一种用于制备L-2-氨基丁酸的重组共表达体系及其应用的制作方法
本发明涉及基因工程技术领域,具体涉及一种用于制备l-2-氨基丁酸的重组共表达体系及其应用。
背景技术:
l-2-氨基丁酸是一种非天然氨基酸,可以提高葡萄糖磷酸酯酶活性,促进脑细胞新陈代谢,医学临床用于无痛去龋和脑血管病后遗症,并且具有降压功效。此外,l-2-氨基丁酸还是一种重要的化工原料和医药中间体,可用于生产抑菌抗结核药乙胺丁醇和抗癫痫药左乙拉西坦,l-2-氨基丁酸的工业化生产技术已成为制药工程研究的热点。
目前,l-2-氨基丁酸的生产方法主要有化学法和生物法。化学法主要有化学拆分法和化学合成法。化学拆分法如用手性拆分试剂与消旋的2-氨基丁酸形成非对应异构体进行拆分得到l-2-氨基丁酸。化学合成法如以正丁酸为起始原料,经多聚磷酸路线或三溴化硼路线合成2-溴丁酸,然后在氨水中合成2-氨基丁酸。这些方法所需试剂价格昂贵,成本较高,反应条件苛刻,易生成副产物,不宜应用于工业化大规模生产,同时大量有机溶剂的使用还会污染环境。
生物法又包括微生物发酵法和酶转化法。微生物发酵法指利用大肠杆菌等微生物直接发酵生产l-2-氨基丁酸,专属性较强,条件温和,环境污染少,但存在反应产物成分复杂、分离繁琐的缺点。酶催化制备方法专一性强,效率高,反应条件温和,对环境友好,常温常压反应,不需要耐高温高压的设备,是一种较为理想的药物中间体制备方法。目前,生物酶法催化制备l-2-氨基丁酸应用比较广泛的是用苏氨酸脱氨酶、亮氨酸脱氢酶和甲酸脱氢酶三酶体系催化生产。以l-苏氨酸为起始原料,添加氧化型辅酶nad+,并以甲酸铵作为辅酶循环辅料。反应过程中,苏氨酸脱氨酶催化l-苏氨酸转化为中间产物2-丁酮酸,然后亮氨酸脱氢酶将其转化为最终产物l-2-氨基丁酸,同时,以甲酸脱氢酶催化甲酸铵来实现辅酶nadh的再生,使反应循环进行。
烟酰胺腺嘌呤二核苷酸(即还原型辅酶ⅰ,nadh)是氧化还原反应中重要的辅酶。作为电子供体,1分子nadh可以提供2分子电子用于羰基等基团的还原。氧化还原酶在制药、食品、精细化工、农药等领域具有重要的用途,主要是用来制备手性化合物。氧化还原酶在进行生物催化时,合成产物同时要消耗一定量的辅酶,大约80%的氧化还原酶需要nad+/nadh作为辅酶,所以辅酶的高效供应是开发氧化还原酶催化反应的关键技术之一。但是辅酶一般稳定性低,价格昂贵,从工业应用的经济性角度来看,在生产过程中大量投加辅酶显然是不可行的。因此,氧化还原酶在应用中除了必须有合适的酶和反应工程技术外,还必须提供高效、低成本的辅酶再生系统。所谓辅酶再生就是把辅酶从氧化态再生为还原态,或者反之,从而使辅酶保持在一定的催化剂量水平。
在上述三酶体系催化生产l-2-氨基丁酸的反应中需要投加辅酶,导致生产成本过高,限制了其工业化应用。因此,建立高效的细胞自身代谢辅酶循环再生系统,以实现反应过程中辅酶的零添加,并且辅酶的氧化和还原在同一个宿主细胞中进行,提高了反应效率,这在l-2-氨基丁酸的工业化生产中具有重要的意义。
技术实现要素:
本发明的目的在于提供一种活性高、辅酶循环再生能力强的亮氨酸脱氢酶和甲酸脱氢酶共表达重组菌,将其应用于l-2-氨基丁酸的工业化生产中,利用细胞自身代谢辅酶循环再生系统,以实现反应过程中辅酶的零添加。
为实现上述目的,本发明采用如下技术方案:
一种用于制备l-2-氨基丁酸的重组共表达体系,所述重组共表达体系为单质粒双基因表达载体或双质粒共表达体系,所述单质粒双基因表达载体为以pcdfduet-1或petduet-1为基础,其多克隆位点中插入甲酸脱氢酶基因和亮氨酸脱氢酶基因;所述双质粒共表达体系为以pcdfduet-1、petduet-1和pet-28b的任意两种为基础,包含亮氨酸脱氢酶基因和甲酸脱氢酶基因;
所述亮氨酸脱氢酶基因的核苷酸序列如seqidno.1所示,所述甲酸脱氢酶基因的核苷酸序列如seqidno.3所示。
本发明通过构建亮氨酸脱氢酶基因和甲酸脱氢酶基因的双基因共表达载体,同时提供生物酶法制备l-2-氨基丁酸中涉及的亮氨酸脱氢酶和甲酸脱氢酶。制备l-2-氨基丁酸的过程中使用含有上述双基因共表达载体的全细胞作为催化剂,可省略超声波破碎细胞的步骤,同时,在同一细胞内共表达亮氨酸脱氢酶和甲酸脱氢酶可减少两个酶的空间位阻,有助于加快反应进程,还可以利用宿主细胞自身代谢产生的nad+/nadh,实现辅酶的循环再生,不需要额外添加辅酶。
本发明所述的重组共表达体系可以通过本领域常规方法将亮氨酸脱氢酶基因和甲酸脱氢酶基因片段连接到各种表达载体(如petduet-1、pcdfduet-1和pet-28b)上构建而成。
作为优选,所述单质粒双基因表达载体包括原始载体pcdfduet-1以及分别在其mcs1和mcs2中插入的甲酸脱氢酶基因和亮氨酸脱氢酶基因;其中mcs1是指质粒的第一个多克隆位点,mcs2是指质粒的第二个多克隆位点。
所述双质粒共表达体系为分别在petduet-1的mcs1和mcs2中插入甲酸脱氢酶基因和亮氨酸脱氢酶基因的重组质粒和分别在pcdfduet1的mcs1和mcs2中插入甲酸脱氢酶基因和亮氨酸脱氢酶基因的重组质粒组成的体系1;或分别在pcdfduet1的mcs1和mcs2中插入甲酸脱氢酶基因和亮氨酸脱氢酶基因的重组质粒和在pet-28b中插入亮氨酸脱氢酶基因的重组质粒组成的体系2;或分别在petduet-1的mcs1和mcs2中插入亮氨酸脱氢酶基因和甲酸脱氢酶基因的重组质粒和在pcdfduet1的mcs1插入甲酸脱氢酶基因的重组质粒组成的体系3;或分别在pcdfduet1的mcs1和mcs2中插入甲酸脱氢酶基因和亮氨酸脱氢酶基因的重组质粒和在petduet-1的mcs1中插入甲酸脱氢酶基因的重组质粒组成的体系4;或分别在pcdfduet1的mcs1和mcs2中插入甲酸脱氢酶基因和亮氨酸脱氢酶基因的重组质粒和在pet-28b中插入甲酸脱氢酶基因的重组质粒组成的体系5;或在pcdfduet1的mcs1插入亮氨酸脱氢酶基因的重组质粒和petduet-1的mcs1中插入甲酸脱氢酶基因的重组质粒的体系6;或在pcdfduet1的mcs1插入亮氨酸脱氢酶基因的重组质粒和在pet-28b中插入甲酸脱氢酶基因的重组质粒组成的体系7。
