新型天冬氨酸激酶变体和使用其生产L-氨基酸的方法与流程
本公开内容涉及天冬氨酸激酶变体,包含该变体的微生物,以及通过使用该微生物生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的方法。
背景技术:
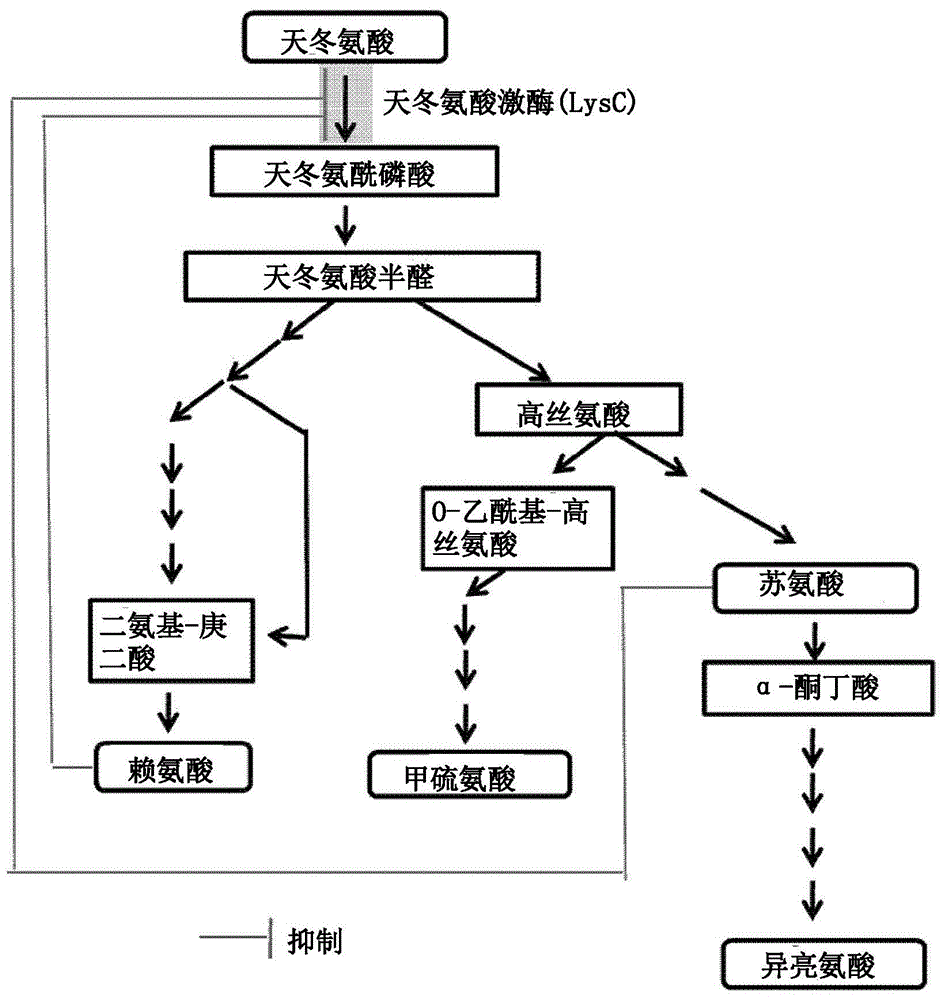
:棒状杆菌属特别是谷氨酸棒杆菌的微生物是广泛用于生产l-氨基酸和其它有用物质的革兰氏阳性微生物。为了生产l-氨基酸和其它有用物质,已经进行了各种研究以开发用于发酵过程的高效生产微生物和技术。例如,主要使用通过增加编码参与l-赖氨酸生物合成酶的基因的表达或通过敲除生物合成不必要的基因来特异性接近靶物质的方法(韩国专利号10-0838038)。同时,在l-氨基酸中,l-赖氨酸、l-苏氨酸、l-甲硫氨酸、l-异亮氨酸和l-甘氨酸是衍生自天冬氨酸(asp)的氨基酸,并且这些氨基酸通常使用通过天冬氨酸激酶(lysc,e.c.2.7.2.4)由asp产生的天冬氨酰磷酸(app)(图1)。因此,为了通过微生物发酵方法生产氨基酸,重要的是使生物合成途径中使用的酶的活性维持在一定水平或更高水平,并且因此在这方面进行了深入研究。具体地,已知通过l-赖氨酸和l-苏氨酸的反馈抑制来调节lysc——其作为天冬氨酸衍生的氨基酸的生物合成途径中的第一种酶——的活性(jmolbiol.2007apr27;368(2):521-36.epub2007年2月20日)。在这方面,尽管已经提交了与反馈抑制相关的应用(美国专利号us8062869b和日本专利号jp3473042b),但是仍需要继续研究以提高天冬氨酸衍生产物的生产力。在这种情况下,本发明人通过确认当使用新型天冬氨酸激酶变体时天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的产生得到改善而完成本公开内容。技术实现要素:技术问题本发明的一个目的是提供一种天冬氨酸激酶变体,其包括在seqidno:1的氨基酸序列中的一个或多个氨基酸取代,其中氨基酸取代包括用l-赖氨酸或l-甲硫氨酸取代seqidno:1的氨基酸序列中第377位的氨基酸。本公开内容的另一个目的是提供编码变体的多核苷酸。本公开内容的仍另一个目的是提供生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的棒状杆菌属的微生物,该微生物包括天冬氨酸激酶变体或具有其增强的活性。本公开内容的仍另一个目的是提供用于生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的方法,包括:在培养基中培养微生物;和从培养的微生物或培养基中回收天冬氨酸衍生的l-氨基酸或其氨基酸衍生物。本公开内容的仍另一个目的是提供用于生产甲硫氨酸的方法,其包括:在培养基中培养微生物;从培养的微生物或培养基生产乙酰高丝氨酸或琥珀酰高丝氨酸;和将乙酰高丝氨酸或琥珀酰高丝氨酸转化为甲硫氨酸。技术方案在下文中将详细描述本公开内容。同时,本文公开的每个解释和示例性实施方式可以应用于其他解释和示例性实施方式。也就是说,本文公开的各种因素的所有组合都属于本公开内容的范围。此外,本公开内容的范围不应被下文提供的具体公开内容所限制。此外,基于常规实验,本领域普通技术人员将能够识别或确认本申请中描述的本公开内容的具体实施方式的许多等同物,并且这些等同物旨在包括在本公开内容中。为了实现上述目的,本公开内容的一个方面提供了一种天冬氨酸激酶变体,其包括在seqidno:1的氨基酸序列中的一个或多个氨基酸取代,其中氨基酸取代包括用另一种氨基酸取代第377位的氨基酸。具体地,本公开内容的目的是提供一种天冬氨酸激酶变体,其中seqidno:1的氨基酸序列中第377位的氨基酸被l-赖氨酸或l-甲硫氨酸取代。如本文所使用,术语“天冬氨酸激酶”是指催化氨基酸、天冬氨酸的磷酸化,并且在生物合成三种必需氨基酸l-甲硫氨酸、l-赖氨酸和l-苏氨酸——其被称为天冬氨酸家族——的第一步中起作用的酶。在本公开内容中,术语“天冬氨酸激酶”可以与“lysc”或“lysc蛋白”互换使用。lysc蛋白可以从已知数据库ncbi的genbank获得。lysc蛋白可以是衍生自棒状杆菌属的lysc,并且具体地,可以是衍生自谷氨酸棒杆菌的具有seqidno:1的氨基酸序列的多肽,但是不限于此。此外,本公开内容中的lysc可以是由seqidno:1的氨基酸序列或与其具有80%、85%、90%、95%或者97%或更高的同源性或同一性的氨基酸序列组成的多肽。此外,显而易见的是,具有缺失、修饰、取代或添加序列的一部分的氨基酸序列的多肽也包括在本公开内容的范围内,只要该氨基酸序列具有这种同源性或同一性并展现出对应于多肽的同源性或同一性的效果。在本公开内容中,本领域众所周知的各种方法可用于获得天冬氨酸激酶(lysc)的方法。这种方法的实例包括基因合成技术,其包括优化密码子以便在棒状杆菌属的微生物中高效获得酶,该基因合成技术通常用于酶的表达,以及使用基于微生物的元基因组的生物信息学方法筛选有用酶资源的方法,但方法不限于此。如本文所使用,术语“变体”是这样的多肽,其中至少一个氨基酸与在保守取代和/或修饰中叙述的多肽不同,使得保留该多肽的功能或性质。变体多肽通过几个氨基酸的取代、缺失或添加与鉴定的序列不同。通常可以通过修饰上述多肽序列中的一个并评估修饰多肽的性质来鉴定这种变体。换句话说,相对于天然蛋白质,变体的能力可以增强、不变或减弱。通常可以通过修饰上述多肽序列中的一个并评估修饰多肽的反应性来鉴定这种变体。此外,一部分变体可以包括其中已除去一个或多个部分——比如,n-末端前导序列或跨膜结构域——的那些。其他变体可以包括其中一小部分已从成熟蛋白的n-末端和/或c-末端除去的变体。如本文所使用,“保守取代”是指一种氨基酸被具有相似结构和/或化学性质的另一种氨基酸取代。例如,变体可以具有保留一种或多种生物活性的至少一种保守取代。这种氨基酸取代通常可以基于残基的极性、电荷、溶解度、疏水性、亲水性和/或两亲性质的相似性来进行。例如,带正电荷的(碱性)氨基酸包括精氨酸、赖氨酸和组氨酸;带负电的(酸性)氨基酸包括天冬氨酸和谷氨酸;芳族氨基酸包括苯丙氨酸、色氨酸和酪氨酸;和疏水性氨基酸包括丙氨酸、缬氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、酪氨酸和色氨酸。典型地,保守取代对所得多肽的活性几乎没有影响。变体还可以包含其他修饰,包括缺失或添加对多肽的性质和二级结构具有最小影响的氨基酸。例如,多肽可以在蛋白质n-末端处与信号(或前导)序列缀合,该信号(或前导)序列共翻译或翻译后指导蛋白质的转移。多肽还可以与接头或其他序列缀合,以便于多肽的合成、纯化或鉴定。例如,本文要求保护的术语“天冬氨酸激酶变体”具体可以指衍生自棒状杆菌属的具有seqidno:1的氨基酸序列的lysc的修饰多肽,并且可以包括含有在seqidno:1的氨基酸序列中的一个或多个氨基酸取代的变体。具体地,它可以是包括用seqidno:1的氨基酸序列中的另一个氨基酸取代第377位的氨基酸残基的变体。“其他氨基酸”没有限制,只要它是l-亮氨酸——其为第377位的氨基酸——以外的氨基酸。具体地,例如,可以用赖氨酸或甲硫氨酸取代第377位的氨基酸残基。l-赖氨酸是碱性氨基酸的实例,并且碱性氨基酸可以是l-赖氨酸、l-精氨酸和l-组氨酸中的一种。l-甲硫氨酸是非极性氨基酸的实例,并且非极性氨基酸可以是l-甲硫氨酸、l-苯丙氨酸、l-丙氨酸、l-半胱氨酸、l-甘氨酸、l-异亮氨酸、l-亮氨酸、l-脯氨酸、l-色氨酸和l-缬氨酸中的一种。然而,这些不限于此。如本文所使用,术语“天冬氨酸激酶变体”可以与“修饰的天冬氨酸激酶”或“修饰的lysc”互换地使用。这种天冬氨酸激酶变体的特征在于,与具有seqidno:1的天冬氨酸激酶活性的多肽相比,具有增强的天冬氨酸激酶活性。此外,即使在本文中将其描述为“具有特定序列号的氨基酸序列的蛋白质”,显而易见的是,具有缺失、修饰、取代或添加序列的一部分的氨基酸序列的多肽也包括在本公开内容的范围内,只要该蛋白质具有与由相应序列号的氨基酸序列组成的肽同一或等同的效果。具体地,这种蛋白质不排除在相应序列号的氨基酸序列的前端或末端添加没有改变蛋白质的功能的序列、天然存在的突变、保守取代或其同义突变。此外,显而易见的是,具有这种序列添加或突变的蛋白质也包括在本公开内容的范围内。如本文所使用,术语“保守取代”是指用具有相似侧链的氨基酸残基取代某一氨基酸残基。具有相似侧链的氨基酸残基家族在本领域中是众所周知的。这种家族的实例可以包括具有碱性侧链的氨基酸(例如,赖氨酸、精氨酸、组氨酸)、具有酸性侧链的氨基酸(例如,天冬氨酸、谷氨酸)、具有不带电荷的极性侧链的氨基酸(例如,天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸)、具有非极性侧链的氨基酸(例如,甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸)、在β位具有分支侧链的氨基酸(例如,苏氨酸、缬氨酸、异亮氨酸)、具有芳族侧链的氨基酸(例如,酪氨酸、苯丙氨酸、色氨酸、组氨酸)、具有含羟基(例如,醇、酚醛)侧链的氨基酸(例如,丝氨酸、苏氨酸、酪氨酸)、和具有含硫侧链的氨基酸(例如,半胱氨酸、甲硫氨酸)。优选地,氨基酸的保守取代可以是天冬氨酸和谷氨酸之间的取代,精氨酸、赖氨酸和组氨酸之间的取代,色氨酸和苯丙氨酸之间的取代,苯丙氨酸和缬氨酸之间的取代,亮氨酸、异亮氨酸和丙氨酸之间的取代,和甘氨酸和丙氨酸之间的取代。