植物调节元件及其使用方法与流程
本申请是于2016年6月24日提交的美国临时申请号62/354,374的部分继续申请,将其内容以其全文通过引用并入本文。以电子方式提交的序列表的引用文件名为“5243wopctseqlist.txt”的序列表创建于2017年5月12日,大小为231千字节,以计算机可读形式与本说明书一并提交。该序列表是本说明书的一部分并且以其全文通过引用并入本文。本公开涉及植物分子生物学领域,更具体地,涉及植物中基因表达的调节。
背景技术:
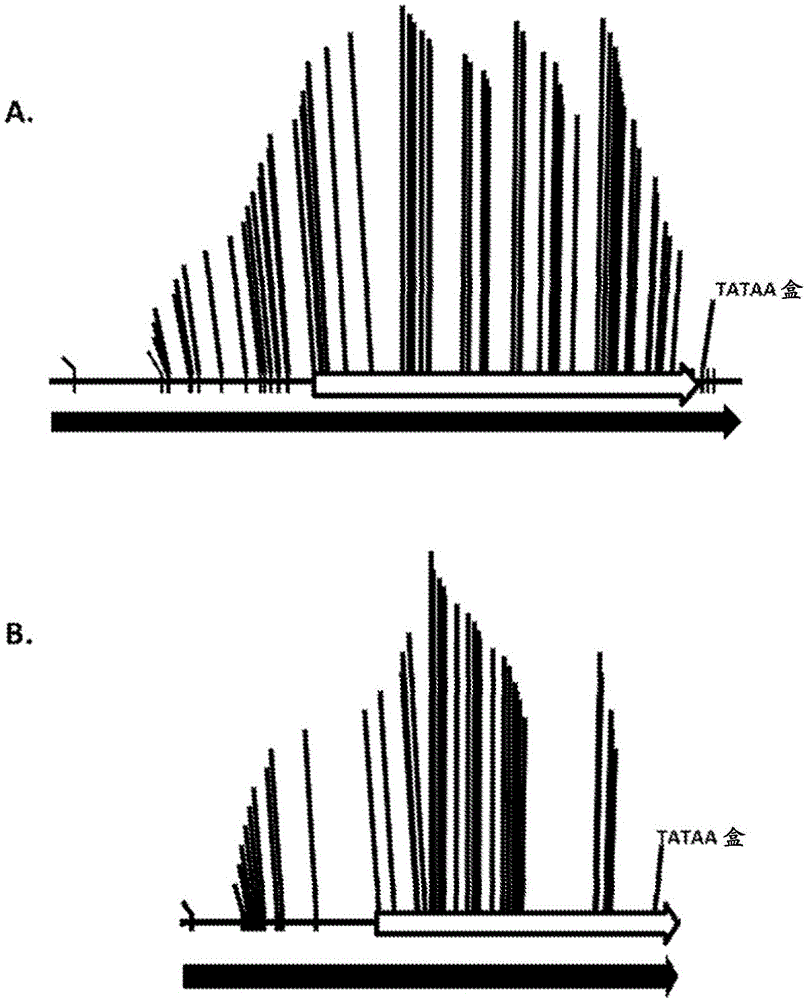
:植物宿主中异源dna序列的表达依赖于可操作地连接的调节元件的存在,这些调节元件在植物宿主内是具有功能的。选择的启动子序列可以决定异源dna序列在生物内表达的时间和位置。如果期望在特定的组织或器官中表达,则可使用组织偏好性启动子。如果期望基因响应刺激而表达,则诱导型启动子是首选调控元件。相比之下,如果期望在植物各细胞中连续表达,则采用组成型启动子。可以在转化载体的表达构建体中包含来自核心启动子序列的上游和/或下游的附加调节序列,以便引起异源核苷酸序列在转基因植物中以不同水平表达。经常,期望的是在植物的特定组织或器官中表达dna序列。例如,可通过遗传操纵植物基因组,使其包含可操作地连接至异源病原体抗性基因的组织偏好性启动子,使得所需的植物组织中产生病原体抗性蛋白,从而提高植物对土壤和空气传播的病原体感染的抗性。可替代地,可能需要抑制植物组织内天然dna序列的表达以实现所需的表型。在这种情况下,可采用下述方式实现这种抑制:转化植物,使植物包含可操作地连接至反义核苷酸序列的组织偏好性启动子,使得反义序列的表达产生干扰天然dna序列的mrna翻译的rna转录物。通过使用基因工程技术遗传地改变植物并且因此产生具有有用性状的植物可能需要获得多种调节元件。启动子和其他调节元件的积累将使研究者能够以所需水平和在所期望细胞位置表达重组dna分子。因此,启动子的集合将允许新性状以所需水平在所期望的组织中表达。因此,可以产生独特的表达模式并用作表达目的异源核苷酸序列的调节区的杂合调节元件的分离、表征和产生可用于植物的遗传操作。技术实现要素:提供了用于调节目的异源多核苷酸序列在植物或植物细胞中的表达的组合物和方法。提供了包含调节元件的新颖多核苷酸序列的合成dna分子,这些调节元件具有基因调节活性。在一些实施例中,这种调节元件具有在植物细胞中引发转录的启动子活性。某些实施例包括seqidno:1-74所示的核苷酸序列。还包括seqidno:1-74所示序列(其中所述序列在植物细胞中启动转录)的功能片段、区段或变体,或包含与seqidno:1-74所示序列(其中所述序列在植物细胞中启动转录)具有至少85%序列同一性的序列的多核苷酸序列。实施例还包括dna构建体,这些dna构建体包含可操作地连接至目的异源核苷酸序列的调节元件,其中所述调节元件能够驱动所述异源核苷酸序列在植物细胞中的表达并且所述调节元件包含本文所公开的核苷酸序列。实施例进一步提供了表达载体,以及如上所述在其基因组中稳定掺入dna构建体的植物或植物细胞。另外,组合物包括此类植物的转基因种子。在一个实施例中,提供了在植物中产生表达模式的杂合调节元件,相对于产生杂合调节元件的任何来源或亲本调节材料,每种杂合调节元件的表达模式是独特的,其中该杂合调节元件含有每个亲本调节元件的一个区段。在一个实施例中,相对于亲本调节元件,该杂合调节元件产生了不同的组织特异性表达模式。在另一个实施例中,相对于由给定组的亲本调节元件表达的较窄的表达模式,这些杂合调节元件在植物组织中提供了更宽的表达模式。在另一个实施例中,这些杂合调节元件可产生组成型表达模式,该表达模式与亲本调节元件的非组成型表达模式不同。提供了涉及杂合调节元件的构建的方法和组合物。在一个实施例中,可以使用一种或多种先前表征的调节元件和先前表征的一种或多种调节元件的组装区段来产生杂合调节元件。在一个另外的实施例中,可以评估表征的调节元件的潜在基序,以帮助定义可能在杂合调节元件中有用的区段。在一个实施例中,该杂合启动子的3’端含有来自亲本调节元件的tata盒。在另一个实施例中,提供的方法涉及在植物或植物细胞中表达核苷酸序列,该方法包括向该植物或植物细胞中引入表达盒,该表达盒包含可操作地连接到目的核苷酸序列的杂合调节元件,其中所述杂合调节元件产生了目的核苷酸序列的组织表达模式,该组织表达模式相对于每种单一的亲本调节元件组织表达模式是独特的。在一个实施例中,可用于制备杂合调节元件的亲本调节元件包含seqidno:1-10、31-34和61-74或其变体。还包括了seqidno:1-10、31-34和61-74所示多核苷酸序列(其中所述多核苷酸序列在植物细胞中启动转录)的功能片段、区段或变体,或包含与seqidno:1-10、31-34和61-74所示多核苷酸序列(其中所述多核苷酸序列在植物细胞中启动转录)具有至少85%序列同一性的序列的多核苷酸序列。在另一个实施例中,可用于制备杂合调节元件的亲本调节元件包含seqidno:1-74中的一个或多个或其变体。实施例还包括dna构建体,这些dna构建体包含可操作地连接至目的异源核苷酸序列的杂合调节元件,其中所述杂合调节元件能够驱动所述异源核苷酸序列在植物细胞中的表达并且所述启动子包含如本文所公开的seqidno:1-74中的一个或其功能片段。实施例进一步提供了表达载体,以及如上所述在其基因组中稳定掺入dna构建体的植物或植物细胞。另外,组合物包括此类植物的转基因种子。在调节元件的转录起始区的下游将是目的序列,其将提供对植物的表型的修饰。这种修饰包括调控内源产物的生成(数量、相对分布等方面)或者外源表达产物的生成,以在植物中提供新颖的或经调控的功能或者产物。例如,涵盖了编码基因产物的异源多核苷酸序列,该基因产物赋予对除草剂、盐、寒冷、干旱、病原体、线虫类或昆虫的抗性或耐性。在一个另外的实施例中,提供了用于调控基因在稳定转化的植物中的表达的方法,该方法包括以下步骤:(a)用包含本文所公开的调节元件或其功能片段的dna构建体转化植物细胞,该调节元件或其功能片段可操作地连接到至少一种异源多核苷酸序列上;(b)使该植物细胞在植物生长条件下生长,以及(c)从其中该连接的核苷酸序列的表达改变该植物的表型的植物细胞再生出稳定转化的植物。在另一个实施例中,该dna构建体进一步包含异源增强子元件。提供了表达盒,这些表达盒包含可操作地连接至目的异源核苷酸序列的seqidno:1-74的调节元件序列。另外,还提供了转化的植物细胞、植物组织、种子和植物。附图说明图1示出了用于辅助定义调节元件区段的区域的基序的分布,所述调节元件区段将被掺入杂合调节元件中。垂直线表示基序位置,白色箭头表示在杂合调节元件中使用的区段,并且黑色箭头表示全长亲本调节元件。a.si-tip2-3b(mod1)调节元件(seqidno:10)和tataa盒上游5′区域(基于基序聚类而选择用作杂合调节元件中的区段)的示意图,和b.bd-grp3pro(mod1)调节元件(seqidno:2)和包括tataa盒的3′下游区域(基于基序聚类而选择用作杂合调节元件中的区段)的示意图。图2示出了用于制备若干根杂合调节元件的区段。图3示出了用于制备各地上部分(aerial)杂合调节元件的区段。具体实施方式本文所用的冠词“一个/种”是指一个/种或多于一个/种(即,指至少一个/种)所述冠词的语法对象。通过举例,“一个元件”是指一个或多个元件。本公开涉及针对植物调节元件的组合物和方法以及它们的使用方法。这些组合物和方法进一步涉及杂合调节元件以及制备和使用杂合调节元件的方法。植物组织是复杂的器官,其可分为不同的区域;例如,在根组织中,组织区域包括根冠、分生组织、伸长区和成熟区。这些区域反映了细胞身份和成熟度的不同阶段。因此,这些细胞中的基因在发育期间被激活和灭活,并且因此可能不会在根或其他组织的所有区域中同时表达。然而,如本文所教导的,组合来自这些基因的调节元件的部件,可以产生合成调节元件,该合成调节元件驱动在特定组织的不同区域中的表达。尽管可以产生独特的表达模式,但所提供的调节元件可以保留组织特异性。本公开涉及针对植物杂合调节元件的组合物和方法以及它们的使用方法。这些组合物包含seqidno:1-74所示的调节元件的多核苷酸序列及其片段、变体和互补序列。在一个实施例中,这些组合物包含seqidno:11-30、35-47和50-59所示的杂合调节元件的多核苷酸序列及其片段、变体和互补序列。组合物可以包括调节元件的核苷酸序列、其片段和变体。在具体实施例中,本文所公开的杂合调节元件序列相对于天然调节元件可用于改变表达模式。这些核苷酸序列还用于构建后续用于在目的植物中表达异源核苷酸序列的表达载体,或用作用于分离其他调节元件的探针。提供了dna构建体的一个实施例,这些dna构建体包含可操作地连接至目的异源多核苷酸序列的seqidno:1-10、31-34和61-74所示的调节元件多核苷酸序列,或seqidno:11-30和35-60所示的杂合调节元件、或其功能片段或变体,及其任何组合。术语“调节元件”是指具有基因调节活性的核酸分子,也就是能够影响可操作地连接的可转录多核苷酸的转录和/或翻译表达模式的核酸分子。因此,术语“基因调节活性”是指通过影响可操作地连接的可转录多核苷酸分子的转录和/或翻译,从而影响这种可操作地连接的可转录多核苷酸分子表达的能力。基因调节活性可为正向和/或负向,并且所述影响可通过其下列特性来表征:时间、空间、发育、组织、环境、生理、病理、细胞周期和/或化学响应,还可通过定量指示或定性指示来表征。调节元件(例如启动子、增强子、前导序列和内含子区域)是具有基因调节活性的核酸分子,并且也是活细胞中基因整体表达的不可缺少的一部分。因此,可通过基因工程方法,使用在植物中起作用的分离的调节元件(如启动子和前导序列)来改变植物表型。启动子可用作调节元件,用来调控可操作地连接的可转录多核苷酸分子的表达。如本文所用,“基因表达模式”为可操作地连接的核酸分子转录成转录的rna分子的任何模式。表达可通过其下列特性来表征:时间、空间、发育、组织、环境、生理、病理、细胞周期和/或化学响应,还可通过定量指示或定性指示来表征。转录的rna分子可被翻译生成蛋白质分子,或可提供反义rna分子或其他调节rna分子,如dsrna、trna、rrna、mirna等。如本文所用,“独特的基因表达模式”是可操作地连接的核酸分子转录成为转录的rna分子的任何模式,该模式与另一种调节元件的差异在于其特定组织或亚组织区域转录表达模式或者细胞类型或细胞状态表达模式,而不仅仅是定量转录量的差异。如本文所用,“独特的基因表达模式”不包括组成型表达模式。在一个实施例中,独特的基因表达模式包括在特定组织中异源核酸的转录的减少或消除。在另一个实施例中,独特的基因表达模式包括在特定组织中异源核酸的转录的增加或产生。如本文所用,“区域转录表达模式”或“亚区域转录表达模式”旨在是指任何非遍在表达模式,该表达模式仅在一个或多个特定组织的某些区域中具有可检测的表达,并且在该一个或多个区域外的任何其他表达都应该处于标称基线水平(“异位表达”)。术语“地上部分表达”是指在植物的一个或多个地上组织中的多核苷酸的基因表达模式。在一个实施例中,目的异源多核苷酸序列以根遍在表达模式或地上部分遍在表达模式表达。在一个实施例中,地上部分表达不包括在植物的花粉中表达。这些调节元件序列或其变体或片段当可操作地连接至目的异源核苷酸序列时,可以驱动该异源多核苷酸序列在表达该构建体的植物组织中的组成型表达。术语“组成型表达”是指在整个植物或植物的大部分组织中发现异源核苷酸序列的表达。如本文所用,术语“蛋白质表达”为转录的rna分子翻译成蛋白质分子的任何模式。蛋白质表达可通过其下列特性来表征:时间、空间、发育或形态等,还可通过定量指示或定性指示来表征。如本文所用,术语“启动子”一般是指参与识别并结合rna聚合酶ii和其他蛋白质(反式作用转录因子)以启动转录的核酸分子。启动子最初可从基因的基因组拷贝的5′侧翼区分离。可替代地,启动子可以是合成生产的dna分子或受操纵的dna分子。调节元件可包含启动子和启动子活性。在一个实施例中,提供了本文公开的调节元件的片段。调节元件片段可表现出启动子活性,并且可单独用于、或与其他调节元件和调节元件片段结合用于例如构建杂合调节元件。在具体实施例中,提供了调节元件的多个片段,这些片段包含本文公开的具有启动子活性的多核苷酸分子中的至少约50、95、150、250、500或约750或更多个连续核苷酸或可替代地由其组成或基本上由其组成。当将此类片段与参考序列进行最佳比对时,可表现出与参考序列的至少约85%、约90%、约95%、约98%、或约99%或更高的同一性。如本文所用,术语“调节元件区段”是调节元件的片段,该片段的特征在于至少使用由higo,k等人(1998)nucleicacidsresearch[核酸研究](以其全文通过引用并入)所述的那些的大量的可识别的调节元件基序,其中当与至少一种其他调节元件区段组合时该调节元件区段产生所希望的或独特的表达模式。如本文所用,“调节元件区段”与“区段”、“限定的区段”和“亲本区段”可互换使用。在一个实施例中,大量的可识别的调节元件基序包括至少25个基序、至少30个基序、至少40个基序或至少50个基序。在具体实施例中,提供了调节元件的多个区段,这些区段包含本文公开的多核苷酸分子的至少约50、95、150、250、500、750、1000、1250、1500、或约1750个连续核苷酸。在另一个实施例中,区段包含多核苷酸分子的至少约50、95、150、250、500、750、1000、1250、1500或约1750个连续核苷酸,但小于亲本多核苷酸分子的全长。在一个实施例中,调节元件区段包含亲本调节元件的片段,其中该片段包含亲本调节元件的不超过约2000、1900、1800、1700、1600、1500、1400、1300、1200、1100、1000或500个连续核苷酸。在一个另外的实施例中,杂合调节元件包含连续多核苷酸区段,其中这些连续多核苷酸区段在植物细胞中启动转录并产生独特的组织表达模式。如本文所用,“组装体”是指两个或更多个可操作地连接的调节元件区段。在一个实施例中,区段的组装体包括连续的区段。在另一个实施例中,区段的组装体包括这些区段的一个或多个,这些区段是不连续的但是仍然会产生表达模式。还可以分析调节元件或调节元件区段中已知的启动子基序,即dna序列特征(例如tataa盒和其他已知的转录因子结合位点基序)的存在。本领域技术人员可以使用此类已知基序的鉴定来设计与来源或亲本调节元件相比具有希望的或独特的表达模式的杂合调节元件。先前已经表征了在调节元件中发现的核苷酸序列基序并且其可获得于place数据库(higo,k等人(1998)nucleicacidsresearch[核酸研究];dna.affrc.go.jp/htdocs/place/,使用“www”前缀可在万维网上访问;还参见pct申请号wo2014/164399)。在一个实施例中,基序包含至少六个核苷酸的序列。在一些实施例中,调节元件区段包含每1000个核苷酸约15、20、25、30、35、40、45、50、75、100、125或150个基序。在一些实施例中,调节元件包含约每5、10、15、20、25、30、35、40、45或50个核苷酸至少一个基序。如本文所用,术语“增强子”或“增强子元件”是指顺式作用转录调节元件,也称为顺式元件,其赋予整体表达模式的一个方面,但单靠其通常不足以驱动可操作地连接的多核苷酸序列的转录。与启动子不同,增强子元件通常不包括转录起始位点(tss)或tata框。调节元件可天然地包含一个或多个增强子元件,这些增强子元件影响可操作地连接的多核苷酸序列的转录。分离的增强子元件还可与异源启动子融合,以产生异源启动子顺式元件,该元件赋予整体调控基因表达的一个方面。本文公开的调节元件或调节元件片段可包含一个或多个增强子元件,这些增强子元件影响可操作地连接的基因的转录。据信,许多增强子元件结合dna结合蛋白和/或影响dna拓扑结构,从而产生局部构象,所述局部构象选择性地允许或限制rna聚合酶接近dna模板或有利于在转录起始位点处选择性打开双螺旋。增强子元件可以用以结合调节转录的转录因子。有些增强子元件结合多于一种转录因子,而且转录因子可以不同的亲和力与多于一种增强子结构域相互作用。可采用多种技术鉴定增强子元件,这些技术包括:缺失分析,也就是从启动子5′端或内部缺失一个或多个核苷酸;使用dnasei足印法、甲基化干扰、电泳迁移率改变测定、通过连接介导pcr执行的体内基因组足印法和其他常规测定进行dna结合蛋白分析;或者使用已知的顺式元件基序或增强子元件作为靶序列或靶基序,采用常规dna序列比较方法(如blast)进行dna序列相似性分析。增强子结构域的精细结构可通过一个或多个核苷酸的诱变(或取代)或通过其他常规方法进一步研究。增强子元件可通过化学合成获得,或从包含此类元件的调节元件中分离获得;并且增强子元件可与另外的含有可用限制性酶切位点的侧翼核苷酸一起合成,以便后续操纵。因此,涵盖了根据本文所公开的方法设计、构建和使用增强子元件,用于调控可操作地连接的可转录多核苷酸分子的表达。如本文所用,术语“5′侧翼区”是指从基因的基因组拷贝中分离的dna分子,并且通常定义为从蛋白质编码序列起始位点开始并5′延伸至5′非翻译区并进入启动子区的多核苷酸区段。这些序列或前导序列可为合成产生的dna元件或受操纵dna元件。前导序列可用作5′调节元件,用来调控可操作地连接的可转录多核苷酸分子的表达。前导序列分子可与异源元件或其天然元件一同使用。如本文所用,术语“杂合”是指通过将第一个dna分子与第二个dna分子融合而生成的单个合成的dna分子,其中第一个dna分子和第二个dna分子通常都不会以该构型(也就是与另一个dna分子融合的构型)存在。因此,杂合dna分子为通常不会天然存在的全新dna分子。如本文所用,术语“杂合调节元件”是指将dna分子这样操纵而产生的调节元件。杂合调节元件可结合两个或更多个dna区段。因此,涵盖了根据本文所公开的方法设计、构建和使用杂合调节元件,用于调控可操作地连接的可转录多核苷酸分子的表达。在一个实施例中,杂合调节元件包含两个或更多个dna区段。在另一个实施例中,杂合调节元件包含三个或更多个dna区段。在另一个实施例中,杂合调节元件包含四个或更多个dna区段。在一个实施例中,dna片段区段可以是亲本区段。如本文所用,“区段”和“亲本区段”是可互换的并且旨在是指天然“亲本调节元件”的片段,分析了这些片段中预测产生区域组织表达模式的基序。亲本区段或其变体的组合可产生杂合调节元件,该杂合调节元件以遍在组织表达模式表达目的基因,该遍在组织表达模式相对于亲本调节元件的每个单独的表达模式是独特的。在一个实施例中,亲本区段可以是亲本调节元件的变体。在一个实施例中,如seqidno:1-10、31-34和61-74所示的亲本调节元件可用作产生亲本区段及其变体的亲本调节元件。作为亲本调节元件还包括seqidno:1-10、31-34和61-74所示多核苷酸序列(其中所述多核苷酸序列在植物细胞中启动转录)的功能片段、区段或变体,和包含与seqidno:1-10、31-34和61-74所示多核苷酸序列(其中所述多核苷酸序列在植物细胞中启动转录)具有至少85%序列同一性的序列的多核苷酸序列。在某些实施例中,亲本调节元件可包括来源于植物的亲本调节元件。在另一个实施例中,亲本调节元件可包括来源于细菌或病毒的亲本调节元件。提供了在植物中产生表达模式的杂合调节元件,该杂合调节元件相对于亲本调节元件是独特的,其中该杂合调节元件含有多于一个亲本调节元件的区段或片段。在一个实施例中,相对于亲本调节元件,该杂合调节元件产生了不同的组织特异性表达模式。