抗EGFR/PD-1双特异性抗体的制作方法
本发明属于肿瘤治疗和生物
技术领域:
,涉及一种抗egfr和pd-1的双特异性抗体分子制备方法和用途。
背景技术:
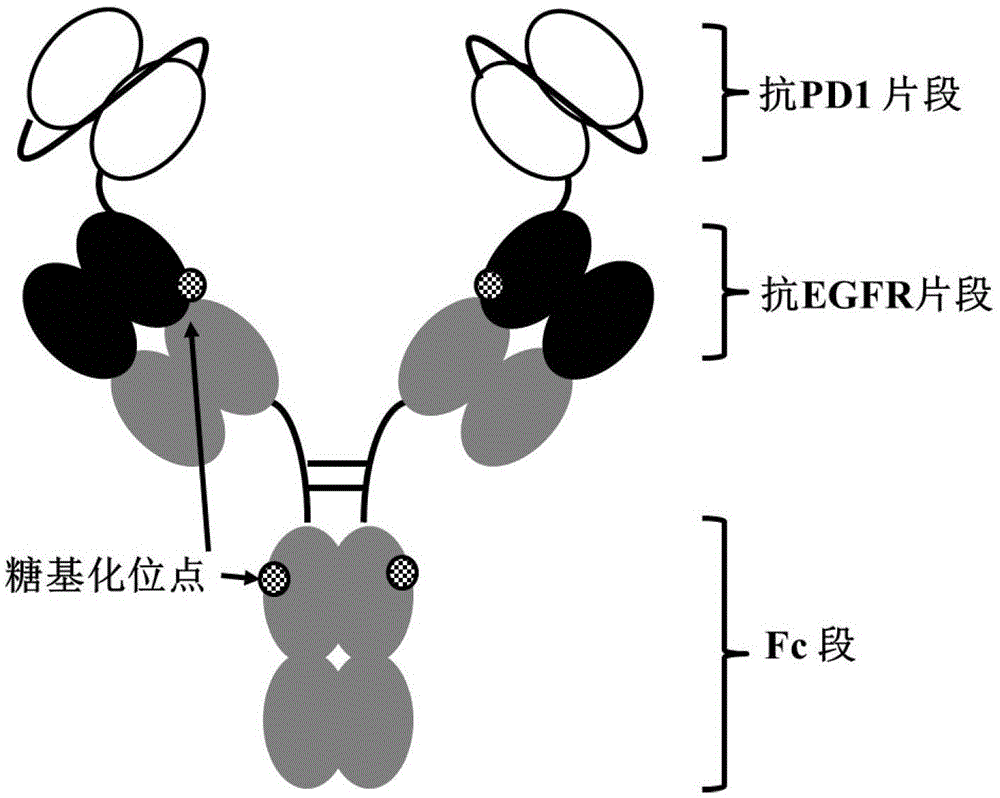
:egfr(epidermalgrowthfactorreceptor)是表皮生长因子(egf)的受体,属于erbb受体家族。egfr是一种分子量为170kda的跨膜糖蛋白,属于受体型酪氨酸激酶,在相关配体如egf和转化生长因子-α(transforminggrowthfactorα,tgfα)的作用下,egfr由单体转化为二聚体而被激活,从而进一步活化下游信号传导通路,调控细胞的增殖。大量研究表明,在大多数肿瘤如胶质细胞癌、肾癌、肺癌、前列腺癌、胰腺癌、乳腺癌等组织中存在egfr的高表达或者异常表达。egfr功能的异常与肿瘤细胞的增殖、血管生成、肿瘤侵袭、转移以及细胞凋亡的抑制有关。其功能的异常主要表现为两个方面:一是在肿瘤组织中的过度异常表达,二是egfr突变体在肿瘤细胞中的持续性激活(不需要配体刺激或形成自循环刺激通路)。在结肠癌病人组织中,egfr的表达率约为25-77%。相关临床数据表明,egfr表达量的多少与肿瘤的恶性程度以及肿瘤患者的预后密切相关。erbitux(cetuximab,imc-c225)是针对egfr的人鼠嵌合单克隆抗体,特异性地与egfr结合,竞争性阻断efgr与其配体结合,从而抑制egfr信号传导。erbitux于2004年2月被批准上市,用于与伊立替康联合治疗egfr阳性、伊立替康化疗无效的转移性结直肠癌和联合放疗用于治疗局部区域性早期头颈部鳞状细胞癌(squamouscellcarcinomaoftheheadandneck,scchn)。由三生国健自主研发的egfr单克隆抗体是按照erbitux的氨基酸序列、用cho细胞表达系统进行表达、并用自主研发的细胞培养生产工艺得到的人鼠嵌合单克隆抗体。体内外生物学活性以及抗肿瘤活性研究显示,三生国健药业自主研发的egfr单克隆抗体与阳性对照药erbitux具有极其相似的生物学活性,且其生物学活性在某些方面略优于阳性对照药erbitux。人程序性细胞死亡受体-1(pd-1)是由288个氨基酸组成的i型膜蛋白,胞外段为负责结合配体的ig可变型(v-型)结构域,胞内段为负责结合信号转导分子的胞质尾区。pd-1胞质尾区含有两个基于酪氨酸的信号转导模体,分别为itim(免疫受体酪氨酸抑制作用模体)和itsm(免疫受体酪氨酸转换作用模体)。pd-1表达在已经激活的t淋巴细胞表面,它与配体pd-l1(程序性死亡受体-配体1,programmedcelldeath-ligand1)和pd-l2(程序性死亡受体-配体2,programmedcelldeath-ligand2)结合可以抑制t淋巴细胞的活性及相关的体内细胞免疫反应。大量研究表明,pd-1和pd-l1的相互作用不仅维持了体内免疫系统的平衡,也是导致pd-l1表达阳性的肿瘤细胞规避免疫监视的主要机制。通过阻断pd-1/pd-l1信号通路,能够激活免疫系统,促进t细胞对肿瘤细胞的杀伤。(pembrolizμmab)是第一个上市的针对pd-1的人源化单克隆抗体,于2014年9月被fda批准用于治疗黑色素瘤,至2018年获批的适应证包括:黑色素瘤、非小细胞肺癌、霍奇金淋巴瘤、头颈部鳞癌、膀胱癌、胃癌和带有msi-h或dmmr的实体肿瘤。(nivolμmab)是百时美施贵宝公司的一款pd-1单克隆抗体,于2014年12月获fda批准上市,适应证包括:黑色素瘤、非小细胞肺癌、肾细胞癌、经典霍奇金淋巴瘤、头颈部鳞癌、膀胱癌、结直肠癌和肝细胞癌。中国专利申请cn201710054783.5公开了一种由三生国健药业自主研发的全新的抗pd-1人源化单克隆抗体。体内外生物学活性以及抗肿瘤活性研究显示,三生国健药业研发的pd-1单克隆抗体的生物学活性介于阳性对照药opdivo和keytruda之间,在某些方面略优于阳性对照药opdivo。双特异性抗体(bispecificantibody,bsab)是指能同时结合两个(或多个)不同抗原表位的抗体分子。与传统的单克隆抗体相比,双特异性抗体具有独特的作用机制:1)双特异性抗体可以同时结合2个或多个不同的抗原分子或相同分子的不同表位,产生协同效应。而抗体联用往往不具备这种效应。2)介导细胞间的相互作用。双特异性抗体可分别结合到效应细胞和靶细胞上的两种抗原上,在效应细胞和靶细胞之间架起桥梁,促进细胞间的相互作用,例如介导免疫细胞对肿瘤细胞的杀伤。因此双特异性抗体具有传统单克隆抗体不具备的独特优势。本专利提供了一种能靶向肿瘤细胞表面分子egfr和t淋巴细胞表面分子pd-1的双特异性抗体。和单克隆抗体相比,该双特异性抗体表现出协同效应,与抗体联用相比,该双特异性抗体抗肿瘤效果优于抗体联用。技术实现要素:本发明提供了一种新的能与egfr和pd-1特异结合的双特异性抗体,还提供了该双特异性抗体的制备方法和应用。因此,本发明的目的在于提供一种能与egfr和pd-1特异结合的双特异性抗体;提供编码所述双特异性抗体的核苷酸分子;提供包含所述核苷酸分子的表达载体;提供所述表达载体的宿主细胞;提供所述双特异性抗体的制备方法;提供包含所述双特异性抗体的药物组合物;提供所述双特异性抗体在制备药物中的应用。为了实现上述目的,本发明采用了如下技术方案:本发明一方面提供了一种能与egfr和pd-1特异结合的双特异性抗体,其包含免疫球蛋白抗体igg和两个相同的单链可变片段scfv,其中每个scfv包含可变区vh和可变区vl,vh通过肽接头l1连接至vl,每个scfv的c末端经由肽接头l2连接至igg重链的n末端。本发明“双特异性抗体”是指拥有两个不同的抗原结合位点,能同时结合至egfr和pd-1的双特异性抗体,其包含两个单链可变片段scfv和与之缀合的免疫球蛋白抗体igg,每个scfv的c末端经由肽接头l2连接至igg每条重链的n末端,形成双特异性抗体的重链融合蛋白,其中每个scfv包含可变区vh和可变区vl,vh通过肽接头l1连接至vl。本发明“单链可变片段scfv”是指包含免疫球蛋白重链vh和轻链vl可变区的融合蛋白,vh通过肽接头融合至vl,其中所述融合蛋白保留了完整免疫球蛋白相同的抗原特异性。本发明“免疫球蛋白抗体igg”是约150kda的分子,它由四条肽链构成,含有两条相同的约50kda的γ重链,和两条相同的约25kda的轻链,从而具有四聚体四级结构。两条重链通过二硫键相互连接,并各自与一条轻链连接。所成的四聚体具有相同的两半,二者形成叉型或者类似y的形状,叉的每一端含有一个相同的抗原结合位点。igg抗体可以基于重链的恒定区中氨基酸序列的微小差异而分为多个亚类(例如igg1、2、3、4)。作为优选的方案,所述vh包含互补决定区hcdr1-3,其中hcdr1的氨基酸序列为seqidno:1,hcdr2的氨基酸序列为seqidno:2,hcdr3的氨基酸序列为seqidno:3;所述vl包含互补决定区lcdr1-3,其中lcdr4的氨基酸序列为seqidno:4,lcdr5的氨基酸序列为seqidno:5,lcdr6的氨基酸序列为seqidno:6;所述igg的重链包含互补决定区hcdr4-6,其中hcdr4的氨基酸序列为seqidno:7,其中hcdr5的氨基酸序列为seqidno:8,其中hcdr6的氨基酸序列为seqidno:9;所述igg的轻链包含互补决定区lcdr4-6,其中lcdr4的氨基酸序列为seqidno:10,其中lcdr5的氨基酸序列为seqidno:11,其中lcdr6的氨基酸序列为seqidno:12。本领域中,抗体的结合区通常均含有一条轻链可变区和一条重链可变区,每一个可变区均含有3个cdr三个结构域。抗体的重链和轻链的cdr结构域分别称为hcdr和lcdr。因此,常规抗体抗原结合位点包含六个cdr,包括分别来自重链和轻链v区的cdr集合。作为优选的方案,vh的氨基酸序列为seqidno:13,vl的氨基酸序列为seqidno:14;所述igg的重链可变区的氨基酸序列为seqidno:15,轻链可变区的氨基酸序列为seqidno:16。作为优选的方案,所述肽接头l1的氨基酸序列为seqidno:17。作为优选的方案,所述肽接头l2的氨基酸序列为seqidno:18。作为优选的方案,所述单链可变片段scfv的氨基酸序列为seqidno:19。作为优选的方案,所述双特异性抗体的重链氨基酸序列为seqidno:20,其轻链氨基酸序列为seqidno:21。在构建本发明的双特异性抗体时,与该双特异性抗体的化学和物理稳定性相关的问题也得到了解决,诸如表达物理稳定的分子、增加热和盐依赖的稳定性、降低聚集、增加在高浓度下的溶解度以及维持分别针对两种抗原egfr和pd-1的亲和力等。本发明另一方面提供了一种核苷酸分子,所述核苷酸分子编码上述任一所述的双特异性抗体。作为优选的方案,所述核苷酸分子编码能与egfr和pd-1特异结合的双特异性抗体的重链的核苷酸序列为seqidno:22,编码其轻链的核苷酸序列为seqidno:23。本发明所述核苷酸分子的制备方法为本领域常规的制备方法,较佳地包括以下制备方法:通过基因克隆技术例如pcr方法等,获得编码上述单克隆抗体的核苷酸分子,或者通过人工全序列合成的方法得到编码上述单克隆抗体的核苷酸分子。本领域技术人员知晓,编码上述双特异性抗体的氨基酸序列的核苷酸序列可以适当引入替换、缺失、改变、插入或增加来提供一个多聚核苷酸的同系物。本发明中多聚核苷酸的同系物可以通过对编码该双特异性抗体基因的一个或多个碱基在保持抗体活性范围内进行替换、缺失或增加来制得。本发明另一方面提供了一种表达载体,所述表达载体含有上述的核苷酸分子。其中所述表达载体为本领域常规的表达载体,是指包含适当的调控序列,例如启动子序列、终止子序列、多腺苷酰化序列、增强子序列、标记基因和/或序列以及其他适当的序列的表达载体。所述表达载体可以是病毒或质粒,如适当的噬菌体或者噬菌粒,更多技术细节请参见例如sambrook等,molecularcloning:alaboratorymanual,第二版,coldspringharborlaboratorypress,1989。许多用于核酸操作的已知技术和方案请参见currentprotocolsinmolecularbiology,第二版,ausubel等编著。本发明所述表达载体较佳地为pdr1,pcdna3.1(+),pcdna3.1/zeo(+),pdhfr,ptt5,pdhff,pgm-csf或pcho1.0,更佳地为ptt5。本发明另外提供了一种宿主细胞,所述宿主细胞含有上述的表达载体。本发明所述的宿主细胞为本领域常规的各种宿主细胞,只要能满足使上述重组表达载体稳定地自行复制,且所携带所述的核苷酸可被有效表达即可。其中所述宿主细胞包括原核表达细胞和真核表达细胞,所述表达载体较佳地包括:cos、cho(中国仓鼠卵巢,chinesehamsterovary)、ns0、sf9、sf21、dh5α、bl21(de3)或tg1,更佳地为e.colitg1、bl21(de3)细胞(表达单链抗体或fab抗体)或者cho-k1细胞(表达全长igg抗体)。将前述表达载体转化至宿主细胞中,即可得本发明优选的重组表达转化体。