一种具有阴离子转运活性的通道蛋白及其基因组修饰调控作物籽粒大小的制作方法
本发明涉及一种基因技术领域,特别地涉及一种具有阴离子转运活性的基因调控籽粒大小。
背景技术
水稻是一年生禾本科粮食作物,全世界近一半人口以稻米为食。提高水稻或其他禾本科作物的产量一直是研究者们关注的热点问题,粒型是与产量直接相关的性状,因而成为研究的直观切入点,其中主要包括粒长,粒宽,粒厚等表型影响籽粒重量,这些表型则是由细胞数量以及细胞形状、大小决定的。
近年来,通过数量性状定位(qtls)等方法,获得了决定籽粒大小的多个基因,主要涉及如下几个方面的生理过程,第一、有机物同化相关酶基因包括淀粉合成相关酶和蔗糖代谢相关酶如gbss、oscin1、gif1/oscin2等(tsaicy,1974,smithametal.,1997,guoyuzhangetal.,2011,ayahikoshomuraetal.,2008,tatsurohiroseetal.,2008,ertaowangetal.,2008)。第二类为糖的转运体及其调控基因如sweet4c、sutfamily、osnf-yb1等(davidesossoetal.,2015,ai-ningbaietal.,2015)。第三类为细胞分裂和激素有关的基因如gs5、gw2、gw8qsw5/gw5、bg1等(yibolietal.,2011,xian-junsongetal.,2007,shaokuiwangetal.,2012,ayahikoshomuraetal.,2008)。在这些已报道的影响粒型的分裂调控基因中,一部分是通过改变细胞数目影响籽粒大小,如gs3,gw2,gw5,gs5,gw8,qgl3,tgw6,gw6a,andbg1等(songetal.,2007,2015;wengetal.,2008;maoetal.,2010;lietal.,2011;wangetal.,2012;zhangetal.,2012;ishimaruetal.,2013;liuetal.,2015);也有一小部分细胞数目变化不大,细胞大小发生改变,如pgl1,gl7,andgs2/gl2等(heangandsassa,2012;cheetal.,2015;duanetal.,2015;huetal.,2015;wangetal.,2015)。其中,无论是影响细胞数目还是细胞大小,大多都是利用激素(ashikarietal.,1999,2005;hongetal.,2003;tanabeetal.,2005;ishimaruetal.,2013)来调控细胞分裂或细胞周期(yibolietal.,2011,xian-junsongetal.,2007,shaokuiwangetal.,2012,liuetal.,2015)从而影响籽粒大小。
技术实现要素:
针对现有技术中存在的技术问题,本发明提出了一种获得适于调整水稻籽粒大小的重组或编辑dna片段的方法,包括:通过基因工程技术手段改造水稻的dna片段,使得水稻或作物中tcbg1基因及其功能类似的同源基因敲除、敲低或基因沉默;其中tcbg1基因的cds序列如seqidno.1所示,氨基酸序列如seqidno.2所示。
如上所述的方法,其中,所述基因工程技术手段为crispr/cas9。
如上所述的方法,其中,crispr/cas9靶序列为如下所述的一个或多个:cgccgtcgccgccgcacgagagg;cctcggccgcctcggcgacctcc;ccgccatggaccccatctgcggg;cccagctgcctcaacacctgcc;gcgggcatggtgtcgatcgtggg;caaggccagacttgggatccagg。
如上所述的方法,其中,重组dna片段的cds序列如seqidno.3所示,氨基酸序列如seqidno.4所示。
如上所述的方法,其中所述基因工程手段为talen技术。
如上所述的方法,其中talen识别区序列为如下所述的一个或多个:ggccgtcaccaccgccttcctcggccgcctcggcgacctccagctcgccgccggc;cgccgtcctcaccggcctctgcgccgccatggaccccatctgcgggcaggcgc;cgcctccgccctggccctggcgctccacgtccccctcaccatgtggatggccagg。
如上所述的方法,其中,所述基因工程技术手段为rnai。
如上所述的方法,其中,rnai靶序列如seqidno.5所示。
如上任一所述的重组dna片段及其功能相似的同源基因。
一种植物基因组,包括如上所述的重组dna片段及其功能相似的同源基因。
一种植物细胞,包括如上所述的重组dna片段。
如上所述的重组dna片段、如上所述的植物基因组或如上所述的植物细胞在调整水稻或作物籽粒大小方面的应用。
一种调整水稻或作物籽粒大小的方法,包括:通过基因工程技术手段处理水稻的dna片段,使得水稻的dna片段经编辑后导致无法转录,致使tcbg1基因沉默;其中tcbg1基因的cds序列如seqidno.1所示,氨基酸序列如seqidno.2所示。