在另一个实施例中,相对于由给定组的亲本调节元件表达的区域组织表达模式,这些杂合调节元件将在植物组织中的表达模式扩展为遍在表达模式。在另一个实施例中,相对于由给定组的亲本调节元件表达的较宽范围的表达模式,这些杂合调节元件表达较窄范围的表达。在另一个实施例中,这些杂合根调节元件可产生组成型表达模式,该组成型表达模式与亲本调节元件的非组成型表达模式不同。在一个实施例中,位于编码区序列的内含子内或3′的本文公开的多核苷酸序列也可有助于调节目的编码区的表达。合适的内含子的实例包括但不限于玉米ivs6内含子或玉米肌动蛋白内含子。调节元件还可包括那些位于转录起始位点的下游(3′)、或转录的区域内、或两者处的元件。转录后调节元件可包括在转录起始之后活跃的元件,例如翻译和转录增强子、翻译和转录阻遏子、以及mrna稳定性决定子。调节元件、或其变体或片段可以与一个或多个异源调节元件可操作地相关联以调控异源调节元件的活性。此类调控包括增强或抑制异源调节元件的转录活性、调控转录后事件、或者增强或抑制异源调节元件的转录活性并调控转录后事件。例如,可将一个或多个调节元件或其片段与组成型、诱导型或组织特异性启动子或其片段可操作地关联,以调控这样的启动子在植物细胞中的所希望组织内的活性。这些组合物可以涵盖分离的或重组的核酸。“分离的”或“重组的”核酸分子在本文用来指不再处于其天然环境中,而是例如处于体外的或异源重组细菌或植物宿主细胞中的核酸序列。当通过重组技术产生时,分离的或重组的核酸分子或其生物活性部分基本上不含其他细胞材料或培养基,或者当用化学法合成时基本上不含化学前体或其他化学物质。分离的或重组的核酸不含在衍生该核酸的生物体的基因组dna中天然位于该核酸的侧翼的序列(也就是位于该核酸5′和3′端的序列)(最佳地是蛋白质编码序列)。例如,在多个实施例中,分离的核酸分子可含有少于约5kb、4kb、3kb、2kb、1kb、0.5kb或0.1kb的核苷酸序列,这些核苷酸序列在衍生该核酸的细胞的基因组dna中天然地位于该核酸分子的侧翼。本文所公开的杂合调节元件序列可以从位于它们各自的转录起始位点侧翼的5′非翻译区分离。如本文所用,术语“多核苷酸”和“核苷酸”都旨在意指一个或多个核苷酸,并且能以单数或复数形式互换使用。本公开还涵盖了所公开的调节元件多核苷酸序列的片段和变体。如本文所用,术语“片段”是指核酸序列的一部分。调节序列的片段可保留启动转录,更具体地以组织特异性或亚组织特异性方式驱动转录的生物活性。可替代地,可用作杂交探针的多核苷酸序列的片段可以不必保留生物活性。调节区的多核苷酸序列的片段范围可从至少约20个核苷酸、约50个核苷酸、约100个核苷酸和高至比seqidno:1-74中任一个的全长少一个核苷酸。调节元件的生物活性部分可通过分离调节序列的一部分并评估该部分的启动子活性来制备。作为调节多核苷酸序列的片段的核酸分子包含至少约16、50、75、100、150、200、250、300、350、400、450、500、550、600、650、700或800个核苷酸或者高至本文所公开的全长调节序列中存在的核苷酸数目。对于多核苷酸序列,变体包含在天然多核苷酸序列中的一个或多个内部位点处的一个或多个核苷酸的缺失和/或添加,和/或在天然多核苷酸中的一个或多个位点处的一个或多个核苷酸的取代。对于多核苷酸序列,可使用熟知的分子生物学技术来鉴定变体,例如,用下文概述的聚合酶链反应(pcr)和杂交技术来鉴定。变体多核苷酸序列可以包括合成来源的多核苷酸序列,例如通过使用定点诱变而产生的那些。通常,如通过本文别处所描述的序列比对程序和参数确定的,本公开的具体核苷酸序列(包括seqidno:1-74)的变体将与该具体的核苷酸序列具有至少约40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高的序列同一性。本公开的多核苷酸序列的生物活性变体可能与该序列相差只有1-15个核酸残基,相差只有1-10个,只有6-10个,只有5个,只有4、3、2个,或甚至只有1个核酸残基。变体多核苷酸序列还涵盖由诱变和重组发生程序(如dna改组)衍生的序列。采用此类程序时,可操纵调节元件多核苷酸序列,以创建新的调节元件。以此方式,由一群相关的序列多核苷酸产生重组多核苷酸文库,这些多核苷酸包含具有实质序列同一性并且能够在体外或体内同源重组的序列区域。这种dna改组的策略在本领域中是已知的。参见例如,stemmet(1994)proc.natl.acad.sci.usa[美国科学院院报]91:10747-10751;stemmer(1994)nature[自然]370:389-391;crameri等人(1997)naturebiotech.[自然生物技术]15:436-438;moore等人(1997)j.mol.biol.[分子生物学杂志]272:336-347;zhang等人(1997)proc.natl.acad.sci.usa[美国科学院院报]94:4504-4509;crameri等人(1998),nature[自然],391:288-291;以及美国专利号5,605,793和5,837,458。本公开的多核苷酸序列可用于分离来自其他生物,尤其是其他植物,更尤其是其他单子叶植物的相应序列。以这种方式,可以使用如pcr、杂交等方法来鉴定此类序列(基于其与本文所述序列的序列同源性)。本公开涵盖了基于与本文所示的完整序列或者与完整序列的片段的序列同一性而分离的序列。在pcr方法中,可以设计寡核苷酸引物,用于在pcr反应中由从任何目的植物中提取的cdna或基因组dna扩增相应的dna序列。用于设计pcr引物与pcr克隆的方法是本领域普遍已知的并公开于sambrook(同上)中。还参见,innis等人编辑(1990)pcrprotocols:aguidetomethodsandapplications[pcr方案:方法和应用指南](academicpress[学术出版社],纽约);innis和gelfand编辑(1995)pcrstrategies[pcr策略](academicpress[学术出版社],纽约);以及innis和gelfand编辑(1999)pcrmethodsmanual[pcr方法手册](academicpress[学术出版社],纽约)。已知的pcr方法包括但不限于:使用成对引物、巢式引物、单特异性引物、简并引物、基因特异性引物、载体特异性引物、部分错配引物等的方法。在杂交技术中,利用已知多核苷酸序列的全部或部分作为探针,该探针选择性杂交来自所选生物体的一群克隆基因组dna片段或cdna片段(即基因组或cdna文库)中存在的其他相应多核苷酸序列。这些杂交探针可以是基因组dna片段、cdna片段、rna片段、或其他寡核苷酸,并且可以用可检测基团如32p或任何其他可检测标记物进行标记。因此,例如,杂交用探针可通过基于本公开的调节元件序列对合成的寡核苷酸进行标记来制备。用于制备杂交用探针和用于构建基因组文库的方法是本领域普遍已知的并且公开于sambrook(同上)中。例如,本文所公开的完整调节元件序列或其一个或多个部分可用作能够与相应的调节元件序列和信使rna特异性杂交的探针。要在多种条件下实现特异性杂交,此类探针包含的序列是调节元件序列中独特的,并且通常其长度为至少约10个核苷酸或者其长度为至少约20个核苷酸。可以使用此类探针通过pcr由选择的植物扩增相应的调节元件序列。可以使用这种技术从所希望的生物体中分离另外的编码序列,或作为用于确定生物体中存在编码序列的诊断试验。杂交技术包括杂交筛选铺板的dna文库(斑块或集落,参见例如sambrook,同上)。一般而言,具有启动子活性并且与本文所公开的多核苷酸序列及其片段杂交的序列将与所公开的序列具有至少40%至50%的同源性,约60%、70%、80%、85%、90%、95%至98%或更高的同源性。即,序列的序列相似性可以在一定范围内,共有至少约40%至50%、约60%至70%、以及约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的序列同一性。相对于参考序列(主题序列),“序列同一性百分比(%)”被确定为在比对序列并引入空位(如果需要)以实现最大百分比序列同一性后,并且不考虑作为序列同一性的一部分的任何氨基酸保守取代,候选序列(查询序列)中与参考序列中的相应氨基酸残基或核苷酸同一的氨基酸残基或核苷酸的百分比。用于确定序列同一性百分比目的而进行的比对能以本领域技术范围内的各种方式实现,例如,使用公共可用的计算机软件,例如blast、blast-2。本领域的技术人员可以确定用于比对序列的适当参数,包括在进行比较的序列的全长度上实现最大比对所需的任何算法。两个序列之间的同一性百分比是序列共有的同一位置的数目的函数(例如,查询序列的同一性百分比=查询序列和主题序列之间的同一位置的数目/查询序列的位置总数×100)。对本公开的分离的调节元件序列进行修饰可以提供异源多核苷酸序列的表达范围。因此,可将这些调节元件序列修饰为弱启动子或强启动子。通常,“弱启动子”是指以低水平驱动编码序列的表达的启动子。“低水平”表达旨在是指以约1/10,000个转录物至约1/100,000个转录物至约1/500,000个转录物的水平表达。相反地,强启动子以高水平或以约1/10个转录物至约1/100个转录物至约1/1,000个转录物的水平驱动编码序列的表达。本文公开的调节元件可用来增加或减少表达,从而导致转化的植物的表型改变。本文所公开的多核苷酸序列及其变体和片段可用于任何植物的遗传操纵。这些调节元件序列在与待控制其表达以实现所需表型响应的异源核苷酸序列可操作地连接时,可用于这个方面。术语“可操作地连接的”,意指异源核苷酸序列的转录或翻译受调节元件序列的影响。以此方式,可将本文公开的调节元件序列与目的异源多核苷酸序列一起在表达盒中提供,以在目的植物中表达,更特别地在转化的植物的生殖组织中表达。可在dna构建体中提供实施例的调节元件,用于在目的生物体中表达。如本文所用,“表达盒”意指包含实施例的调节元件的dna构建体,其中该调节元件可操作地连接至表达转录物或目的基因的异源多核苷酸。此类表达盒将包含转录起始区,该转录起始区包含可操作地连接至异源核苷酸序列的本公开的调节元件多核苷酸序列之一或其变体或片段。此类表达盒可具有多个限制位点,用于使该多核苷酸序列的插入受调节区的转录调节。表达盒可另外含有选择性标记基因以及3’终止区。表达盒在转录的5’-3’方向上可包含转录起始区(即本公开的杂合启动子或其变体或片段)、翻译起始区、目的异源多核苷酸序列、翻译终止区、以及任选地在宿主生物中有功能的转录终止区。实施例的调节区(即启动子、增强子、转录调节区和翻译终止区)和/或多核苷酸对于宿主细胞或彼此之间可以是天然的/类似的。可替代地,实施例的调节区和/或多核苷酸对于宿主细胞或彼此之间可以是异源的。如本文所用,关于序列的“异源性”是指该序列源于外来物种,或者,如果源于相同物种的话,则是通过蓄意人为干预从其在组合物和/或基因组基因座中的天然形式进行实质性修饰得到的序列。例如,可操作地连接至异源多核苷酸的调节元件来自与从其衍生该多核苷酸的物种不同的物种,或者,如果来自相同/类似的物种,那么一方或双方基本上由它们的原来形式和/或基因组基因座进行实质性修饰得到,或者该调节元件不是被可操作地连接的多核苷酸的天然调节元件。终止区对于转录起始区可以是天然的,对于可操作地连接的目的dna序列可以是天然的,对于植物宿主可以是天然的,或者可能源自另一种来源(即,对于调节元件、被表达的dna序列、植物宿主或其任何组合来说为外源的或异源的)。本文所公开的调节元件及其变体和片段可用于植物的基因工程,例如,用于产生转化的或转基因的植物,来表达目的表型。如本文所用,术语“经转化的植物”和“转基因植物”是指在其基因组内包含异源多核苷酸的植物。通常,异源多核苷酸稳定整合于转基因或转化的植物基因组内,这样使得多核苷酸得以传递至连续世代。异源多核苷酸可以单独地或作为重组dna构建体的部分整合进基因组中。应当理解的是,如本文所用,术语“转基因的”包括其基因型已经通过异源核酸的存在而改变的任何细胞、细胞系、愈伤组织、组织、植物部分或植物,包括最初如此改变的那些转基因以及通过有性杂交或无性繁殖从初始转基因产生的那些。通过以下来产生转基因事件:用包含核酸表达盒的异源dna构建体转化植物细胞,该核酸表达盒包含目的转基因;再生由该转基因插入该植物的基因组中所产生的植物群体;并且选择表征为插入特定基因组位置的特定植物。事件在表型上表征为转基因的表达。在遗传水平上,事件是植物遗传组成的一部分。术语“事件”也指转化株和另一个植物之间有性杂交所产生的后代,其中所述后代包含异源dna。如本文所用,术语植物包括全植物、植物器官(例如,叶、茎、根等)、植物细胞、植物原生质体、可再生植物的植物细胞组织培养物、植物愈伤组织、植物块和在植物或植物部分(如胚、花粉、胚珠、种子、叶、花、枝、果、核、穗、穗轴、壳、茎、根、根尖、花药等)中完好的植物细胞。籽粒意指由商业种植者出于栽培或繁殖物种之外的目的所生产的成熟种子。再生的植物的子代、变体和突变体也包括在本公开的范围内,其条件是这些部分包含引入的多核苷酸。如本文所用,术语tis包括本文公开的组合物和方法可用于任何植物物种(包括但不限于单子叶植物和双子叶植物)的转化。植物物种的实例包括玉米(corn,zeamays),芸苔属物种(brassicasp.)(例如,甘蓝型油菜(b.napus)、芜菁(b.rapa)、芥菜(b.juncea))(特别是可用作种子油来源的那些芸苔属物种),苜蓿(紫花苜蓿(medicagosativa)),稻(rice,oryzasativa),黑麦(rye,secalecereale),高梁(sorghum,sorghumbicolor,sorghumvulgare)),粟米(例如,珍珠粟(御谷(pennisetumglaucum)),黍(prosomillet,panicummiliaceum),粟(foxtailmillet,setariaitalica),穇子(fingermillet,eleusinecoracana),向日葵(sunflower,helianthusannuus),红花(safflower,carthamustmctorius),小麦(wheat,triticumaestivum),大豆(soybean,glycinemax),烟草(tobacco,nicotianatabacum),马铃薯(potato,solanumtuberosum),花生(peanut,arachishypogaea),棉花(海岛棉(gossypiumbarbadense)、陆地棉(gossypiumhirsutum)),甘薯(sweetpotato,ipomoeabatatas),木薯(cassava,manihotesculenta),咖啡(咖啡属物种(coffeaspp.)),椰子(coconut,cocosnucifera),菠萝(pineapple,ananascomosus),柑橘树(柑橘属物种(citrusspp.)),可可(cocoa,theobromacacao),茶树(tea,camelliasinensis),香蕉(芭蕉属物种(musaspp.)),鳄梨(avocado,perseaamericana),无花果(fig,ficuscasica),番石榴(guava,psidiumguajava),芒果(mango,mangiferaindica),橄榄(olive,oleaeuropaea),木瓜(番木瓜(caricapapaya)),腰果(cashew,anacardiumoccidentale),澳洲坚果(macadamia,macadamiaintegrifolia),巴旦杏(almond,prunusamygdalus),甜菜(sugarbeets,betavulgaris),甘蔗(甘蔗属物种(saccharumspp.)),燕麦,大麦,蔬菜,观赏植物和针叶树。蔬菜包括番茄(tomatoes,lycopersiconesculentum)、莴苣(例如,莴苣(lactucasativa))、青豆(菜豆(phaseolusvulgaris))、利马豆(limabean,phaseoluslimensis)、豌豆(香豌豆属物种(lathyrusspp.))和黄瓜属(cucumis)的成员诸如黄瓜(cucumber,c.sativus)、香瓜(cantaloupe,c.cantalupensis)和甜瓜(muskmelon,c.melo)。观赏植物包括杜鹃(杜鹃花属物种(rhododendronspp.))、绣球花(hydrangea,macrophyllahydrangea)、木槿(hibiscus,hibiscusrosasanensis)、玫瑰(蔷薇属物种(rosaspp.))、郁金香(郁金香属物种(tulipaspp.))、水仙(水仙属物种(narcissusspp.))、矮牵牛(petunias,petuniahybrida)、康乃馨(carnation,dianthuscaryophyllus)、一品红(poinsettia,euphorbiapulcherrlma)和菊花。可以使用的针叶树包括例如松树,诸如火炬松(loblollypine,pinustaeda)、湿地松(slashpine,pinuselliotii)、西黄松(ponderosapine,pinusponderosa)、黑松(lodgepolepine,pinuscontorta)和辐射松(montereypine,pinusradiata);花旗松(douglas-fir,pseudotsugamenziesii);西方铁杉(westernhemlock,tsugacanadensis);北美云杉(sitkaspruce,piceaglauca);红杉(redwood,sequoiasempervirens);冷杉,如银杉(胶冷杉(abiesamabilis))和香脂冷杉(balsamfir,abiesbalsamea);以及雪松,如西部红雪松(thujaplicata)和阿拉斯加黄雪松(黄扁柏(chamaecyparisnootkatensis))。在特定实施例中,植物可以是作物植物(例如,玉米、苜蓿、向日葵、芸苔属植物、大豆、棉花、红花、花生、高粱、小麦、粟米、烟草等)。在其他实施例中,玉米和大豆植物是最佳的,并且在又其他的实施例中,玉米植物是最佳的。其他目的植物包括提供目的种子的谷物类植物、油料种子植物和豆科植物。目的种子包括谷物种子,例如玉米、小麦、大麦、稻、高粱、黑麦等。油料种子植物包括棉花、大豆、红花、向日葵、芸苔属、玉蜀黍、苜蓿、棕榈、椰子等。豆科植物包括豆类和豌豆。豆类包括瓜耳豆、槐豆、胡芦巴、大豆、四季豆、豇豆、绿豆、利马豆、蚕豆、小扁豆、鹰嘴豆等。本文所公开的调节元件序列所表达的异源编码序列可用于改变植物的表型。表型的各种变化是所关注的,包括修饰基因在植物中的表达,改变植物对病原体或昆虫的防御机制,提高植物对除草剂的耐受性,改变植物发育以响应环境胁迫,调控植物对盐、温度(热和寒冷)、干旱等的响应。这些结果可通过表达包含适当基因产物的目的异源多核苷酸序列来获得。在特定实施例中,目的异源多核苷酸序列是在植物或植物部分中表达水平提升的植物内源序列。可以通过提供改变的一个或多个内源基因产物(特别是激素、受体、信号分子、酶、转运蛋白或辅因子)的表达或通过影响植物中的营养摄取来获得结果。这些改变导致转化的植物的表型的变化。在某些实施例中,本文公开的调节元件的表达模式可用于许多类型的筛选。