本发明还提供了一种包含所述的重组共表达体系的重组基因工程菌。
作为优选,所述重组基因工程菌的宿主细胞为大肠杆菌e.colibl21(de3)。
本发明还提供了所述的重组基因工程菌在催化生产l-2-氨基丁酸中的应用。
所述的应用包括:以所述的重组基因工程菌经发酵培养获得的活细胞为催化剂,以l-苏氨酸为起始原料,甲酸铵作为辅酶循环辅料,并添加苏氨酸脱氢酶粗酶液,去离子水为反应介质构成反应体系,进行酶催化反应,反应结束后,得到含l-2-氨基丁酸的反应液,反应液经过分离纯化,得到l-2-氨基丁酸。
作为优选,重组基因工程菌的发酵培养方法为:将重组基因工程菌接种于含有抗生素的培养基中培养,获得种子液,经扩大培养获得od600达到0.6~0.8的菌液,加入诱导剂,18-28℃诱导8-20h,发酵培养结束后,收集菌体。
培养基可为本领域任何可使菌体生长并产生本发明的培养基,优选为lb培养基,lb培养基:蛋白胨10g/l,酵母粉5g/l,nacl10g/l,蒸馏水溶解,ph7.0。
种子液培养的条件为37℃,150r/min条件下培养12h。扩大培养的条件为37℃,180r/min。
摇瓶培养重组基因工程菌时,采用iptg作为诱导剂,终浓度0.1mmol/l,诱导温度22℃,诱导时间14h。
发酵罐培养基因工程菌时,采用乳糖作为诱导剂,往菌液中加到质量终浓度≥8g/l的乳糖,22℃诱导培养15-19h。作为优选,诱导时间为17h。
作为优选,反应体系中,l-苏氨酸质量浓度为100~200g/l,甲酸铵质量浓度为50~150g/l,苏氨酸脱氨酶浓度为10000~20000u/l,重组基因工程菌活细胞以菌体湿重计为20~80g/l;酶催化温度为30~40℃,时间为2-8h。
更为优选,反应体系中,l-苏氨酸质量浓度为180g/l,甲酸铵质量浓度为95.3g/l,苏氨酸脱氨酶浓度为13000u/l,重组基因工程菌活细胞以菌体湿重计为40g/l。酶催化的温度为35℃,时间为8h。
与现有技术相比,本发明有益效果主要体现在:
(1)本发明通过构建亮氨酸脱氢酶基因和甲酸脱氢酶基因共表达体系,转化至表达菌中得到重组基因工程菌,以经发酵培养的重组基因工程菌活细胞作为催化剂应用于制备l-2-氨基丁酸的催化反应体系中,生产效率高,当l-苏氨酸投料量为180g/l时,反应8h,转化率达99%以上,时空产率19.3g/(l·h)。
(2)本发明使用全细胞作为催化剂,避免了酶的纯化,操作简便;两个酶在一个细胞中,可减少空间位阻,加快反应速度;细胞耐受性强,可适应较大范围反应温度、ph等因素;以去离子水为反应介质构成反应体系,反应产物提纯方便;反应过程中无需添加辅酶,节约成本。
附图说明
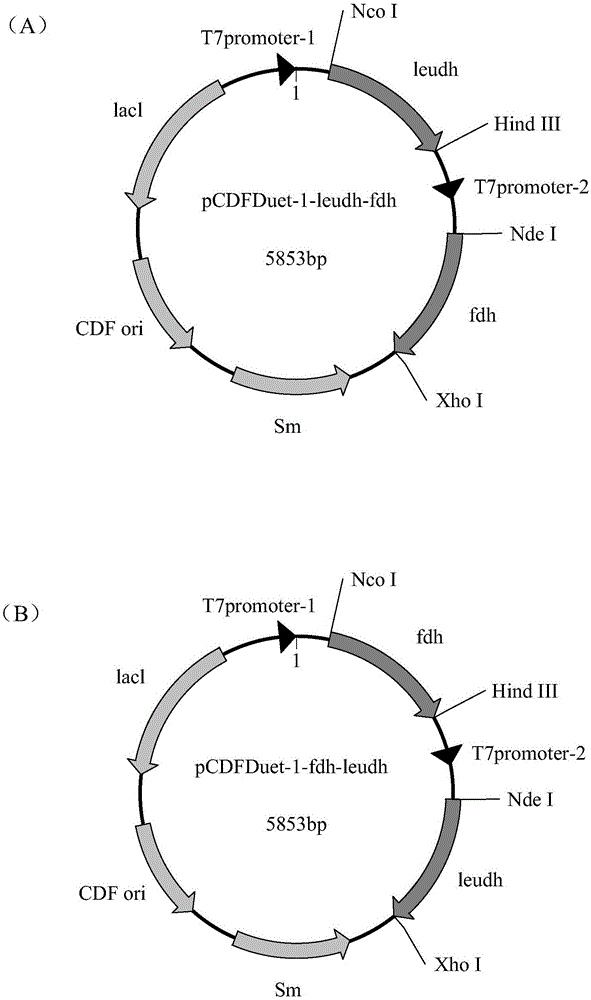
图1为共表达质粒构建示意图,其中(a)为pcdfduet-1-leudh-fdh,(b)为pcdfduet-1-fdh-leudh。
图2为l-2-氨基丁酸的hplc分析图,(a)是l-2-氨基丁酸的标准品溶液(1.20mol/l)的hplc分析图,(b)是共表达菌株催化体系反应液的hplc分析图。
图3为pcdfduet-1-leudh(mcs1)/pet-28b-fdh构建示意图。
图4为诱导温度对pcdfduet-1-leudh(mcs1)/pet-28b-fdh的sds-page(a)和底物转化率(b)的影响,(a)中a-f分别是在诱导温度18、20、22、24、26和28℃下诱导培养的菌体破碎上清物,a-f相对应的沉淀物。
图5为诱导剂浓度对pcdfduet-1-leudh(mcs1)/pet-28b-fdh的sds-page(a)和底物转化率(b)的影响,(a)中a-f分别是在诱导剂浓度0.06、0.08、0.1、0.12、0.14和0.16mmol/l下诱导培养的菌体破碎上清物,a-f相对应的沉淀物。
图6为诱导时间对pcdfduet-1-leudh(mcs1)/pet-28b-fdh的sds-page(a)和底物转化率(b)的影响,(a)中a-g分别是在诱导时间8、10、12、14、16、18和20h下诱导培养的菌体破碎上清物,a-g相对应的沉淀物。
图7为反应温度对制备l-2-氨基丁酸的影响。
图8为ph变化对制备l-2-氨基丁酸的影响,(a)是在去离子水中反应体系的ph变化曲线,(b)是反应体系ph对制备l-2-氨基丁酸的影响。
图9为辅底物甲酸铵浓度对制备l-2-氨基丁酸的影响。
图10为发酵罐培养乳糖浓度对催化反应进程的影响。
图11为发酵罐培养诱导时间对催化反应进程的影响。
图12利用e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh菌株制备l-2-氨基丁酸的反应进程。
具体实施方式
为了使本发明的目的、技术方案及优点更加清楚明白,下面结合具体实施例对本发明作进一步描述,所描述的具体实施例仅用以解释本发明,并不用于限定本发明。