如本文所使用,术语“同源性”或“同一性”是指两个给定氨基酸序列或核苷酸序列之间的相关程度,并且可以表示为百分比。术语“同源性”和“同一性”通常彼此互换使用。保守多核苷酸或多肽序列的序列同源性或同一性可以通过标准比对算法来确定,并且可以结合由所使用的程序建立的默认缺口罚分一起使用。通常预期基本上同源或相同的多核苷酸或多肽在适度或高度严格条件下与靶多核苷酸或多肽的全长度的至少约50%、约60%、约70%、约80%或约90%杂交。含有简并密码子而不是密码子的多核苷酸也被认为用于杂交。可以使用已知的计算机算法,比如使用默认参数的“fasta”程序(例如,pearsonetal.,(1988)proc.natl.acad.sci.usa85:2444)来确定任意两个多核苷酸或多肽序列是否彼此具有同源性、相似性或同一性。可选地,可以使用needleman-wunsch算法(needleman和wunsch,1970,j.mol.biol.48:443-453)来确定,该算法在emboss包的needleman程序中进行(emboss:theeuropeanmolecularbiologyopensoftwaresuite,riceetal.,2000,trendsgenet.16:276-277)(优选地为5.0.0版本或其后的版本)(gcg程序包(devereux,j.,etal.,nucleicacidsresearch12:387(1984)),blastp,blastn,fasta(atschul,[s.][f.,][etal.,jmolecbiol215]:403(1990);guidetohugecomputers,martinj.bishop,[ed.,]academicpress,sandiego,1994,and[carilloeta/.](1988)siamjappliedmath48:1073)。例如,可以使用国家生物技术信息中心(ncbi)的blast或clustalw来确定同源性或同一性。可以通过使用例如公布的gap计算机程序(例如,smith和waterman,adv.appl.math(1981)2:482)比较序列信息来确定多核苷酸或多肽序列的同源性、相似性或同一性。总之,gap程序将同源性或同一性定义为通过将相似比对的符号(即核苷酸或氨基酸)的数量除以两个序列中较短的符号的总数获得的值。gap程序的默认参数可以包括(1)如schwartz和dayhoff,eds.,atlasofproteinsequenceandstructure,nationalbiomedicalresearchfoundation,第353-358页,1979中所公开的gribskovetal.(1986),nucl.acidsres.14:6745中的一元比较矩阵(包含对于同一性值为1和对于非同一性值为0)和重量比较矩阵;(2)每个缺口罚分为3.0,并且每个缺口中的每个符号为另外的0.10罚分(或缺口开放罚分为10和缺口延伸惩罚为0.5);和(3)对于末端缺口不罚分。因此,如本文所使用,术语“同源性”或“同一性”是指序列之间的相关性。本公开内容的另一方面提供了编码天冬氨酸激酶变体的多核苷酸。天冬氨酸激酶变体如上所述。如本文所使用,术语“多核苷酸”是指由共价键合为链的核苷酸单体组成的核苷酸聚合物,并且其实例是具有预定或更长长度的dna或rna链。具体地,多核苷酸是指编码修饰多肽的多核苷酸片段。编码天冬氨酸激酶变体的多核苷酸可以是编码包括在seqidno:1的氨基酸序列中的一个或多个氨基酸取代的多肽的多核苷酸,其中氨基酸取代包括用另一个氨基酸取代第377位的氨基酸残基。具体地,作为代表性实例,编码天冬氨酸激酶变体的多核苷酸可以是编码多肽的多核苷酸,在该多肽中用在seqidno:1的氨基酸序列中的另一个氨基酸——l-赖氨酸或l-甲硫氨酸——取代第377位的氨基酸残基。更具体地,多核苷酸可以是编码lysc蛋白的多核苷酸,该lysc蛋白具有由seqidno:3或seqidno:5组成的氨基酸序列。例如,多核苷酸可以是由seqidno:4或seqidno:6的核苷酸序列组成的多核苷酸。在编码区中可以进行各种修饰,条件是它们不改变由编码区表达的蛋白质的氨基酸序列,这是由于密码子简并性或考虑到要表达蛋白质的生物体所优选的密码子。因此,显而易见的是,由于密码子简并性,由seqidno:3或seqidno:5的氨基酸序列组成的多肽或能够翻译成与其具有同源性或同一性的多核苷酸的多核苷酸也可以包括在本公开内容的范围内。可选地,可以包括通过在严格条件下由已知基因序列——例如,编码具有由seqidno:1的氨基酸序列组成的酶的活性的蛋白质的序列——与全部或部分多核苷酸序列的互补序列杂交制备的探针,并且没有限制。“严格条件”是指允许多核苷酸之间的特异性杂交的条件。在文献(例如,j.sambrook等)中具体公开了这些条件。例如,严格条件可以包括具有高同源性或同一性(例如,80%或更高、85%或更高、具体地90%或更高、更具体地95%或更高、甚至更具体地97%或者更高、甚至更具体地99%或更高)的基因可以在它们之间杂交,而具有其较低同源性或同一性的基因不能彼此杂交的条件;或常规的dna杂交条件(即,条件为在盐浓度和温度对应于60℃、1×ssc和0.1%sds;具体地在60℃下,0.1×ssc和0.1%sds;更具体地在68℃下,0.1×ssc和0.1%sds下漂洗一次,并且具体地两次或三次)。杂交需要两个核酸具有互补序列,尽管碱基之间的错配是可能的,这取决于杂交的严重性。术语“互补的”用于描述能够彼此杂交的核苷酸碱基之间的关系。例如,就dna而言,腺苷与胸腺嘧啶互补,和胞嘧啶与鸟嘌呤互补。因此,本公开内容还可以包括基本上相似的核酸序列以及与整个序列互补的分离的核酸片段。具体地,可以使用杂交条件检测具有同源性或同一性的多核苷酸,该杂交条件包括tm值为55℃,并且使用上述条件的杂交步骤。此外,tm值可以是60℃、63℃或65℃,但是不限于此。本领域普通技术人员可以根据其目的适当地调整tm值。杂交多核苷酸的适当严格性取决于多核苷酸的长度和互补程度,并且变量是本领域众所周知的(sambrook等,同上,9.50-9.51,11.7-11.8)。本公开内容的仍另一方面提供了包含天冬氨酸激酶变体的宿主细胞和用包含编码天冬氨酸激酶变体的多核苷酸的载体转化的微生物。具体地,本公开内容提供了生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的棒状杆菌属的微生物,其包括天冬氨酸激酶变体。包含本公开内容的天冬氨酸激酶变体的微生物的特征在于与野生型或未修饰的微生物相比,天冬氨酸激酶的活性增强。如本文所使用,术语“活性的增强”意思是引入蛋白质的活性,或者与微生物的内在活性或预修饰活性相比活性增强。活性的“引入”意思是微生物最初没有的特定蛋白质的活性被天然地或人工地表达。“内在活性”是指当通过天然或人工因素的遗传变异改变微生物性状时,在转化之前最初存在于亲本菌株中的特定蛋白质的活性。如本文所使用,术语“载体”是指包括编码靶蛋白的多核苷酸的核苷酸序列的dna构建体,该多核苷酸可操作地连接至适当的调节序列以在适当的宿主中表达靶蛋白。调节序列可以包括可启动转录的启动子、控制转录的任何操纵基因序列、编码适当mrna核糖体结合位点的序列以及控制转录和翻译终止的序列。载体可以转染到适当的宿主中,并且然后可以独立于宿主基因组复制或起作用,并且可以整合到基因组本身中。本公开内容中使用的载体没有特别限制,只要它可以在宿主中表达,并且可以使用本领域已知的任何载体。常用载体的实例是天然状态或重组状态的质粒、黏粒、病毒和噬菌体(bacteriophage)。例如,pwe15、m13、mbl3、mbl4、ixii、ashii、apii、t10、t11、charon4a和charon21a可以用作噬菌体(phage)载体或黏粒载体;和pbr系统、puc系统、pbluescriptii系统、pgem系统、ptz系统、pcl系统和pet系统可以用作质粒载体。具体地,可以使用pdz、pacyc177、pacyc184、pcl、peccg117、puc19、pbr322、pmw118和pcc1bac载体,但是这些不限于此。可以在本公开内容中使用的载体没有特别限制,并且可以使用已知的表达载体。此外,编码靶蛋白的多核苷酸可以通过用于细胞内染色体插入的载体插入染色体中。将多核苷酸插入染色体中可以通过本领域已知的任何方法比如同源重组进行,但不特别限于此。载体可以进一步包括用于指示载体插入宿主染色体中的选择标记。选择标记用于选择用载体转化的细胞,即,确认是否插入了靶核酸分子。因此,选择标记可赋予细胞显示抗药性、细胞毒性剂抗性、自养或可选择的表型表达比如表面蛋白的表达的能力。在选择剂存在下,可以选择转化细胞,因为只有表达选择标记的细胞表现存活或显示另一种表型。如本文所使用,术语“转化”是指以由多核苷酸编码的蛋白质在宿主细胞中表达的方式将包括编码靶蛋白的多核苷酸的载体引入宿主细胞中。只要转化的多核苷酸可以在宿主细胞中表达,它就可以被整合到宿主细胞的染色体中或置于宿主细胞的染色体中,或者存在于染色体外。此外,多核苷酸包括编码靶蛋白的dna和rna。多核苷酸可以以任何形式引入,只要它可以被引入宿主细胞中并在其中表达。例如,多核苷酸可以以表达盒的形式引入宿主细胞中,表达盒是包括其自主表达所需的所有元件的基因构建体。表达盒可以包括可操作地连接至多核苷酸的启动子、转录终止子、核糖体结合位点或翻译终止子。表达盒可以是可自我复制的表达载体的形式。此外,可以将多核苷酸原样引入宿主细胞中并可操作地连接至宿主细胞中的表达所需的序列,但是不限于此。