其中所述转化方法为本领域常规转化方法,较佳地为化学转化法,热激法或电转法。作为优选的方案,所述宿主细胞是真核细胞。优选自cho细胞和293e细胞。本发明另一方面提供了上述能与egfr和pd-1特异结合的双特异性抗体的制备方法,所述制备方法包括以下步骤:a)在表达条件下,培养上述的宿主细胞,从而表达能与egfr和pd-1特异结合的双特异性抗体;b)分离并纯化步骤a)所述的双特异性抗体。本发明所述的宿主细胞的培养方法、所述抗体的分离和纯化方法为本领域常规方法,具体操作方法请参考相应的细胞培养技术手册以及抗体分离纯化技术手册。本发明中公开的抗egfr/pd-1双特异性抗体的制备方法包括:在表达条件下,培养上述的宿主细胞,从而表达能与egfr和pd-1特异结合的双特异性抗体;分离和纯化所述的抗egfr/pd-1双特异性抗体。利用上述方法,可以将重组蛋白纯化为基本均一的物质,例如在sds-page电泳上为单一条带。可以利用亲和层析的方法对本发明公开的抗egfr/pd-1双特异性抗体进行分离纯化,根据所利用的亲和柱的特性,可以使用常规的方法例如高盐缓冲液、改变ph等方法洗脱结合在亲和柱上的抗egfr/pd-1双特异性抗体。本发明的发明人对所得抗egfr/pd-1双特异性抗体进行了检测实验,实验结果表明该抗egfr/pd-1双特异性抗体能很好地与靶细胞和抗原结合,具有较高的亲和力。本发明另一方面提供了一种组合物,所述组合物包含上述所述的能与egfr和pd-1特异结合的双特异性抗体和一种或多种药学上可接受的载体、稀释剂或赋形剂。本发明提供的双特异性抗体,可以和药学上可以接受的载体一起组成药物制剂组合物从而更稳定地发挥疗效,这些制剂可以保证本发明公开的双特异性抗体的氨基酸核心序列的构像完整性,同时还保护蛋白质的多官能团防止其降解(包括但不限于凝聚、脱氨或氧化)。通常情况下,对于液体制剂,通常可以在2℃-8℃条件下保存至少稳定一年,对于冻干制剂,在30℃至少六个月保持稳定。所述双特异性抗体制剂可为制药领域常用的混悬、水针、冻干等制剂。对于本发明公开的双特异性抗体的水针或冻干制剂,药学上可以接受的载体较佳地包括但不限于:表面活性剂、溶液稳定剂、等渗调节剂和缓冲液之一或其组合。其中表面活性剂较佳地包括但不限于:非离子型表面活性剂如聚氧乙烯山梨醇脂肪酸酯(吐温20或80);poloxamer(如poloxamer188);triton;十二烷基硫酸钠(sds);月桂硫酸钠;十四烷基、亚油基或十八烷基肌氨酸;pluronics;monaquattm等,其加入量应使抗egfr/pd-1双特异性抗体的颗粒化趋势最小。溶液稳定剂较佳地包括但不限于以下列举之一或其组合:糖类,例如,还原性糖和非还原性糖;氨基酸类,例如,谷氨酸单钠或组氨酸;醇类,例如:三元醇、高级糖醇、丙二醇、聚乙二醇等,溶液稳定剂的加入量应该使最后形成的制剂在本领域的技术人员认为达到稳定的时间内保持稳定状态。等渗调节剂较佳地包括但不限于氯化钠、甘露醇之一或其组合。缓冲液较佳地包括但不限于:tris、组氨酸缓冲液、磷酸盐缓冲液之一或其组合。本发明另一方面提供了上述能与egfr和pd-1特异结合的双特异性抗体、或上述药物组合物在制备药物中的应用,其特征在于所述药物用于治疗癌症或肿瘤。本发明所称的用于治疗癌症或肿瘤的药物,指具有抑制和/或治疗肿瘤的药物,可以包括伴随肿瘤相关症状发展的延迟和/或这些症状严重程度的降低,进一步还包括已存在的肿瘤伴随症状的减轻并防止其他症状的出现,还包括减少或防止肿瘤的转移等。本发明所述的药物所针对的肿瘤较佳地包括但不限于:肺癌、骨癌、胃癌、胰腺癌、皮肤癌、头颈癌、子宫癌、卵巢癌、睾丸癌、子宫癌、输卵管癌、子宫内膜癌、子宫颈癌、阴道癌、外阴癌、直肠癌、结肠癌、肛门区癌、乳腺癌、食管癌、小肠癌、内分泌系统癌、甲状腺癌、甲状旁腺癌、肾上腺癌、尿道癌、阴茎癌、前列腺癌、胰腺癌、脑癌、睾丸癌、淋巴癌、移行细胞癌、膀胱癌、肾癌或输尿管癌、肾细胞癌、肾盂癌、霍奇金病、非霍奇金淋巴瘤、软组织肉瘤、儿童实体瘤、淋巴细胞性淋巴瘤、中枢神经系统(cns)肿瘤、原发性中枢神经系统淋巴瘤、肿瘤血管生成、脊柱肿瘤、脑干神经胶质瘤、垂体腺瘤、黑素瘤、卡波西肉瘤、表皮样癌、鳞状细胞癌、t细胞淋巴瘤、慢性或急性白血病和所述癌的组合。本发明中双特异性抗体及其组合物在对包括人在内的动物给药时,给药剂量因病人的年龄和体重,疾病特性和严重性,以及给药途径而异,可以参考动物实验的结果和种种情况,总给药量不能超过一定范围。具体讲静脉注射的剂量是1-1800mg/天。本发明公开的双特异性抗体及其组合物还可以和其他的抗肿瘤药联合给药以达到更加有效治疗肿瘤的目的,这些抗肿瘤药包括但不限于:1、细胞毒类药物:1)作用于核酸化学结构的药物:烷化剂如氮芥类、亚硝脲类、甲基磺酸酯类;铂类化合物如顺铂(cisplatin)、卡铂(carboplatin)和草酸铂(oxaliplatin)等;抗生素类如阿霉素(adriamycin/doxorubicin)、放线菌素d(dactinomycind)、柔红霉素(daunorubicin)、表阿霉素(epirubicin)、光辉霉素(mithramycin)等;2)影响核酸代谢的药物:二氢叶酸还原酶抑制剂如甲氨喋呤(mtx)和培美曲塞(pemetrexed)等;胸腺核苷合成酶抑制剂如氟尿嘧啶类(5-氟尿嘧啶、卡培他滨)等;嘌呤核苷合成酶抑制剂如6-巯基嘌呤等;核苷酸还原酶抑制剂如羟基脲(hydroxycarbamide)等;dna多聚酶抑制剂如阿糖胞苷(cytosinearabinoside)和吉西他滨(gemcitabine)等;3)作用于微管蛋白的药物:多西他赛(docetaxel)、长春花碱(vincristine)、长春瑞滨(vinorelbine)、鬼臼硷类、高三尖杉酯碱等;2、激素类药物:抗雌激素如他莫昔芬(tamoxifen)、屈洛昔芬(droloxifene)、依西美坦(exemestane)等;芳香化酶抑制剂如氨鲁米特(aminoglutethimide)、福美司坦(formestane)、来曲唑(letrozle)、阿那曲唑(anastrozole)等;抗雄激素:氟它氨rh-lh激动剂/拮抗剂:诺雷德、依那通等;3、生物反应调节剂类药物:此类药物主要通过调节机体免疫功能以到抗肿瘤的效果,如干扰素类(interferon);白细胞介素-2(interleukin-2);胸腺肽类(thymosins)等;4、单克隆抗体类药物:曲妥昔单抗(trastuzumab)、利妥昔单抗(rituximab)、西妥昔单抗(cetuximab)、贝伐单抗(bevacizumab)等;5、其他类抗肿瘤药物:包括一些目前机制尚不明确、有待进一步研究的药物等。本发明公开的双特异性抗体及其组合物可以和上述的抗肿瘤药物之一或其组合联合用药。在符合本领域常识的基础上,上述各优选条件,可任意组合,即得本发明各较佳实例。本发明所用试剂和原料均市售可得。本发明的积极进步效果在于:1、与单克隆抗体相比,本发明提供的抗egfr/pd-1双特异性抗体表现出协同效应,该双特异性抗体能同时结合egfr和pd-1,对靶细胞和抗原的亲和力与单克隆抗体相近,并且具有良好的生物活性,能抑制结肠癌细胞sw-48和皮肤鳞癌细胞a431的生长,并激活t淋巴细胞,促进t细胞对肿瘤细胞的杀伤。2、与抗体联用相比,抗体联用需要注射两种分开的产品或单次注射两种不同抗体的共制剂,而共制剂需要找到(在相对高浓度下)具有可接受的粘度且能促进两种抗体的化学和物理稳定性的配制条件,这是较为困难的,并且,这两种抗体联用的方式涉及两种药物的累加成本,都增加了患者的支付成本。本发明提供的抗egfr/pd-1双特异抗体不仅能同时靶向肿瘤细胞表面分子egfr和t淋巴细胞表面分子pd-1,而且对肿瘤细胞杀伤效果优于抗体联用。3、本发明的抗egfr/pd-1双特异性抗体的fab去糖基化对活性没有影响,降低了后续工艺产业化难度。4、该抗egfr/pd-1双特异性抗体可以单独或与其他抗肿瘤药物联合应用在肿瘤或癌症治疗中,即能够运用于抗肿瘤或癌症药物的制备中。附图说明图1:本发明抗egfr/pd-1双特异性抗体分子结构示意图。图2a:本发明抗egfr/pd-1双特异性抗体hplc检测图谱。图2b:本发明抗egfr/pd-1双特异性抗体的sds-page检测结果。从左至右三个泳道分别为非还原型样品抗体、分子量标记、还原型样品抗体。图3a:elisa检测本发明抗egfr/pd-1双特异性抗体与egfr-ecd-fc的结合。图3b:elisa检测本发明抗egfr/pd-1双特异性抗体与pd-1-ecd-fc的结合。图3c:elisa检测本发明抗egfr/pd-1双特异性抗体同时与egfr-ecd-fc和pd-1-ecd-fc的结合。图4a:本发明抗egfr/pd-1双特异性抗体与细胞表面抗原egfr结合活性。图4b:本发明抗egfr/pd-1双特异性抗体与细胞表面抗原pd-1结合活性。图5a:本发明抗egfr/pd-1双特异性抗体与egfr-ecd-his结合的动力学参数特征检测结果。图5b:本发明抗egfr/pd-1双特异性抗体与pd-1-ecd-his结合的动力学参数特征检测结果。图6:本发明抗egfr/pd-1双特异性抗体对a431细胞体外增殖抑制实验结果。图7:本发明抗egfr/pd-1双特异性抗体阻断pd-1与pd-l1结合的细胞实验结果。图8a:本发明抗egfr/pd-1双特异性抗体对a431肿瘤细胞的adcc效应实验结果。图8b:本发明抗egfr/pd-1双特异性抗体对cd4+t淋巴细胞的adcc效应实验结果。图9:本发明抗egfr/pd-1双特异性抗体在裸小鼠sw48结肠癌模型上抗egfr的药效学研究。图10:本发明抗egfr/pd-1双特异性抗体在人源化pd-1小鼠mc38结肠癌模型上的药效学研究。图11:本发明抗egfr/pd-1双特异性抗体介导pbmc对sw-48肿瘤细胞的杀伤研究。用*表示和不加抗体组比较存在显著性差异,用#表示和联合用药组比较存在显著性差异,其中*p<0.05,**p<0.01,***p<0.001,#p<0.05,##p<0.01,###p<0.001。图12:本发明抗egfr/pd-1双特异性抗体在大鼠体内药代动力学参数结果。图13a:elisa检测本发明抗egfr/pd-1双特异性抗体b、抗egfr/pd-1双特异性抗体c与egfr-ecd-fc的结合。图13b:elisa检测本发明抗egfr/pd-1双特异性b抗体、抗egfr/pd-1双特异性抗体c与pd-1-ecd-fc的结合。图13c:本发明抗egfr/pd-1双特异性抗体b和抗egfr/pd-1双特异性抗体c的fab分子量去卷积图谱。图13d:本发明抗egfr/pd-1双特异性抗体的毛细管等电聚焦图谱。图13e:本发明抗egfr/pd-1双特异性抗体b的毛细管等电聚焦图谱。图13f:本发明抗egfr/pd-1双特异性抗体c的毛细管等电聚焦图谱。具体实施方式以下实施例、实验例是对本发明进行进一步的说明,不应理解为是对本发明的限制。实施例不包括对传统方法的详细描述,如那些用于构建载体和质粒的方法,将编码蛋白的基因插入到这样的载体和质粒的方法或将质粒引入宿主细胞的方法.这样的方法对本领域中具有普通技术的人员是众所周知的,并且在许多出版物中都有所描述,包括sambrook,j.,fritsch,e.f.andmaniais,t.