变形以其非限制性含义使用,意为包括该单词之后的项目,但不排除未具体提及的项目。此外,如果需要,动词“由……组成”可以由“基本上由......组成”代替,意为本文限定的产品可以包含除具体标识组份之外的其它组分,所述其它组分不改变本发明的独特性。此外,如本文所限定的方法可以包含除具体标识的步骤之外的其它步骤,所述其它步骤不改变本发明的独特特性。此外,不定冠词“一个/一种(a/an)”对元素的引用不排除存在多于一个/多于一种元素的可能性,除非上下文明确要求存在一个/一种且仅一个/一种元素。因此,不定冠词“一个/一种(a/an)”通常意为“至少一个/一种”。如本文所用的术语“约”或“大约”意为所呈现的值可以变化+/-10%。该值也可以作为精确值被读取,因此可以省略术语“约”。例如,术语“约100”涵盖90-110,并且也涵盖100。在此将本说明书中引用的所有专利和参考文献均通过引用整体并入。提供以下实施例仅用于说明目的,并不旨在以任何方式限制本发明的范围。术语“肠”涵盖结肠和小肠。附图描述图1.egfr抑制诱导肠类器官中的细胞周期退出。(a)实验设置。在bme中涂铺后一周,将类器官用egfr(或mek/erk抑制剂)或dmso处理。处理后1(d1)、2(d2)、4(d4)或7(d7)收集样品。从处理步骤开始重复该程序以进行重新涂铺实验。(b)对照(enr)或egfr抑制(egfri)4天后的肠类器官。egfri处理后lgr5gfpirescreer荧光增加。rfp通道用于显示本底。下面图是明场图像。(c)肠类器官的细胞周期的分析。在处死前1小时施用edu。对照(enr)类器官连续掺入edu(上图)并表达ki67(下图),而egfri处理的类器官随时间退出细胞周期。(d)c的定量。(e)对照或egfri处理的类器官的dna含量的hoechst分析。左手峰是egfri处理的类器官,并且右手峰是enr培养基中的对照类器官。右边的条显示3个独立实验的定量。条形的顶部条带代表g2/m期的细胞,条形的中间条带代表s期的细胞,并且条形的底部条带代表g0/g1期的细胞。(f)在培养基中重新引入egf后肠类器官的细胞周期的分析。(g)lgr5gfpirescreer+细胞在egfri处理4天后退出细胞周期。磷酸化-组蛋白h3(ph3)染色用于可视化m期。底部的图表显示了定量。dapi用于可视化细胞核。比例尺=50μm。图2.egfr信号传导诱导的细胞周期退出是由mapk信号传导通路介导的。(a)egfr抑制后erk磷酸化的组织学分析。perk的快速损失逐渐超过24小时,但在48小时内保持低水平(上图)。该时期与类器官的细胞周期退出一致(ki67染色,下图)。(b)使用阿法替尼对mek或erk的单一抑制以及同时抑制egfr和erbb-2产生与吉非替尼诱导的egfr抑制相似的结果。在处死前1小时将edu加入培养基中。中图显示来自lgr5gfpirescreer等位基因的内源性gfp表达。下图是明场图像。dapi用于可视化细胞核。比例尺=50μm。图3.谱系追踪表明qlgr5+细胞是干细胞。(a)实验设置。在解离后7天,将类器官重新涂铺在enr(对照)或egfri培养基中的bme内。处理后4天,用4’oh他莫昔芬(t)诱导重组16小时,并恢复egf信号传导。将类器官在他莫昔芬诱导后4或12天收集,或在enr中重新涂铺2轮。(b)重组(yfp+)细胞产生chga+eec、lyz+帕内特细胞(左图)或tuft细胞(通过其顶端肌动蛋白和乙酰化微管蛋白致密束鉴定)(右图)。追踪活性(上图)和静止(下图)lgr5+细胞。(c)b的定量。每个条中的底部带是cbc,紧接在底部带上方的带是eec,上面的下一个带是paneth,上面的下一个带是tuft并且每个条中的顶部带是rest。(d)重组的(通过x-gal染色鉴定)活性(左上图)和静止(左下图)lgr5+细胞产生显示多能性的整个类器官。重组的dclk1+细胞(右图)在任一条件下均未扩增。比例尺=50μm。图4.rna测序鉴定了qlgr5+和algr5+干细胞之间的关键分子差异。(a)使用来自对照(enr)或egfr抑制(egfri)条件的dclk1gfpirescreer(dclk1)等位基因,使用lgr5gfpdtr(lgr5dtr)、lgr5gfpirescreer(lgr5gfp)和tuft细胞对分选的lgr5+细胞的整个转录物进行分层聚类。加入对照类器官作为参考。(b)主组分分析(pca)。(c)显示活性和静止lgr5细胞的比较的火山图。x轴显示调整的p值(q值,以-log10计),并且y轴显示倍数变化(以log2计)。灰点表示错误发现率(fdr)小于0.01的基因,黑点表示没有显著变化的基因。(d)显示在lgr5+细胞中通过egfr抑制差异调控的基因的热图。颜色表示每行(基因)的z值。(e)显示标志基因的标准化表达值的箱形图。(f)显示单个活性和静止lgr5+细胞的整个转录组的pearson相关性的k均值聚类的热图。图5.高纯度eec培养的推导。(a)eec(chga)和帕内特细胞(lyz)的标志物分析。将类器官用notch抑制剂dapt(d)、wnt分泌抑制剂iwp-2(i)、吉非替尼(egfri)或这些处理的组合处理4天。dmso用作对照。(b)抑制mek信号传导(meki)连同wnt和notch信号传导通路同样地增加eec细胞数。深灰色和浅灰色v形箭头分别指向图的左半部分中高sct和高gip表达的区域。深灰色和浅灰色v形箭头分别指向图的左半部分中高cck和高sst表达的区域。胃抑制蛋白(gip)、分泌素(sct)、生长激素抑制素(sst)和胆囊收缩素(cck)阳性细胞数量急剧增加。(c)在类器官中eec相关标志基因表达的qpcr分析。ei:egfri和mi:meki。比例尺=50μm。误差棒表示标准偏差。图6.单细胞转录组分析显示诱导的eec之间的异质性。(a)热图,其显示来自meki和egfri实验的个体活类器官细胞的整个转录组的pearson相关性的k均值聚类。数字表示聚类。颜色代码为整个细胞转录组的pearson相关性。(b)描绘由细胞群表达的个体细胞和标志基因的t-sne图。(c)显示各基因的log2转化的颜色编码转录物计数的t-sne图。(d)显示聚类2、5、6和7中各基因的彩色编码转录物计数的热图。图7.生态位信号传导通路在lgr5+细胞增殖中的作用。(a)facs图,其显示内源性lgr5gfpdtr荧光(上图,x轴)和ki67efluor660免疫染色(下图,x轴)。使用pl3通道(y轴)来区分本底。门显示出就野生型对照而言的阳性细胞。(b)使用lgr5gfpirescreer和rosa-topcfp等位基因的对照(enr)和egfr抑制的(egfri)类器官的明场(上图)和荧光(下图)图像。rfp通道用于区分本底。(c)egfri处理后4天lgr5gfpirescreer+细胞的标志基因分析。浅灰色和深灰色v形箭头分别指向左手图像中的高chga和lyz表达的区域。在中间和右手图像中,浅灰色v形箭头指向高鬼笔环肽表达的区域。(d)4天egfri处理的类器官中标志基因表达的qpcr分析。