转铁蛋白受体转基因模型的制作方法
血脑屏障(bbb)阻止大多数大分子从外周进入脑,并因此限制了需要脑暴露的大分子治疗剂的使用。转铁蛋白受体(tfr)在bbb处高度表达,并且可以用于经由受体介导的转胞吞作用将此类治疗剂穿过bbb转运。先前已经开发了小鼠模型,其中小鼠tfr被全长人tfrcdna取代,目的是评估潜在治疗剂穿过bbb的能力。然而,这些转基因小鼠是不健康的并且表现出异常高的tfr表达,低红血细胞计数和高血清铁浓度。yu等人,sciencetrans.med.,6(261):261ra154(2014)。结果,这些现有的小鼠模型不适合用作评估能够穿过bbb治疗脑疾病的治疗剂的工具;需要更能代表内源性tfr表达和表型的模型。
发明概述
在一个方面,本公开提供了包含编码嵌合转铁蛋白受体(tfr)多肽的核酸序列的多核苷酸,所述嵌合转铁蛋白受体(tfr)多肽包含非人哺乳动物转铁蛋白结合位点和具有与seqidno:1至少80%相同的氨基酸序列的异源性顶端结构域。在一些实施方案中,异源性顶端结构域包含seqidno:1的氨基酸序列。在一些实施方案中,异源性顶端结构域包含seqidno:7、seqidno:8或seqidno:9的氨基酸序列。
在一些实施方案中,非人哺乳动物转铁蛋白结合位点是天然的(例如,来自与tfr的跨膜和/或细胞内结构域相同的物种)转铁蛋白结合位点,例如天然小鼠转铁蛋白结合位点。在一些实施方案中,嵌合tfr多肽与seqidno:3具有至少80%的氨基酸序列同一性、或至少85%、90%或95%的同一性。在一些实施方案中,嵌合tfr多肽包含seqidno:3的氨基酸序列。在一些实施方案中,编码嵌合tfr多肽的异源性顶端结构域的核酸序列的区域与seqidno:2具有至少70%的核苷酸序列同一性。在一些实施方案中,编码嵌合tfr多肽的异源性顶端结构域的核酸序列的区域包含seqidno:2的核苷酸序列。在一些实施方案中,编码嵌合tfr的多核苷酸包含小鼠转铁蛋白受体基因的外显子和内含子,并且编码异源性顶端结构域的核酸序列位于小鼠转铁蛋白受体基因的第四外显子之后,以取代小鼠转铁蛋白受体基因的顶端结合结构域。
另一方面,本文提供了嵌合tfr多肽,其包含非人哺乳动物转铁蛋白结合位点和具有与seqidno:1至少80%相同的氨基酸序列的异源性顶端结构域。在一些实施方案中,嵌合tfr多肽包含天然tfr多肽,其中仅天然顶端结构域被异源性顶端结构域取代。在一些实施方案中,嵌合tfr多肽包含天然tfr结合位点和与天然tfr结合位点是异源的顶端结合结构域,例如,其中除顶端结构域之外,还有至少一个结构域或其区域包含非天然氨基酸序列。在一些实施方案中,异源性顶端结构域包含seqidno:1的氨基酸序列。在一些实施方案中,异源性顶端结构域包含seqidno:7、seqidno:8或seqidno:9的氨基酸序列。在一些实施方案中,嵌合tfr与seqidno:3具有至少80%、90%、95%或98%的氨基酸序列同一性。在一些实施方案中,嵌合tfr多肽包含seqidno:3的氨基酸序列。
在另一方面,本文提供了表达如上所述的嵌合转铁蛋白受体的宿主细胞。在一些实施方案中,宿主细胞包含编码嵌合转铁蛋白受体多肽的多核苷酸。在一些实施方案中,宿主细胞是小鼠细胞。在一些实施方案中,宿主细胞表达的嵌合tfr多肽包含(a)取代tfr多肽的内源性顶端结构域的异源性顶端结构域和(b)内源性转铁蛋白结合位点。在一些实施方案中,异源性顶端结构域具有与seqidno:1至少80%相同的氨基酸序列。在一些实施方案中,宿主细胞表达嵌合tfr,其中仅内源性tfr的顶端结构域被异源性顶端结构域取代。在一些实施方案中,宿主细胞表达包含内源性tfr结合位点和异源性顶端结构域的嵌合tfr,例如,其中除顶端结构域之外,还有至少一个结构域或其区域包含非天然氨基酸序列。在一些实施方案中,异源性顶端结构域包含seqidno:1的氨基酸序列。在一些实施方案中,编码细胞中异源性顶端结构域的核酸序列包含seqidno:2的核苷酸序列。在一些实施方案中,宿主细胞是小鼠细胞。在一些实施方案中,编码细胞中顶端结构域的核酸序列位于小鼠转铁蛋白受体基因的第四外显子之后。在一些实施方案中,宿主细胞是离体的。在一些实施方案中,宿主细胞是胚胎干细胞。在一些实施方案中,宿主细胞的基因组包含天然tfr的顶端结构域的缺失。
在另一方面,本公开提供了表达嵌合tfr多肽的非人转基因动物,其中嵌合tfr多肽包含异源性顶端结构域,其取代非人转基因动物内源性tfr多肽的顶端结构域。在一些实施方案中,非人转基因动物的基因组包含转铁蛋白受体基因,其编码取代非人转基因动物的内源性tfr的顶端结构域的异源性顶端结构域。在一些实施方案中,非人转基因动物表达嵌合tfr,其包含取代非人转基因动物的tfr的天然结构域的异源性顶端结构域和天然转铁蛋白结合位点。在一些实施方案中,非人转基因动物表达嵌合转铁蛋白受体,其中仅内源性转铁蛋白受体的顶端结构域被异源性顶端结构域取代。在一些实施方案中,非人转基因动物表达嵌合tfr多肽,其包含内源性tfr结合位点和与内源性tfr结合位点异源的顶端结合结构域,例如,其中除顶端结构域之外,还有至少一个结构域或其区域包含非天然氨基酸序列。在一些实施方案中,非人转基因动物包含如上所述的宿主细胞。在一些实施方案中,转基因动物是啮齿动物。在一些实施方案中,转基因动物是小鼠或大鼠。在一些实施方案中,转基因动物是嵌合tfr纯合的。在一些实施方案中,转基因动物是嵌合tfr杂合的。
