抗体结构域的优选配对的制作方法
本发明涉及抗原结合分子(abm),其包含人抗体结构域序列,特别是包含由vl和cl抗体结构域组成的抗体轻链的同二聚体,与至少包含vh和ch1抗体结构域的抗体重链缔合,该缔合通过配对vl和vh结构域以及cl和ch1结构域实现,其中优选的配对由cl和ch1结构域中的某些点突变支持。
背景技术:
单克隆抗体已被广泛用作治疗性抗原结合分子。这里将使用完整的igg1免疫球蛋白作为实例来解释基本的抗体结构。
两条相同的重(h)链和两条相同的轻(l)链结合,形成y形抗体分子。重链各有四个结构域。氨基端可变结构域(vh)位于y的末端。其后是三个恒定结构域:ch1、ch2和位于y茎底部的羧基端ch3。短拉伸、开关连接重链变量区和恒定区。铰链将ch2和ch3(fc片段)连接至抗体的剩余部分(fab片段)。可通过在完整抗体分子中蛋白水解切割铰链来产生一个fc和两个相同的fab片段。轻链由可变(vl)和恒定(cl)两个结构域构造,由开关分开。
铰链区中的二硫键连接两条重链。轻链通过另外的二硫键与重链偶联。根据免疫球蛋白的类别,asn连接的碳水化合物部分附接在恒定结构域中的不同位置。对于igg1,铰链区中的在cys235和cys238对之间两个二硫键将两条重链合并在一起。轻链通过ch1结构域中的cys220(eu索引编号)或cys233(根据kabat编号)与cl结构域中的cys214(eu索引和kabat编号)之间的两个另外的二硫键与重链偶联。碳水化合物部分与每个ch2的asn306附接,在y的茎中产生明显的凸起。
这些特征具有深远的功能性结果。重链和轻链(vh)和(vl)的可变区位于n-端区域,即y的“末端”,其中它们被定位以与抗原反应。分子的该末端是氨基酸序列的n-端所在的一侧。y的茎以有效介导效应子功能的方式投射,例如补体的激活和与fc受体或adcc和adcp的相互作用。其ch2和ch3结构域凸起以促进与效应蛋白的相互作用。氨基酸序列的c端位于末端的可被称为y的“底部”的相对侧。
在抗体中发现了两种类型的轻链,称为兰姆达(λ)和卡帕(κ)。给定的免疫球蛋白具有κ链或λ链,而不是每种一个。在具有λ或κ轻链的抗体之间未发现功能差异。
抗体分子中的每个结构域具有在压缩的反平行β桶中彼此紧密堆积的两个β片(sheet)的相似结构。这种保守结构称为免疫球蛋白折叠。恒定结构域的免疫球蛋白折叠含有针对4-链片堆积的3-链片。通过每个片的β链之间的氢键,通过内部中相对片的残基之间的疏水键合,以及通过片之间的二硫键来稳定折叠。3-链片包含链c、f和g,并且4-链片具有链a、b、e和d。字母a至g表示β链沿着免疫球蛋白折叠的氨基酸序列的连续位置。
可变结构域的折叠具有排列在4和5链的两片中的9个β链。5-链片与恒定结构域的3-链片在结构上同源,但含有额外的链c’和c”。链(a、b、c、d、e、f、g)的剩余部分具有与其恒定结构域免疫球蛋白折叠中的对应物相同的拓扑结构和相似的结构。二硫键将相反的片中的链b和f连接起来,如在恒定结构域中那样。
轻和重免疫球蛋白链的可变结构域含有三个高变环或互补决定区(cdr)。v结构域的三个cdr(cdr1、cdr2、cdr3)聚集在β桶的一端。cdr是连接免疫球蛋白折叠的β链b-c、c’-c”和f-g的环。cdr中的残基从一个免疫球蛋白分子到下一个免疫球蛋白分子不同,赋予每个抗体抗原特异性。
抗体分子末端的vl和vh结构域紧密堆积,使得6个cdr(每个结构域上3个)配合构建用于抗原特异性结合的表面(或空腔)。因此,抗体的天然抗原结合位点由连接轻链可变结构域的链b-c、c’-c”和f-g以及重链可变结构域的链b-c、c’-c”和f-g的环组成。
不是天然免疫球蛋白中的cdr环或不是由cdr环和cdr环区域中任选相邻的环确定的抗原结合袋的一部分的环,不具有抗原结合或表位结合特异性,但有助于整个免疫球蛋白分子和/或其效应子或其他功能的正确折叠,因此称为结构环。
现有技术文献表明,迄今为止,为了操纵现有的抗原结合位点,已采用免疫球蛋白样支架,从而引入了新的结合特性。在大多数情况下,cdr区经工程化用于抗原结合,换言之,在免疫球蛋白折叠的情况下,仅修饰天然抗原结合位点以改变其结合亲和力或特异性。存在大量文献,其描述了这种被操纵的免疫球蛋白的不同形式,通常以单链fv片段(scfv)或fab片段的形式表达,或者在噬菌体颗粒的表面上展示或者在各种原核或真核表达系统中可溶性表达。
目前正在开发用于识别两种不同靶标的改进治疗方法的抗体构建体。
davis等人(蛋白质工程(proteinengineering),design&selection2010,23(4)195-202)描述了异二聚体fc平台,该异二聚体fc平台通过使用链交换工程化结构域(seed)ch3异二聚体来支持双特异性和不对称融合蛋白的设计。这些人igg和igach3结构域的衍生物产生互补的人seedch3异二聚体,该异二聚体由人iga和igg序列的交替区段组成。在wo2007/110205a2和ep1999154b1中进一步描述了seed工程化。wo2010/136172a1公开了三或四特异性抗体,该抗体包含一个或两个与抗体的fc部分的c端连接的单链fab。
beck等人(naturereviewsimmunology,第10卷,第5号,2010年5月1日,第345-352页)描述了下一代治疗性抗体,并且特别是指不同类型的双特异性抗体。
ridgway等人,(proteinengineering,第9卷,第7号,1996,第617-621页)描述了用于重链异二聚化的抗体ch3结构域的“隆突-入-穴(knobsinto-holes)”工程化。
vonkreudenstein等人,(landesbioscience,第5卷,第5号,2013,第646-654页)描述了基于被工程化以用于稳定性的异二聚体fc的双特异性抗体支架。
liu等人,(journalofbiologicalchemistry2015,290:7535-7562)描述了通过静电机制制备单价双特异性异二聚体igg抗体的策略。通过瞬时和稳定转染的哺乳动物细胞产生衍生自具有正确轻链(lc)和重链(hc)配对的抗her2和抗egfr抗体的异二聚体igg分子。通过在vh-vl和ch1-cl界面处切换极性或疏水性残基来驱动lc和hc的特异性配对。每种经工程化的变体的特征在于一系列的点突变,其中在vh和vl结构域中。另外,点突变在ch1结构域(例如k147d)和在cl(cκ或ck)结构域(例如t180k)中被工程化(根据eu指数编号)。一些变体尤其包含ch1结构域中147位和cκ结构域中180或131位的点突变。
wo2014/081955进一步公开了这样的异二聚体抗体,该异二聚体抗体在以下结构域中的每一个中包含一个或多个取代:第一和第二ch3结构域、ch1结构域、cl结构域、vh结构域和vl结构域。
lewis等人,(naturebiotechnology2014,32:191-198)描述了通过基于结构的正交fab界面设计产生双特异性igg抗体。产生具有改进的hc-lc配对的双特异性igg。发现可变结构域支配重链和轻链的特异性组装。两种不同的设计在vh、vl、ch1和cl结构域的每一个中采用点突变。其中一个设计尤其包含ch1结构域146位和cλ结构域129位的点突变(根据kabat编号)。
dillon等人,(mabs2016;doi:10.1080/19420862.2016.1267089)描述了单个哺乳动物细胞中不同同种型和起源物种的双特异性igg的产生,以及促进与先前描述的用于优选重链异二聚化的隆突-入-穴突变结合的选择性fab臂组装的设计。
上述设计的双特异性抗体必然结合一系列点突变以稳定igg结构,包括位于vh和vl结构域中的主要点突变。期望工程化双特异性抗体,其中正确的配对已经仅由ch1和cl结构域中的点突变支持。
技术实现要素:
本发明的目的是提供抗体重链和轻链的改进配对,其支持hc和lc的正确配对,同时保持vh和vl结构域的框架不变。这种改进的配对将有利于双特异性抗体的产生。
所述目的通过本发明的主题解决。
根据本发明,提供了一抗原结合分子(abm),该抗原结合分子包含抗体轻链(lc)的同源lc/hc二聚体,该抗体轻链由vl和cl抗体结构域组成,与包含至少vh和ch1抗体结构域的抗体重链(hc)缔合,该缔合通过配对vl和vh结构域以及cl和ch1结构域实现,其中cl结构域中的位置18处和ch1结构域中的位置26处的氨基酸具有相反的极性,其中根据imgt编号。
具体地,所述同源lc/hc二聚体的特征在于同源结构域,其配对以形成同源(结构域)对。具体理解的是,lc/hc二聚体是同源的,因为单体cl和ch1结构域是同源的或匹配的对应物,优选彼此识别以产生相比于野生型结构域的一对cl和ch1结构域。具体地,如本文所述的cl结构域优选地与同源ch1结构域配对;并且如本文所述的ch1结构域优选地与同源cl结构域配对。
根据一具体方面,abm的特征在于同源cl和ch1抗体结构域,该同源cl和ch1抗体结构域优选地通过吸引力彼此配对,并且由于排斥力,优选地不与非同源的或野生型的其他对应结构域配对。因此,大大降低了作为野生型抗体结构域的对应抗体结构域的错配对,或通过各自的点突变使其成为非同源(排斥以降低组装的可能性)的错配对。
具体地,同源cl和ch1结构域的特征在于在所述氨基酸位置处的极性相反,特别是使得
a)cl结构域中18位的氨基酸残基具有正极性,特别是r、h或k中的任何一个;ch1结构域中26位的氨基酸残基具有负极性,特别是d或e中的任何一个;或
b)cl结构域中18位的氨基酸残基具有负极性,特别是d或e中的任何一个;且ch1结构域中26位的氨基酸残基具有正极性,特别是r、h或k中的任何一个。
具体地,abm包含一个或两个点突变,该一个或两个点突变是cl结构域中18位的点突变和ch1结构域中26位的点突变中的任一个或两个。
除非另有说明,否则在本文中位置根据imgt系统编号(lefranc等人,1999,nucleicacidsres.27:209-212)。权利要求中指出的位置的编号对应于根据kabat和kabat的eu索引的编号,如下表所示。kabat编号方案的解释可以在kabat,ea等人,sequencesofproteinsofimmunologicalinterest(nih出版号91-3242,第5版(1991))中找到。
当构建fab臂时,所指出的位置令人惊讶地证明是显性的,其中hc和lc以改善的亲和力组装(配对)。现有技术构建体涉及位于不同位置的ch1和cl点突变的不同配对,其除了显性vh和vl点突变之外被工程化。通过在指定的cl和ch1位置建立相反的极性,优选产生突变的cl和ch1结构域的同源对(在本文中理解为同源结构域或同源配对)。同时,与野生型cl和ch1结构域的假同源配对或配对显著减少。
具体地,abm的特征如下:
a
a)cl结构域是cκ,该cκ包含与seqid1具有至少90%序列同一性的氨基酸序列,其至少含有点突变t18x,其中x是r、h或k中的任何一个;和
b)ch1结构域包含与seqid3具有至少90%序列同一性的氨基酸序列,其至少含有点突变k26x,其中x是d或e中的任一个;
或b
a)cl结构域是cλ,该cλ包含与seqid2具有至少90%序列同一性的氨基酸序列,其至少含有点突变k18x,其中x是d或e中的任何一个;和
b)ch1结构域包含与seqid3具有至少90%序列同一性的氨基酸序列,其中26位的k未被任何其他氨基酸取代,或者其至少含有点突变k26x,其中x是r或h中的任一个;
或c
a)cl结构域是cλ,该cλ包含与seqid2具有至少90%序列同一性的氨基酸序列,其中18位的k未被任何其他氨基酸取代,或者其至少含有点突变k18x,其中x是r或h中的任一个;和
b)ch1结构域包含与seqid3具有至少90%序列同一性的氨基酸序列,其至少含有点突变k26x,其中x是d或e中的任一个;
其中根据imgt编号。
