一种酶改质甜菊糖的制备方法和制备用酶及应用与流程
本发明涉及生物医药技术领域,特别涉及一种酶改质甜菊糖的制备方法和制备用酶及应用。
背景技术:
甜菊糖,又名甜菊糖苷,甜度为蔗糖的200-350倍,具有无毒副作用,食用安全,并且研究表明甜菊糖可用于预防高血压、糖尿病、肥胖症、心脏病和龋齿等病症,是一种可替代蔗糖非常理想的甜味剂。甜菊糖是继甘蔗、甜菜糖之外第三种有开发价值和健康推崇的天然蔗糖替代品,被国际上誉为“世界第三蔗糖”。
甜菊糖是从菊科草本植物甜叶菊的叶子中提取出来的一类含多种成分的双萜糖苷的混合物。其中甜菊苷(stevioside,st)、莱包迪甙a(rebaudiosidea,ra)和莱包迪甙c(rebaudiosidec,rc)的含量较高,共占90%以上。天然甜菊糖由于其本身的结构苷元甜菊醇无甜味且具有苦涩味,会带来不愉快的后味,严重影响了甜菊糖的味质,限制了其更为广泛的工业应用。因此,改善甜菊糖的甜味特征具有重要意义。
目前,有报道利用酶法或发酵法在甜菊糖组分中引入一些新的糖分子,得到的衍生物(或称酶改质甜菊糖)的甜味特性有较大的改善,并减弱后苦涩味。然而这些生产方法大多能耗高,耗时长,纯度与产率均偏低。因此,有必要发展一种工艺简单、耗时短、成本低、产率高且绿色环保的酶改质甜菊糖的制备方法。
技术实现要素:
为了解决上述技术问题,本发明提供了一种酶改质甜菊糖的制备方法和制备用酶及应用;本发明所述制备方法工艺简单、耗时短、产量高和绿色安全。
第一方面,本发明提供了一种酶改质甜菊糖的制备方法,包括:
在溶解有甜菊糖原料和蔗糖(sucrose)的溶液中加入β-果糖苷酶(ffase)得到反应液,调节所述反应液的ph为5.0-8.0,反应温度维持在20-45℃,搅拌反应后,收集得到酶改质甜菊糖,所述甜菊糖原料包括甜菊苷和莱包迪甙a中的一种或多种,所述β-果糖苷酶来源于嗜酸微杆菌属(microbacteriumsaccharophilum)或日本曲霉菌属(aspergillusjaponicus)。
本发明中,所述甜菊苷(st),或称甜菊甙,分子式为c38h60o18,化学结构如式ⅰ所示。所述莱包迪甙a(ra),或称甜菊a苷,分子式为c44h70o23,化学结构如式ⅱ所示:
所述酶改质甜菊糖的制备方法的具体工艺路线如式(1)所示:
其中,所述工艺采用生物酶法,所述甜菊糖原料的化学结构通式如式ⅲ所示,所述酶改质甜菊糖的化学结构通式如式ⅳ所示,其中r基包括二糖基和三糖基。所述二糖基可为β-glc-β-glc-;所述三糖基课可以为(β-glc)2-β-glc-;所述glc为葡萄糖(glucose)。所述蔗糖(sucrose)经β-果糖苷酶(ffase)催化作用下分解得到葡萄糖(glucose)和果糖(fructose,f),其中果糖(fructose,f)分子在β-果糖苷酶催化作用下以β-2,6糖苷键连接至19-o-β-葡萄糖基的6-oh上。
本发明中,所述甜菊糖原料为甜菊苷(st)时,所述酶改质甜菊糖的制备方法的具体工艺路线如式(2)所示:
其中,果糖(f)分子在β-果糖苷酶催化作用下以β-2,6糖苷键连接至甜菊苷的19-o-β-葡萄糖基的6-oh上,得到甜菊苷衍生物(st-f)如式ⅴ所示。所述反应过程中还包括所述β-果糖苷酶催化蔗糖水解并得到葡萄糖。
本发明中,所述甜菊糖原料为莱包迪甙a(ra)时,所述酶改质甜菊糖的制备方法的具体工艺路线如式(3)所示:
其中,果糖(f)分子在β-果糖苷酶催化作用下以β-2,6糖苷键连接至莱包迪甙a的19-o-β-葡萄糖基的6-oh上,得到莱包迪甙a衍生物(ra-f)如式ⅵ所示。所述反应过程中还包括所述β-果糖苷酶催化蔗糖水解并得到葡萄糖。可选地,所述酶改质甜菊糖包括酶改质的甜菊苷和酶改质莱包迪甙a中的一种或多种。具体地,所述酶改质甜菊糖包括所述甜菊苷衍生物(st-f)和所述莱包迪甙a衍生物(ra-f)中的一种或多种。
由于甜菊苷的19位连接的糖基与苦味密切相关,而甜菊苷13-糖基与甜味相关。本发明所述的制备方法通过所述β-果糖苷酶可以快捷、有效地改性甜菊糖原料,以改善甜菊糖的后苦味。通过所述酶改质甜菊糖的制备方法在甜菊糖原料(如st或ra)的19-o-β-葡萄糖基的6-oh上引入果糖基,得到酶改质甜菊糖(如st-f或ra-f)的甜味特性有较大改善。
可选地,所述β-果糖苷酶包括第一β-果糖苷酶或第二β-果糖苷酶,所述第一β-果糖苷酶的基因编码序列包括如seqidno:1所示的核苷酸序列,所述第二β-果糖苷酶的基因编码序列包括如seqidno:2所示的核苷酸序列。
可选地,所述β-果糖苷酶通过微生物表达生成,所述微生物包括大肠杆菌、毕赤酵母和枯草芽孢杆菌中的一种或多种。进一步地,可选地,所述β-果糖苷酶通过大肠杆菌表达生成。所述β-果糖苷酶在所述大肠杆菌表达生成的体系中均属于异源表达。本发明优选地大肠杆菌表达系统,简单可行,培养周期短,发酵成本低,酶产量高。本发明所述β-果糖苷酶可以以冻干粉形式或粗酶液形式添加至反应体系中。
可选地,本发明所述制备方法中,调节所述反应液的ph为5.0-8.0。进一步可选地,所述反应液的ph为7.0-8.0。优选地,所述反应液的ph为7.2-8.0。本发明所述反应液的ph可偏碱性,有助于提升所述β-果糖苷酶反应活性,提高所述酶改质甜菊糖的产率,缩短反应时间。
可选地,所述反应液的反应温度维持在20-45℃。进一步地,可选地,所述反应液的反应温度维持在20-30℃。优选地,所述反应液的反应温度维持在20-28℃。例如,所述反应液的反应温度维持在20℃,或为25℃,或为28℃,或为35℃。
可选地,所述搅拌反应的反应时间为2-5小时。进一步可选地,所述搅拌反应的反应时间为2-4小时。本发明所述制备方法耗时短,所述反应时间为优选反应时间,当增加反应时间时,所述制备方法的得到的产品的转化率会降低。