在另一个方面,本文提供了筛选能与嵌合tfr结合的顶端结构域结合多肽(adbp)的方法,该方法包括使候选adbp与如上所述的嵌合tfr多肽接触;并确定与嵌合tfr多肽结合的候选adbp的量。在一些实施方案中,使候选adbp与嵌合tfr多肽接触的步骤包括使adbp与表达嵌合tfr多肽的宿主细胞接触。在一些实施方案中,使候选adbp与嵌合tfr多肽接触的步骤包括使adbp与表达嵌合tfr多肽的内皮细胞接触。在一些实施方案中,内皮细胞是血脑屏障内皮细胞。在一些实施方案中,通过免疫测定法确定结合嵌合tfr多肽的候选adbp的量。在一些实施方案中,通过表面等离子体共振法确定结合嵌合tfr多肽的候选adbp的量。在一些实施方案中,其中在体内进行接触步骤。在一些实施方案中,候选adbp与效应分子连接。在一些实施方案中,效应分子是小分子、rna、dna或多肽。在一些实施方案中,效应分子是多肽。在一些实施方案中,多肽是抗体或其抗原结合片段。
在另一个方面,本文提供了测量与嵌合tfr多肽结合的adbp的量的方法,该方法包括使adbp与上文公开的嵌合tfr多肽接触;并通过免疫测定法或表面等离子体共振法确定与嵌合tfr多肽结合的adbp的量。
在另一个方面,本文提供了筛选能穿过血脑屏障的adbp的方法,该方法包括:(a)向如本文所公开的非人转基因动物施用能结合顶端结构域的adbp,所述顶端结构域与seqidno:1具有至少80%的氨基酸序列同一性;和(b)测量非人转基因动物的脑中adbp的存在或活性。在一些实施方案中,adbp与效应分子连接。在一些实施方案中,效应分子是小分子、rna、dna或多肽。在一些实施方案中,多肽是抗体或其抗原结合片段。在一些实施方案中,确定步骤包括进行定量免疫测定法。在一些实施方案中,测量步骤包括使动物的脑或脑组织与能结合效应分子的试剂接触,以确定脑中效应分子的水平。在一些实施方案中,测量步骤包括测量效应分子的药效动力学(pd)效应。在一些实施方案中,效应分子是抗bace1抗体或其抗原结合片段,并且测量步骤包括测量脑中可溶性aβ40的水平。在一些实施方案中,效应分子是能结合脑中的靶标的抗体或其抗原结合片段。
在另一个方面,本文提供了监测能穿过血脑屏障的adbp的方法,该方法包括:(a)向如本文所公开的非人转基因动物施用能结合顶端结构域的adbp,所述顶端结构域与seqidno:1具有至少80%的氨基酸序列同一性;和(b)测量非人转基因动物的脑中adbp的存在或活性。在一些实施方案中,adbp与效应分子连接。在一些实施方案中,效应分子是小分子、rna、dna或多肽。在一些实施方案中,多肽是抗体或其抗原结合片段。在一些实施方案中,确定步骤包括进行定量免疫测定法。在一些实施方案中,确定步骤包括使效应分子与能结合效应分子的试剂接触,并确定脑中存在的效应分子的水平。在一些实施方案中,效应分子是能结合脑中的靶标的抗体或其抗原结合片段。在一些实施方案中,测量步骤包括测量与靶标的结合的效应分子的pd效应。在一些实施方案中,效应分子是抗bace1抗体或其抗原结合片段,并且测量步骤包括测量脑中可溶性aβ40的水平。
在另一个方面,本文提供了产生能表达嵌合转铁蛋白受体(tfr)多肽的转基因非人单细胞胚胎的方法,该方法包括用与seqidno:1具有至少80%同一性的异源性顶端结构域取代非人单细胞胚胎中内源性tfr的顶端结构域。在一些实施方案中,通过同源重组进行顶端结构域的取代。在一些实施方案中,该方法包括接触cas9蛋白、至少一种单向导rna(sgrna)和包含编码异源性顶端结构域的核酸序列的供体dna,其中异源性顶端结构域的侧翼为左同源臂和右同源臂,使得异源性顶端结构域的编码序列取代非人单细胞胚胎的基因组中内源性tfr的顶端结构域。在一些实施方案中,异源性顶端结构域是为在非人单细胞胚胎中表达而密码子优化的。在一些实施方案中,非人单细胞胚胎是小鼠胚胎。在一些实施方案中,供体dna位于小鼠转铁蛋白受体基因的第四外显子之后。
在另一方面,本文提供了产生非人转基因动物的方法,其包括(a)将上文所公开的转基因非人单细胞胚转移至与非人单细胞相同动物物种的假孕雌性动物,和(b)从雌性动物产生的后代中选择非人转基因动物,其中非人转基因动物包含嵌合转铁蛋白受体(tfr)多肽,其中内源性tfr的顶端结构域已被具有与seqidno:1至少80%同一的氨基酸序列的异源性顶端替换结构域取代。
在另一个方面,本文提供了产生能表达嵌合转铁蛋白受体(tfr)多肽的非人转基因动物的方法,该方法包括:(a)将编码与seqidno:1具有至少80%同一性的顶端结构域的多核苷酸引入动物的胚胎细胞中,其中多核苷酸靶向内源性tfr基因的编码内源性tfr顶端结构域的区域,并且其中编码与seqidno:1具有至少80%同一性的顶端结构域的多核苷酸取代内源性tfr基因的编码内源性顶端结构域的区域,和(b)使所述细胞或其子代发育成非人转基因动物。
前面的一般性描述和以下的详细描述是示例性和说明性的,并且旨在提供对所要求保护的本发明的进一步说明。从以下对本发明的详细描述,本领域技术人员将容易明白其他目的、优点和新特征。
附图简述