实施例1:亮氨酸脱氢酶基因(leudh)和甲酸脱氢酶基因(fdh)克隆及共表达重组体系构建
亮氨酸脱氢酶的基因来源于中间型嗜热放线菌(thermoactinomycesintermedius)的全基因组序列。为了使该基因连接到载体pet-28b后可以表达带有his-tag的蛋白,切除其终止密码子,并以b.subtilis168的密码子偏好性为参照对其序列和常用限制性内切酶识别位点bamhi、xhoi、psti、hindiii和ncoi进行序列优化,获得新的亮氨酸脱氢酶基因(核苷酸序列为seqidno.1,氨基酸序列为seqidno.2),已在专利申请(201610867380.8)中公开,并已将新的亮氨酸脱氢酶基因连接于表达载体pet-28b上,即为pet-28b-leudh。
甲酸脱氢酶基因(核苷酸序列为seqidno.3,氨基酸序列为seqidno.4)来源于禾谷镰刀菌(fusariumgraminearum),已连接于表达载体pet-28b上,即为pet-28b-fdh。
以pet-28b-leudh为模板,pcr扩增得到亮氨酸脱氢酶基因片段。
上游引物(引物1):5’-ccatgggtaaaatcttcgactacatg-3’(seqidno.5),
下游引物(引物2):5’-aagcttttatttgttgttgaagttga-3’(seqidno.6);
以pet-28b-fdh为模板,pcr扩增得到甲酸脱氢酶基因片段。
上游引物(引物3):5’-catatgggtaaaattgtactggttctgtatgac-3’(seqidno.7),
下游引物(引物4):5’-ctcgagttatttcttgtcgtgtttaccgtac-3’(seqidno.8)。
pcr反应体系(50μl)为:10×taqpolymerasebuffer5μl,dntpmixture4μl;模板质粒1μl;上下游引物各0.5μl;taqpolymerasedna聚合酶0.5μl;无菌水38.5μl。
采用bio-radpcr仪,反应的程序为94℃预变性5min;94℃变性45s,50℃退火45s,72℃延伸1min(30个循环);72℃延伸10min。
获得pcr扩增产物采用回收试剂盒(axygen,美国爱思进)回收,纯化后的片段分别标记为片段leudh,片段fdh,分别与pgem-teasy载体连接,获得重组载体pgem-teasy-leudh、pgem-teasy-fdh。利用限制性核酸内切酶ncoi和hindiii双酶切重组载体pgem-teasy-leudh和质粒pcdfduet-1,因此形成的亮氨酸脱氢酶基因片段与质粒pcdfduet-1的第一个多克隆位点具有相同的粘性末端,用t4连接酶连接,获得质粒pcdfduet-1-leudh(mcs1)。然后再利用限制性核酸内切酶ndei和xhoi双酶切重组载体pgem-teasy-fdh和质粒pcdfduet-1-leudh(mcs1),因此形成的甲酸脱氢酶基因片段与质粒pcdfduet-1-leudh(mcs1)的第二个多克隆位点具有相同的粘性末端,用t4连接酶连接,即构建得到含有本发明的亮氨酸脱氢酶基因片段与甲酸脱氢酶基因片段的单质粒重组共表达载体pcdfduet-1-leudh-fdh,将leudh和fdh交换多克隆位点,得到质粒pcdfduet-1-fdh-leudh,如图1所示。
采用相同策略构建得到的单质粒共表达体系为:pcdfduet-1-leudh-fdh、pcdfduet-1-fdh-leudh、petduet-1-leudh-fdh、petduet-1-fdh-leudh;双质粒共表达体系为:petduet-1-leudh-fdh/pcdfduet-1-leudh-fdh、petduet-1-leudh-fdh/pcdfduet1-fdh-leudh、petduet-1-fdh-leudh/pcdfduet-1-leudh-fdh、petduet-1-fdh-leudh/pcdfduet1-fdh-leudh、petduet-1-leudh-fdh/pcdfduet-1-leudh(mcs1)、petduet-1-leudh-fdh/pet-28b-leudh、petduet-1-fdh-leudh/pcdfduet-1-leudh(mcs1)、petduet-1-fdh-leudh/pet-28b-leudh、pcdfduet-1-leudh-fdh/petduet-1-leudh(mcs1)、pcdfduet-1-leudh-fdh/pet-28b-leudh、pcdfduet1-fdh-leudh/petduet-1-leudh(mcs1)、pcdfduet1-fdh-leudh/pet-28b-leudh、petduet-1-leudh-fdh/pcdfduet-1-fdh(mcs1)、petduet-1-leudh-fdh/pet-28b-fdh、petduet-1-fdh-leudh/pcdfduet-1-fdh(mcs1)、petduet-1-fdh-leudh/pet-28b-fdh、pcdfduet-1-leudh-fdh/petduet-1-fdh(mcs1)、pcdfduet-1-leudh-fdh/pet-28b-fdh、pcdfduet1-fdh-leudh/petduet-1-fdh(mcs1)、pcdfduet1-fdh-leudh/pet-28b-fdh、petduet-1-leudh(mcs1)/pcdfduet-1-fdh(mcs1)、petduet-1-leudh(mcs1)/pet-28b-fdh、pcdfduet-1-leudh(mcs1)/petduet-1-fdh(mcs1)、pcdfduet-1-leudh(mcs1)/pet-28b-fdh、pet-28b-leudh/petduet-1-fdh(mcs1)、pet-28b-leudh/pcdfduet-1-fdh(mcs1)。