(1989)molecularcloning:alaboratorymanual,2ndedition,coldspringharborlaboratorypress.以下实施例中使用的实验材料和来源以及实验试剂的配制方法具体说明如下。实验材料:cho细胞:购自thermofisher公司,货号a29133。293e细胞:来自nrcbiotechnologyresearchinstitute。结肠癌细胞a431:购自atcc。pd-l1aapc/cho-k1细胞:购自promega公司,货号j1252。cd4+t细胞:购自allcells,货号lp180329。nk细胞:购自allcells公司,货号pb012-c。sw48结肠癌细胞株:购自中国科学院上海生命科学研究院。balb/c裸小鼠:购自上海灵畅生物科技有限公司。mc38小鼠结肠癌细胞株:购自和元生物技术(上海)股份有限公司。小鼠品系c57bj/6j-pdcd1em1(hpdcd1)/smoc:购自上海南方模式生物科技股份有限公司,产品编号:nm-ki-00015。pbmc细胞:购自allcells公司,货号pb005-c。sd大鼠:购自浙江维通利华实验动物技术有限公司,生产许可证scxk(浙)2018-0001。实验试剂:pbs:购自生工生物工程(上海)股份有限公司,货号b548117。bsa:购自生工生物工程(上海)股份有限公司,货号a60332。elisa包被液:0.1mnahco3,ph9.5。elisa封闭液:pbs+2%bsa。elisa稀释液:pbs+0.5%bsa。elisa终止液:50μl2mh2so4。pbst:pbs+0.05%tween20。cck-8显色液:购自dojindo,货号ck04。bio-glo:购自promega,货号g7940。fbs:购自gibco,货号10099。tmb:购自bd公司,货号555214。streptavidin:购自bd公司,货号554061。hrp标记的鼠抗人fab抗体:购自sigma,货号a0293。抗cd28抗体:购自abcam,货号ab213043。抗cd3抗体:r&d,#mab100。il-2:购自r&d,货号202-il。羊抗人igg-fitc:购自sigma,货号f4143。抗mhci阳性对照抗体:购自invivogen公司,货号hla-c1。proteina芯片:labelno:29139131-aa;lot:10261132。hbs-ep工作液:购自lifescience,br-1006-69。实验仪器:spectramaxi3x酶标仪:购自moleculardevices公司。spectramaxm5酶标仪:购自moleculardevices公司。beckmancoμltercytoflex流式细胞仪:购自beckman公司。hitrapmabselectsure柱:购自ge公司。本发明实施例中所述的egfr单克隆抗体均是指三生国健药业按照erbitux的氨基酸序列、用cho细胞表达系统进行表达、自主研发的细胞培养生产工艺得到的人鼠嵌合单克隆抗体。本发明实施例中所述的pd-1单克隆抗体均是指中国专利申请cn201710054783.5公开的由三生国健药业自主研发的全新的抗pd-1人源化单克隆抗体。实施例1抗egfr/pd-1双特异性抗体的分子构建本发明采用了scfv和igg串联的方式构建了抗egfr/pd-1双特异性抗体,具体形式为scfv-l2-igg,如图1所示。其中,scfv采用了vh-l1-vl的分子形式。将抗pd-1单克隆抗体的重链可变区vh(seqidno:13)和轻链可变区vl(seqidno:14)通过l1(seqidno:17)连接起来,得到抗pd-1的单链抗体片段scfv(seqidno:19)。利用l2(seqidno:18)将该单链抗体片段和抗egfr单克隆抗体的重链连接起来,从而得到抗egfr/pd-1双特异性抗体分子的重链(seqidno:20),egfr单抗的轻链(seqidno:21)则保持不变。为了提高抗体分子在cho细胞中的表达效率,委托金唯智公司对抗egfr/pd-1双特异性抗体分子的核酸序列进行密码子优化。优化主要考虑密码子的偏好性、gc含量、mrna二级结构、重复序列等因素,随后委托金唯智公司合成。抗egfr/pd-1双特异性抗体重链核酸序列为seqidno:22,轻链核酸序列为seqidno:23。实施例2抗egfr/pd-1双特异性抗体的表达与纯化将抗egfr/pd-1双特异性抗体重链和轻链的dna片段分别亚克隆到ptt5载体中,提取重组质粒共转染cho细胞和/或293e细胞。细胞培养7天后,将培养液通过高速离心、微孔滤膜抽真空过滤后上样至hitrapmabselectsure柱,用100mm柠檬酸,ph3.5的洗脱液一步洗脱蛋白,回收目标样品并透析换液至pbs。将纯化后的蛋白用hplc检测,如图2a所示,抗体分子状态均一,单体纯度达到97%以上。分别加入还原型蛋白电泳上样缓冲液和非还原型蛋白电泳上样缓冲液,煮沸后进行检测,结果如图2b所示,全长蛋白分子在250kd处(理论分子量198kd),重链在76kd处,轻链在23kd处。实施例3酶联免疫吸附法(elisa)测定抗egfr/pd-1双特异性抗体对抗原的亲和力3.1elisa检测抗egfr/pd-1双特异性抗体与egfr的亲和力将重组egfr-ecd-fc蛋白用包被液稀释至3μg/ml,以50μl/孔加入酶标板,4℃过夜。pbst洗板3次,加入200μl/孔封闭液,37℃放置1小时后pbst洗板1次待用。用稀释液稀释抗egfr/pd-1双特异性抗体至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。pbst洗板3次,加入hrp标记的鼠抗人fab抗体,37℃放置30分钟。pbst洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的tmb,室温(20±5℃)避光放置5分钟;每孔加入终止液终止底物反应,酶标仪450nm处读取od值,graphpadprism6进行数据分析,作图并计算ec50。实验结果如图3a所示,抗egfr/pd-1双特异性抗体和阳性对照egfr单抗与egfr-ecd结合的ec50分别为0.2091nm和0.2090nm,两者亲和力相当。3.2elisa检测抗egfr/pd-1双特异性抗体与pd-1的亲和力为了检测抗egfr/pd-1双特异性抗体与pd-1的结合能力,将重组pd-1-ecd-fc蛋白用包被液稀释至0.4μg/ml,50μl/孔加入酶标版,4℃过夜。pbst洗板3次,加入200μl/孔封闭液,37℃放置1小时后pbst洗板1次待用。用稀释液稀释抗egfr/pd-1双特异性抗体至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。pbst洗板3次,加入hrp标记的鼠抗人fab抗体,37℃放置30分钟。pbst洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的tmb,室温(20±5℃)避光放置5分钟;每孔加入50μl2mh2so4终止液终止底物反应,酶标仪450nm处读取od值,graphpadprism6进行数据分析,作图并计算ec50。实验结果如图3b所示,抗egfr/pd-1双特异性抗体和阳性对照pd-1单抗与pd-1-ecd结合的ec50分别为0.228nm和0.149nm,抗egfr/pd-1双特异性抗体亲和力与pd-1单抗相当。3.3elisa检测抗egfr/pd-1双特异性抗体同时结合pd-1和egfr的能力空间位阻可能会影响抗egfr/pd-1双特异性抗体同时结合两种抗原的能力。为了检测抗egfr/pd-1双特异性抗体同时结合pd-1和egfr的能力,将重组pd-1-ecd-fc蛋白用包被液稀释至0.4μg/ml,50μl/孔加入酶标版,4℃过夜。pbst洗板3次,加入200μl/孔封闭液,37℃放置1小时后pbst洗板1次待用。用稀释液稀释抗egfr/pd-1双特异性抗体至100μg/ml,4倍比稀释形成12个浓度梯度(最高浓度为100000ng/ml,最低浓度为0.02ng/ml),依次加入封闭后的酶标板,100μl/孔,37℃放置1小时。pbst洗板3次,按150ng/孔加入egfr-ecd-fc-biotin,37℃放置1小时。pbst洗板3次后加入hrp标记的streptavidin,37℃放置30分钟。pbst洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的tmb,室温(20±5℃)避光放置5分钟;每孔加入50μl2mh2so4终止液终止底物反应,酶标仪450nm处读取od值,graphpadprism6进行数据分析,作图并计算ec50。实验结果如图3c所示,抗egfr/pd-1双特异性抗体能同时结合egfr和pd-1,且ec50为0.2863nm。实施例4facs法测定抗egfr/pd-1双特异性抗体对靶细胞表面抗原的结合亲和力4.1facs法测定抗egfr/pd-1双特异性抗体对a431细胞表面egfr的结合亲和力本实验以细胞表面egfr高表达的结肠癌细胞a431作为靶细胞,将a431细胞按照2×105/孔接种于96孔板,用含有0.5%bsa的pbs洗涤三次,每次300g离心5分钟,弃上清。将100μl按照3倍梯度从3μg/ml连续稀释12个梯度的抗egfr/pd-1双特异性抗体作为一抗,并取按照3倍梯度从1μg/ml连续稀释12个梯度的egfr单抗作为阳性对照加入96孔板,将细胞悬浮后于4℃孵育1h。pbs洗涤细胞两次以去除未结合的抗egfr/pd-1双特异性抗体,再将细胞与100μl的1g/ml羊抗人igg-fitc于4℃孵育30分钟。300g离心5分钟,pbs洗涤细胞两次以去除未结合的二抗,最后将细胞重悬在200μlpbs中,通过beckmancoμltercytoflex流式细胞仪测定抗egfr/pd-1双特异性抗体对该细胞的结合亲和力。所得数据通过graphpadprism6软件拟合分析。实验结果如图4a所示,抗egfr/pd-1双特异性抗体和阳性对照egfr单抗能特异性的结合细胞表面表达的egfr,ec50分别为4.377nm和4.976nm,两者亲和力相当。4.2facs法测定抗egfr/pd-1双特异性抗体对pd-1稳转细胞株表面pd-1的结合亲和力同样的,以细胞表面表达pd-1的cho稳转细胞作为靶细胞,通过流式细胞仪测定抗egfr/pd-1双特异性抗体对该细胞表面pd-1的结合亲和力,方法如前所述,所得数据通过graphpadprism6软件拟合分析。实验结果如图4b所示,抗egfr/pd-1双特异性抗体和阳性对照pd-1单抗可以特异性的结合细胞表面表达的pd-1,ec50分别为1.528nm和1.618nm,两者亲和力相当。实施例5biacoretm8k测定抗egfr/pd-1双特异性抗体对抗原的亲和力使用proteina捕获法测定抗egfr/pd-1双特异性抗体和抗原egfr-ecd-his结合的动力学参数。