(e)与各对照(dmso处理的,培养相同天数)相比,重新引入egf信号传导1(d1)、3(d3)或5(d5)天后标志蛋白阳性细胞的相对数量。(f)egf撤除(nr)或egfri处理(2xnr+egfri)的重复循环后egf引入后的类器官的细胞周期进入。(g)f的定量。比例尺=50μm。误差棒表示标准偏差。要点:enr=egf、头蛋白和r-spondin1;-r=r-spondin1撤除;-n=头蛋白撤除;egfri=egfr信号传导的抑制(使用吉非替尼,同时从培养基中撤除egf)。图8.egfri处理后类器官的细胞组成。(a)对照(enr)或egfri处理的类器官的石蜡切片上的阿尔新蓝、pas和粘蛋白2(muc2)染色。(b)对帕内特细胞(lyz)和eec(chga)标志物进行的整个类器官染色的3d重建。与对照相比,两种细胞类型在egfri处理的类器官中更集中(下图)。右边的图表提供了定量。(c)与对照(左图)相比,使用dclk1gfpirescreer等位基因的内源荧光可视化的tuft细胞数在egfri处理后增加(右图)。gfp荧光不与eec(chga)或帕内特细胞标志物(lyz)重叠。比例尺=50μm。图9.lgr5+细胞特征的基因本体(go)注释和单细胞分析。(a)在lgr5细胞中egfri处理后下调的基因的go注释分析(使用revigo)。与静止lgr5+细胞相比,细胞周期和分裂相关注释以及小分子生物合成(均与细胞周期进程相关)的活性显著更高。x轴表示p值(-log10),y轴表示go注释集的大小。(b)t-sne图显示测序的lgr5+细胞的分布。(c)t-sne图,其显示分别用于鉴定paneth和tuft细胞的lyz和dclk1mrna的log2转化的彩色编码转录物水平。图10.诱导的eec类器官的单细胞测序的初步分析。(a)t-sne图,其显示由raceid鉴定的聚类的分布。(b)t-sne图,其比较源自idegfri和idmeki实验的细胞分布。(c)t-sne图,其显示用于鉴定肠上皮细胞的apoa1mrna的log2转化的彩色编码转录物水平。肠祖细胞命运概述。肠祖细胞可以分化成许多细胞类型,如肠上皮细胞、杯状细胞、帕内特细胞或肠内分泌细胞。先前已知notch激活与wnt抑制相结合促进肠上皮细胞分化。还已知notch抑制与wnt激活相结合可促进帕内特细胞分化。此外,已知wnt和notch抑制可以促进杯状细胞分化。然而,之前对如何增强肠内分泌细胞分化缺乏了解。图11.eec培养物的组成。用idmi分化培养基处理导致分化主要偏向肠嗜铬细胞分化。每个类器官的eec绝对增加主要是肠嗜铬细胞。产生glp1、cck、nts、stt和gip的细胞存在的程度较小。图12.qpcr结果,显示不同eec分化方案中的信使rna水平。条形图显示相对于对照的各种eec标志物的倍数mrna表达。bmp通路的激活以tac1(肠嗜铬细胞的标志物)为代价选择性地增强分泌素信使rna。bmp4用于激活bmp通路,并且以10μg/ml的浓度存在。基本条件:idmi(iwp2、dapt和mek抑制剂)。mhy1485是mtor激活剂,其在5μm的浓度下进行测试。vismogenib是一种hedgehog抑制剂(特别是smoothened抑制剂),其在5μm的浓度下进行测试。图13.不同肠内分泌分化方案中分泌素和gip的染色。bmp通路的激活大大增加了s细胞的数量。图14.不同肠内分泌分化方案中血清素的染色。bmp通路的激活减少了肠嗜铬细胞的数量。图15.qpcr结果显示与标准分化培养基相比,eec分化方案中不同eec标志物的信使水平的log2-变化。eec分化培养基大大增加了人类器官中eec的数量,但不会增加帕内特细胞中eec的数量。图16.在不同肠内分泌分化方案中的chga表达。图17.notch、wnt和mek的三重抑制产生富含eec的培养物。观察到每个类器官30-80%chga+细胞。图18.eec培养物中的激素表达。eec的所有不同亚型都存在于idmi培养物中并表达单一激素。有些细胞是chga-但对激素是阳性的。图19.eec在培养中分泌激素。收集2天培养基用于-毛喉素结果。然后用毛喉素诱导培养物1小时,并收集培养基用于+毛喉素结果。图20.tac1、glp1和分泌素免疫反应细胞在肠隐窝和绒毛中的差异定位。a-c)pyy-glp1双阳性细胞位于隐窝中,而绒毛中的l细胞则失去了glp1的表达。d-e)表达血清素的隐窝中的肠嗜铬细胞共表达tac1。绒毛中的tac1表达缺失,其中肠嗜铬细胞开始共表达分泌素。图21.bmp信号传导是绒毛激素特征的驱动因子。a)肠内分泌细胞分化(wnt、notch和mapk的抑制)培养基,其中bog信号传导被头蛋白的存在所抑制,产生一种令人联想到隐窝的激素特征:分泌素不存在,并且血清素+eec总是共表达tac1。排除头蛋白和将bmp4引入该分化混合物(定义为eecbmp高),大大降低了glp1数量以及tac1。单一血清素+肠内分泌细胞,以及分泌素+细胞,出现在该培养基中。b)a)的定量c)bmp低或高eec分化培养基中小肠类器官的明场图像。gcg-venus报告子用于跟踪glp1阳性细胞。尽管不同的分化方案产生形态学上难以区分的类器官,但是在bmp激活的背景中glp1表达大大降低。图22.体内bmp信号传导的抑制导致glp1+区室的扩增,并抑制分泌素表达。a)通过用ldn193189口服管饲小鼠进行的60小时治疗导致glp1数量增加。glp1由绒毛中的l-细胞广泛表达,其不总是共表达pyy。b)与对照处理相比,bmpr抑制的小鼠的绒毛中的分泌素大大减少。实施例材料和方法类组织培养基础培养基(先进的杜尔贝科改良伊格尔培养基/f12,补充有青霉素/链霉素、10mmhepes、glutamax、b27[lifetechnologies,carlsbad,ca]和1mmn-乙酰半胱氨酸[sigma])补充有50ng/ml鼠重组表皮生长因子(egf;peprotech,hamburg,germany)、r-spondin1(条件培养基,5%终体积)和头蛋白(条件培养基,5%终体积),称为‘’enr”培养基。使用用ha小鼠rspo1-fc(来自斯坦福大学(stanforduniversity)的calvinkuo的馈赠)稳定转染的hek293t细胞或用小鼠头蛋白-fc表达载体瞬时转染后产生条件培养基。先进的杜尔贝科改良伊格尔培养基/f12(补充有青霉素/链霉素和glutamax)适应1周。将细胞涂铺于bme(trevigen)中。为了抑制egf信号传导,用吉非替尼(5μm;santacruzbiotechnology)处理细胞,并从培养基中取出egf。用iwp-2(1.5μm;sigmaaldrich)抑制wnt分泌并用dapt(1mm,sigmaaldrich)抑制notch。所有处理均在传代类器官后5天进行。对于egfr再激活实验,将类器官重新涂铺于新鲜bme和enr培养基中以确保洗去egfr抑制剂。为了诱导cre-ert活性,用4-羟基它莫西芬(1um)处理类器官过夜。用相似浓度的化合物溶剂二甲基亚砜(dmso)处理所有对照类器官。在处理期间,使用evos显微镜(electronmicroscopysciences)对细胞成像。