将共表达体系共转入到e.colibl21(de3)感受态细胞中,转化方法为:质粒1~2μl,加入到100μle.colibl21(de3)感受态细胞中,充分混匀后,冰浴30min;将装有混合物的eppendorf管置于42℃水浴热击90s,然后立即转移到冰上冷却2min;向管中加入600μllb液体培养基,置于37℃、200r/min恒温摇床上培养1h,然后涂布于含有相应抗性的lb固体平板上,37℃培养12~18h。
实施例2:不同共表达体系亮氨酸脱氢酶和甲酸脱氢酶的表达
将上述实施例1构建的不同共表达菌株,接种至装有50mllb液体培养基的摇瓶,并加入相应的抗性,在37℃,150r/min条件下摇床培养12h,获得种子液,种子液以体积分数1%(v/v)的接种量转接至100mllb液体培养基中,并添加抗性,37℃,180r/min培养至od600达到0.6~0.8,加入诱导剂iptg至终浓度0.1mmol/l,诱导温度22℃,诱导时间14h。发酵结束后4℃,8000r/min,10min离心,弃上清,生理盐水洗涤两次,收集菌体,4℃保存备用。
实施例3:不同共表达菌株生产l-2-氨基丁酸的效率及比酶活对比
酶活定义:在标准条件下,每分钟生成1μmoll-2-氨基丁酸所需的酶量为1单位酶活,表观比酶活即为每克湿菌体所具有的酶活单位数。
样品检测方法:美国waters高效液相色谱仪,色谱柱eclipsexd8-c18(5μm4.6mm×250mm),流动相:0.02mol/l的磷酸氢二钠(ph7.2):乙腈=70:30,流速:1.0ml/min,柱温:30℃,紫外检测波长:360nm。
样品衍生化条件:取100μl待测样品,与100μl0.5mol/l的nahco3溶液和50μl1%(v/v)2,4-二硝基氟苯乙腈溶液混合,60℃避光保温1h,反应结束后冷却至室温,再加入750μlnah2po4/na2hpo4缓冲液(0.2mol/l,ph7.0)。原理:氨基酸的游离末端nh2在碱性环境中可与2,4-二硝基氟苯发生亲核芳环取代反应,可用hplc定量检测生成的二硝基苯氨基酸衍生物。在上述条件下,2,4-二硝基氟苯和l-2-氨基丁酸的保留时间分别为4.1分钟和5.8分钟,如图2所示。
优势菌株的筛选:反应体系为180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l菌体(实施例2获得的各个菌体),于去离子水中构成30ml反应体系,在35℃、600r/min条件下进行酶催化反应8h。反应过程中定时取样,用浓盐酸终止反应,样品用于液相色谱分析。具体测定结果示于表1。
通过表观比酶活及生产l-2-氨基丁酸的效率来衡量共表达菌株的优劣,催化效率较高的共表达菌株如下:pcdfduet-1-fdh-leudh、petduet-1-fdh-leudh/pcdfduet1-fdh-leudh、pcdfduet1-fdh-leudh/pet-28b-leudh、petduet-1-leudh-fdh/pcdfduet-1-fdh(mcs1)、pcdfduet1-fdh-leudh/petduet-1-fdh(mcs1)、pcdfduet1-fdh-leudh/pet-28b-fdh、pcdfduet-1-leudh(mcs1)/petduet-1-fdh(mcs1)、pcdfduet-1-leudh(mcs1)/pet-28b-fdh,其中pcdfduet-1-leudh(mcs1)/pet-28b-fdh的催化效率最高,表观比酶活达到78.25u/g,底物转化率达99.5%。故选择e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh为优选工程菌用于后续研究,双质粒菌株pcdfduet-1-leudh(mcs1)/pet-28b-fdh构建方法如图3所示。
表1不同共表达体系构建菌株的催化性能
注:a表示在反应体系为180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l菌体(实施例2获得的各个菌体),于去离子水中构成30ml反应体系,在35℃、600r/min条件下进行酶催化反应8h;b表示在标准条件下测得的表观比酶活。
实施例4:e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh菌株摇瓶培养发酵条件优化
反应体系同上述优势菌株的筛选条件,优化e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh摇瓶培养的发酵条件,选择sds-page和底物转化率作为参考指标。首先优化了诱导温度,诱导时加入0.1mmol/l的iptg,分别在18、20、22、24、26、28℃条件下诱导16h,结果见图4,由图可知,22℃为最佳的诱导温度。
进一步考察iptg浓度对产酶的影响,诱导时分别添加0.06、0.08、0.1、0.12、0.14、0.16mmol/l诱导剂iptg,22℃条件下诱导16h,结果见图5,最终确定最佳iptg浓度为0.1mmol/l。