将浓度为1μg/ml的抗egfr/pd-1双特异性抗体结合在proteina芯片上,将抗原egfr-ecd-his用1xhbs-ep工作液从50nm往下2倍稀释设6个浓度梯度与抗体结合,于hbs-ep工作液中解离。使用proteina捕获法测定抗egfr/pd-1双特异性抗体和抗原pd-1-ecd-his结合的动力学参数。将浓度为1μg/ml的抗egfr/pd-1双特异性抗体结合在proteina芯片上,将抗原pd-1-ecd-his用1xhbs-ep工作液从250nm往下2倍稀释设5个浓度梯度与抗体结合,于hbs-ep工作液中解离。抗egfr/pd-1双特异性抗体和egfr-ecd-his、pd-1-ecd-his结合的动力学参数见表1,动力学特征参数检测结果分别如图5a、5b所示。表1.抗egfr/pd-1双特异性抗体的动力学参数kd为亲和力常数;ka为抗原抗体结合速率;kd为抗原抗体解离速率;kd=kd/ka。文献anticancerresearch27:3355-3366(2007)中报道,biacore测定的cetuximab结合动力学参数为ka:2.2e+05m-1s-1,kd:1.1e-03s-1,kd:5.2e-09m。本发明公开的抗egfr/pd-1双特异性抗体的抗egfr部分包含了和cetuximab抗体可变区一致的氨基酸序列,实验结果表明,抗egfr/pd-1双特异性抗体与抗原egfr的结合动力学参数与文献报道一致。专利文献ep2161336a1中报道,biacore测定的nivolumab(编号5c4)结合动力学参数为ka:4.32e+05m-1s-1,kd:3.15e-04s-1,kd:0.73e-09m。实验结果表明,抗egfr/pd-1双特异性抗体与抗原pd-1的结合动力学参数与已上市药物nivolumab大致相当。综上所述,抗egfr/pd-1双特异性抗体与抗原egfr和pd-1具有良好的结合动力学性质。实施例6抗egfr/pd-1双特异性抗体对鳞癌a431细胞的体外增殖抑制作用a431细胞表面高表达egfr,抗egfr抗体能有效抑制a431细胞的体外增殖。根据细胞生长速率,将处于对数生长期的a431细胞以2.5×103/孔接种于96孔培养板,贴壁生长24h后,加入不同浓度的egfr单抗或抗egfr/pd-1双特异性抗体。egfr单抗的最高作用浓度为60nm,三倍稀释,共10个不同浓度,每个浓度设双复孔,并设空白对照及无细胞调零孔。细胞在37℃、5%co2条件下继续培养72h后,加入cck-8显色液20μl/孔,室温孵育1.5h。在spectramaxm5酶标仪上,以450nm为检测波长,650nm为参比波长,进行读数,用graphpadprism6进行数据分析,作图并计算ic50。实验结果如图6所示,抗egfr/pd-1双特异性抗体和阳性对照egfr单抗对a431细胞的ic50分别为0.809nm和0.931nm,两者抑制活性相当。实施例7抗egfr/pd-1双特异性抗体阻断pd-1与pd-l1结合的细胞实验取对数期生长的pd-l1aapc/cho-k1细胞,胰酶消化成单个细胞后转移到白色底透96孔板,100μl/孔,40000细胞/孔,置于37℃,5%co2,孵育过夜。将抗egfr/pd-1双特异性抗体、阳性对照pd-1单抗和同型阴性对照抗体igg1按三倍梯度稀释成2×工作液,最高浓度600nm,最低浓度0.09nm,共9个浓度梯度。同时,取密度在1.4-2×106/ml,细胞活率在95%以上的pd-1效应细胞胰酶消化成1.25×106细胞/ml的单细胞悬液。取前一天铺好的pd-l1aapc/cho-k1细胞,弃掉上清,加入40μl梯度稀释的抗体工作液,再加入等体积的pd-1效应细胞。置于37℃,5%co2,孵育6小时。细胞于37℃孵育6小时后,每孔加入80μl检测试剂bio-glo。室温孵育10分钟后,用spectramaxi3x读取luminescence。所有数据均为双复孔,所得信号值取平均值后用4-parameter法拟合,用graphpadprism6进行数据分析,作图并计算ic50。实验结果如图7所示,抗egfr/pd-1双特异性抗体和阳性对照pd-1单抗阻断pd-1与pd-l1结合的ic50分别为1.425nm和0.891nm,抗egfr/pd-1双特异性抗体的抑制活性与pd-1单抗相当。实施例8抗egfr/pd-1双特异性抗体对a431肿瘤细胞和t淋巴细胞的adcc效应8.1抗egfr/pd-1双特异性抗体对a431肿瘤细胞的adcc效应将a431细胞用含5%fbs的1640培养基稀释至2×105个/ml后,取50μl至96孔平底板中过夜。将最高浓度40μg/ml的抗egfr/pd-1双特异性抗体,和最高浓度30μg/ml的egfr单抗按4倍梯度稀释共9个浓度梯度,取50μl不同浓度的阳性对照egfr单抗或抗egfr/pd-1双特异性抗体于96孔板中,每个浓度设双复孔。细胞在37℃、5%co2条件下继续培养15分钟后,按照效应细胞(本实验中为nk细胞):靶细胞(本实验中为a431细胞)=5:1的比例加入nk细胞,在37℃、5%co2条件下继续培养3小时。300g离心后取100μl上清,加入50μl的ldh室温避光反应15分钟。酶标仪490nm处读取od值,graphpadprism6进行数据分析,作图并计算杀伤率。实验结果如图8a所示,抗egfr/pd-1双特异性抗体对a431肿瘤细胞具有明显的抗体依赖的细胞杀伤效果,和阳性对照egfr单抗相比,抗egfr/pd-1双特异性抗体和egfr单抗的ec50分别为1.367nm和1.731nm,两者活性相当。8.2抗egfr/pd-1双特异性抗体对cd4+t细胞的adcc效应cd4+t细胞的活化:将5μg/ml的抗cd3抗体包被到24孔板中,接种0.5m/孔的cd4+t细胞,并加入2μg/ml的抗cd28抗体和100u/ml的il-2,于37℃、5%co2条件下培养72小时。收集激活的cd4+t细胞,用facs检测pd-1表达水平。将激活的cd4+t细胞用含5%fbs的1640培养基稀释至2×105个/ml后,取50μl至96孔平底板中。将最高浓度80μg/ml的抗egfr/pd-1双特异性抗体,和最高浓度10μg/ml的抗mhci阳性对照抗体按4倍梯度稀释共10个浓度梯度,取50μl不同浓度的样品于96孔板中,每个浓度设双复孔。细胞在37℃、5%co2条件下继续培养15分钟后,按照效应细胞(本实验中为nk细胞):靶细胞(本实验中为cd4+t细胞)=5:1的比例加入nk细胞,在37℃、5%co2条件下继续培养3小时。300g离心后取100μl上清,加入50μl的ldh室温避光反应15分钟。酶标仪490nm处读取od值,graphpadprism6进行数据分析。实验结果如图8b所示,抗egfr/pd-1双特异性抗体对激活的cd4+t细胞没有明显的抗体依赖的细胞杀伤效果。实施例9抗egfr/pd-1双特异性抗体在sw48移植瘤模型上的抗肿瘤作用本实验用于评价抗egfr/pd-1双特异性抗体在动物模型上的抗egfr活性。sw48结肠癌细胞表面表达egfr,抗egfr抗体能有效抑制sw48细胞的增殖。阳性对照egfr单抗的剂量设置为25mg/kg,受试样品抗egfr/pd-1双特异性抗体的剂量设计为与egfr单抗等摩尔剂量,即34mg/kg,并设置一个低剂量组为6.8mg/kg。对照组给以相同体积的生理盐水。收集体外培养的结肠癌sw48细胞,将细胞悬液浓度调整为3×107/ml。在无菌条件下,接种100μl细胞悬液于balb/c裸小鼠右侧肋部皮下。移植瘤用游标卡尺测量移植瘤直径,待平均肿瘤体积生长至100-200mm3后将动物随机分组。egfr单抗和抗egfr/pd-1双特异性抗体按照上述剂量给药,对照组给等量生理盐水,每周腹腔注射给药3次,连续给药3周。整个实验过程中,每周2次测量移植瘤直径,同时称小鼠体重。肿瘤体积(tumorvolume,tv)的计算公式为:tv=1/2×a×b2。其中a、b分别表示长、宽。根据测量的结果计算出相对肿瘤体积(relativetumorvolume,rtv),计算公式为:rtv=vt/v0。其中v0为分组给药时(即d0)测量所得肿瘤体积,vt为每一次测量时的肿瘤体积。抗肿瘤活性的评价指标为相对肿瘤增殖率t/c(%),计算公式如下:t/c(%)=(trtv/crtv)×100(trtv:治疗组rtv;crtv:阴性对照组rtv)。疗效评价标准:t/c(%)>40%为无效;t/c(%)≤40,并经统计学处理p≤0.05为有效。实验重复两次。实验结果如图9所示,抗egfr/pd-1双特异性抗体在34mg/kg剂量时抑瘤率为53.7%,大致相同摩尔剂量下的egfr单抗(25mg/kg)抑瘤率为63.6%,抗egfr/pd-1双特异性抗体在低剂量为6.8mg/kg时抑瘤率为48.2%。结果表明,在sw48移植瘤模型上,抗egfr/pd-1双特异性抗体通过阻断egfr抑制肿瘤生长。在相等摩尔剂量下,抗egfr/pd-1双特异性抗体和阳性对照egfr单抗的抑瘤效果相当,低剂量时(6.8mg/kg),抗egfr/pd-1双特异性抗体也有抑瘤活性。实施例10抗egfr/pd-1双特异性抗体在人源化pd-1小鼠mc38移植瘤模型上的抗肿瘤作用本实验用于评价抗egfr/pd-1双特异性抗体在动物模型上的抗pd-1活性。采用人源化pd-1小鼠品系c57bj/6j-pdcd1em1(hpdcd1)/smoc。阳性对照pd-1单抗的剂量设置为10mg/kg,受试抗egfr/pd-1双特异性抗体的剂量设计为与pd-1单抗等摩尔剂量,即16mg/kg,对照组给以相同体积的生理盐水。收集体外培养的mc38细胞,将细胞悬液浓度调整为1×107/ml。在无菌条件下,接种100μl细胞悬液于人源化pd-1小鼠右侧肋部皮下。人源化pd-1小鼠皮下移植瘤用游标卡尺测量移植瘤直径,待平均肿瘤体积生长至100-200mm3后将动物随机分组。pd-1单抗、抗egfr/pd-1双特异性抗体按剂量给药、对照组给等量生理盐水,每周腹腔注射给药2次,连续给药3周。整个实验过程中,每周2次测量移植瘤直径,同时称小鼠体重。肿瘤体积(tumorvolume,tv)的计算公式为:tv=1/2×a×b2。其中a、b分别表示长、宽。根据测量的结果计算出相对肿瘤体积(relativetumorvolume,rtv),计算公式为:rtv=vt/v0。其中v0为分组给药时(即d0)测量所得肿瘤体积,vt为每一次测量时的肿瘤体积。抗肿瘤活性的评价指标为相对肿瘤增殖率t/c(%),计算公式如下:t/c(%)=(trtv/crtv)×100(trtv:治疗组rtv;crtv:阴性对照组rtv)。疗效评价标准:t/c(%)>40%为无效;t/c(%)≤40,并经统计学处理p≤0.05为有效。实验重复两次。实验结果如图10所示,第21天时,抗egfr/pd-1双特异性抗体在16mg/kg剂量时抑瘤率为91.5%,大致相同摩尔剂量下的pd-1单抗(10mg/kg)抑瘤率为85.2%。结果表明,在人源化pd-1小鼠mc38移植瘤模型上,抗egfr/pd-1双特异性抗体可以通过阻断pd-1抑制肿瘤生长,且抑瘤效果与阳性对照pd-1单抗相当。