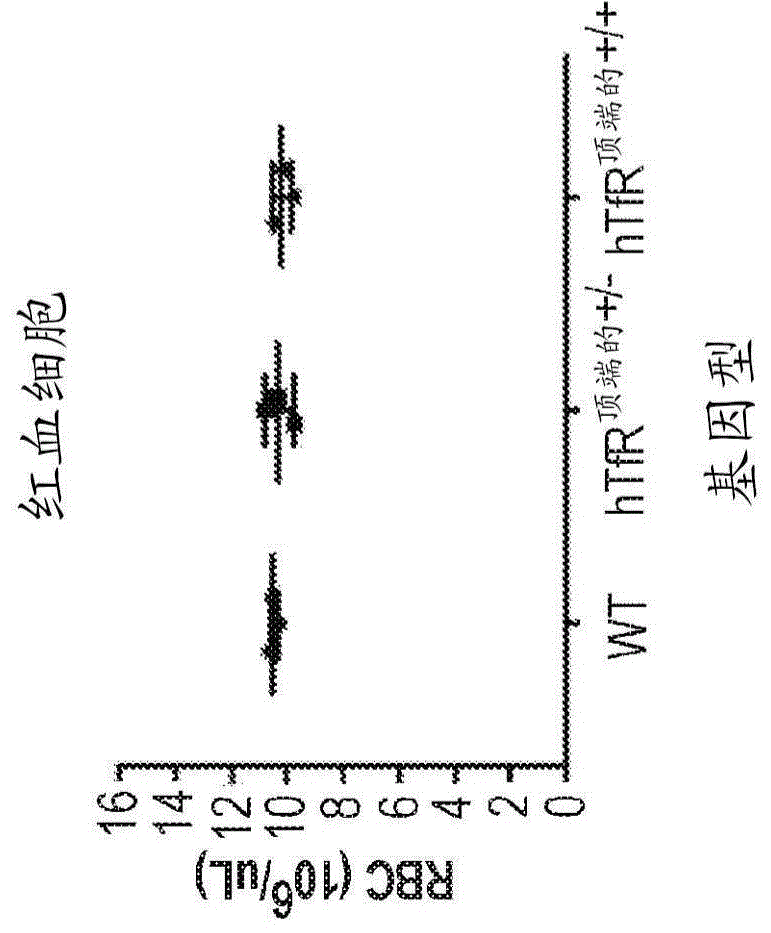
图1a-1c显示了野生型、hutfr顶端的+/-和hutfr顶端的+/+小鼠的全血细胞计数分析的结果。在总的红血细胞、血红蛋白或血细胞比容中未观察到基因型特异性差异。附图中的图表示平均值±sd,每组n=3。
图2a-2b显示全身施用的抗tfr在hutfr顶端的+/-小鼠中的脑血管定位。将5mg/kg人顶端特异性抗tfr静脉内施用于c57b16野生型或嵌合hutfr顶端的+/-杂合小鼠中。一小时后,用pbs灌注小鼠,并对脑进行抗体分布染色。代表性图像显示在hutfr顶端的+/-杂合小鼠,而不是野生型小鼠中全身注射的抗tfr(顶端结构域特异性的)的显著的血管定位。这表明嵌合hutfr蛋白在bbb处被表达。
图3a-3b显示了全身施用的抗tfr/bace1在hutfr顶端的+/+小鼠中的脑实质分布。来自hutfr小鼠的脑皮质切片的代表性图像显示静脉内注射的抗tfr/bace1(50mg/kg,给药后24hrs)的广泛实质分布。相反,在来自注射抗bace1的hutfr顶端的+/+hutfr小鼠的脑切片中未观察到明显的染色。
图4a-4d显示了在hutfr顶端的+/+小鼠中施用抗-tfr/bace1双特异性抗体后,该双特异性抗体的脑摄取和a-β的减少。图4a显示给小鼠施用50mg/kg抗tfr/bace1或抗bace1后24小时,hutfr顶端的+/+小鼠中的血浆huigg1浓度。结果显示,相对于抗bace1的清除,tfr介导的抗tfr/bace1的清除增强。图4b显示全身给予抗体后抗tfr/bace1的平均脑摄取。结果显示,与小鼠脑中的抗bace1的积聚相比,抗tfr/bace1的积聚增加约28倍。图4c显示与用抗bace1处理的小鼠相比,用抗tfr/bace1处理的小鼠中脑中的a-β减少了49%。图4d显示与未处理的野生型小鼠相比,已经用抗bace1或抗tfr/bace1处理的hutfr顶端的+/+小鼠中血浆a-β水平降低。所有图代表平均值±sd,每组n=8(对于未处理的野生型小鼠,n=2)。
图5a-5d显示与野生型小鼠相比,hutfr顶端的+/+小鼠的各种组织中tfr的表达;没有观察到脑(图5a)、肝脏(图5b)、肾脏(图5c)和肺(图5d)中总tfr表达的显著差异。所有图代表平均值±sd,每组n=4-8。
发明详述
我们已经开发了嵌合形式的转铁蛋白受体,其包括非人(例如小鼠)哺乳动物转铁蛋白结合位点和与含有转铁蛋白结合位点的结构域异源的顶端结构域。这些嵌合受体可以在转基因动物中被表达,特别是在转铁蛋白结合位点来源于转基因动物物种并且顶端结构域来源于灵长类动物(例如人或猴)的情况下。因此,本发明提供了编码嵌合转铁蛋白受体的多核苷酸,该嵌合转铁蛋白受体包含非人哺乳动物转铁蛋白结合位点和具有与seqidno:1至少80%相同的氨基酸序列的顶端结构域。本发明还提供了表达此类嵌合tfr的非人(例如非灵长类动物)转基因动物,以及非人转基因动物在体内筛选可通过与人转铁蛋白受体(hutfr)结合而穿过bbb的多肽中的用途。在一些实施方案中,非人转基因动物含有天然转铁蛋白受体(诸如小鼠转铁蛋白受体(mtfr)),其中顶端结构域被具有与seqidno:1至少80%相同的氨基酸序列的直系同源顶端结构域取代,从而使得天然转铁蛋白结合位点和编码转铁蛋白受体的序列的大部分(例如至少70%或至少75%)完整。因此,这种非人转基因动物最大程度地保留了非人动物的内源性转铁蛋白受体的转铁蛋白结合功能,包括维持适当的铁内稳态以及结合和转运转铁蛋白的能力。结果,转基因动物是健康的并且适合用于治疗脑疾病的治疗剂的发现和开发。
术语
如本文所用,单数形式“一个/一种(a/an)”和“所述(the)”包括复数指示物,除非内容另有明确说明。因此,例如,提及“抗体”任选地包括两种或更多种此类分子的组合等。
如本文所用,当用于修饰数值或范围中指定的量时,术语“约”和“大约”表示数值以及本领域技术人员已知的与所述值的合理偏差,例如,±20%,±10%或±5%在所述值的预期含义内。
如本文所用的“转铁蛋白受体”是指转铁蛋白受体蛋白1。人转铁蛋白受体1多肽序列如seqidno:6所示。来自其他物种的转铁蛋白受体蛋白1序列也是已知的(例如,黑猩猩,登录号xp_003310238.1;猕猴,np_001244232.1;狗,np_001003111.1;牛,np_001193506.1;小鼠,np_035768.1;大鼠,np_073203.1;和鸡,np_990587.1)。术语“转铁蛋白受体”还包括示例性参考序列例如人序列的等位基因变体,其由转铁蛋白受体蛋白1染色体基因座处的基因编码。全长运铁蛋白受体蛋白包括短的n-末端细胞内区、跨膜区和大的细胞外结构域。细胞外结构域的特征在于三个结构域:蛋白酶样结构域、螺旋结构域和顶端结构域。
如本文所用的术语“嵌合tfr”是指转铁蛋白受体蛋白,其具有顶端结构域的被来自异源性转铁蛋白受体的相应顶端结构域区取代的全部或亚区。
如本文所用的“转铁蛋白结合位点”是指tfr蛋白的螺旋和蛋白酶样结构域中的区域,其介导转铁蛋白(例如铁结合的转铁蛋白)与受体的结合。转铁蛋白结合位点位于顶端结构域的远端。