具体地,cl和ch1结构域是人源的,特别是人igg或igg1分子,特别是功能活性变体,该功能活性变体的特征在于与天然存在的人序列具有至少90%的序列同一性和一个或多个点突变,例如本文描述的,并且具体地特征还在于类似于人igg、igm或ige结构(特别是人igg1结构)中各个结构域的结构的抗体结构域的β-桶结构。
如本文所描述的任何cκ、cλ或ch1结构域的功能活性变体具体地特征在于能够与相应的匹配抗体结构域配对的抗体结构域结构,特别是其中
a
a)cl结构域变体是cκ变体,该cκ变体包含与seqid1具有至少90%序列同一性的氨基酸序列,并且其含有点突变t18x,其中x是r、h或k中的任一个;能够与下列配对
b)ch1结构域,该ch1结构域由鉴定为seqid3的氨基酸序列组成,除了点突变k26x,其中x是d或e中的任一个;
或b
a)cl结构域变体是cλ变体,该cλ变体包含与seqid2具有至少90%序列同一性的氨基酸序列,并且其含有点突变k18x,其中x是d或e中的任一个;能够与下列配对
b)ch1结构域,该ch1结构域由鉴定为seqid3或除了点突变k26x的seqid3中任一个的氨基酸序列组成,其中x是r或h中的任何一个;
或c
a)cl结构域变体是cλ变体,该cλ变体包含与seqid2具有至少90%序列同一性的氨基酸序列,其中18位的k未被任何其他氨基酸取代,或其含有点突变k18x,其中x是r或h中的任一个;能够与下列配对
b)ch1结构域,该ch1结构域由鉴定为seqid3的氨基酸序列组成,除了点突变k26x,其中x是d或e中的任一个。
具体地,cκ氨基酸序列(本文也称为野生型或亲本)由seqid1鉴定。
具体地,cλ氨基酸序列(本文也称为野生型或亲本)由seqid2鉴定。
具体地,ch1氨基酸序列(本文也称为野生型或亲本)由seqid3鉴定。
具体地,cl结构域的特征在于人igg1的cl序列或其工程化的功能活性变体,所述工程化的功能活性变体包含一个或多个点突变,优选多达10个点突变,特别是1、2、3、4、5、6、7、8、9或10个点突变中的任一个。
具体地,ch1结构域的特征在于人igg1的ch1序列或其工程化的功能活性变体,该工程化的功能活性变体包含一个或多个点突变,优选多达10个点突变,特别是1、2、3、4、5、6、7、8、9或10个点突变中的任一个。
具体地,cl和ch1结构域中的任何一个或每个的特征在于相应的人igg1序列或其工程化的功能活性变体,该工程化的功能活性变体包含一个或多个点突变,优选多达10个点突变,特别是1、2、3、4、5、6、7、8、9或10个点突变中的任一个。
具体地,二聚体包含cl和ch1结构域之间的至少一个域间二硫桥。具体地,域间二硫桥桥接ch1结构域中的cys220(eu索引编号)或cys233(根据kabat编号)和cl结构域中的cys214s。(eu索引和kabat编号)。
具体地,cl结构域还包含点突变f7x,其中x是s、a或v中的任一个,并且其ch1结构域还包含点突变a20l,其中根据imgt编号。这种进一步的点突变另外支持同源cl和ch1结构域的优选配对。
具体地,abm中的vl和vh结构域不含有改变界面区域中氨基酸的极性的任何点突变,该界面区域在配对vl和vh结构域时提供域间接触,从而形成抗原结合位点。
具体地,abm包含由vh/vl结构域配对组成的功能性抗原结合位点,能够以高亲和力结合靶标,且kd小于10-6m、10-7m、10-8m、10-9m或10-10m中的任一个。具体地,abm是靶向两种不同抗原的双特异性或异二聚体抗体,其中每种抗原被抗体识别,其kd小于10-6m、10-7m、10-8m、10-9m或10-10m中的任一个。
具体地,hc还包含至少一个ch2和至少一个ch3结构域。特别地,hc通过ch2结构域延伸并进一步被ch3结构域延伸,即ch2-ch3结构域的序列进一步与包含ch2-ch3结构域的另一抗体链配对,例如形成由ch2-ch3结构域和各自的链组成的fc区。具体地,通过使用或不使用接头或铰链区将ch2结构域融合至ch1结构域的c-端来延伸hc。具体地,通过使用或不使用接头将ch3结构域融合至ch2结构域的c-端来进一步延伸hc。在一些情况下,通过使用或不使用接头将ch4结构域融合至ch3结构域的c-端来进一步延伸hc。
具体地,abm包括铰链区,优选人铰链区,例如人igg1铰链区,例如包含鉴定为seqid4的氨基酸序列或由鉴定为seqid4的氨基酸序列组成。
结构域的连接特异性地通过重组融合或化学连接。特异性连接可以通过将一个结构域的c端连接到另一个结构域的n-端,例如,其中端区域中的一个或多个氨基酸残基被删除以缩短结构域大小,或延伸以增加结构域的灵活性。
具体地,缩短的结构域序列包括c-端和/或n-端区域的删除,例如删除至少1、2、3、4或5,多达6、7、8、9或10个氨基酸。
具体地,可以使用连接序列,其是免疫球蛋白的接头或铰链区或至少部分铰链区,例如肽接头,例如包括至少1、2、3、4或5个氨基酸,多达10、15或20个氨基酸。连接序列在本文中也称为“连接点”。结构域可以通过接头延伸,例如通过源自免疫球蛋白结构域的n-或c-末端区域的氨基酸序列,其天然地位于结构域附近,例如包括结构域之间的天然连接点。或者,接头可含有源自铰链区的氨基酸序列。但是,接头也可以是人工序列,例如,由连续的gly和/或ser氨基酸组成,优选长度为5至20个氨基酸,优选为8至15个氨基酸。
根据具体方面,abm是包含天然存在的免疫球蛋白或免疫球蛋白样支架的结构的抗体或免疫球蛋白,其中abm的特征在于至少一个(优选两个)抗原结合位点和在有或没有合适的连接序列的情况下由与重链和轻链相互连接的抗体结构域组成的结构,其中hc二聚化成lc以形成至少一个抗原结合位点,并且任选地其中两个hc二聚化形成fc区。
具体地,abm是抗体fab或(fab)2片段,或包含fc部分或fc区的全长抗体中的任一种,优选其中abm是全长igg、igm或ige抗体,特别是igg1、igg2、igg3或igg4中的任何一种。具体地,abm以任何合适的顺序包含一个或两个fab臂或fab片段(fab部分)。根据具体实施方案,abm甚至可以包含两个以上的fab臂,例如,三个或四个fab臂,其中fab臂中的至少一个或仅一个包含如本文所描述的同源lc/hc对和同源cl和ch1结构域。根据具体实施方案,其他fab臂包含野生型(天然存在的)cl和ch1结构域。
根据具体实施方案,abm仅包含一个同源lc/hc二聚体,其中hc进一步与包含ch2-ch3的fc链二聚化,任选地还包含ch4,从而获得fc区。这种abm特别是单价的单特异性抗体,其特征在于仅有一个fab臂和fc区。
根据另一具体实施方案,abm包含至少两个lc/hc二聚体,其中仅一个lc/hc二聚体的特征在于同源lc/hc二聚体和同源cl和ch1结构域(即,同源cl/ch1对),如本文所描述的。或者,abm由包含第一同源lc/hc二聚体的第一lc/hc二聚体和包含第二同源cl/ch1对的第二同源lc/hc二聚体组成,该第一同源lc/hc二聚体包含以本文所描述的点突变为特征的第一同源cl/ch1对,该第二同源cl/ch1对的特征在于本文所描述的点突变,它们与第一同源cl/ch1对的那些不同,使得第一同源cl和ch1结构域优选彼此配对,并且第二同源cl和ch1结构域优选彼此配对,但是,第一同源cl/ch1对的cl结构域优选不与第二同源cl/ch1对的ch1结构域配对(或甚至与其排斥),并且第一同源cl/ch1对的ch1结构域优选不与第二同源cl/ch1对的cl结构域配对(或甚至与其排斥)。
根据一具体实施方案,abm包含两个不同的fab臂,从而提供两种不同的fv结构,每种fv结构具有特异性结合特征。具体地,abm是靶向两种不同抗原或两种不同抗原表位的异二聚体或双特异性抗体。
本发明进一步提供了异二聚体或双特异性抗体,其包含识别不同抗原或表位的第一和第二fab臂,其中第一和第二fab臂中仅一个包含如本文所述的abm的同源lc/hc二聚体。具体地,异二聚体抗体是双特异性抗体或免疫球蛋白,或其抗原结合片段,例如双特异性全长免疫球蛋白或(fab)2。
具体地,abm是双特异性抗体,其中第一靶标是cd3、cd16或her2neu中的任一种,且第二靶标是egfr。
fab臂在本文中特别理解为由vh-ch1结构域序列组成的hc的二聚体和由vl-cl(κ或λ)结构域序列组成的lc,具有或不具有任何二硫桥、铰链结构域和/或连接抗体结构域的接头序列。当从抗体切割时,fab臂通常理解为fab片段(或fab部分)。fab臂具体地特征在于只有一个通过配对vh和vl结构域形成的抗原结合位点,其能够仅单特异性地和单价地结合靶标。
具体地,双特异性抗体中的第一和第二fab臂中的仅一个包含
a)cl结构域中的点突变f7x,其中x是s、a或v中的任一个;和
b)ch1结构域中的点突变a20l;
其中根据imgt编号。
上文指出的位置7和20处的这种点突变在本文中理解为支持性点突变,其实际上不改变氨基酸残基的极性,但是匹配对应氨基酸残基维度的空间特征。
具体地,包含上述支持性f7x点突变的cl结构域,其中x是s、a或v中的任一个,吸引并优选与包含支持性a20l点突变的对应ch1结构域配对,但是不优选地与野生型ch1结构域或不含a20l点突变的ch1结构域配对。
具体地,包含上述支持性a20l点突变的ch1结构域吸引并优选与包含上述支持性f7x点突变的对应cl结构域配对,其中x是s、a或v中的任一个,但是不优选地与野生型cl结构域或不含上述f7x点突变的cl结构域配对,其中x是s、a或v中的任何一个。
具体地,异二聚体抗体的特征在于
a
a)所述第一fab臂包含本文所描述的同源lc/hc二聚体,其具体特征在于上文鉴定的点突变,特别是一个或两个点突变,其提供cl结构域中18位的氨基酸残基和具有相反极性的ch1结构域中26位的氨基酸残基,其中cl和ch1结构域还包含上文鉴定的支持性点突变,特别是cl结构域中的点突变f7x,其中x是s、a或v中的任一个;和ch1结构域中的点突变a20l;且
b)所述第二fab臂不包含a)的点突变中的任一个,或
b
a)所述第一fab臂包含本文所描述的同源lc/hc二聚体,其具体特征在于上文鉴定的点突变,特别是一个或两个点突变,其提供cl结构域中18位的氨基酸残基和具有相反极性的ch1结构域中26位的氨基酸残基,其中cl和ch1结构域不进一步包含上文鉴定的支持性点突变,特别是cl结构域中的点突变f7x,其中x是s、a或v中的任一个;和ch1结构域中的点突变a20l;和
b)所述第二fab臂包含上文鉴定的支持性点突变,特别是cl结构域中的点突变f7x,其中x是s、a或v中的任一个;和ch1结构域中的点突变a20l。
a的这种双特异性构建体具体地特征在于本文所描述的点突变,用于优选配对同源lc/hc二聚体的同源cl和ch1结构域,其仅在两个fab臂(即,第一fab臂)中的一个中工程化,因此不利地配对或附着到另一个fab臂(即,第二fab臂)的hc或lc中的任一个。
b的这种双特异性构建体具体地特征在于第一fab臂,其包含如本文所描述的同源lc/hc二聚体,其特征在于在cl结构域中的18位和在ch1结构域中的26位的一个或两个点突变,从而获得在具有相反极性的这些位置上的氨基酸残基和包含支持性点突变的第二fab臂,从而
a)第一fab臂的hc有利地与第一fab臂的lc配对或连接,并且不利地与第二fab臂的lc配对或连接;和
a)第一fab臂的lc有利地与第一fab臂的hc配对或连接,并且不利地与第二fab臂的hc配对或连接;
反之亦然,意思是
c)第二fab臂的hc有利地与第二fab臂的lc配对或连接,并且不利地与第一fab臂的lc配对或连接;和
d)第二fab臂的lc有利地与第二fab臂的hc配对或连接,并且不利地与第一fab臂的hc配对或连接。
根据一具体实施方案,第一和第二fab臂均在cl结构域中的18位和ch1结构域中的26位包含一个或两个点突变,从而在具有相反极性的这些位置处获得氨基酸残基。但是,其中第一和第二fab臂中的点突变是不同的,从而产生
a)包含cl结构域的第一fab臂,其中18位的氨基酸残基具有特异性识别ch1结构域的正极性,其中26位的氨基酸残基具有负极性;和