实施例11抗egfr/pd-1双特异性抗体介导pbmc对sw-48肿瘤细胞的杀伤收集培养好的sw-48细胞,用rpmi1640培养基重悬稀释,于96孔板中每孔接种1×104细胞,37℃、5%co2条件下培养过夜。pbmc用rpmi1640培养基重悬稀释(pbmc捐赠者编号分别为#d180198和#d180199),按照效应细胞(本实验中为pbmc细胞):靶细胞(本实验中为sw-48细胞)=20:1的比例,每孔加入2×105pbmc细胞。实验组加入样品抗egfr/pd-1双特异性抗体使终浓度为100nm、10nm和1nm,抗体联用组为egfr单抗和pd-1单抗按1:1混合且终浓度均为100nm,单独用药组为终浓度100nm的egfr单抗或pd-1单抗,空白对照组不加任何抗体,在37℃、5%co2条件下继续培养5天。用pbs清洗3遍,每孔加入200μlpbs和50μl检测试剂bio-glo。室温孵育5分钟后,用spectramaxi3x读取luminescence,用graphpadprism6进行数据分析,用*表示和不加抗体组比较存在显著性差异,用#表示和联合用药组比较存在显著性差异。实验数据如图11所示,和阴性空白对照组相比,100nm的抗egfr/pd-1双特异性抗体能显著介导pbmc对sw-48肿瘤细胞的杀伤(***p<0.001,p为偏差),并且杀伤效果呈现剂量依赖,高剂量组(终浓度为100nm)明显优于低剂量组(终浓度为10nm和1nm)。和单独用药组相比,抗egfr/pd-1双特异性抗体明显优于egfr单抗和pd-1单抗,和抗体联用组相比,100nm的抗egfr/pd-1双特异性抗体也明显优于egfr单抗加pd-1单抗的药物组合(两次实验分别为#p<0.05和###p<0.001),表明抗egfr/pd-1双特异性抗体在介导pbmc对sw-48肿瘤细胞的杀伤过程中具有协同效应。实施例12抗egfr/pd-1双特异性抗体的药代动力学研究大致实验方法和步骤如下,每组4只sd大鼠,体重200g左右,每只大鼠通过尾静脉注射剂量为2mg的抗egfr/pd-1双特异性抗体;分别在给药后的特定时间眼眶取血,血液自然凝固后8000rpm/min离心取血清。血清中抗体药物浓度采用三种测量方法检测,方法1和方法2的原理依赖抗egfr/pd-1双特异性抗体同时结合两种抗原的能力,能反映血浆中完整的抗体分子的含量变化,方法3则用来检测血浆中总体的抗egfr/pd-1双特异性抗体和抗egfr/pd-1双特异性抗体抗体片段含量变化。方法1)包被egfr,检测pd-1。用egfr-ecd-fc包被elisa板4℃过夜,包被量为150ng/孔;包被完毕后用pbst洗板3次,然后用pbs+2%bsa于37℃封闭2小时;然后加入2000倍稀释的大鼠血清孵育1小时,取大鼠空白血清作为对照;pbst洗板3次后加入pd-1-ecd-fc-biotin孵育1小时,包被量为7.5ng/孔;洗板并最终加入hrp标记的鼠抗人抗体,37℃放置30分钟;pbst洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的tmb,室温(20±5℃)避光放置5分钟;每孔加入50μl2m的h2so4终止液终止底物反应,酶标仪450nm处读取od值,数据处理时扣除空白血浆的od值。方法2)包被pd-1,检测egfr。用pd-1-ecd-fc包被elisa板4℃过夜,包被量为20ng/孔;包被完毕后用pbst洗板3次,然后用pbs+2%bsa于37℃封闭2小时;然后加入2000倍稀释的大鼠血清孵育1小时,取大鼠空白血清作为对照;pbst洗板3次洗板后加入hrp标记的streptavidin,37℃放置30分钟;pbst洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的tmb,室温(20±5℃)避光放置5分钟;每孔加入50μl2mh2so4终止液终止底物反应,酶标仪450nm处读取od值,数据处理时扣除空白血浆的od值。方法3)包被proteina,检测抗体fab段。用proteina包被elisa板4℃过夜,包被量为100ng/孔;包被完毕后用pbst洗板3次,然后用pbs+2%bsa于37℃封闭2小时;然后加入2000倍稀释的大鼠血清孵育1小时,取大鼠空白血清作为对照;pbst洗板3次洗板后加入hrp标记的鼠抗人fab抗体,37℃放置30分钟;pbst洗板3次后,在吸水纸上尽量拍干残留液滴,每孔加入100μl的tmb,室温(20±5℃)避光放置5分钟;每孔加入50μl2m的h2so4终止液终止底物反应,酶标仪450nm处读取od值,数据处理时扣除空白血浆的od值。用phoenix软件计算抗体药物在大鼠体内的半衰期,用graphpadprism6进行数据整理分析和作图。药代动力学参数见表2,实验结果如表2和图12所示,方法1和方法2检测的大鼠体内半衰期分别为173和171小时,方法3检测的大鼠体内半衰期为298小时。方法3检测的半衰期较长,可能的解释是:1)抗egfr/pd-1双特异性抗体在血浆中可能降解为scfv和igg等抗体片段,运用方法3检测到的是血浆中抗egfr/pd-1双特异性抗体和igg抗体片段的总体含量变化;2)方法3检测方法更加敏感,实验结果包含系统误差。表2.抗egfr/pd-1双特异性抗体在大鼠体内药代动力学参数grouphl_lambda_z(hr)cmax(ng/μl)auclast(hr*ng/μl)aucinf_obs(hr*ng/μl)vz_obs(μl/kg)cl_obs(μl/hr/kg)mrtlast(hr)1173.82488554848065872.9592578278.554532461.149130.8323552171.458103365656840.575046.3792545154.8225137.52142159.20419253298.3044351264.5191849.7225286251.69514937.1462535.2596775181.62501实施例13抗egfr/pd-1双特异性抗体的fab去糖基化对活性的影响抗egfr/pd-1双特异性抗体分子重链的第347位天冬酰胺存在糖基化,增加了抗体分子的电荷异质性。为了提高抗体分子的均一性,方便生产制备工艺的开发,将347位天冬酰胺突变为谷氨酸,得到抗体分子抗egfr/pd-1双特异性抗体b(重链氨基酸序列seqidno:24),将347位天冬酰胺突变为天冬氨酸,得到抗体分子抗egfr/pd-1双特异性抗体c(重链氨基酸序列seqidno:25)。质谱鉴定表明突变后347位无糖基化,实验数据如图13c和表4。蛋白表达纯化方法与实施例2相同,elisa检测抗egfr/pd-1双特异性抗体b和抗egfr/pd-1双特异性抗体c与抗原的结合方法与实施例3相同。抗原亲和力实验结果见表3和图13a、13b。和抗egfr/pd-1双特异性抗体相比,抗egfr/pd-1双特异性抗体b和抗egfr/pd-1双特异性抗体c与抗原的亲和力没有明显变化,表明fab去糖基化对抗egfr/pd-1双特异性抗体的活性没有明显的影响。毛细管等电聚焦(i-cief)实验如图13d、13e、13f显示,与抗egfr/pd-1双特异性抗体相比,抗egfr/pd-1双特异性抗体b和抗egfr/pd-1双特异性抗体c的电荷异质性明显减少,降低了后续工艺开发的难度,为后续的规模化大生产提供了可能性。表3.elisa检测抗原亲和力结果表4.抗egfr/pd-1双特异性抗体b和抗egfr/pd-1双特异性抗体c的fab分子量测定样品名称重链fab理论分子量重链fab测试分子量抗egfr/pd-1双抗b50582.2750581.70抗egfr/pd-1双抗c50581.2950580.80由上述实验可知,本发明的抗egfr/pd-1双特异性抗体在对靶细胞和抗原的亲和力、对鳞癌a431细胞的抑制活性、在sw48移植瘤模型上的抑瘤率都与阳性抗egfr单抗或抗pd-1单抗相当;本发明抗egfr/pd-1双特异性抗体对激活的cd4+t细胞没有明显的抗体依赖的细胞杀伤效果;在人源化pd1小鼠mc38移植瘤模型上,抗egfr/pd-1双特异性抗体可以阻断pd1移植肿瘤生长,且抑瘤率略优于阳性对照。并且,100nm的抗egfr/pd-1双特异性抗体能显著介导pbmc对sw-48肿瘤细胞的杀伤(***p<0.001),和抗体联用组相比,100nm的抗egfr/pd-1双特异性抗体也明显优于egfr单抗加pd-1单抗的药物组合(两次实验分别为#p<0.05和###p<0.001),并且杀伤效果呈现剂量依赖,高剂量组(终浓度为100nm)明显优于低剂量组(终浓度为10nm和1nm)。表明抗egfr/pd-1双特异性抗体在介导pbmc对sw-48肿瘤细胞的杀伤过程中具有协同效应。本发明的抗egfr/pd-1双特异性抗体的fab去糖基化对活性没有影响,降低了后续工艺开发的难度,为后续的规模化大生产提供了可能性。序列表<110>沈阳三生制药有限责任公司<120>抗egfr/pd-1双特异性抗体<130>201811<160>25<170>siposequencelisting1.0<210>1<211>5<212>prt<213>composite<400>1sertyraspmetser15<210>2<211>16<212>prt<213>composite<400>2thrileserglyglyglyargtyrthrtyrtyrproaspthrvallys151015<210>3<211>8<212>prt<213>composite<400>3protyrglyglytyrpheaspval15<210>4<211>11<212>prt<213>composite<400>4argalaserglnserileserasnpheleuhis1510<210>5<211>7<212>prt<213>composite<400>5tyralaserglnserileser15<210>6<211>9<212>prt<213>composite<400>6glnglnserasnsertrpprohisthr15<210>7<211>7<212>prt<213>composite<400>7leuthrasntyrglyvalhis15<210>8<211>21<212>prt<213>composite<400>8valiletrpserglyglyasnthrasptyrasnthrprophethrser151015argleuserileasn20<210>9<211>13<212>prt<213>composite<400>9alaargalaleuthrtyrtyrasptyrgluphealatyr1510<210>10<211>11<212>prt<213>composite<400>10argalaserglnserileglythrasnilehis1510<210>11<211>7<212>prt<213>composite<400>11tyralasergluserileser15<210>12<211>9<212>prt<213>composite<400>12glnglnasnasnasntrpprothrthr15<210>13<