一种提高山羊产羔数的多基因聚合效应分析的育种方法与流程
本发明属于山羊分子生物技术和分子标记技术领域,特别涉及到一种提高山羊产羔数的多基因聚合效应分析的育种方法。本发明将所述的分子标记与山羊的产羔数进行聚合效应分析以应用到山羊产羔数性状的早期育种工作中。
背景技术
目前山羊产业的集约化程度不高,多为散养模式,且良种化程度低,生产力水平不高,经济效益很不理想。因此,养羊业健康、持续发展是当务之急。繁殖性状是重要的经济性状,对于种畜而言,繁殖力就是生产力,繁殖性能直接关系到肉羊产业的经济效应,因而繁殖力低是制约高效养羊业发展的主要瓶颈,而从遗传本质上研究提高繁殖力是最高效的方式。种羊的繁殖力属于多基因调控的数量性状,研究中,如果仅考虑单一基因或单一位点对繁殖力的影响,可能会导致统计结果较大程度的偏离实际结果,而同时将调控目的性状的主效基因一起进行统计学分析,可能会获得更准确的结果,从而实现对目的性状更可靠的选择。而本发明所采用的以分子辅助标记基因为基础的多基因聚合效应分析方法,就是同时将多个基因的目的基因座上表现优良的纯合基因型同时集中到同一个体中,从而培育出由超级经济性状的个体组成的新品种,这对优质高繁山羊新品种(系)的培育具有重要意义。
促性腺激素释放激素受体(gonadotropinreleasinghormonereceptor,gnrhr)基因是位于垂体促性腺细胞膜上的一种高亲和性g蛋白偶联受体(campionce,turzilloam,claycm.thegeneencodingtheovinegonadotropin-releasinghormone(gnrh)receptor:cloningandinitialcharacterization.[j].gene.1996,170(2):277-280.),可转导产生于下丘脑的gnrh的信号,调节垂体前叶合成与释放fsh和lh,从而促进性腺组织的生长、成熟和调控生物的繁殖性能。因此,gnrhr基因在调节哺乳动物繁殖力中起着关键作用,是繁殖性状的一个重要候选基因(储明星,肖杰文,狄冉,等.促性腺激素释放激素受体(gnrhr)基因多态性及其与济宁青山羊高繁殖力关系[j].农业生物技术学报.2009,17(2):218-223.)。近年来,国内外研究者对gnrhr基因的多态性进行了一定的研究,孙洁等(孙洁,储明星,陈宏权,等.gnrhr基因多态性及其与小尾寒羊高繁殖力关系[j].农业生物技术学报.2008,16(2):230-236.)对gnrhr基因在几个绵羊群体中的多态性研究了研究,结果发现,山羊在外显子1片段中存在5个碱基突变,其中有3个引起了氨基酸的改变;在山羊和小尾寒羊的外显子3区域内,存在2个突变,均引起了氨基酸的改变,且小尾寒羊的突变纯合型的平均产羔数较之野生型多0.81只,差异极显著(p<0.01),由此推测,gnrhr基因可能是影响小尾寒羊繁殖力的主效基因。
促卵泡素受体(folliclestimulatinghormonereceptor,fshr)基因属于g蛋白偶联受体超家族中的糖蛋白。魏伍川等(魏五川,许尚忠.牛fshr基因5′端侧翼序列的分析和多态性研究[j].畜牧兽医学报.2002,33(5):417-423.)在fshr基因5′端侧翼区发现了多态位点,并且存在该多态性的牛,其排卵率和双胎率均比其它牛要高,而在小尾寒羊中不存在该多态位点。宁立娜[10]对冀中山羊fshr基因的5′端侧翼区进行了克隆、测序及pcr-rflp分析,结果发现其所研究的山羊群体均表现为bb型,推测bb基因型可能与哺乳动物的高产仔数呈正相关。朱吉等[11]在湘东黑山羊、南江黄羊和贵州黑山羊3个山羊品种中,研究了fshr基因5′端侧翼区的多态性和产羔数的相关性,结果发现,fshr基因对贵州黑山羊的产羔数具有显著影响(p<0.05),推断fshr基因可能是影响山羊繁殖性能的候选基因。
骨形态发生蛋白6(bonemorphogeneticprotein6,bmp6)基因的cdna序列最先是由lyons等(lyonskm,peltonrw,hoganbl.patternsofexpressionofmurinevgr-1andbmp-2arnasuggestthattransforminggrowthfactor-beta-likegenescoordinatelyregulateaspectsofembryonicdevelopment[j].genes&development.1989,3(11):1657.)在小鼠的胚胎中分离出来的。近年来的研究发现,bmp6基因在体内通过抑制或促进相关酶类或生殖激素的活性而直接参与调控与繁殖相关激素的合成与分泌,从而影响繁殖器官的形成与发育,例如bmp6基因可通过自分泌或旁分泌途径,刺激fsh的合成(juengeljl,readerkl,bibbyah,etal.theroleofbonemorphogeneticproteins2,4,6and7duringovarianfolliculardevelopmentinsheep:contrasttorat[j].reproduction.2006,131(3):501.)。
