一种特异性结合短肽及其应用的制作方法
本发明属于蛋白质多肽技术领域,具体涉及一种特异性结合短肽及其应用。
背景技术:
突触是神经元之间相互接触并进行信息传递的关键部位,在神经系统正常活动中起着十分重要的调节作用。根据对下一级神经元活动的影响,可以把突触分为兴奋性突触和抑制性突触。皮层肌动蛋白(cortactin)是一种微丝结合蛋白,在神经突触位置表达丰富,在突触发生、突触成熟和突触重塑过程发挥重要作用。
皮层肌动蛋白由ems1基因编码,在sds-聚丙烯酰胺凝胶电泳中显示2条电泳条带,分别位于80kda和85kda,可能与蛋白质翻译后修饰有关,磷酸化和乙酰化修饰是其发挥调节作用的前提。其中,乙酰化修饰受乙酰转移酶pcaf和去乙酰转移酶hdac6调节,皮层肌动蛋白的乙酰化修饰能够增加其与f-actin的结合能力。在神经元中,皮层肌动蛋白在突触的分布也受到乙酰化的调节。当它乙酰化水平降低时,破坏兴奋性突触形态、结构和功能,阻碍神经元之间信息传递。
技术实现要素:
有鉴于此,本发明的目的在于提供一种特异性结合短肽及其应用,可通过与皮层肌动蛋白的特异结合,有效介导皮层肌动蛋白乙酰化,维持兴奋性突触形态结构与功能的稳定,起神经保护作用。
为了实现上述发明目的,本发明提供以下技术方案:
本发明提供了一种特异性结合短肽,氨基酸序列如seqidno.1所示。
优选的,所述特异性结合短肽由氨基端的穿膜肽和羧基端的结合肽组成。
优选的,所述穿膜肽的氨基酸序列如seqidno.2所示;所述结合肽为dip2a氨基酸序列的273-302位,序列如seqidno.3所示。
本发明还提供了上述特异性结合短肽在制备神经保护药物中的应用。
优选的,所述神经保护表现在介导皮层肌动蛋白乙酰化,维持兴奋性突触形态结构与功能稳定。
本发明提供了一种特异性结合短肽,包括41位氨基酸,由穿膜肽和结合肽组成,穿膜肽是广泛使用的穿透细胞膜关键氨基酸,结合肽来自于dip2a氨基酸序列273-302位,是dip2a与皮层肌动蛋白结合的关键结构。所述特异性结合短肽通过与皮层肌动蛋白的特异结合,有效降低皮层肌动蛋白乙酰化,破坏兴奋性突触形态结构与功能的稳定,起效快,作用明显。该结合肽可在体外合成,规模化工业生产。该结合短肽可应用于改善突触形态和功能紊乱药物组合,可用于制备神经保护类药物。
附图说明
图1为结合短肽发现与鉴定;
图2为本发明实施例2中结合短肽关键氨基酸的鉴定;
图3为结合短肽抑制cortactin乙酰化;
图4为结合短肽对神经元突触功能的影响;
图5为cortactin持续乙酰化改善突触兴奋性
具体实施方式
本发明提供了一种特异性结合短肽,氨基酸序列如seqidno.1所示。
本发明所述特异性结合短肽,其氨基酸序列如seqidno.1所示:ygrkkrrqrrrrpkrpplkeffvddfeellevqqpdpnqpk。本发明所述特异性结合短肽优选由氨基端的穿膜肽和羧基端的结合肽组成,所述穿膜肽的氨基酸序列优选如seqidno.2所示:ygrkkrrqrrr。本发明对所述穿膜肽的来源并没有特殊限定,选用本领域广泛使用的穿透细胞膜关键氨基酸即可;穿膜肽的作用是携带多肽、蛋白质及核酸等进入细胞,并且导入后的物质仍保持生物活性。本发明选择的是人类艾滋病病毒hiv-1编码tat蛋白47-57位的氨基酸序列作为穿膜肽,该片段在本领域已被广泛应用。本发明所述结合肽优选为dip2a氨基酸序列的273-302位,序列如seqidno.3所示:rpkrpplkeffvddfeellevqqpdpnqpk。本发明所述结合肽是dip2a与皮层肌动蛋白结合的关键结构。本发明对所述结合肽的来源并没有特殊限定,优选利用人工合成所述序列。
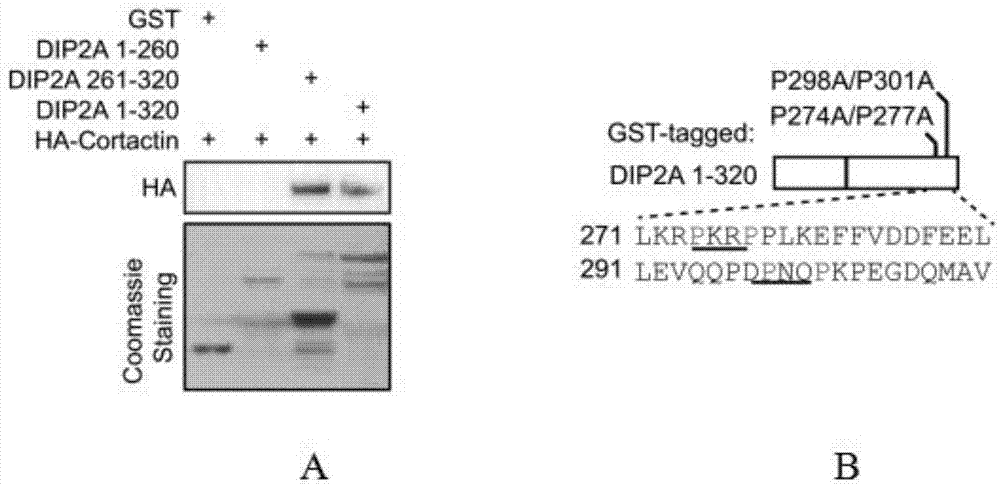
