一种利用lncRNA来诊断和治疗前列腺肿瘤的方法与流程
本发明涉及分子检测技术领域,尤其涉及lncrna检测和治疗前列腺癌(pca)的方法。本发明提供了pcat3/pcat9-mir-203-snai2轴可作为pca的诊断和治疗靶点。
背景技术:
前列腺癌(pca)是世界上最常见的上皮癌之一,也是仅次于肺癌的第二大癌症死亡原因。前列腺癌的发生和进展是涉及多基因突变和基因调控异常的多步过程。越来越多的证据表明,特定基因调控异常,包括micrornas(mirnas)和长链非编码rna(lncrnas),是pca的发病和发展的主要原因。尽管如此,在pca中mirnas和lncrnas之间的交叉效应仍然不是很清楚。研究发生在pca中的mirna-lncrna的效应机制不仅可以提高我们对pca的认知,也将有助于确定治疗pca的靶点。
长链非编码rna(incrnas)是一类长度大于200的非编码rna,涉及多种生物学过程,包括肿瘤的发展和转移,以及在转录和翻译水平调节蛋白的表达,。据多篇文献报道,incrnas的异常表达可以指示pca的某些特定阶段,并在pca的发病和进展中起着重要的作用。pcat3,又称pca3,是一种最具特异性的前列腺癌诊断生物标志物,参与控制前列腺癌细胞的存活和调节雄性激素受体信号通路。pcat9,也被称为pcgemi,是由雄性激素控制的pca前列腺中高度特异性的incrnas之一。然而,pca3和pcat9在前列腺癌中迁移、血管生成以及与相关疾病中的作用,并没有被完全阐明。
mirnas是一种单链非编码rna,长度约为20-24个核苷酸,通过与蛋白质编码mrna的3'端未翻译区域(3'-utrs)结合,在转录后抑制目标基因的表达。毫无疑问,mirnas在调控各种生理和病理过程中起着重要的作用,包括肿瘤发生。越来越多的证据证明mirnas在pca发展中的关键作用。例如,肿瘤抑制因子mirna,包括let-7family,mir-200family,mir-203,mir-205,mir-15a/mir-16-1,mir-101,mir-99,mir-99,mir-146a,mir-146a,mir-330和mir-34a,mir-222,mir-222,mir-21,mir-141,mir-141,mir-18a和mir-125b等,已被证实为前列腺癌相关的mirna。据报道,mir-203在前列腺癌中被下调,并通过靶向作用于bm11、lasp1、zeb2和snai2(24-28)影响癌症的转移。尽管在pca中广泛地被认为是一种肿瘤抑制因子,但根据我们所研究的pca中mir-203的上游调控机制的数据,并没得到有充分印证。
作为snai/slug超级家族的一员,snai2与不同癌症的发病机制有关。snai2在pca中通过调节细胞周期蛋白d1促进前列腺癌移植瘤的生长。与此同时,pca中的snai2抑制了癌症干细胞的活性,减少了多种多功能基因的表达。一些研究也表明,mir-203/snai2体系在各种类型的癌症中发挥了广泛的作用。然而,在前列腺癌的形成和发展过程中lncrnas、mirnas和snai2之间的交叉效应却没有被报道过。重要的是,lncrnas被证实可以作为mirnas的分子海绵。例如,zhum.等人认为lncrnah19-mir-675可以通过靶向作用于tgfp1抑制前列腺癌转移。pcat9通过充当mir-770的海绵,刺激骨关节炎滑膜细胞增殖。在本研究中,我们证明pcat3和pcat9通过调节mir-203/snai2调节前列腺癌的肿瘤发生、迁移、血管生成和转移。这些发现为pca提供了一个潜在的诊断方法和治疗靶点。
技术实现要素:
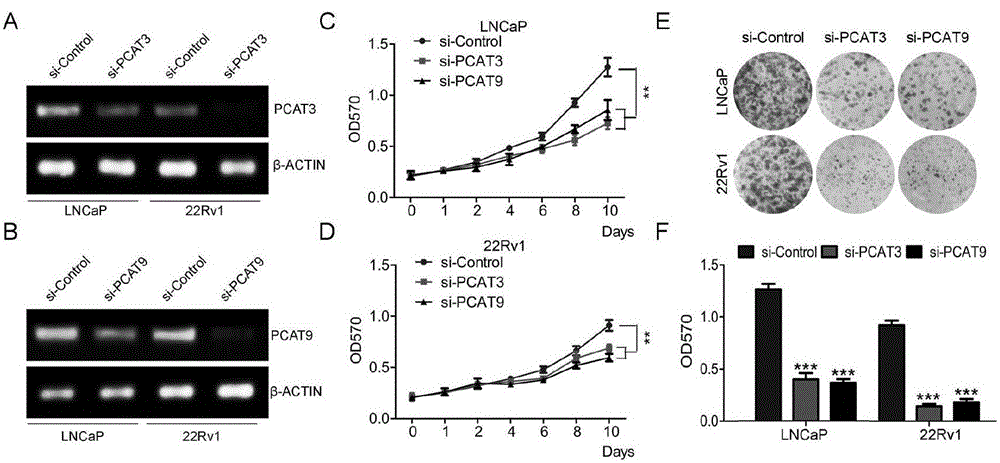
本发明确定了雄性激素依赖的incrnaspcat3和pcat9在pca中的潜在作用。我们使用的说雄性激素敏感型细胞lncap和22rv1。在进行平板克隆形成试验、mtt分析和transwell迁移试验之前使用pcat3或pcat9sirna对细胞转染48h。转染效果通过半定量rt-pcr(图1a和1b)分析。如图1c和1d所示,与阴性对照相比,pcat3或pcat9的功能丧失导致lncap和22rv1细胞增殖显著下降。在平板克隆形成试验中也观察到类似的结果,即pcat3或pcat9的沉默抑制了lncap和22rv1细胞的增值(图1e和1f)。此外,pcat3或pcat9的下调也显著降低了细胞的入侵和迁移(图2a-c)。这些发现表明pcat3和pcat9在前列腺癌的发生和进展中起到了促进肿瘤的作用。