可以与本文所公开的调节序列一起使用的目的多核苷酸序列的大体种类包括例如涉及信息的那些基因(如锌指)、涉及通信的那些基因(如激酶)和涉及管家的那些基因(如热休克蛋白)。转基因的更具体的类别例如包括编码农艺学、昆虫抗性、抗病性、除草剂抗性、环境胁迫抗性(对寒冷、盐、干旱等的耐受性改变)和谷物特征的重要性状的基因。仍另外的转基因种类包括从植物和其他真核生物以及原核生物诱导外源性产物(如酶、辅因子及激素)的表达的基因。应认识到,任何目的基因可以可操作地连接至本公开的调节元件并在植物中表达。以说明的方式,而非意图限制,以下是可与本文公开的调节元件一起使用的基因类型的其他实例的列表。1.转基因,其赋予对昆虫或疾病的抗性并编码:(a)植物抗病性基因。通常通过植物中抗病性基因(r)的产物与病原体中相应的无毒性(avr)基因的产物之间的特异性相互作用来激活植物防御。可以用经克隆的抗性基因来转化植物变种,以工程化对特定病原菌株具有抗性的植物。参见,例如jones等人,(1994)science[科学]266:789(cloningofthetomatocf-9geneforresistancetocladosporiumfulvum[克隆番茄cf-9基因以抵抗番茄叶霉病菌]);martin等人,(1993)science[科学]262:1432(tomatoptogeneforresistancetopseudomonassyringaepv.tomatoencodesaproteinkinase[用于抵抗丁香假单胞菌番茄致病变体的番茄pto基因编码蛋白激酶]);mindrinos等人,(1994)cell[细胞]78:1089(arabidopsisrsp2geneforresistancetopseudomonassyringae[用于抵抗丁香假单胞菌的拟南芥属rsp2基因]);mcdowell和woffenden,(2003)trendsbiotechnol.[生物技术趋势]21(4):178-83以及toyoda等人,(2002)transgenicres.[转基因研究]11(6):567-82。与野生型植物相比,对疾病具有抗性的植物对病原体更具抗性。(b)编码苏云金芽孢杆菌(bacillusthuringiensis)蛋白质、其衍生物或其上建模的合成多肽的基因。参见,例如,geiser等人,(1986)gene[基因]48:109,其公开了btδ-内毒素基因的克隆和核苷酸序列。此外,编码δ-内毒素基因的dna分子可购自美国典型培养物保藏中心(americantypeculturecollection)(美国马里兰州罗克韦尔市(roekville,md.)),例如登录号40098、67136、31995和31998下。经遗传工程化的苏云金芽孢杆菌转基因的其他非限制性实例在以下专利和专利申请中给出,并且特此通过引用并入本文中:美国专利号5,188,960;5,689,052;5,880,275;5,986,177;6,023,013、6,060,594、6,063,597、6,077,824、6,620,988、6,642,030、6,713,259、6,893,826、7,105,332;7,179,965、7,208,474;7,227,056、7,288,643、7,323,556、7,329,736、7,449,552、7,468,278、7,510,878、7,521,235、7,544,862、7,605,304、7,696,412、7,629,504、7,705,216、7,772,465、7,790,846、7,858,849和wo1991/14778;wo1999/31248;wo2001/12731;wo1999/24581和wo1997/40162。也可以堆叠编码杀有害生物蛋白的基因,所述蛋白包括但不限于:来自假单胞菌属物种(pseudomonassp.)的杀昆虫蛋白,例如pseen3174(monalysin,(2011)plospathogens[plos病原体],7:1-13),来自假单胞菌蛋白菌(pseudomonasprotegens)菌株cha0和pf-5(之前为荧光假单胞菌(fluorescens))(pechy-tarr,(2008)environmentalmicrobiology[环境微生物学]10:2368-2386:基因库登录号eu400157);来自台湾假单胞菌(pseudomonastaiwanensis)(liu等人,(2010)j.agric.foodchem.[农业食品化学学报]58:12343-12349)和来自假产碱假单胞菌(pseudomonaspseudoalcaligenes)(zhang等人,(2009)annalsofmicrobiology[微生物学年报]59:45-50和li等人,(2007)plantcelltiss.organcult.[植物细胞组织和器官培养]89:159-168);来自发光杆菌属物种(photorhabdussp.)和致病杆菌属物种(xenorhabdussp.)的杀昆虫蛋白(hinchliffe等人,(2010)theopentoxinologyjournal[开放性毒理学杂志]3:101-118和morgan等人,(2001)appliedandenvir.micro.[应用与环境微生物学]67:2062-2069),美国专利号6,048,838和美国专利号6,379,946;美国序列号13792861的pip-1多肽;美国序列号13/800233的afip-1a和/或afip-1b多肽;美国序列号13/839702的phi-4多肽;pct序列号pct/us14/51063的pip-47多肽;pct序列号pct/us14/55128的pip-72多肽;pct公开号wo2015/120270的ptip-50多肽和ptip-65多肽;pct公开号wo2015/120276的ptip-83多肽;pct序列号pct/us15/55502的ptip-96多肽;以及δ-内毒素,包括但不限于cry1、cry2、cry3、cry4、cry5、cry6、cry7、cry8、cry9、cry10、cry11、cry12、cry13、cry14、cry15、cry16、cry17、cry18、cry19、cry20、cry21、cry22、cry23、cry24、cry25、cry26、cry27、cry28、cry29、cry30、cry31、cry32、cry33、cry34、cry35、cry36、cry37、cry38、cry39、cry40、cry41、cry42、cry43、cry44、cry45、cry46、cry47、cry49、cry51和cry55类的δ-内毒素基因和苏云金芽孢杆菌溶细胞cyt1和cyt2基因。苏云金芽孢杆菌杀昆虫蛋白的这些类别的成员包括但不限于cry1aa1(登录号aaa22353);cry1aa2(登录号aaa22552);cry1aa3(登录号baa00257);cry1aa4(登录号caa31886);cry1aa5(登录号baa04468);cry1aa6(登录号aaa86265);cry1aa7(登录号aad46139);cry1aa8(登录号i26149);cry1aa9(登录号baa77213);cry1aa10(登录号aad55382);cry1aa11(登录号caa70856);cry1aa12(登录号aap80146);cry1aa13(登录号aam44305);cry1aa14(登录号aap40639);cry1aa15(登录号aay66993);cry1aa16(登录号hq439776);cry1aa17(登录号hq439788);cry1aa18(登录号hq439790);cry1aa19(登录号hq685121);cry1aa20(登录号jf340156);cry1aa21(登录号jn651496);cry1aa22(登录号kc158223);cry1ab1(登录号aaa22330);cry1ab2(登录号aaa22613);cry1ab3(登录号aaa22561);cry1ab4(登录号baa00071);cry1ab5(登录号caa28405);cry1ab6(登录号aaa22420);cry1ab7(登录号caa31620);cry1ab8(登录号aaa22551);cry1ab9(登录号caa38701);cry1ab10(登录号a29125);cry1ab11(登录号i12419);cry1ab12(登录号aac64003);cry1ab13(登录号aan76494);cry1ab14(登录号aag16877);cry1ab15(登录号aao13302);cry1ab16(登录号aak55546);cry1ab17(登录号aat46415);cry1ab18(登录号aaq88259);cry1ab19(登录号aaw31761);cry1ab20(登录号abb72460);cry1ab21(登录号abs18384);cry1ab22(登录号abw87320);cry1ab23(登录号hq439777);cry1ab24(登录号hq439778);cry1ab25(登录号hq685122);cry1ab26(登录号hq847729);cry1ab27(登录号jn135249);cry1ab28(登录号jn135250);cry1ab29(登录号jn135251);cry1ab30(登录号jn135252);cry1ab31(登录号jn135253);cry1ab32(登录号jn135254);cry1ab33(登录号aas93798);cry1ab34(登录号kc156668);cry1ab样(登录号aak14336);cry1ab样(登录号aak14337);cry1ab样(登录号aak14338);cry1ab样(登录号abg88858);cry1ac1(登录号aaa22331);cry1ac2(登录号aaa22338);cry1ac3(登录号caa38098);cry1ac4(登录号aaa73077);cry1ac5(登录号aaa22339);cry1ac6(登录号aaa86266);cry1ac7(登录号aab46989);cry1ac8(登录号aac44841);cry1ac9(登录号aab49768);cry1ac10(登录号caa05505);cry1ac11(登录号caa10270);cry1ac12(登录号i12418);cry1ac13(登录号aad38701);cry1ac14(登录号aaq06607);cry1ac15(登录号aan07788);cry1ac16(登录号aau87037);cry1ac17(登录号aax18704);cry1ac18(登录号aay88347);cry1ac19(登录号abd37053);cry1ac20(登录号abb89046);cry1ac21(登录号aay66992);cry1ac22(登录号abz01836);cry1ac23(登录号caq30431);cry1ac24(登录号abl01535);cry1ac25(登录号fj513324);cry1ac26(登录号fj617446);cry1ac27(登录号fj617447);cry1ac28(登录号acm90319);cry1ac29(登录号dq438941);cry1ac30(登录号gq227507);cry1ac31(登录号gu446674);cry1ac32(登录号hm061081);cry1ac33(登录号gq866913);cry1ac34(登录号hq230364);cry1ac35(登录号jf340157);cry1ac36(登录号jn387137);cry1ac37(登录号jq317685);cry1ad1(登录号aaa22340);cry1ad2(登录号caa01880);cry1ae1(登录号aaa22410);cry1af1(登录号aab82749);cry1ag1(登录号aad46137);cry1ah1(登录号aaq14326);cry1ah2(登录号abb76664);cry1ah3(登录号hq439779);cry1ai1(登录号aao39719);cry1ai2(登录号hq439780);cry1a样(登录号aak14339);cry1ba1(登录号caa29898);cry1ba2(登录号caa65003);cry1ba3(登录号aak63251);cry1ba4(登录号aak51084);cry1ba5(登录号abo20894);cry1ba6(登录号abl60921);cry1ba7(登录号hq439781);cry1bb1(登录号aaa22344);cry1bb2(登录号hq439782);cry1bc1(登录号caa86568);cry1bd1(登录号aad10292);cry1bd2(登录号aam93496);cry1be1(登录号aac32850);cry1be2(登录号aaq52387);cry1be3(登录号acv96720);cry1be4(登录号hm070026);cry1bf1(登录号cac50778);cry1bf2(登录号aaq52380);cry1bg1(登录号aao39720);cry1bh1(登录号hq589331);cry1bi1(登录号kc156700);cry1ca1(登录号caa30396);cry1ca2(登录号caa31951);cry1ca3(登录号aaa22343);cry1ca4(登录号caa01886);cry1ca5(登录号caa65457);cry1ca6[1](登录号aaf37224);cry1ca7(登录号aag50438);cry1ca8(登录号aam00264);cry1ca9(登录号aal79362);cry1ca10(登录号aan16462);cry1ca11(登录号aax53094);cry1ca12(登录号hm070027);cry1ca13(登录号hq412621);cry1ca14(登录号jn651493);cry1cb1(登录号m97880);cry1cb2(登录号aag35409);cry1cb3(登录号acd50894);cry1cb样(登录号aax63901);cry1da1(登录号caa38099);cry1da2(登录号i76415);cry1da3(登录号hq439784);cry1db1(登录号caa80234);cry1db2(登录号aak48937);cry1dc1(登录号abk35074);cry1ea1(登录号caa37933);cry1ea2(登录号caa39609);cry1ea3(登录号aaa22345);cry1ea4(登录号aad04732);cry1ea5(登录号a15535);cry1ea6(登录号aal50330);cry1ea7(登录号aaw72936);cry1ea8(登录号abx11258);cry1ea9(登录号hq439785);cry1ea10(登录号adr00398);cry1ea11(登录号jq652456);cry1eb1(登录号aaa22346);cry1fa1(登录号aaa22348);cry1fa2(登录号aaa22347);cry1fa3(登录号hm070028);cry1fa4(登录号hm439638);cry1fb1(登录号caa80235);cry1fb2(登录号baa25298);cry1fb3(登录号aaf21767);cry1fb4(登录号aac10641);cry1fb5(登录号aao13295);cry1fb6(登录号acd50892);cry1fb7(登录号acd50893);cry1ga1(登录号caa80233);cry1ga2(登录号caa70506);cry1gb1(登录号aad10291);cry1gb2(登录号aao13756);cry1gc1(登录号aaq52381);cry1ha1(登录号caa80236);cry1hb1(登录号aaa79694);cry1hb2(登录号hq439786);cry1h样(登录号aaf01213);cry1ia1(登录号caa44633);cry1ia2(登录号aaa22354);cry1ia3(登录号aac36999);cry1ia4(登录号aab00958);cry1ia5(登录号caa70124);cry1ia6(登录号aac26910);cry1ia7(登录号aam73516);cry1ia8(登录号aak66742);cry1ia9(登录号aaq08616);cry1ia10(登录号aap86782);cry1ia11(登录号cac85964);cry1ia12(登录号aav53390);cry1ia13(登录号abf83202);cry1ia14(登录号acg63871);cry1ia15(登录号fj617445);cry1ia16(登录号fj617448);cry1ia17(登录号gu989199);cry1ia18(登录号adk23801);cry1ia19(登录号hq439787);cry1ia20(登录号jq228426);cry1ia21(登录号jq228424);cry1ia22(登录号jq228427);cry1ia23(登录号jq228428);cry1ia24(登录号jq228429);cry1ia25(登录号jq228430);cry1ia26(登录号jq228431);cry1ia27(登录号jq228432);cry1ia28(登录号jq228433);cry1ia29(登录号jq228434);cry1ia30(登录号jq317686);cry1ia31(登录号jx944038);cry1ia32(登录号jx944039);cry1ia33(登录号jx944040);cry1ib1(登录号aaa82114);cry1ib2(登录号abw88019);cry1ib3(登录号acd75515);cry1ib4(登录号hm051227);cry1ib5(登录号hm070028);cry1ib6(登录号adk38579);cry1ib7(登录号jn571740);cry1ib8(登录号jn675714);cry1ib9(登录号jn675715);cry1ib10(登录号jn675716);cry1ib11(登录号jq228423);cry1ic1(登录号aac62933);cry1ic2(登录号aae71691);cry1id1(登录号aad44366);cry1id2(登录号jq228422);cry1ie1(登录号aag43526);cry1ie2(登录号hm439636);cry1ie3(登录号kc156647);cry1ie4(登录号kc156681);cry1if1(登录号aaq52382);cry1ig1(登录号kc156701);cry1i样(登录号aac31094);cry1i样(登录号abg88859);cry1ja1(登录号aaa22341);cry1ja2(登录号hm070030);cry1ja3(登录号jq228425);cry1jb1(登录号aaa98959);cry1jc1(登录号aac31092);cry1jc2(登录号aaq52372);cry1jd1(登录号cac50779);cry1ka1(登录号aab00376);cry1ka2(登录号hq439783);cry1la1(登录号aas60191);cry1la2(登录号hm070031);cry1ma1(登录号fj884067);cry1ma2(登录号kc156659);cry1na1(登录号kc156648);cry1nb1(登录号kc