通过以上材料我们可以得出上述3个基因可能参与调控动物的相关繁殖过程并发挥着重要作用。而山羊的产羔数性状受微效多基因控制,且呈加性-显性基因作用模式,目前,国内外学者已对影响绵羊产羔数众多基因的单基因效应进行了大量的分析,但在山羊方面的研究还较少,且也未对多个候选基因之间的多基因聚合效应进行深入研究。
对于繁殖性状的基因调控,目前主要有两种主流观点,一种认为繁殖性状是由某一个或几个主效基因调控,同时由多个微效基因协同调控;另一观点认为繁殖性状是由多个基因共同调控的数量性状。对于这两个观点,多数研究者对第二个观点持肯定态度。目前已报道的除了这3个基因外,bmpr-ib、bmp15、gdf9、bmp4和bmpr-ia等基因也影响动物的繁殖性状,但目前尚未有研究证实哪一个基因为主效基因。
另外,关于繁殖性状的分子标记国内外研究很多但涉及多基因聚合效应的研究很少,而生物机体的某一性状或功能都是由不同层次的多个基因进行调控的,属于一个由多个基因及其产物组成的调控网络。因此,单个遗传标记效应相对较少,往往不能直接进行生产实际的应用,而多基因聚合是打开这一复杂性状调控网络的一个重要步骤。
为此我们结合了影响山羊产羔数的多个重要基因,运用多基因聚合效应分析模型,将显著影响山羊产羔数性状的多态位点的优势基因型聚合到同一个基因组中,以提高选育的准确性和可靠性,从而选育出具有优良产羔数性状的种母羊,进而快速高繁力山羊新品系的培育进程。
技术实现要素:
本发明的目的在于克服现有技术对产羔数性状遗传选育的缺陷和不足,提供一种影响山羊产羔数性状的多基因聚合分子标记以及提高山羊产羔数性状的多基因聚合育种方法,以克服常规育种技术耗时长、育种效率低和单个基因分子标记对于复杂经济性状的选择效果差的不足。
本发明的目的通过下述技术方案得以实现:
一方面,本发明提供了一种影响山羊产羔数性状的多基因聚合分子标记,所述多基因聚合分子标记包括gnrhr、fshr和bmp6基因的分子标记。在这3个基因中各存在1个多态位点,它们所对应的核苷酸序列如seqidno.1-3所示,其中序列中的m是g或a;n是c或t;l是t或c,它们的差异导致了山羊产羔数性状的不同。
发明人采用大量的山羊样本进行了基因型检测,利用统计分析软件进行单基因和多基因效应分析。结果显示上述3个基因的单基因效应对产羔数性状均具有显著的影响,而多基因效应对产羔数性状也有显著的影响,且多基因效应远优于单基因效应,表明通过多基因聚合效应分析的育种方法能够大大提高母羊的产羔数性状,说明本发明的实用性较好。
其中gnrhr基因的分子标记如seqidno.1所示:
tcttgaagctgtatcagccataaaggatggcaaacggtgactctcctgatcagaatgaaaaccactgttcagcgatcaacagcagcatcctactaacaccm(g/a)ggcagcctccccaccctgaccctatctggaaagatccgagtgactgttactttcttcctttttctactctccacaattttcaacac
在该序列的第101位的m为g或a,该突变导致在山羊群体中出现产羔数性状的多态性;
其中,fshr基因的分子标记如seqidno.2所示:
taccagctcccaacgcagacctcttctcaaaagggctcagtgtggagcctctgaaatctgggcaggattgtgtctgcagaagcagaagcaagcaggtggatggataagtaaacatggcn(c/t)ttgttcctggtggccttgctcgcattcctgagcttgggctcaggatgccaccatcgactctgtc
在该序列的第119位的n为c或t,该突变导致在山羊群体中出现产羔数性状的多态性;
其中,bmp6基因的分子标记如seqidno.3所示:
tgtgccctcacctgctgtctcctctctcttccagagactcggacctgtttctgctgggcacgcgtgctgtgtgggcctcagaggcgggctggctggagtttgacatcacggccaccagcaacctgtgggtcctgactccgcagcacaacatggggctgcagctgagcgtggttacgcgl(c/t)gacggtgagttcagggactcgggcctttgcctccttggcggcaggtgtgggcagccgtggtccccaggcgggccccccgtttcccgacctcctggagccttggtggacaagcagagtcctgccccagggcctcctggcaaggactcagggacggttagatggtccgggagcctcagagaggggtaggagctgggtgctgagttaaagaggaaggccggg
在该序列的第179位的l为t或c,该突变导致在山羊群体中出现产羔数性状的多态性。
另一方面,本发明还提供了一种用于检测gnrhr、fshr和bmp6基因聚合分子标记的引物对。
其中,检测gnrhr基因的引物对如seqidno.4和seqidno.5所示;
seqidno.4:
gnrhr的上游引物pcr-f:5'-tcttgaagctgtatcagccata-3';
seqidno.5:
gnrhr的下游引物pcr-r:5'-gtgttgaaaattgtggagagtaga-3'。
其中,检测fshr基因的引物对如seqidno.6和seqidno.7所示;
seqidno.6:
fshr的上游引物pcr-f:5'-taccagctcccaacgcagac-3';
seqidno.7:
fshr的下游引物pcr-r:5'-gacagagtcgatggtggcat-3'。
其中,检测bmp6基因的引物对如seqidno.8和seqidno.9所示。
seqidno.8:
bmp6的上游引物pcr-f:5'-tgtgccctcacctgctgtctc-3';
seqidno.9:
bmp6的下游引物pcr-r:5'-cccggccttcctctttaactc-3'。
另一方面,本发明还提供了一种试剂盒,其含有seqidno.4和seqidno.5,seqidno.6和seqidno.7,以及seqidno.8和seqidno.9所示的引物对。
另一方面,本发明还提供上述的多基因聚合分子标记或引物对或试剂盒在山羊产羔性状检测或山羊育种中的应用。
另一方面,本发明还提供了一种山羊的遗传育种改良方法,该方法为确定山羊如上述的分子标记,并对seqidno.1、seqidno.2和seqidno.3所述的3个分子标记同时进行聚合效应分析,采用gnrhr-fshr-bmp6多基因聚合效应分析模型,选育aattcc型个体,以逐代提高山羊的产羔数性能。
具体地,所述的方法包括以下步骤:
(1)提取山羊的基因组dna;
(2)采用seqidno.4和seqidno.5,seqidno.6和seqidno.7,以及seqidno.8和seqidno.9所示的引物对分别对gnrhr、fshr和bmp6基因进行pcr扩增和基因分型;
(3)根据步骤(2)的基因分型结果,选育gnrhr基因如seqidno.1所述的分子标记第101位为aa型,fshr基因如seqidno.2所述的分子标记第179位为tt型,以及bmp6基因如seqidno.3所述的分子标记第101位为cc型的个体作为种羊。
作为优选的实施方式,步骤(2)中,基因分型采用高分辨率熔解曲线(hrm)技术或限制性内切酶酶切反应(rflp)技术。除了这两种基因分型技术,目前用于基因分型的方法主要有:限制性片段长度多态性(restrictionfragmentlengthpolymorphism,rflp)、末端限制性长度多态性(terminalrestrictionfragmentlengthpolymorphism,t-rflp)、扩增片段长度多态性(amplifiedfragmentlengthpolymorphisms,aflp)、多重连接探针扩增(multiplexligation-dependentprobeamplification,mlpa)、高分辨率溶解曲线(high-resolutionmelting,hrm)和单链构象多态性(single-strandconformationpolymorphism,sscp)以及测序技术等等,均可以用于本发明以实现基因分型。
聚合酶链式反应-限制性内切酶酶切反应(pcr-rflp)技术是在pcr技术的基础上发展起来的。dna碱基置换正好发生在某种限制性内切酶的识别位点上,使酶切位点增加或者消失,利用这一酶切性质的改变,pcr特异扩增包含碱基置换的这段dna,经某一限制酶切割,再利用琼脂糖凝胶电泳分离酶切产物,与正常pcr产物进行比较来确定是否变异。