如本文所用的“非人哺乳动物转铁蛋白结合位点”是指来自非人哺乳动物的天然转铁蛋白受体的转铁蛋白结合位点的序列,或其能够与天然非人哺乳动物转铁蛋白结合的功能衍生物。在一些实施方案中,非人哺乳动物转铁蛋白结合位点包含与非人类哺乳动物的天然转铁蛋白受体的转铁蛋白结合位点至少80%、至少90%、至少95%或至少98%相同的氨基酸序列。非人哺乳动物的实例包括小鼠、大鼠、兔、牛、羊、犬、猫、马、猪、非人灵长类动物等。
如本文所用,“hutfr顶端的+/+小鼠”是指转基因小鼠,其中小鼠转铁蛋白受体的顶端结构域已经被人转铁蛋白受体的顶端结构域取代,并且转基因小鼠是转基因纯合的。
如本文所用,“hutfr顶端的+/-小鼠”是指转基因小鼠,其中小鼠转铁蛋白受体的顶端结构域已被人转铁蛋白受体的顶端结构域取代;并且转基因小鼠是转基因杂合的。
如本文所用,关于转铁蛋白受体或其结构域的术语“野生型”、“天然的”和“天然存在的”是指具有在自然界存在的序列的转铁蛋白受体或其结构域。
如本文所用的“内源性”转铁蛋白受体或其结构域是指在细胞或非人动物中天然存在的转铁蛋白受体,即,在不对细胞或动物进行遗传修饰的情况下。
如本文所用,关于转铁蛋白受体的结构域(例如顶端结构域)的术语“异源性”是指在其天然背景之外表达的转铁蛋白受体的结构域,例如,从它通常在自然界中接近的转铁蛋白受体序列分离,或与它通常不接近的转铁蛋白受体序列邻近(或连续)。
术语“氨基酸”是指天然存在的和合成的氨基酸,以及以与天然存在的氨基酸类似的方式起作用的氨基酸类似物和氨基酸模拟物。
天然存在的氨基酸是由遗传密码编码的那些氨基酸,以及后来被修饰的那些氨基酸,例如羟脯氨酸,γ-羧基谷氨酸和o-磷酸丝氨酸。天然存在的α-氨基酸包括但不限于丙氨酸(ala)、半胱氨酸(cys)、天冬氨酸(asp)、谷氨酸(glu)、苯丙氨酸(phe)、甘氨酸(gly)、组氨酸(his)、异亮氨酸(ile)、精氨酸(arg)、赖氨酸(lys)、亮氨酸(leu)、甲硫氨酸(met)、天冬酰胺(asn)、脯氨酸(pro)、谷氨酰胺(gln)、丝氨酸(ser)、苏氨酸(thr)、缬氨酸(val)、色氨酸(trp),酪氨酸(tyr)及它们的组合。天然存在的α-氨基酸的立体异构体包括但不限于d-丙氨酸(d-ala)、d-半胱氨酸(d-cys)、d-天冬氨酸(d-asp)、d-谷氨酸(d-glu)、d-苯丙氨酸(d-phe)、d-组氨酸(d-his)、d-异亮氨酸(d-ile)、d-精氨酸(d-arg)、d-赖氨酸(d-lys)、d-亮氨酸(d-leu)、d-甲硫氨酸(d-met)、d-天冬酰胺(d-asn)、d-脯氨酸(d-pro)、d-谷氨酰胺(d-gln)、d-丝氨酸(d-ser)、d-苏氨酸(d-thr)、d-缬氨酸(d-val)、d-色氨酸(d-trp)、d-酪氨酸(d-tyr)及它们的组合。
氨基酸在本文中可以通过它们通常已知的三字母符号或由iupac-iub生物化学命名委员会推荐的单字母符号表示。
术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,指氨基酸残基的聚合物。该术语适用于其中一个或多个氨基酸残基是相应天然存在的氨基酸的人工化学模拟物的氨基酸聚合物,以及天然存在的氨基酸聚合物和非天然存在的氨基酸聚合物。氨基酸聚合物可以包含全部是l-氨基酸、全部是d-氨基酸,或l和d氨基酸的混合物。
“保守修饰的变体”是指导致用可归类为具有相似特征的另一种氨基酸取代氨基酸的改变。以这种方式限定的保守氨基酸组的类别的实例可包括:“带电荷/极性组”,包括glu(谷氨酸或e)、asp(天冬氨酸或d)、asn(天冬酰胺或n)、gln(谷氨酰胺或q)、lys(赖氨酸或k)、arg(精氨酸或r)和his(组氨酸或h);“芳香族组”,包括phe(苯丙氨酸或f)、tyr(酪氨酸或y)、trp(色氨酸或w)和(组氨酸或h);和“脂肪族组”,包括gly(甘氨酸或g)、ala(丙氨酸或a)、val(缬氨酸或v)、leu(亮氨酸或l)、ile(异亮氨酸或i)、met(甲硫氨酸或m)、ser(丝氨酸或s)、thr(苏氨酸或t)和cys(半胱氨酸或c)。在每一组中,也可以鉴定亚组。例如,可以将带电荷或极性氨基酸组细分为亚组,其包括:包含lys、arg和his的“带正电荷的亚组”;含有glu和asp的“带负电荷的亚组”;以及包含asn和gln的“极性亚组”。在另一个实例中,可以将芳香族或环状组细分为亚组,其包括:包含pro、his和trp的“氮环亚组”;以及包含phe和tyr的“苯基亚组”。在另一个实例中,可以将脂肪族组细分为亚组,例如包含val、leu、gly和ala的“脂肪族非极性亚组”;和包含met、ser、thr和cys的“脂肪族轻微极性亚组”。保守突变的类别的实例包括上述亚组内的氨基酸的氨基酸取代,诸如但不限于:lys取代arg,或反之亦然,使得可以维持正电荷;glu取代asp,或反之亦然,使得可以维持负电荷;ser取代thr,或反之亦然,使得可以维持游离的-oh;以及gln取代asn,或反之亦然,使得可以维持游离的-nh2。在一些实施方案中,例如在活性位点疏水性氨基酸取代天然存在的疏水性氨基酸以保持疏水性。