如上所述的重组dna片段、如上所述的植物基因组或如上所述的植物细胞在调整细胞膨压或渗透势方面的应用。
一种调整水稻或作物细胞膨压或渗透势的方法,包括:通过基因工程技术手段处理水稻的dna片段,使得水稻的dna片段经翻译后tcbg1基因沉默;其中tcbg1基因的cds序列如seqidno.1所示,氨基酸序列如seqidno.2所示。
细胞膨压或渗透势是促使细胞生长与伸长的重要因素。细胞膨压或渗透势可以维持细胞大小和形状。细胞内水分、糖类、有机酸、各种无机离子含量等维持了细胞的膨压或渗透势。如:细胞膜上的水通道蛋白(maurelc,etal.2015),糖转运体蛋白,氨基酸等有机物转运体(zifarellig,puschm,2010;zhangetal.,2016),无机离子通道(beauzamyetal.2014)都直接影响细胞的膨压或渗透势。然而,通过调控作物籽粒相关细胞的渗透势或细胞膨压,增加干物质积累的效率和籽粒大小,无论机理还是实例迄今为止尚无报道。
本发明的一些实例中,通过直接调控细胞渗透势或膨压,成功地改变籽粒细胞的库容量,增加了作物籽粒的大小。这无疑为作物的增产提供了基础,有着广泛的应用前景。
附图说明
下面,将结合附图对本发明的优选实施方式进行进一步详细的说明,其中:
图1是tcbg1基因的cds序列;
图2是tcbg1基因的氨基酸序列;
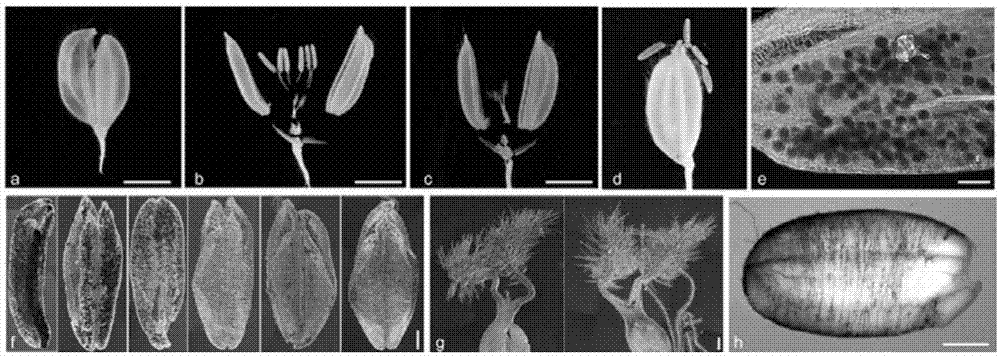
图3是tcbg1的表达模式分析;图中显示为protcbg1-gus转基因水稻中不同组织的gus染色;其中a-d是小穗壳,标尺=0.2mm;e和f是花粉囊,标尺=1mm;g是雌蕊,标准尺=0.5mm;h是种子,标准尺=1mm;
图4是sgrna::cas9诱导转基因水稻tcbg1突变体植株的原理图;图a为图解sgrna::cas9靶序列(除双划线部分)和相应的pams[protospaceradjacentmotifsequence(基序序列),为三个核苷酸ngg序列](双划线)的tcbg1基因,划线区域为一个ncoⅰ限制性酶切位点;图b为pcr/re检测转基因水稻植株中sgrna::cas9诱导的突变,对提取的独立转基因水稻幼苗的dna样本进行分析的结果;图c中红色短线表示删除,右侧数字表示突变的类型和涉及的核苷酸的数量;图d为第1,2和7号植株等位基因克隆的桑格法测序结果;
图5是利用转录激活因子效应蛋白核酸酶(transcriptionactivator-likeeffectornuclease,talen)技术诱导转基因水稻tcbg1突变体植株的原理图;图a为选取的靶序列在tcbg1中位置,其中双下划线部分碱基分别代表左侧靶序列和右侧靶序列;b中短线表示删除,右侧数字表示突变的类型和涉及的核苷酸的数量;
图6是利用rnai(rna干扰)技术沉默水稻tcbg1基因;图a为选取的rnai靶序列在tcbg1基因中的位置;图b为构建rnai转基因水稻的重组载体图谱;图c为rt-rcr(荧光定量pcr)方法鉴定转基因植株。
图7是tcbg1和wt糙米粒的比较;图a为tcbg1和wt糙米粒的大小及形态对比,标尺=1cm;图b为tcbg1和wt糙米粒的500粒覆盖面积统计结果;图c为tcbg1和wt糙米粒的千粒重统计结果;
图8是tcbg1和wt籽粒形态的比较;图a和c是wt、tcbg1-1和tcbg1-2的谷粒粒长比较及统计结果;图b和d是wt、tcbg1-1和tcbg1-2的谷粒粒宽比较及统计结果;图e是wt、tcbg1-1和tcbg1-2的谷粒粒厚比较及统计结果;图f和h是wt、tcbg1-1和tcbg1-2的糙米粒粒长比较及统计结果;图g和i是wt、tcbg1-1和tcbg1-2的糙米粒粒宽比较及统计结果;图j是wt、tcbg1-1和tcbg1-2的糙米粒粒厚比较及统计结果;其中图a、b、f和g的标尺均为1cm;
图9是tcbg1和wt灌浆速度比较;图a和图b分别是对比wt和tcbg1突变体受精以后干重及湿重的结果,其中n=20;以及
图10是利用超极化脉冲的典型全细胞电流记录系统获得的tcbg1氯离子外排特性;图a和图b中,向非洲爪蟾卵母细胞中注射不含有rna酶的水(作为对照),灌流液为nd96溶液;图c和图d分别以nd96(图c)和改变的nd96(图d)作为灌流液记录表达tcbg1的卵母细胞的电流,其中改变的nd96为将nacl替换为naglu2;图e和图f分别以上述的nd96(图e)和上述的改变的nd96(图f)作为灌流液记录表达tcbg1突变体tcbg1(删除251t)的卵母细胞的电流。
具体实施方式
为使本发明实施例的目的、技术方案和优点更加清楚,下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出tcbg1蛋白功能创造性结果前提下所获得的所有其他实施例,都属于本发明保护的范围。
在以下的详细描述中,可以参看作为本申请一部分用来说明本申请的特定实施例的各个说明书附图。在附图中,相似的附图标记在不同图式中描述大体上类似的组件。本申请的各个特定实施例在以下进行了足够详细的描述,使得具备本领域相关知识和技术的普通技术人员能够实施本申请的技术方案。应当理解,还可以利用其它实施例或者对本申请的实施例进行结构、逻辑或者tcbg1蛋白性质的改变。