转化方法包括将核酸引入细胞的任何方法,并且可以通过选择本领域已知的适当标准技术来进行,这取决于宿主细胞。方法的实例包括电穿孔、磷酸钙(capo4)沉淀、氯化钙(cacl2)沉淀、显微注射、聚乙二醇(peg)技术、deae-葡聚糖技术、阳离子脂质体技术、乙酸锂-dmso技术等,但是不限于此。此外,术语“可操作的连接”意思是多核苷酸序列与启动子序列功能性连接,该启动子序列启动和介导编码本公开内容的靶蛋白的多核苷酸的转录。可操作的连接可以使用本领域已知的基因重组技术制备,并且可以使用已知的裂解酶和连接酶制备位点特异性dna切割和连接,但是这些不限于此。如本文所使用,术语“生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的微生物”是指天然生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的微生物,或指其中将天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的生产力赋予缺乏生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的能力的亲本菌株的微生物。如本文所使用,术语“天冬氨酸衍生的l-氨基酸或其氨基酸衍生物”是指能够使用天冬氨酸(天冬氨酸盐)作为前体进行生物合成的材料,并且可以与术语“天冬氨酸衍生产物”互换使用。“氨基酸衍生物”是指可以由l-氨基酸产生的材料和包括l-氨基酸的前体的材料。此外,氨基酸衍生物没有限制,只要它是可以通过使用天冬氨酸作为前体的生物合成产生的材料。具体地,包括通常用乙酰磷酸(acetylphosphate)合成的材料,而没有限制。例如,氨基酸衍生物可以是l-赖氨酸、l-苏氨酸、l-甲硫氨酸、l-甘氨酸、高丝氨酸、o-乙酰高丝氨酸、o-琥珀酰高丝氨酸、o-磷酸高丝氨酸、l-异亮氨酸和尸胺,但是不限于此。可以使用天冬氨酸作为前体直接生物合成尸胺,或者可以通过赖氨酸脱羧酶从赖氨酸生产尸胺。可以使用天冬氨酸作为前体直接生物合成l-甲硫氨酸,或者可以通过从o-乙酰高丝氨酸或o-琥珀酰高丝氨酸转化来生产。如本文所使用,术语“天冬氨酸盐(天冬氨酸)”缩写为asp或d,并且是指天冬氨酸,其是用于蛋白质生物合成的α-氨基酸。与所有其他氨基酸一样,天冬氨酸包括氨基和羧酸基团。通常,天冬氨酸通过天冬氨酸激酶(lysc)产生为天冬氨酰磷酸,并且然后在细胞中转化为l-赖氨酸、l-甲硫氨酸、l-高丝氨酸、l-苏氨酸、l-异亮氨酸等。为了增强天冬氨酸衍生产物的生物合成,可以使用本公开内容的天冬氨酸激酶变体。例如,为了增强l-赖氨酸、l-苏氨酸、l-甲硫氨酸、l-甘氨酸、高丝氨酸、o-乙酰高丝氨酸、o-琥珀酰高丝氨酸、o-磷酸高丝氨酸、l-异亮氨酸和尸胺的生物合成,可以引入本公开内容的天冬氨酸激酶变体或可以增强其活性。此外,通过引入或增强特定蛋白质的活性或通过灭活特定蛋白质的活性,可以进一步提高天冬氨酸衍生产物的生产力。可以使用本领域已知的适当方法进行引入、增强和灭活特定蛋白质活性的方法,这取决于微生物的特征。生产天冬氨酸衍生产物、l-氨基酸或其氨基酸衍生物的微生物可以是通过包括本公开内容的天冬氨酸变体能够生产天冬氨酸衍生的“氨基酸”或其氨基酸衍生物的任何微生物。其具体实例可以包括埃希氏菌属、沙雷氏菌属、欧文氏菌属、肠道菌属、沙门氏菌属、链霉菌属、假单胞菌属、短杆菌属或棒杆菌属,并且更具体地,微生物可以是谷氨酸棒杆菌,但是不限于此。本公开内容的仍另一方面提供了用于生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的方法,其包括:培养上述微生物;和从培养的微生物或培养基中回收天冬氨酸衍生的l-氨基酸或其氨基酸衍生物。在本领域已知的优化培养条件和酶活性条件下,本领域普通技术人员可以容易地确定用于生产天冬氨酸衍生的l-氨基酸或其氨基酸衍生物的方法。具体地,尽管没有特别限制,但是培养微生物的步骤可以通过本领域已知的分批培养、连续培养和补料分批培养来进行。在此,培养条件没有特别限制,但是可以通过使用碱性化学品(例如,氢氧化钠、氢氧化钾或氨)或酸性化学品(例如,磷酸或硫酸)来维持最佳ph(例如,ph5至9,具体地ph6至8,和最具体地ph6.8)。此外,可以通过向细胞培养物中添加氧气或含氧气体混合物来维持需氧条件。培养温度可以维持在20℃至45℃,具体地在25℃至40℃,并且培养可以进行约10小时至160小时,但是这些不限于此。通过培养产生的天冬氨酸衍生的氨基酸或其氨基酸衍生物可以分泌到培养基中或者可以保留在细胞中。此外,用于培养的培养基可以包括单独地或组合地作为碳源的糖和碳水化合物(例如,葡萄糖、蔗糖、乳糖、果糖、麦芽糖、糖蜜、淀粉和纤维素)、油和脂肪(例如,大豆油、葵花籽油、花生油和椰子油)、脂肪酸(例如,棕榈酸、硬脂酸和亚油酸)、醇(例如,甘油和乙醇)和有机酸(例如,乙酸);单独地或组合地作为氮源的含氮有机化合物(例如,蛋白胨、酵母提取物、肉汁、麦芽汁、玉米溶液、大豆粕粉和尿素)或无机化合物(例如,硫酸铵、氯化铵、磷酸铵、碳酸铵和硝酸铵);和单独地或组合地作为磷源的磷酸二氢钾、磷酸氢二钾或其相应的含钠盐;但是这些不限于此。此外,培养基可以包含必需的生长促进物质,比如其他金属盐(例如,硫酸镁或硫酸铁)、氨基酸和维生素。根据培养方法,可以进行用于回收在本公开内容的培养方法中产生的天冬氨酸衍生的氨基酸或其氨基酸衍生物的方法,以通过使用本领域已知的适当方法从培养基收集靶氨基酸。例如,可以使用离心、过滤、阴离子交换色谱、结晶和hplc,并且可以通过使用本领域已知的适当方法从培养基或微生物中回收靶天冬氨酸衍生的氨基酸或其氨基酸衍生物。此外,回收方法可以进一步包括纯化步骤,并且可以使用本领域已知的适当方法来进行。本公开内容的仍另一方面提供了用于生产l-甲硫氨酸的方法,其包括:培养本公开内容的微生物;从培养的微生物或培养基生产o-乙酰高丝氨酸或o-琥珀酰高丝氨酸;和将o-乙酰高丝氨酸或o-琥珀酰高丝氨酸转化为l-甲硫氨酸。具体地,“转化为l-甲硫氨酸的步骤”可以包括在硫化物存在下使o-乙酰高丝氨酸或o-琥珀酰高丝氨酸与o-乙酰高丝氨酸硫化氢解酶或o-琥珀酰高丝氨酸硫化氢解酶反应的步骤。“o-乙酰高丝氨酸或o-琥珀酰高丝氨酸”可以是包含由本公开内容的微生物产生的o-乙酰高丝氨酸或o-琥珀酰高丝氨酸的发酵液,或者可以是纯化形式。此外,例如,“硫化物”可以是甲硫醇,并且甲硫醇可指甲硫醇钠(ch3s-na)的形式以及气体或液化状态的甲硫醇(ch3sh)和包含国际专利公开号wo2010/098629中描述的形式的二甲基硫(dms)的甲硫醇;并且甲硫醇还可指包含能够提供硫原子的形式的甲硫醇衍生物。此外,“o-乙酰高丝氨酸硫化氢解酶或o-琥珀酰高丝氨酸硫化氢解酶”可以是产生其的微生物的发酵液,或者可以是纯化形式。在本领域已知的优化培养条件和酶活性条件下,本领域普通技术人员可以容易地确定用于生产l-甲硫氨酸的方法。具体的培养方法和培养基如上所描述。有益效果与包括野生型lysc的微生物相比,根据本公开内容的包含天冬氨酸激酶变体的微生物可以以高生产力获得天冬氨酸衍生的l-氨基酸或天冬氨酸衍生产物,而不抑制宿主细胞的生长。附图说明图1显示了谷氨酸棒杆菌的天冬氨酸衍生的氨基酸和其氨基酸衍生物的生物合成途径。具体实施方式在下文中,将结合所附示例性实施方式详细地描述本公开内容。然而,本文公开的示例性实施方式仅用于说明目的,并且不应被解释为限制本公开内容的范围。实施例1:制备引入lysc变体的菌株通过天冬氨酸激酶的结构建模选择与野生型相比其活性可以增强的变体,并且如下所述制备引入该变体的菌株。在编码衍生自谷氨酸棒杆菌atcc13032(下文称为wt)的天冬氨酸激酶(seqidno:1)的基因lysc(seqidno:2)中,选择第377位的氨基酸作为变异位点,并选择碱性氨基酸l-赖氨酸和非极性氨基酸l-甲硫氨酸作为其他氨基酸的代表性实例进行取代。为了制备引入变异的载体,以该变异位点为中心,设计了用于扩增5′上游区域的一对引物(seqidno:7和8或seqidno:7和10)和用于扩增3′下游区域的一对引物(seqidno:9和12或seqidno:11和12)(表1)。在seqidno:7和12的引物中,在每个末端插入xbai限制酶位点(加下划线的)。此外,在seqidno:8和9的一对引物或seqidno:10和11的一对引物中,将核苷酸取代变异(加下划线的)置于设计为彼此交叉的位点处。表1使用wt的染色体作为模板,利用seqidno:7和8、seqidno:7和10、seqidno:9和12或seqidno:11和12的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合30秒。其后,在72℃下进行聚合反应,持续7分钟。结果,以基因lysc的变异位点为中心,分别获得5'上游区域的dna片段(509bp)和3'下游区域的dna片段(520bp)。使用两个扩增的dna片段作为模板,利用seqidno:7和12的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合60秒。其后,在72℃下进行聚合反应,持续7分钟。结果,包括编码天冬氨酸激酶变体(seqidno:3)——其中第377位的亮氨酸被赖氨酸取代——的基因lysc的变体(seqidno:4)的dna片段(1011bp)被扩增。此外,为了确认第377位氨基酸的重要性,获得包括编码天冬氨酸激酶变体(seqidno:5)——其中第377位的亮氨酸被甲硫氨酸取代——的基因lysc的变异(seqidno:6)的dna片段(1011bp)。