可选地,所述搅拌反应的搅拌速率为200-300rpm。
可选地,所述收集得到酶改质甜菊糖的过程包括:对所述反应液加热以使所述β-果糖苷酶变性,过滤并收集滤液,将所述滤液纯化处理后得到酶改质甜菊糖,其中,所述加热的温度为85-100℃,时间为0.3-1小时。可选地,所述加热的温度还可以为90-100℃,时间为0.5-1小时。
可选地,所述甜菊糖原料在所述反应液中的质量分数为1%-20%。进一步地,可选地,所述甜菊糖原料在所述反应液中的质量分数为10%-20%。例如,所述甜菊糖原料在所述反应液中的质量分数为10%,或为12%,或为15%,或为20%。
可选地,所述甜菊糖原料与所述β-果糖苷酶的质量比为1:(0.1-2)。进一步地,可选地,所述甜菊糖原料与所述β-果糖苷酶的质量比为1:(0.5-2)。
可选地,所述甜菊糖原料与所述蔗糖的质量比为1:(1-10)。进一步地,可选地,所述甜菊糖原料与所述蔗糖的质量比为1:(2-5)。
可选地,,所述反应液中还包括缓冲液,所述缓冲液包括磷酸盐缓冲液和tris-hcl缓冲液中的任意一种或多种。可选地,所述缓冲液还包括其他种类缓冲液。具体地,所述缓冲液包括磷酸钠缓冲液。可选地,所述缓冲液的浓度为10-1000mmol/l。进一步可选地,所述缓冲液的浓度为10-1000mmol/l。优选地,所述缓冲液的浓度为100-500mmol/l。例如,所述缓冲液的浓度为100mmol/l,或为200mmol/l,或为500mmol/l。
由于不同微生物种属来源的β-果糖苷酶的酶学性质存在差异,包括酶的比活性、酶作用的底物范围、最适ph、最适温度、作用时间和酶的稳定性等方面。本发明所述β-果糖苷酶包括第一β-果糖苷酶或第二β-果糖苷酶;其中,第一β-果糖苷酶来源于嗜酸微杆菌属,第二β-果糖苷酶来源于日本曲霉菌属。本发明所述β-果糖苷酶既具有水解酶性质,可以将蔗糖中的果糖分子水解下来,有具有逆向催化功能,能够将果糖分子加合上所述甜菊苷和莱包迪甙a的19-o-β-葡萄糖基的6-oh上。
本发明第一方面所提供的酶改质甜菊糖的制备方法,所述制备方法工艺简单,成本低廉,耗时短,绿色环保;由所述制备方法制得的酶改质甜菊糖具有极高的产率。在底物(甜菊糖原料)和酶的投料比相同的情况下,现有技术仅支持底物浓度在千分之几的酶催化反应进行,一般转化率仅为40-50%,且仅在底物浓度接近最低值时,转化率才能达到60%左右,或者通过补加原料用于提高转化率;而本发明所述的制备方法可支持较高的底物浓度(如5%-20%)下,转化率可接近90%左右;因此,本发明所述的制备成本低,适于工业化生产。
第二方面,本发明还提供了一种酶改质甜菊糖的制备用酶,其中,所述制备用酶包括β-果糖苷酶,所述β-果糖苷酶包括第一β-果糖苷酶或第二β-果糖苷酶,所述第一β-果糖苷酶的基因编码序列包括如seqidno:1所示的核苷酸序列,所述第二β-果糖苷酶的基因编码序列包括如seqidno:2所示的核苷酸序列;所述第一β-果糖苷酶来源于嗜酸微杆菌属,所述第二β-果糖苷酶来源于日本曲霉菌属。
可选地,所述第一β-果糖苷酶的氨基酸序列包括如seqidno:3所示的氨基酸序列。所述第二β-果糖苷酶的氨基酸序列包括如seqidno:4所示的氨基酸序列。
其中,所述如seqidno:3所示的氨基酸序列的基因编码序列如seqidno:1所示;可选的,所述第一β-果糖苷酶的氨基酸序列的基因编码序列应该考虑简并碱基,即如seqidno:3所示的氨基酸序列的编码基因包括如seqidno:1所示的核苷酸序列,保护范围还应该保护与seqidno:1具有碱基简并性质的核苷酸序列,这些核苷酸序列对应的氨基酸序列仍然为seqidno:3。同样的,对于所述第二β-果糖苷酶,所述如seqidno:4所示的氨基酸序列的编码基因同样应该考虑简并碱基。
可选地,所述β-果糖苷酶通过构建重组质粒在微生物中表达,所述重组质粒的载体质粒为pet28a(+)载体质粒。将所述第一β-果糖苷酶和/或第二所述β-果糖苷酶的基因编码序列插入至所述pet28a(+)载体质粒中得到重组质粒,所述重组质粒可以高效、高产的在微生物细胞中的异源表达得到所述第一β-果糖苷酶和/或所述第二β-果糖苷酶。
可选地,所述第一β-果糖苷酶和/或第二所述β-果糖苷酶的基因编码序列上增设his标签(组氨酸标签)的核苷酸序列,能使表达后的蛋白带上his标签,his标签有利于表达后蛋白的分离纯化,及在实验中的分析和追踪,比如用于免疫印迹实验时的分析。
本发明第二方面提供的酶改质甜菊糖的制备用酶—β-果糖苷酶,具有很好的生物活性,纯度高,可广泛应用于食品甜味剂制备、生物制药等领域。相比于传统其他发酵体系,本发明优选的β-果糖苷酶具有更高的产率,耗时短,具有更强的生物活性及特异性。
本发明第三方面提供了β-果糖苷酶以及含有所述β-果糖苷酶基因的微生物菌株在生物催化中的应用,其中,所述β-果糖苷酶由来源于嗜酸微杆菌属或日本曲霉菌属的β-果糖苷酶基因编码,所述β-果糖苷酶催化式ⅲ所示的化合物转化成式ⅳ所示的化合物,