本文中除非另外说明,核酸是以5′到3′方向从左往右书写。
在本发明中使用了很多术语。为了清楚一致的理解本发明,提供以下定义:
基因:是指表达功能性分子诸如但不限于特定蛋白质的核酸片段,包括位于编码序列之前(5′非编码序列)和之后(3′非编码序列)的调控序列。“天然基因”是指如自然界中存在那样具有其自身调控序列的基因。
基因组:组成生物基因组的总dna或整组基因,由染色体或染色体组携带。应用于植物细胞时,不仅涵盖细胞核内存在的染色体dna,也涵盖细胞的亚细胞组分(例如线粒体或者质粒)中存在的细胞器dna。
orf:开放阅读框(openreadingframe),从起始密码子开始,是dna序列中具有编码蛋白质潜能,一段无终止密码子打断的碱基序列。
突变基因:是已通过人工干预被改变的基因。这种突变基因的序列与相应的非突变基因的序列的不同之处在于包含至少一个核苷酸添加、缺失或置换。在本发明的某些实施方案中,突变基因包含由如本文所公开的crispr/cas内切核酸酶系统引起的改变。突变的植物为包含突变基因的植物。
靶向突变:是天然基因中的通过如下方式产生的突变:使用涉及本文所公开的或者本领域已知的能够在靶序列的dna中诱导双链改变的方法,对天然基因内的靶序列进行改变。
重组dna:是指在自然界中通常不会一起存在的核酸片段的组合。因此,重组dna可以包含源于不同来源的调控序列和编码序列,或源于相同来源但以不同于通常天然存在的方式排列的调控序列和编码序列。
crispr(clusteredregularlyinterspacedshortpalindromicrepeats)/cas(crispr相关):是经工程改造的核酸酶系统,为可用于基因组工程改造的细菌系统,是许多细菌和古菌的适应性免疫应答的一部分。
cas9:涉及一种cas蛋白编码的内切核酸酶cas9基因,能够将双链断裂引入dna靶序列中。其通常与侧翼crispr基因座偶联、相关或接近或在侧翼crispr基因座的附近。
限制性内切酶:一种在特殊核甘酸序列处水解双链dna的内切酶。ncoi是一种限制性内切酶,特异性识别并切割序列ccatgg。
靶位点,靶序列,目标序列:在本文可互换使用,是指在基因组的多核苷酸序列(包括叶绿体和线粒体dna)中被内切核酸酶基因特异识别的一段序列。例如,ccatgg为限制性内切酶ncoi的靶位点。
转化:将外源基因转移到植物基因组中,称为转化。涵盖可将核酸分子导入这种细胞中的所有技术。实例包括但不限于:用病毒载体转染;用质粒载体转化;电穿孔;脂质体转染;显微注射(mueller等人,(1978)cell15:579-85);农杆菌属介导的转移;直接dna摄入;whiskerstm介导的转化;以及微粒轰击。这些技术可用于植物细胞的稳定转化和瞬时转化二者。
talen:即transcriptionactivator-like(tal)effectornucleases,转录激活因子效应蛋白核酸酶。talen是一种可靶向修饰特异dna序列的酶,它借助于tal效应子——一种由植物细菌分泌的天然蛋白来识别特异性dna碱基对。tal效应子可被设计识别和结合所有的目的dna序列。对tal效应子附加一个核酸酶就生成了talen。tal效应核酸酶可与dna结合并在特异位点对dna链进行切割,从而导入新的遗传物质。
rnai:双链rna对基因表达的阻断作用被称为rna干预(rnainterference,rnai)双链rna经酶切后会形成很多小片段,称为sirna,这些小片段一旦与信使rna(mrna)中的同源序列互补结合,就会导致mrna失去功能,使该蛋白的表达水平降低,也就是使基因“沉默”了。
本文中增加,提高或增强是可互换的。例如产量或者籽粒大小等相比于野生型提高至少3%,4%,5%,6%,7%,8%,9%或10%或以上。
表型、表型性状:可指基因或系列基因的可观测表达。表型对于肉眼或本领域任何其它已知评估手段而言可以是可观测的,例如称重、计数、测量(长度、宽度、角度等)、显微法、生化分析等。在某些情况下,表型由单个基因或基因座直接控制,即“单基因性状”或“简单遗传性状”。在无较大水平的环境变化时,可在群体中分离单基因性状以给出“定性”或“离散”分布,即表型属于离散类。在其它情况下,表型是若干个基因的结果并且可被视为“多基因性状”或“复杂性状”。在群体中分离多基因性状以给出“定量的”或“连续”分布,即表型不可分为离散类。单一基因和多基因性状均可受到其表达所处的环境的影响,但是多基因性状与环境关系更紧密。
植株:可为整个植株、其任何部分、或来源于植株的细胞或组织培养物。因此,术语“植株”可指如下中的任何一种:整个植株、植株组分或器官(例如叶、茎、根等)、植株组织、种子、植株细胞、和/或它们的子代。植株细胞是从植株中获得的植株的细胞或来源于从植株中所获得的细胞的培养物。
本文中将具体描述水稻中单基因tcbg1的缺失或沉默影响植株灌浆,进而直接或间接影响产量。
一、目标基因的描述