在两条或更多条多肽序列的上下文中,术语“相同的”或“同一性”百分比是指,在比较窗上进行最大对应比较和比对时的指定区域内,或者如使用序列比较算法或通过手动比对和视觉检查所测量的指定区域内是相同的,相同或具有指定百分比(例如,至少60%同一性、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%或至少95%或更大)的氨基酸残基的两条或更多条序列或子序列。
对于多肽的序列比较,通常一条氨基酸序列充当参考序列,候选序列与其进行比较。可以使用本领域技术人员可获得的各种方法(例如视觉比对),或使用公众可获得的软件,使用已知算法进行比对以实现最大比对。这些程序包括blast程序、align、align-2(genentech,南旧金山,calif.)或megalign(dnastar)。用于实现最大比对的比对所用的参数可以由本领域技术人员确定。为了本申请的目的,对于多肽序列的序列比较,使用用于比对两条蛋白质序列的具有缺省参数的blastp算法标准蛋白blast。
术语“包含”旨在意指组合物和方法包括所列举的要素,但不排除其他要素。当用于限定组合物和方法时,“基本上由......组成”是指特定的材料或步骤以及不会实质上影响所要求保护的本发明的基本的和新的特性的那些材料或步骤。“由......组成”意指排除多于痕量的其他成分和所列举的实质方法步骤。由这些过渡术语中的每一个限定的实施方案都在本发明的范围内。
术语“多核苷酸”、“核酸”和“寡核苷酸”可互换使用,并且是指任何长度的聚合形式的核苷酸(脱氧核糖核苷酸或核糖核苷酸或它们的类似物)。多核苷酸可以具有任何三维结构,并且可以执行已知或未知的任何功能。以下是多核苷酸的非限制性实例:基因或基因片段(例如,探针、引物、est或sage标签)、外显子、内含子、信使rna(mrna)、转运rna、核糖体rna、核酶、cdna、重组多核苷酸、分支多核苷酸、质粒、载体、任何序列的分离的dna、任何序列的分离的rna、核酸探针和引物。多核苷酸可包含修饰的核苷酸,诸如甲基化的核苷酸和核苷酸类似物。如果存在,可以在多核苷酸组装之前或之后赋予对核苷酸结构的修饰。核苷酸序列可被非核苷酸组分中断。聚合后可以进一步修饰多核苷酸,诸如通过与标记组分缀合。该术语还指双链和单链分子。除非另有说明或要求,否则作为多核苷酸的本发明的任何实施方案包括双链形式和已知或预测构成双链形式的两种互补单链形式中的每一种。
多核苷酸由四种核苷酸碱基的特定序列组成:腺嘌呤(a);胞嘧啶(c);鸟嘌呤(g);胸腺嘧啶(t);以及当多核苷酸是rna时,尿嘧啶(u)取代胸腺嘧啶。因此,术语“多核苷酸序列”是多核苷酸分子的字母表示。
术语“敲入”是指预定遗传基因座中的dna序列信息的一对一替换或插入在该基因座内未发现的序列信息。本领域技术人员将容易理解如何使用各种遗传方法(例如crispr/cas9系统、zfn、talen、转座子介导的插入)在基因组的特定基因座中敲入的靶多核苷酸序列。
术语“血脑屏障”或“bbb”是指高度选择性的半透膜屏障,其将循环血液与中枢神经系统(cns)中的脑细胞外液分开。血脑屏障由通过紧密连接而连接的脑内皮细胞形成。
转铁蛋白受体
转铁蛋白受体经由受体介导的配体占据的转铁蛋白受体的内吞作用介导铁的细胞摄取。tfr存在于人和非人物种中,例如非人灵长类动物和啮齿动物。天然人tfr(hutfr)(uniprotp02786,seqidno:6)是同型二聚体ii型跨膜蛋白;它具有细胞质结构域、跨膜区和细胞外结构域,其包含顶端结构域和转铁蛋白结合结构域。hutfr的每个单体具有三个结构上不同的结构域:靠近膜的蛋白酶样结构域,负责所有二聚体接触的螺旋结构域和膜远端的顶端结构域(lawrence等人,science,286(1999),第779-782页)。hutfr二聚体的分子量约为190,000道尔顿。具有seqidno:1的序列(由seqidno:2编码)的hutfr的顶端结构域不参与转铁蛋白和tfr之间的相互作用。据推测,该结构域可为其他蛋白质提供接触表面以结合tfr。天然食蟹猴tfr、天然猕猴tfr和天然黑猩猩tfr也是已知的,例如分别通过登录号xp_005545315、np_001244232.1和xp_003310238.1表示。天然食蟹猴tfr、天然猕猴tfr和天然黑猩猩tfr的顶端结构域分别与seqidno:1的天然人tfr的顶端结构域共有约96%、95%和98%的序列同一性。
天然小鼠tfr(mtfr)(uniprotq62351,seqidno:5)与hutfr具有约77%的氨基酸序列同一性。天然mtfr的顶端结构域与天然hutfr的顶端结构域约74%同一。mtfr包含与人类对应物相似的三个结构上不同的结构域。具有注释外显子和内含子的小鼠tfr的完整基因序列可以从ncbi数据库(基因id:22042)中找到。在染色体16上发现小鼠tfr(ncbi参考序列nc_000082.6)。
一方面包括嵌合tfr多肽。在一些实施方案中,嵌合tfr包含非人哺乳动物转铁蛋白结合位点和与hutfr的顶端结构域(seqidno:1)共有例如,至少75%、至少77%、至少80%、至少85%、至少90%或至少95%的氨基酸序列同一性的异源性顶端结构域。在一些实施方案中,异源性顶端结构域具有seqidno:1、seqidno:7、seqidno:8或seqidno:9的序列。