用限制酶xbai处理不能在谷氨酸棒杆菌中复制的载体pdz(韩国专利号0924065)和dna片段(1011bp),并且用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为pdz-lysc(l377k)和pdz-lysc(l377m)。使用电脉冲方法(appl.microbiol.biotechnol.(1999)52:541-545)将载体pdz-lysc(l377k)和pdz-lysc(l377m)各自转化到wt中,并且然后在含有卡那霉素(25mg/l)的lb培养基中获得转化株。获得wt::lysc(l377k)和wt::lysc(l377m),其为通过经由二次重组过程(交叉)插入染色体上的dna片段将核苷酸变异引入基因lysc中的菌株,并且这些分别命名为谷氨酸棒杆菌ca01-2307和ca01-2308。ca01-2307和ca01-2308于2017年3月29日保藏于韩国微生物保藏中心,该中心是布达佩斯条约下的国际保藏机构,并且分配的登记号为kccm12000p和kccm12001p。实施例2:确认引入lysc变体的菌株用于生产天冬氨酸衍生的氨基酸的能力为了比较从实施例1获得的菌株ca01-2307和ca01-2308以及菌株wt用于产生主要天冬氨酸衍生的氨基酸的能力,使用下述方法培养菌株,并分析培养基中的组分。将各菌株接种到包含种子培养基(25ml)的角挡板烧瓶(250ml)中,并且在37℃下培养20小时,同时以200rpm振荡。将种子培养液(1ml)接种到包含生产培养基(24ml)的角挡板烧瓶(250ml)中,并且然后在37℃下培养24小时,同时以200rpm振荡。通过hplc分析l-赖氨酸和l-苏氨酸的浓度,l-赖氨酸和l-苏氨酸是衍生自l-天冬氨酸和天冬氨酸的代表性氨基酸,并将分析的浓度显示在表2中。<种子培养基(ph7.0)>葡萄糖(20g)、蛋白胨(10g)、酵母提取物(5g)、尿素(1.5g)、kh2po4(4g)、k2hpo4(8g)、mgso4·7h2o(0.5g)、生物素(100μg)、硫胺素hcl(1000μg)、泛酸钙(2000μg)、烟酰胺(2000μg)(在1l蒸馏水中)<生产培养基(ph7.0)>葡萄糖(100g)、(nh4)2so4(40g)、大豆蛋白(2.5g)、玉米浆(cornsteepsolid)(5g)、尿素(3g)、kh2po4(1g)、mgso4·7h2o(0.5g)、生物素(100μg)、硫胺素氯化物(1000μg)、泛酸钙(2000μg)、烟酰胺(3000μg)、caco3(30g)(在1l蒸馏水中)表2由ca01-2307和ca01-2308产生的天冬氨酸衍生的氨基酸的浓度作为分析天冬氨酸衍生的氨基酸的浓度的结果,确认当引入lysc变异时,与wt相比,l-赖氨酸的浓度显著增加,并且l-天冬氨酸和l-苏氨酸的浓度也增加。基于上述结果,以如上所述相同的方式培养菌株ca01-2307、ca01-2308和wt,以便详细比较其生产赖氨酸的能力。此外,以如上所述相同的方式分析培养基中l-赖氨酸的浓度(表3)。表3由菌株wt、ca01-2307和ca01-2308产生的l-赖氨酸的浓度作为分析l-赖氨酸的浓度的结果,与之前的评价一样,确认与由菌株wt产生的l-赖氨酸的生产力相比,由包括lysc(l377k)变异的ca01-2307产生的l-赖氨酸的生产力大大增加。此外,确认与由菌株wt产生的l-赖氨酸的生产力相比,由包括lysc(l377m)变异的ca01-2308产生的l-赖氨酸的生产力也增加。基于上述结果,确认天冬氨酸衍生的氨基酸中l-赖氨酸的生产力由于天冬氨酸激酶变体(seqidno:3)和由于天冬氨酸激酶变体(seqidno:5)大大增加,在天冬氨酸激酶变体(seqidno:3)中在本公开内容中选择的第377位的亮氨酸被赖氨酸取代,在天冬氨酸激酶变体(seqidno:5)中第377位的亮氨酸被甲硫氨酸取代。实施例3:制备l-苏氨酸增强菌株并确认l-苏氨酸的生产力为了通过引入lysc(l377k)变异——其具有实施例2中确认的较高的赖氨酸生产力——清楚地确认l-苏氨酸生产力的变化,将变异引入到编码生产高丝氨酸的高丝氨酸脱氢酶的基因中,该基因是l-苏氨酸、l-异亮氨酸、l-甲硫氨酸和高丝氨酸衍生物的生物合成途径中的常见中间体。具体地,通过将本领域已知的hom(g378e)变异(appl.microbiol.biotechnol.45,612-620(1996))引入到实施例1中制备的菌株ca01-2307中来制备菌株。此外,还制备了将hom(g378e)变异引入wt中的菌株作为对照组。为了制备引入hom(g378e)的载体,使用wt基因组dna作为模板,利用seqidno:13和14和seqidno:15和16进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合30秒。其后,在72℃下进行聚合反应,持续7分钟。结果,以基因hom的变异为中心,获得了5'上游区域的dna片段(220bp)和3'下游区域的dna片段(220bp)。使用这两种pcr产物作为模板,利用seqidno:13和16进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合30秒。其后,在72℃下进行聚合反应,持续7分钟。结果,扩增了包含基因hom变异的dna片段(440bp)。表4seqidno:引物序列(5′-3′)13引物7tcctctagactggtcgcctgatgttctac14引物8gccaaaacctccacgcgatc15引物9atcgcgtggaggttttggct16引物10gactctagattagtccctttcgaggcgga用限制酶xbai处理先前在实施例1中使用的载体pdz和dna片段(440bp),并且使用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为pdz-hom(g378e)。使用电脉冲方法(appl.microbiol.biotechnol.(1999,52:541-545))将载体pdz-hom(g378e)引入菌株wt和ca01-2307中,并且然后在含有卡那霉素(25mg/l)的选择性培养基中获得转化株。获得wt::hom(g378e)和ca01-2307::hom(g378e),其为通过经由二次重组过程(交叉)插入染色体上的dna片段将核苷酸变异引入基因hom的菌株。为了比较wt::hom(g378e)和ca01-2307::hom(g378e)用于生产苏氨酸的能力,以与实施例2中相同的方式培养菌株,并分析培养基中苏氨酸的浓度。表5由菌株wt::hom(g378e)和ca01-2307::hom(g378e)产生的苏氨酸的浓度作为分析l-苏氨酸的浓度的结果,确认在包括lysc变异的菌株中l-苏氨酸的生产力显著增加(表5)。实施例4:l-异亮氨酸增强菌株的制备和l-异亮氨酸生产力的比较为了确认lysc(l377k)变异的引入对l-异亮氨酸生产力的影响,制备了用于增强编码本领域已知的l-苏氨酸脱水酶的基因的ilva(v323a)变异(appl.enviro.microbiol.,dec.1996,p.4345-4351)的表达的载体。为了制备引入基因ilva的变异的载体,以变异位点为中心,设计了用于扩增5′上游区域的一对引物(seqidno:17和18)和用于扩增3′下游区域的一对引物(seqidno:19和20)。在seqidno:17和20的引物中,在每个末端插入bamhi限制酶位点(加下划线的)。此外,在seqidno:18和19的引物中,核苷酸取代变异(加下划线的)置于设计为彼此交叉的位点处。表6seqidno:引物序列(5′-3′)17引物11acggatcccagactccaaagcaaaagcg18引物12acaccacggcagaaccaggtgcaaaggaca19引物13ctggttctgccgtggtgtgcatcatctctg20引物14acggatccaaccaaacttgctcacactc使用wt的染色体作为模板,利用seqidno:17和18以及seqidno:19和20的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合30秒。其后,在72℃下进行聚合反应,持续7分钟。结果,以基因ilva的变异位点为中心,获得5'上游区域的dna片段(627bp)和3'下游区域的dna片段(608bp)。使用两个扩增的dna片段作为模板,利用seqidno:17和20的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合60秒。其后,在72℃下进行聚合反应,持续7分钟。结果,扩增了包括编码ilva变体的基因ilva的变异的dna片段(1217bp),在该ilva变体中第323位的缬氨酸被丙氨酸取代。用限制酶bamhi处理载体peccg117(韩国专利号10-0057684)和dna片段(1011bp),并且使用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为peccg117-ilva(v323a)。使用电脉冲方法将载体peccg117-ilva(v323a)引入在实施例3中制备的菌株wt::hom(g378e)和ca01-2307::hom(g378e),并且然后涂抹在含有卡那霉素(25mg/l)的选择性培养基上以获得转化株。