156678);cry1样(登录号aac31091);cry2aa1(登录号aaa22335);cry2aa2(登录号aaa83516);cry2aa3(登录号d86064);cry2aa4(登录号aac04867);cry2aa5(登录号caa10671);cry2aa6(登录号caa10672);cry2aa7(登录号caa10670);cry2aa8(登录号aao13734);cry2aa9(登录号aao13750);cry2aa10(登录号aaq04263);cry2aa11(登录号aaq52384);cry2aa12(登录号abi83671);cry2aa13(登录号abl01536);cry2aa14(登录号acf04939);cry2aa15(登录号jn426947);cry2ab1(登录号aaa22342);cry2ab2(登录号caa39075);cry2ab3(登录号aag36762);cry2ab4(登录号aao13296);cry2ab5(登录号aaq04609);cry2ab6(登录号aap59457);cry2ab7(登录号aaz66347);cry2ab8(登录号abc95996);cry2ab9(登录号abc74968);cry2ab10(登录号ef157306);cry2ab11(登录号cam84575);cry2ab12(登录号abm21764);cry2ab13(登录号acg76120);cry2ab14(登录号acg76121);cry2ab15(登录号hm037126);cry2ab16(登录号gq866914);cry2ab17(登录号hq439789);cry2ab18(登录号jn135255);cry2ab19(登录号jn135256);cry2ab20(登录号jn135257);cry2ab21(登录号jn135258);cry2ab22(登录号jn135259);cry2ab23(登录号jn135260);cry2ab24(登录号jn135261);cry2ab25(登录号jn415485);cry2ab26(登录号jn426946);cry2ab27(登录号jn415764);cry2ab28(登录号jn651494);cry2ac1(登录号caa40536);cry2ac2(登录号aag35410);cry2ac3(登录号aaq52385);cry2ac4(登录号abc95997);cry2ac5(登录号abc74969);cry2ac6(登录号abc74793);cry2ac7(登录号cal18690);cry2ac8(登录号cam09325);cry2ac9(登录号cam09326);cry2ac10(登录号abn15104);cry2ac11(登录号cam83895);cry2ac12(登录号cam83896);cry2ad1(登录号aaf09583);cry2ad2(登录号abc86927);cry2ad3(登录号cak29504);cry2ad4(登录号cam32331);cry2ad5(登录号cao78739);cry2ae1(登录号aaq52362);cry2af1(登录号abo30519);cry2af2(登录号gq866915);cry2ag1(登录号ach91610);cry2ah1(登录号eu939453);cry2ah2(登录号acl80665);cry2ah3(登录号gu073380);cry2ah4(登录号kc156702);cry2ai1(登录号fj788388);cry2aj(登录号);cry2ak1(登录号kc156660);cry2ba1(登录号kc156658);cry3aa1(登录号aaa22336);cry3aa2(登录号aaa22541);cry3aa3(登录号caa68482);cry3aa4(登录号aaa22542);cry3aa5(登录号aaa50255);cry3aa6(登录号aac43266);cry3aa7(登录号cab41411);cry3aa8(登录号aas79487);cry3aa9(登录号aaw05659);cry3aa10(登录号aau29411);cry3aa11(登录号aaw82872);cry3aa12(登录号aby49136);cry3ba1(登录号caa34983);cry3ba2(登录号caa00645);cry3ba3(登录号jq397327);cry3bb1(登录号aaa22334);cry3bb2(登录号aaa74198);cry3bb3(登录号i15475);cry3ca1(登录号caa42469);cry4aa1(登录号caa68485);cry4aa2(登录号baa00179);cry4aa3(登录号cad30148);cry4aa4(登录号afb18317);cry4a样(登录号aay96321);cry4ba1(登录号caa30312);cry4ba2(登录号caa30114);cry4ba3(登录号aaa22337);cry4ba4(登录号baa00178);cry4ba5(登录号cad30095);cry4ba样(登录号abc47686);cry4ca1(登录号eu646202);cry4cb1(登录号fj403208);cry4cb2(登录号fj597622);cry4cc1(登录号fj403207);cry5aa1(登录号aaa67694);cry5ab1(登录号aaa67693);cry5ae1(登录号i34543);cry5ad1(登录号abq82087);cry5ba1(登录号aaa68598);cry5ba2(登录号abw88931);cry5ba3(登录号afj04417);cry5ca1(登录号hm461869);cry5ca2(登录号zp_04123426);cry5da1(登录号hm461870);cry5da2(登录号zp_04123980);cry5ea1(登录号hm485580);cry5ea2(登录号zp_04124038);cry6aa1(登录号aaa22357);cry6aa2(登录号aam46849);cry6aa3(登录号abh03377);cry6ba1(登录号aaa22358);cry7aa1(登录号aaa22351);cry7ab1(登录号aaa21120);cry7ab2(登录号aaa21121);cry7ab3(登录号abx24522);cry7ab4(登录号eu380678);cry7ab5(登录号abx79555);cry7ab6(登录号aci44005);cry7ab7(登录号adb89216);cry7ab8(登录号gu145299);cry7ab9(登录号add92572);cry7ba1(登录号abb70817);cry7bb1(登录号kc156653);cry7ca1(登录号abr67863);cry7cb1(登录号kc156698);cry7da1(登录号acq99547);cry7da2(登录号hm572236);cry7da3(登录号kc156679);cry7ea1(登录号hm035086);cry7ea2(登录号hm132124);cry7ea3(登录号eem19403);cry7fa1(登录号hm035088);cry7fa2(登录号eem19090);cry7fb1(登录号hm572235);cry7fb2(登录号kc156682);cry7ga1(登录号hm572237);cry7ga2(登录号kc156669);cry7gb1(登录号kc156650);cry7gc1(登录号kc156654);cry7gd1(登录号kc156697);cry7ha1(登录号kc156651);cry7ia1(登录号kc156665);cry7ja1(登录号kc156671);cry7ka1(登录号kc156680);cry7kb1(登录号bam99306);cry7la1(登录号bam99307);cry8aa1(登录号aaa21117);cry8ab1(登录号eu044830);cry8ac1(登录号kc156662);cry8ad1(登录号kc156684);cry8ba1(登录号aaa21118);cry8bb1(登录号cad57542);cry8bc1(登录号cad57543);cry8ca1(登录号aaa21119);cry8ca2(登录号aar98783);cry8ca3(登录号eu625349);cry8ca4(登录号adb54826);cry8da1(登录号bac07226);cry8da2(登录号bd133574);cry8da3(登录号bd133575);cry8db1(登录号baf93483);cry8ea1(登录号aaq73470);cry8ea2(登录号eu047597);cry8ea3(登录号kc855216);cry8fa1(登录号aat48690);cry8fa2(登录号hq174208);cry8fa3(登录号afh78109);cry8ga1(登录号aat46073);cry8ga2(登录号abc42043);cry8ga3(登录号fj198072);cry8ha1(登录号aaw81032);cry8ia1(登录号eu381044);cry8ia2(登录号gu073381);cry8ia3(登录号hm044664);cry8ia4(登录号kc156674);cry8ib1(登录号gu325772);cry8ib2(登录号kc156677);cry8ja1(登录号eu625348);cry8ka1(登录号fj422558);cry8ka2(登录号acn87262);cry8kb1(登录号hm123758);cry8kb2(登录号kc156675);cry8la1(登录号gu325771);cry8ma1(登录号hm044665);cry8ma2(登录号eem86551);cry8ma3(登录号hm210574);cry8na1(登录号hm640939);cry8pa1(登录号hq388415);cry8qa1(登录号hq441166);cry8qa2(登录号kc152468);cry8ra1(登录号afp87548);cry8sa1(登录号jq740599);cry8ta1(登录号kc156673);cry8样(登录号fj770571);cry8样(登录号abs53003);cry9aa1(登录号caa41122);cry9aa2(登录号caa41425);cry9aa3(登录号gq249293);cry9aa4(登录号gq249294);cry9aa5(登录号jx174110);cry9aa样(登录号aaq52376);cry9ba1(登录号caa52927);cry9ba2(登录号gu299522);cry9bb1(登录号aav28716);cry9ca1(登录号caa85764);cry9ca2(登录号aaq52375);cry9da1(登录号baa19948);cry9da2(登录号aab97923);cry9da3(登录号gq249293);cry9da4(登录号gq249297);cry9db1(登录号aax78439);cry9dc1(登录号kc156683);cry9ea1(登录号baa34908);cry9ea2(登录号aao12908);cry9ea3(登录号abm21765);cry9ea4(登录号ace88267);cry9ea5(登录号acf04743);cry9ea6(登录号acg63872);cry9ea7(登录号fj380927);cry9ea8(登录号gq249292);cry9ea9(登录号jn651495);cry9eb1(登录号cac50780);cry9eb2(登录号gq249298);cry9eb3(登录号kc156646);cry9ec1(登录号aac63366);cry9ed1(登录号aax78440);cry9ee1(登录号gq249296);cry9ee2(登录号kc156664);cry9fa1(登录号kc156692);cry9ga1(登录号kc156699);cry9样(登录号aac63366);cry10aa1(登录号aaa22614);cry10aa2(登录号e00614);cry10aa3(登录号cad30098);cry10aa4(登录号afb18318);cry10a样(登录号dq167578);cry11aa1(登录号aaa22352);cry11aa2(登录号aaa22611);cry11aa3(登录号cad30081);cry11aa4(登录号afb18319);cry11aa样(登录号dq166531);cry11ba1(登录号caa60504);cry11bb1(登录号aac97162);cry11bb2(登录号hm068615);cry12aa1(登录号aaa22355);cry13aa1(登录号aaa22356);cry14aa1(登录号aaa21516);cry14ab1(登录号kc156652);cry15aa1(登录号aaa22333);cry16aa1(登录号caa63860);cry17aa1(登录号caa67841);cry18aa1(登录号caa67506);cry18ba1(登录号aaf89667);cry18ca1(登录号aaf89668);cry19aa1(登录号caa68875);cry19ba1(登录号baa32397);cry19ca1(登录号afm37572);cry20aa1(登录号aab93476);cry20ba1(登录号acs93601);cry20ba2(登录号kc156694);cry20样(登录号gq144333);cry21aa1(登录号i32932);cry21aa2(登录号i66477);cry21ba1(登录号bac06484);cry21ca1(登录号jf521577);cry21ca2(登录号kc156687);cry21da1(登录号jf521578);cry22aa1(登录号i34547);cry22aa2(登录号cad43579);cry22aa3(登录号acd93211);cry22ab1(登录号aak50456);cry22ab2(登录号cad43577);cry22ba1(登录号cad43578);cry22bb1(登录号kc156672);cry23aa1(登录号aaf76375);cry24aa1(登录号aac61891);cry24ba1(登录号bad32657);cry24ca1(登录号caj43600);cry25aa1(登录号aac61892);cry26aa1(登录号aad25075);cry27aa1(登录号baa82796);cry28aa1(登录号aad24189);cry28aa2(登录号aag00235);cry29aa1(登录号cac80985);cry30aa1(登录号cac80986);cry30ba1(登录号bad00052);cry30ca1(登录号bad67157);cry30ca2(登录号acu24781);cry30da1(登录号ef095955);cry30db1(登录号bae80088);cry30ea1(登录号acc95445);cry30ea2(登录号fj499389);cry30fa1(登录号aci22625);cry30ga1(登录号acg60020);cry30ga2(登录号hq638217);cry31aa1(登录号bab11757);cry31aa2(登录号aal87458);cry31aa3(登录号bae79808);cry31aa4(登录号baf32571);cry31aa5(登录号baf32572);cry31aa6(登录号bai44026);cry31ab1(登录号bae79809);cry31ab2(登录号baf32570);cry31ac1(登录号baf34368);cry31ac2(登录号ab731600);cry31ad1(登录号bai44022);cry32aa1(登录号aag36711);cry32aa2(登录号gu063849);cry32ab1(登录号gu063850);cry32ba1(登录号bab78601);cry32ca1(登录号bab78602);cry32cb1(登录号kc156708);cry32da1(登录号bab78603);cry32ea1(登录号gu324274);cry32ea2(登录号kc156686);cry32eb1(登录号kc156663);cry32fa1(登录号kc156656);cry32ga1(登录号kc156657);cry32ha1(登录号kc156661);cry32hb1(登录号kc156666);cry32ia1(登录号kc156667);cry32ja1(登录号kc156685);cry32ka1(登录号kc156688);cry32la1(登录号kc156689);cry32ma1(登录号kc156690);cry32mb1(登录号kc156704);cry32na1(登录号kc156691);cry32oa1(登录号kc156703);cry32pa1(登录号kc156705);cry32qa1(登录号kc156706);cry32ra1(登录号kc156707);cry32sa1(登录号kc156709);cry32ta1(登录号kc156710);cry32ua1(登录号kc156655);cry33aa1(登录号aal26871);cry34aa1(登录号aag50341);cry34aa2(登录号aak64560);cry34aa3(登录号aat29032);cry34aa4(登录号aat29030);cry34ab1(登录号aag41671);cry34ac1(登录号aag50118);cry34ac2(登录号aak64562);cry34ac3(登录号aat29029);cry34ba1(登录号aak64565);cry34ba2(登录号aat29033);cry34ba3(登录号aat29031);cry35aa1(登录号aag50342);cry35aa2(登录号aak64561);cry35aa3(登录号aat29028);cry35aa4(登录号aat29025);cry35ab1(登录号aag41672);cry35ab2(登录号aak64563);cry35ab3(登录号ay536891);cry35ac1(登录号aag50117);cry35ba1(登录号aak64566);cry35ba2(登录号aat29027);cry35ba3(登录号aat29026);cry36aa1(登录号aak64558);cry37aa1(登录号aaf76376);cry38aa1(登录号aak64559);cry39aa1(登录号bab72016);cry40aa1(登录号bab72018);cry40ba1(登录号bac77648);cry40ca1(登录号eu381045);cry40da1(登录号acf15199);cry41aa1(登录号bad35157);cry41ab1(登录号bad35163);cry41ba1(登录号hm461871);cry41ba2(登录号zp_04099652);cry42aa1(登录号bad35166);cry43aa1(登录号bad15301);cry43aa2(登录号bad95474);cry43ba1(登录号bad15303);cry43ca1(登录号kc156676);cry43cb1(登录号kc156695);cry43cc1(登录号kc156696);cry43样(登录号bad15305);cry44aa(登录号bad08532);cry45aa(登录号bad22577);cry46aa(登录号bac79010);cry46aa2(登录号bag68906);cry46ab(登录号bad35170);cry47aa(登录号aay24695);cry48aa(登录号caj18351);cry48aa2(登录号caj86545);cry48aa3(登录号caj86546);cry48ab(登录号caj86548);cry48ab2(登录号caj86549);cry49aa(登录号cah56541);cry49aa2(登录号caj86541);cry49aa3(登录号caj86543);cry49aa4(登录号caj86544);cry49ab1(登录号caj86542);cry50aa1(登录号bae86999);cry50ba1(登录号gu446675);cry50ba2(登录号gu446676);cry51aa1(登录号abi14444);cry51aa2(登录号gu570697);cry52aa1(登录号ef613489);cry52ba1(登录号fj361760);cry53aa1(登录号ef633476);cry53ab1(登录号fj361759);cry54aa1(登录号aca52194);cry54aa2(登录号gq140349);cry54ba1(登录号gu446677);cry55aa1(登录号abw88932);cry54ab1(登录号jq916908);cry55aa2(登录号aae33526);cry56aa1(登录号acu57499);cry56aa2(登录号gq483512);cry56aa3(登录号jx025567);cry57aa1(登录号anc87261);cry58aa1(登录号anc87260);cry59ba1(登录号jn790647);cry59aa1(登录号acr43758);cry60aa1(登录号acu24782);cry60aa2(登录号eao57254);cry60aa3(登录号eem99278);cry60ba1(登录号gu810818);cry60ba2(登录号eao57253);cry60ba3(登录号eem99279);cry61aa1(登录号hm035087);cry61aa2(登录号hm132125);cry61aa3(登录号eem19308);cry62aa1(登录号hm054509);cry63aa1(登录号bai44028);cry64aa1(登录号baj05397);cry65aa1(登录号hm461868);cry65aa2(登录号zp_04123838);cry66aa1(登录号hm485581);cry66aa2(登录号zp_04099945);cry67aa1(登录号hm485582);cry67aa2(登录号zp_04148882);cry68aa1(登录号hq113114);cry69aa1(登录号hq401006);cry69aa2(登录号jq821388);cry69ab1(登录号jn209957);cry70aa1(登录号jn646781);cry70ba1(登录号ado51070);cry70bb1(登录号eel67276);cry71aa1(登录号jx025568);cry72aa1(登录号jx025569)。δ-内毒素的实例还包括但不限于美国专利号5,880,275和7,858,849的cry1a蛋白;美国专利号8,304,604和8.304,605的dig-3或dig-11毒素(cry蛋白(如cry1a)的α螺旋1和/或α螺旋2变体的n末端缺失),美国专利申请序列号10/525,318的cry1b;美国专利号6,033,874的cry1c;美国专利号5,188,960、6,218,188的cry1f;美国专利号7,070,982、6,962,705和6,713,063的cry1a/f嵌合体;美国专利号7,064,249的cry2蛋白如cry2ab蛋白;cry3a蛋白,包括但不限于通过融合至少两种不同cry蛋白的可变区和保守区的独特组合产生的工程化杂合杀昆虫蛋白(ehip)(美国专利申请公开号2010/0017914);cry4蛋白;cry5蛋白;cry6蛋白;美国专利号7,329,736、7,449,552、7,803,943、7,476,781、7,105,332、7,378,499和7,462,760的cry8蛋白;cry9蛋白,如cry9a、cry9b、cry9c、cry9d、cry9e、和cry9f家族的成员;cry15蛋白,描述于naimov等人,(2008)appliedandenvironmentalmicrobiology[应用与环境微生物学]74:7145-7151中;美国专利号6,127,180、6,624,145和6,340,593的cry22、cry34ab1蛋白;美国专利号6,248,535、6,326,351、6,399,330、6,949,626、7,385,107和7,504,229的cryet33和cryet34蛋白;美国专利公开号2006/0191034、2012/0278954,和pct公开号wo2012/139004的cryet33和cryet34同源物;美国专利号6,083,499、6,548,291和6,340,593的cry35ab1蛋白;cry46蛋白、cry51蛋白、cry二元毒素;tic901或相关毒素;us2008/0295207的tic807;pctus2006/033867的et29、et37、tic809、tic810、tic812、tic127、tic128;美国专利申请公开号2015/0047076的tic3131、tic3400、和tic3407;美国专利号8,236,757的axmi-027、axmi-036和axmi-038;us7,923,602的axmi-031、axmi-039、axmi-040、axmi-049;wo2006/083891的axmi-018、axmi-020和axmi-021;wo2005/038032的axmi-010;wo2005/021585的axmi-003;us2004/0250311的axmi-008;us2004/0216186的axmi-006;us2004/0210965的axmi-007;us2004/0210964的axmi-009;us2004/0197917的axmi-014;us2004/0197916的axmi-004;wo2006/119457的axmi-028和axmi-029;wo2004/074462的axmi-007、axmi-008、axmi-0080rf2、axmi-009、axmi-014和axmi-004;美国专利号8,084,416的axmi-150;us20110023184的axmi-205;us2011/0263488的axmi-011、axmi-012、axmi-013、axmi-015、axmi-019、axmi-044、axmi-037、axmi-043、axmi-033、axmi-034、axmi-022、axmi-023、axmi-041、axmi-063、和axmi-064;us2010/0197592的axmi-r1和相关蛋白;wo2011/103248的axmi221z、axmi222z、axmi223z、axmi224z和axmi225z;wo11/103247的axmi218、axmi219、axmi220、axmi226、axmi227、axmi228、axmi229、axmi230、和axmi231;美国专利号8,334,431的axmi-115、axmi-113、axmi-005、axmi-163和axmi-184;us2010/0298211的axmi-001、axmi-002、axmi-030、axmi-035、和axmi-045;us20090144852的axmi-066和axmi-076;美国专利号8,318,900的axmi128、axmi130、axmi131、axmi133、axmi140、axmi141、axmi142、axmi143、axmi144、axmi146、axmi148、axmi149、axmi152、axmi153、axmi154、axmi155、axmi156、axmi157、axmi158、axmi162、axmi165、axmi166、axmi167、axmi168、axmi169、axmi170、axmi171、axmi172、axmi173、axmi174、axmi175、axmi176、axmi177、axmi178、axmi179、axmi180、axmi181、axmi182、axmi185、axmi186、axmi187、axmi188、axmi189;us2010/0005543的axmi079、axmi080、axmi081、axmi082、axmi091、axmi092、axmi096、axmi097、axmi098、axmi099、axmi100、axmi101、axmi102、axmi103、axmi104、axmi107、axmi108、axmi109、axmi110、axmi111、axmi112、axmi114、axmi116、axmi117、axmi118、axmi119、axmi120、axmi121、axmi122、axmi123、axmi124、axmi1257、axmi1268、axmi127、axmi129、axmi164、axmi151、axmi161、axmi183、axmi132、axmi138、axmi137;和美国专利号8,319,019的具有修饰的蛋白水解位点的cry蛋白如cry1a和cry3a;以及美国专利申请公开号2011/0064710的来自苏云金芽孢杆菌菌株vbts2528的cry1ac、cry2aa和cry1ca毒素蛋白。其他cry蛋白是本领域技术人员熟知的(参见crickmore等人,“bacillusthuringiensistoxinnomenclature[苏云金芽孢杆菌毒素命名法]”(2011),网址为lifesci.sussex.ac.uk/home/neil_crickmore/bt/,可以使用“www”前缀在万维网上访问)。cry蛋白的杀昆虫活性是本领域技术人员所熟知的(综述参见vanfrannkenhuyzen,(2009)j.invert.path.[无脊椎动物病理学杂志]101:1-16)。使用cry蛋白作为转基因植物性状是本领域技术人员所熟知的,并且cry转基因植物(包括但不限于cry1ac、cry1ac+cry2ab、cry1ab、cry1a.105、cry1f、cry1fa2、cry1f+cry1ac、cry2ab、cry3a、mcry3a、cry3bb1、cry34ab1、cry35ab1、vip3a、mcry3a、cry9c和cbi-bt)已获得监管部门的批准(参见,sanahuja,(2011)plantbiotechjournal[植物生物技术杂志]9:283-300和cera(2010)转基因作物数据库环境风险评估中心(cera)(gmcropdatabasecenterforenvironmentalriskassessment),ilsi研究基金会,华盛顿特区,网址为cera-gmc.org/index.php?action=gm_crop_database,可以使用“www”前缀在万维网上访问)。本领域技术人员熟知的多种杀有害生物蛋白也可以在植物中表达,如vip3ab&cry1fa(us2012/0317682)、cry1be&cry1f(us2012/0311746)、cry1ca&cry1ab(us2012/0311745)、cry1f&cryca(us2012/0317681)、cry1da&cry1be(us2012/0331590)、cry1da&cry1fa(us2012/0331589)、cry1ab&cry1be(us2012/0324606)以及cry1fa&cry2aa、cry1i或cry1e(us2012/0324605));cry34ab/35ab以及cry6aa(us20130167269);cry34ab/vcry35ab和cry3aa(us20130167268);cry3a和cry1ab或vip3aa(us20130116170);以及cry1f、cry34ab1和cry35ab1(pct/us2010/060818)。杀有害生物蛋白还包括杀昆虫脂肪酶,这些杀昆虫脂肪酶包括美国专利号7,491,869的脂质酰基水解酶,和胆固醇氧化酶,如来自链霉菌属(purcell等人(1993)biochembiophysrescommun[生物化学与生物物理学研究通讯]15:1406-1413)。杀有害生物蛋白还包括美国专利号5,877,012、6,107,279、6,137,033、7,244,820、7,615,686和8,237,020中的vip(营养性杀昆虫蛋白)毒素等。其他vip蛋白质是本领域技术人员熟知的(参见,lifesci.sussex.ac.uk/home/neil_crickmore/bt/vip.html,其可以使用“www”前缀在万维网上访问)。杀有害生物蛋白还包括可从如下生物体获得的毒素复合物(tc)蛋白质:致病杆菌属、发光杆菌属和类芽孢杆菌属(paenibacillus)(参见美国专利号7,491,698和8,084,418)。一些tc蛋白具有“独立”杀昆虫活性并且其他tc蛋白增强由相同给定生物体产生的独立毒素(stand-alonetoxin)的活性。可以通过源自不同属的来源生物体的一种或多种tc蛋白“增效剂”来增强“独立”tc蛋白(例如来自发光杆菌属、致病杆菌属或类芽孢杆菌属)的毒性。有三种主要类型的tc蛋白。如本文所提及的,a类蛋白(“蛋白a”)是独立毒素。b类蛋白(“蛋白b”)和c类蛋白(“蛋白c”)提高了a类蛋白的毒性。a类蛋白的实例是tcba、tcda、xpta1和xpta2。b类蛋白的实例是tcac、tcdb、xptb1xb和xptc1wi。c类蛋白的实例是tccc、xptc1xb和xptb1wi。杀有害生物蛋白还包括蜘蛛、蛇和蝎毒蛋白。蜘蛛肽的实例包括但不限于莱科毒素-1肽及其突变体(美国专利号8,334,366)。(c)昆虫特异性激素或信息素,例如蜕皮激素和保幼激素、其变体、基于其的模拟物、或其拮抗剂或激动剂。参见,例如hammock等人,(1990)nature[自然]344:458,该文献公开了经克隆的保幼激素酯酶(保幼激素的灭活剂)的杆状病毒表达。(d)昆虫特异性肽,其表达时破坏受影响害虫的生理学。例如,参见regan,(1994)j.biol.chem.[生物化学杂志]269:9的公开内容(表达克隆产生编码昆虫利尿激素受体的dna);pratt等人,(1989)biochem.biophys.res.comm.[生物化学与生物物理研究通讯]163:1243(在diplopterapuntata中识别出了阿洛斯德汀(allostatin));chattopadhyay等人,(2004)criticalreviewsinmicrobiology[微生物学评论]30(1):33-54;zjawiony,(2004)jnatprod[天然产物杂志]67(2):300-310;carlini和grossi-de-sa,(2002)toxicon[毒素]40(11):1515-1539;ussuf等人,(2001)currsci.[当代科学]80(7):847-853以及vasconcelos和oliveira,(2004)toxicon[毒素]44(4):385-403。还参见,tomalski等人的美国专利号5,266,317,他们公开了编码昆虫特异性毒素的基因。(e)引起单萜、倍半萜、类固醇、异羟肟酸、类苯基丙烷衍生物或具有杀昆虫活性的另一种非蛋白质分子超积累的酶。(f)参与生物活性分子的修饰(包括翻译后修饰)的酶;例如糖酵解酶、蛋白水解酶、脂肪分解酶、核酸酶、环化酶、转氨酶、酯酶、水解酶、磷酸酶、激酶、磷酸化酶、聚合酶、弹性蛋白酶、几丁质酶和葡聚糖酶,无论是天然还是合成的。参见,scott等人的pct申请号wo93/02197,该文献公开了愈创葡聚糖酶(callase)基因的核苷酸序列。含有几丁质酶编码序列的dna分子可以例如从atcc登录号39637和67152下获得。还参见,kramer等人,(1993)insectbiochem.molec.biol.[昆虫生物化学与分子生物学]23:691,他们教导了编码烟草钩虫几丁质酶的cdna的核苷酸序列;以及kawalleck等人,(1993)plantmolec.biol.[植物分子生物学]21:673,他们提供了欧芹ubi4-2聚泛素基因的核苷酸序列;美国专利申请序列号10/389,432、10/692,367以及美国专利号6,563,020。(g)刺激信号转导的分子。例如,参见botella等人,(1994)plantmolec.biol.[植物分子生物学]24:757,该文献公开了绿豆钙调素cdna克隆的核苷酸序列,以及griess等人,(1994)plantphysiol.[植物生理学]104:1467,他们提供了玉米钙调素cdna克隆的核苷酸序列。(h)疏水性瞬时肽。参见,pct申请号wo95/16776和美国专利号5,580,852(公开了抑制真菌植物病原体的鲎抗菌肽(tachyplesin)的肽衍生物)以及pct申请号wo95/18855和美国专利号5,607,914(教导了赋予抗病性的合成抗微生物肽)。(i)膜透性酶,一种通道形成剂或通道阻断剂。例如,参见jaynes等人,(1993)plantsci.[植物科学]89:43,该文献公开了天蚕素-β裂解肽类似物的异源表达,以提供对烟草假单胞菌(pseudomonassolanacearum)具有抗性的转基因烟草植物。(j)病毒侵入性蛋白或其衍生的复合毒素。例如,病毒外壳蛋白在转化的植物细胞中的积累赋予由外源蛋白基因来源的病毒以及由相关病毒导致的病毒侵染和/或疾病发展的抗性。参见,beachy等人,(1990)ann.rev.phytopathol.[植物病理学年评]28:451。外壳蛋白介导的抗性已赋予经转化的植物抵抗苜蓿花叶病毒、黄瓜花叶病毒、烟草线条病毒、马铃薯x病毒、马铃薯y病毒、烟草蚀纹病毒、烟草脆裂病毒和烟草花叶病毒。同上。(k)昆虫特异性抗体或其衍生的免疫毒素。因此,靶向昆虫肠道中关键代谢功能的抗体将使受影响的酶失活,杀灭昆虫。参看taylor等人,摘要#497,seventhint′lsymposiumonmolecularplant-microbeinteractions[关于分子植物-微生物相互作用的第七届国际研讨会](苏格兰爱丁堡,1994)(通过生产单链抗体片段进行转基因烟草中的酶失活)。(l)病毒特异性抗体。参见,例如tavladoraki等人,(1993)nature[自然]366:469,他们提出表达重组抗体基因的转基因植物不受病毒侵袭。(m)在自然界中由病原体或寄生虫产生的发育阻滞蛋白(developmental-arrestiveprotein)。因此,真菌内α-1,4-d-多聚半乳糖醛酸酶通过溶解植物细胞壁均-α-1,4-d-半乳糖醛酸酶来促进真菌定植和植物营养释放。参见lamb等人,(1992)bio/technology[生物技术]10:1436。以下文献描述了编码豆内聚半乳糖醛酸酶抑制蛋白的基因的克隆和表征:toubart等人,(1992)plantj.[植物杂志]2:367。