在本发明两个示范性的实施例中,发明人以山羊血液基因组dna为模板,pcr扩增gnrhr和bmp6基因的特异性片段产物,通过在线软件nebcutterv2.0(http://nc2.neb.com/nebcutter2/)对酶切位点进行分析,确定gnrhr和bmp6基因的特异性片段产物分别含有mspⅰ(c↓cgg)和hhai(gcg↓c)限制性内切酶识别位点,分别经mspⅰ和hhai酶切后,以3%的琼脂糖凝胶电泳检测等位片段的长度,进而判定每个样品的基因型,并以此为遗传标记进行相关联分析,从而判断此分子标记是否影响山羊的产羔数。
高分辨熔解曲线(hrm)技术的主要原理是基于核酸分子物理性质的不同,通过检测dna双链序列之间的解链温度差异来判断序列之间的差异。hrm利用特定的染料(lcgreen、evagreen等)可以插入dna双链中的特性,再通过高精度pcr仪的高分辨率及灵敏度实时监测温度变化过程中双链dna荧光染料与pcr扩增产物的结合情况,收集荧光信号,记录高分辨率熔解曲线,从而实现对dna序列差异的分析。hrm的精度可达到检测出单个碱基的差异特性,所以hrm分析技术成为近年来进行基因分型的一种新型工具。
在本发明的一个示范性的实施例中,以山羊血液基因组dna为模板,pcr扩增fshr基因的特异性片段产物,并通过hrm技术记录的高分辨率溶解曲线,检测出不同dna样本间fshr基因的单个碱基差异,从而对每个样本进行基因判型,并以此为遗传标记进行相关联分析,从而判断此分子标记是否影响重庆黑山羊的产羔数。
本发明中,作为优选的实施方式,所述的山羊为重庆黑山羊。本发明相对于现有技术具有如下的优点及效果:
(1)本发明研究并确定影响山羊产羔数相关的分子标记,使用这些分子标记对山羊的产羔数性状进行标记辅助选择,能够极大地增加山羊的产羔数,加快育种进程。
(2)本发明提供了一种用于鉴定影响山羊产羔数性状分子标记的引物对,通过这些分子标记及引物对,可建立高效准确的分子标记辅助选择育种技术,将其应用于山羊繁殖性状的遗传改良中,从而提高山羊的产羔数,挖掘山羊的繁殖潜力,进而提高企业肉羊养殖的经济效益,增加其核心竞争力。
(3)本发明通过多基因聚合效应分析,优选最佳组合基因型作为分子标记,能够增加山羊产羔数性状的遗传进展,减少山羊繁殖性状的育种时间,从而有效提高种羊育种的经济效益,其中,通过该聚合育种方法,本发明将优选aattcc组合基因型的个体,则每只羊的平均产羔数较gactcc组合基因型可增加0.67只,由此可见,优良的繁殖性能在养羊产业中发挥着巨大的收益潜力。
(4)本研究应用多基因聚合效应分析模型对山羊的繁殖性状进行遗传研究,不仅能够丰富山羊繁殖性状的研究方法,而且可以为繁殖性状的遗传效应研究、与分子标记的挖掘和分子标记辅助选择奠定理论基础。
附图说明
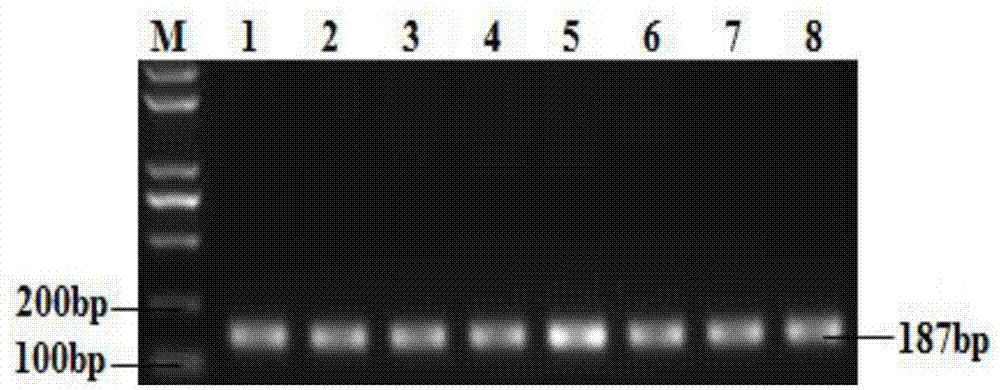
图1:是本发明中山羊gnrhr基因的pcr扩增产物凝胶电泳图。
图2:是本发明中山羊fshr基因的pcr扩增产物凝胶电泳图。
图3:是本发明中山羊bmp6基因的pcr扩增产物凝胶电泳图。
图4:是本发明中山羊gnrhr基因mspⅰ–rflp的三种基因型(gg,ga,aa)的凝胶电泳图。
图5:是本发明中山羊fshr基因三种基因型(cc,ct,tt)的高分辨率溶解曲线图。
图6:是本发明中山羊bmp6基因hhai–rflp的三种基因型(tt,tc,cc)的凝胶电泳图。
图7:是本发明中山羊gnrhr基因的gg基因型的测序峰图。
图8:是本发明中山羊gnrhr基因的ga基因型的测序峰图。
图9:是本发明中山羊gnrhr基因的aa基因型的测序峰图。
图10:是本发明中山羊fshr基因的cc基因型的测序峰图。
图11:是本发明中山羊fshr基因的ct基因型的测序峰图。
图12:是本发明中山羊fshr基因的tt基因型的测序峰图。
图13:是本发明中山羊bmp6基因的tt基因型的测序峰图。
图14:是本发明中山羊bmp6基因的tc基因型的测序峰图。
图15:是本发明中山羊bmp6基因的cc基因型的测序峰图。
具体实施方式
下面结合实施例及附图对本发明作进一步详细的阐述,但本发明的实施方式不限于此。
基本实验方法:
(1)实验样本:选取有完整产羔记录的重庆黑山羊母羊91只,颈静脉抽血5ml并做抗凝处理,于-20℃保存备用。本研究所使用的重庆黑山羊均来自广东省云浮市温氏新旺羊业有限公司的簕竹种羊场,该场全年实行高床舍饲饲养,按统一饲养标准饲喂日粮,自由采食饮水。