本发明所针对的目标基因是tcbg1基因。图1所示即为seqidno.1所示的tcbg1基因cds序列。该基因的orf全长1440个碱基,gc含量达70.49%,无内含子。tcbg1基因的氨基酸序列如图2或者seqidno.2所示,该基因编码480个氨基酸,具有九个跨膜结构域,具备典型的mate家族二级结构特征。
tcbg1属于广泛存在于细菌,真菌,动植物中的dtx家族。根据进化关系可以把该家族分成四个亚家族,tcbg1属于第三个亚家族,具备典型的mate家族二级结构特征。拟南芥中已有十多个成员被报道且功能多样、迥异。近年研究证实此家族的一些成员介导有机酸及阴离子的转运调控拟南芥细胞膨压具有耐旱性质。水稻中目前只有osfrdl1和osfrdl2被报道具有转运柠檬酸的功能与铁营养吸收有关,其他大部分成员的功能尚未解析。
二、tcbg1的表达模式分析
一个基因的表达模式决定了基因的功能。为了探究tcbg1的表达模式,我们将该基因的3204bp启动子(3.2kbupstreamofthestartcodon)protcbg1与β-glucuronidase(gus)受体融合,并将重组质粒转入野生型水稻(wt)中。对protcbg1-gus转基因植株中tcbg1启动子的活性进行了分析,其结果显示:随着颖壳的发育,tcbg1的表达水平逐步增加。
如图3所示,在水稻wt植株的生殖器官(图3a-g)的不同发育时期均检测到gus活性。图中蓝色表示具有gus活性的组织,且蓝色越深表示该组织中的tcbg1基因表达量越高。图a、b和d显示tcbg1在水稻雄蕊中有较高的表达。进一步将雄蕊放大,如图e所示,可以清晰看出tcbg1主要表达在雄蕊的花药中。如图f所示,进一步观察tcbg1在不同时期花药中的表达量发现,随着花药的发育,tcbg1在花药中的表达量升高。图f中,从右向左为花药的成熟程度逐渐提高。图b、c和g显示的是tcbg1在雌蕊中的表达模式分析,可以看出,随着雄蕊的发育,tcbg1的表达量逐渐降低。图h为种子,如图所示,tcbg1在种子中表达量较高。此结果证实tcbg1在水稻生殖发育过程中高表达,预示tcbg1会参与籽粒的发育过程。
在本发明的一些实施例中,在筛选水稻突变体的过程中,针对tcbg1基因与其编码的蛋白,通过改变细胞膨压或渗透势增加水稻籽粒的灌浆速度和时间,从而实现了增加水稻的籽粒大小。这一发现与以往报道的籽粒大小控制基因不同,这一家族既不调控籽粒的细胞分裂,也不调控籽粒同化物的合成和运输。因此,本发明在增加水稻产量方面具备开创性。
在以下一些具体的实施例中,通过水稻tcbg1突变体进一步说明本发明的技术方案。
实施例一:
利用crispr/cas9敲除tcbg1以及突变体的鉴定:
利用crispr/cas9技术对tcbg1基因进行编辑。选取该基因orf的几个合适位点作为靶序列,通过序列分析,得到如表1中所示的6个合适位点。以下将以第3号靶序列为例,简要说明利用crispr/cas9技术敲除tcbg1基因及突变体的鉴定。
表1.合适的crispr/cas9的靶序列。其中单划线部分碱基为合适的酶切位点,双划线部分碱基为pam。
第242bp-265bp作为crispr靶序列(图4a),其中包含一个ncoi(ccatgg)限制性酶切位点(单划线部分碱基),方便后续阳性植株的鉴定。利用农杆菌介导的遗传转化系统,将构建好的融合质粒转入野生型水稻中,并将收获的种子重新种下。对获得的阳性植株编号(如1-12)并提取基因组dna。在该碱基序列中选取该crispr靶序列两端设计引物,使该crispr靶序列与两端引物间距离相差至少几十个碱基,且两个引物区间内只能含有一个该限制性内切酶,此处为ncoi。利用上述设计的引物将crispr靶序列区域进行pcr扩增,然后用ncoi对pcr产物在37℃过夜酶切。电泳检测发现编号1,2,7的植株完全不能被切开,说明该酶切位点被破坏,即为纯合突变体;编号为3,4,5,8,9,10,11,12号的植株虽然也有部分碱基片段没有被切开,但是还有一部分可以被ncoi切开,说明该植株为杂合突变体;最后一列为对照组,即没有将含有融合质粒的农杆菌转入该植株,其相应碱基片段可以完全被该限制性内切酶切成两段(如图4b)。对编号1,2,7植株进行测序分析,发现这三个植株均发生一个碱基的缺失(figure4candd),突变后的cds序列如seqidno.3所示,其cds碱基序列251位胸腺嘧啶(t)缺失。
实施例二:
利用talen技术敲除tcbg1以及突变体的鉴定:
利用转录激活因子效应蛋白核酸酶(transcriptionactivator-likeeffectornuclease,talen)技术实现对水稻tcbg1基因进行定点修饰,获得功能缺失的突变体。
选择合适的靶序列作为talen的识别位点。本发明中,选取该基因orf的几个合适位点作为识别区序列,通过序列分析,设计几组talen识别区序列,得到如表2中所示的3个合适位点。
表1.合适的talen的识别序列。其中同一序列中双下划线部分碱基分别为左侧靶序列和右侧靶序列。
以下将以第2号识别区序列作为talen识别位点为例,简要说明利用talen技术敲除tcbg1基因及其突变体的鉴定。如图5a所示,选取第219-271bp作为talen的识别区序列,其中左侧双下划线部分碱基组成左侧靶序列,右侧双下划线部分碱基组成右侧靶序列。左、右靶序列可特异识别并结合至目的基因tcbg1的指定位置,并对其之间部分碱基进行编辑。