在本发明中,所述dip2a氨基酸序列的结构域如图1-b所示,在dip2a序列的261-320氨基酸部分如图1-b所示存在两个pxxp序列,其中序列中的p代表脯氨酸,而x代表其他的任何氨基酸。这种结合对于保持蛋白质的正确折叠和空间构象具有重要意义,序列中的p是保证结合的重要氨基酸位点,pxxp氨基酸序列中p的单点突变可能对结合的影响不大,但如果同时突变两个p,或插入/删除x而破坏pxxp序列,则会极大地减弱pxxp和sh3结构的结合。
本发明还提供了上述特异性结合短肽在制备神经保护药物中的应用。
在本发明所述应用中,所述神经保护优选表现在介导皮层肌动蛋白乙酰化,维持兴奋性突触形态结构与功能稳定。
下面结合实施例对本发明提供的特异结合短肽及其应用进行详细的说明,但是不能把它们理解为对本发明保护范围的限定。
实施例1
结合短肽发现与鉴定
将构建正确的gst标签融合蛋白dip2a1-260、261-320、1-320质粒转化到大肠杆菌bl21中,均匀涂布于含有amp的lb固体培养基上,37℃培养过夜。挑取单克隆菌落,接种到4ml含有amp的lb液体培养基中,37℃振荡培养8~10h。取菌液接种于10倍体积的新鲜lb液体培养基,37℃振荡培养至菌液od值~0.6。加入iptg(1mm),16℃振荡培养过夜。将菌液4000rpm离心10min,弃上清,以适量菌体裂解液重悬菌体。以300w功率超声破碎菌体至溶液澄清,注意操作过程保持低温。4℃,12000rpm离心30min,取上清即为gst融合蛋白溶液。取50μl谷胱甘肽-琼脂糖凝胶4b珠子原液,以500μlpbs重悬。4℃,500g离心5min,弃上清,以1mlpbs重悬,4℃,垂直混悬仪平衡1h。4℃,500g离心5min,弃上清,以相同珠子体积的pbs重悬。向gst融合蛋白溶液中加入np40(10%),终浓度为0.5%。按照2mg蛋白:50μl珠子的比例,加入平衡后的谷胱甘肽-琼脂糖凝胶4b珠子。4℃,垂直混悬仪结合1h。4℃,500g离心5min,弃上清。分别用含np40(0.5%)和不含np40的细菌裂解液重悬沉淀,达到洗涤的目的。最终获得gst融合蛋白-琼脂糖凝胶4b珠子,以相同珠子体积的pbs重悬。取5μlgst融合蛋白-琼脂糖凝胶4b珠子,通过sds电泳和考马斯亮蓝染色,确定珠子所带gst融合蛋白的浓度。hek293细胞过表达ha-cortactin,48h后用裂解液buffera(20mmtris-hcl[ph8.0],10mmnacl,1mmedta,0.5%np40,蛋白酶抑制剂cocktail)将细胞裂解,提取细胞全蛋白,调整蛋白浓度为1mg/ml,并分装为1mg/tube。取1mg细胞蛋白,加入5μggst融合蛋白-琼脂糖凝胶4b珠子,4℃,垂直混悬仪结合过夜。4℃,2000rpm离心5min,弃上清,buffera裂解液洗涤沉淀3次,10min/次。将获得的免疫沉淀复合物-琼脂糖珠沉淀用2xloadingbuffer重悬,煮沸5min,常规方法westernblot检测目的蛋白,anti-ha抗体免疫结果如图1-a,富含脯氨酸(p)的dip2a261-320能够结合ha-cortactin。在dip2a氨基酸序列271-291位置,共有两处pxxp序列,如图1-b下划线所示。
实施例2
结合短肽关键氨基酸位点的鉴定