进一步考察了诱导时间对产酶的影响,诱导时加入0.1mmol/l的iptg,22℃条件下分别诱导8、10、12、14、16、18、20h,由图6可知,最佳诱导时间为14h。
实施例5:反应温度和反应体系ph对制备l-2-氨基丁酸的影响
首先考察反应温度对制备l-2-氨基丁酸的影响,反应体系为180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l摇瓶培养的菌体,于去离子水中构成30ml反应体系,分别在25、30、35、40、45、50℃,600r/min条件下进行酶催化反应8h,结果见图7。在35℃之前,随着反应温度的提高,转化率也提高,当反应温度为35℃时,转化率达到最高,继续升高反应温度,转化率急剧下降,可能是因为过高的反应温度使酶失活,综合实验结果,确定35℃为最佳反应温度。
进一步考察反应体系ph对制备l-2-氨基丁酸的影响,反应体系为180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l摇瓶培养的菌体,于去离子水中构成30ml反应体系,在35℃,600r/min条件下进行酶催化反应8h,ph变化曲线见图8a。初始反应体系ph为6.5,反应8h后ph为8.5,转化率达99%以上。故实验组于0.2mmol/l磷酸钠缓冲液构成30ml反应体系,使用ph-stat(梅特勒,瑞士)通过添加体积分数为50%的磷酸水溶液和氨水水溶液分别控制反应液的ph为6.0、6.5、7.0、7.5、8.0、8.5、9.0,在35℃,600r/min条件下进行酶催化反应8h,结果见图8b。过高或过低的ph都会影响酶活,当ph为6.5~8.0时,转化率达99%以上,当ph为8.5时,转化率达98%,且控制反应体系ph时,反应进程并没有明显提高,故为了节约成本和方便提纯,以去离子水为反应介质构成反应体系,不控制反应液的ph。
实施例6:底物浓度和辅底物浓度对制备l-2-氨基丁酸的影响
首先考察辅底物甲酸铵浓度对制备l-2-氨基丁酸的影响,固定底物l-苏氨酸质量浓度为180g/l,分别添加与l-苏氨酸的摩尔比1:1、1.05:1、1.1:1、1.15:1、1.2:1、1.25:1的甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l摇瓶培养的菌体,于去离子水中构成30ml反应体系,在35℃、600r/min条件下进行酶催化反应8h,结果见图9,因反应体系中甲酸铵的添加量最少应与l-苏氨酸的摩尔量相同,且当甲酸铵:l-苏氨酸=1:1时,底物转化率可以达99%以上,故为了节约成本,选择添加与l-苏氨酸的摩尔量相同的甲酸铵,即为95.3g甲酸铵。
底物l-苏氨酸的初始质量浓度分别为120g/l、150、180、210g/l,甲酸铵的摩尔量与l-苏氨酸的摩尔量相同,13000u/l苏氨酸脱氨酶粗酶液,40g/l摇瓶培养的菌体,于去离子水中构成30ml反应体系,在35℃、600r/min条件下进行酶催化反应8h,结果见表2。如表2所示,在最优条件下,最高底物投料量为180g/l,8h左右反应结束,底物转化率99%以上,产物产量达1.51mol/l,时空产率19.3g/(l·h)。
表2l-苏氨酸浓度对制备l-2-氨基丁酸的影响
实施例7:e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh菌株发酵罐培养发酵条件优化
在摇瓶培养发酵的基础上,进行大规模发酵罐培养发酵,以验证e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh菌株工业化应用能力。首先考察乳糖浓度对催化反应进程的影响,将共表达亮氨酸脱氢酶和甲酸脱氢酶的重组基因工程菌接种至装有100mllb液体培养基的摇瓶,并分别添加终浓度为30mg/l的链霉素和卡那霉素,在37℃,150r/min条件下摇床培养12h,作为一级种子液,一级种子液以体积分数10%(v/v)的接种量转接至1llb液体培养基中,并分别添加终浓度为30mg/l的链霉素和卡那霉素,于37℃,150r/min条件下摇床培养6h,作为二级种子液,二级种子液以体积分数5%(v/v)的接种量转接至30l发酵培养基中,并分别添加终浓度为30mg/l的链霉素和卡那霉素,通气量1.1vvm,罐压0.05mpa,ph7.0(用体积分数50%的磷酸水溶液和氨水水溶液调节),37℃,350r/min培养至od600达到8~10,分别添加质量浓度为4、6、8、10、12.5g/l的乳糖,诱导温度22℃,诱导时间17h。发酵结束后4℃,8000r/min,10min离心,弃上清,生理盐水洗涤两次,收集菌体,4℃保存备用。
反应体系为180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l发酵罐培养的菌体,于去离子水中构成100ml反应体系,在35℃、600r/min条件下进行酶催化反应8h,结果见图10,由图10可知,过低的乳糖添加量不利于反应的进行,使底物完全转化的最低乳糖浓度为8g/l,继续提高乳糖浓度对反应的进程没有明显影响,确定最适的乳糖质量浓度为8g/l。
进一步考察诱导时间对催化反应进程的影响,随着诱导时间的延长,酶蛋白的表达量会有一定程度的增加,为了确定最适诱导时间,诱导时添加质量浓度为8g/l的乳糖,诱导温度22℃,分别在降温诱导11、13、15、17、19h后取样检测,其余培养条件和反应条件同上,结果见图11,由图11可知,随着诱导时间的延长,目的蛋白酶的催化能力逐渐升高,诱导17h后,催化能力基本达到稳定,其后增长不明显。最终确定诱导时间为17h。