为了诱导肠内分泌分化,将细胞培养在enr或enr加丙戊酸和chir99021中(yin等人(2014)naturemethods11:106-112)。培养5天后,除去培养基,并用pbs洗涤类器官。用于eec分化的混合物包括:iwp2(1.5μm;sigmaaldrich)、dapt(1mm,sigmaaldrich)和mek抑制剂pd0325901(5μm;sigmaaldrich)。免疫染色通过将基质胶轻轻溶解在冰冷的pbs中收集整个类器官,随后在4℃下在4%多聚甲醛中固定过夜。接下来,将类器官透化并在含有0,5%tritonx-100和2%正常驴血清(jacksonimunoresearch)的pbs中在室温下封闭30分钟。将类器官在室温下在含有一抗的封闭缓冲液中孵育2小时。使用的一抗是兔抗溶菌酶(1:500;dako)、山羊抗嗜铬粒蛋白a(1:500;santacruz)、小鼠抗ki67(1:250;bdpharmingen)、兔抗磷酸化-组蛋白3(ph3ser10,1:1000;millipore)、小鼠抗细胞角蛋白20(1:1000;dako)、山羊抗胆囊收缩素(sc-21617,1:100;santacruz)、兔抗神经降压素(sc-20806,1:100;santacruz)、山羊抗分泌素(sc-26630,1:100;santacruz)、山羊抗生长激素抑制素(sc-7819,1:100;santacruz)、兔抗胃抑制蛋白(ab22624-50,1:500,abcam)、兔抗胰高血糖素如肽1(ab22625,1;500,abcam)和小鼠抗乙酰化微管蛋白(1:100;santacruz)。将类器官与相应的二抗alexa488、568和647缀合的抗兔、抗山羊和抗小鼠(1:1000;molecularprobes)一起在含有dapi(1;1000,invitrogen)封闭缓冲液中孵育,或与鬼笔环肽德克萨斯红(phalloidintexasred,1:1000;lifetechnologies)一起孵育。在与edu(10um)预孵育1小时后,使用click-it测定试剂盒(thermofisher)可视化edu掺入。如前所述进行lacz染色(barker等人.(2007)nature449(7165):1003-7)。将切片包埋在vectashield(vectorlabs)中,并使用sp5和sp8共聚焦显微镜(leica)成像。使用imagej软件进行图像分析。facs分选对于lgr5和ki67表达的facs分析,在37℃下胰蛋白酶处理15分钟后(trypleexpress;lifetechnologies,carlsbad,ca),lgr5gfpdtr类器官首先通过机械破碎解离成单细胞。使用4%多聚甲醛将单细胞固定在冰上30分钟,并在pbs中洗涤3次。将细胞在含有0,5%tritonx-100的pbs中透化30分钟,并用efluor-660缀合的大鼠抗ki67(1:1000;ebioscience)抗体在冰上染色30分钟。对于细胞周期分析,将细胞在1ug/mlhoechst33342(thermofisher)中染色。随后,在bdfacscalibur(bdbiosciences)上分析染色的细胞。为了进行表达分析,将类器官解离并立即在bdfacsaria(bdbiosciences)上分选。将细胞在96孔板中的trizol内分选为单细胞,或在eppendorf管中的trizol内批量分选。rna分离对于rna测序,将细胞分选到trizol(lifetechnologies)中,并根据具有以下改变的制造商的说明分离总rna。将rna在-20℃下用2ug糖原(lifetechnologies)沉淀过夜。没有额外的rna分离步骤用于分选到384孔中的细胞。对于定量pcr分析,使用rnaeasy试剂盒(qiagen)按照制造商的方案中指导从类器官中分离rna。定量pcr使用如所述的sybr-green和bio-rad系统进行pcr分析(munoz等人(2012)theembojournal31:3079-3091)。pcr反应一式三份进行,每个引物具有标准曲线。使用cfx管理器软件(bio-rad)计算表达的变化。使用ncbi引物设计工具设计引物。单细胞和批量测序如前所述,使用修改形式的cel-seq方案制备rna样品(grun等人(2015)nature525:251-255;hashimshony等人(2012)cellreports2:666-673)。将ercc加标物加入trizol溶液中。将rna沉淀溶解在引物混合物中并在70℃下孵育2分钟。将分选到384孔中的细胞在65℃下直接裂解5分钟。使用75-bp配对末端测序在illuminanextseq500上对cdna文库进行测序。rna测序数据的分析.如前所述(grun等人(2015)nature525:251-255)除了以下例外定量配对末端读段。与多个位置没有对齐或对齐的读段被丢弃。对于批量测序的分析,忽略了umi;相反,每个转录物的读段计数由唯一作图至该转录物的读段确定。这个计数除以作图至所有转录物的读段总数,再乘以100万,以生成每百万读段(rpm)计数。使用rpm优先于rpkm,因为cel-seq仅允许3'末端测序。使用r平台中的deseq包评估差异基因表达(anders和huber(2010)genomebiology11:r106)。使用的截止值是调整的p值<0,1和fdr<0,1并且与比较的群体的差异至少为2倍。为了防止没有读段的样品禁用比率分析,在比率计算和log2转换之前将所有0读段转换为0,1读段。使用revigo(supek等人(2011)plosone6:e21800)和gorilla(eden等人(2009)bmcbioinformatics10:48)软件进行基因本体分析。如前所述分析单细胞测序数据(grun等人(2015)nature525:251-255)。实施例1为了理解lgr5+干细胞是如何保持循环的,我们操纵关键信号传导通路在腺窝生态位中活跃。用抗ki67抗体(在所有细胞周期阶段中循环细胞的标志物)对lgr5gfpdtr类器官进行的流式细胞术分析证实大多数lgr5+细胞循环(图7)。我们使用两种独立的方法抑制wnt信号传导;i)iwp-2处理抑制帕内特细胞分泌wnt3,和ii)从培养基中撤除r-spondin1导致卷曲受体从细胞表面丧失。r-spondin1撤除立即导致lgr5gfp表达缺失(图7a)。iwp处理引起较温和的wnt抑制,其依赖于通过增殖稀释配体。lgr5gfp表达逐渐下调,而干细胞分化。然而,剩余的lgr5gfp细胞维持ki67表达(63.5±2.8%对比对照中的94.4±2.1%)。撤除bmp抑制剂头蛋白或加入notch抑制剂dapt均诱导gfp+细胞数量迅速减少,但不影响剩余lgr5细胞的增殖(头蛋白撤除中为82.3±1.4%,dapt中为45.1±10%)(图7)。接下来,我们使用吉非替尼抑制egfr信号传导,同时从培养基中撤除egf(egfri处理,图7)。虽然lgr5gfp表达持续存在,但lgr5细胞最终丧失ki67表达(13.1±1.0%),表明从细胞周期退出。