b)包含cl结构域的第二fab臂,其中18位的氨基酸残基具有特异性识别ch1结构域的负极性,其中26位的氨基酸残基具有正极性;
任选地,其中支持性点突变位于第一fab臂或第二fab臂中。
其他实施方案涉及双特异性构建体,其中
a)包含cl结构域的第一fab臂,其中18位的氨基酸残基具有特异性识别ch1结构域的正极性,其中26位的氨基酸残基具有负极性;和
b)第二fab臂,其中cl结构域中18位的氨基酸残基和/或ch1结构域中26位的氨基酸残基是不带电荷的,特别是n、c、q、g、s、t、w或y中的任何一个;或非极性的,特别是a、i、l、m、f、p或v中的任何一个;
任选地,其中支持性点突变位于第一fab臂或第二fab臂中。
根据具体方面,本文所描述的abm,特别是本文所描述的异二聚体抗体,包含两个hc,每个hc包含ch2和ch3结构域,和任选的ch4结构域,其中hc二聚化成fc区。
fc区具体地特征在于fc链的二聚体,每个fc链的特征在于包含ch2-ch3抗体结构域的链,该二聚体可以是同二聚体或异二聚体,例如,其中第一fc链与ch2和/或ch3结构域中的至少一个点突变中的第二fc链不同。
具体地,fc区包含两个ch3结构域,所述两个ch3结构域被工程化为引入和/或表征为以下一种或多种:
a)链交换工程化结构域(seed)ch3异二聚体,其由人iga和iggch3序列的交替区段组成;
b)一种或多种隆突或穴突变,优选为t366y/y407’t、f405a/t394’w、t366y:f405a/t394’w:y407’t、t366w/y407’a和s354c:t366w/y349’c:t366’s:l368’a:y407’v中的任一种;
c)第一ch3结构域中的半胱氨酸残基,其与第二ch3结构域中的半胱氨酸残基共价连接,从而引入域间二硫桥,优选连接两个ch3结构域的c-端;
d)一种或多种突变,其中排斥电荷抑制异二聚体形成,优选为以下任何一种:k409d/d399’k、k409d/d399’r、k409e/d399’k、k409e/d399’r、k409d:k392d/d399’k:e356’k或k409d:k392d:k370d/d399’k:e356’k:e357’k;和/或
e)选择用于异二聚体形成和/或热稳定性的一种或多种突变,优选为以下任何一种:
t350v:l351y:f405a:y407v/t350v:t366l:k392l:t394w,
t350v:l351y:f405a:y407v/t350v:t366l:k392m:t394w,
l351y:f405a:y407v/t366l:k392m:t394w,
f405a:y407v不366l:k392m:t394w,或
f405a:y407v/t366l:t394w,
其中根据kabat的eu索引编号。
这样的ch3突变被工程化以分别产生两种不同的fc链和hc(不同之处至少为ch3结构域的不同序列),其优选彼此配对,从而获得fc链或hc的异二聚体,显著减少产生hc同二聚体的趋势,即,相同序列的两个hc的二聚体。
在本文所描述的ch3点突变的说明中,“斜线”区分一条链或一个结构域上的点突变与来自相应对的另一条链或其他结构域的点突变;氨基酸位置编号中的“缩进”表示异二聚体的第二链或二聚体。“冒号”分别识别链或结构域之一上的点突变的组合。
可以使用如上所述选择用于异二聚体形成和/或热稳定性的任何突变或根据vonkreudenstein等人的公开内容的其他突变(landesbioscience,vol.5,no.5,2013,pp646-654)。
优选地,(i)隆突;或(ii)穴突变,或(iii)隆突和穴突变,在一个链或结构域上工程化,对应物(i)穴,或(ii)隆突突变,或(iii)穴和隆突突变,是在异二聚体的另一条链上工程化的。
具体地,包含一个或两个工程化ch3结构域的一对ch3结构域可包含一个以上(另外的)的域间二硫桥,例如,2或3个,连接两个ch3域的配对。
具体地,在相应的ch3结构域对的两个ch3结构域中工程化不同的突变(根据上述a))以产生同源(匹配)对,其中一个结构域包含β-片区中接触表面的空间修饰,其优选地,通过互补的空间修饰连接到另一个结构域的相应接触表面。这种空间修饰主要来自不同的氨基酸残基和侧链,例如,产生“隆突”或“穴”结构,它们互补以形成“隆突入穴”二聚体。
根据一具体方面,fc区中的每个ch3结构域是igg型,其氨基酸序列被鉴定为seqid5或seqid5的功能变体,其被工程化以通过掺入至少一个具有至少2个氨基酸长度的β链iga区段获得链交换,并且fc区优选包含通过将第一ch3结构域的iga区段与第二ch3结构域的iga区段配对的ch3结构域的同源对。这种链交换的ch3结构域具体地可以包含iga和igg氨基酸序列的交替片段,例如,包含至少1、2、3、4或5个不同的iga区段,每个区段位于不同的位置并通过非iga区段彼此分开,例如igg区段。
根据具体方面,abm是效应功能感受态抗体,该抗体包含fcγ受体结合位点和/或c1q结合位点,任选地在fc区中。
具体地,抗体的特征在于adcc和/或cdc活性中的任一种。
根据另一具体方面,abm是效应阴性(en)抗体,该抗体包含缺乏与fcγ受体和/或c1q结合的fc区。
具体地,抗体是效应缺陷的(本文也称为效应阴性),通过fc区显著降低或不结合fcγ受体或cd16a。
具体地,效应阴性抗体的特征在于人igg2ch2序列或其工程化变体,其包含us8562986中描述的修饰的人igg2ch2结构域(f296a、n297q),融合至c-端ch3结构域的n-端(根据eu的kabat索引编号)。
具体地,en抗体具有显著降低的或没有adcc和/或cdc。
具体地,abm包含抗体的fc部分,其包含在ch2与ch3结构域的相互连接处的fcrn结合位点,和/或在ch2结构域的n端区域内的fcγ受体结合位点,和/或ch2结构域的n-端区域内的c1q结合位点。
根据具体方面,abm包含位于ch2和/或ch3结构域中的ph依赖性fcrn结合位点(如果有的话)。具体地,fcrn结合位点具有以ph依赖性方式结合fcrn的亲和力,kd小于10-4m或小于10-5m、10-6m、10-7m或10-8m。
具体地,与在生理ph(ph7.4)下的相同结合亲和力相比,以ph依赖性方式结合fcrn的结合亲和力在ph5-6下增加至少1-log,优选为至少2-log或3-log。
根据另一方面,abm被工程化以改变ph依赖性fcrn结合。例如,至少一个ch3结构域被工程化以在fcrn结合位点包含至少一个突变以降低ph依赖性fcrn结合,特别是h433a或h435a突变中的至少一个,或h433a和h435a突变二者,其中根据eu的kabat索引编号。ph依赖性fcrn结合的降低可以使以ph依赖性方式结合fcrn的结合亲和力在ph5-6下小于1-log,优选大约相同或更小,与在生理ph(ph7.4)下相同的结合亲和力相比。
具体实施方案涉及本文示例的任何abm,或包含实施例部分中描述的任何重链和轻链或任何重链和轻链对。具体地,如本文所描述的abm可包含实施例部分中描述的重链和轻链或由实施例部分中描述的重链和轻链组成。
具体地,提供本文描述的abm用于医学、诊断或分析用途。
本发明进一步提供了包含本文所述abm的药物制剂,优选在肠胃外或粘膜制剂中,任选地含有药学上可接受的载体或赋形剂。
本发明还提供了编码本文所述abm的分离的核酸。
本发明进一步提供了表达盒或质粒,其包含或包括本文所述的核酸和任选的表达由核酸序列编码的abm的其他序列,例如调节序列。
具体地,表达盒或质粒包含表达本文所描述的abm的编码序列,或本文所描述的abm的hc和/或lc。
根据具体实例,abm由一个或多个hc和lc组成,其中每个hc的特征在于相同的hc氨基酸序列,并且每个lc的特征在于相同的lc氨基酸序列,并且编码hc和lc的序列用于产生单价或同二聚体抗体。
根据另一具体实例,abm由两个不同的hc和两个不同的lc组成,并且两个不同的hc和两个不同的lc的编码序列用于产生异二聚体或双特异性抗体。
本发明进一步提供了生产宿主细胞,该生产宿主细胞包含至少一个表达盒或质粒,所述表达盒或质粒包含一个或多个编码本文所述abm的核酸分子。
具体地,宿主细胞瞬时或稳定地表达abm。根据具体实例,宿主细胞是真核宿主细胞,优选为任何酵母或哺乳动物细胞。
本发明还提供了生产本文所描述的abm的方法,其中在生产所述abm的条件下培养或维持本文所描述的宿主细胞。
具体地,可以从细胞培养上清液中分离和/或纯化abm。根据具体实例,abm是双特异性全长抗体,该抗体是包含两个不同hc和两个不同lc的异二聚体,并且abm分别包含同源hc/lc对和同源cl和ch1结构域的正确配对,abm由宿主细胞产生,其中少于10%的产生的抗体配对错误,优选小于5%,如通过质谱法(lc-esi-ms)比较最大峰强度测量的。
附图说明
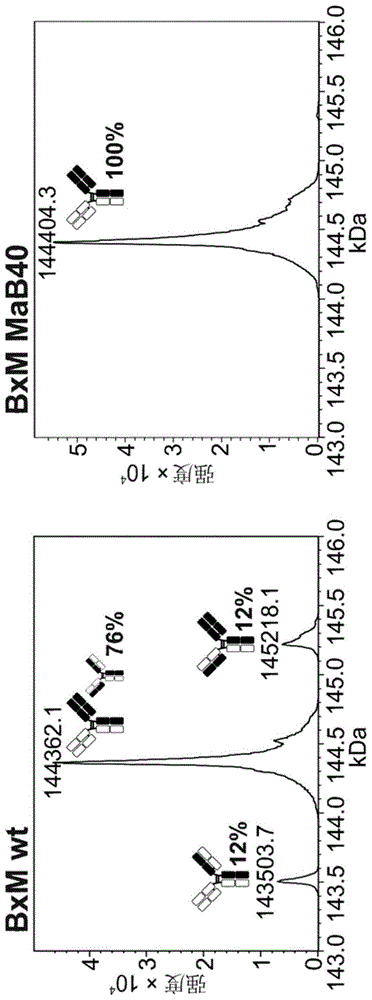
图1:双特异性iggbxm在expi293f中瞬时产生,其不携带界面突变(左图)或携带mab40的界面突变(右图)。将两种抗体去糖基化并通过lc-esi-ms分析。b10v5轻链和重链以白色显示,且hu225m轻链和重链以黑色显示。每种检测到的链配对变体的相对丰度表示为所有检测到的完整igg的百分比。在bxmwt中,fab中错配的两种变体都可检测到相当大的量(当计算最大峰强度时各自为12%)。因此,正确配对变体的峰值还将包含其中轻链具有交换位置的错配变体。bxmmab40的生产仅产生正确配对的变体。由于界面工程化,错配的变体消失;
图2:纯化的bxm野生型和bxmmab40的分析尺寸排阻色谱。两种igg均在16.3分钟的预期时间洗脱。没有检测到界面工程化对sec曲线的负面影响;
图3:双特异性iggbxo在hek293-6e中瞬时产生,其不携带界面突变(bxowt,左上图)或携带mab40的界面突变(bxomab40,右上图)。引入b10v5fab中的支持性突变,其导致产生双特异性抗体bxomab5/40、bxomab21/40和bxomab45/40(剩余的下图)。将所有抗体去糖基化并通过lc-esi-ms分析。b10v5轻链和重链以白色显示,且okt3轻链和重链以黑色显示。每种检测到的链配对变体的相对丰度表示为所有检测到的完整igg的百分比。在bxowt中,fab中错配的两种变体都可以不同的量检测到,累积到大于40%的错配抗体。在bxomab40中,错配显著降低,但仍可检测到。bxo不仅含有mab40的突变,还含有任一支持性突变,显示出改善的配对行为。所有检测到的完整igg中超过90%是正确配对的bxo;
图4:纯化的bxo野生型、bxomab40、bxomab5/40和bxomab45/40的分析尺寸排阻色谱。所有igg均在15.4min的预期时间洗脱。没有检测到界面工程化对sec曲线的负面影响;
图5:序列
seqid1:人igg1的cκ结构域的氨基酸序列
seqid2:人igg1的cλ结构域的氨基酸序列
seqid3:人igg1的ch1结构域的氨基酸序列
seqid4:人igg1铰链区的氨基酸序列
seqid5:人igg1的ch3结构域的氨基酸序列
具体实施方式
在整个说明书中使用的具体术语具有以下含义。
如本文所用,术语“抗原结合分子”或abm应该是指包含结合结构域的分子,所述结合结构域是具有特定结合亲和力和/或亲合力的特异性识别或结合抗原或其表位的多肽。根据abm的具体实例,结合结构域是免疫球蛋白型结合区,该免疫球蛋白型结合区包含选自下组的多肽:单结构域抗体、单链可变结构域、fd片段、犰狳(armadillo)重复多肽、iii型纤连蛋白结构域、iii型腱生蛋白结构域、锚蛋白重复基序结构域、脂质运载蛋白、kunitz结构域、fyn衍生的sh2结构域、小蛋白、c型凝集素样结构域支架、工程化抗体模拟物,以及保留抗原结合功能的任何前述遗传操作的对应物。
abm的具体实施方案包含抗体或其抗原结合片段或由抗体或其抗原结合片段组成。
如本文所用,术语“抗体”定义为抗原结合多肽,其为免疫球蛋白或免疫球蛋白样分子,或显示模块抗体形式的其他蛋白质,例如,由一个或多个抗体结构域组成并具有与免疫球蛋白或抗体相似的抗原结合特性,特别是可表现出单特异性或双特异性或单价、双价或多价结合特性的蛋白质,用于例如,抗原、效应分子或结构的表位的至少两个特异性结合位点,特别是病原体起源或人体结构,如自身抗原,包括细胞相关或血清蛋白。术语“抗体”和“免疫球蛋白”在本文中可互换使用。
抗体通常由抗体结构域组成或包含抗体结构域,其被理解为免疫球蛋白的重链和/或轻链的恒定和/或可变结构域,具有一个或多个或不具有接头序列。抗体特别理解为由可变和/或恒定抗体结构域的组合组成或包含可变和/或恒定抗体结构域的组合,其具有或不具有连接序列或铰链区,包括可变抗体结构域对,例如一个或两个vh/vl对。多肽被理解为抗体结构域,如果包含β-桶结构,该β-桶结构由通过环序列连接的抗体结构域结构的至少两个β链组成。抗体结构域可以是天然结构或通过诱变或衍生化修饰,例如,以修饰抗原结合特性或任何其他特性,例如稳定性或功能特性,例如与fc受体fcrn和/或fcγ受体的结合。
如本文所用,术语“抗体”特别地包括全长抗体,包括免疫球蛋白样结构的抗体。具体地,抗体可以是全长抗体,例如igg型(例如,igg1、igg2、igg3或igg4亚型)、iga1、iga2、igd、ige或igm抗体。
术语还包括抗体、抗体结构域或抗体片段的衍生物、组合或融合中的任一种。
术语“全长抗体”用于指包含抗体的fc区或至少大部分fc部分的任何抗体分子,其特异性地包括重链二聚体。术语“全长抗体”在本文用于强调特定抗体分子不是抗体片段。
据此,抗体通常被理解为蛋白质(或蛋白质复合物),其包括基本上由免疫球蛋白基因或免疫球蛋白基因片段编码的一种或多种多肽。公认的免疫球蛋白基因包括κ、λ、α、γ、δ、ε和μ恒定区基因,以及免疫球蛋白可变区基因。轻链(lc)分类为κ(包括vl和cλ结构域)或λ(包括vl和cκ结构域)。重链(hc)分类为γ、μ、α、δ或ε,其反过来分别定义免疫球蛋白类别,igg、igm、iga、igd和ige。
hc或lc各自由至少两个彼此连接的结构域组成,以产生结构域的链。特别理解的是,抗体hc包括vh抗体结构域和至少一个与vhc-端结合的抗体结构域,即,在有或没有连接序列的情况下,至少一个抗体结构域连接到vh结构域的c端。抗体lc包括vl抗体结构域和至少一个与vlc-端结合的抗体结构域,即,在有或没有连接序列的情况下,至少一个抗体结构域连接到vl结构域的c端。
所述定义还包括可变区的重链和轻链的结构域(例如dab、fd、vi、vk、vh、vhh)和完整抗体的恒定区或单个结构域,例如ch1、ch2、ch3、ch4、ci和ck,以及由结构环连接的抗体结构域的至少两个β链组成的小结构域。通常,具有通过特定cdr结构的抗原结合位点的抗体能够通过一对vh/vl结构域的cdr环结合靶抗原。
术语“抗体”应具体包括分离形式的抗体,其基本上不含针对不同靶抗原的其他抗体和/或包含抗体结构域的不同结构排列。仍然,分离的抗体可以包含在组合制剂中,其含有分离的抗体的组合,例如,具有至少一种其他抗体,例如具有不同特异性的单克隆抗体或抗体片段。
术语“抗体”应适用于动物来源的抗体,包括人类物种,如哺乳动物,包括人、鼠、兔、山羊、骆驼科动物、美洲驼、牛和马,或禽类,如母鸡,该术语应特别包括基于动物来源序列的重组抗体,例如,人类序列。
术语“抗体”特别适用于人抗体。
关于抗体使用的术语“人”应理解为包括具有衍生自人种系免疫球蛋白序列的可变区和恒定区的抗体。人抗体可包括不由人种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机或位点特异性诱变或通过体内体细胞突变引入的突变),例如在cdr中。人抗体包括从人免疫球蛋白文库分离的抗体或从一种或多种人免疫球蛋白转基因动物中分离的抗体。
人抗体优选选自或衍生自iga1、iga2、igd、ige、igg1、igg2、igg3、igg4和igm。
鼠抗体优选选自或衍生自iga、igd、ige、lgg1、igg2a、igg2b、igg2c、igg3和igm。
术语“抗体”进一步适用于嵌合抗体,例如嵌合抗体,具有不同物种的起源序列,例如鼠和人来源的序列。
关于抗体使用的术语“嵌合”是指其中重链和轻链的每个氨基酸序列的一部分与衍生自特定物种或属于特定类别的免疫球蛋白中的相应序列同源的那些分子,而链的剩余部分与另一物种或类别中的相应序列同源。通常,轻链和重链的可变区模拟衍生自一种哺乳动物物种的免疫球蛋白的可变区,而恒定部分与衍生自另一种的免疫球蛋白的序列同源。例如,可变区可以源自目前已知的来源,使用来自非人宿主生物的易于获得的b细胞或杂交瘤与衍生自例如人细胞制剂的恒定区组合。
术语“抗体”可以进一步适用于人源化抗体。
关于抗体使用的术语“人源化”是指具有抗原结合位点的分子,其基本上衍生自来自非人物种的免疫球蛋白,其中分子的剩余免疫球蛋白结构基于人免疫球蛋白的结构和/或或序列。抗原结合位点可以包含融合到恒定结构域上的完整可变结构域或仅包含移植到可变结构域中的适当框架区上的互补决定区(cdr)。抗原结合位点可以是野生型或修饰的,例如,通过一个或多个氨基酸取代,优选被修饰以更接近地类似于人免疫球蛋白。一些形式的人源化免疫球蛋白保留所有cdr序列(例如含有来自小鼠抗体的所有六个cdr的人源化小鼠抗体)。其他形式具有相对于原始抗体改变的一个或多个cdr。
根据一具体实施方案,如本文所述的abm中包含的所有抗体结构域是人源或其人源化或功能活性变体,具有至少60%的序列同一性,或至少70%、80%、90%或95%的序列同一性,优选地其中抗体结构域的来源是igg1、igg2、igg3、igg4、iga、igm或ige抗体中的任何一种。具体地,所有抗体结构域源自相同的基本免疫球蛋白折叠,尽管b-片形式可以不同,并且连接环当然是可变的,尤其是在v结构域中。
术语“抗体”还适用于单克隆或多克隆抗体,特别是重组抗体,该术语包括通过重组方法制备、表达、产生或分离的所有抗体和抗体结构,例如来自动物,例如哺乳动物包括人的抗体,其包括来自不同来源的基因或序列,例如嵌合、人源化抗体或杂交瘤衍生的抗体。进一步的实例是指从转化以表达抗体的宿主细胞中分离的抗体,或从抗体或抗体结构域的重组组合文库中分离的抗体,或通过涉及抗体基因序列到其他dna序列剪接的任何其他方式制备、表达、产生或分离的抗体。
术语“抗体”应理解为包括新的或现有的功能活性变体,例如,天然存在的抗体。还应理解,术语抗体的变体,特别是抗体样分子的变体或抗体变体,也应包括这些分子的衍生物。
衍生物是一种或多种抗体和/或融合蛋白的任何组合,其中抗体的任何结构域或小结构域可以在任何位置与一种或多种其他蛋白质融合,例如与其他抗体或抗体片段融合,但也可以与配体、酶、毒素等融合。本文描述的abm或抗体可特异性地用作分离的多肽或组合分子,例如,通过与其他肽或多肽的重组、融合或缀合技术。肽优选与抗体结构域序列同源,且优选长度为至少5个氨基酸,更优选长度为至少10个氨基酸或甚至至少为50或100个氨基酸,并且至少部分构成抗体结构域的环区。
抗体的衍生物也可以通过各种化学方法(例如,共价偶联、静电相互作用、二硫化物键合等)与其他物质缔合或结合而获得。与抗体结合的其他物质可以是脂质、碳水化合物、核酸、有机和无机分子或其任何组合(例如peg、前药或药物)。衍生物还包含具有相同氨基酸序列但完全或部分由非天然或化学修饰的氨基酸制备的抗体。在一具体实施方案中,抗体是包含允许与生物学上可接受的化合物特异性相互作用的另外的标签的衍生物。对于可用的标签没有特定限制,只要它对抗体与其靶标的结合没有或具有可接受的负面影响即可。合适的标签的实例包括his标签、myc标签、flag标签、strep标签、钙调蛋白标签、gst标签、mbp标签和s标签。在另一具体实施方案中,抗体是包含标记的衍生物。如本文所用,术语“标记”是指可检测的化合物或组合物,其直接或间接与抗体缀合以产生“标记的”抗体。标签可以自身检测,例如,放射性同位素标记或荧光标记,或者在酶标记的情况下,可以催化可检测的底物化合物或组合物的化学改变。
抗体的衍生物例如是,衍生自亲本抗体或抗体序列,例如亲本抗原结合(例如cdr)或框架(fr)序列,例如在硅化或重组工程中或通过化学衍生化或合成获得的突变体或变体。
如本文所用,术语“变体”应具体包括如本文所述的任何“突变体”、“同源物”或“衍生物”。术语“变体”应具体包括功能活性变体,其特征在于某种功能。
本文所述的abm或抗体的功能特别在于特定的抗原结合特性(特别是表位特异性)和cl和ch1结构域的优选配对,其中cl结构域中18位的氨基酸和/或ch1结构域中26位的氨基酸的特征在于相反的极性(根据imgt编号)。抗体恒定结构域的功能变体的功能在本文中被理解为与对应抗体结构域配对以产生抗体结构域对的能力。特别地,本文所述的cl和ch1结构域的功能变体包含优选配对的主要点突变,其中cl结构域中18位的氨基酸和/或ch1结构域中26位的氨基酸特征在于相反极性(“显性点突变”;根据imgt编号),以及任选地支持优选配对以产生cl/ch1二聚体的另外点突变,但不降低配对的可能性。
术语“变体”应特别指抗体,例如突变体抗体或抗体片段,例如通过诱变方法获得的,特别地是指将插入物删除、交换、引入特定抗体氨基酸序列或区域或化学衍生化氨基酸序列中,例如在恒定结构域中以工程化抗体稳定性,效应子功能或半衰期,或在可变结构域中改善抗原结合特性,例如通过本领域可获得的亲和力成熟技术获得的。可以使用任何已知的诱变方法,包括在所需位置的点突变,例如,通过随机化技术获得的。在某些情况下,随机选择位置,例如用任何可能的氨基酸或选择的优选氨基酸随机化抗体序列。术语“诱变”是指用于改变多核苷酸或多肽序列的任何本领域公认的技术。优选的诱变类型包括易错pcr诱变、饱和诱变或其他定点诱变。
术语“功能变体”在本文中也称为“功能活性变体”,可以例如包括通过插入、删除或取代一个或多个氨基酸,或氨基酸序列中一个或多个氨基酸残基的化学衍生,或核苷酸序列内的核苷酸,或例如在cdr或fr序列中的序列远端的任一端或两端修饰亲本序列(例如来自亲本抗体)产生的序列,哪种修饰不影响,特别是损害该序列的活性。在结合位点对所选靶抗原具有特异性的情况下,抗体的功能活性变体仍然具有预定的结合特异性,尽管这可以改变,例如,改变对特定表位的精细特异性、亲和力、亲合力、kon或koff率等。例如,亲和力成熟的抗体特别理解为功能活性变体抗体。因此,亲和力成熟的抗体中的修饰的cdr序列被理解为功能活性变体。
与母体分子相比,功能活性优选由变体的结构和功能决定,例如在用于以ph依赖性方式确定结合靶抗原和/或分子的期望体内半衰期和/或fcrn结合的特异性的测定中,例如,通过测量抗体的功能在标准测定中测定的。
当抗原在细胞表面上表达时,抗体在抗原结合方面的功能活性通常在elisa测定、biacore测定、octetbli测定或基于facs的测定中测定。
可以获得功能活性变体,例如,通过改变亲本抗体的序列,例如具有抗体的特异性天然结构的单克隆抗体,例如igg1结构,以获得在识别靶抗原时具有相同特异性但具有与母体结构不同的结构的变体,例如,以修饰任何抗体结构域以引入特定突变,产生双特异性构建体或产生母体分子的片段。
通常,可修饰亲本抗体或序列以产生在抗原结合位点之外或在结合位点内的序列区域内掺入突变的变体,其不损害抗原结合,并且优选具有与亲本抗体类似的生物学活性,包括结合抗原的能力,例如具有基本相同的生物活性,如通过特异性结合测定或靶向抗原的功能试验所确定的。
如本文所用,术语“基本上相同的生物活性”是指由基本上相同的活性表示的活性为如对于可比较的或亲本抗体测定的活性的至少20%、至少50%、至少75%、至少90%,例如至少20%、至少100%,或至少125%,或至少150%,或至少175%,或例如高达200%。