选取构建相关的talen敲除载体,并利用实施例一中方法将其转入野生型水稻中。收取种子,并将收获的种子重新种下。筛选得到的第二代转基因水稻,从筛选到的140株转基因植株中鉴定目的基因被破坏的阳性植株,根据所设计识别区序列以及所使用载体的不同,可以有不同的鉴定方法,例如利用实施例一种所述的酶切方法、抗生素筛选方法等,此处不再赘述。
提取转基因植株的基因组dna,扩增目的基因tcbg1的dna片段,并对其进行测序分析,最终鉴定到八种(十一株)基因组被不同程度编辑的阳性植株。图5b中显示了这十一株突变体类型,为不同程度或不同位置的碱基缺失。
实施例三:
rnai株系的获得及分析
利用rna干扰的技术抑制tcbg1的表达从而研究目的基因的功能,本研究选取该基因orf靠近5’端位置第241个碱基至第540个碱基序列,即长度为300bp的片段作为靶序列(如图6a),其碱基序列详见seqidno.5。将该段靶序列重组到水稻表达载体ptck309(如图6b)中,然后转入野生型水稻中。
首先对获得的阳性植株进行dna水平的鉴定,检测转基因植株中是否包括表达载体ptck309上特有的序列,以确定转基因是否成功。然后对其中的转基因阳性植株中tcbg1的表达水平进行分析,定量检测rnai技术编辑后tcbg1基因的表达量。通过利用rt-pcr对tcbg1的表达检测,最终筛选得到tcbg1的表达被抑制了百分之六十以上的植株,将其命名为ri-1和ri-2(如图6c)。如图6c所示,纵轴表示tcbg1的相对表达量,黑色柱型表示野生型植株,灰色表示与野生型植株具有相同生长时期的rnai技术得到的tcbg1突变体植株,其中中间灰色柱型为ri-1,右侧为ri-2。
实施例一、实施例二和实施例三的表型性状及分析
此处以crispr/cas9技术获得的碱基缺失突变体和rnai技术获得的基因沉默突变体为例,分析tcbg1的表型性状。其中将crispr/cas9技术获得的突变体命名为tcbg1-1,rnai技术获得的突变体命名为tcbg1-2。
a、野生型和突变体粒型比较分析:tcbg1突变体籽粒千粒重较野生型显著提高
粒型直接决定了籽粒的重量,包括粒长、粒宽和粒厚。tcbg1突变体谷粒的长度,宽度和厚度较野生型均有不同程度的增加,其中谷粒长增加了10.2%左右(如图8a和c),谷粒宽度增加了约7.1%(如图8b和d),粒厚也提高了6.5%(如图8e),同时对糙米粒也进行了统计,发现糙米粒的长、宽和厚度分别增加8%,9%和3%(如图8f-j),因此使千粒重较野生型提高了13%左右(如图7c)。
tcbg1的rna干扰株系表型与tcbg1突变体相似,株高也较野生型稍矮,籽粒较野生型变大,tcbg1的表达水平较野生型下降百分之六十左右,籽粒大小相应变大。
b、tcbg1突变体灌浆速率明显快于野生型。
为了探究tcbg1影响了灌浆过程的那一阶段导致突变体和野生型之间出现的个体大小差别,对野生型和突变体的灌浆过程进行了跟踪记录,每隔三天取样,测量了稻谷和糙米粒的湿重和干重。如图9a和b所示,稻谷和糙米相似,在授粉十二天后突变体的灌浆速度明显比野生型快,突变体的干重和湿重均明显高于野生型,授粉21天后灌浆达到最高值,突变体稻谷的湿重和干重较野生型分别提高18.1%和21.1%,糙米粒的湿重和干重分别增加16.0%和18.2%,为了探究突变体灌浆加快的原因,对灌浆过程中淀粉合成相关的ap2/erebp、agpl2、gbssi、glucan、bei五个基因的表达水平进行检测,发现在突变体中这五个基因的表达水平明显比野生型高两倍以上。因此可以得出,在灌浆的中后期突变体灌浆速率明显加快使得最终突变体积累的有机物明显比野生型增多。
c、tcbg1的分子功能:tcbg1具有氯离子外向转运活性
为探究tcbg1的电生理活性,用微注射的方法将体外转录的tcbg1注入非洲爪蟾卵母细胞(下称蛙卵)中。对照组蛙卵细胞使用同样的方法注射等体积的不含rna酶的水。将注射crna和水的蛙卵在18℃环境中培养约36小时后进行电生理活性实验。对实验结果分析发现tcbg1与dtx33/35功能类似,具有氯离子外向转运活性。如图10所示,对照组蛙卵(注射不含有rna酶的水)未检测到明显的电流活性(如图10a和b),而表达tcbg1的蛙卵(注射tcbg1体外转录的crna)在灌流液中加入96mmnacl时可检测到明显的电流活性(如图10c)。
我们利用上述技术,将第251位t删除的tcbg1突变体crna表达在蛙卵中,由于该蛋白在翻译过程中提前终止,其氨基酸序列如seqidno.4所示,也未检测到明显的电流活性(如图10e和f),说明该位点的突变导致了tcbg1功能的丧失。
进一步地,基于tcbg1的转运活性以及底物特异性,为进一步确认该电流活性由氯离子产生,用谷氨酸根取代氯离子,在灌流液中加入96mm谷氨酸钠,同样的方法记录注射crna的蛙卵细胞的电流活性。此时并未检测到明显电流的产生(如图10d),初步确认tcbg1电流的产生是由于氯离子的外向转运产生的。因此,突变体中缺失了该通道使得氯离子不能从胞内排除,增大了细胞的渗透势从而改变细胞的膨压。