我们进一步分析了与egfr抑制相关的早期事件(图1a)。尽管类器官的‘绒毛’区室广泛凋亡,但类似于含有lgr5+(lgr5gfpirescreer)细胞的隐窝结构的芽持续长达一周(图1b)。我们使用lgr5gfpdtr等位基因(tian等人(2011)nature478:255-259)证实了这些结果,证实了cbc在没有egf信号传导的情况下存活(图7)。在egfri处理4天后,lgr5+细胞组成44.4±0.8%(对照中13.6±6.5%)的类器官(图7)。tcfcfp(rosatcf-cfp)wnt报告子等位基因(serup等人(2012)diseasemodels&mechanisms5:956-966)证实wnt信号在非增殖性lgr5细胞中保持较高(图7)。ki67蛋白在最初的24小时内持续存在,但从48小时开始丢失(图1c和图1d)。使用edu的短脉冲作为复制其dna的细胞的量度,我们发现egfri早在24小时就导致dna复制快速停止,持续至少一周(图1c和图1d)。与退出s期并最终退出细胞周期一致,使用hoechstdna染色标记egfri处理的类器官的dna含量证实所有细胞处于g0/g1期(图1e)。在egfri处理后4天,egf信号传导的重建在24小时内诱导快速细胞周期进入(ki67+)并在48小时内进展至s期(edu+)(图1f和图7)。即使在第二轮egfri处理后,静止的lgr5细胞也进入细胞周期(图7)。然后,我们进一步细化了非分裂的egfri诱导的lgr5+细胞的分析。它们缺乏细胞周期标志物ki67和m期标志物ph3并且不掺入edu,排除在egfri处理期间持续存在的罕见分裂细胞(图1g)。缺乏溶菌酶(lyz)和嗜铬粒蛋白a(chga)意味着lgr5+细胞分别地不是分化的帕内特细胞或肠内分泌细胞(eec)(图7)。tuft细胞(肠m细胞)是罕见的机械感觉细胞,也参与对寄生虫侵袭的响应(howitt等人(2016)science351:1329-1333)。它们可以通过其典型的顶端肌动蛋白束来区分,其标记为乙酰化微管蛋白和鬼笔环肽(hofer和drenckhahn(1996)histochemistryandcellbiology105:405-412)。绝大多数lgr5+细胞不显示tuft细胞形态,并且-类似地-tuft细胞主要是lgr5-(图7)。定量pcr分析证实lgr5细胞不分化成肠上皮细胞、paneth、eec、杯状或tuft细胞(图7)。muc2(粘蛋白2)以及pas和阿尔新蓝(alcianblue)染色以可视化粘液结构显示egfri处理后杯状细胞的数量显著减少(图s2)。大多数分裂ta祖细胞产生成熟的肠上皮细胞。虽然我们发现egfri处理后4天每芽的lyz+帕内特细胞数和chga+肠内分泌细胞数增加,但任一细胞类型的总数(每个类器官)没有显著改变(图8)。在egfri处理后,tuft细胞的总数增加(图s2)。我们使用dclk1gfpirescreer等位基因证实了这些结果(nakanishi等人(2013)),证实egfri处理使dclk1+tuft细胞的绝对数量增加3.2倍(enr中为11.3±6.6,egfri中为35.8±8.8)(图9)。简言之,虽然egfr抑制影响类器官中的细胞类型组成,但它驱使lgr5+细胞进入静止而不诱导分化成肠谱系之一。mapk信号传导是egfr信号通路的主要下游靶标并调控细胞周期进程。mapk激酶(mek)磷酸化mapk(erk)以诱导其核定位和激活。egfri处理早在1小时就减少了erk磷酸化(图2a)。然而,尽管持续静止,我们观察到48小时内磷酸-erk(perk)水平逐渐恢复(图2a)。我们使用mek(pd0325901;meki)或erk(sch772984;erki)的小型抑制剂探询mek/erk信号传导是否对肠干细胞的细胞周期进展至关重要。两种抑制剂都诱导lgr5+细胞的静止,这意味着egfr下游的erk通路调控lgr5+细胞的增殖(图2b)。使用抑制egfr和erbb2的阿法替尼产生了类似的结果(图2b)。我们得出结论,通过mapk信号传导的egfr抑制足以在肠类器官干细胞中诱导可逆的静止状态。为了测试静止的lgr5细胞是否保留干细胞潜能,我们使用了lgr5gfpirescreerrosa26yfp/lacz和lgr5gfpirescreerrosa26tdtomato类器官以跟踪lgr5+细胞的命运(图3a)。使用4-oh他莫昔芬(tmx)的creer诱导导致快速重组,其可通过yfp(或tdtomato)荧光或lacz表达可视化(图3b和图3c)。标记的、分裂的lgr5细胞在egfri处理后产生静止的lgr5细胞,表明后者确实来源于活性lgr5细胞(图3b)。此外,标记的帕内特细胞和eec的比例在egfri处理后增加(图3b和图3c)。在非egfri处理的对照中,重组细胞在传代后产生完整的类器官,表明干细胞的有效标记(图3d)。在传代和egf信号传导的再激活后,重组静息lgr5细胞的后代持续数代传代并产生类器官(图3d)。这些结果表明,在egfri处理后诱导的静止lgr5细胞表现为真正的干细胞。tuft细胞是非分裂的,并且可能通过在损伤和肿瘤生长后促进组织再生而显示干细胞样特性(nakanishi等人(2013)naturegenetics45:98-103)。我们使用源自dclk1gfpirescreerrosayfp/lacz小鼠模型的类器官跟踪tuft细胞的命运。在正常和egfri处理的类器官中,标记的细胞随时间保持为单细胞(图3c)。当在egf存在下传代时,标记的细胞丢失并且对类器官生成没有贡献(图3c),反驳了在器官样环境中dclk1+tuft细胞具有干细胞潜能。为了更好地理解静止lgr5细胞的分子特征,我们对facs-分离的活性(dmso对照)和静止(egfri处理,d4)lgr5+干细胞进行了rna测序。在我们的研究中,我们包括lgr5gfpirescreer(n=2)和lgr5gfpdtr(n=2)等位基因。我们还包括分选的dclk1gfptuft细胞和大量的类器官用于比较。分层聚类和主组分分析(pca)显示lgr5+细胞彼此更相似,而不是整个类器官或tuft细胞(图4a和图4b)。与其细胞周期的差异一致,活性和静止的lgr5+干细胞分别聚类(图4a和图4b)。活性和静止lgr5细胞之间的差异基因表达分析揭示了533个差异调控的基因,其中290个富含静息lgr5细胞(fdr<0.01,图5c)。erk通路的转录效应子(etv4[7.7x,p-adj<0.001]和etv5[7.7x,p-adj<0.001])在静止的lgr5+干细胞中被下调,证实了有效的erk抑制(图4c和4d)。类似地,数种细胞周期相关基因,如ccnb1(2.1x,p-adj<0.005)和ccnb2(1.9x,p-adj<0.05)减少,与g0停滞一致(图4c和s9)。对egfri处理后下调的基因的go分析证实了细胞周期相关基因的明显缺失(图s9)。静止lgr5细胞中定义的转录物之一是lef1(未在活跃的lgr5中检测到,p-adj<0,001),一种wnt信号传导通路的组分。