其中,当所述r基为β-glc-β-glc-时,所述式ⅲ所示的化合物为甜菊苷(stevioside);当所述r基为(β-glc)2-β-glc-时,所述式ⅲ所示的化合物为莱包迪甙a(rebaudiosidea);当所述r基为(β-glc,α-rha-)-β-glc-时,所述式ⅲ所示的化合物为莱包迪甙c(rebaudiosidec);当所述r基为α-rha-β-glc-时,所述式ⅲ所示的化合物为杜克甙a(dulcosidea);所述r基为β-glc-时,所述式ⅲ所示的化合物为甜茶苷(rubusoside);当所述r基为(β-glc,β-xyl)-β-glc-时,所述式ⅲ所示的化合物为莱包迪甙f(rebaudiosidef)。具体地,所述β-果糖苷酶可以将果糖基以β-2,6糖苷键连接至所述式ⅲ所示的化合物的19-o-β-葡萄糖基的6-oh上得到所述式ⅳ所示的化合物。例如,所述β-果糖苷酶可以将果糖基以β-2,6糖苷键连接至所述甜菊苷的19-o-β-葡萄糖基的6-oh上得到甜菊苷衍生物;或将果糖基以β-2,6糖苷键连接至所述甜菊苷的19-o-β-葡萄糖基的6-oh上得到莱包迪甙a衍生物等。
可选地,所述β-果糖苷酶具有如seqidno:3或seqidno:4所示的氨基酸序列。所述β-果糖苷酶基因编码序列包括如seqidno:1或seqidno:2所示的核苷酸序列。本发明还提供了一种重组质粒,所述重组质粒包括第一β-果糖苷酶或第二β-果糖苷酶的基因编码序列,其中,所述第一β-果糖苷酶的基因编码序列包括如seqidno:1所示的核苷酸序列,所述第二β-果糖苷酶的基因编码序列包括如seqidno:2所示的核苷酸序列。
可选地,所述第一β-果糖苷酶或所述第二β-果糖苷酶的基因编码序列插入到pet28a(+)载体质粒的bamhi和hindiii酶切位点之间。所述第一β-果糖苷酶或所述第二β-果糖苷酶的基因编码序列插入到pet28a(+)载体质粒时,所述第一β-果糖苷酶或所述第二β-果糖苷酶的基因编码序列的5’端可加入起始密码子(如atg)与pet28a(+)载体质粒中bamhⅰ酶切位点相连,3’端可加入终止密码子(如taa)与pet28a(+)载体质粒中hindiii酶切位点相连。
本发明还提供了一种重组质粒的制备方法,包括:
(1)提供上游引物和下游引物,所述上游引物和下游引物的碱基序列分别如seqidno:5-seqidno:8所示;
(2)提供或制备第一β-果糖苷酶或第二β-果糖苷酶基因模板,并以步骤(1)所述上游引物和所述下游引物为pcr引物,扩增所述第一β-果糖苷酶或所述第二β-果糖苷酶基因片段;
(3)取pet28a(+)载体质粒,将步骤(2)扩增得到的所述第一β-果糖苷酶或所述第二β-果糖苷酶基因片段和所述pet28a(+)载体质粒利用相同的内切酶分别进行双酶切反应,纯化回收后进行连接,得到所述重组质粒。
可选地,所述步骤(3)中,所示双酶切反应的酶切位点可以为bamhi和hindiii内切酶。
本发明的有益效果包括以下几个方面:
1、本发明所述的制备方法采用生物酶法,操作简便,耗时短、转化率高和绿色安全,可以广泛适用于工业化规模生产;
2、本发明所述的制备方法,底物终浓度可高到达1%-20%,远远高于传统工艺底物终浓度;
3、由本发明所述的制备方法制备的酶改质甜菊糖,性质稳定,甜度高,后苦涩味极大改善,热值低,可广泛应用在食品工业及制药领域;
4、本发明所述制备用酶—β-果糖苷酶的生物活力好,特异性强。
附图说明
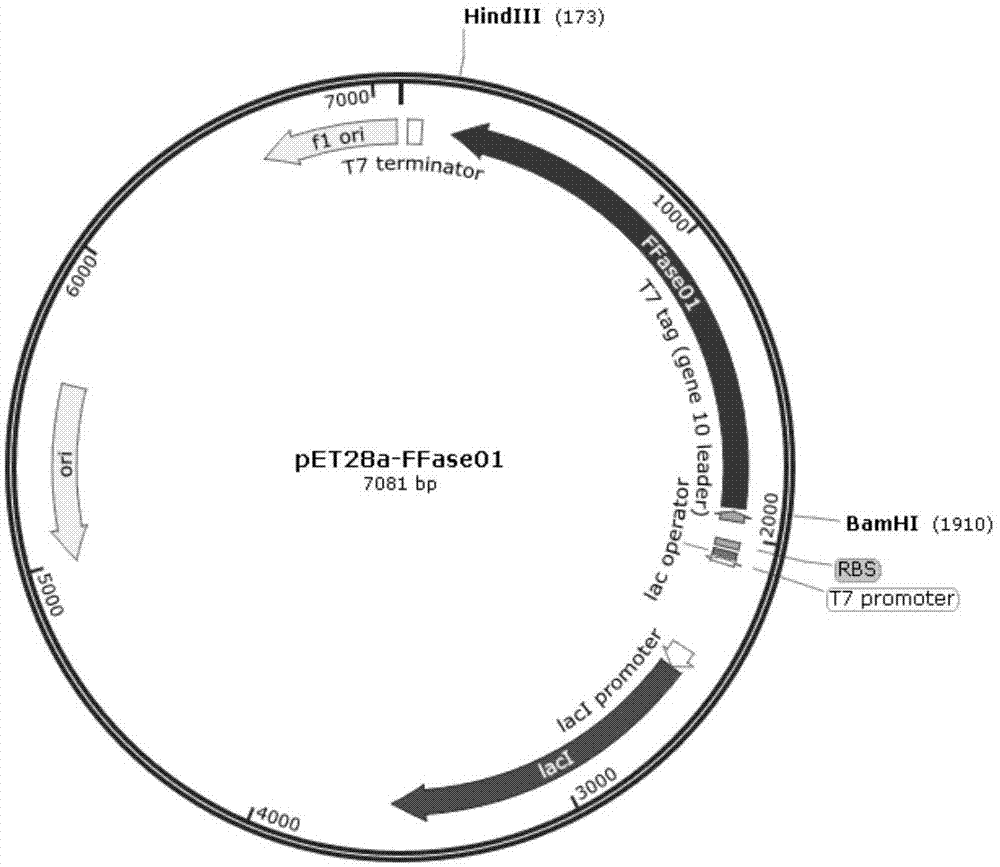
图1为本发明一实施例提供的pet28a-ffase01重组质粒的质粒图谱;
图2为本发明一实施例提供的pet28a-ffase02重组质粒的质粒图谱;
图3为本发明一实施例提供的ra-f的质谱图;
图4为本发明一实施例提供的st-f的质谱图。
具体实施方式
以下所述是本发明实施例的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明实施例原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也视为本发明实施例的保护范围。
若无特别说明,本发明实施例所采用的原料及其它化学试剂皆为市售商品。
(1)构建pet28a-ffase01、pet28a-ffase02重组质粒
a)提供上游引物和下游引物,并通过实验得到的β-果糖苷酶的基因编码序列。其中,所述β-果糖苷酶(ffase)包括第一β-果糖苷酶(ffase01)或第二β-果糖苷酶(ffase02)。所述第一β-果糖苷酶的基因编码序列包括如seqidno:1所示的核苷酸序列,所述第一β-果糖苷酶来源于嗜酸微杆菌属;所述第二β-果糖苷酶的基因编码序列包括如seqidno:2所示的核苷酸序列,所述第二β-果糖苷酶来源于日本曲霉菌属。所述第一β-果糖苷酶对应的上游引物的碱基序列如seqidno:5所示,下游引物的碱基序列如seqidno:6所示。所述第二β-果糖苷酶对应的上游引物的碱基序列如seqidno:7所示,下游引物的碱基序列如seqidno:8所示。
b)将所述ffase01或ffase02的基因编码序列插入到pet28a(+)载体质粒的bamhi和hindiii酶切位点之间。所述ffase01或ffase02的基因编码序列插入到pet28a(+)载体质粒时,所述ffase01或ffase02的基因编码序列的5’端添加起始密码子(如atg)与pet28a(+)载体质粒中bamhⅰ酶切位点相连,3’端还添加有终止密码子(如taa)与pet28a(+)载体质粒中hindiii酶切位点相连。然后转入大肠杆菌感受态细胞dh5α,进行阳性克隆pcr鉴定和测序鉴定。经过pcr产物凝胶电泳检测和测序鉴定符合目的片段大小和序列,成功构建pet28a-ffase01或pet28a-ffase02重组质粒。如图1和图2分别是所示为pet28a-ffase01或pet28a-ffase02重组质粒图谱。
(2)表达β-果糖苷酶ffase01或ffase02