如本文所述的优选变体在抗原结合方面具有功能活性,优选地其具有特异性结合单个抗原的效力,并且不显著结合非靶抗原的其他抗原,例如具有至少2log,优选至少3log的kd值差异。通过功能活性变体的抗原结合通常不受损害,对应于与亲本抗体或序列基本上相同的结合亲和力,或包含序列变体的抗体,例如,具有小于2log,优选小于3log的kd值差异,但是,具有甚至提高的亲和力的可能性,例如,具有至少1log,优选为至少2log的kd值差值。
在一优选的实施方案中,亲本抗体a)的功能活性变体是抗体的生物活性片段,该片段包含至少50%的分子序列,优选至少60%、至少70%、至少80%、至少90%,或至少95%,且最优选为至少97%、98%或99%;
b)通过至少一个氨基酸取代、添加和/或删除衍生自抗体,其中功能活性变体与分子或其部分具有序列同一性,例如具有至少50%序列同一性,优选至少60%,更优选至少70%,更优选至少80%,还更优选至少90%,甚至更优选至少95%,且最优选至少97%、98%或99%的抗体;和/或
c)由抗体或其功能活性变体以及另外至少一个与多肽或核苷酸序列异源的氨基酸或核苷酸组成。
在一个实施方案中,如本文所述的abm或抗体的功能活性变体与上述变体基本相同,但分别与其多肽或编码核苷酸序列不同,因为它衍生自不同物种的同源序列。这些被称为天然存在的变体或类似物。
术语“功能活性变体”还包括天然存在的等位基因变体,以及突变体或任何其他非天然存在的变体。如本领域所知,等位基因变体是(多)肽的替代形式,其特征在于具有一个或多个氨基酸的取代、删除或添加,其基本上不改变多肽的生物学功能。
功能活性变体可以通过多肽或核苷酸序列中的序列改变来获得,例如,通过一个或多个点突变,其中当如本文所述使用时,序列改变保留或改善未改变的多肽或核苷酸序列的功能。此类序列改变可包括但不限于(保守的)取代、添加、删除、突变和插入。
具体的功能活性变体是cdr变体。cdr变体包括由cdr区中的至少一个氨基酸修饰的氨基酸序列,其中所述修饰可以是氨基酸序列的化学或部分改变,该修饰允许该变体保留未修饰序列的生物学特征。cdr氨基酸序列的部分改变可以通过删除或取代一个至几个氨基酸,例如,1、2、3、4或5个氨基酸,或通过添加或插入一个或几个氨基酸,例如,1、2、3、4或5个氨基酸,或通过一至几个氨基酸的化学衍生,例如,1、2、3、4或5个氨基酸,或其组合。氨基酸残基中的取代可以是保守取代,例如,用一个疏水氨基酸取代另一个疏水氨基酸。
保守取代是在与其侧链和化学性质相关的氨基酸家族中发生的取代。这类家族的实例是具有碱性侧链、具有酸性侧链、具有非极性脂肪族侧链、具有非极性芳族侧链、具有不带电荷的极性侧链、具有小侧链、具有大侧链等的氨基酸。
点突变特别理解为多核苷酸的工程化,对于不同氨基酸其导致在一个或多个单个(非连续的)或双联体氨基酸的取代或交换、删除或插入中不同于非工程化氨基酸序列的氨基酸序列的表达。
根据某一方面,如本文所描述的针对抗体cl和ch1结构域的优选配对的cl和ch1结构域的点突变改变了cl结构域中18位的氨基酸残基和/或具有相反的极性的ch1结构域中26位的氨基酸的极性,其中根据imgt编号。具体实施方案涉及以下内容:
a)其中cl结构域中18位的氨基酸残基具有正极性,ch1结构域中26位的氨基酸残基具有负极性;或
b)其中cl结构域中18位的氨基酸残基具有负极性,ch1结构域中26位的氨基酸残基具有正极性。
上述点突变在本文中称为“显性”点突变,因为通过在指定位置中相反极性的这种点突变,可以产生abm或抗体,其特征在于具有这种突变的cl和ch1结构域的优选配对,即使在cl或ch1结构域或任何相邻的vl或vh结构域中没有其他点突变。
除了显性点突变之外,还可以存在其他点突变,其甚至改善lc和hc的优选配对,例如本文称为“支持性”点突变的点突变。这种支持性点突变可以在任何cl和/或对应ch1结构域中,或在vl和/或对应vh结构域中进行工程化。示例性支持性点突变如下:cl结构域中的点突变f7x,其中x是s、a或v中的任何一个;和对应ch1结构域中的点突变a20l,其中根据imgt编号。通常,支持性点突变是保守性点突变,其特征在于氨基酸残基的取代,其中氨基酸残基的极性不会被这种取代改变。
如本文所述的abm或抗体的变体可包括点突变,所述点突变是指相同极性和/或电荷的氨基酸的交换。在这方面,氨基酸是指由64个三联体密码子编码的20种天然存在的氨基酸。这20种氨基酸可分为具有中性电荷、正电荷和负电荷的那些氨基酸:
20种天然存在的氨基酸如下表所示,以及它们各自的三字母和单字母代码和极性:
关于多肽序列的“百分比(%)氨基酸序列同一性”定义为在比对序列和引入缺口后的候选序列中与特定多肽序列中的氨基酸残基相同的氨基酸残基的百分比,必要时,实现最大百分比序列同一性,而不考虑任何保守取代作为序列同一性的一部分。本领域技术人员可以确定用于测量比对的适当参数,包括在所比较的序列的全长上实现最大比对所需的任何算法。
abm或抗体变体特别理解为包括具有特定糖基化模式的同源物、类似物、片段、修饰或变体,例如,通过糖基化工程产生的,其是功能性的并且可以用作功能等同物,例如,结合到特定目标并且具有功能性质。abm或抗体可以是糖基化的或非糖基化的。例如,如本文所述的重组abm或抗体可以在合适的哺乳动物细胞中表达,以允许分子的特异性糖基化,如由表达抗体的宿主细胞所确定的。
术语抗体结构域的“β-片”或“β链”,特别是恒定抗体结构域如cl或ch1结构域,在本文中以下列方式理解。抗体结构域通常由至少两条β链组成,所述β链通过至少两个或三个主链氢键侧向连接,形成通常扭曲的褶皱片。β链是通常3至10个氨基酸长度的单个连续氨基酸段,采用这种延伸的构象并包括至少一个其他链的主链氢键,使得它们形成β片。在β片中,大多数β链与其他链相邻排列,并与其邻域形成广泛的氢键网络,其中一条链骨架中的n-h基团与相邻链骨架中的c=o基团形成氢键。
抗体恒定结构域(例如cl或ch1结构域)的结构类似于可变结构域的结构,其由通过环连接的β-链组成,其中一些包含短的α-螺旋状延伸。从各种fc晶体结构的b因子可以看出,该框架主要是刚性的并且环相对更柔韧。抗体cl或ch1结构域通常具有七条形成β-片(a-b-c-d-e-f-g)的β链,其中β链通过环连接,三个环位于结构域的n-端末端(a-b、c-d、e-f),并且另外三个环位于结构域的n端-末端(b-c、d-e、f-g)。结构域的“环区”是指位于β链区域之间的蛋白质部分(例如,每个cl或ch1结构域包含7个β片,a至g,从n-端到c-端)。
优选地,一对抗体结构域,例如通过连接涉及每个结构域的a、b和/或e链的结合表面产生(异)二聚体的一对cl和ch1结构域(在本文中称为结合界面)。通过cl结构域的β-片区域与ch1结构域的β-片区域的这种接触,产生二聚体(称为cl/ch1)。
具体地,如本文所描述的cl和ch1结构域包含或组成于人igg1抗体的氨基酸序列。
特别地,cκ结构域的特征在于鉴定为seqid1的氨基酸序列,或其功能变体,例如具有一定的序列同一性。
特别地,cλ结构域的特征在于鉴定为seqid2的氨基酸序列,或其功能变体,例如具有一定的序列同一性。
特别地,ch1结构域的特征在于鉴定为seqid3的氨基酸序列,或其功能变体,例如具有一定的序列同一性。
或者,如本文所述的cl和ch1抗体结构域包含或组成于任何人lgg2、lgg3、lgg4、iga、igm、ige、igd或其功能变体的氨基酸序列,例如具有一定的序列同一性。
抗体的fv部分通常被理解为通过连接涉及每个结构域的c、c’和f链的结合表面(结合界面)产生(异)二聚体的vl和vh结构域对。通过vl结构域的β-片区域与vh结构域的β-片区域的这种接触,产生二聚体(称为vl/vh)。
fab臂在本文中理解为第一和第二抗体链对,其中第一链包含vl结构域和cl结构域或由vl结构域和cl结构域组成,其连接到vl结构域的c端(轻链,lc),并且第二链包含vh结构域和ch1结构域或由vh结构域和ch1结构域组成,其连接到vh结构域的c端(重链,hc),其中vl通过结合界面连接到vh(与之配对),且cl通过结合界面连接到ch1(与之配对),从而产生lc和hc的(异)二聚体(也称为lc/hc)。
抗体的fc部分在本文中理解为一对抗体链,各自包含ch2结构域和ch3结构域,其连接至ch2结构域的c端(fc链),其中每个抗体链的ch2结构域通过涉及每个ch2结构域的a、b和/或e链的结合表面(结合界面)彼此连接,并且其中每个抗体链的ch3结构域通过涉及每个ch3结构域的a、b和/或e链的结合表面(结合界面)彼此连接(配对),从而产生fc链的(同)二聚体。本文描述的fc部分可以来自igg、iga、igd、ige或igm。
在本文所描述的一个实施方案中,fc部分包含突变的ch3结构域,例如,其具有被异源序列取代的一条或多条β链的至少一部分,例如包括一个或多个点突变,或隆突或穴突变。在这种情况下,fc区包含fc链的异二聚体,其特征在于组装两个不同的ch3结构域。
特定的隆突突变是一个或多个氨基酸取代以通过掺入一个或多个氨基酸来增加两个结构域之间的接触表面,所述氨基酸提供β链结构的额外突起,例如,选自由t366y、t366w、t394w、f405a组成的组中的一种或多种ch3隆突突变。特定的隆突修饰表示抗体的ch3结构域中的突变t366w(根据eu的kabat索引编号)。隆突突变特异性地提供匹配(同源)表面以结合另一抗体结构域,例如,其经修饰以掺入穴突变。
特定的穴突变是一个或多个氨基酸取代以通过掺入一个或多个氨基酸来增加两个结构域之间的接触表面,所述氨基酸提供β链结构的额外洞穴,例如,选自由t366s、l368a和y407v组成的组中的一种或多种ch3穴突变。特定的穴-修饰表示抗体的ch3结构域中的任何突变t366s、l368a、y407v、y407t(根据eu的kabat索引编号)。穴突变特异性地提供匹配(同源)表面以结合另一抗体结构域,例如,其经修饰以掺入隆突突变。
将隆突匹配到穴突变中,是例如,一个ch3结构域上的t366y和ch3结构域对的第二ch3结构域上的匹配的y407t,在本文中称为t366y/y407’t。进一步匹配的突变是
t366y/y407’t、
f405a/t394’w、
t366y:f405a/t394’w:y407’t、
t366w/y407’a,和/或
s354c:t366w/y349’c:t366’s:l368’a:y407’v。
特异性ch3突变包括分子间β-链交换,例如,其中ch3β链内的一个或多个区段或序列被突变以掺入与原始ch3结构域不同的抗体结构域的区段或序列,例如,不同类型或亚型的抗体结构域。通过链交换获得特异性突变体,其中igg型的ch3结构域掺入iga型的ch3结构域的一个或多个区段或序列。如果两个链交换的ch3结构域突变形成同源对,则每个ch3结构域的iga区段或序列产生同源的域间接触表面,使得突变的ch3结构域优选在野生型ch3结构域上彼此配对。掺入区段交换的抗体结构域的这种修饰的具体实例可以是链交换工程化结构域(seed)。通过优选配对重链的seed修饰的ch3结构域,这些修饰可用于产生不对称或双特异性抗体。这基于在保守的ch3结构域内交换结构相关的序列。来自seedch3结构域中的人iga和igg的交替序列产生两个不对称但互补的结构域,称为ag和ga。seed设计允许有效产生ag/ga异二聚体,同时不利于ag和gaseedch3结构域的同源二聚化。
通过域内或域间二硫桥可进一步支持抗体结构域或lc/hc或fc链的连接。二硫键通常由两个半胱氨酸的硫醇基团的氧化形成,从而连接s原子以在两个半胱氨酸残基之间形成二硫桥。
根据一具体实施方案,抗体结构域包括掺入半胱氨酸残基的突变,该半胱氨酸残基能够形成二硫桥以通过另外的域内二硫桥稳定抗体结构域,或通过另外的域间二硫桥形成一对抗体结构域。具体地,半胱氨酸可以在ch3结构域的c-端区域或c-端插入(通过另外的氨基酸或氨基酸取代)。通过ch3对之间的二硫键形成,可以稳定带有额外半胱氨酸修饰的一对ch3,从而产生ch3/ch3二聚体。在一些实施方案中,二硫连接的抗体结构域是同二聚体或异二聚体,因此是相同或不同结构域的对。