与我们的报告子表达一致,我们观察到一些众所周知的wnt靶基因的显著增加,包括rnf43(2.3x,p-adj<0.005)和lgr5(2x,padj<0.05)(图4d)。独立实验中的定量pcr分析证实了这些结果(图9)。我们还注意到静息lgr5细胞中ap-1转录因子家族(junb、fos、fosb)成员的强烈增加(图4d)。与组织学分析一致,帕内特细胞、肠上皮细胞和杯状细胞特异性基因表达保持不变。与活性lgr5+干细胞相比,由eec及其前体表达的chga在静止时高7.3倍,尽管低表达水平的高变异阻碍了进一步的结论(p-val=0.019和padj=0.28)。类似地,虽然dclk1(6x,p-adj<0.05)和一些其他tuft细胞标志物在egfri处理后增加,但它们在静止lgr5细胞中的水平显著低于在tuft细胞中的水平(图4d)。基因表达的全局变化可能由所有静止的lgr5干细胞共享,或者是特定亚群变化的结果。我们最近表明,单个活性lgr5细胞是相当同源的(grun等人(2015)nature525:251-255)。我们对来自对照或egfri处理的类器官的总共192个facs纯化的单活性或静止lgr5细胞进行单细胞测序分析。使用raceid(grun等人(2015)nature525:251-255),我们鉴定了含有来自对照和含有活性和静止的lgr5+细胞(图9)的egfri处理的细胞(图4f)的细胞的单个显著群体(簇1)。次要簇2(四个细胞)和4(一个细胞)表达paneth(lyz1)和tuft(dclk1)细胞相关基因,而簇3(一个细胞)类似于簇1(图9)。因此,静止lgr5群体是同源的,并且尽管基因表达有一些显著变化,但非常类似于静止lgr5细胞。实施例2与增加的chga和lgr5表达相结合的静止(如实施例1中所述)使人想起winton及其同事描述的保留标记的分泌前体(buczacki等人(2013)nature495:65-69)。这些细胞有效地分化成eec,表明lgr5细胞的细胞周期退出可促进eec的产生。eec表达的激素调控多种生理响应,如胃排空、胰酶释放、情绪和葡萄糖耐受。它们还用于定义亚型(参见介绍)。g蛋白偶联的味觉受体已被鉴定为激素分泌的调控剂(janssen和depoortere(2013)tem24:92-100)。eec可以直接接触管腔并用微绒毛感知内容物。其他eec,即所谓的封闭型细胞,没有管腔内衬并需要刺激其他来源(janssen和depoortere(2013)tem24:92-100)。它们的基础过程的长度也变化,并且可以与肠神经元形成突触接触以连接到神经系统。为了建立eec分化的方案,我们使用静止的lgr5细胞作为起始点并调节参与分泌分化的notch和wnt信号传导通路。通过dapt处理(d)抑制notch信号传导增强了分泌分化,导致lyz+帕内特细胞数量的大量增加(图5a)。使用iwp-2(i)与dapt组合抑制wnt分泌消除了帕内特细胞分化并诱导了eec和杯状细胞(图5a)。egfri处理抑制了杯状细胞分化,但帕内特细胞和eec免于此(图5a)。egfr/wnt/notch通路(idegfri)的联合抑制导致eec的大量增加,同时抑制肠上皮细胞、杯状细胞和帕内特细胞分化(图5a)。类似地,抑制mek与wnt和notch信号通路(idmeki)一起增加chga+细胞数量。不同的eec亚型在正常肠类器官培养物中很少见(图5b)。idmeki处理导致这些eec细胞类型数量稳健增加。我们使用qpcr来证实这些结果。泛-eec(pan-eec)标志物chga的表达在idegfri中高25倍,并且在idmeki处理的类器官中高超过100倍(图5c)。巧合的是,在idegfri处理后,sst(55x)、gip(14x)、sct(5x)、胆囊收缩素(15x)和胰高血糖素(gcg/胰高血糖素原,4x)mrna的表达上调,与idmeki趋势类似(图5c)。nts是分析的唯一激素,以对照水平表达。因此,我们的方案成功地产生了大量的eec和表达一组调控哺乳动物生理学的激素的各种亚型(egerod等人(2012)endocrinology153:5782-5795)。为了阐明eec诱导的类器官的细胞组成和各个ecc的激素表达的异质性程度,我们进行了单细胞rna测序分析。我们从idegfri和idmeki处理的类器官中分选了活的单细胞(没有额外的标志物)。在通过我们过滤的289个细胞中,我们鉴定了94个细胞的簇为富含aldob(4.9x,p-adj<0.001)、apoa1(12.6x,p-adj<0.001)和alpi(5.6x,p-adj<0.001,图10)的肠上皮细胞。这些被排除在进一步分析之外,以更好地表征剩余群。源自idegfri或idmeki处理的类器官的细胞在t-sne空间中类似地分布并一起分析(图s4)。使用raceid,我们鉴定了12个不同的细胞簇(图6a)。细胞转录组的pearson相关性的k均值聚类揭示了簇之间的明显分离以及簇内可能的异质性(例如,7和8,图6a)。差异基因表达分析揭示了每个簇的特征基因,我们用它们对细胞类型进行分类(表s1)。最突出的簇是3(53个细胞)和4(35个细胞),其表达泛-eec标志物chga和chgb(图6c)。chga和reg4表达形成梯度,在簇4中均更高。这些chgb高簇中的激素产生最好由tac1和tph1表达定义,这两者都是肠嗜铬细胞的标志物。tac1编码激素物质p,而tph1编码血清素合成中的限速酶(egerod等人(2012)endocrinology153:5782-5795;grun等人(2015)nature525:251-255)。两者都可以充当激发连接的肠神经元的神经递质(latorre等人(2016)neurogastroenterologyandmotility:theofficialjournaloftheeuropeangastrointestinalmotilitysociety28(5):620-630)。其他簇具有相对低水平的chga和chgb转录物,但包括表达激素的细胞(图6c)。通过由k细胞表达的gip表达(74x)标记簇2(21个细胞)。fabp5也在该簇中高度富集(12,6x),与其在gip分泌中的作用一致(shibue等人(2015)americanjournalofphysiology,endocrinologyandmetabolism308:e583-591)。簇5(9个细胞)成员表达非常高水平的sst(182x),从而将它们鉴定为d-细胞(图6c)。ghrelin(ghrl)表达分布于多于一个簇,但在簇6中最高(19x,3个细胞)。我们还注意到islet1(isl-1,9,7x)在这些细胞中与ghr1共表达。islet1在细胞命运决定中起重要作用,并且其损失导致葡萄糖稳态受损(terry等人(2014)americanjournalofphysiology,gastrointestinalandliverphysiology307:g979-991)。簇7中的细胞(18个细胞)均高度表达cck(55.7x),并且该簇中的亚群也富含其他激素,如gcg(28,2x)、ghrl(5,3x)和pyy(11,4x)。