(n)在自然界中由植物产生的发育阻滞蛋白。例如,已在以下文献中表明表达大麦核糖体失活基因的转基因植物具有增加的对真菌疾病的抗性:logemann等人,(1992)bio/technology[生物技术]10:305。(o)参与系统获得抗性(sar)应答的基因和/或发病机制相关基因。briggs,(1995)currentbiology[当代生物学]5(2):128-131,pieterse和vanloon,(2004)curr.opin.plantbio.[植物生物学新观点]7(4):456-64,以及somssich,(2003)cell[细胞]113(7):815-6。(p)抗真菌基因(cornelissen和melchers,(1993)pl.physiol.[植物生理学]101:709-712;和parijs等人,(1991)planta[植物]183:258-264;以及bushnell等人,(1998)can.j.ofplantpath.[加拿大植物病理学杂志]20(2):137-149)。还参见,美国专利申请号09/950,933。(q)解毒基因,如伏马菌素、白僵菌素、念珠菌素和玉米赤霉烯酮及其结构相关的衍生物的基因。例如,参见美国专利号5,792,931。(r)胱抑素和半胱氨酸蛋白酶抑制剂。参见,美国申请序列号10/947,979。(s)防御素基因。参见,wo03/000863和美国申请序列号10/178,213。(t)赋予对线虫的抗性的基因。参见wo03/033651,以及urwin等人,(1998)planta[植物]204:472-479,williamson(1999)curropinplantbio.[植物生物学新观点]2(4):327-31。(u)赋予对炭疽茎腐病(由真菌禾生炭疽菌引起)的抗性的基因,如rcg1。参见,jung等人,generation-meansanalysisandquantitativetraitlocusmappingofanthracnosestalkrotgenesinmaize[玉米中炭疽茎腐病基因的代平均值分析和数量性状基因座谱图],theor.appl.genet.[理论与应用遗传学](1994)89:413-418,以及美国临时专利申请号60/675,664。(v)核酸,所述核酸涉及通过干扰核糖核酸(rna)分子而下调昆虫有害生物物种中靶基因的表达,这些核糖核酸分子防治昆虫有害生物物种。pct公开wo2011/025860和wo2014/153254描述了抑制无脊椎动物植物有害生物(包括叶甲属(diabrotica)植物有害生物)中靶基因表达的方法。此外,pct公开wo2009/091864描述了用于抑制来自昆虫有害生物物种(包括来自草盲蝽属(lygus)的有害生物)的靶基因的组合物和方法。2.赋予对除草剂的抗性的转基因,例如:(a)抑制生长点或分生组织的除草剂,如咪唑啉酮或磺酰脲。这个类别的实例性基因编码突变体als和ahas酶,分别如以下文献中所述:lee等人,(1988)emboj.[欧洲分子生物学学会杂志]7:1241和miki等人,(1990)theor.appl.genet.[理论与应用遗传学]80:449。还参见,美国专利号5,605,011;5,013,659;5,141,870;5,767,361;5,731,180;5,304,732;4,761,373;5,331,107;5,928,937和5,378,824,以及国际公开wo96/33270。(b)草甘膦(分别由突变体5-烯醇丙酮酰-3-磷酸莽草酸合酶(epsp)和aroa基因赋予的抗性)和其他膦酰基化合物如草铵膦(草丁膦乙酰转移酶(pat)和吸水链霉菌草丁膦乙酰转移酶(bar)基因)、以及吡啶氧基或苯氧基丙酸和环己酮(acc酶抑制剂编码基因)。参见,例如shah等人的美国专利号4,940,835,其公开了epsps形式的能赋予草甘磷抗性的核苷酸序列。barry等人的美国专利号5,627,061也描述了编码epsps酶的基因。还参见,美国专利号6,566,587;6,338,961;6,248,876b1;6,040,497;5,804,425;5,633,435;5,145,783;4,971,908;5,312,910;5,188,642;4,940,835;5,866,775;6,225,114b1;6,130,366;5,310,667;4,535,060;4,769,061;5,633,448;5,510,471;re.36,449;re37,287e和5,491,288以及国际公开ep1173580;wo01/66704;ep1173581和ep1173582。还给予植物草甘磷抗性,使该植物表达编码草甘磷氧化还原酶的基因,这在美国专利号5,776,760和5,463,175中进行了更全面地描述,将这些文献通过引用以其全文并入本文。另外,可通过过量表达编码草甘磷n-乙酰转移酶的基因,来赋予植物草甘磷抗性。参见例如美国专利申请序列号11/405,845和10/427,692以及pct申请号us01/46227。编码突变aroa基因的dna分子可以在atcc登录号39256下获得,并且该突变基因的核苷酸序列公开于comai的美国专利号4,769,061中。欧洲专利申请号0333033(kumada等人)和美国专利号4,975,374(goodman等人)公开了赋予除草剂(如l-草铵膦)抗性的谷氨酰胺合成酶基因的核苷酸序列。欧洲专利号0242246和0242236(leemans等人)提供了草胺膦-乙酰-转移酶基因的核苷酸序列;以下文献中描述了表达编码草胺膦乙酰转移酶活性的嵌合bar基因的转基因植物的产生:degreef等人,(1989)在bio/technology[生物技术]7:61。还参见,美国专利号5,969,213;5,489,520;5,550,318;5,874,265;5,919,675;5,561,236;5,648,477;5,646,024;6,177,616b1和5,879,903。赋予苯氧基丙酸和环己酮(如稀禾啶和吡氟氯禾灵)抗性的示例性基因是acc1-s1、acc1-s2和acc1-s3基因,其描述于以下文献中:marshall等人,(1992)theor.appl.genet.[理论与应用遗传学]83:435。(c)抑制光合作用的除草剂,如三嗪(psba基因和gs+基因)和苄腈(腈水解酶基因)。przibilla等人,(1991)plantcell[植物细胞]3:169,描述了用编码突变体psba基因的质粒对衣藻进行转化。stalker的美国专利号4,810,648中公开了腈水解酶基因的核苷酸序列,并且包含这些基因的dna分子可在atcc登录号53435、67441和67442下获得,将该文献通过引用以其全文并入本文。编码谷胱甘肽s-转移酶的dna的克隆和表达描述于以下文献中:hayes等人,(1992)biochem.j.[生物化学杂志]285:173。(d)乙酰羟酸合酶,已经发现其使表达该酶的植物对多种类型的除草剂具有抗性,已经被引入到各种植物中(参见,例如,hattori等人,(1995)molgengenet.[分子遗传学和普通遗传学]246:419)。赋予除草剂抗性的其他基因包括:编码大鼠细胞色素p4507a1和酵母nadph-细胞色素p450氧化还原酶的嵌合蛋白的基因(shiota等人,(1994)plantphysiol.[植物生理学]106(1):17-23),针对谷胱甘肽还原酶和超氧化物歧化酶的基因(aono等人,(1995)plantcellphysiol[植物细胞生理学]36:1687)和各种磷酸转移酶的基因(datta等人,(1992)plantmolbiol[植物分子生物学]20:619)。(e)原卟啉原氧化酶(protox)是叶绿素的生成所必需的,而叶绿素是所有植物存活所必需的。原卟啉原氧化酶用作多种除草剂化合物的靶标。这些除草剂还抑制存在的所有不同种类的植物的生长,导致其完全破坏。对这些除草剂具有抗性的、含有经改变的原卟啉原氧化酶活性的植物的开发描述于以下文献中:美国专利号6,288,306b1;6,282,837b1和5,767,373;以及国际公开号wo01/12825。3.赋予或贡献于改变的谷物特征的转基因,如:(a)改变的脂肪酸,例如,通过以下方式:(1)下调硬脂酰-acp去饱和酶以提高植物的硬脂酸含量。参见,knultzon等人,(1992)proc.natl.acad.sci.usa[美国科学院院报]89:2624和wo99/64579(genesfordesaturasestoalterlipidprofilesincorn[改变玉米中脂质谱的去饱和酶的基因]);(2)通过fad-2基因修饰提高油酸和/或通过fad-3基因修饰降低亚麻酸(参见,美国专利号6,063,947、6,323,392、6,372,965和wo93/11245),(3)改变共轭亚麻酸或亚油酸含量,例如在wo01/12800中,(4)改变lec1、agp、dek1、superal1、mi1ps、各种lpa基因(如lpa1、lpa3、hpt或hggt)。例如,参见,wo02/42424、wo98/22604、wo03/011015,美国专利号6,423,886、美国专利号6,197,561、美国专利号6,825,397,美国专利申请公开号2003/0079247、2003/0204870,wo02/057439、wo03/011015以及rivera-madrid等人,(1995)proc.natl.acad.sci.[美国科学院院报]92:5620-5624。(b)改变的磷含量,例如,通过(1)引入植酸酶编码基因将促进植酸盐的分解,向经转化的植物中添加更多的游离磷酸根。例如,参见vanhartingsveldt等人,(1993)gene[基因]127:87,该文献公开了黑曲霉(aspergillusniger)植酸酶基因的核苷酸序列。(2)上调降低植酸盐含量的基因。在玉米中,这例如可通过以下方式来实现:将与一种或多种等位基因如lpa等位基因(在以低水平植酸为特征的玉米突变株中鉴定到)相关的dna进行克隆并再引入,如raboy等人,(1990)maydica[美迪卡杂志]35:383中所述,和/或通过改变肌醇激酶活性,如wo02/059324、美国专利申请公开号2003/0009011、wo03/027243、美国专利申请公开号2003/0079247、wo99/05298、美国专利号6,197,561、美国专利号6,291,224、美国专利号6,391,348、wo2002/059324、美国专利申请公开号2003/0079247、wo98/45448、wo99/55882、wo01/04147中所述。(c)例如,通过改变影响淀粉分支模式的酶的基因,或改变硫氧还蛋白如ntr和/或trx(参见,美国专利号6,531,648)和/或γ玉米醇溶蛋白敲除或突变体如cs27或tusc27或en27的基因(参见,美国专利号6,858,778和美国专利申请公开号2005/0160488和2005/0204418)来产生改变的碳水化合物。参见,shiroza等人,(1988)j.bacteriol.[细菌学杂志],170:810(链球菌(streptococcus)突变果糖基转移酶基因的核苷酸序列),steinmetz等人,(1985),mol.gen.genet.[分子遗传学和普通遗传学],200:220(枯草芽孢杆菌(bacillussubtilis)果聚糖蔗糖酶基因的核苷酸序列),pen等人,(1992)bio/technology[生物/技术],10:292(生产表达地衣芽孢杆菌(bacilluslicheniformis)α-淀粉酶的转基因植物),elliot等人,(1993),plantmolec.biol.[植物分子生物学],21:515(番茄转化酶基因的核苷酸序列),等人,(1993),j.biol.chem.[生物化学杂志],268:22480(大麦α-淀粉酶基因的定点诱变)和fisher等人,(1993),plantphysiol.[植物生理学],102:1045(玉米胚乳淀粉分支酶ii),wo99/10498(通过修饰udp-d-木糖4-差向异构酶、fragile1和2、ref1、hchl、c4h改进的可消化性和/或淀粉提取),美国专利号6,232,529(通过改变淀粉水平(agp)生产高油种子的方法)。以上提到的脂肪酸修饰基因还可用来通过淀粉途径和油途径的相互关系影响淀粉含量和/或组成。(d)改变的抗氧化剂含量或组成,如改变生育酚或生育三烯酚。例如,参见涉及通过改变植基异戊烯基转移酶(ppt)来操纵抗氧化剂水平的美国专利号6,787,683、美国专利申请公开号2004/0034886和wo00/68393,涉及通过改变尿黑酸香叶基香叶基转移酶(hggt)来操纵抗氧化剂水平的wo03/082899。(e)改变的必需种子氨基酸。例如,参见美国专利号6,127,600(增加种子中必需氨基酸的积累的方法)、美国专利号6,080,913(增加种子中必需氨基酸的积累的二元方法)、美国专利号5,990,389(高赖氨酸)、wo99/40209(改变种子中氨基酸组成)、wo99/29882(用于改变蛋白质的氨基酸含量的方法)、美国专利号5,850,016(改变种子中氨基酸组成)、wo98/20133(具有升高水平的必需氨基酸的蛋白质)、美国专利号5,885,802(高甲硫氨酸)、美国专利号5,885,801(高苏氨酸)、美国专利号6,664,445(植物氨基酸生物合成酶)、美国专利号6,459,019(赖氨酸和苏氨酸增加)、美国专利号6,441,274(植物色氨酸合酶β亚基)、美国专利号6,346,403(甲硫氨酸代谢酶)、美国专利号5,939,599(高硫)、美国专利号5,912,414(甲硫氨酸增加)、wo98/56935(植物氨基酸生物合成酶)、wo98/45458(具有较高百分比的必需氨基酸的工程化的种子蛋白)、wo98/42831(赖氨酸增加)、美国专利号5,633,436(增加含硫氨基酸含量)、美国专利号5,559,223(具有含可编程水平的必需氨基酸的限定的结构的合成储存蛋白,用于改进植物的营养价值)、wo96/01905(苏氨酸增加)、wo95/15392(赖氨酸增加)、美国专利申请公开号2003/0163838、美国专利申请公开号2003/0150014、美国专利申请公开号2004/0068767、美国专利号6,803,498、wo01/79516和wo00/09706(cesa:纤维素合酶)、美国专利号6,194,638(半纤维素)、美国专利号6,399,859和美国专利申请公开号2004/0025203(udpgdh)、美国专利号6,194,638(rgp)。4.控制雄性不育的基因有几种赋予遗传性雄性不育的方法,例如在基因组内单独位置处赋予雄性不育的多个突变基因,如brar等人在美国专利号4,654,465和4,727,219中所公开的,以及染色体易位,如patterson在美国专利号3,861,709和3,710,511中所述。除这些方法之外,albertsen等人的美国专利号5,432,068描述了核雄性不育的系统,其包括:鉴定对雄性能育性至关重要的基因;沉默这种对雄性能育性至关重要的天然基因;从基本的雄性能育性基因中除去天然启动子并用诱导型启动子替换;将这个遗传工程化的基因插回植物中,并因此产生雄性不育的植物,因为诱导型启动子不是“开”的,导致雄性能育性基因不被转录。通过诱导或将启动子打“开”来恢复能育性,该启动子反过来又允许赋予雄性能育性的基因被转录。(a)在绒毡层特异性启动子的控制下以及应用化学品n-ac-ppt下引入脱乙酰酶基因(wo01/29237)。(b)引入各种雄蕊特异性启动子(wo92/13956、wo92/13957)。(c)引入barnase和barstar基因(paul等人,(1992)plantmol.biol.[植物分子生物学]19:611-622)。关于细胞核雄性和雌性不育系统和基因的另外的实例,也可参见美国专利号5,859,341;6,297,426;5,478,369;5,824,524;5,850,014和6,265,640中找到。5.创建用于位点特异性dna整合的位点的基因这包括引入可以在flp/frt系统中使用的frt位点和/或可以在cre/loxp系统中使用的lox位点。例如,参见lyznik等人(2003)plantcellrep[植物细胞报告]21:925-932和wo99/25821。可以使用的其他系统包括噬菌体mu的gin重组酶(maeser等人,1991;vickichandler,themaizehandbook[玉米手册],第118章(springer-verlag[施普林格出版社]1994)、大肠杆菌的pin重组酶(enomoto等人,1983)和psr1质粒的r/rs系统(araki等人,1992)。6.影响非生物胁迫抗性(包括但不限于开花、穗和种子发育、提高氮利用效率、改变氮响应性、抗旱性或耐旱性、抗寒性或耐寒性、抗盐性或耐盐性)和胁迫下产量提高的基因。例如,参见wo00/73475,其中通过改变苹果酸盐而改变水利用效率;描述基因(包括cbf基因)和转录因子(这些基因和转录因子有效于减轻冻害、高盐度和干旱对植物的负面影响,并对植物表型赋予其他正向影响)的美国专利号5,892,009、美国专利号5,965,705、美国专利号5,929,305、美国专利号5,891,859、美国专利号6,417,428、美国专利号6,664,446、美国专利号6,706,866、美国专利号6,717,034、wo2000060089、wo2001026459、wo2001035725、wo2001034726、wo2001035727、wo2001036444、wo2001036597、wo2001036598、wo2002015675、wo2002017430、wo2002077185、wo2002079403、wo2003013227、wo2003013228、wo2003014327、wo2004031349、wo2004076638、wo9809521、和wo9938977;美国专利申请公开号2004/0148654和wo01/36596,其中植物中的脱落酸被改变,导致植物表型改进,如产量提高和/或对非生物胁迫的耐受性提高;wo2000/006341、wo04/090143、美国专利申请序列号10/817483和美国专利号6,992,237,其中细胞分裂素表达被改变,产生胁迫耐受性(如耐旱性)提高和/或产量提高的植物,将这些文献通过引用以其全文并入本文。还参见wo0202776、wo2003052063、jp2002281975、美国专利号6,084,153、wo0164898、美国专利号6,177,275和美国专利号6,107,547(氮利用率的提高和氮响应性的改变)。对于乙烯改变,参见美国专利申请公开号2004/0128719、美国专利申请公开号2003/0166197和wo2000/32761。对于非生物胁迫的植物转录因子或转录调节子,参见,例如,美国专利申请公开号2004/0098764或美国专利申请公开号2004/0078852。