为了允许抗体链或结构域的正确配对,可以特别使用任何ch3突变,例如,可以使用隆突-入-穴技术、seed技术、电荷排斥技术、二硫连接或交叉mab技术来减少不正确相关分子的数量。
“一对”抗体结构域在本文中理解为一组两个抗体结构域,其中一个抗体结构域在其表面上或在空腔中具有与另一个区域上的区域特异性结合并因此互补的区域。抗体结构域可以通过β-片区域的接触而结合并组装以形成一对抗体结构域。这种结构域对也称为二聚体,其例如通过静电相互作用、重组融合或共价连接相缔合,将两个结构域置于直接物理缔合中,例如,包括固体和液体形式。本文具体描述的是cl/ch1二聚体,其可以是通过本文鉴定的位置处的某些点突变的优选的同源抗体结构域对。
“优选配对”在本文中理解为抗体结构域或抗体链的二聚体的形成,从而获得抗体结构域或抗体链对,其中该对通过抗体结构域的结合界面的增加的亲和力或亲合力和结构域对或hc/lc对的增加的(热)稳定性形成。同源抗体结构域可以通过界面区域中的修饰产生,例如本文所描述的,其优选地在相同类型的任何野生型结构域上彼此配对。
在一对抗体结构域中,抗体结构域在本文中称为“对应”结构域。在本文所述的抗体中,以下结构域被认为是适合形成一对抗体结构域的对应物(通过斜线(/)分开的对应物):
vl/vh;
cl(cλ或cκ)/ch1;
ch2/ch2;
ch3/ch3。
关于一对结构域或结构域二聚体的术语“同源”理解为具有匹配的结合点或结构的结构域,以在每个结构域上获得接触表面以优选形成一对这样的结构域。如果修饰至少一个结构域以优选结合其同源(对应)结合配偶体以产生结构域对,则将特定结构域理解为“同源”或同源结构域对。优选地,两个同源结构域都被工程化以掺入匹配的突变,例如,突变引入相反极性的氨基酸残基,隆突-入-穴突变、seed突变、用于二硫桥形成的额外半胱氨酸残基,或采用电荷排斥技术的修饰。
关于如本文所述的abm或抗体的术语“多价”是指具有至少两个结合位点以结合相同靶抗原的分子,特异性结合这种靶抗原的相同或不同表位。该术语应包括二价抗体或具有2个或更多个结合靶抗原的化合价的分子,例如,通过至少2、3、4或甚至更多的结合位点。例如,二价抗体可以通过两对vh/vl结构域具有两个抗原结合位点,两者都结合相同的靶抗原。
关于如本文所述的abm或抗体的术语“多特异性”应指具有至少两个特异性结合至少两种不同靶抗原的结合位点的分子。该术语应包括双特异性抗体或具有2种或更多种特异性的分子,以结合多于一种靶抗原,例如,通过至少2、3、4或甚至更多的结合位点。
例如,双特异性抗体可以通过一对vh/vl结构域(fv区)结合一种靶抗原,且通过第二对vh/vl结构域(fv区)结合另一种靶抗原。双特异性抗体通常由四种不同的抗体链组成,即两种hc和两种lc,使得通过具有第一lc的第一hc和具有第二lc的第二hc的异二聚化(配对)形成两个不同的cdr结合位点。
如本文所用,术语“抗原”或“靶标”应特别包括能够被抗体的结合位点(也称为互补位)识别的所有抗原和靶分子。如本文所描述的,结合分子所靶向的特别优选的抗原是已经证明具有或能够具有免疫学或治疗相关性的那些抗原,尤其是已经测试了临床功效的那些抗原。如本文所用,术语“靶”或“抗原”应特别包括选自(人或其他动物)肿瘤相关受体和可溶性肿瘤相关抗原的分子,它们是自身抗原,例如位于肿瘤细胞表面的受体或大量存在于癌症患者的循环中并与此类肿瘤相关的细胞因子或生长因子。其他抗原可能是病原体来源,例如微生物或病毒病原体。
靶抗原被识别为整个靶分子或这种分子的片段,尤其是子结构,例如,靶的多肽或碳水化合物结构,通常称为“表位”,例如b细胞表位、t细胞表位,其是免疫学相关的,即也可被天然或单克隆抗体识别。如本文所用,术语“表位”应特别指分子结构,其可完全构成特异性结合配偶体或是如本文所述的abm或抗体的结合位点的特异性结合配偶体的一部分。术语表位也可以指半抗原。在化学上,表位可以由碳水化合物,肽,脂肪酸,有机、生物化学或无机物质或它们的衍生物及其任何组合组成。如果表位是多肽,则其在肽中通常包括至少3个氨基酸,优选8至50个氨基酸,且更优选为约10-20个氨基酸。肽的长度没有严格的上限,其几乎可以包含蛋白质多肽序列的全长。表位可以是线性或构象表位。线性表位由多肽或碳水化合物链的一级序列的单个区段组成。线性表位可以是连续的或重叠的。构象表位由通过折叠多肽以形成三级结构而聚集在一起的氨基酸或碳水化合物组成,并且氨基酸不一定在线性序列中彼此相邻。具体地,表位至少是诊断相关分子的一部分,即样品中表位的缺失或存在与疾病或患者的健康状况或制造过程中的过程状态或环境和食物的状况定性或定量相关。表位也可以是治疗相关分子的至少一部分,即可以通过改变疾病进程的特异性结合结构域靶向的分子。
具体实施方案涉及天然存在的抗原或表位,或表位的合成(人工)抗原。作为天然存在的抗原的衍生物的人工抗原可具有增加的抗原性或稳定性的优点,这与被认为是特异性abm或抗体的结合配偶体有关。
如本文所用,术语“特异性”或“特异性结合”是指结合反应,所述结合反应决定异源分子群中的目标同源配体。因此,在指定条件下(例如,免疫测定条件),本文所述的abm或抗体结合其特定靶标,并且不会以显著量结合到样品中存在的其他分子。特异性结合意指结合在选择的靶标识别,高、中或低结合亲和力或亲合力方面是选择性的。如果结合常数或结合动力学至少10倍差异,通常实现选择性结合,优选地差异为至少100倍,且更优选为至少1000倍。
如本文所用,术语“可变结合区”也称为“cdr区”是指具有能够结合与抗原相互作用的不同结构的分子。这些分子可以原样使用或整合在较大的蛋白质中,从而形成具有结合功能的这种蛋白质的特定区域。不同的结构可以衍生自结合蛋白的天然库,例如衍生自免疫球蛋白或抗体。变化的结构也可以通过随机化技术产生,特别是本文所述的那些。这些包括诱变的cdr或非cdr区(例如,恒定抗体结构域的结构环区),抗体可变结构域或恒定结构域的环区,特别是抗体的cdr环。通常,本文所述的abm或抗体的结合结构由这种可变结合区形成。
用于本文所述的abm或抗体的术语“细胞毒性”或“细胞毒活性”应指任何针对细胞抗原的特定分子,当与抗原结合时,其激活程序性细胞死亡并引发细胞凋亡。特异性抗体通过其对效应细胞的活性而有效,导致细胞毒性t细胞或介导抗体依赖性细胞毒性(adcc)、补体依赖性细胞毒性(cdc)和/或细胞吞噬作用(adcp)的细胞的激活。特异性抗体通过细胞凋亡诱导程序性细胞死亡和/或通过结合介导adcc和/或cdc活性的效应细胞的fc受体来杀死抗体包被的靶细胞。
本文描述的abm或抗体可以表现出或可以不表现出fc效应子功能。fc可以通过形成免疫复合物结合表面抗原来募集补体并帮助消除靶抗原或靶细胞。
特异性抗体可以缺乏活性fc部分或fc效应功能,因此,由不含抗体fc部分或不含fcγ受体结合位点的抗体结构域组成,或者包含缺乏fc效应功能的抗体结构域,例如通过修饰来减少fc效应功能,特别是消除或减少adcc和/或cdc活性。可以工程化替代抗体以掺入修饰来增加fc效应功能,特别是增强adcc和/或cdc活性。
这些修饰可以通过诱变实现,例如,fcγ受体结合位点的突变或通过衍生物或试剂以干扰抗体形式的adcc和/或cdc活性,从而实现fc效应功能的降低或增加。
术语“抗原结合位点”或“结合位点”是指参与抗原结合的abm或抗体的部分。抗体的抗原结合位点通常由重链(“h”)和/或轻链(“l”)的n-端可变区(“v”)或其可变结构域的氨基酸残基形成。重链和轻链的v区内的三个高度发散的延伸(称为“高变区”)介于称为框架区的更保守的侧翼延伸之间。抗原结合位点提供与结合的表位或抗原的三维表面互补的表面,并且高变区称为“互补决定区”或“cdr”。结合在cdr中的结合位点在本文中也称为“cdr结合位点”。
术语“表达”以下列方式理解。含有表达产物的所需编码序列的核酸分子,诸如例如,如本文所述的abm或抗体,和控制序列,诸如例如可操作连接的启动子可用于表达目的。用这些序列转化或转染的宿主能够产生编码的蛋白质。为了实现转化,表达系统可以包含在载体中;但是,相关的dna也可以整合到宿主染色体中。具体地,所述术语是指在合适条件下的宿主细胞和相容载体,例如,用于表达由载体携带的并导入宿主细胞的外源dna编码的蛋白质。
编码dna是编码特定多肽或诸如例如抗体的蛋白质的特定氨基酸序列的dna序列。启动子dna是启动、调节或以其他方式介导或控制编码dna的表达的dna序列。启动子dna和编码dna可以来自相同基因或来自不同基因,并且可以来自相同或不同的生物体。重组克隆载体通常包括一个或多个用于克隆或表达的复制系统,一个或多个用于在宿主中选择的标记,例如,抗生素抗性,和一个或多个表达盒。
本文使用的“载体”定义为克隆的重组核苷酸序列的转录所需的dna序列,即重组基因的转录和它们在合适的宿主生物体中的转译。
“表达盒”是指编码表达产物的dna编码序列或dna区段,所述表达产物可以在限定的限制性位点插入到载体中。盒式限制位点设计成确保盒插入适当的阅读框架中。通常,外源dna插入载体dna的一个或多个限制性位点,然后连同可传递的载体dna由载体携带进入宿主细胞中。具有插入或添加的dna的片段或序列,例如表达载体,也可称为“dna构建体”。
表达载体包含表达盒,并且另外通常包含在宿主细胞或基因组整合位点中自主复制的起点,一种或多种可选择的标记(例如,氨基酸合成基因或赋予抗生素抗性的基因,例如博来霉素、卡那霉素、g418或潮霉素),许多限制酶切割位点,合适的启动子序列和转录终止子,这些组分可操作地连接在一起。如本文所用,术语“载体”包括自主复制的核苷酸序列以及基因组整合核苷酸序列。常见类型的载体是“质粒”,其通常是双链dna的自含分子,其可以容易地接受另外的(外源)dna并且其可以容易地引入合适的宿主细胞中。质粒载体通常含有编码dna和启动子dna,并具有一个或多个适于插入外源dna的限制性位点。具体地,术语“载体”或“质粒”是指可以将dna或rna序列(例如外源基因)引入宿主细胞的载体,以转化宿主并促进引入序列的表达(例如,转录和转译)。
如本文所用,术语“宿主细胞”应指转化以产生特定重组蛋白的原代受试细胞,例如本文所述的abm或抗体,及其任何后代。应该理解的是,并非所有后代都与亲本细胞完全相同(由于故意或无意的突变或环境差异),但是,这些改变后代包括在这些术语中,只要后代保留与最初转化的细胞相同的功能即可。术语“宿主细胞系”是指宿主细胞的细胞系,用于表达重组基因以产生重组多肽,例如重组抗体。如本文所用,术语“细胞系”是指已经获得长时间增殖能力的特定细胞类型的已建立克隆。可以将这种宿主细胞或宿主细胞系维持在细胞培养物中和/或培养以产生重组多肽。
如本文所用,关于核酸、抗体或其他化合物的术语“分离的”或“分离”应指与其天然缔合的环境充分分离的这种化合物,以便以“基本纯粹的”形式存在。“分离的”不一定意味着将人工或合成混合物与其他化合物或材料排除,或者存在不干扰基本活性的杂质,并且可能存在,例如,由于不完全的纯化。特别地,编码本文所述的abm或抗体的分离的核酸分子也意味着包括天然存在的核酸序列的密码子优化的变体以改善某些宿主细胞中的表达,或化学合成的那些。
关于核酸,有时使用术语“分离的核酸”。当应用于dna时,该术语是指与其中其起源的生物体的天然存在的基因组中与其立即邻接的序列分离的dna分子。例如,“分离的核酸”可以包含插入载体(例如,质粒或病毒载体)中的dna分子,或整合到原核或真核细胞或宿主生物的基因组dna中。当应用于rna时,术语“分离的核酸”主要指由如上定义的分离的dna分子编码的rna分子。或者,该术语可以指已经从以其天然状态(即,在细胞或组织中)与之缔合的其他核酸充分分离的rna分子。“分离的核酸”(dna或rna)可以进一步代表通过生物或合成方法直接产生的分子,并与其产生过程中存在的其他组分分离。
关于多肽或蛋白质,例如分离的抗体,术语“分离的”应特别指不含或基本上不含与它们天然缔合的物质的化合物,例如在其天然环境中发现它们的其他化合物,或当这种制备是通过体外或体内实施的重组dna技术时,其中制备它们的环境(例如,细胞培养)。分离的化合物可以与稀释剂或佐剂一起配制,并且仍然为了实际目的而分离-例如,当用于诊断或治疗时,多肽或多核苷酸可以与药学上可接受的载体或赋形剂混合。