一致地,据报道i细胞共表达cck与不同水平的其他激素(egerod等人(2012)endocrinology153:5782-5795)。eec分化的早期诱导物之一是神经原素-3(neurog3),其后是neurod1。neurog3(5,2x)表达在簇9(6个细胞)和簇3的与簇9最相似的一些细胞中最高。几乎所有eec簇都表达neurod1。鉴于这些转录因子的时间表达,我们建议簇9代表eec祖细胞,然后通过neurod1产生一组eec。簇1(18个细胞)富含杯细胞和帕内特细胞相关基因,如agr2(33x)、muc2(26x)、ttf3(23x)和defa24(28x)。即使在过滤后,仍然可见一些表达aldob和mt1/2的剩余肠上皮细胞样细胞(簇8,7个细胞)。这些簇可以是在eec诱导之前产生的,因为我们没有看到egfr或mek抑制后其数量增加。dclk1和trpm5表达将簇10(15个细胞)鉴定为tuft细胞(图6b和6c)。总之,分析的145/289个细胞(所有细胞的50%)是eec或它们的祖细胞,证实了我们的诱导方案的高效率。由于多种激素可以在同一细胞中共表达,我们进一步研究了单细胞水平的激素表达的异质性(图6d)。我们专注于表达多种激素的簇2、5、6和7。基于gip、sst、cck和ghrelin表达的4个主要组是可见的。cck+细胞进一步分裂成gcg+ghrl+、gcg+ghrl-、gcg-ghrl+和gcg-ghrl-簇,其中一些还共表达低水平的sst和/或gip。sst+细胞的转录组更均一,共表达低水平的gip和cck,而一个细胞仅共表达ghr1。我们先前已经报道了cck+和tac1+细胞之间的部分重叠。一致地,簇3和簇4中的一些tac1+细胞表达低水平的cck(图6b和图6c)。总之,我们的单细胞分析表明我们的方案基于标记基因表达将eec富集至培养物的约50%。此外,我们生成了一组eec亚型,包括一些具有复杂激素表达谱的稀有细胞。实施例3使用含有wnt、notch和mapk信号传导的抑制剂的鼠eec分化培养基来产生类器官。这些类器官含有所有eec亚型的混合物。产生血清素的肠嗜铬细胞是这些培养物中最丰富的细胞类型。研究了产生不同eec亚型的机制。这是为了扩大培养物的用途并了解这些细胞的发展。在可能控制eec亚型决定的信号传导通路的第一次筛选中,发现bmp、hedgehog和mtor通路的调节影响eec亚型的相对比率。分泌素-产生s细胞在wnt、notch和mapk抑制期间似乎特别罕见。s细胞通常位于近端十二指肠,与本研究中的类器官从中分离出来的相同的位置。通过从eec分化培养基中撤除头蛋白并向培养基中加入bmp4(10μg/ml)来激活bmp通路,改变了eec亚型的相对丰度。在bmp通路激活后,观察到s细胞数量(在信使上,相对于对照是400倍)以及每个细胞的分泌素水平(基于ihc)急剧增加。s细胞的这种增加似乎以肠嗜铬细胞的数量为代价,这表明潜在的重叠发育通路。参见实施例5中bmp信号传导的作用的进一步讨论。实施例4首先在扩增培养基中培养人小肠(si)类器官(如sato等人(2011)gastroenterology141(5):1762-72中所述)以增加干细胞数,然后在分化培养基(缺乏wnt条件培养基、tgfβ-抑制剂、p38抑制剂和烟酰胺的扩增培养基)中重新涂铺以指导分化。重新涂铺涉及在冷培养基中破坏基质胶(不解离类器官),用基础培养基洗涤类器官(也可以使用pbs),然后在新鲜的基质胶中涂铺(不解离类器官)。然后将类器官在分化培养基中培养1天。第二天,eec特异性肠内分泌分化方案。这涉及加入eec分化培养基5天,其与标准分化培养基(缺乏wnt条件培养基、tgfβ-抑制剂、p38抑制剂和烟酰胺的扩增培养基)相同,加入1.5μm的iwp2、10mm的dapt、100nm的mekipd0325901。在该实验中,将这些人细胞分化为eec命运所需的mapk抑制剂水平要远低于实施例3中将鼠细胞分化为eec命运所需的水平(在小鼠系统中,100-500nm对比1-5μm),可能是由于缺乏在人体系统中产生egf的帕内特细胞。讨论在这里,我们鉴定了egf信号传导是在类器官中lug5干细胞增殖的必不可少的驱动因素。在wnt信号传导未受影响但egf信号传导被阻断的条件下,活跃分裂的lgr5干细胞转化为静止的lgr5细胞,其保留各种wnt靶基因的表达。这种细胞状态可以维持长达一周。然而,egf信号传导的简单恢复将静止细胞转变回其正常的活性干细胞状态。差异表达分析揭示了众所周知的增殖诱导转录因子如ets样因子etv4和-5的丢失,表明它们在干细胞分裂中起关键作用。因此,egfr信号传导的化学抑制阻止lgr5+干细胞而不影响其干细胞潜能。我们相信高水平的wnt信号传导在诱导静止期间维持这种干细胞潜能。在类器官以及隐窝中,lgr5+细胞始终是分泌wnt3的帕内特细胞的紧邻邻居(sato等人(2011)nature469:415-418)。在这种设置中,wnt3不会在距离上扩散,而是直接负载到lgr5干细胞上(farin等人(2016)nature530:340-343)。静止的lgr5干细胞与egfri处理的类器官中的帕内特细胞保持并置,并因此暴露于高的局部wnt信号。实际上,3个独立的wnt靶基因等位基因以及基因表达分析证实了egfr抑制后wnt信号传导增加。总之,我们的结果表明维持干细胞命运需要wnt而不是egf,而干细胞增殖依赖于wnt和egf的组合。先前的研究已经鉴定出静止细胞接近具有干细胞潜能的‘+4’位置处的分化区(sangiorgi和capecchi(2008)naturegenetics40:915-920;takedaetal.(2011)science334:1420-1424;yan等人(2012)proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica109:466-471)。我们已经报道了在该位置专门产生分泌细胞的dll1+前体的存在(vanes等人(2012)naturecellbiology14:1099-1104)。使用组蛋白标记保留测定法,dougwinton小组鉴定了具有分泌分化潜能的类似群体。这些标记保留细胞与cbc共享特征(包括lgr5表达),但表达一些分泌谱系基因如chga的良好水平(buczacki等人(2013)nature495:65-69)。总之,这些分泌前体代表瞬时状态,但在需要时可以去分化为干细胞,因此可以被认为是兼性干细胞(buczacki等人(2013)nature495:6569;vanes等人(2012)naturecellbiology14:1099-1104)。对于隐窝中丰富的肠上皮细胞前体存在类似的情况(tetteh等人(2016)cellstemcell18:203-213)。令我们惊讶的是,我们注意到静止lgr5细胞略微偏向于eec标志物如chga的表达,这使得它们让人想起dougwinton鉴定的lgr5+标记-保留细胞。