影响植物生长和农艺性状(如产量、开花、植物生长和/或植物结构)的其他基因和转录因子可被引入或渗入到植物中,参见例如wo97/49811(lhy)、wo98/56918(esd4)、wo97/10339和美国专利号6,573,430(tfl)、美国专利号6,713,663(ft)、wo96/14414(con)、wo96/38560、wo01/21822(vrn1)、wo00/44918(vrn2)、wo99/49064(gi)、wo00/46358(fri)、wo97/29123、美国专利号6,794,560、美国专利号6,307,126(gai)、wo99/09174(d8和rht)以及wo2004076638和wo2004031349(转录因子)。“rnai”是指用于降低基因表达的一系列相关技术(参见例如美国专利号6,506,559)。这些名称包括“反义抑制”,即产生能够抑制目标蛋白质的表达的反义rna转录物,以及“共抑制”或“正义抑制”,指产生能够抑制相同的或者基本上相似的外来基因或者内源基因的表达的正义rna转录物(美国专利号5,231,020)。此类技术依赖于使用能导致积累这样的双链rna的构建体,该双链rna中的一条链与待沉默的靶基因互补。本文公开的调节序列可用来驱动将会产生rna干扰的构建体(包括微rna和sirna)的表达。一些实施例涉及通过干扰核糖核酸(rna)分子下调昆虫有害生物物种中靶基因的表达。pct公开wo2007/074405描述了抑制无脊椎动物有害生物(包括科罗拉多马铃薯甲虫)中靶基因表达的方法。pct公开wo2005/110068描述了抑制无脊椎动物有害生物(特别是包括西方玉米根虫)中靶基因表达的方法,该方法作为防治昆虫侵染的手段。此外,pct公开wo2009/091864描述了用于抑制来自昆虫有害生物物种(包括来自草盲蝽属的有害生物)的靶基因的组合物和方法。核酸分子包括用于靶向液泡atp酶h亚基的沉默元件,该沉默元件可用于防治如美国专利申请公开2012/0198586中所述的鞘翅目有害生物群体和侵袭。pct公开wo2012/055982描述了抑制或下调编码以下的靶基因表达的核糖核酸(rna或双链rna):昆虫核糖体蛋白,如核糖体蛋白l19、核糖体蛋白l40或核糖体蛋白s27a;昆虫蛋白酶体亚基,如rpn6蛋白、pros25、rpn2蛋白、蛋白酶体β1亚基蛋白或prosβ2蛋白;copi囊泡的昆虫β-外被体、copi囊泡的γ-外被体、copi囊泡的β′-外被体蛋白或ζ-外被体;昆虫跨膜四蛋白(tetraspanin)2a蛋白(推定的跨膜结构域蛋白);属于肌动蛋白家族的昆虫蛋白,例如肌动蛋白5c;昆虫泛素-5e蛋白;昆虫sec23蛋白,其是参与细胞内蛋白质转运的gtp酶活化剂;涉及运动活性的作为非常规肌球蛋白的昆虫皱纹蛋白质;涉及核替代mrna剪接调节的昆虫曲颈蛋白;昆虫囊泡h+-atp酶g亚基蛋白和昆虫tbp-1如tat结合蛋白。pct公开wo2007/035650描述了抑制或下调编码snf7的靶基因表达的核糖核酸(rna或双链rna)。美国专利申请公开2011/0054007描述了靶向rps10的多核苷酸沉默元件。美国专利申请公开2014/0275208和us2015/0257389描述了靶向ryanr、hp2和pat3的多核苷酸沉默元件。美国专利申请公开2011/0054007描述了靶向rps10的多核苷酸沉默元件。pct公开wo2016/205445描述了减少生育力的多核苷酸沉默元件,包括nclb、mael、boule、和vgr。pct公开wo/2016/138106、wo2016/060911、wo2016/060912、wo2016/060913、和wo2016/060914描述了靶向赋予对鞘翅目和半翅目有害生物的抗性的copi外被体亚单位核酸分子的多核苷酸沉默元件。美国专利申请公开2012/029750、us20120297501和2012/0322660描述了干扰核糖核酸(rna或双链rna),所述干扰核糖核酸在被昆虫有害生物物种摄取时起作用以下调所述昆虫有害生物中靶基因的表达,其中所述rna包含至少一个沉默元件,其中所述沉默元件是包含经退火的互补链的双链rna区域,所述双链rna区域的一条链包含或由如下核苷酸序列组成,所述核苷酸序列至少部分地与靶基因中的靶标核苷酸序列互补。美国专利申请公开2012/0164205描述了用于干扰双链核糖核酸(用于抑制无脊椎动物有害生物)的潜在靶标,包括:chd3同源序列、β-微管蛋白同源序列、40kdav-atp酶同源序列、ef1α同源序列、26s蛋白质体亚基p28同源序列、保幼激素环氧化物酶水解酶同源序列、溶胀依赖氯通道蛋白同源序列、葡萄糖-6-磷酸1-脱氢酶蛋白同源序列、act42a蛋白同源序列、adp-核糖因子1同源序列、转录因子iib蛋白同源序列、几丁质酶同源序列、泛素缀合酶同源序列、甘油醛-3-磷酸脱氢酶同源序列、泛素b同源序列、保幼激素酯酶同源物、和α微管蛋白同源序列。可对本文公开的分离的调节元件序列进行修饰,以提供所述异源核苷酸序列的一系列水平表达。因此,可利用完整调节元件区域的一部分,且驱动目的核苷酸序列表达的能力得到保持。应认识到,可采用不同方式缺失启动子序列的一部分,来改变mrna的表达水平。例如,如果在截短过程中移除(阻遏蛋白的)负调节元件,则因调节元件缺失,mrna表达水平可能降低,或者可替代地,其表达可能增加。通常,使用至少约20个核苷酸的分离的调节元件序列来驱动多核苷酸序列表达。方便的终止区可获得自根癌农杆菌(a.tumefaciens)的ti质粒,如章鱼碱合酶和胭脂碱合酶终止区。还参见,guerineau等人,(1991)mol.gen.genet.[分子遗传学和普通遗传学]262:141-144;proudfoot,(1991)cell[细胞]64:671-674;sanfacon等人,(1991)genesdev.[基因与发育]5:141-149;mogen等人,(1990)plantcell[植物细胞]2:1261-1272;munroe等人,(1990)gene[基因]91:151-158;ballas等人,(1989)nucleicacidsres.[核酸研究]17:7891-7903;以及joshi等人,(1987)nucleicacidres.[核酸研究]15:9627-9639。包含本文公开的序列的表达盒还可含有待共转化到该生物体中的基因的至少一个另外的核苷酸序列。可替代地,所述一个或多个另外的序列可以在另一表达盒中提供。在适当的情况下,可以优化其表达处于本公开的杂合调节元件序列控制下的多核苷酸序列和任何另外的一个或多个核苷酸序列,以便在转化的植物中增加表达。也就是说,可使用植物偏好性密码子来合成这些核苷酸序列,从而改进表达。对于宿主偏好性密码子使用的讨论,参见,例如campbell和gowri,(1990)plantphysiol.[植物生理学]92:1-11。本领域中可获得用于合成植物偏好的基因的方法。参见例如,murray等人,(1989)nucleicacidsres.[核酸研究]17:477-498。已知有另外的序列修饰能增强细胞宿主中的基因表达。这些包括消除以下序列:编码假聚腺苷酸化信号、外显子-内含子剪接位点信号、转座子样重复的序列,及可能不利于基因表达的其他经充分表征的序列。可以将异源多核苷酸序列的g-c含量调整至给定细胞宿主的平均水平,所述平均水平通过参考该宿主细胞中表达的已知基因来计算。当可能时,修饰序列以避免出现可预见的发夹二级mrna结构。表达盒可以另外包含5′前导序列。这些前导序列可以起到增强翻译的作用。翻译前导序列是本领域已知的并且包括但不限于:小核糖核酸病毒前导序列,例如,emcv前导序列(脑心肌炎5’非编码区域)(elroy-stein等人,(1989)proc.nat.acad.sci.usa[美国科学院院报]86:6126-6130);马铃薯y病毒组前导序列,例如,tev前导序列(烟草蚀纹病毒)(allison等人,(1986)virology[病毒学]154:9-20);mdmv前导序列(玉米矮花叶病毒);人类免疫球蛋白重链结合蛋白(bip)(macejak等人,(1991)nature[自然]353:90-94);来自苜蓿花叶病病毒的外壳蛋白mrna的非翻译前导序列(amvrna4)(jobling等人,(1987)nature[自然]325:622-625);烟草花叶病毒前导序列(tmv)(gallie等人,(1989)molecularbiologyofrna[rna的分子生物学],第237-256页);和玉米褪绿斑驳病毒前导序列(mcmv)(lommel等人,(1991)virology[病毒学]81:382-385)。还参见,della-cioppa等人,(1987),plantphysiology[植物生理学],84:965-968。也可采用已知的用来提高mrna稳定性的方法,例如内含子,如玉米泛素内含子(christensen和quail,(1996)transgenicres.[转基因研究]5:213-218;christensen等人,(1992)plantmolecularbiology[植物分子生物学]18:675-689)或者玉米adhi内含子(kyozuka等人,(1991)mol.gen.genet.[分子遗传学和普通遗传学]228:40-48;kyozuka等人,(1990)maydica[美迪卡杂志]35:353-357)等。在制备表达盒时,可以操作各种dna片段,以提供处于适当取向以及合适时,处于适当阅读框中的dna序列。为此,可采用衔接子(adapter)或接头以连接dna片段,或可以涉及其他操作以提供方便的限制位点、移除多余的dna、移除限制位点等。出于这个目的,可以涉及体外诱变、引物修复、限制性酶切(restriction)、退火、再取代(例如转换和颠换)。还可以在表达盒中包含报道基因或者选择性标记基因。本领域已知的合适报道基因的实例可以见于例如以下文献:jefferson等人,(1991)plantmolecularbiologymanual[植物分子生物学手册],gelvin等人编辑(kluweracademicpublishers[克吕韦尔学术出版集团]),第1-33页;dewet等人,(1987)mol.cell.biol.[分子细胞生物学]7:725-737;goff等人,(1990)emboj.[欧洲分子生物学学会杂志]9:2517-2522;kain等人,(1995)biotechniques[生物技术]19:650-655;和chiu等人,(1996)currentbiology[当代生物学]6:325-330。用于选择转化细胞或者组织的选择性标记基因可以包括赋予抗生素抗性或除草剂抗性的基因。合适的选择性标记基因的实例包括但不限于编码对以下具有抗性的基因:氯霉素(herreraestrella等人,(1983)emboj.[欧洲分子生物学学会杂志]2:987-992);甲氨蝶呤(herreraestrella等人,(1983)nature[自然]303:209-213;meijer等人,(1991)plantmol.biol.[植物分子生物学]16:807-820);潮霉素(waldron等人,(1985)plantmol.biol.[植物分子生物学]5:103-108和zhijian等人,(1995)plantscience[植物科学]108:219-227);链霉素(jones等人,(1987)mol.gen.genet.[分子遗传学和普通遗传学]210:86-91);壮观霉素(bretagne-sagnard等人,(1996)transgenicres.[转基因研究]5:131-137);博莱霉素(hille等人,(1990)plantmol.biol.[植物分子生物学]7:171-176);磺酰胺(guerineau等人(1990)plantmol.biol.[植物分子生物学]15:127-36);溴苯腈(stalker等人,(1988)science[科学]242:419-423);草甘膦(shaw等人,(1986)science[科学]233:478-481和美国专利申请序列号10/004,357和10/427,692);草丁膦(deblock等人,(1987)emboj.[欧洲分子生物学学会杂志]6:2513-2518)。其他可在转基因事件的恢复中发挥实用的基因将包括但不限于以下实例,如:gus(β-葡糖醛酸糖苷酶;jefferson,(1987)plantmol.biol.rep.[植物分子生物学报告]5:387)、gfp(绿色荧光蛋白;chalfie等人,(1994)science[科学]263:802)、荧光素酶(riggs等人,(1987)nucleicacidsres.[核酸研究]15(19):8115和luehrsen等人,(1992)methodsenzymol.[酶学方法]216:397-414)以及编码用于花青素生产的玉米基因(ludwig等人,(1990)science[科学]247:449)。包含可操作地连接至目的多核苷酸序列的杂合调节元件的表达盒可用于转化任何植物。在另一个实施例中,包含可操作地连接至目的多核苷酸序列的seqidno:1-74的序列的表达盒可用于转化任何植物。以此方式,可以获得经基因修饰的植物、植物细胞、植物组织、种子、根等。本文公开的某些方法涉及将多核苷酸引入到植物中。如本文所用,“引入”旨在表示将多核苷酸呈递给植物,以这样的方式使得序列进入植物细胞的内部。本公开的方法不取决于将序列引入植物中的具体方法,只要多核苷酸进入植物的至少一个细胞的内部即可。将多核苷酸引入植物的方法是本领域已知的,所述方法包括但不限于稳定转化法、瞬时转化法和病毒介导法。“稳定转化”是其中引入到植物中的多核苷酸构建体整合到植物的基因组中并能由其子代继承的转化。“瞬时转化”意指将多核苷酸引入植物中并且它不整合到该植物的基因组中。转化方案以及将核苷酸序列引入植物中的方案,可根据要靶向转化的植物或者植物细胞的类型(即单子叶植物或者双子叶植物)而变化。将核苷酸序列引入到植物细胞中并随后插入到植物基因组中的合适方法包括显微注射(crossway等人,(1986)biotechniques[生物技术]4:320-334)、电穿孔(riggs等人,(1986)proc.natl.acad.sci.usa[美国科学院院报]83:5602-5606)、农杆菌介导的转化(townsend等人,美国专利号5,563,055和zhao等人,美国专利号5,981,840)、直接基因转移(paszkowski等人,(1984)emboj[欧洲分子生物学学会杂志]3:2717-2722)以及弹道粒子加速(参见例如,美国专利号4,945,050;5,879,918;5,886,244和;5,932,782;tomes等人,(1995),在plantcell,tissue,andorganculture:fundamentalmethods[植物细胞、组织和器官培养:基本方法],gamborg和phillips编辑(springer-verlag,berlin[德国柏林施普林格出版公司]);mccabe等人(1988)biotechnology[生物技术]6:923-926);以及lec1转化法(wo00/28058)。还参见weissinger等人,(1988)ann.rev.genet.[遗传学年鉴]22:421-477;sanford等人,(1987)particulatescienceandtechnology[微粒科学与技术]5:27-37(洋葱);christou等人,(1988)plantphysiol.[植物生理学]87:671-674(大豆);mccabe等人,(1988)bio/technology[生物/技术]6:923-926(大豆);finer和mcmullen,(1991)invitrocelldev.biol.[体外细胞与发育生物学]27p:175-182(大豆);singh等人,(1998)theor.appl.genet.[理论与应用遗传学]96:319-324(大豆);datta等人,(1990)biotechnology[生物技术]8:736-740(稻);klein等人,(1988)proc.natl.acad.sci.usa[美国科学院院报]85:4305-4309(玉米);klein等人,(1988)biotechnology[生物技术]6:559-563(玉米);美国专利号5,240,855、5,322,783、和5,324,646;klein等人,(1988)plantphysiol.[植物生理学]91:440-444(玉米);fromm等人,(1990)biotechnology[生物技术]8:833-839(玉米);hooykaas-vanslogteren等人,(1984)nature[自然](伦敦)311:763-764;美国专利号5,736,369(谷类);bytebier等人,(1987)proc.natl.acad.sci.usa[美国科学院院报]84:5345-5349(百合科);dewet等人,(1985)在theexperimentalmanipulationofovuletissues[卵巢组织的实验操作]中,chapman等人编辑(纽约朗文出版社(longman,newyork)),第197-209页(花粉);kaeppler等人,(1990)plantcellreports[植物细胞报告]9:415-418和kaeppler等人,(1992)theor.appl.genet.[理论与应用遗传学]84:560-566(晶须介导的转化);d′halluin等人,(1992)plantcell[植物细胞]4:1495-1505(电穿孔);li等人,(1993)plantcellreports[植物细胞报告]12:250-255以及christou和ford(1995)annalsofbotany[植物学年鉴]75:407-413(稻);osjoda等人,(1996)naturebiotechnology[自然生物技术]14:745-750(经由根癌农杆菌的玉米)。在一个实施例中,可以使用各种瞬时转化方法将包含杂合调节元件的dna构建体提供给植物。在另一个实施例中,可以使用各种瞬时转化方法将包含所公开的序列seqidno:1-74的dna构建体提供给植物。这类瞬时转化方法包括但不限于病毒载体系统,和以阻止dna后续释放的方式沉淀多核苷酸。因此,可以从微粒结合的dna进行转录,但其被释放以整合至基因组的频率大大降低了。这种方法包括使用包被有聚乙烯亚胺(pei;sigma#p3143)的颗粒。在其他实施例中,可以通过使植物与病毒或病毒核酸接触而将多核苷酸引入植物中。通常,这类方法涉及将本公开的多核苷酸构建体掺入病毒dna或rna分子内。涉及病毒dna或rna分子、用于将多核苷酸引入植物中并表达其中所编码的蛋白质的方法是本领域已知的。参见例如,美国专利号5,889,191、5,889,190、5,866,785、5,589,367、5,316,931和porta等人,(1996)molecularbiotechnology[分子生物技术]5:209-221。用于在植物基因组的特定位置靶向插入多核苷酸的方法是本领域已知的。在一个实施例中,利用位点特异性重组系统实现多核苷酸在所希望的基因组位置处的插入。参见例如wo99/25821、wo99/25854、wo99/25840、wo99/25855和wo99/25853。简言之,可以在转移盒中包含本公开的多核苷酸,所述转移盒侧翼为两个不相同的重组位点。将该转移盒引入植物中,该植物已经将靶位点稳定地掺入其基因组中,该靶位点侧翼为与转移盒的位点相对应的两个不相同的重组位点。提供适当的重组酶,并将该转移盒整合到靶位点。由此,目的多核苷酸被整合在植物基因组中的具体染色体位置处。可依据常规方式将已转化的细胞培育成植株。