肿瘤发展过程中,肿瘤引发的血管再生和癌症的干细胞性被认为是非常重要的。我们研究了在pca中下调内源性pcat3或pcat9的表达是否能调节内皮细胞的血管形成。lncap和22rv1细胞经对照sirna、pcat3sirna或pcat9sirna转染48h。收集条件细胞培养基收集并添加huvecs进行血管生成分析。如图3a和3b所示,与对照sirna相比,被pcat3或pcat9sirna转染的lncap和22rv1细胞在受到刺激时,血管生成明显减少。此外,sox2、oct4和nanog是在维持胚胎干细胞和癌症干细胞的多能性方面发挥关键作用的转录因子。westernblot检测显示pcat3或pcat9的沉默降低了lncap和22rv1细胞中oct4、sox2、nanog的蛋白水平,揭示了pcat3和pcat9可以调节pca中肿瘤干细胞的性质(图3c和3d)。这些数据进一步证实了pcat3和pcat9在pca中的关键作用。
本发明探讨pcat3和pcat9在前列腺癌发生和进展中的调控机制。我们使用在线软件starbasev2.0来识别可能与pcat3和pcat9交互的mirnas。starbasev2.0预测pcat3和pcat9都包含两个与mir-203s区结合的互补序列(图4a和4b)。同时,荧光素酶报告实验表明,mir-203模拟物减少pcat3或pcat9的mrna上标记mir-203质粒携带的荧光素酶活动的结合位点,但这个变化减少了变异(图4f),这表明mir-203可以作为海绵压制pca中的pcat3和pcat9。在我们之前的研究和其他文献中,转录因子snai2是mir-203在多种癌症中的直接靶点(图4g)。在此,我们也进行了荧光素酶的试验,以确定mir-203对lncap和22rv1细胞的snai2表达的影响。正如所料,荧光素酶实验表明,mir-203模拟物的荧光素酶活性抑制野生型snai23'-utr-wt,但对突变型snai23'-utr-mut没有影响(图4h和4i)。此外,pcat3和pcat9在含mir-203模拟物的转染细胞中的表达明显减少,qrt-pcr(图5a-d)和rna-fish(图5e)已证实了这一表达。此外,我们发现,与对照sirna相比,mir-203的表达水平在被pcat3或pcat9sirna转染的lncap和22rv1细胞中表达上调(图5f和5g)。同时,pcat3sirna和pcat9sirna抑制snai2蛋白水平(图5h和5i)。因此,我们总结出pcat3和pcat9在mir-203的作用下,通过调控snai2表达来调节前列腺癌的增殖和进展。
本发明中,pcat3和pcat9通过利用mir-203-snal2通路调控pca的肿瘤细胞增殖和进展。为了进一步测试pcat3(或pcat9),mir-203和snai2之间的联系,我们转染对照sirna,pcat3sirna,pcat9sirna,pcat3sirna+对照mirna抑制剂,pcat9sirna+对照mirna抑制剂,pcat3sirna+mir-203抑制剂,pcat9sirna+mir-203抑制剂,pcat3sirna+psin空慢病毒,pcat9sirna+psin空慢病毒,pcat3sirna+psin-snai2慢病毒或pcat9sirna+psin-snai2慢病毒于lncap和22rv1细胞中调查其对细胞增殖的影响。mtt分析表明,mir-203的抑制或snai2的过度表达能显著减缓pcat3sirna和pcat9sirna对细胞活力的抑制作用(图6a和6b)。此外,transwell迁移实验表明,mir-203的沉默或snai2的过表达可挽救被pcat3或pcat9sirna抑制lncap和22rv1细胞的迁移能力(图6c-e)。在f-actin染色中也有类似的观察,对照sirna,pcat3sirna+mir-203抑制剂,pcat9sirna+mir-203抑制剂,sirna+psin-snai2慢病毒和pcat3sirna+psin-snai2慢病毒组中lncap细胞形状比其他组的细胞更长(图6f)。