在gst-dip2a261-320的基础上,通过核苷酸点突变试剂盒(quickchangelightningsite-directedmutagenesis试剂盒,agilenttechnologies)单点突变p为a,分别或同时破坏两个pxxp结构,得到三个突变质粒gst-p274a/p277a、gst-p298a/p301a、和double。原核表达并纯化上述三种gst标签融合蛋白,以gst标签蛋白作为阴性对照,dip2a261-320作为阳性对照。hek293细胞过表达ha-cortactin并提取蛋白,进行gst-pulldown实验。将融合蛋白沉淀复合物进行westernblot检测,anti-ha抗体免疫印迹结果如图2所示:当有一个pxxp序列被破坏后,dip2a261-320与ha-cortactin的显著减弱;当同时破坏两个pxxp序列后,dip2a261-320几乎不能与ha-cortactin结合。
实施例3
结合短肽对cortactin乙酰化水平的抑制作用
体外合成含有dip2a蛋白序列中含有pxxp的最短多肽ygrkkrrqrrrrpkrpplkeffvddfeellevqqpdpnqpk(下划线所示为tat穿膜序列)。该多肽(dip2a273-302)可与cortactin结合,特异地阻断细胞内源dip2a与cortactin的相互结合。
以不同浓度的多肽孵育hek293细胞,12h后提取细胞总蛋白并进行westernblot检测cortactin、tubulin乙酰化修饰水平。结果如图3-a和图3-b所示:随着多肽浓度的增加,ac-cortactin水平呈现下降的趋势,而ac-tub水平基本不变。
用50μm多肽孵育hek293细胞不同时间,westernblot检测cortactin、tubulin乙酰化修饰水平。结果如图3-c和图3-d所示:随着时间的延长,ac-cortactin水平呈现明显降低,而ac-tub水平基本不变。
实施例4
结合短肽抑制突触兴奋性
培养出生当天小鼠原代神经元:提前用多聚赖氨酸(pll)包被培养皿或载玻片,室温包被过夜或37℃包被2h。回收pll,无菌水漂洗3次。预冷hanks缓冲溶液(hbss),注意保持低温无菌操作。将乳鼠冰晕,75%酒精消毒,迅速取脑。于hbss中剥离海马/皮层组织,轻轻转移到装有新鲜hbss的ep管中。剪成小块,37℃胰酶(0.125%)消化30min,期间每隔10min将组织块轻柔重悬。用含有10%fbs的f12培养基终止消化,800rpm离心1min,去上清。加入f12+10%fbs培养基重悬,37℃静置5min。用移液器轻柔吹打细胞团块,静置2min,上清即为神经元混悬液,铺板。
以相同的多肽(1μm)孵育体外培养的神经元(div15),12h后电生理技术检测神经元微小兴奋性突触后电流(miniatureexcitatorypostsynapticcurrents,mepsc)。采用epc-10/2放大器(heka,德国)以电压钳模式进行全细胞膜片钳记录。记录电极由硼硅酸盐玻璃毛细管(warnerinstruments)自制,电阻为3~5mω。只有串联电阻<15mω的全细胞贴片用于记录。膜电位保持在-70mv,电极内溶液组成:130mm葡萄糖酸钾,1mmegta,5mmna2atp,2mmmgatp,0.3mmna2gtp,5mmqx-314和10mmhepes(ph7.3)。浴液组成:25mmhepes(ph7.3),128mmnacl,30mm葡萄糖,5mmkcl,5mmcacl2,1mmmgcl2,50μmd-ap5,20μm荷包牡丹碱。d-ap5、荷包牡丹碱和qx-314来自tocrisbioscience,其他化学品来自sigma-aldrich。当记录mepsc时,将1μm河豚毒素(tocrisbioscience)加入浴液中。使用patchmaster软件(heka,germany)获取数据,并使用minianalysis软件(synaptosoft)、clampfit(moleculardevices)和igor(wavemetrics)进行分析。