(2)dna提取:庆黑山羊血液基因组dna的抽提是参照血液基因组dna提取试剂盒(购自天根生化科技有限公司),以1%的琼脂糖凝胶电泳检测,无弥散或拖尾现象;并用紫外分光光度计对抽提的dna进行质量检测和浓度测定。将a260/280比值在1.8~2.0,a260/230比值在1.7~1.9的dna样判为合格。最后将合格的dna样品统一稀释成50ng/μl,并于-20℃冰箱内进行低温保存。
(3)dnapool测序:随机抽取30个不同重庆黑山羊个体的dna样品,按等量混合的原则做成两个dna池,并以此为模板进行pcr扩增,并对pcr产物进行测序,核酸序列如seqidno.1-3所述。利用dnastar软件对测序结果进行序列整理并与ncbi上基因组序列进行比对,发现在序列seqidno.1的第101bp处存在g→a突变;在序列seqidno.2的第119bp处存在c→t突变;在序列seqidno.3的第179bp处存在t→c突变.
实施例1gnrhr基因的pcr-rflp检测方法的建立
(1)引物设计
根据山羊gnrhr基因序列(ncbi上的geneid:100860755),利用primer5.0软件设计一对引物,序列如下:
gnrhr:
seqidno.4:上游引物pcr-f:5'-tcttgaagctgtatcagccata-3';
seqidno.5:下游引物pcr-r:5'-gtgttgaaaattgtggagagtaga-3'。
(2)pcr扩增体系和程序设置
pcr反应体系:10μl体系中含基因组dna模板1μl,上、下游引物各0.2μl,5μltaqdna聚合酶,以去离子双蒸水补充至终体积。
pcr反应程序:94℃预变性2min,35个循环(94℃变性30s,56℃退火30s,72℃延伸30s),72℃后延伸5min,4℃保存。
pcr反应产物用2%琼脂糖凝胶电泳检测,得到187bp的特异性扩增片段(如图1)。
(3)pcr-rflp基因分型
pcr反应结束后即进行不同snps位点扩增产物的酶切。酶切体系:15μl体系中含10×buffer1μl,双蒸水8.5μl,限制性内切酶mspⅰ(10u/μl)0.5μl,pcr反应产物5μl,震荡混匀,37℃水浴30min。酶切产物经3%的琼脂糖凝胶电泳检测,得到三种基因型:gg(99bp、88bp),ga(187bp、99bp和88bp),aa(187bp)(如图4)。
(4)不同基因型测序结果判定:序列测序在深圳华大基因科技有限公司进行,基因片段进行正反两个方向的测序。将gnrhr多态位点不同基因型的pcr产物送测,测序峰图的结果如图7~9所示。
测序结果如下所示:
seqidno.1:
tcttgaagctgtatcagccataaaggatggcaaacggtgactctcctgatcagaatgaaaaccactgttcagcgatcaacagcagcatcctactaacaccm(g/a)
ggcagcctccccaccctgaccctatctggaaagatccgagtgactgttactttcttcctttttctactctccacaattttcaacac
注:序列表中标注的m处为突变位点(括号中为突变碱基,为等位基因突变),在序列的首尾加粗显示的为引物序列。
(5)山羊gnrhr基因与产羔数性状的关联分析
试验共检测了91只重庆黑山羊母羊,通过上述操作确定每个个体的基因型,并将不同基因型与产羔数进行关联分析。数据采用spss20.0统计分析软件中的glm程序配合构建的单位点效应模型对所述分子标记位点进行统计分析,采用lsd和dunnett’st3法进行均数间的多重比较,研究胎次和基因型对多胎性状(产羔数)的影响,建立如下所示线性模型:
yijk=μ+gi+bj+pk+εijk(模型1)
式中,yijk是性状的观察值;μ为总体平均值;gi为i标记的基因型效应;bj为品种效应(在合并群体中需考虑此效应影响);pk为第k个胎次的固定效应;εijk为随机残差效应。所有数据结果均以平均值±标准差(mean±sd)的形式呈现。
由表1可知,gnrhr基因mspⅰ–rflp酶切位点的多态性对重庆黑山羊产羔数有显著影响(p<0.05),在胎次3和平均产羔数方面,aa型个体的产羔数均显著高于gg型和ga型个体(p<0.05),aa型为优势等位基因型。
表1重庆黑山羊gnrhr基因的多态性对产羔数的影响
注:同列中标有不同小写字母的数据间差异显著(p<0.05),不同大写字母的数据间差异极显著(p<0.01),无肩标或肩标字母相同的数据间差异不显著(p>0.05)。
实施例2fshr基因的pcr-hrm检测方法的建立
(1)高分辨率熔解曲线(hrm)的引物设计
根据山羊fshr基因序列(ncbi上的geneid:100861291),利用primer5.0软件设计一对引物,序列如下:
fshr:
seqidno.6:上游引物pcr-f:5'-taccagctcccaacgcagac-3';
seqidno.7:下游引物pcr-r:5'-gacagagtcgatggtggcat-3'。
(2)高分辨率熔解曲线(hrm)反应条件
使用55-65℃温度梯度pcr以了解引物的最适退火温度从而得到特异的pcr产物,以提高高分辨率熔解的灵敏度。
pcr反应体系(10μl):dna模板1μl,上、下游引物各0.3μl,
pcr反应程序为:94℃预变性2min,40个循环(94℃变性10s,60℃退火10s,72℃延伸5s)。
hrm反应程序为:95℃15s,55℃15s,55℃-95℃(0.1℃/s的温度上升速度),持续监测荧光变化,采集数据,获得熔解曲线,整个过程大概10分钟。应用ecotmreal-timepcrsystemsoftwarev3.0.16.0软件分析结果。
(3)pcr-hrm基因分型
用ecotmreal-timepcrsystemsoftwarev3.0.16.0软件分析后,统计每个样本的基因型,共存在三种基因型:cc,ct,tt(如图5)。
(4)不同基因型测序结果判定:序列测序在深圳华大基因科技有限公司进行,基因片段进行正反两个方向的测序。将fshr多态位点不同基因型的pcr产物送测,测序峰图的结果如图10~12所示。
测序结果如下所示:
seqidno.2:
taccagctcccaacgcagacctcttctcaaaagggctcagtgtggagcctctgaaatctgggcaggattgtgtctgcagaagcagaagcaagcaggtggatggataagtaaacatggcn(c/t)
ttgttcctggtggccttgctcgcattcctgagcttgggctcaggatgccaccatcgactctgtc
注:序列表中标注的n为突变位点(括号中为突变碱基,为等位基因突变),在该序列的首尾加粗显示的为引物序列。
(5)山羊fshr基因与产羔数性状的关联分析
试验共检测了91只重庆黑山羊母羊,通过上述操作确定每个个体的基因型,并将不同基因型与产羔数进行关联分析。数据采用spss20.0统计分析软件中的glm程序配合构建的单位点效应模型对所述分子标记位点进行统计分析,采用lsd和dunnett’st3法进行均数间的多重比较,研究胎次和基因型对多胎性状(产羔数)的影响,建立如下所示线性模型:
yijk=μ+gi+bj+pk+εijk(模型1)
式中,yijk是性状的观察值;μ为总体平均值;gi为i标记的基因型效应;bj为品种效应(在合并群体中需考虑此效应影响);pk为第k个胎次的固定效应;εijk为随机残差效应。所有数据结果均以平均值±标准差(mean±sd)的形式呈现。
由表2可知,fshr基因pcr–hrm多态位点对重庆黑山羊产羔数有极显著影响(p<0.01),在胎次3、在胎次4和平均产羔数方面,tt型个体的产羔数均极显著高于ct型和cc型个体(p<0.01)。
表2重庆黑山羊fshr基因的多态性对产羔数的影响
注:同列中标有不同小写字母的数据间差异显著(p<0.05),不同大写字母的数据间差异极显著(p<0.01),无肩标或肩标字母相同的数据间差异不显著(p>0.05)。
实施例3bmp6基因的pcr-rflp检测方法的建立
(1)引物设计
根据山羊bmp6基因序列(ncbi上的geneid:100860793),利用primer5.0软件设计一对引物,序列如下:
bmp6:
seqidno.8:上游引物pcr-f:5'-tgtgccctcacctgctgtctc-3';
seqidno.9:下游引物pcr-r:5'-cccggccttcctctttaactc-3'。
(2)pcr扩增体系和程序设置
pcr反应体系:10μl体系中含基因组dna模板1μl,上、下游引物各0.2μl,5μltaqdna聚合酶,以去离子双蒸水补充至终体积。
pcr反应程序:94℃预变性2min,35个循环(94℃变性30s,59℃退火30s,72℃延伸30s),72℃后延伸5min,4℃保存。
pcr反应产物用2%琼脂糖凝胶电泳检测,得到398bp的特异性扩增片段(如图3)。
(3)pcr-rflp基因分型
pcr反应结束后即进行不同snps位点扩增产物的酶切。酶切体系:15μl体系中含10×buffer1μl,双蒸水8.5μl,限制性内切酶hhai(10u/μl)0.5μl,pcr反应产物5μl,震荡混匀,37℃水浴30min。酶切产物经3%的琼脂糖凝胶电泳检测,得到三种基因型:tt(398bp),tc(398bp、220bp和178bp),cc(220bp、178bp)(如图6)。