嵌合tfr的非人哺乳动物转铁蛋白结合位点允许非哺乳动物转铁蛋白与嵌合tfr的特异性结合。在一些实施方案中,非人哺乳动物转铁蛋白结合位点是天然转铁蛋白结合位点,例如小鼠转铁蛋白受体结合位点。
在一些实施方案中,嵌合tfr多肽包含天然tfr多肽,其中仅天然顶端结构域被异源性顶端结构域取代。在一些实施方案中,嵌合tfr多肽包含天然tfr结合位点和与天然tfr结合位点异源的顶端结合结构域,例如,其中除顶端结构域之外,还有至少一个结构域或其区域具有非天然氨基酸序列。
在一些实施方案中,嵌合tfr多肽与seqidno:3具有至少80%、至少85%、至少85%、至少92%、至少95%或至少98%的氨基酸序列同一性。在一个实施方案中,编码嵌合tfr多肽的多核苷酸包含小鼠运铁蛋白受体基因的外显子和内含子以及编码hutfr顶端结构域的核酸序列。在一个实施方案中,非人哺乳动物tfr顶端结构域被hutfr顶端结构域的编码序列取代,例如通过用hutfr顶端结构域序列取代非人哺乳动物tfr基因中的相应外显子。在示例性实施方案中,非人哺乳动物tfr基因是小鼠tfr基因。在一个实施方案中,mtfr顶端结构域被hutfr顶端结构域的编码序列取代,所述hutfr顶端结构域的编码序列位于例如小鼠运铁蛋白受体基因的第四外显子之后以产生嵌合tfr。
在一些方面,本发明提供了分离的核酸,其包含编码包含本文所述的嵌合tfr多肽的任何多肽的核酸序列。在一些实施方案中,编码嵌合tfr多肽中的异源性顶端结构域的核酸序列的区域与天然hutfr的顶端结构域的编码序列(seqidno:2)共有至少75%、至少77%、至少80%、至少85%、至少90%或至少95%的核酸序列同一性。
在另一个方面,提供了多核苷酸,其包含编码本文所述的嵌合转铁蛋白受体的核苷酸序列。多核苷酸可以是单链或双链的。在一些实施方案中,多核苷酸是dna。在具体的实施方案中,多核苷酸是cdna。在一些实施方案中,多核苷酸是rna。
密码子优化
在一些实施方案中,嵌合tfr的编码序列(尤其是编码hutfr顶端结构域的序列)经过密码子优化,以改善嵌合tfr在小鼠中的表达。用于密码子优化的方法是容易获得的,例如,在http://genomes.urv.es/optimizer可获得的优化程序和来自dna2.0(newark,加利福尼亚)的
用所期望的顶端结构域取代非人哺乳动物转铁蛋白受体的顶端结构域的方法
可以使用多种方法(例如锌指核酸酶(zfn)、tale效应结构域核酸酶(talen)、转座子介导的系统和以及cripsr/cas9系统)产生包含如本文所公开的异源性顶端结构域的敲入的非人转基因动物。这些方法通常包括向细胞施用一种或多种编码一种或多种核酸酶的多核苷酸,使得核酸酶通过剪切dna以在dna链中产生5'和3'切割末端来介导内源性基因的修饰。在存在供体序列(其侧翼为与从5'末端延伸5'的序列和从3'末端延伸3'的序列基本同源的左同源臂和右同源臂)的情况下,所述供体被整合到经由同源定向修复(hdr)由核酸酶靶向的内源性基因。在一些实施方案中,使用crispr/cas9系统进行敲入。例如,将编码异源性顶端结构域的核酸序列引入内源性tfr基因以产生嵌合tfr,这导致编码顶端结构域的天然存在的序列被取代,但基因的整体结构得以维持。
crispr
在一些实施方案中,使用cripsr/cas9系统进行与seqidno:1至少80%相同的顶端结构域的敲入。crispr/cas9系统包括cas9蛋白和至少一至两种核糖核酸,其能够将cas9蛋白导向至待取代的转铁蛋白受体的顶端结构域中的靶基序并与之杂交。这些核糖核酸通常被称为“单向导rna”或“sgrna”。然后cas9蛋白切割靶基序,这导致双链断裂或单链断裂。在包含侧翼为两个同源臂的hutfr顶端结构域编码序列的供体dna存在下,供体dna被插入靶转铁蛋白受体dna中,取代顶端结构域。
用于本发明的cas9蛋白可以是天然存在的cas9蛋白或其功能衍生物。天然序列多肽的“功能衍生物”是与天然序列多肽具有共同的定性生物学性质的化合物。“功能衍生物”包括但不限于天然序列的片段和天然序列多肽的衍生物及其片段,条件是它们与相应的天然序列多肽具有共有的生物学活性。本文考虑的生物学活性是cas9的功能衍生物将dna底物水解成片段的能力。cas9多肽或其片段的合适功能衍生物包括但不限于突变体、融合物、cas9蛋白的共价修饰或其片段。
在一些实施方案中,cas9蛋白来自酿脓链球菌(streptococcuspyogenes)。cas9含有2个核酸内切酶结构域,包括剪切与sgrna非互补的靶dna的ruvc样结构域,以及剪切与sgrna互补的靶dna的hnh核酸酶结构域。cas9的双链核酸内切酶活性还需要被称为前间隔相关基序(pam)的短的保守序列(2-5个核苷酸),其紧跟靶序列中靶基序的3'。在一些实施方案中,pam基序是ngg基序。在一个示例性实施方案中,通过使用cas9蛋白取代小鼠中的顶端结构域,所述cas9蛋白由sgrna导向至小鼠基因的外显子4和9之间的区域。将供体dna引入反应中。供体dna包含人顶端结构域编码序列,其位于与外显子4上游起始的小鼠tfr序列同源的左同源臂和与外显子9内起始的小鼠tfr序列同源的右同源臂之间。在某些实施方案中,左同源臂与外显子4上游起始的小鼠tfr序列重叠817个核苷酸,并且右同源臂与外显子9内起始的小鼠tfr序列重叠807个核苷酸。结果,可以在第四小鼠外显子后插入编码所期望的顶端结构域(诸如具有与seqidno:1至少80%相同的氨基酸序列的顶端结构域)的核苷酸序列,并且插入的核苷酸序列在3'末端的侧翼为恰当跟随的小鼠外显子。在一些实施方案中,插入小鼠tfr基因的人顶端结构域编码序列是为小鼠表达而密码子优化的。