将构建的重组质粒pet28a-ffase01和pet28a-ffase02中的一种或多种转入大肠杆菌bl21(de3)中,并以1%的接种量接种至含有4ml的lb培养基中,维持恒定的37℃,200rpm的摇晃速率,过夜培养后,将菌液以1%的接种量转接到含有1l的lb培养基(50μg/ml卡那霉素)的2l三角瓶中,继续37℃恒温培养至培养基中的od600值达到0.6左右,加入终浓度为度0.1mm-1mm的诱导剂iptg,在20-37℃条件培养12-16小时后离心收集菌体。将菌体用50mm磷酸缓冲液(ph=7.4)进行重悬并经超声破碎和离心,收集上清液得到含有ffase01或ffase02的粗酶液。
将表达获得的含有ffase01或ffase02的粗酶液进行sds-聚丙烯酰胺凝胶电泳(sds-page)鉴定。本实施方式表达获得的粗酶液中,所述ffase01和ffase02的分子大小均与相应蛋白的理论计算值相近,其中,ffase01的理论分子量为64kda,ffase02的理论分子量为70kda。此外,对收集得到的粗酶液可进行进一步纯化,可制得ffase01或ffase02的冻干粉。
(3)ffase01或ffase02催化活性比较
按上述步骤(1)及步骤(2)的操作过程,获取其他来源的β-果糖苷酶ffase03-ffase05;其中,β-果糖苷酶ffase03来源于许旺氏酵母(schwanniomycesoccidentalis),β-果糖苷酶ffase04来源于卷曲乳杆菌(lactobacilluscrispatus),β-果糖苷酶ffase05来源于菊苣(cichoriumintybus)。设计反应体系:在1ml的磷酸钠缓冲液中,分别加入甜菊苷或莱包迪甙a5mg,加入蔗糖20mg,搅拌至完全溶解;继续加入上述ffase01-ffase05中的任意一种β-果糖苷酶的粗酶液200μl(50mg),调节ph至7.4;在20℃、200rpm搅拌速率下反应5h,收集得到酶改质甜菊糖,并评估每种β-果糖苷酶的酶活情况,见下表所示:
实施例1
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入莱包迪甙a50g和蔗糖100g,搅拌至完全溶解;继续加入10gffase01冻干酶粉,调节ph至7.4;在35℃、250rpm搅拌速率下反应2h,反应完成后将反应液加热至100℃热处理0.5h,使ffase01蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖ra-f51.73g,纯度>95%。本实施例中,每隔一定时间取反应液进行质谱分析;图3是实验过程中检测的酶改质甜菊糖ra-f的质谱图,并根据液相色谱对反应液进行测定的数据计算得到转化率为91.5%。
实施例2
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷50g和蔗糖100g,搅拌至完全溶解;继续加入10gffase01冻干酶粉,调节ph至7.4;在35℃、250rpm搅拌速率下反应2h,反应完成后将反应液加热至100℃热处理0.5h,使ffase01蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f51.85g,纯度>95%。本实施例中,每隔一定时间取反应液进行质谱分析;图4是实验过程中检测的酶改质甜菊糖st-f的质谱图,并根据液相色谱对反应液进行测定的数据计算得到转化率为92.0%。
实施例3
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷50g和蔗糖100g,搅拌至完全溶解;继续加入10gffase02冻干酶粉,调节ph至7.4;在28℃、250rpm搅拌速率下反应2h,反应完成后将反应液加热至100℃热处理0.5h,使ffase02蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f52.64g,纯度>95%;实验测得转化率为92.4%。
实施例4
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷50g和蔗糖200g,搅拌至完全溶解;继续加入5gffase02冻干酶粉,调节ph至7.4;在28℃、250rpm搅拌速率下反应2h,反应完成后将反应液加热至100℃热处理0.5h,使ffase02蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f53.27g,纯度>95%;实验测得转化率为93.5%。
实施例5
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷50g和蔗糖100g,搅拌至完全溶解;继续加入5gffase02冻干酶粉,调节ph至8.0;在28℃、250rpm搅拌速率下反应2h,反应完成后将反应液加热至100℃热处理0.5h,使ffase02蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f51.98g,纯度>95%;实验测得转化率为91.3%。
实施例6
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷50g和蔗糖100g,搅拌至完全溶解;继续加入5gffase02冻干酶粉,调节ph至7.4;在28℃、250rpm搅拌速率下反应4h,反应完成后将反应液加热至100℃热处理0.5h,使ffase02蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f52.72g,纯度>95%;实验测得转化率为92.6%。
实施例7