本发明中,pcat3/pcat9-mir-203-snai2轴参与了pca体内异种移植生长的调控。为了进一步描述pcat3/pcat9-mir-203-snai2轴在pca的体内功能,在balb/c裸鼠中建立了异种移植瘤模型。将被对照sirna,pcat3sirna,pcat9sirna,pcat3sirna+mir-203抑制剂,pcat9sirna+mir-203抑制剂,pcat3sirna+psin-snai2或pcat9sirna+psin-snai2转染的lncap细胞皮下注射到裸鼠后肢的上部区域。一旦肿瘤明显,每隔一天进行测量。如预期一样,被转染pcat3或pcat9sirna的小鼠比对照sirna组异种移植增长速度慢(图7a)。在实验结束时能明显观察到肿瘤的增大和体重的减少(p<0.01)(图7b和7c)。这些结果表明mir-203-snai2轴可能在pcat3/pcat9引导的前列腺肿瘤生长中具有重要意义。
综上所述,我们的发明系统地显示pcat3/pcat9-mir-203-snai2轴在前列腺癌肿瘤发生、迁移、血管生成以及干细胞化中起着关键作用(图8)。
附图说明
图1lncrnaspcat3或pcat9下调抑制前列腺癌lncap和22rv1细胞增殖。a和b:半定量rt-pcr检测被pcat3或pcat9sirna转染的lncap(a)和22rv1(b)细胞中pcat3或pcat9sirna的表达。c和d:采用mtt法测定被pcat3或pcat9sirna转染后lncap(c)和22rv1(d)细胞的相对细胞存活率和生长速率,结果表示为平均值±标准误差(n=6)。lncap和22rv1细胞中pcat3和pcat9的沉默,显著降低了平板克隆形成。菌落数量测定结果表示为均值±标准误差(f)。
图2pcat3或pcat9的沉默显著抑制了lncap和22rv1细胞的迁移。a.被对照sirna,pcat3sirna或pcat9sirna转染后,lncap和22rv1细胞的迁移能力的代表性图像。b和c:通过清除结晶紫后测定od570,对细胞迁移能力进行了测定。结果表示为数据平均值±标准误差(n=3)。
图3lncap和22rv1细胞中pcat3或pcat9的功能缺失降低了内皮细胞的血管形成,并抑制了干细胞化的相关因子。a和b:对添加了被对照sirna,pcat3sirna或pcat9sirna转染后的lncap和22rv1细胞的huvecs细胞进行血管形成试验(a)。对于血管生成分析来说,管长(b)用imagej软件以100像素为单位进行测量。c和d:westernblot测定被对照sirna,pcat3sirna或pcat9sirna转染后的lncap和22rv1细胞中nanog、sox2和oct4的表达。
图4pcat3和pcat9与mir-203在pca中的相互抑制。a、b和g:假定mir-203结合位点在pcat3(a)和pcat9(b)的mrna上,以及snai2(g)的3'-utr上。报告基团显示了野生型(wt)结合位点和突变型序列。c、d、e和f:mir-203模拟物转染抑制了pcat3或pcat9mrna结合位点上携带mir-203的报告质粒荧光素酶活性。h和i:采用含3'-utr区的snai2荧光素酶报告基团检测mir-203和snai23'-utr之间的相互作用。。
图5pcat3和pcat9均为mir-203-snai2轴的诱导物。a、b、c、d、e:使用qrt-pcr(a-d)和rna-fish(e)检测pcat3和pcat9在被mir-203模拟物转染的lncap和22rv1细胞中的表达。f和g:半定量rt-pcr检测被对照sirna、pcat3sirna或pcat9sirna转染的lncap(f)和22rv1(g)细胞中mir-203的表达。h和i:westernblot检测被对照sirna、pcat3sirna或pcat9sirna转染的lncap(h)和22rv1(i)细胞中snai2的表达。
图6pcat3/pcat9-mir-203-snai2调节轴调节前列腺癌细胞的增殖和迁移。a.mtt法测定lncap(a)和22rv1(b)在指示组中的细胞活力。c、d、e:指示组的细胞迁移能力(c),采用transwell迁移试验检测合格。f.使用免疫荧光法检测lncap细胞中的f-actin。
图7pcat3/pcat9-mir-203-snai2轴控制体内lncap细胞的异种移植生长。a.从对照sirna、pcat3sirna、pcat9sirna、pcat3sirna+mir-203抑制剂、pcat9sirna+mir-203抑制剂、pcat9sirna+psin-snai2和pcat9sirna+psin-snai2组lncap细胞移植肿瘤具有代表性的图片。b、c:肿瘤生长曲线(b)及lncap异种移植瘤体重(c)。肿瘤体积和重量表示为平均值±标准误差(n=4)。