可以根据所用的特定crispr/cas9系统和靶多核苷酸的序列选择sgrna。在一些实施方案中,设计一至两种核糖核酸以与紧邻cas9蛋白识别的脱氧核糖核酸基序的靶基序杂交。在一些实施方案中,设计一至两种核糖核酸中的每一种以与紧邻cas9蛋白识别的脱氧核糖核酸基序的靶基序杂交,其中靶基序位于待取代的基因组序列的侧翼。可以使用例如,在http://crispr.mit.edu容易获得的软件设计向导rna。可用于产生嵌合tfr转基因小鼠的示例性sgrna包括seqidno:10-11。
如本文所公开的供体dna包含编码与seqidno:1至少75%相同的氨基酸序列的核苷酸序列。在一些实施方案中,供体dna包含编码seqidno:1或编码与seqidno:1共有至少75%、至少77%、至少80%、至少85%、至少90%或至少95%的氨基酸序列同一性的序列的序列。在一些实施方案中,供体dna包含seqidno:2的核苷酸序列或与seqidno:2共有至少60%、至少70%、至少77%、至少80%、至少85%、至少90%的序列或至少95%的序列同一性的序列。如本文所公开的供体dna还包含左同源臂和右同源臂,其位于顶端结构域编码序列的侧翼,并被设计为与相对于cas9蛋白的剪切位点与5'和3'外显子序列重叠。同源臂可以延伸超出5'和3'外显子序列,并且每个同源臂的长度可以是至少20、30、40、50、100或150个核苷酸。本领域技术人员可以容易地确定实验所需的同源臂的最佳长度。在一个示例性实施方案中,供体dna的左同源臂跨越seqidno:4的第1-817位核苷酸,并且右同源臂跨越seqidno:4的第1523-2329位核苷酸。在一些实施方案中,左同源臂与seqidno:4的第1-817位核苷酸共有至少85%、至少90%、至少95%或至少98%的序列同一性。在一些实施方案中,右同源臂与seqidno:4的第1523-2329位核苷酸共有至少85%、至少90%、至少95%或至少98%的序列同一性。
在一些实施方案中,还可以选择sgrna以最小化与除靶多核苷酸序列之外的核酸序列的杂交。在一些实施方案中,设计一至两种核糖核酸以与靶基序杂交来最小化crispr/cas9系统的脱靶效应,所述靶基序当与细胞中的所有其他基因组核苷酸序列相比时含有至少两个错配。本领域技术人员将理解,可以使用多种技术来选择合适的靶基序以最小化脱靶效应(例如,生物信息学分析)。使用crispr/cas9系统来降低基因表达的方法被描述于各种出版物中,例如美国专利公开第2014/0170753号和第2016/0257974号,其公开内容通过引用整体并入本文。
锌指核酸酶(zfn)
在一些实施方案中,通过使用zfn敲入hutfr顶端结构域来产生嵌合tfr。zfn是融合蛋白,其包含foki内切核酸酶的非特异性剪切结构域(n)和锌指蛋白(zfp)。涉及一对znf以识别靶基因中的特定基因座:一个识别待修饰位点的上游序列,并且另一个识别待修饰位点的下游序列。zfn的核酸酶部分在特定基因座处切割。然后可以将如上所述的供体dna插入特定基因座。使用zfn以降低基因表达的方法是众所周知的,例如,如美国专利第9,045,763号以及durai等人,“zincfingernucleases:custom-designedmolecularscissorsforgenomeengineeringofplantandmammaliancells,”nucleicacidresearch,33(18):5978-5990(2005)中所公开的,它们的公开内容通过引用整体并入。
转录激活因子样效应物核酸酶(talen)
在一些实施方案中,通过用talen敲入hutfr顶端结构域来产生嵌合tfr。talen类似于zfn,因为它们在基因组位点周围成对结合并引导相同的非特异性核酸酶foki在特定位点剪切基因组,但不是识别dna三联体,而是每个结构域识别单个核苷酸。使用zfn以降低基因表达的方法也是众所周知的,例如,如美国专利第9,005,973号以及christian等人,“targetingdnadouble-strandbreakswithtaleffectornucleases,”genetics,186(2):757-761(2010)中所公开的,它们的公开内容通过引用整体并入。
表达嵌合tfr的宿主细胞/转基因动物
在一些实施方案中,本发明提供了表达嵌合tfr的宿主细胞,例如,其包含编码上述嵌合转铁蛋白受体的核酸序列。在一些实施方案中,宿主细胞是非人哺乳动物细胞。上述任何敲入方法(即crispr、talen、锌指核酸酶)可被用于用具有与seqidno:1至少80%相同的氨基酸序列的异源性顶端结构域取代宿主细胞中天然转铁蛋白受体的顶端结构域。在一些实施方案中,宿主细胞是与seqidno:1至少80%相同的真核细胞,例如小鼠细胞。在一些情况下,使宿主细胞与sgrna和cas9、包含编码异源性顶端结构域的核酸序列的供体dna接触,所述核酸序列侧翼为左同源臂和右同源臂。sgrna和同源臂具有这样的序列,使得异源性顶端结构域编码序列插入基因组中的位置以取代宿主细胞中天然转铁蛋白受体的顶端结构域的编码序列。在一些实施方案中,宿主细胞是来自非灵长类哺乳动物(诸如小鼠、大鼠、兔、牛、羊、犬、猫、马、猪等)的细胞。