实施例8:分批补料方式对制备l-2-氨基丁酸的影响
反应体系为180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氨酶粗酶液,40g/l发酵罐培养的菌体,于去离子水中构成100ml反应体系,在35℃、600r/min条件下进行酶催化反应8h,l-苏氨酸和甲酸铵的添加方式见表3。由表3可知甲酸铵分批添加不利于催化反应的进行,初始l-苏氨酸添加量越大,最终转化率越高,且一次性投料也可以达到完全转化,综合实验结果,选择一次性投料,即初始l-苏氨酸质量浓度为180g/l,甲酸铵质量浓度为95.3g/l。
表3l-苏氨酸和甲酸铵的添加方式对制备l-2-氨基丁酸的影响
注:a表示在0h投入18gl-苏氨酸和9.53g甲酸铵;b表示分别在0h、0.5h、1h投入6gl-苏氨酸和3.17g甲酸铵;c表示分别在0h、0.5h、1h投入6gl-苏氨酸,在0h投入9.53g甲酸铵;d表示分别在0h、0.5h、1h投入8、5、5gl-苏氨酸,在0h投入9.53g甲酸铵;e表示分别在0h、0.5h、1h投入10、4、4gl-苏氨酸,在0h投入9.53g甲酸铵;f表示分别在0h、0.5h、1h投入12、3、3gl-苏氨酸,在0h投入9.53g甲酸铵。
实施例9:利用e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh菌株制备l-2-氨基丁酸的反应进程
将e.colibl21(de3)pcdfduet-1-leudh(mcs1)/pet-28b-fdh接种至装有100mllb液体培养基的摇瓶,并分别添加终浓度为30mg/l的链霉素和卡那霉素,在37℃,150r/min条件下摇床培养12h,作为一级种子液,一级种子液以体积分数10%(v/v)的接种量转接至1llb液体培养基中,并分别添加终浓度为30mg/l的链霉素和卡那霉素,于37℃,150r/min条件下摇床培养6h,作为二级种子液,二级种子液以体积分数5%(v/v)的接种量转接至30l发酵培养基中,并分别添加终浓度为30mg/l的链霉素和卡那霉素,通气量1.1vvm,罐压0.05mpa,ph7.0(用体积分数50%的磷酸水溶液和氨水水溶液调节),37℃,350r/min培养至od600达到8~10,加入质量浓度为8g/l的诱导剂乳糖,诱导温度22℃,诱导时间17h。发酵结束后4℃,8000r/min,10min离心,弃上清,生理盐水洗涤两次,收集菌体,4℃保存备用。
反应体系1l:180g/l底物l-苏氨酸,95.3g/l辅底物甲酸铵,13000u/l苏氨酸脱氢酶粗酶液,40g/l上述菌体,以去离子水为反应介质构成反应体系,在35℃、600r/min条件下进行酶催化反应8h。反应过程中定时取样,用浓盐酸终止反应,样品用于液相色谱分析。反应进程见图12,l-2-氨基丁酸在反应2h内生成的速度较快,之后反应速度放缓,可能是因为随着反应的进行酶活性不断降低,并且产物的积累对酶活性也有一定的抑制作用。反应至8h基本结束,此时底物转化率99%以上,e.e.值99.5%以上,时空产率19.3g/(l·h)。
序列表
<110>浙江工业大学
<120>一种用于制备l-2-氨基丁酸的重组共表达体系及其应用
<160>8
<170>siposequencelisting1.0
<210>1
<211>1104
<212>dna
<213>人工序列(artificialsequence)
<400>1
atgggtaaaatcttcgactacatggaaaaatacgactacgaacaactggtgatgtgccag60
gacaaagaatccggtctgaaggctatcatctgcatccacgtgactaccctgggtccggct120
ctgggtggtatgcgtatgtggacgtacgcttccgaagaagaggcgattgaagatgctctg180
cgcctgggtcgtggcatgacctataaaaacgcggctgcaggtctgaacctgggtggcggc240
aaaaccgtaattattggtgacccgcgtaaagacaaaaacgaggcgatgtttcgtgctctg300
ggtcgcttcatccagggtctgaacggtcgttacattaccgccgaggatgttggtaccact360
gtcgaagacatggacatcatccacgaagaaacccgttacgtaaccggcgtttccccagcc420
tttggctcttctggcaacccgagcccggtgactgcctacggcgtttatcgtggtatgaaa480
gcagcagcgaaagaagcattcggcgatgactctctggaaggtaaagtcgttgctgttcaa540
ggtgtgggccacgtcgcgtatgaactgtgcaaacatctgcacaacggcggtgcgaaactg600
atcgtaacggacatcaacaaagaaaacgcggatcgtgcggtgcaagaatttggtgcggag660
ttcgtgcacccggataaaatctatgatgttgaatgcgacatcttcgccccgtgcgcactg720
ggtgctatcatcaacgacgaaactatcgaacgcctgaaatgtaaagtagtcgctggctcc780
gcgaacaaccagctgaaagaagaacgtcatggcaaaatgctggaggaaaaaggtattgta840