如本文所用,术语“重组”应意指“由基因工程制备或由基因工程产生的结果”。或者,使用术语“工程化”。例如,可以通过工程化相应的亲本序列来工程化抗体或抗体结构域以产生修饰的抗体或结构域来产生变体。重组宿主特异性地包含表达载体或克隆载体,或者它已经过基因工程化以含有重组核酸序列,特别是使用宿主外源的核苷酸序列。通过在宿主中表达相应的重组核酸来产生重组蛋白。如本文所用,术语“重组抗体”包括通过重组方法制备、表达、产生或分离的抗体,例如(a)从动物(例如小鼠)分离的抗体,其是人免疫球蛋白基因或由其制备的杂交瘤的转基因或转染色体,(b)从转化以表达抗体的宿主细胞分离的抗体,例如来自转染瘤,(c)从重组,组合人抗体文库分离的抗体,和(d)通过涉及将人免疫球蛋白基因序列剪接到其他dna序列的任何其他方式制备、表达、产生或分离的抗体。此类重组抗体包含经工程化以包括重排和突变的抗体,所述重排和突变例如在抗体成熟过程中发生。
一旦鉴定出具有所需结构的抗体,就可以通过本领域熟知的方法产生这种抗体,包括例如杂交瘤技术或重组dna技术。
在杂交瘤方法中,免疫小鼠或其他合适的宿主动物,例如仓鼠,以引发产生或能够产生特异性结合用于免疫的蛋白质的抗体的淋巴细胞。或者,可以在体外免疫淋巴细胞。然后使用合适的融合剂(例如聚乙二醇)将淋巴细胞与骨髓瘤细胞融合,以形成杂交瘤细胞。
测定在其中生长杂交瘤细胞的培养基以产生针对抗原的单克隆抗体。优选地,由杂交瘤细胞产生的单克隆抗体的结合特异性通过免疫沉淀或通过体外结合测定法测定,例如放射免疫测定法(ria)或酶联免疫吸附测定法(elisa)。
例如,重组单克隆抗体可以通过分离编码所需抗体链的dna并用进行表达的编码序列转染重组宿主细胞来产生,使用公知的重组表达载体,例如包含编码本文所述的abm或抗体的核苷酸序列的质粒或表达盒。重组宿主细胞可以是原核和真核细胞,例如上述那些。
根据具体方面,核苷酸序列可用于遗传操作以使抗体人源化或改善抗体的亲和力或其他特征。例如,如果抗体用于人类的临床试验和治疗,则恒定区可以工程化为更接近人类恒定区以避免免疫应答。可能需要遗传操作抗体序列以获得对靶抗原的更大亲和力。对于本领域技术人员显而易见的是,可以对抗体进行一种或多种多核苷酸改变,并仍然保持其对靶抗原的结合能力。
通过各种方法产生抗体分子通常是很好理解的。例如,美国专利6331415(cabilly等人)描述了重组产生抗体的方法,其中重链和轻链同时从单个载体或单个细胞中的两个单独载体表达。wibbenmeyer等人,(1999,biochimbiophysacta1430(2):191-202)和lee和kwak(2003,j.biotechnology101:189-198)描述了使用在单独的大肠杆菌培养物中表达的质粒由单独产生的重链和轻链生产单克隆抗体。在例如harlow等人,antibodies:alaboratorymanual,coldspringharborlaboratorypress,coldspringharbor,n.y.,(1988)中提供了与抗体产生相关的各种其他技术。
使用通过培养中的连续细胞系生产抗体分子的任何方法产生单克隆抗体。用于制备单克隆抗体的合适方法的实例包括kohler等人,(1975,nature256:495-497)的杂交瘤方法和(kozbor,1984,j.immunol.133:3001;和brodeur等人,1987,单克隆抗体生产技术与应用(monoclonalantibodyproductiontechniquesandapplications)(marceldekker,inc.,newyork),pp.51-63)的人b细胞杂交瘤方法。
如本文所述的abm或抗体可用于施用以治疗有此需要的受试者。
如本文所用,术语“受试者”应是指温血哺乳动物,特别是人类或非人动物。因此,术语“受试者”还可特别指动物,包括狗、猫、兔、马、牛、猪和家禽。特别地,本文所述的abm或抗体被提供用于医学用途,以治疗需要预防或治疗疾病状况的受试者或患者。术语“患者”包括接受预防性或治疗性治疗的人和其他哺乳动物受试者。因此,术语“治疗”意味着包括预防性和治疗性治疗。
具体地,本文所述的abm或抗体以基本上纯的形式提供。如本文所用,术语“基本上纯的”或“纯化的”是指包含至少50%(w/w),优选至少60%、70%、80%、90%或95%的化合物的制剂,例如核酸分子或抗体。通过适合于化合物的方法测量纯度(例如,色谱法、聚丙烯酰胺凝胶电泳、hplc分析等)。
术语“治疗有效量”,在本文中可与化合物的任何术语“有效量”或“足够量”互换使用,例如,本文所述的abm或抗体是足以在向受试者施用有益或期望结果(包括临床结果)时的量或活性,并且因此,其有效量或同义词取决于其中使用它所处的背景。
有效量旨在表示足以治疗、预防或抑制此类疾病或病症的化合物的量。在疾病的背景下,如本文所述的治疗有效量的abm或抗体特别用于治疗、调节、减弱、逆转或影响受益于抗体与其靶抗原相互作用的疾病或病症。
对应于这种有效量的化合物的量将根据各种因素而变化,例如给定的药物或化合物、药物制剂、施用途径、疾病或病症的类型、受治疗的受试者或宿主的身份等,但仍可由本领域技术人员常规确定。
本文所述的abm或抗体可特异性地用于药物组合物中。因此,提供了药物组合物,其包含如本文所述的abm或抗体和药学上可接受的载体或赋形剂,例如,人造载体或赋形剂,其在体液中不与免疫球蛋白一起天然存在,或者其与免疫球蛋白一起天然存在,但在含有不同量或比例的载体或赋形剂的制剂中提供。
本文所述的药物组合物可以作为推注注射或输注或通过连续输注施用。适于促进这种给药方式的药物载体在本领域中是众所周知的。
药学上可接受的载体通常包括与本文所述的abm或抗体在生理学上相容的任何和所有合适的固体或液体物质、溶剂、分散介质、包衣、等渗和吸收延迟剂等。药学上可接受的载体的其他实例包括无菌水、盐水、磷酸盐缓冲盐水、右旋糖、甘油、乙醇等,以及其任何组合。
在一个这样的方面,abm或抗体可以与适合所需施用途径的一种或多种载体组合。抗体可以,例如与乳糖、蔗糖、淀粉、烷酸的纤维素酯、硬脂酸、滑石、硬脂酸镁、氧化镁、磷酸和硫酸的钠盐和钙盐、阿拉伯胶、明胶、海藻酸钠、聚乙烯吡咯烷、聚乙烯醇的任一种混合,并且任选进一步压片或包封用于常规施用。或者,abm或抗体可以溶解在盐水、水、聚乙二醇、丙二醇、羧甲基纤维素胶体溶液、乙醇、玉米油、花生油、棉籽油、芝麻油、黄蓍胶和/或各种缓冲液中。其他载体、佐剂和施用方式在制药领域中是众所周知的。载体可包括控释材料或延时材料,例如单独或与蜡一起的单硬脂酸甘油酯或二硬脂酸甘油酯,或本领域熟知的其他材料。
另外的药学上可接受的载体是本领域已知的并且在例如,雷米顿的医药科学(remington’spharmaceuticalsciences)中有描述。液体制剂可以是溶液、乳液或悬浮液,并且可以包括赋形剂,例如悬浮剂、增溶剂、表面活性剂、防腐剂和螯合剂。
考虑了药物组合物,其中配制了本文所述的abm或抗体和一种或多种治疗活性剂。通过将所述具有所需纯度的abm或抗体与任选的药学上可接受的载体、赋形剂或稳定剂以冻干制剂或水溶液的形式混合,制备稳定的制剂用于储存。用于体内施用的制剂是特别无菌的,优选以无菌水溶液的形式。这可以通过无菌过滤膜或其他方法过滤而容易地完成。本文公开的abm或抗体和其他治疗活性剂也可以配制成免疫脂质体,和/或包埋在微胶囊中。
包含本文所述的abm或抗体的药物组合物的施用可以多种方式进行,包括口服、皮下、静脉内、鼻内、耳内(intraotically)、透皮、粘膜、局部,例如凝胶、药膏、洗剂、乳膏等、腹膜内、肌肉内、肺内、阴道、胃肠外、直肠或眼内。
用于肠胃外给药的示例性制剂包括适合于皮下、肌肉内或静脉内注射的那些,如例如无菌溶液、乳液或悬浮液。
本发明特别提供了如本文提供的实施例中详述的示例性abm和抗体。其他抗体变体是可行的,例如,包括示例性抗体的功能变体,例如,其中fc被进一步工程化以改善分子的结构和功能,或者其中产生包含不同cdr结合位点或具有不同特异性的抗体,特别是其中获得两个不同的fv区。
参考以下实施例将更全面地理解前面的描述。但是,这些实施例仅代表实施本发明的一个或多个实施方案的方法,不应理解为限制本发明的范围。
实施例
实施例1:b10v5xhu225mseed
描述了具有igg结构的双特异性抗体。b10v5-fab与人c-met结合,而hu225m-fab与人egfr(表皮生长因子受体)结合。hu225m轻链和hu225m重链之间的界面具有突变,其将双特异性igg的轻链指向其同源重链。抗体的ch3结构域被seed结构域取代(称为seed-ag或seed-ga,davis等人,2010和us20070287170a1)以实施重链的异二聚化。lc-esi-ms分析用于确认所有四条链的正确组装。在下文中,术语bxm将用于描述该双特异性igg。
将所有随后的链分别克隆到载体ptt5(加拿大国家研究委员会)中以在哺乳动物系统中表达。
具有seed-ga的hu225m重链被称为hu225m_hc_ga(seqid6):
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
hu225m轻链被称为hu225m_lc(seqid8):
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
将具有seed-ag的b10v5重链称为b10v5_hc_ag(seqid9):
加下划线的:信号肽metdtlllwvlllwvpgstg(seqid10)
b10v5轻链称为b10v5_lc(seqid11):
加下划线的:信号肽metdtlllwvlllwvpgstg(seqid10)
将界面突变引入hu225m
根据制造商的方案,使用quikchangelightning定点诱变试剂盒(quikchangelightningsite-directedmutagenesiskit)(#210519,agilenttechnologies)通过定点诱变引入突变。将突变k26d引入hu225m_hc_ag的ch1中,并将突变t18r引入hu225m_lc的cl中。通过对目的基因进行测序证实了突变被成功引入。
具有突变k26d的hu225m重链被称为hu225m_hc_resq28_ga(seqid12):
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
具有突变t18r的hu225m轻链被称为hu225m_lc_mb40(seqid13)
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
bxm野生型和突变体bxmmab40在expi293fti细胞中的表达和纯化
根据制造商的方案,使用expi293ftm细胞和expifectaminetm293转染试剂盒(thermofisher,a14525)表达bxm。设置了两种不同的转染:
编码每条链的dna位于四种不同的质粒上。转染期间质粒的摩尔比为2:1:1:1(b10v5重链:b10v5轻链:hu225m重链:hu225m轻链)。
突变体bxmmab40在hu225m(hu225m_hc_resq28_ga,seqid12)的ch1上含有突变k26d,并且在hu225m(hu225m_lc_mb40,seqid13)的cl上含有t18r,而bxm野生型不含任何突变。将培养物旋转减慢,并将含有感兴趣的蛋白质的上清液通过0.22μm过滤器过滤,并使用montage抗体纯化试剂盒和使用prosep-a培养基的旋转柱(merck-millipore,lsk2aba20)根据制造商的说明书纯化。使用amiconultra-15,10kdamwco浓缩纯化的bxm野生型和bxmmab40,然后使用slide-a-赖氨酸透析盒(slide-a-lyserdialysiscassettes)0.5-3ml7,000mwco(thermofisher,#66370)对pbs进行透析。总之,bxm野生型和bxmmab40均被独立表达、纯化和分析两次。两次重复都导致了类似的结果。