eec占肠上皮细胞的不到1%,但在细胞数量方面共同形成人体内最大内分泌器官(latorre等人(2016)neurogastroenterologyandmotility:theofficialjournaloftheeuropeangastrointestinalmotilitysociety28(5):620-630)。已经提出eec用作如营养物和微生物的管腔内容物的传感器,并通过激素的分泌调控生理响应如葡萄糖敏感性、胃排空、情绪和食欲(janssen和depoortere(2013)tem24:92-100)。eec的功能研究受到缺乏大量产生这些细胞的稳健体外系统的阻碍。我们先前已经报道了通过notch抑制将干细胞定向分化成分泌细胞(vanes等人(2005)nature435:959-963)。帕内特细胞形成需要活跃的wnt信号传导结合notch抑制(vanes等人(2012)naturecellbiology14:1099-1104;vanes等人(2005)nature435:959-963),并且联合抑制wnt和notch信号传导主要产生杯状细胞(vanes等人(2005)nature435:959-963;yin等人(2014)naturemethods11:106-112)。通过将egfr通路的抑制与wnt和notch阻断相结合,在原代细胞培养物中有效地形成多种类型的eec。已显示egfr信号传导对于杯状细胞的产生是重要的(heuberger等人(2014)proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica111:3472-3477)。我们认为,分别通过抑制notch、wnt和egfr信号传导同时抑制肠上皮细胞、paneth和杯状细胞的命运是eec产生的关键。基于特定肽的产生,可以区分多达20种eec亚型。这些细胞在近端至远端胃肠道中以不同频率存在。已经发现某些eec在转录物和蛋白质水平上表达数种功能相关的激素(egerod等人(2012)endocrinology153:5782-5795;grun等人(2015)nature525:251-255),尽管其中一些可能成熟的中间阶段。我们通过单细胞转录组谱分析证明了同时产生eec的几种主要亚型。确认我们以前的工作(grun等人(2015)nature525:251-255),我们检测到chga-高和低-eec群。前者富含表达tac1/tph1的eec,其中一些细胞低表达cck。后者在激素表达方面非常多样化。具有总体相似的转录组谱的细胞聚类在一起,即使它们可以基于激素表达细分为数个类别。激素表达水平的广泛分布表明eec亚型之间的分离并不明确。很有可能推测它们的表达至少部分是随机的并且甚至可能是暂时动态的。实施例5肠内分泌细胞改变隐窝向绒毛迁移过程中的激素表达多种肠内分泌细胞激素在肠隐窝和绒毛中差异富集。l细胞在隐窝中共表达glp1和pyy,但在绒毛中大多缺乏glp1表达(图20a-图20c)。肠嗜铬细胞沿整个隐窝-绒毛轴产生血清素,但选择性地共表达隐窝中的tac1和绒毛中的分泌素(图20d-图20e)。这表明eec可以在从隐窝迁移到绒毛期间转换激素表达,并且隐窝和绒毛eec之间存在潜在的谱系关系。然而,对该观察结果的另一种解释是,对绒毛中的分泌素或pyy具有免疫反应性的细胞来源于隐窝中的无关祖细胞,对血清素和tac1或glp1呈阴性。为了证明这种转分化事件可以在eec的生命周期中发生,我们分析了来自tac1-cre/rosa26tdtomato小鼠的肠。这种报告子忠实地标记了共表达tac1和血清素的隐窝中的细胞。此外,追踪绒毛中的血清素+细胞和大部分分泌素+细胞,而对tac1呈阴性。其他激素,包括cck,不是来源于tac1/血清素+祖细胞。这些数据表明,个体eec可以在隐窝和绒毛中表达不同的激素组。大多数产生分泌素的细胞是肠嗜铬细胞谱系的一部分。eec转换激素表达的能力意味着eec的无关分化途径可能比先前预期的少,但是由有限数量的细胞产生的激素类型的相当改变产生了过多的eec状态。bmp信号传导在肠内分泌细胞中诱导绒毛激素特征我们接下来探询从隐窝到绒毛迁移发生的激素转换是否是eec中默认成熟过程的一部分,或者这仅仅反映了eec暴露的不同生态位信号。已知从隐窝离开到绒毛的多种形态发生素,包括上升水平的bmp和hedgehog,以及下降水平的wnt信号。早期实施例中描述的鼠肠类器官系统中肠内分泌细胞的分化方案主要基于wnt、mapk和notch信号传导通路的三重抑制。我们使用这种分化培养基并将干细胞因子头蛋白添加到分化混合物中,从而产生bmp低环境。令人惊讶地,我们观察到该培养物中的所有肠嗜铬细胞总是共同表达血清素和tac1,而缺乏分泌素表达。因此,这种培养物反映了eec激素特征,让人想起隐窝状态,暗示生态位信号在时间驱动的eec成熟中占主导地位。为了解决生态位信号是否确实可以协调eec激素特征,我们使用我们先前定义的eec分化系统作为起点,并在该混合物的背景下调节信号传导通路。加入vismogenib以抑制hedgehog信号传导,撤掉头蛋白以激活bmpr1a/bmpr2信号传导,并加入alk5/4/7抑制剂a83或tgf-β1以调节不同的tgf-β家族受体通路。虽然wnt已被porcupine抑制剂iwp2限制在eec分化混合物中,但我们在顶部除去r-spondin以获得更明显和快速的wnt抑制。我们分别使用sct和gcg转录物水平作为绒毛和隐窝激素特征的代理,而chga作为具有从隐窝到绒毛的恒定表达的转录物被包括。抑制hedghehog信号传导不会调节任何评估的激素,这与其在肠间充质中的主要作用一致。a83和tgf-β1均抑制所有eec激素的表达。r-spondin去除同样损害所评估的所有激素的eec分化。当wnt信号被抑制而不是在失去notch信号时选择分泌命运时,由于干细胞快速分化成吸收谱系,这可能是相关的。最后,在eec分化期间撤除头蛋白诱导sct的表达同时抑制gcg转录,这与从隐窝到绒毛表达谱的转换一致。我们将外源性bmp4添加到该eec分化混合物中以进一步放大bmp信号传导,并与我们之前定义的“eecbmp低”培养基相比,将该组合称为“eecbmp高”。用这种“eecbmp高”分化培养基刺激类器官诱导产生对分泌素具有免疫反应性的细胞,以及表达没有tac1的血清素的嗜铬细胞。在该方案中glp1+细胞数量大大减少(图21a-图21c),在类器官中没有明显的形态变化。从不同肠区域建立的类器官在激素特征方面保持其区域性质,其中在近端si中富集gip并且在远端si中富集pyy、nts和gcg。显示从隐窝到绒毛的均匀分布的其他激素或eec标志物包括chga、tph1、cck和十二指肠gip,仅受到不同bmp水平的轻微影响。我们确实观察到pyy和nts的上调,其在绒毛中表达最高。bmp的作用总结在下表中。总的来说,这些数据指出了bmp信号在诱导与绒毛中eec相关的激素中的作用。eecbmp高(bmp激活剂)eecbmp低(bmp抑制剂)绒毛样eec特性隐窝样eec特性高分泌素表达低分泌素表达低tac1和gcg表达高tac1和gcg表达低glp1+细胞数目高glp1+细胞数目实施例6在60小时的处理中,通过口服管饲将35mg/kg的ldn193189(selleckchem目录号s2618)施用给小鼠。