tatgcccctgattacgttatcaacgctggtggcgtgatcaacgtagcggacgagctgctg900
ggctataaccgcgaacgcgcgatgaagaaggttgaaggcatctacgacaaaattctgaaa960
gttttcgaaatcgcaaaacgcgatggtatcccgagctacctggctgccgaccgcatggcc1020
gaagaacgtatcgaaatgatgcgtaagacccgcagcacctttctgcaggaccaacgtaac1080
ctgatcaacttcaacaacaaataa1104
<210>2
<211>367
<212>prt
<213>人工序列(artificialsequence)
<400>2
metglylysilepheasptyrmetglulystyrasptyrgluglnleu
151015
valmetcysglnasplysgluserglyleulysalaileilecysile
202530
hisvalthrthrleuglyproalaleuglyglymetargmettrpthr
354045
tyralaserglugluglualailegluaspalaleuargleuglyarg
505560
glymetthrtyrlysasnalaalaalaglyleuasnleuglyglygly
65707580
lysthrvalileileglyaspproarglysasplysasnglualamet
859095
pheargalaleuglyargpheileglnglyleuasnglyargtyrile
100105110
thralagluaspvalglythrthrvalgluaspmetaspileilehis
115120125
glugluthrargtyrvalthrglyvalserproalapheglyserser
130135140
glyasnproserprovalthralatyrglyvaltyrargglymetlys
145150155160
alaalaalalysglualapheglyaspaspserleugluglylysval
165170175
valalavalglnglyvalglyhisvalalatyrgluleucyslyshis
180185190
leuhisasnglyglyalalysleuilevalthraspileasnlysglu
195200205
asnalaaspargalavalglnglupheglyalagluphevalhispro
210215220
asplysiletyraspvalglucysaspilephealaprocysalaleu
225230235240
glyalaileileasnaspgluthrilegluargleulyscyslysval
245250255
valalaglyseralaasnasnglnleulysglugluarghisglylys
260265270
metleugluglulysglyilevaltyralaproasptyrvalileasn
275280285
alaglyglyvalileasnvalalaaspgluleuleuglytyrasnarg
290295300
gluargalametlyslysvalgluglyiletyrasplysileleulys
305310315320
valphegluilealalysargaspglyileprosertyrleualaala
325330335
aspargmetalaglugluargileglumetmetarglysthrargser
340345350
thrpheleuglnaspglnargasnleuileasnpheasnasnlys
355360365
<210>3
<211>1098
<212>dna
<213>人工序列(artificialsequence)
<400>3
atgggtaaaattgtactggttctgtatgacgctggtaaacacgctgcagatgaggaaaaa60
ctgtatggctgtaccgaaaacaaactgggcattgccaactggctgaaagaccagggtcat120
gagctgatcactaccagcgacaaagaaggtgaaacctctgaactggacaaacatatcccg180
gacgccgatatcatcatcaccacgccgttccacccggcctatatcaccaaggagcgtctg240
gataaagccaaaaacctgaagctggttgttgttgctggcgtgggcagcgaccacatcgac300
ctggattacattaaccaaactggtaagaagatttctgttctggaagtcaccggttccaac360
gtagtttctgtcgcagaacacgtggtgatgacgatgctggtcctggtgcgtaacttcgta420
ccggcgcacgaacagatcatcaaccatgattgggaagttgctgcaatcgctaaggacgcg480
tacgacattgaaggcaaaaccatcgctaccatcggtgcaggtcgtatcggttaccgcgta540