图8前列腺癌中pcat3/pcat9-mir-203-snai2轴的表达及功能。
具体实施方式
1.细胞培养
两种前列腺癌细胞lncap和22rv1购买于americantypicalculturecenter(atcc;manassas,va,usa),使用含10%胎牛血清和1%青霉素/链霉素的rpmi1640培养基于5%co2培养箱中37℃培养。人类脐静脉内皮细胞(huvecs)购买于lifetechnologies(carlsbad,california,usa),根据说明书进行培养。
2.sirna转染
抗pcat3和pcat9的sirnas由riobiotechnlogy(广州,广东,中国)合成。sirna的序列如下:5'phos/rcrurargrcrarcrarcrargrcrarurgrarurcrarururarcgg(si-pcat3);5'phos/rgrurgrgrcrararcrargrgrcrarargrcrargrargrgrgraaa(si-pcat9)。sirnas的转染使用lipofectaminernaimax试剂(invitrogen,carlsbad,ca,usa)。
3.质粒构造和慢病毒产生
snai2的orf从du145细胞的cdna获得,然后克隆到spel和ecorl限制位点的psin/ef1α-ires-puro病毒表达载体上。lentivirus由psin表达载体、包装质粒pspax2和包膜质粒pmd2共转染的hek-293t细胞使用fugenehd(roche,basel,switzerland)产生。用于克隆的引物为:5’-ggactagtatgccgcgctccttcctggtc-3’(psin-snai2,forward);5’-cggaattctcagtgtgctacacagcagccagattc-3’(psin-snai2,reverse)。
4.mtt法测定细胞活力
细胞以2×103的密度播种于96-孔板中,经过一段时间的潜伏期后,每孔中添加20μl的mtt试剂(beyotime;5mg/ml),37℃培养4h。之后,吸去培养基每孔添加200μldmso(beyotime)轻轻摇晃10min,使用elisareader(moeculardevices,sunnyvaleca,usa)。
5.平板克隆形成
含有0.6%琼脂糖凝胶和10%fbs的基本培养基是添加到6孔板室温孵育1h。sirna转染40h后,细胞(5×103)重新悬浮于含0.3%琼脂糖凝胶和10%fbs的rpmi1640培养基中,接种于预处理的6孔板中,培养14d形成平板克隆。随后,使用pbs将其冲洗过两次,用70%的乙醇和0.1%的结晶紫染色,然后用解剖显微镜进行计数。数据进行三次平行,显示平均数±标准差。
6.半定量rt-pcr和qrt-pcr
半定量rt-pcr和qrt-pcr均按之前描述进行。引物序列如下:5’-agatttgtgtggctgcagc-3’(pcat3,forward),5’-tcctgcccatcctttaagg-3’(pcat3,reverse);5’-ggtaggcacgtggaggacta-3’(pcat9,forward),5’-tgcttttgtgggtttgttca-3’(pcat9,reverse);5’-atgaggaatctggctgctgt-3’(snai2,forward),5’-caggagaaaatgcctttgga-3’(snai2,reverse);5’-gtcgtatccagtgcagggtccgaggtattcgcactggatacgacctagtg-3’(specificrtprimerformir-203),5’-gtgcagggtccgaggt-3’(realtimepcr,mir-203,forward),5’-gccgcgtgaaatgtttagg-3’(realtimepcr,mir-203,reverse);5’-ctcgcttcggcagcaca-3’(realtimepcr,u6,forward),5"-aacgcttcacgaatttgcgt-3’(realtimepcr,u6,reverse)。
7.boyden小室侵袭模型