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷50g和蔗糖100g,搅拌至完全溶解;继续加入5gffase02冻干酶粉,调节ph至8.0;在20℃、250rpm搅拌速率下反应4h,反应完成后将反应液加热至100℃热处理0.5h,使ffase02蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f52.39g,纯度>95%;实验测得转化率为92.1%。
实施例8
一种酶改质甜菊糖的制备方法,包括:
在500ml的磷酸钠缓冲液中,分别加入甜菊苷100g和蔗糖200g,搅拌至完全溶解;继续加入20gffase02冻干酶粉,调节ph至7.4;在28℃、250rpm搅拌速率下反应2h,反应完成后将反应液加热至100℃热处理0.5h,使ffase02蛋白变性并过滤除去蛋白,收集滤液后进行喷雾干燥得到酶改质甜菊糖粗品,将所述酶改质甜菊糖粗品经硅胶树脂分离、结晶等后处理纯化后得到酶改质甜菊糖st-f107.21g,纯度>95%;实验测得转化率为91.3%。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
序列表
<110>邦泰生物工程(深圳)有限公司
江西邦泰绿色生物合成生态产业园发展有限公司
<120>一种酶改质甜菊糖的制备方法和制备用酶及应用
<150>pct/cn2018/088358
<151>2018-05-25
<160>8
<170>siposequencelisting1.0
<210>1
<211>1731
<212>dna
<213>人工序列(artificialsequence)
<400>1
acccatagcacccgcggccgcgtgcgccgcgtgctggcgggcggcctggcgaccagcacc60
ctggcggcggcggtgctgattgcgggcgcggcgccggcgaccgcgcagagcggcctgcag120
gatggcccggaaccgaccattcatacccagcaggcgtatgcgccggaagatgattttacc180
gcgaaatggacccgcgcggatgcgcgccagctgcagcgcatgagcgatccgaccgcgccg240
agccgcgaaaacagcatgccggcgagcgtgaccatgccgaccgtgccgcaggattttccg300
gatatgagcaacgaacaggtgtgggtgtgggatacctggccgctgaccgatgaagatgcg360
aaccagtatagcgtgaacggctgggaaattatttttagcctggtggcggatcgcaacctg420
ggctttgatgatcgccatgtgtttgcgaaaattggctatttttatcgcccggcgggcgtg480
ccggcggcggaacgcccggaaaacggcggctggacctatggcggcctggtgtttaaagaa540
ggcgtgaccggccagatttttgaagatcagagctttagccatcagacccagtggagcggc600
agcgcgcgcgtgagcaaaaacggcgaaattaaactgttttttaccgatgtggcgttttat660
cgcaacagcgatggcaccaacattaaaccgtatgatccgcgcattgcgctgagcgtgggc720
aaagtgaaagcgaacaaaaaaggcgtgaccctgaccggctttaacaaagtgaccgatctg780
ctgcaggcggatggcacctattatcagaccggcgcgcagaacgaattttttaactttcgc840
gatccgtttacctttgaagatccggcgcatccgggcgaaacctttatggtgtttgaaggc900
aacagcgcgatgcagcgcgaaaccgcgacctgcaacgaagcggatctgggctatcgccag960
ggcgatccgtatgcggaaaccgtggatgatgtgaacgcgagcggcgcgacctatcagatt1020
ggcaacgtgggcctggcgaaagcgaaaaacaaacagctgaccgaatgggaatttctgccg1080
ccgattctgagcgcgaactgcgtgaccgatcagaccgaacgcccgcagatttattttaaa1140
gatggcaaaagctatctgtttaccattagccatcgcggcacctttgcggcgggcctggat1200
ggcccggaaggcgtgtatggctttgtgggcgatggcattcgcagcgattatcagccgctg1260
aacggcggcagcggcctggcgctgggcaacccgaccaacctgaactttctgggcggccag1320
ccgtttgcgccggattttaaccagcatccgggccattttcaggcgtatagccattatgtg1380
atgccgggcggcctggtgcagagctttattgataccattggcacccatgatgattttgtg1440
cgcggcggcaccctggcgccgaccgtgaaaatggatattggcgtgggcggcgatccgacc1500
aaaaccgcggtggattatagctatggcagcgaaggcctgggcggctgggcggatattccg1560
gcgaacaaacatctgtttaccaacggcaaatttggcgtggcggtgagcgatgaagcggcg1620