结果如图4所示,其中图4-a表示体外培养神经元mepsc代表电流,多肽浓度1μm,孵育时间12h,mepsc在div15时的记录;图4-b表示两组神经元mepsc振幅,图4-c表示两组神经元mepsc频率。与对照组相比,特异阻断dip2a-cortactin相互作用,显著降低了神经元mepsc的振幅(amplitude)和频率(frequency)。
实施例5
cortactin持续乙酰化改善突触兴奋性
含有该特异结合序列结合序列的cortactin,持续乙酰化形式有效改善突触兴奋性。利用改变电荷的策略,我们设计构建了模拟ac-cortactin形式的cortactin-8kq,即把8个乙酰化残基赖氨酸(k,pi=9.74)突变为带负电荷的谷氨酰胺(q,pi=5.65),模拟乙酰化状态下的电荷中和;同理,将8个乙酰化残基k突变为等电点相近的精氨酸(r,pi=10.76),构建得到模拟cortactin非乙酰化形式cortactin-8kr。
按照前述方法体外培养出生当天小鼠原代神经元,至第21天(div21)进行免疫荧光染色,anti-map2标记神经元树突,psd95为突触后标记物。如图5-a和b,缺失dip2a时,即cortactin乙酰化下降时,psd95定位异常,更靠近树突;在缺失dip2a-cortactin结合状态下,神经元div15时慢病毒(lentivirus,lv)侵染介导持续cortactin乙酰化形式-8kq过表达。与对照组相比,过表达cortactin-8kq能够促使神经元psd95向远离树突位置伸展,说明模拟乙酰化形式的cortactin能够补救dip2a-cortactin结合缺失造成的psd95定位异常。同样地,这种结合缺失造成的神经元电生理活性异常,也能被持续乙酰化形式cortactin(8kq)补救,如图5-c和d。
综上所述,本发明提供了一种特异性结合短肽及其应用,dip2a-cortactin相互作用是dip2a调节cortactin乙酰化的前提,进而影响神经元突触功能;阻断这一相互作用即cortactin乙酰化下降,神经元突触功能受损。恢复cortactin与dip2a结合,有效改善cortactin乙酰化和突触的结构域功能。本发明通过特异结合,有效介导皮层肌动蛋白乙酰化,维持兴奋性突触形态结构与功能的稳定。该结合肽可在体外合成,规模化工业生产,应用于改善突触形态和功能紊乱,起神经保护作用。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
序列表
<110>东北师范大学
<120>一种特异性结合短肽及其应用
<160>3
<170>siposequencelisting1.0
<210>1
<211>41
<212>prt
<213>人工序列(artificialsequence)
<400>1
tyrglyarglyslysargargglnargargargargprolysargpro
151015
proleulysgluphephevalaspasppheglugluleuleugluval
202530
glnglnproaspproasnglnprolys
3540
<210>2
<211>11
<212>prt
<213>人工序列(artificialsequence)
<400>2
tyrglyarglyslysargargglnargargarg
1510
<210>3
<211>30
<212>prt
<213>人工序列(artificialsequence)
<400>3
argprolysargproproleulysgluphephevalaspasppheglu
151015
gluleuleugluvalglnglnproaspproasnglnprolys
202530
- 还没有人留言评论。精彩留言会获得点赞!