细胞(5×104)悬浮于200μ1含1%fbs的基本培养基中并接种于transwell上室中。下室为500μl含10%fbs的培养基。孵化24h后,迁移的细胞用4%多聚甲醛固定(beyotime)15min,用0.1%结晶紫染色15min,用显微镜(olympus)拍照。然后用200μ1的醋酸(33%,v/v)消除染色的结晶紫,并用elisareader(moeculardevices,sunnyvaleca,usa)在od570测量吸光值。
8.内皮细胞血管形成分析
huvecs以每孔1×105浓度接种含生长因子的培养基于24孔板中。在培养中培养8h后,用显微镜(olympus)拍照。血管的形成是通过imagej软件测量10×的累积管长度。
9.质粒构造和荧光素酶报告分析
扩增lncap细胞的cdna和基因组dna含mir-203结合位点的pcat3、pcat9和3'-utr的cdna片段,并克隆到psicheck-2的renilla荧光素酶基因下游的sgfl或pmel位点。这些结构之后根据说明书使用定点诱变系统(beyotime)进行突变。用于克隆和定向诱变的引物如下:5'-gaccgcgatcgcccttaaaggatgggcagg-3'(pcat3-bindingsite-1,forward),5'-cttagtttaaaccgtaatgatcatgctgtgtg-3'(pcat3-bindingsite-1,reverse);5'-gaccgcgatcgcatcatcacatgagacagcaa-3'(pcat3-bindingsite-2,forward),5'-cttagtttaaacgtggtgatacatcattggca-3'(pcat3-bindingsite-2,reverse);5'-gaccgcgatcgctactacgagatgcactggga-3'(pcat9-bindingsite,forward),5'-cttagtttaaacaatcatatttgcacacaacg-3'(pcat9-bindingsite,reverse);5'-gaccgcgatcgctgacaaataaagtccaaaggc-3'(snai2-3’-utr,forward),5'-cttagtttaaacaatcatgaagcaagtaaagtctc-3'(snai2-3’-utr,reverse);5'-cgttcataaccaaattgccctgtatttctaac-3'(pcat3-bindingsite-l-mut,forward),5'-cagggcaatttggttatgaacgcacagtttag-3'(pcat3-bindingsite-l-mut,reverse);5'-aaggtaacctttatctgccctgtggtgagtgc-3'(pcat3-bindingsite-2-mut,forward),5'-cagggcagataaaggttacctttggggatttg-3'(pcat3-bindingsite-2-mut,reverse);5'-ctgtctctgtaaaagtgccctgtatttacaag-3'(pcat9-bindingsite-l-mut,forward),5'-cagggcacttttacagagacagagaatttca-3'(pcat9-bindingsite-l-mut,reverse);5'-aaagagatcatgaggtgccctggagtgcactg-3'(pcat9-bindingsite-2-mut,forward),5'-cagggcacctcatgatctcttttccctctgc-3'(pcat9-bindingsite-2-mut,reverse);5'-ttttgctgccaatattgccctgatctgaaaag-3'(snai2-3’-utr-mut,forward),5'-cagggcaatattggcagcaaaaaaaaatgta-3'(snai2-3’-utr-mut,reverse)。
10.体内实验
所有动物实验都是在动物护理和使用中心的批准下进行的。实验中使用的是4-6周龄的雄性balb/c。lncap细胞密度达1×107重悬于含50%基质胶的pbs中,皮下注射的裸鼠右下肢,每组4只小鼠。有明显的肿瘤出现的第二天测量肿瘤体积。在注射后32d后,取出肿瘤称重拍照。
- 还没有人留言评论。精彩留言会获得点赞!