以与实施例2中显示的烧瓶培养方法相同的方式培养菌株,并分析培养基中l-异亮氨酸的浓度(表7)。表7作为分析l-异亮氨酸的浓度的结果,确认在包括lysc变异的菌株中l-异亮氨酸的生产力大大增加。实施例5:o-乙酰高丝氨酸增强菌株的制备和o-乙酰高丝氨酸的生产力的比较为了确定lysc(l377k)变异的引入对o-乙酰高丝氨酸的生产的影响,缺失了编码参与o-乙酰高丝氨酸降解途径的胱硫醚γ-合酶的基因metb和编码o-乙酰高丝氨酸(巯基)-裂解酶的基因mety,并且然后过表达编码高丝氨酸o-乙酰转移酶——其为o-乙酰高丝氨酸生物合成酶——的基因metx,并且从而制备生产o-乙酰高丝氨酸的菌株。首先,为了缺失基因metb,基于该基因——衍生自wt的metb——的核苷酸序列信息,设计了用于扩增基因metb的5′上游区域的一对引物(seqidno:21和22)和用于扩增3′下游区域的一对引物(seqidno:23和24)。表8seqidno:引物序列(5′-3′)21引物15tctagatgcgctgattatctcacc22引物16actggtgggtcatggttgcatatgagatcaactcctgtaa23引物17ttacaggagttgatctcatatgcaaccatgacccaccagt24引物18tctagaccttgaagttcttgactg使用wt的染色体作为模板,利用seqidno:21和22、seqidno:23和24的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,在72℃下聚合90秒。其后,在72℃下进行聚合反应,持续7分钟。结果,获得了基因metb的5'上游区域的dna片段(450bp)和3'下游区域的dna片段(467bp)。使用两个扩增的dna片段作为模板,利用seqidno:21和24的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合3分钟。其后,在72℃下进行聚合反应,持续7分钟。结果,缺失了基因metb的中心区域,并且因此扩增了包括仅上游和下游末端的dna片段(917bp)。用限制酶xbai处理载体pdz和dna片段(917bp),并且使用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为pdz-δmetb。使用电脉冲方法将载体pdz-δmetb引入在实施例3中制备的菌株wt::hom(g378e)和ca01-2307::hom(g378e),并且然后从含有卡那霉素(25mg/l)的选择性培养基获得转化株。获得wt::hom(g378e)δmetb和ca01-2307::hom(g378e)δmetb,其为通过经由二次重组过程(交叉)插入染色体上的dna片段缺失metb基因的菌株。为了缺失参与o-乙酰高丝氨酸的另一降解途径的基因mety,基于该基因——衍生自wt的mety——的核苷酸序列信息,设计了用于扩增基因mety的5′上游区域的一对引物(seqidno:25和26)和用于扩增3′下游区域的一对引物(seqidno:27和28)。表9使用wt的染色体作为模板,利用seqidno:25和26、seqidno:27和28进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,在72℃下聚合90秒。其后,在72℃下进行聚合反应,持续7分钟。结果,获得了基因mety的5'上游区域的dna片段(512bp)和3'下游区域的dna片段(520bp)。使用两个扩增的dna片段作为模板,利用seqidno:25和28的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合3分钟。其后,在72℃下进行聚合反应,持续7分钟。结果,缺失了基因mety的中心区域,并且因此扩增了包括仅上游和下游末端的dna片段(1032bp)。用限制酶xbai处理载体pdz和dna片段(1032bp),并且使用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为pdz-δmety。使用电脉冲方法将载体pdz-δmety引入上述制备的菌株wt::hom(g378e)δmetb和ca01-2307::hom(g378e)δmetb的每一个中,并且然后在含有卡那霉素(25mg/l)的选择性培养基中获得转化株。获得wt::hom(g378e)δmetbδmety和ca01-2307::hom(g378e)δmetbδmety,其为通过经由二次重组过程(交叉)插入染色体上的dna片段缺失基因mety的菌株。为了使o-乙酰高丝氨酸的生产最大化,制备了用于增强基因metx的表达的载体。为了扩增编码高丝氨酸o-乙酰转移酶(metx)的基因,基于报道的wt衍生序列,通过在引物(seqidno:29和30)的两端插入bamhi限制酶区域来设计载体用于从启动子区(起始密码子的上游约300bp)扩增到终止子区(终止密码子的下游约100bp)。表10seqidno:引物序列(5′-3′)29引物23ggatcccctcgttgttcacccagcaacc30引物24ggatcccaaagtcacaactacttatgttag使用wt的染色体作为模板,利用seqidno:25和26的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合90秒。其后,在72℃下进行聚合反应,持续7分钟。结果,获得了包括基因metx的dna片段(1546bp)。用限制酶bamhi处理载体peccg117(韩国专利号10-0057684)和metx的dna片段,并且使用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为peccg117-metx。使用电脉冲方法将载体peccg117-metx引入上述制备的菌株wt::hom(g378e)δmetbδmety和ca01-2307::hom(g378e)δmetbδmety,并且然后涂抹在含有卡那霉素(25mg/l)的选择性培养基上以获得转化株。为了比较上述制备的菌株用于生产o-乙酰高丝氨酸(下文称为o-ah)的能力,使用下述方式培养菌株并分析培养基中o-乙酰高丝氨酸的浓度。将菌株的一个铂环(platinumloop)接种到包含o-ah生产培养基(25ml)的角挡板烧瓶(250ml)中,并在37℃下培养20小时,同时以200rpm振荡。通过hplc分析o-乙酰高丝氨酸的浓度,并将分析的浓度显示在表11中。<o-ah生产培养基(ph7.0)>葡萄糖(100g)、(nh4)2so4(40g)、大豆蛋白(2.5g)、玉米桨(5g)、尿素(3g)、kh2po4(1g)、mgso4·7h2o(0.5g)、生物素(100μg)、硫胺素氯化物(1000μg)、泛酸钙(2000μg)、烟酰胺(3000μg)、caco3(30g)、l-甲硫氨酸(0.3g)(在1l蒸馏水中)。表11作为分析上表所显示的o-乙酰高丝氨酸的浓度的结果,确认通过lysc变异生产的o-乙酰高丝氨酸的浓度增加。实施例6:制备o-琥珀酰高丝氨酸增强菌株和o-琥珀酰高丝氨酸的生产力的比较为了确定lysc(l377k)变异的引入对o-琥珀酰高丝氨酸的生产的影响,制备了生产o-琥珀酰高丝氨酸的菌株。由于野生型谷氨酸棒杆菌不能天然产生o-琥珀酰高丝氨酸,因此通过取代o-乙酰转移酶的底物结合区域的氨基酸,将实施例5中制备的菌株修饰为具有o-琥珀酰转移酶(metx)的活性,以便生产o-琥珀酰高丝氨酸。因此,基于基因——wt衍生的metx——的序列,设计了一对引物(seqidno:31和32)以制备用于引入metx变异的载体。表12seqidno:引物序列(5′-3′)31引物25tctagaatgcccaccctcgcgccttc32引物26tctagattagatgtagaactcgatg使用wt的染色体作为模板,利用seqidno:27和28的引物进行pcr。在95℃下变性5分钟后,在下述条件下进行总共30个循环的pcr:在95℃下变性30秒,在55℃下退火30秒,和在72℃下聚合90秒。其后,在72℃下进行聚合反应,持续7分钟。结果,获得了基因metx的编码区的dna片段(1146bp)。用限制酶xbai处理载体pdz和dna片段(1146bp),并且使用dna连接酶连接这些,并且然后克隆以获得质粒。质粒被命名为pdz-metx。基于载体pdz-metx,为了制备其中第176位的氨基酸被天冬酰胺取代和第313位的氨基酸被精氨酸取代的变异载体,设计了引起第176位氨基酸变异的一对引物(seqidno:33和34)和引起第313位氨基酸变异的一对引物(seqidno:35和36)。表13使用定点诱变试剂盒(stratagene,usa)与上述每种引物一起制备变体基因metx。将基于现有野生型质粒pdz-metx的转化株质粒命名为pdz-metx(q176n,l313r)。使用电脉冲方法将上述制备的载体pdz-metx(q176n,l313r)转化入实施例5中制备的菌株wt::hom(g378e)δmetbδmety和ca01-2307::hom(g378e)δmetbδmety,并且然后在含有卡那霉素(25mg/l)的选择性培养基中获得转化株。获得wt::hom(g378e)metx(q176n、l313r)δmetbδmety和ca01-2307::hom(g378e)metx(q176n,l313r)δmetbδmety,其为通过经由二次重组过程(交叉)插入染色体上的dna片段将基因metx替换为metx(q176n,l313r)的菌株。