(4)不同基因型测序结果判定:序列测序在深圳华大基因科技有限公司进行,基因片段进行正反两个方向的测序。将bmp6多态位点不同基因型的pcr产物送测,测序峰图的结果如图13~15所示。
测序结果如下所示:
seqidno.3:
tgtgccctcacctgctgtctcctctctcttccagagactcggacctgtttctgctgggcacgcgtgctgtgtgggcctcagaggcgggctggctggagtttgacatcacggccaccagcaacctgtgggtcctgactccgcagcacaacatggggctgcagctgagcgtggttacgcgl(c/t)
gacggtgagttcagggactcgggcctttgcctccttggcggcaggtgtgggcagccgtggtccccaggcgggccccccgtttcccgacctcctggagccttggtggacaagcagagtcctgccccagggcctcctggcaaggactcagggacggttagatggtccgggagcctcagagaggggtaggagctgggtgctgagttaaagaggaaggccggg
注:序列表中标注的l为突变位点(括号中为突变碱基,为等位基因突变),在该序列的首尾加粗显示的为引物序列。
(5)山羊bmp6基因与产羔数性状的关联分析
试验共检测了91只重庆黑山羊母羊,通过上述操作确定每个个体的基因型,并将不同基因型与产羔数进行关联分析。数据采用spss20.0统计分析软件中的glm程序配合构建的单位点效应模型对所述分子标记位点进行统计分析,采用lsd和dunnett’st3法进行均数间的多重比较,研究胎次和基因型对多胎性状(产羔数)的影响,建立如下所示线性模型:
yijk=μ+gi+bj+pk+εijk(模型1)
式中,yijk是性状的观察值;μ为总体平均值;gi为i标记的基因型效应;bj为品种效应(在合并群体中需考虑此效应影响);pk为第k个胎次的固定效应;εijk为随机残差效应。所有数据结果均以平均值±标准差(mean±sd)的形式呈现。
由表3可知,bmp6基因hhai–rflp酶切位点的多态性对重庆黑山羊产羔数有极显著影响(p<0.01),在平均产羔数方面,cc型和ct型个体的产羔数均极显著高于tt型个体(p<0.01)。
表3重庆黑山羊bmp6基因的多态性对产羔数的影响
注:同列中标有不同小写字母的数据间差异显著(p<0.05),不同大写字母的数据间差异极显著(p<0.01),无肩标或肩标字母相同的数据间差异不显著(p>0.05)。
实施例4多基因聚合效应分析模型的建立
首先运用实施例1和2和3中所述方法分别检测出gnrhr、fshr和bmp6基因的多态位点,并与重庆黑山羊的产羔数进行单基因效应分析,上述结果显示这3个基因均对重庆黑山羊的产羔数具有显著影响。为研究上述3个基因之间的聚合效应以及在育种工作中进一步提高对山羊产羔数性状的选育效率,本发明建立了山羊产羔数性状的多位点聚合效应分析模型。
(1)多基因聚合效应分析模型
试验共检测了91只重庆黑山羊,通过pcr-rflp和pcr-hrm方法确定了gnrhr-fshr-bmp6组合模型,并进行了组合基因型与产羔数的关联分析。建立模型如下:
yijk=μ+ci+bj+pk+εijk
其中,yijk是性状的观察值;μ为总体平均值;ci为第i个个体的组合基因型效应;bj为品种效应(在合并群体中需考虑此效应影响);pk为第k个胎次的固定效应;εijk为随机残差效应。
(2)多基因聚合效应分析在山羊产羔数性状关联分析中的应用,分析结果见表4。
由表4可知,gnrhr-fshr-bmp6组合模型对山羊的产羔数有显著影响(p<0.05),aactcc组合基因型的第1胎产羔数显著高于ggcctc、gacttc、aacttc和gactcc组合基因型;在胎次2中,aattcc和ggctcc组合基因型的产羔数均极显著高于aactcc组合基因型(p<0.01),同时又显著高于gacctc组合基因型(p<0.05);在胎次3和平均产羔数方面,aattcc组合基因型均极显著高于ggcctc、gacccc和gacctc组合基因型(p<0.01),且aattcc组合基因型在第2、3、4胎的产羔数以及平均产羔数均高于其它组合基因型,分别高达2.40只、2.80只、2.60只和2.42只,所以aattcc可视为最佳组合基因型。在育种工作中采用gnrhr-fshr-bmp6多基因聚合效应分析模型,对aattcc组合基因型的个体进行优选,则选育效果均高于其他组合基因型和单位点效应,这一研究结果将有助于进一步加快重庆黑山羊的育种进程。