ctggagcgcctgctgcctttcaacccgaaagaactgctgtactacgactaccaggcgctg600
ccgaaagaagcggaagaaaaggtaggtgctcgccgtgtagagaacatcgaagaactggtt660
gcccaggcggatatcgtgaccgttaacgcaccgctgcacgcgggtactaaaggcctgatc720
aacaaagagctgctgtctaaattcaagaaaggcgcgtggctggtcaacactgcccgcggt780
gcaatctgcgttgcggaagatgttgcggctgctctggaatccggtcagctgcgcggttac840
ggtggtgatgtctggttcccgcagccggcaccgaaggatcatccgtggcgtgacatgcgt900
aacaaatacggtgcaggtaacgcaatgaccccgcactactctggcaccaccctggacgcc960
cagacccgttatgcggagggcactaaaaacatcctggaatctttcttcaccggcaaattc1020
gactatcgcccgcaggacattatcctgctgaacggtgaatacgtgactaaagcgtacggt1080
aaacacgacaagaaataa1098
<210>4
<211>365
<212>prt
<213>人工序列(artificialsequence)
<400>4
metglylysilevalleuvalleutyraspalaglylyshisalaala
151015
aspgluglulysleutyrglycysthrgluasnlysleuglyileala
202530
asntrpleulysaspglnglyhisgluleuilethrthrserasplys
354045
gluglygluthrsergluleuasplyshisileproaspalaaspile
505560
ileilethrthrprophehisproalatyrilethrlysgluargleu
65707580
asplysalalysasnleulysleuvalvalvalalaglyvalglyser
859095
asphisileaspleuasptyrileasnglnthrglylyslysileser
100105110
valleugluvalthrglyserasnvalvalservalalagluhisval
115120125
valmetthrmetleuvalleuvalargasnphevalproalahisglu
130135140
glnileileasnhisasptrpgluvalalaalailealalysaspala
145150155160
tyraspilegluglylysthrilealathrileglyalaglyargile
165170175
glytyrargvalleugluargleuleupropheasnprolysgluleu
180185190
leutyrtyrasptyrglnalaleuprolysglualagluglulysval
195200205
glyalaargargvalgluasnileglugluleuvalalaglnalaasp
210215220
ilevalthrvalasnalaproleuhisalaglythrlysglyleuile
225230235240
asnlysgluleuleuserlysphelyslysglyalatrpleuvalasn
245250255
thralaargglyalailecysvalalagluaspvalalaalaalaleu
260265270
gluserglyglnleuargglytyrglyglyaspvaltrppheprogln
275280285
proalaprolysasphisprotrpargaspmetargasnlystyrgly
290295300
alaglyasnalametthrprohistyrserglythrthrleuaspala
305310315320
glnthrargtyralagluglythrlysasnileleugluserphephe
325330335
thrglylyspheasptyrargproglnaspileileleuleuasngly
340345350
glutyrvalthrlysalatyrglylyshisasplyslys
355360365
<210>5
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>5
ccatgggtaaaatcttcgactacatg26
<210>6
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>6
aagcttttatttgttgttgaagttga26
<210>7
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>7
catatgggtaaaattgtactggttctgtatgac33
<210>8
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>8
ctcgagttatttcttgtcgtgtttaccgtac31
- 还没有人留言评论。精彩留言会获得点赞!