为了最大化o-琥珀酰高丝氨酸的生产,制备了用于增强变体基因metx(q176n,l313r)的表达的载体。基于实施例5中制备的peccg117-metx,使用seqidno:33至36的引物制备过表达变体metx的载体,其中第176位的变体氨基酸——其具有o-琥珀酰转移酶的活性——被天冬酰胺取代,和第313位氨基酸被精氨酸取代。具体地,使用定点诱变试剂盒(stratagene,usa)与每种引物一起制备载体,并将制备的载体命名为peccg117-metx(q176n,l313r)。使用电脉冲方法将peccg117-metx(q176n,l313r)和空载体peccg117引入wt::hom(g378e)metx(q176n、l313r)δmetbδmety和ca01-2307::hom(g378e)metx(q176n,l313r)δmetbδmety中,它们是上述制备的生产o-琥珀酰高丝氨酸的菌株,并涂抹在含有卡那霉素(25mg/l)的选择性培养基上以获得转化株。为了比较菌株用于生产o-琥珀酰高丝氨酸的能力,使用下述方法培养菌株并分析培养基中o-琥珀酰高丝氨酸的浓度。将菌株的一个铂环接种到包含25ml实施例5中使用的相同o-ah生产培养基组合物的角挡板烧瓶(250ml)中,并且在37℃下培养20小时,同时以200rpm振荡。通过hplc分析o-琥珀酰高丝氨酸的浓度,并将分析的浓度显示在表14中。表14作为分析如上表中显示的o-琥珀酰高丝氨酸的浓度的结果,确认通过lysc变异产生的o-琥珀酰高丝氨酸的产量增加。这些结果表明,本公开内容的变体可以增加天冬氨酸衍生的氨基酸和/或其衍生物的产量。由前述本公开所属领域的普通技术人员将能够理解,本公开内容可以以其他特定形式体现,而无需修改本公开内容的技术概念或基本特征。在这方面,本文公开的示例性实施方式仅用于说明目的,并且不应解释为限制本公开内容的范围。相反,本公开内容旨在不仅涵盖示例性实施方式,而且涵盖包括在由所附权利要求限定的本公开内容的精神和范围内的各种替换、修改、等同物和其他实施方式。<110>cj第一制糖株式会社<120>新型天冬氨酸激酶变体和使用其生产l-氨基酸的方法<130>opa18147<150>10-2017-0083437<151>2017-06-30<160>36<170>kopatentin3.0<210>1<211>421<212>prt<213>谷氨酸棒状杆菌<400>1valalaleuvalvalglnlystyrglyglyserserleugluserala151015gluargileargasnvalalagluargilevalalathrlyslysala202530glyasnaspvalvalvalvalcysseralametglyaspthrthrasp354045gluleuleugluleualaalaalavalasnprovalproproalaarg505560glumetaspmetleuleuthralaglygluargileserasnalaleu65707580valalametalailegluserleuglyalaglualaglnserphethr859095glyserglnalaglyvalleuthrthrgluarghisglyasnalaarg100105110ilevalaspvalthrproglyargvalargglualaleuaspglugly115120125lysilecysilevalalaglypheglnglyvalasnlysgluthrarg130135140aspvalthrthrleuglyargglyglyseraspthrthralavalala145150155160leualaalaalaleuasnalaaspvalcysgluiletyrseraspval165170175aspglyvaltyrthralaaspproargilevalproasnalaglnlys180185190leuglulysleuserphegluglumetleugluleualaalavalgly195200205serlysileleuvalleuargservalglutyralaargalapheasn210215220valproleuargvalargsersertyrserasnaspproglythrleu225230235240ilealaglysermetgluaspileprovalgluglualavalleuthr245250255glyvalalathrasplysserglualalysvalthrvalleuglyile260265270serasplysproglyglualaalalysvalpheargalaleualaasp275280285alagluileasnileaspmetvalleuglnasnvalserservalglu290295300aspglythrthraspilethrphethrcysproargseraspglyarg305310315320argalametgluileleulyslysleuglnvalglnglyasntrpthr325330335asnvalleutyraspaspglnvalglylysvalserleuvalglyala340345350glymetlysserhisproglyvalthralagluphemetglualaleu355360365argaspvalasnvalasnilegluleuileserthrsergluilearg370375380ileservalleuilearggluaspaspleuaspalaalaalaargala385390395400leuhisgluglnpheglnleuglyglygluaspglualavalvaltyr405410415alaglythrglyarg420<210>2<211>1266<212>dna<213>谷氨酸棒状杆菌<400>2gtggccctggtcgtacagaaatatggcggttcctcgcttgagagtgcggaacgcattaga60aacgtcgctgaacggatcgttgccaccaagaaggctggaaatgatgtcgtggttgtctgc120tccgcaatgggagacaccacggatgaacttctagaacttgcagcggcagtgaatcccgtt180ccgccagctcgtgaaatggatatgctcctgactgctggtgagcgtatttctaacgctctc240gtcgccatggctattgagtcccttggcgcagaagcccaatctttcacgggctctcaggct300ggtgtgctcaccaccgagcgccacggaaacgcacgcattgttgatgtcactccaggtcgt360gtgcgtgaagcactcgatgagggcaagatctgcattgttgctggtttccagggtgttaat420aaagaaacccgcgatgtcaccacgttgggtcgtggtggttctgacaccactgcagttgcg480ttggcagctgctttgaacgctgatgtgtgtgagatttactcggacgttgacggtgtgtat540accgctgacccgcgcatcgttcctaatgcacagaagctggaaaagctcagcttcgaagaa600atgctggaacttgctgctgttggctccaagattttggtgctgcgcagtgttgaatacgct660cgtgcattcaatgtgccacttcgcgtacgctcgtcttatagtaatgatcccggcactttg720attgccggctctatggaggatattcctgtggaagaagcagtccttaccggtgtcgcaacc780gacaagtccgaagccaaagtaaccgttctgggtatttccgataagccaggcgaggctgcg840aaggttttccgtgcgttggctgatgcagaaatcaacattgacatggttctgcagaacgtc900tcttctgtagaagacggcaccaccgacatcaccttcacctgccctcgttccgacggccgc960cgcgcgatggagatcttgaagaagcttcaggttcagggcaactggaccaatgtgctttac1020gacgaccaggtcggcaaagtctccctcgtgggtgctggcatgaagtctcacccaggtgtt1080accgcagagttcatggaagctctgcgcgatgtcaacgtgaacatcgaattgatttccacc1140tctgagattcgtatttccgtgctgatccgtgaagatgatctggatgctgctgcacgtgca1200ttgcatgagcagttccagctgggcggcgaagacgaagccgtcgtttatgcaggcaccgga1260cgctaa1266<210>3<211>421<212>prt<213>谷氨酸棒状杆菌<400>3valalaleuvalvalglnlystyrglyglyserserleugluserala151015gluargileargasnvalalagluargilevalalathrlyslysala202530glyasnaspvalvalvalvalcysseralametglyaspthrthrasp354045gluleuleugluleualaalaalavalasnprovalproproalaarg505560glumetaspmetleuleuthralaglygluargileserasnalaleu65707580valalametalailegluserleuglyalaglualaglnserphethr859095glyserglnalaglyvalleuthrthrgluarghisglyasnalaarg100105110ilevalaspvalthrproglyargvalargglualaleuaspglugly115120125lysilecysilevalalaglypheglnglyvalasnlysgluthrarg130135140aspvalthrthrleuglyargglyglyseraspthrthralavalala145150155160leualaalaalaleuasnalaaspvalcysgluiletyrseraspval165170175aspglyvaltyrthralaaspproargilevalproasnalaglnlys180185190leuglulysleuserphegluglumetleugluleualaalavalgly195200205serlysileleuvalleuargservalglutyralaargalapheasn210215220valproleuargvalargsersertyrserasnaspproglythrleu225230235240ilealaglysermetgluaspileprovalgluglualavalleuthr245250255glyvalalathrasplysserglualalysvalthrvalleuglyile260265270serasplysproglyglualaalalysvalpheargalaleualaasp275280285alagluileasnileaspmetvalleuglnasnvalserservalglu290295300aspglythrthraspilethrphethrcysproargseraspglyarg305310315320argalametgluileleulyslysleuglnvalglnglyasntrpthr325330335asnvalleutyraspaspglnvalglylysvalserleuvalglyala340345350glymetlysserhisproglyvalthralagluphemetglualaleu355360365argaspvalasnvalasnileglulysileserthrsergluilearg370375380ileservalleuilearggluaspaspleuaspalaalaalaargala385390395400leuhisgluglnpheglnleuglyglygluaspglualavalvaltyr405410415alaglythrglyarg420<210>4<211>1266<212>dna<213>谷氨酸棒状杆菌<400>4gtggccctggtcgtacagaaatatggcggttcctcgcttgagagtgcggaacgcattaga60aacgtcgctgaacggatcgttgccaccaagaaggctggaaatgatgtcgtggttgtctgc120tccgcaatgggagacaccacggatgaacttctagaacttgcagcggcagtgaatcccgtt180ccgccagctcgtgaaatggatatgctcctgactgctggtgagcgtatttctaacgctctc240gtcgccatggctattgagtcccttggcgcagaagcccaatctttcacgggctctcaggct300ggtgtgctcaccaccgagcgccacggaaacgcacgcattgttgatgtcactccaggtcgt360gtgcgtgaagcactcgatgagggcaagatctgcattgttgctggtttccagggtgttaat420aaagaaacccgcgatgtcaccacgttgggtcgtggtggttctgacaccactgcagttgcg480ttggcagctgctttgaacgctgatgtgtgtgagatttactcggacgttgacggtgtgtat540accgctgacccgcgcatcgttcctaatgcacagaagctggaaaagctcagcttcgaagaa600atgctggaacttgctgctgttggctccaagattttggtgctgcgcagtgttgaatacgct660cgtgcattcaatgtgccacttcgcgtacgctcgtcttatagtaatgatcccggcactttg720attgccggctctatggaggatattcctgtggaagaagcagtccttaccggtgtcgcaacc780gacaagtccgaagccaaagtaaccgttctgggtatttccgataagccaggcgaggctgcg840aaggttttccgtgcgttggctgatgcagaaatcaacattgacatggttctgcagaacgtc900tcttctgtagaagacggcaccaccgacatcaccttcacctgccctcgttccgacggccgc960cgcgcgatggagatcttgaagaagcttcaggttcagggcaactggaccaatgtgctttac1020gacgaccaggtcggcaaagtctccctcgtgggtgctggcatgaagtctcacccaggtgtt1080accgcagagttcatggaagctctgcgcgatgtcaacgtgaacatcgaaaagatttccacc1140tctgagattcgtatttccgtgctgatccgtgaagatgatctggatgctgctgcacgtgca1200ttgcatgagcagttccagctgggcggcgaagacgaagccgtcgtttatgcaggcaccgga1260cgctaa1266<210>5<211>421<212>prt<213>谷氨酸棒状杆菌<400>5valalaleuvalvalglnlystyrglyglyserserleugluserala151015gluargileargasnvalalagluargilevalalathrlyslysala202530glyasnaspvalvalvalvalcysseralametglyaspthrthrasp354045gluleuleugluleualaalaalavalasnprovalproproalaarg505560glumetaspmetleuleuthralaglygluargileserasnalaleu65707580valalametalailegluserleuglyalaglualaglnserphethr859095glyserglnalaglyvalleuthrthrgluarghisglyasnalaarg100105110ilevalaspvalthrproglyargvalargglualaleuaspglugly115120125lysilecysilevalalaglypheglnglyvalasnlysgluthrarg130135140aspvalthrthrleuglyargglyglyseraspthrthralavalala145150155160leualaalaalaleuasnalaaspvalcysgluiletyrseraspval165170175aspglyvaltyrthralaaspproargilevalproasnalaglnlys180185190leuglulysleuserphegluglumetleugluleualaalavalgly195200205serlysileleuvalleuargservalglutyralaargalapheasn210215220valproleuargvalargsersertyrserasnaspproglythrleu225230235240ilealaglysermetgluaspileprovalgluglualavalleuthr245250255glyvalalathrasplysserglualalysvalthrvalleuglyile260265270serasplysproglyglualaalalysvalpheargalaleualaasp275280285alagluileasnileaspmetvalleuglnasnvalserservalglu290295300aspglythrthraspilethrphethrcysproargseraspglyarg305310315320argalametgluileleulyslysleuglnvalglnglyasntrpthr325330335asnvalleutyraspaspglnvalglylysvalserleuvalglyala340345350glymetlysserhisproglyvalthralagluphemetglualaleu355360365argaspvalasnvalasnileglumetileserthrsergluilearg370375380ileservalleuilearggluaspaspleuaspalaalaalaargala385390395400leuhisgluglnpheglnleuglyglygluaspglualavalvaltyr405410415alaglythrglyarg420<210>6<211>1266<212>dna<213>谷氨酸棒状杆菌<400>6gtggccctggtcgtacagaaatatggcggttcctcgcttgagagtgcggaacgcattaga60aacgtcgctgaacggatcgttgccaccaagaaggctggaaatgatgtcgtggttgtctgc120tccgcaatgggagacaccacggatgaacttctagaacttgcagcggcagtgaatcccgtt180ccgccagctcgtgaaatggatatgctcctgactgctggtgagcgtatttctaacgctctc240gtcgccatggctattgagtcccttggcgcagaagcccaatctttcacgggctctcaggct300ggtgtgctcaccaccgagcgccacggaaacgcacgcattgttgatgtcactccaggtcgt360gtgcgtgaagcactcgatgagggcaagatctgcattgttgctggtttccagggtgttaat420aaagaaacccgcgatgtcaccacgttgggtcgtggtggttctgacaccactgcagttgcg480ttggcagctgctttgaacgctgatgtgtgtgagatttactcggacgttgacggtgtgtat540accgctgacccgcgcatcgttcctaatgcacagaagctggaaaagctcagcttcgaagaa600atgctggaacttgctgctgttggctccaagattttggtgctgcgcagtgttgaatacgct660cgtgcattcaatgtgccacttcgcgtacgctcgtcttatagtaatgatcccggcactttg720attgccggctctatggaggatattcctgtggaagaagcagtccttaccggtgtcgcaacc780gacaagtccgaagccaaagtaaccgttctgggtatttccgataagccaggcgaggctgcg840aaggttttccgtgcgttggctgatgcagaaatcaacattgacatggttctgcagaacgtc900tcttctgtagaagacggcaccaccgacatcaccttcacctgccctcgttccgacggccgc960cgcgcgatggagatcttgaagaagcttcaggttcagggcaactggaccaatgtgctttac1020gacgaccaggtcggcaaagtctccctcgtgggtgctggcatgaagtctcacccaggtgtt1080accgcagagttcatggaagctctgcgcgatgtcaacgtgaacatcgaaatgatttccacc1140tctgagattcgtatttccgtgctgatccgtgaagatgatctggatgctgctgcacgtgca1200ttgcatgagcagttccagctgggcggcgaagacgaagccgtcgtttatgcaggcaccgga1260cgctaa1266<210>7<211>29<212>dna<213>人工序列<220><223>引物1<400>7tcctctagagctgcgcagtgttgaatacg29<210>8<211>30<212>dna<213>人工序列<220><223>引物2<400>8tggaaatcttttcgatgttcacgttgacat30<210>9<211>30<212>dna<213>人工序列<220><223>引物3<400>9acatcgaaaagatttccacctctgagattc30<210>10<211>30<212>dna<213>人工序列<220><223>引物4<400>10tggaaatcatttcgatgttcacgttgacat30<210>11<211>30<212>dna<213>人工序列<220><223>引物5<400>11acatcgaaatgatttccacctctgagattc30<210>12<211>29<212>dna<213>人工序列<220><223>引物6<400>12gactctagagttcacctcagagacgatta29<210>13<211>29<212>dna<213>人工序列<220><223>引物7<400>13tcctctagactggtcgcctgatgttctac29<210>14<211>20<212>dna<213>人工序列<220><223>引物8<400>14gccaaaacctccacgcgatc20<210>15<211>20<212>dna<213>人工序列<220><223>引物9<400>15atcgcgtggaggttttggct20<210>16<211>29<212>dna<213>人工序列<220><223>引物10<400>16gactctagattagtccctttcgaggcgga29<210>17<211>28<212>dna<213>人工序列<220><223>引物11<400>17acggatcccagactccaaagcaaaagcg28<210>18<211>30<212>dna<213>人工序列<220><223>引物12<400>18acaccacggcagaaccaggtgcaaaggaca30<210>19<211>30<212>dna<213>人工序列<220><223>引物13<400>19ctggttctgccgtggtgtgcatcatctctg30<210>20<211>28<212>dna<213>人工序列<220><223>引物14<400>20acggatccaaccaaacttgctcacactc28<210>21<211>24<212>dna<213>人工序列<220><223>引物15<400>21tctagatgcgctgattatctcacc24<210>22<211>40<212>dna<213>人工序列<220><223>引物16<400>22actggtgggtcatggttgcatatgagatcaactcctgtaa40<210>23<211>40<212>dna<213>人工序列<220><223>引物17<400>23ttacaggagttgatctcatatgcaaccatgacccaccagt40<210>24<211>24<212>dna<213>人工序列<220><223>引物18<400>24tctagaccttgaagttcttgactg24<210>25<211>24<212>dna<213>人工序列<220><223>引物19<400>25tctagaagtagcgttgctgtacac24<210>26<211>43<212>dna<213>人工序列<220><223>引物20<400>26atcaatggtctcgatgcccatatggcatttggaggtccttaag43<210>27<211>43<212>dna<213>人工序列<220><223>引物21<400>27cttaaggacctccaaatgccatatgggcatcgagaccattgat43<210>28<211>24<212>dna<213>人工序列<220><223>引物22<400>28tctagatggaaccgttgcaaccac24<210>29<211>28<212>dna<213>人工序列<220><223>引物23<400>29ggatcccctcgttgttcacccagcaacc28<210>30<211>30<212>dna<213>人工序列<220><223>引物24<400>30ggatcccaaagtcacaactacttatgttag30<210>31<211>26<212>dna<213>人工序列<220><223>引物25<400>31tctagaatgcccaccctcgcgccttc26<210>32<211>25<212>dna<213>人工序列<220><223>引物26<400>32tctagattagatgtagaactcgatg25<210>33<211>35<212>dna<213>人工序列<220><223>引物27<400>33acgcgccagcgcctggaacatcggcattcaatccg35<210>34<211>35<212>dna<213>人工序列<220><223>引物28<400>34cggattgaatgccgatgttccaggcgctggcgcgt35<210>35<211>33<212>dna<213>人工序列<220><223>引物29<400>35gtagataccgatattcggtacccctaccaccag33<210>36<211>33<212>dna<213>人工序列<220><223>引物30<400>36ctggtggtaggggtaccgaatatcggtatctac33当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1