cagaaaattcgcaaaattctgggcagcaaatttgatgattatctggatggcaaaccggtg1680
agcgcgaccgtgcgcgcgctgattgaaaaactgctggcgcagtatggcggc1731
<210>2
<211>1911
<212>dna
<213>人工序列(artificialsequence)
<400>2
aaactgaccaccaccaccctggcgctggcgaccggcgcggcggcggcggaagcgagctat60
catctggataccaccgcgccgccgccgaccaacctgagcaccctgccgaacaacaccctg120
tttcatgtgtggcgcccgcgcgcgcatattctgccggcggaaggccagattggcgatccg180
tgcgcgcattataccgatccgagcaccggcctgtttcatgtgggctttctgcatgatggc240
gatggcattgcgggcgcgaccaccgcgaacctggcgacctataccgataccagcgataac300
tttctgattcagccgggcggcaaaaacgatccggtggcggtgtttgatggcgcggtgatt360
ccggtgggcgtgaacaacaccccgaccctgctgtataccagcgtgagctttctgccgatt420
tggagcattccgtatacccgcggcagcgaaacccagagcctggcggtggcgcgcgatggc480
ggccgccgctttgataaactggatcagggcccggtgattgcggatcatccgtttgcggtg540
gatgtgaccgcgtttcgcgatccgtttgtgtttcgcagcgcgaaactggatgtgctgctg600
gatgaagaagtggcgcgcaacgaaaccgcggtgcagcaggcggtggatggctggaccgaa660
aaaaacgcgccgtggtatgtggcggtgagcggcggcgtgcatggcgtgggcccggcgcag720
tttctgtatcgccagaacggcggcaacgcgagcgaatttcagtattgggaatatctgggc780
gaatggtggcaggaagcgaccaacagcagctggggcgatgaaggcacctgggcgggccgc840
tggggctttaactttgaaaccggcaacgtgctgtttctgaccgaagaaggccatgatccg900
cagaccggcgaagtgtttgtgaccctgggcaccgaaggcagcggcctgccgccgcaggtg960
agcagcattcatgatatgctgtgggcggcgggcgaagtgggcgtgggcagcgaacaggaa1020
aaagtggaatttagcccgagcatggcgggctttctggattggggctttagcgcgtatgcg1080
gcggcgggcaaagtgctgccggcgagcagcgcggtgagcaaaaccagcggcgtggaagtg1140
gatcgctatgtgagctttgtgtggctgaccggcgatcagtatgaacaggcggatggcttt1200
ccgaccgcgcagcagggctggggcagcctgctgctgccgcgcgaactgaaaaccgtggaa1260
aacgtggtggataacgaactggtgcgcgaagaaggcgtgagctgggtggtgggcgaaagc1320
gataaccagaccgcgcgcctgcgcaccctgggcattaccattgcgcgcgaaaccaaagcg1380
gcgctgctggcgaacggcagcgtgaccgcggaagaagatcgcaccctgcagaccgcggcg1440
gtggtgccgtttgcgcagagcccgagcagcaaattttttgtgctgaccgcgcagctggat1500
gcgagcgcgcgcagcagcccgctgcagagcggctttgaaattctggcgagcgaactggaa1560
cgcaccgcgatttattatcagtttagcaacgaaagcctggtggtggatcgcagccagacc1620
agcgcggcggcgccgaccaacccggatagctttaccgaaagcggcaaactgcgcctgttt1680
gatgtgattgaaaacggccaggaacaggtggaaaccctggatctgaccgtggtggtggat1740
aacgcggtggtggaagtgtatgcgaacggccgctttgcgctgagcacctgggcgcgcagc1800
tggtatgataacagcacccagattcgcttttttcataacggcgaaggcgaagtgcagttt1860
cgcaacgtgagcgtgagcgaaggcctgtataacgcgtggccggaacgcaac1911
<210>3
<211>577
<212>prt
<213>嗜酸微杆菌属(microbacteriumsaccharophilum)
<400>3
thrhisserthrargglyargvalargargvalleualaglyglyleu
151015
alathrserthrleualaalaalavalleuilealaglyalaalapro