211>117<212>prt<213>composite<400>13gluvallysleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphealaphesersertyr202530aspmetsertrpvalargglnalaproglylysargleuglutrpval354045alathrileserglyglyglyargtyrthrtyrtyrproaspthrval505560lysglyargphethrileserargaspasnalalysasnserhistyr65707580leuglnmetasnserleuargalagluaspthralavaltyrphecys859095alaserprotyrglyglytyrpheaspvaltrpglyglnglythrleu100105110valthrvalserser115<210>14<211>107<212>prt<213>composite<400>14gluilevalleuthrglnserproalathrleuserleuserprogly151015gluargalathrleusercysargalaserglnserileserasnphe202530leuhistrptyrglnglnlysproglyglnalaproargleuleuile354045lystyralaserglnserileserglyileproalaargphesergly505560serglyserglythraspphethrleuthrileserserleuglupro65707580gluaspphealavaltyrphecysglnglnserasnsertrpprohis859095thrpheglyglnglythrlysvalgluilelys100105<210>15<211>119<212>prt<213>composite<400>15glnvalglnleulysglnserglyproglyleuvalglnprosergln151015serleuserilethrcysthrvalserglypheserleuthrasntyr202530glyvalhistrpvalargglnserproglylysglyleuglutrpleu354045glyvaliletrpserglyglyasnthrasptyrasnthrprophethr505560serargleuserileasnlysaspasnserlysserglnvalphephe65707580lysmetasnserleuglnserasnaspthralailetyrtyrcysala859095argalaleuthrtyrtyrasptyrgluphealatyrtrpglyglngly100105110thrleuvalthrvalserala115<210>16<211>107<212>prt<213>composite<400>16aspileleuleuthrglnserprovalileleuservalserprogly151015gluargvalserphesercysargalaserglnserileglythrasn202530ilehistrptyrglnglnargthrasnglyserproargleuleuile354045lystyralasergluserileserglyileproserargphesergly505560serglyserglythraspphethrleuserileasnservalgluser65707580gluaspilealaasptyrtyrcysglnglnasnasnasntrpprothr859095thrpheglyalaglythrlysleugluleulys100105<210>17<211>20<212>prt<213>composite<400>17glyglyglyglyserglyglyglyglyserglyglyglyglysergly151015glyglyglyser20<210>18<211>15<212>prt<213>composite<400>18glyglyglyglyserglyglyglyglyserglyglyglyglyser151015<210>19<211>244<212>prt<213>composite<400>19gluvallysleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphealaphesersertyr202530aspmetsertrpvalargglnalaproglylysargleuglutrpval354045alathrileserglyglyglyargtyrthrtyrtyrproaspthrval505560lysglyargphethrileserargaspasnalalysasnserhistyr65707580leuglnmetasnserleuargalagluaspthralavaltyrphecys859095alaserprotyrglyglytyrpheaspvaltrpglyglnglythrleu100105110valthrvalserserglyglyglyglyserglyglyglyglysergly115120125glyglyglyserglyglyglyglysergluilevalleuthrglnser130135140proalathrleuserleuserproglygluargalathrleusercys145150155160argalaserglnserileserasnpheleuhistrptyrglnglnlys165170175proglyglnalaproargleuleuilelystyralaserglnserile180185190serglyileproalaargpheserglyserglyserglythraspphe195200205thrleuthrileserserleugluprogluaspphealavaltyrphe210215220cysglnglnserasnsertrpprohisthrpheglyglnglythrlys225230235240valgluilelys<210>20<211>708<212>prt<213>composite<400>20gluvallysleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphealaphesersertyr202530aspmetsertrpvalargglnalaproglylysargleuglutrpval354045alathrileserglyglyglyargtyrthrtyrtyrproaspthrval505560lysglyargphethrileserargaspasnalalysasnserhistyr65707580leuglnmetasnserleuargalagluaspthralavaltyrphecys859095alaserprotyrglyglytyrpheaspvaltrpglyglnglythrleu100105110valthrvalserserglyglyglyglyserglyglyglyglysergly115120125glyglyglyserglyglyglyglysergluilevalleuthrglnser130135140proalathrleuserleuserproglygluargalathrleusercys145150155160argalaserglnserileserasnpheleuhistrptyrglnglnlys165170175proglyglnalaproargleuleuilelystyralaserglnserile180185190serglyileproalaargpheserglyserglyserglythraspphe195200205thrleuthrileserserleugluprogluaspphealavaltyrphe210215220cysglnglnserasnsertrpprohisthrpheglyglnglythrlys225230235240valgluilelysglyglyglyglyserglyglyglyglyserglygly245250255glyglyserglnvalglnleulysglnserglyproglyleuvalgln260265270proserglnserleuserilethrcysthrvalserglypheserleu275280285thrasntyrglyvalhistrpvalargglnserproglylysglyleu290295300glutrpleuglyvaliletrpserglyglyasnthrasptyrasnthr305310315320prophethrserargleuserileasnlysaspasnserlyssergln325330335valphephelysmetasnserleuglnserasnaspthralailetyr340345350tyrcysalaargalaleuthrtyrtyrasptyrgluphealatyrtrp355360365glyglnglythrleuvalthrvalseralaalaserthrlysglypro370375380servalpheproleualaproserserlysserthrserglyglythr385390395400alaalaleuglycysleuvallysasptyrpheprogluprovalthr405410415valsertrpasnserglyalaleuthrserglyvalhisthrphepro420425430alavalleuglnserserglyleutyrserleuserservalvalthr435440445valproserserserleuglythrglnthrtyrilecysasnvalasn450455460hislysproserasnthrlysvalasplysargvalgluprolysser465470475480cysasplysthrhisthrcysproprocysproalaprogluleuleu485490495glyglyproservalpheleupheproprolysprolysaspthrleu500505510metileserargthrprogluvalthrcysvalvalvalaspvalser515520525hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu530535540valhisasnalalysthrlysproargglugluglntyrasnserthr545550555560tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn565570575glylysglutyrlyscyslysvalserasnlysalaleuproalapro580585590ileglulysthrileserlysalalysglyglnproarggluprogln595600605valtyrthrleuproproserarggluglumetthrlysasnglnval610615620serleuthrcysleuvallysglyphetyrproseraspilealaval625630635640glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro645650655provalleuaspseraspglyserphepheleutyrserlysleuthr660665670valasplysserargtrpglnglnglyasnvalphesercysserval675680685methisglualaleuhisasnhistyrthrglnlysserleuserleu690695700serproglylys705<210>21<211>214<212>prt<213>composite<400>21aspileleuleuthrglnserprovalileleuservalserprogly151015gluargvalserphesercysargalaserglnserileglythrasn202530ilehistrptyrglnglnargthrasnglyserproargleuleuile354045lystyralasergluserileserglyileproserargphesergly505560serglyserglythraspphethrleuserileasnservalgluser65707580gluaspilealaasptyrtyrcysglnglnasnasnasntrpprothr859095thrpheglyalaglythrlysleugluleulysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucys210<210>22<211>2124<212>dna<213>composite<400>22gaagtcaagctggtggaatccggaggcggactcgtgcagcccggaggatctttaaggctg60tcttgtgctgccagcggctttgccttctccagctacgacatgagctgggtgcggcaagcc120cccggtaagaggctcgaatgggtggctaccatttccggcggcggacggtacacctactat180cccgacacagtcaagggtcgtttcaccatctcccgggataacgccaagaattcccactat240ttacagatgaatagcctccgggccgaggataccgccgtgtatttttgcgcctccccctac300ggcggctactttgacgtgtggggtcaaggtacactggtcacagtgtctagcggaggcgga360ggaagcggaggtggcggcagcggcggaggtggtagcggaggaggaggatccgaaatcgtg420ctgacccagagccccgctacactgtctttatcccccggagagagggctaccctcagctgt480cgggccagccagtccatcagcaacttcctccactggtatcagcagaaacccggccaagct540cctaggttattaatcaagtacgccagccagagcatctccggcattcccgctcgtttttcc600ggcagcggaagcggaaccgactttactttaaccatctccagcctcgaacccgaagacttc660gctgtgtacttttgccagcagtccaacagctggccccacacattcggccaaggaaccaag720gtcgagatcaaaggaggcggcggctctggtggcggaggctccggtggcggaggatcccaa780gttcagctgaagcagagcggacccggtttagtgcaacccagccaatctttatccatcact840tgtacagtctccggcttcagcctcacaaactacggcgtgcactgggtgaggcaatctccc900ggtaagggactggagtggctgggagtgatttggtccggcggcaacaccgattacaacacc960cccttcacatctcgtctgagcatcaacaaggacaatagcaagagccaagttttcttcaaa1020atgaactctttacagtccgaggacaccgccatttactattgcgctcgtgctctcacctac1080tatgactatgaattcgcctactggggccaaggcactttagtcaccgtgtccgccgctagc1140acaaaaggcccctccgtgtttcctctcgcccctagcagcaaatccaccagcggaggcaca1200gctgctctgggctgcctcgtgaaggactattttcccgagcccgtgaccgtctcttggaac1260tccggagctctcaccagcggcgtgcacacattccccgctgtgctgcagagcagcggttta1320tactctttatccagcgtggtcaccgtgcccagcagctctttaggaacccagacatacatc1380tgcaacgtgaatcacaagcccagcaacaccaaagtcgataagcgggtggaacctaaatct1440tgtgataagacccatacttgtcctccttgtcccgctcccgaactcctcggcggcccttcc1500gtgttcctcttccctcccaagcctaaggataccctcatgatcagccggacacccgaagtg1560acttgtgtggtggtcgatgtgagccatgaggaccccgaggtgaagttcaactggtacgtg1620gacggcgtggaggtgcacaacgctaagaccaagcctcgggaagagcagtacaacagcaca1680tatagggtggtcagcgtcctcaccgtgctccatcaagattggctgaacggcaaggaatac1740aagtgtaaggtgtccaacaaggctttacccgcccccattgagaagaccatctccaaagct1800aagggccaacctcgggaaccccaagtttataccctcccccctagccgggaggagatgacc1860aaaaatcaagtttctttaacatgtctggtgaagggcttctaccccagcgacatcgccgtg1920gaatgggagtccaacggccaacccgagaacaactataagaccaccccccccgttttagac1980tccgatggttctttttttttatatagcaagctcaccgtggataagagccggtggcagcaa2040ggtaatgtgttttcttgttccgtgatgcacgaggctttacacaatcactacacccagaag2100tctttatccctctcccccggtaag2124<210>23<211>642<212>dna<213>composite<400>23gacattttactgacccagtcccccgtgattctctccgtgtcccccggtgagcgggtgagc60ttctcttgtcgtgccagccagtccatcggcaccaacatccactggtaccagcagaggacc120aacggatccccccggctgctgatcaagtacgccagcgagtccatcagcggcatcccttct180cgtttctccggatccggatccggcaccgacttcactttaagcatcaacagcgtggagagc240gaggacatcgccgattactactgccagcagaacaacaactggcccaccaccttcggcgct300ggcaccaagctggagctgaaaaggaccgtggccgccccctccgtgttcatcttccccccc360tccgatgaacagctgaagagcggaaccgcctccgtggtgtgtttactgaacaacttctac420ccccgggaggccaaggtgcagtggaaggtggacaacgctttacagagcggcaactcccaa480gaaagcgtgaccgagcaagatagcaaggacagcacctactctttatccagcactttaact540ttaagcaaggccgactacgagaaacacaaggtgtacgcttgtgaggtgacacaccaaggt600ttatcctcccccgttacaaagtccttcaatcggggcgagtgc642<210>24<211>708<212>prt<213>composite<400>24gluvallysleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphealaphesersertyr202530aspmetsertrpvalargglnalaproglylysargleuglutrpval354045alathrileserglyglyglyargtyrthrtyrtyrproaspthrval505560lysglyargphethrileserargaspasnalalysasnserhistyr65707580leuglnmetasnserleuargalagluaspthralavaltyrphecys859095alaserprotyrglyglytyrpheaspvaltrpglyglnglythrleu100105110valthrvalserserglyglyglyglyserglyglyglyglysergly115120125glyglyglyserglyglyglyglysergluilevalleuthrglnser130135140proalathrleuserleuserproglygluargalathrleusercys145150155160argalaserglnserileserasnpheleuhistrptyrglnglnlys165170175proglyglnalaproargleuleuilelystyralaserglnserile180185190serglyileproalaargpheserglyserglyserglythraspphe195200205thrleuthrileserserleugluprogluaspphealavaltyrphe210215220cysglnglnserasnsertrpprohisthrpheglyglnglythrlys225230235240valgluilelysglyglyglyglyserglyglyglyglyserglygly245250255glyglyserglnvalglnleulysglnserglyproglyleuvalgln260265270proserglnserleuserilethrcysthrvalserglypheserleu275280285thrasntyrglyvalhistrpvalargglnserproglylysglyleu290295300glutrpleuglyvaliletrpserglyglyasnthrasptyrasnthr305310315320prophethrserargleuserileasnlysaspasnserlyssergln325330335valphephelysmetasnserleuglnsergluaspthralailetyr340345350tyrcysalaargalaleuthrtyrtyrasptyrgluphealatyrtrp355360365glyglnglythrleuvalthrvalseralaalaserthrlysglypro370375380servalpheproleualaproserserlysserthrserglyglythr385390395400alaalaleuglycysleuvallysasptyrpheprogluprovalthr405410415valsertrpasnserglyalaleuthrserglyvalhisthrphepro420425430alavalleuglnserserglyleutyrserleuserservalvalthr435440445valproserserserleuglythrglnthrtyrilecysasnvalasn450455460hislysproserasnthrlysvalasplysargvalgluprolysser465470475480cysasplysthrhisthrcysproprocysproalaprogluleuleu485490495glyglyproservalpheleupheproprolysprolysaspthrleu500505510metileserargthrprogluvalthrcysvalvalvalaspvalser515520525hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu530535540valhisasnalalysthrlysproargglugluglntyrasnserthr545550555560tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn565570575glylysglutyrlyscyslysvalserasnlysalaleuproalapro580585590ileglulysthrileserlysalalysglyglnproarggluprogln595600605valtyrthrleuproproserarggluglumetthrlysasnglnval610615620serleuthrcysleuvallysglyphetyrproseraspilealaval625630635640glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro645650655provalleuaspseraspglyserphepheleutyrserlysleuthr660665670valasplysserargtrpglnglnglyasnvalphesercysserval675680685methisglualaleuhisasnhistyrthrglnlysserleuserleu690695700serproglylys705<210>25<211>708<212>prt<213>composite<400>25gluvallysleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphealaphesersertyr202530aspmetsertrpvalargglnalaproglylysargleuglutrpval354045alathrileserglyglyglyargtyrthrtyrtyrproaspthrval505560lysglyargphethrileserargaspasnalalysasnserhistyr65707580leuglnmetasnserleuargalagluaspthralavaltyrphecys859095alaserprotyrglyglytyrpheaspvaltrpglyglnglythrleu100105110valthrvalserserglyglyglyglyserglyglyglyglysergly115120125glyglyglyserglyglyglyglysergluilevalleuthrglnser130135140proalathrleuserleuserproglygluargalathrleusercys145150155160argalaserglnserileserasnpheleuhistrptyrglnglnlys165170175proglyglnalaproargleuleuilelystyralaserglnserile180185190serglyileproalaargpheserglyserglyserglythraspphe195200205thrleuthrileserserleugluprogluaspphealavaltyrphe210215220cysglnglnserasnsertrpprohisthrpheglyglnglythrlys225230235240valgluilelysglyglyglyglyserglyglyglyglyserglygly245250255glyglyserglnvalglnleulysglnserglyproglyleuvalgln260265270proserglnserleuserilethrcysthrvalserglypheserleu275280285thrasntyrglyvalhistrpvalargglnserproglylysglyleu290295300glutrpleuglyvaliletrpserglyglyasnthrasptyrasnthr305310315320prophethrserargleuserileasnlysaspasnserlyssergln325330335valphephelysmetasnserleuglnserglnaspthralailetyr340345350tyrcysalaargalaleuthrtyrtyrasptyrgluphealatyrtrp355360365glyglnglythrleuvalthrvalseralaalaserthrlysglypro370375380servalpheproleualaproserserlysserthrserglyglythr385390395400alaalaleuglycysleuvallysasptyrpheprogluprovalthr405410415valsertrpasnserglyalaleuthrserglyvalhisthrphepro420425430alavalleuglnserserglyleutyrserleuserservalvalthr435440445valproserserserleuglythrglnthrtyrilecysasnvalasn450455460hislysproserasnthrlysvalasplysargvalgluprolysser465470475480cysasplysthrhisthrcysproprocysproalaprogluleuleu485490495glyglyproservalpheleupheproprolysprolysaspthrleu500505510metileserargthrprogluvalthrcysvalvalvalaspvalser515520525hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu530535540valhisasnalalysthrlysproargglugluglntyrasnserthr545550555560tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn565570575glylysglutyrlyscyslysvalserasnlysalaleuproalapro580585590ileglulysthrileserlysalalysglyglnproarggluprogln595600605valtyrthrleuproproserarggluglumetthrlysasnglnval610615620serleuthrcysleuvallysglyphetyrproseraspilealaval625630635640glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro645650655provalleuaspseraspglyserphepheleutyrserlysleuthr660665670valasplysserargtrpglnglnglyasnvalphesercysserval675680685methisglualaleuhisasnhistyrthrglnlysserleuserleu690695700serproglylys705当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 与CD56分子特异性结合的多肽及其应用的制造方法与工艺
- 用于治疗HBV感染和相关病症的三特异性结合分子的制造方法与工艺
- 对磺胺类药物具有特异性的固相微萃取纤维的制备方法与流程
- 避免信号干扰的肌原纤维蛋白丝氨酸磷酸化水平的测定方法与流程
- 一种检测猪初乳中抗PEDV特异性IgA抗体的夹心ELISA试剂盒的制造方法与工艺
- 用于施用CD19xCD3双特异性抗体的给药方案的制造方法与工艺
- 双特异性抗‑VEGF/抗‑ANG‑2抗体及其在治疗眼血管疾病中的应用的制造方法与工艺
- 特异性人EWS‑FLI1融合蛋白抗体的制备方法及在制备尤文肉瘤诊断抗体试剂的应用与制造工艺
- 活化人CD40的抗体和针对人PD‑L1的抗体的组合疗法的制造方法与工艺
- 双特异性四价抗体及其制造和使用方法与制造工艺