将ldn193189溶于ph3.1的柠檬酸中,并每天两次给予口服管饲。与对照小鼠相比,这导致在肠绒毛中表达glp1的细胞扩增。与对照小鼠相比,肠绒毛中的细胞内分泌素表达显著降低(参见图22)。使用的剂量较高,并且预计更低剂量也是有效的。假设这些结果可以外推到人类身上。ldn193189是一种选择性bmp信号转导抑制剂,其抑制bmpi型受体alk2和alk3的转录活性(在c2c12细胞中ic50分别为5nm和30nm),对bmp展现出相对于tgf-β的选择性的200倍。bmp信号传导的其他抑制剂,如本文所述的那些,也被设想在体内具有类似的效果。可以得出结论,bmp抑制剂(和相反的bmp激活剂)具有用于其中需要体内调节glp1和/或分泌素的疗法的潜力。实例包括但不限于治疗糖尿病、肥胖、胃酸过少或胃酸过多。ldn193189的结构:序列表<110>荷兰皇家科学院(koninklijkenederlandseakademievanwetenschappen)<120>改进的分化方法<130>p068547wo<141>2017-06-20<150>gb1610748.4<151>2016-06-20<160>28<170>seqwin2010,version1.0<210>1<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>1ccagctgcctgtagtgtcaa20<210>2<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>2tcttgaggctcgccttgatg20<210>3<211>21<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>3gccaagagccatgtgactatc21<210>4<211>22<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>4cagagctggtactttggtgttc22<210>5<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>5acccgccagtctcctacatc20<210>6<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>6gcatctaggcgcagggattg20<210>7<211>18<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>7gctgtgcaagctgaaggg18<210>8<211>18<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>8gctgtgcaagctgaaggg18<210>9<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>9cagctcgtccactctttccg20<210>10<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>10cctctcgtctccttggaggg20<210>11<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>11ggaatggatggctaccgtgg20<210>12<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>12catgccacccatgctcgaat20<210>13<211>21<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>13actaaggtggcctacctccaa21<210>14<211>22<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>14ggaggtgacagtcaaggtgaga22<210>15<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>15aggatccatctgtcctttgg20<210>16<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>16acgttgtatgtcttggacag20<210>17<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>17gaccccaagacactcagacg20<210>18<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>18ttttctgtgtcctgctcgct20<210>19<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>19cttcccagaagaagtcgcca20<210>20<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>20gtgactggcacgagatgttg20<210>21<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>21gaagagcggcgtatgtctgt20<210>22<211>19<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>22ccagaaggagctttgcgga19<210>23<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>23gacctgcgactagactgacc20<210>24<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>24ccagttcctgtttcccggtg20<210>25<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>25aactgttggctaggggacac20<210>26<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>26tgatgaaagtcccctctgcg20<210>27<211>20<212>dna<213>人工序列(artificialsequence)<220><221><222><223>引物<400>27tgctgaccatcttccagctc20<210>28<211>20<212>dna<213>引物(primer)<400>28gaatgtagggccttctgggt20当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1