202530
alathralaglnserglyleuglnaspglyprogluprothrilehis
354045
thrglnglnalatyralaprogluaspaspphethralalystrpthr
505560
argalaaspalaargglnleuglnargmetseraspprothralapro
65707580
serarggluasnsermetproalaservalthrmetprothrvalpro
859095
glnasppheproaspmetserasngluglnvaltrpvaltrpaspthr
100105110
trpproleuthraspgluaspalaasnglntyrservalasnglytrp
115120125
gluileilepheserleuvalalaaspargasnleuglypheaspasp
130135140
arghisvalphealalysileglytyrphetyrargproalaglyval
145150155160
proalaalagluargprogluasnglyglytrpthrtyrglyglyleu
165170175
valphelysgluglyvalthrglyglnilephegluaspglnserphe
180185190
serhisglnthrglntrpserglyseralaargvalserlysasngly
195200205
gluilelysleuphephethraspvalalaphetyrargasnserasp
210215220
glythrasnilelysprotyraspproargilealaleuservalgly
225230235240
lysvallysalaasnlyslysglyvalthrleuthrglypheasnlys
245250255
valthraspleuleuglnalaaspglythrtyrtyrglnthrglyala
260265270
glnasngluphepheasnpheargaspprophethrphegluasppro
275280285
alahisproglygluthrphemetvalphegluglyasnseralamet
290295300
glnarggluthralathrcysasnglualaaspleuglytyrarggln
305310315320
glyaspprotyralagluthrvalaspaspvalasnalaserglyala
325330335
thrtyrglnileglyasnvalglyleualalysalalysasnlysgln
340345350
leuthrglutrpglupheleuproproileleuseralaasncysval
355360365
thraspglnthrgluargproglniletyrphelysaspglylysser
370375380
tyrleuphethrileserhisargglythrphealaalaglyleuasp
385390395400
glyprogluglyvaltyrglyphevalglyaspglyileargserasp
405410415
tyrglnproleuasnglyglyserglyleualaleuglyasnprothr
420425430
asnleuasnpheleuglyglyglnprophealaproasppheasngln
435440445
hisproglyhispheglnalatyrserhistyrvalmetproglygly
450455460
leuvalglnserpheileaspthrileglythrhisaspasppheval
465470475480
argglyglythrleualaprothrvallysmetaspileglyvalgly
485490495
glyaspprothrlysthralavalasptyrsertyrglyserglugly
500505510
leuglyglytrpalaaspileproalaasnlyshisleuphethrasn
515520525
glylyspheglyvalalavalseraspglualaalaglnlysilearg
530535540
lysileleuglyserlyspheaspasptyrleuaspglylysproval
545550555560
seralathrvalargalaleuileglulysleuleualaglntyrgly
565570575
gly
<210>4
<211>637
<212>prt
<213>日本曲霉菌属(aspergillusjaponicus)
<400>4
lysleuthrthrthrthrleualaleualathrglyalaalaalaala
151015
glualasertyrhisleuaspthrthralaproproprothrasnleu
202530
serthrleuproasnasnthrleuphehisvaltrpargproargala
354045
hisileleuproalagluglyglnileglyaspprocysalahistyr
505560
thraspproserthrglyleuphehisvalglypheleuhisaspgly
65707580
aspglyilealaglyalathrthralaasnleualathrtyrthrasp