因此,tcbg1基因介导氯离子的转运。通过将tcbg1基因敲除或使基因沉默可以调控细胞膨压或渗透势,从而增加水稻籽粒的大小。在上述的实施例一和二中,tcbg1基因的突变体中缺失了氯离子外排通道,使得氯离子不能从胞内排除,增大了细胞的渗透势从而改变细胞的膨压,进而改变了水稻生长的生理过程,使得种子的细胞变大,积累更多的有机物,增大种子的重量。本发明代表了控制水稻籽粒大小的全新机制,在增加水稻产量方面具备开创性。
上述实施例仅供说明本发明之用,而并非是对本发明的限制,有关技术领域的普通技术人员,在不脱离本发明范围的情况下,还可以做出各种变化和变型,因此,所有等同的技术方案也应属于本发明公开的范畴。
sequencelisting
<110>首都师范大学
<120>一种具有阴离子转运活性的通道蛋白及其基因组修饰调控作物籽粒大小
<130>90
<160>5
<170>patentinversion3.3
<210>1
<211>1440
<212>dna
<213>水稻(oryzasativa)
<400>1
atgacgcctccaccgccgtcgccgccgcacgagaggaaaacgtgggcggagtcggtggcc60
agcgagtttcgggcgcagcgcggcatcgcgttccctctcatcgccatgaacctcacctgg120
ttcgccaagctggccgtcaccaccgccttcctcggccgcctcggcgacctccagctcgcc180
gccggcaccctcggcttcagcttcgccaatgtcaccggcttcgccgtcctcaccggcctc240
tgcgccgccatggaccccatctgcgggcaggcgcacggcgccagcaacgggaagctcctc300
cgcaagacgctggtgatggccaccatcctgctgctgggcgcgtccatccccatcgccttc360
ctgtggctgcacgtggacgccgtcctcctccggttcggacagcaggcggacatgagcagc420
aacgcacgcagctacgtggtgtgcctcctcccggacctcgccgtcacctccttcgtcaac480
ccgctcaagtcgtacctgagcgcgcagggggtgacgctccccacgctgttcgcctccgcc540
ctggccctggcgctccacgtccccctcaccatgtggatggccaggaccaggggcatccag600
ggcgtcgccaccgccgtgtgggtcagcgacctggccgtggccgtcatgctcgccggctac660
gtgctcgtctcggagcgacgacggaaggcgggagggggcggcggatgggtggagcagacg720
aggggtgagtgggtccggctcctccggctggccgttcccagctgcctcaacacctgcctg780
gagtggtggtgctacgagatactggtgctcctgacgggacgcctcccggacgcccggcgc840
acggtggcggtgatggccgtgacgctcaacttcgactacctgctgttcgcggggatgctg900
tccctgtcggtgagcgcgtcggtgcgcgtgtcgaacgagctgggcgcgggggaggcgtgg960
gcggcgaggcgcgcgggcatggtgtcgatcgtgggcggcgcggtgggcggggtgggcggc1020
ggggtggcgatggtggcggcgcggcgggcgtgggggagcatatacagctcagacgccggg1080
gtgcgggagggggtggggagggcgatggaggtgatggcggtgctggaggtggtgaacttc1140
ccgctgaacgtgtgcggggggatagtgcgagggacggcgaggccggcggtggggatgtac1200
gccgtggtggccggcttctacgtgctggcgctgccgctcggggtcgcgctcgccttcaag1260
gccagacttgggatccagggcctcctcctgggcttcctggtgggcgccgcggccagcttg1320
gcggtgctcctcaccttcatcgcgcgcatggattggcccgccgaggcccaaaaggcgcgg1380
actagaaccacagcaaccgtggcccaattccaccaacacgacgaggtcgtccagccttga1440
<210>2
<211>479
<212>prt
<213>水稻(oryzasativa)
<400>2
metthrproproproproserproprohisgluarglysthrtrpala
151015
gluservalalaserglupheargalaglnargglyilealaphepro
202530
leuilealametasnleuthrtrpphealalysleualavalthrthr