表4多基因聚合效应对重庆黑山羊产羔数的影响
注:同列中标有不同字母的数据间差异显著(p<0.05),无肩标或肩标字母相同的数据间差异不显著(p>0.05);个体数小于10的组合基因型未在表中列出。
实施例5多基因聚合育种方法的效应分析
本发明提供一个能够显著增加重庆黑山羊产羔数的分子辅助育种方法,使用多基因聚合效应分析的育种方法对重庆黑山羊进行标记辅助选择,能够极大地增加山羊产羔数性状的育种进程。
本发明中影响重庆黑山羊产羔数的分子标记全部选育成aattcc组合基因型个体,则每只种母羊的平均产羔数较gactcc组合基因型的个体最多可以增加0.67只,由此可见对繁殖性能高的种母羊进行选育,可提高养羊业的经济效益并有助于加速我国肉羊产业的发展进程。
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、替代、简化、组合,均应为等效的置换方式,都包含在本发明的保护范围之内。
序列表
<110>华南农业大学
<120>一种提高山羊产羔数的多基因聚合效应分析的育种方法
<141>2018-06-27
<160>9
<170>siposequencelisting1.0
<210>1
<211>187
<212>dna
<213>caprahircusl
<400>1
tcttgaagctgtatcagccataaaggatggcaaacggtgactctcctgatcagaatgaaa60
accactgttcagcgatcaacagcagcatcctactaacaccmggcagcctccccaccctga120
ccctatctggaaagatccgagtgactgttactttcttcctttttctactctccacaattt180
tcaacac187
<210>2
<211>183
<212>dna
<213>caprahircusl
<400>2
taccagctcccaacgcagacctcttctcaaaagggctcagtgtggagcctctgaaatctg60
ggcaggattgtgtctgcagaagcagaagcaagcaggtggatggataagtaaacatggcnt120
tgttcctggtggccttgctcgcattcctgagcttgggctcaggatgccaccatcgactct180
gtc183
<210>3
<211>398
<212>dna/rna
<213>caprahircusl
<400>3
tgtgccctcacctgctgtctcctctctcttccagagactcggacctgtttctgctgggca60
cgcgtgctgtgtgggcctcagaggcgggctggctggagtttgacatcacggccaccagca120
acctgtgggtcctgactccgcagcacaacatggggctgcagctgagcgtggttacgcgmg180
acggtgagttcagggactcgggcctttgcctccttggcggcaggtgtgggcagccgtggt240
ccccaggcgggccccccgtttcccgacctcctggagccttggtggacaagcagagtcctg300
ccccagggcctcctggcaaggactcagggacggttagatggtccgggagcctcagagagg360
ggtaggagctgggtgctgagttaaagaggaaggccggg398
<210>4
<211>22
<212>dna
<213>artificialsequence
<400>4
tcttgaagctgtatcagccata22
<210>5
<211>24
<212>dna
<213>artificialsequence
<400>5
gtgttgaaaattgtggagagtaga24
<210>6
<211>20
<212>dna
<213>artificialsequence
<400>6
taccagctcccaacgcagac20
<210>7
<211>20
<212>dna
<213>artificialsequence
<400>7
gacagagtcgatggtggcat20
<210>9
<211>21
<212>dna
<213>artificialsequence
<400>9
tgtgccctcacctgctgtctc21
<210>9
<211>21
<212>dna
<213>artificialsequence
<400>9
cccggccttcctctttaactc21
- 还没有人留言评论。精彩留言会获得点赞!