参见例如,mccormick等人,(1986)plantcellreports[植物细胞报告]5:81-84。然后可以培育这些植株,并用相同的经转化的株系或者不同的株系进行授粉,并鉴定出具有所希望的表型特征的表达的所得后代。可以生长两代或两代以上以确保希望的表型特征的表达稳定保持并且遗传,然后收获种子以确保希望的表型特征已经实现表达。以此方式,本公开提供了其基因组中稳定掺入了多核苷酸构建体(例如,包含seqidno:1-74之一的表达盒)的转化种子(也称为“转基因种子”)。有各种各样的方法用于从植物组织再生植物。特定的再生方法将取决于起始植物组织和待再生的特定植物种类。来自单一植物原生质体转化体或来自各种转化的外植体的植物的再生、发育和培养是本领域所熟知的(weissbach和weissbach(编辑),(1988)在methodsforplantmolecularbiology[植物分子生物学方法]中,academicpress,inc.[学术出版社有限公司],加利福尼亚州圣地亚哥(sandiego,calif.)。这种再生和生长过程通常包括如下步骤:选择经转化的细胞,通过胚性发育的通常阶段、通过生根苗阶段培养那些个体化细胞。以同样的方式再生转基因胚和种子。然后将所得的转基因生根芽苗种植在合适的植物生长培养基(如土壤)中。优选地,再生植物自花授粉以提供纯合的转基因植物。或者,将得自再生植物的花粉与农学上重要的品系的产生种子的植物进行杂交。相反地,将来自这些重要品系的植物的花粉用于给再生植物授粉。使用本领域技术人员熟知的方法培养含有所希望的多核苷酸的各实施例的转基因植物。这些实施例提供了用于筛选化合物的组合物,所述化合物调节植物内的表达。载体、细胞和植物可用于筛选seqidno:1-74的调节元件序列的激动剂和拮抗剂的候选分子。例如,可以将报道基因与调节元件序列可操作连接并作为转基因在植物中表达。添加待检验的化合物并测量报道基因的表达以确定其对启动子活性的影响。在一个实施例中,可以使用crispr/cas9系统通过基因组编辑来将调节元件例如序列seqidno:1-74编辑或插入到植物中。在一个方面,可以使用基因组编辑技术将所公开的调节元件引入植物的基因组中,或者可以使用基因组编辑技术编辑植物基因组中先前引入的调节元件。例如,可以通过使用双链断裂技术(如talen、大范围核酸酶、锌指核酸酶、crispr-cas等)将公开的调节元件引入植物基因组中期望的位置中。例如,为了位点特异性插入的目的,可以使用crispr-cas系统将所公开的调节元件引入基因组中期望的位置中。植物基因组中期望的位置可以是任何对于插入来说期望的靶位点,例如适于育种的基因组区域,或者可以是位于具有现有的目的性状的基因组窗口中的靶位点。现有的目的调节元件可能是内源调节元件或先前引入的调节元件。在另一个方面,在所公开的调节元件先前已经被引入到基因组中的情况下,可以使用基因组编辑技术来改变或修饰引入的调节元件序列。可以引入到所公开的调节元件组合物中的位点特异性修饰包括使用用于引入位点特异性修饰的任何方法产生的修饰,该方法包括但不限于通过使用基因修复寡核苷酸(例如美国公开2013/0019349),或通过使用双链断裂技术,如talen、大范围核酸酶、锌指核酸酶、crispr-cas等。此类技术可用于通过在引入的多核苷酸内的核苷酸的插入、缺失或取代来修饰先前引入的多核苷酸。可替代地,可以使用双链断裂技术向引入的多核苷酸中添加另外的核苷酸序列。“改变的靶位点”、“改变的靶序列”、“修饰的靶位点”和“修饰的靶序列”在本文中可互换地使用,并且意指如本文公开的靶序列,当与未改变的靶序列相比时,该靶序列包含至少一种改变。此类“改变”包括,例如:(i)至少一个核苷酸的替代、(ii)至少一个核苷酸的缺失、(iii)至少一个核苷酸的插入、或(iv)(i)-(iii)的任何组合。本说明书中提到的所有出版物、专利和专利申请指示了本公开所属领域技术人员的水平。将所有出版物、专利和专利申请通过引用并入本文,其程度就像明确且单独地指出每一个单独的出版物、专利或专利申请通过引用结合一样。对本公开的不同所示实施例的上述描述并不旨在是详尽的或者限制范围于所公开的精确形式。虽然为了说明目的而在此描述了具体实施例和实例,但是在本公开范围内,如相关领域的技术人员将认识到的,不同的等效修饰是可能的。本文提供的教导可以应用于除了上述实例之外的其他目的。根据上述教导,许多修改和变化是可能的,因此它们在所附权利要求书的范围内。可以根据上述详细描述进行这些改变和其他改变。通常,在以下权利要求书中,所使用的术语不应被解释为将范围限制于说明书和权利要求书中公开的具体实施例。就使用的数字(例如量、温度、浓度等)而言,已努力确保其准确性,但仍应允许有一些实验误差和偏差。除非另有说明,份为重量份;分子量为平均分子量;温度是摄氏度;并且压力为大气压或接近大气压。实验实例1:根杂合调节元件根是复杂的器官,其可分为不同的区域;例如根冠、分生组织、伸长区和成熟区。这些区域反映了根细胞身份和成熟度的不同阶段。这些细胞中的基因被激活和灭活,并且因此可能不会在根的所有区域中同时表达。调节元件控制这些基因的表达。鉴定具有所需表达特征的天然存在的调节元件在许多不同水平上提出了挑战,并且可导致转基因植物中未预料到的表达水平和模式。天然存在的调节元件的替代物是通过组合不同调节元件的区段而产生的杂合调节元件。可以组合在玉米根的不同区域中起作用的调节元件的区段,使得产生在根的全部或大部分区域中起作用的杂合调节元件。优选地,尽管可能发生异位表达,但根特异性得到维持。实例2:杂合根调节元件序列的构建使用先前已经表征了表达的调节元件的区段构建了各杂合调节序列(seqidno:11-30和35-60)(步骤1;表1)。通过表达模式、根尖和成熟区将天然调节元件粗略地分为两组。根尖包括根冠、分生组织和邻近分生组织的伸长区的区域。成熟区包括根的成熟区和邻近成熟区的伸长区的部分。使用公共数据库和专有数据库评估属于任一类别的调节元件的潜在基序,以帮助定义可用于杂合调节元件中的区段(步骤2)。用于在该过程中进行筛选的基序算法是本领域已知的,例如higo,k等人(1998)nucleicacidsresearch[核酸研究]。通过评估基序的位置、基序的大小和类型以及基序的聚类来启动该过程。tata盒与tata盒上游约1kb之间的基序聚类有利于作为用于杂合启动子的区段而不是更远端的区段。第三个步骤(步骤3)包括贡献该杂合调节元件的3’端(包括tata盒)的所选区段和含有该杂合调节元件的5’端的区段的组装(表2)。组装过程通常将根尖区段置于该调节元件的3’端。将seqidno:11-18各自克隆到各自的表达载体中以引入玉米植物中。将β-葡糖醛酸糖苷酶(gus)基因用作报道基因以表征各杂合调节元件指导的表达。每个载体再生大约10个玉米转基因事件。表1:转基因玉米植物中根启动子的表达模式基于组织化学染色zm=玉米;sb=高梁;bd=二穗短柄草(brachypodiumdistachyon);si=粟(setariaitalica)表2:向根杂合启动子贡献区段的亲本调节元件zm=玉米;sb=高粱;bd=二穗短柄草;si=粟实例3:转基因玉米中调节元件的表达分析表3示出了根中各杂合调节元件的表达结果。总体而言,各杂合调节元件驱动表达并提供通常在根中普遍存在的表达模式。在分生组织、伸长区和成熟区中的事件之间的表达几乎一致。根冠中的表达更加可变,因为仅一个天然存在的启动子(seqidno:1;表1)明确地驱动在根冠中的表达。因此,在这些实验中,在不存在seqidno:1的情况下,根冠中的表达是异位的。杂合调节元件的表达水平通常比个体亲本高2倍或更多;然而,并非所有杂合调节元件都以较高水平表达,并且要么等同于表达最高的亲本,要么略低于表达最高的亲本。使用来自亲本的区段(seqidno:8和seqidno:4)构建seqidno:11,所述亲本的区段具有最小重叠的空间表达特征。将来自seqidno:8的区段的三个拷贝与来自seqidno:4的区段的一个拷贝融合。组织化学染色的转基因玉米根的结果示出表达大部分都位于转基因根的分生组织和成熟区,这体现了两个亲本调节区段。在更高的表达事件中,该表达模式混合在一起形成了在分生组织、伸长区和成熟区的更一致的表达。seqidno:11的亲本区段均未在根冠中驱动表达;然而,一些事件确实示出了在根冠中的异位表达。另外的观察结果是在茎中表达,所述表达由seqidno:4所示的亲本调节元件表现出。seqidno:12表明来自不同物种的区段能以杂合形式一起起作用。seqidno:6来源于高梁并且在伸长区驱动比seqidno:8更高水平的表达。将来自seqidno:6的亲本调节元件区段的三个拷贝以类似于seqidno:11的构型置于来自seqidno:4的亲本调节元件区段的上游。在组织化学染色的转基因根中,构建的结果是相对于seqidno:11更一致的表达模式。定量数据示出seqidno:12指导的表达水平分别超过seqidno:6和seqidno:4的个体亲本调节元件的水平。增加约为3倍。观察根冠中的异位表达,并且观察茎中的表达。seqidno:13通过分别组合来自seqidno:8、6和4的各区段的一个拷贝来消除重复区段。组织化学染色结果在根冠、分生组织、伸长区和成熟区示出一致表达。组织化学染色是将取样的组织浸入含有底物x-gluc(钠盐)的溶液中的结果,其导致在表达β-葡糖醛酸糖苷酶的组织中的部位处出现蓝色沉淀。seqidno:14和15是用不同于来自seqidno:8、6和4的那些区段构建的杂合调节元件。在seqidno:14中,来自seqidno:4的区段被来自seqidno:2的区段替换。seqidno:2具有与seqidno:4类似的表达模式,但其来自二穗短柄草,而seqidno:4来自玉米。在来自seqidno:2和seqidno:6的区段之间添加来自seqidno:5的区段。seqidno:15由来自三个物种的区段组成。seqidno:9来源于玉米,seqidno:5来自高粱,并且seqidno:2获得自短柄草属(表2)。seqidno:15与seqidno:14非常相似,区别是seqidno:6的区段取代了来自seqidno:9的区段。seqidno:14和seqidno:15均驱动在玉米根中的表达(表3)。在根冠中观察到异位表达。其他表达位点包括具有低水平茎和外壳表达的一些seqidno:14和seqidno:15事件,这些事件与具有茎表达和低水平叶表达的seqidno:13不同。seqidno:16通过将具有相似表达模式的区段(用于主要在分生组织中的表达,例如seqidno:2和seqidno:4)替换为来自亲本调节元件的区段(驱动在根尖的不同部分的表达)来证明额外的灵活性。出于seqidno:1在转基因玉米植物中的根冠偏好而选择seqidno:1,并将其用于代替seqidno:14构型中的seqidno:2。seqidno:16驱动在根冠和根的其他区域中的表达(表3)。未检测到在根以外的表达。seqidno:17探索了使用另一个来自短柄草属的调节元件区段seqidno:3代替seqidno:2以及用来自粟的区段seqidno:10来替换玉米区段seqidno:9。seqidno:18与seqidno:17相似,除了将来自高粱的seqidno:5用来自高梁的另一区段seqidno:7替换(表2)。seqidno:17和seqidno:18均在转基因玉米根中起作用,其中seqidno:17比seqidno:18在根冠中产生更多的异位表达(表3)。seqidno:17和18的几个事件具有某些茎表达。来自seqidno:11-18的组合结果表明,可以组装来自玉米、高粱、短柄草属和粟的在玉米根的不同区域起作用的区段以产生在玉米根的所有区域中起作用的杂合调节元件(表3)。表3:在转基因玉米根中各杂合调节元件的表达结果杂合调节元件根冠分生组织伸长区成熟区seqid11++++++++++seqid12++++++++++++++seqid13++++++++++++++++seqid14++++++++++++++seqid15++++++++++++seqid16++++++++++++++seqid17++++++++++++++++seqid18++++++++++基于组织化学染色在玉米植物中测试了另外的区段组合,其包含杂合启动子(seqidno:13和17)的区段组分。将各亲本调节元件的三个区段分配到一个位置中,其中“a”表示5’区段,“b”表示中间区段并且“c”表示3’区段(参见图2)。然后将区段置于重新排序的组合中,如图2和表4和表5所示,以测试杂合启动子中各区段的相对位置。改变seqidno:13的三个区段的顺序导致三种组合中的两种的亲本类型表达(表4)。未示出亲本类型表达的组合在根冠中的表达降低。这些结果表明该调节元件内的区段的位置的灵活性,但并非所有组合都会产生相同的结果。这些结果由seqidno:17的重新排序支持,seqidno:17由与seqidno:13不同的区段组成。三种组合中的两种导致亲本类型表达,并且与seqidno:13一样,一种组合没有导致亲本类型表达(表5)。与seqidno:13相比(cba对比cab),未示出亲本表型的组合也具有不同的区段排列。表4:转基因玉米根中杂合调节元件的相对定向区段的表达基于组织化学染色表5:转基因玉米根中杂合调节元件的相对定向区段的表达为了测试仅使用亲本调节元件的两个区段以在杂合启动子中产生独特的表达模式,使用seqidno:13的两个区段进行成对组合,并将其与seqidno:13的表达进行比较(表6)。结果示出可以制备反映seqidno:13表达的组合,但是不制备不反映seqidno:13表达的组合。在那些不反映表达的组合中,一个组合(ca)并不完全反映各区段的贡献。结果表明,两个区段足以产生在根的不同区域起作用的启动子,但是区段组合在一起以及它们如何组合在一起可能影响表达结果。表6:转基因玉米根中杂合调节元件的相对定向区段的表达实例4:地上部分杂合调节元件用于产生根杂合调节元件的方法还可用于其他的杂合调节元件类型,包括表达地上组织的杂合调节元件(“地上部分调节元件”)。组合来自玉米须偏好性亲本调节元件(seqidno:48)和短柄草属绿色组织亲本调节元件(seqidno:49)的区段,如在实例3中针对seqidno:11所述,其中将来自须偏好性调节元件的重复区段置于为seqidno:48提供tata盒的绿色组织区段的上游。还进行了相反的情况,其中来自玉米须偏好性亲本调节元件(seqidno:48)的须表达区段提供tata盒,并且短柄草属绿色组织亲本调节元件(seqidno:49)的多个区段位于地上部分杂合调节元件的上游。表7和图3所示的梳理这些元件的结果是所得的杂合调节元件驱动在玉米植物的叶和须中的转基因表达。在其他组织例如根和茎中的表达与亲本相比没有显著的差异。总之,营养性绿色组织启动子区段可与生殖性非绿色组织区段组合以在玉米中产生所希望的表达模式。来自亲本调节元件的区段可以源自不同物种。预期组合这些区段未必使目的组织中的表达增加远远超过亲本提供的水平。因此,为了帮助调节增加水平方面的表达,使用三个不同植物病毒启动子的-20区代替为地上部分杂合调节元件提供tata盒的区段的-20区。病毒启动子通常是植物中的强启动子,因此这些启动子的-20区(由tata盒区域和与基础转录机制相互作用的其他序列组成)可被视为表达的“优化”。因此,将上游调节区段与这些“优化的”序列元件连接可以导致杂合调节元件的表达水平增加。所选择的提供-20区的三个病毒启动子是bydv(豆黄矮病毒,seqidno:67)、hlcv(蜀葵皱叶病毒,seqidno:66)和香蕉条纹病毒(ay,seqidno:65)。这些代表双生病毒科(geminiyiridae)和花椰菜花叶病毒科(caulimoviridae)病毒家族,尽管可以使用其他病毒启动子和来自除双生病毒科和花椰菜花叶病毒科之外的其他家族的启动子。表7示出-20区中的每一个都正向影响在叶和须中的表达。在根和茎中的表达未以同样的方式受到影响。bsv(ay)-20区提供了最显著增加的表达,其包括外壳表达的增加。表7:双生病毒科和花椰菜花叶病毒科-20区正向影响在叶和须中的表达制备了一组杂合调节元件,其由非冗余亲本区段组成。提供tata盒的区段(seqidno:48)产生在叶和须组织中的表达,但其中须表达低于叶表达(表8)。为了增加须表达,将来自两种不同的须偏好性启动子的区段(seqidno:61和62)与seqidno:48组合进行测试。在第一种杂合调节元件中,添加了来自seqidno:61的一个区段(seqidno:47)。观察到须表达的增加。叶和根表达保持大约相同,并且茎表达略微增加。添加seqidno:62(seqidno:56)导致须表达的更加显著的增加。叶和茎表达略有下降,并且根表达没有变化。在第三种杂合调节元件中,将来自具有seqidno:56的杂合调节元件的5’须区段用绿色组织区段(seqidno:64)替换以产生具有seqidno:57的地上部分杂合调节元件。结果是叶表达增加和须表达恢复到以前的水平。茎表达略有增加并且根表达保持不变。这些结果证明了使用非冗余亲本调节元件区段以产生在玉米的靶组织中起作用的杂合调节元件的能力,其中这些非冗余区段可能来自不同的物种(在这种情况下来自二穗短柄草)。表8:在转基因玉米中各杂合调节元件的表达结果与根杂合调节元件类似,彼此相对定向的区段可影响设计用于在其他组织中表达的杂合调节元件的表达结果(关于区段定向,请参见图2)。表9示出了表达结果,绿色组织区段位于杂合调节元件(seqidno:54)的5’端,相对于提供tata盒(seqidno:55)的3’位置。当绿色组织区段位于5′位置时,叶表达没有增加(seqidno:54;表9)。然而,当绿色组织区段位于3′位置时,叶表达增加。位置也影响了外壳表达,但有一些相反的作用。表9:转基因玉米中杂合调节元件的相对定向区段的表达在另一个实例中,表7中使用的须区段的两个拷贝(来自具有seqidno:60的亲本调节元件)位于也在表7中的绿色组织区段的两个拷贝(来自具有seqidno:49的亲本调节元件)的上游。在该构型中,叶表达和须表达相对于亲本增加约两倍(seqidno:53;表10)。然而,逆转该排列使得绿色组织区段的两个拷贝位于须区段的两个拷贝的上游,这导致无叶表达(seqidno:59;表10)。这些合并的结果说明提供tata盒的区段或3’区段可影响杂合调节元件提供的表达。表10:转基因玉米中杂合调节元件的相对定向区段的表达实例10:玉米的农杆菌介导的转化和转基因植物的再生对于本公开的玉米多核苷酸的农杆菌介导的转化,采用了zhao提出的方法(美国专利号5,981,840和pct专利公开wo98/32326)。简而言之,从玉米中分离未成熟的胚,并且在特定条件下将胚与农杆菌悬浮液接触,借此该细菌能够将本公开的调节元件序列转移到至少一个未成熟的胚的至少一个细胞中(步骤1:侵染步骤)。在该步骤中,将未成熟的胚浸泡在农杆菌悬浮液中,引发接种。使这些胚与农杆菌共培养一段时间(步骤2:共培养步骤)。在侵染步骤后,可以在固体培养基上培养未成熟的胚。在该共培养期之后,进行任选的“静息”步骤。在该静息步骤中,将这些胚在已知抑制农杆菌生长的至少一种抗生素的存在下孵育,而不添加植物转化子的选择剂(步骤3:静息步骤)。接着,在含有选择剂的培养基上培养经接种的胚,并且回收生长的经转化的愈伤组织(步骤4:选择步骤)。在转移到温室之前,从愈伤组织中再生小植株(步骤5:再生步骤)。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1