354045
alapheleuglyargleuglyaspleuglnleualaalaglythrleu
505560
glypheserphealaasnvalthrglyphealavalleuthrglyleu
65707580
cysalaalametaspproilecysglyglnalahisglyalaserasn
859095
glylysleuleuarglysthrleuvalmetalathrileleuleuleu
100105110
glyalaserileproilealapheleutrpleuhisvalaspalaval
115120125
leuleuargpheglyglnglnalaaspmetserserasnalaargser
130135140
tyrvalvalcysleuleuproaspleualavalthrserphevalasn
145150155160
proleulyssertyrleuseralaglnglyvalthrleuprothrleu
165170175
phealaseralaleualaleualaleuhisvalproleuthrmettrp
180185190
metalaargthrargglyileglnglyvalalathralavaltrpval
195200205
seraspleualavalalavalmetleualaglytyrvalleuvalser
210215220
gluargargarglysalaglyglyglyglyglytrpvalgluglnthr
225230235240
argglyglutrpvalargleuleuargleualavalprosercysleu
245250255
asnthrcysleuglutrptrpcystyrgluileleuvalleuleuthr
260265270
glyargleuproaspalaargargthrvalalavalmetalavalthr
275280285
leuasnpheasptyrleuleuphealaglymetleuserleuserval
290295300
seralaservalargvalserasngluleuglyalaglyglualatrp
305310315320
alaalaargargalaglymetvalserilevalglyglyalavalgly
325330335
glyvalglyglyglyvalalametvalalaalaargargalatrpgly
340345350
seriletyrserseraspalaglyvalarggluglyvalglyargala
355360365
metgluvalmetalavalleugluvalvalasnpheproleuasnval
370375380
cysglyglyilevalargglythralaargproalavalglymettyr
385390395400
alavalvalalaglyphetyrvalleualaleuproleuglyvalala
405410415
leualaphelysalaargleuglyileglnglyleuleuleuglyphe
420425430
leuvalglyalaalaalaserleualavalleuleuthrpheileala
435440445
argmetasptrpproalaglualaglnlysalaargthrargthrthr
450455460
alathrvalalaglnphehisglnhisaspgluvalvalglnpro
465470475
<210>3
<211>1439
<212>dna
<213>水稻(oryzasativa)
<400>3
atgacgcctccaccgccgtcgccgccgcacgagaggaaaacgtgggcggagtcggtggcc60
agcgagtttcgggcgcagcgcggcatcgcgttccctctcatcgccatgaacctcacctgg120
ttcgccaagctggccgtcaccaccgccttcctcggccgcctcggcgacctccagctcgcc180
gccggcaccctcggcttcagcttcgccaatgtcaccggcttcgccgtcctcaccggcctc240
tgcgccgccaggaccccatctgcgggcaggcgcacggcgccagcaacgggaagctcctcc300
gcaagacgctggtgatggccaccatcctgctgctgggcgcgtccatccccatcgccttcc360
tgtggctgcacgtggacgccgtcctcctccggttcggacagcaggcggacatgagcagca420
acgcacgcagctacgtggtgtgcctcctcccggacctcgccgtcacctccttcgtcaacc480