859095
thrseraspasnpheleuileglnproglyglylysasnaspproval
100105110
alavalpheaspglyalavalileprovalglyvalasnasnthrpro
115120125
thrleuleutyrthrservalserpheleuproiletrpserilepro
130135140
tyrthrargglysergluthrglnserleualavalalaargaspgly
145150155160
glyargargpheasplysleuaspglnglyprovalilealaasphis
165170175
prophealavalaspvalthralapheargaspprophevalphearg
180185190
seralalysleuaspvalleuleuaspglugluvalalaargasnglu
195200205
thralavalglnglnalavalaspglytrpthrglulysasnalapro
210215220
trptyrvalalavalserglyglyvalhisglyvalglyproalagln
225230235240
pheleutyrargglnasnglyglyasnalaserglupheglntyrtrp
245250255
glutyrleuglyglutrptrpglnglualathrasnsersertrpgly
260265270
aspgluglythrtrpalaglyargtrpglypheasnphegluthrgly
275280285
asnvalleupheleuthrglugluglyhisaspproglnthrglyglu
290295300
valphevalthrleuglythrgluglyserglyleuproproglnval
305310315320
serserilehisaspmetleutrpalaalaglygluvalglyvalgly
325330335
sergluglnglulysvalglupheserprosermetalaglypheleu
340345350
asptrpglypheseralatyralaalaalaglylysvalleuproala
355360365
serseralavalserlysthrserglyvalgluvalaspargtyrval
370375380
serphevaltrpleuthrglyaspglntyrgluglnalaaspglyphe
385390395400
prothralaglnglnglytrpglyserleuleuleuproarggluleu
405410415
lysthrvalgluasnvalvalaspasngluleuvalarggluglugly
420425430
valsertrpvalvalglygluseraspasnglnthralaargleuarg
435440445
thrleuglyilethrilealaarggluthrlysalaalaleuleuala
450455460
asnglyservalthralaglugluaspargthrleuglnthralaala
465470475480
valvalprophealaglnserproserserlysphephevalleuthr
485490495
alaglnleuaspalaseralaargserserproleuglnserglyphe
500505510
gluileleualasergluleugluargthralailetyrtyrglnphe
515520525
serasngluserleuvalvalaspargserglnthrseralaalaala
530535540
prothrasnproaspserphethrgluserglylysleuargleuphe
545550555560
aspvalilegluasnglyglngluglnvalgluthrleuaspleuthr
565570575
valvalvalaspasnalavalvalgluvaltyralaasnglyargphe
580585590
alaleuserthrtrpalaargsertrptyraspasnserthrglnile
595600605
argphephehisasnglygluglygluvalglnpheargasnvalser
610615620
valsergluglyleutyrasnalatrpprogluargasn
625630635
<210>5
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>5
cgcggatccatgacccatagcacccgcgg29
<210>6
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>6
cccaagcttcggcggtatgacgcggtcgt29
<210>7
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>7
cgcggatccatgaaactgaccaccaccacc30
<210>8
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>8
cccaagcttcaacgcaaggccggtgcgcaat31
- 还没有人留言评论。精彩留言会获得点赞!