lc-esi-ms分析以分析链配对
在用lc-esi-ms系统测量之前,使用pngasef释放两个样品的n-聚糖。计算所有十种可能的链配对变体的质量,并分析其存在的质谱。含有四条不同链但具有与其非同源重链结合的轻链(即具有交换轻链的bxm)的错配变体具有与正确组装的bxm相同的质量,因此不能与正确配对的bxm区分。但是,如果不能检测到一条或两条轻链与其非同源重链的错配,则可以统计地排除具有交换的轻链的bxm的存在。类似地,如果仅以少量(<5%相对丰度)检测到两条轻链的错配,则所述错配的变体将仅以可忽略的量(<1%)存在。
在任何样品中均未检测到重链同二聚体(图1)。在bxm野生型中,两条轻链能够以相似的量结合其非同源重链(在两种情况下相对丰度为12%),导致仅76%的正确配对bxm(不计算具有交换轻链的bxm)。在bxm中,mab40错配不可检测,仅检测到正确配对的双特异性igg。
总之,界面突变的引入导致轻链与重链错配的完全消失。
尺寸排阻色谱hplc(hplc-sec)
使用sec分析bxm野生型和bxmmab40。两个色谱图在1.56分钟均显示出主峰,这是igg预期的。在突变体和野生型中都可检测到聚集的迹象(图2)。
热位移分析以确定热稳定性
使用实时pcr系统步骤1+(steponeplus)进行热位移分析。bxm野生型和bxmmab40的浓度在pbs中为1μm,并且使用20×终浓度的染料铜染及橙染(invitrogen)。一式三份测量两种样品。bxm野生型的热分析图显示在64.8℃和74.5℃下有两个展开事件。bxmmab40的热分析图显示在64.6℃和74.6℃下发生两次展开事件。因此,界面突变不会损害蛋白质的热稳定性。
bxm对其抗原的亲和力
使用具有涂有蛋白a的生物传感器的octet系统分析bxm野生型和mab40的亲和力。作为抗原,使用cmet和egfr的细胞外结构域。测试三种不同浓度的抗体以确定亲和力。bxmwt对cmet的kd为0.35nm,对egfr为5.3nm。bxmmab40对cmet的kd为0.42nm,对egfr为2.9nm,证实界面突变不损害抗体的亲和力。
两种抗原的同时结合
使用具有链霉亲和素包被的生物传感器的octet系统证实bxm同时结合其两种抗原的能力。首先,将生物传感器浸没在含有生物素化的cmet的溶液中。在猝灭和缓冲液更换后,将生物传感器浸没在含有bxm野生型或bxmmab40的溶液中。在两种情况下,都检测到抗体与其第一抗原的结合。此后,将生物传感器浸没在含有egfr的溶液中,并检测野生型和mab40与第二抗原的结合。
界面突变对hek293-6e产率的影响
根据标准技术,使用聚乙烯亚胺(pei)瞬时转染,在hek293-6e细胞(加拿大国家研究委员会)中表达bxm野生型和bxmmab40。通过蛋白质a亲和层析纯化表达的igg,并对pbs透析。在280nm处测量两种蛋白质样品的吸光度以确定浓度。总之,将两种蛋白质独立地表达、纯化和测量三次,并计算平均产率。bxm野生型的产率为57.3mg/l(±13.7标准偏差),且bxmmab40的产率为58.9mg/l(±7.3标准偏差),表明界面工程化对蛋白质产率没有不利影响。
实施例2:b10v5xokt3seed
描述了具有与实施例1中所述相似的igg结构的双特异性抗体。b10v5-fab与人c-met结合,而okt3-fab与人cd3结合。okt3轻链和okt3重链之间的界面具有与实施例1中所述相同的突变。此外,引入b10v5-fab中的突变以进一步强化轻链与重链的正确配对。如上所述,seed技术应用于重链的异二聚化。lc-esi-ms分析用于确认所有四条链的正确组装。在下文中,术语bxo将用于描述该双特异性igg。
构建体的克隆
b10v5重链(seqid9)和轻链(seqid11)描述于实施例1中。
将所有随后的链分别克隆到载体ptt5(加拿大国家研究委员会)中以在哺乳动物系统中表达。
okt3重链被称为okt3_hc_ga(seqid14)
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
okt3轻链被称为okt3_lc(seqid15)
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
将界面突变引入okt3和b10v5
根据如上所述的制造商的方案,使用quikchangelightning定点诱变试剂盒(#210519,agilenttechnologies)通过定点诱变引入突变。在okt3中,引入ch1中的突变k26d和cl中的t18r。在b10v5中,引入ch1中的突变a20l和cl中的f7s、f7a或f7v。通过对目的基因进行测序证实了突变被成功引入。
具有突变k26d的okt3重链被称为okt3_hc_resq28_ga(seqid16)
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
具有突变t18r的okt3轻链被称为okt3_lc_mb40(seqid17)
加下划线的:信号肽mklpvrllvlmfwipasls(seqid7)
具有突变a20l的b10v5重链被称为b10v5_hc_resq203_ag(seqid18)
加下划线的:信号肽metdtlllwvlllwvpgstg(seqid10)
具有突变f7s的b10v5轻链被称为b10v5_lc_mb9(seqid19)
加下划线的:信号肽metdtlllwvlllwvpgstg(seqid10)
具有突变f7a的b10v5轻链被称为b10v5_lc_mb21(seqid20)
加下划线的:信号肽metdtlllwvlllwvpgstg(seqid10)
具有突变f7v的b10v5轻链被称为b10v5_lc_mb45(seqid21)
加下划线的:信号肽metdtlllwvlllwvpgstg(seqid10)
bxo野生型和突变体bxomab40、bxomab5/40、bxomab21/40和bxomab45/40在hek293-6e细胞中的表达和纯化
根据标准技术,使用聚乙烯亚胺(pei)瞬时转染,在hek293-6e细胞中表达双特异性抗体。设置了四种不同的转染:
编码每条链的dna位于四种不同的质粒上。转染期间质粒的摩尔比为2:1:1:1(b10v5重链:b10v5轻链:okt3重链:okt3轻链)。转染后5天通过离心收获培养物,并通过蛋白a亲和层析纯化上清液。针对pbs透析所有样品。
lc-esi-ms分析以分析链配对。
在测量之前,使用pngasef释放两个样品的n-聚糖。如实施例1中所述进行分析。界面突变的引入导致轻链与重链的可检测错配显著降低(图3)。
尺寸排阻色谱hplc(hplc-sec)
使用sec分析bxo野生型、bxomab5/40和bxomab45/40。所有色谱图显示15.5分钟的主峰,这是针对igg的预期(图4)。可以在突变体中检测到聚集的迹象以及类似量的野生型。
讨论
实施例1描述了具有称为bxm的igg结构的双特异性抗体的产生,在hu225mfab中具有或不具有突变。通过lc-esi-ms对bxm野生型的分析证明了产生双特异性igg的问题,而没有任何工程化来强化轻链与其同源重链的正确配对。两条轻链都能够与其非同源重链结合至24%的总量,这反过来意味着仅76%的纯化蛋白质样品是正确配对的双特异性igg。
为了产生突变体bxmmab40,仅引入两个点突变,ch1中的k26d和cl中的t18r,两者都在hu225mfab中。这些突变足以完全抑制不正确的轻-至-重链配对。与之前的报道相反(lewis等人,2014年,liu等人,2015),不需要对可变结构域进行工程化。因此,本发明的突变具有广泛适用于各种其他双特异性抗体的潜力。另外,省略可变结构域中的任何突变限制了影响抗体对其抗原的亲和力的风险。
进一步的研究表明,bxmmab40中引入的突变对热稳定性和蛋白质产率没有不利影响。此外,bxmmab40对其抗原的亲和力与bxm野生型相似,并且能够同时结合两种抗原,如通过生物层干涉测定所证明的。尺寸排阻色谱显示bxmmab40和bxm野生型之间没有差异,这证明突变不会导致抗体的聚集或降解增加。
为了评估鉴定的突变是否是通用的,如实施例2中所示构建了不同的双特异性抗体bxo。与实施例1类似,将没有突变的bxo(野生型)与具有ch1中的突变k26d的bxo和okt3fab(称为bxomab40)的cl中的t18r比较。通过lc-esi-ms分析bxo野生型显示轻链与其非同源重链的错配对比在bxm野生型中检测到的错配更普遍。总共,有超过40%的检测到的igg在fab中表现出错配,导致少于60%的正确配对的bxo。bxomab40的分析表明,上述突变的引入再次对轻链的配对行为具有相当大的影响。bxomab40中74%的检测到的igg正确配对。为了进一步实施链的正确组装,在bxo的另一个fab即b10v5fab中引入支持性突变,导致产生bxomab5/40、bxomab21/40和bxomab45/40。在含有支持性突变的所有三种突变体中,正确配对的双特异性igg的量得到极大改善(lc-esi-ms中相对丰度>90%)。
通过尺寸排阻色谱法分析bxo野生型、bxomab5/40和bxomab45/40表明本发明的突变对抗体的聚集或降解没有不利影响。
实施例3:在ch1和cl之间的界面处的位置中的氨基酸侧链的表面暴露
getarea程序(fraczkiewicz等人,1998,j.comp.chem.,19,319-333)用于计算溶剂可及的表面积或蛋白质的溶剂化能量。人igg1fab片段1dfb.pdb的原子坐标,其是针对具有野生型ch1和cl结构域的人免疫缺陷病毒1型的gp41的人单克隆抗体fab片段(he等人,1992,proc.natl.acad.sci.usa89:7154-7158),作为输入提供给程序。应用1.4埃的探针半径。ch1结构域中残基ala20和lys26以及cl结构域中phe7和thr18的程序输出显示在下表i中。
骨架和侧链原子的贡献分别列在第4和第5列中。下一列列出了侧链表面积与每个残基的“无规卷曲(randomcoil)”值的比。残基x的“无规卷曲”值是30个无规构象的整体中三肽gly-x-gly中x的平均溶剂可及表面积。如果比值超过50%则认为残留物是溶剂暴露的,如果该比例小于20%则被埋入,并且如果该比例为至少20%则不被埋入。20个氨基酸的“无规卷曲”值列于下表ii中。
表1:用于chi和cl之间界面处的位置中的氨基酸侧链的表面暴露的getarea输出。根据imgt编号
表ii:20个氨基酸的无规卷曲值
从上表i中所示的结果可以看出,四个残基中的三个被埋入(比(%)值小于5%),而cl结构域中的残基thr18具有38.3的比(%)值,因此不被埋入。
令人惊讶的是,发现cl结构域中的突变位置thr18(其未埋入在ch/ch1界面内),导致cl/ch1结构域的配对改善。当cl结构域与ch1结构域配对时特别发现改进的配对,其中cl结构域中18位的氨基酸残基与ch1结构域中26位的氨基酸残基具有相反的极性,如本文进一步描述的。
这更令人惊讶,因为现有技术的工程化方法应用某些标准来选择沿重链和轻链界面的残基对,以被具有相反极性的带电残基取代。根据现有技术的这种标准(例如,liu等人,journalofbiologicalchemistry2015,290:7535-7562;和wo2014/081955a1),认为所有位置都被埋入是必要的。
因此,cl结构域中的18位是该现有技术规则的例外,并且令人惊讶地有助于cl/ch1结构域对的稳定性,尽管没有被埋入在ch1和cl之间的界面内。
- 还没有人留言评论。精彩留言会获得点赞!