cgctcaagtcgtacctgagcgcgcagggggtgacgctccccacgctgttcgcctccgccc540
tggccctggcgctccacgtccccctcaccatgtggatggccaggaccaggggcatccagg600
gcgtcgccaccgccgtgtgggtcagcgacctggccgtggccgtcatgctcgccggctacg660
tgctcgtctcggagcgacgacggaaggcgggagggggcggcggatgggtggagcagacga720
ggggtgagtgggtccggctcctccggctggccgttcccagctgcctcaacacctgcctgg780
agtggtggtgctacgagatactggtgctcctgacgggacgcctcccggacgcccggcgca840
cggtggcggtgatggccgtgacgctcaacttcgactacctgctgttcgcggggatgctgt900
ccctgtcggtgagcgcgtcggtgcgcgtgtcgaacgagctgggcgcgggggaggcgtggg960
cggcgaggcgcgcgggcatggtgtcgatcgtgggcggcgcggtgggcggggtgggcggcg1020
gggtggcgatggtggcggcgcggcgggcgtgggggagcatatacagctcagacgccgggg1080
tgcgggagggggtggggagggcgatggaggtgatggcggtgctggaggtggtgaacttcc1140
cgctgaacgtgtgcggggggatagtgcgagggacggcgaggccggcggtggggatgtacg1200
ccgtggtggccggcttctacgtgctggcgctgccgctcggggtcgcgctcgccttcaagg1260
ccagacttgggatccagggcctcctcctgggcttcctggtgggcgccgcggccagcttgg1320
cggtgctcctcaccttcatcgcgcgcatggattggcccgccgaggcccaaaaggcgcgga1380
ctagaaccacagcaaccgtggcccaattccaccaacacgacgaggtcgtccagccttga1439
<210>4
<211>104
<212>prt
<213>水稻(oryzasativa)
<400>4
metthrproproproproserproprohisgluarglysthrtrpala
151015
gluservalalaserglupheargalaglnargglyilealaphepro
202530
leuilealametasnleuthrtrpphealalysleualavalthrthr
354045
alapheleuglyargleuglyaspleuglnleualaalaglythrleu
505560
glypheserphealaasnvalthrglyphealavalleuthrglyleu
65707580
cysalaalaargthrproseralaglyargargthralaproalathr
859095
glyserserseralaargargtrp
100
<210>5
<211>300
<212>dna
<213>水稻(oryzasativa)
<400>5
tgcgccgccatggaccccatctgcgggcaggcgcacggcgccagcaacgggaagctcctc60
cgcaagacgctggtgatggccaccatcctgctgctgggcgcgtccatccccatcgccttc120
ctgtggctgcacgtggacgccgtcctcctccggttcggacagcaggcggacatgagcagc180
aacgcacgcagctacgtggtgtgcctcctcccggacctcgccgtcacctccttcgtcaac240
ccgctcaagtcgtacctgagcgcgcagggggtgacgctccccacgctgttcgcctccgcc300
- 还没有人留言评论。精彩留言会获得点赞!
- 使用阴离子氧化多糖用于保护和修复角蛋白纤维的方法
- 电转运递送阴离子型药物的阴极的制作方法
- *酮类化合物在制备治疗胆汁淤积的药物中的应用的制作方法
- 重组人二价金属离子转运蛋白腺病毒、制备及应用的制作方法
- 一种钾离子转运相关蛋白系统及其编码基因簇与应用的制作方法
- 一种slc22a6基因多态性检测特异性引物和液相芯片的制作方法
- 一种新的多肽——人组织阴离子转运多肽41和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人组织阴离子转运多肽13.75和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人组织阴离子转运多肽11.33和编码这种多肽的多核苷酸的制作方法
- 一种多肽——人组织阴离子转运多肽13.93和编码这种多肽的多核苷酸的制作方法