在一些实施方案中,在胚胎干(es)细胞中进行敲入的方法以产生表达嵌合转铁蛋白受体多肽的es细胞。然后胚胎干细胞可以发育成子代细胞或非人转基因动物,其基因组包含编码嵌合转铁蛋白受体多肽的核酸。在一些实施方案中,将es细胞引入囊胚中并转移到假孕雌性动物中。在一些情况下,可以选择携带转基因的建群雄性动物并与野生型雌性动物交配以产生f1杂合小鼠。随后可以通过f1代杂合非人动物的交配产生纯合的非人动物。用于培养es细胞和引入核苷酸序列以靶向es细胞的基因组来产生转基因动物的方法是众所周知的,例如,如ramirez-solis等人,“genetargetinginmouseembryonicstemcells,”methodsenzymol.,225:855-878(1993);和美国专利公开第2013/0318643号中所描述的,它们的公开内容通过引用整体并入。在一些实施方案中,来自具有本发明的嵌合tfr的转基因动物的胚胎干细胞可被用作提供转基因动物后代的来源。
在一些实施方案中,敲入的方法在单细胞非人动物中进行。在一个示例性实施方案中,经由原核显微注射将sgrna、cas9和包含与seqidno:1至少80%相同的顶端结构域编码序列的供体多核苷酸引入单细胞胚胎中,其中编码序列侧翼为左同源臂和右同源臂。然后将受体胚胎转移到假孕雌性动物中。sgrna与cas9蛋白形成复合物,然后其靶向非人动物胚胎中转铁蛋白受体的顶端结构域的编码序列。结果,非人动物转铁蛋白受体顶端结构域被剪切并被来自供体多核苷酸的转铁蛋白受体顶端结构域编码序列取代。在一些情况下,可以选择携带转基因的建群雄性动物并与野生型雌性动物交配以产生f1杂合小鼠。随后可以通过f1代杂合非人动物的交配产生纯合的非人动物。本文公开的转基因动物可以是啮齿动物,例如小鼠或大鼠。
在一个示例性实施方案中,部分由于非人转基因动物(例如非灵长类哺乳动物)保留天然tfr的内含子和转铁蛋白结合结构域的事实,通过敲入具有与seqidno:1至少80%相同的氨基酸序列的顶端结构域产生的转基因动物通常是健康的,并且表现出与相同物种的野生型小鼠的那些生理状态相似的生理状态。在一个实施方案中,保留tfr的顶端结构域外的所有内含子。例如,tfr表达水平类似于同一物种的野生型动物;转基因小鼠的表达水平小于野生型小鼠的表达水平的不超过10%、20%、30%、40%、50%、60%、70%、80%或90%,或大于野生型小鼠的表达水平的不超过10%、20%、30%、40%、50%、75%、100%、150%、200%、300%或500%。红血细胞计数、血红蛋白水平和/或血细胞比容水平也与相同物种的野生型动物相似;差值不大于50%,例如,不大于40%、不大于30%、不大于20%或不大于10%。在典型的实施方案中,根据本发明的转基因动物保留选择性bbb转运(其能够输入营养物和蛋白质)并保留保护cns免受毒素的能力;转基因的存在不会干扰转铁蛋白结合或fcrn结合能与顶端结构域结合的抗体,如下所述。通常,转基因动物中的tfr介导的细胞运输也与那些野生型动物相似。与完全缺乏人tfr的野生型小鼠或表达整个hutfr细胞外结构域(例如,表达整个hutfr蛋白质)的转基因动物模型相比,根据本发明的转基因动物作为人bbb穿透药物的药代动力学或药效动力学研究的模型更相关。
尽管如实施例中所示在小鼠中说明了本发明,但本领域普通技术人员将理解其他非人哺乳动物,例如啮齿动物、兔、牛、羊、犬、猫、马、猪、骆驼科动物、非人灵长类动物和其他哺乳动物也可以被工程化以类似方式表达嵌合tfr,并且这些转基因动物也可以用于本文公开的应用。
顶端结构域结合多肽
如本文所用的“顶端结构域结合多肽”或“adbp”是指能与具有与seqidno:1至少80%相同的氨基酸序列的顶端结构域结合的多肽。adbp可以是能够与嵌合tfr的hutfr的顶端结构域结合的抗体或任何多肽。在一些实施方案中,adbp是将穿过血脑屏障递送的试剂。在一些实施方案中,adbp还包含例如通过共价连接与其连接的效应分子。效应分子可以是治疗剂、标记试剂或诊断试剂。在某些实施方案中,效应分子是多肽,诸如治疗或诊断抗体,或对酶或信号转导分子具有酶活性或抑制活性的多肽。在某些实施方案中,效应分子包括小分子、rna、dna或蛋白质。
在一些实施方案中,adbp是双特异性抗体,其中顶端结构域结合区是识别顶端结构域的抗体,并且效应分子是识别不同抗原(例如酶或信号转导分子)的抗体,并且效应部分的结合激活或抑制酶或信号转导分子。
筛选能结合嵌合tfr的adbp
本文所公开的嵌合tfr可被用于筛选能够结合tfr的adbp。筛选方法包括使候选adbp与上文所公开的嵌合tfr接触并确定与嵌合tfr结合的候选adbp的量。在一些实施方案中,使候选adbp与嵌合tfr接触的步骤包括使adbp与表达嵌合tfr的宿主细胞接触。在一些情况下,使候选adbp与嵌合tfr接触的步骤包括使adbp与表达嵌合tfr的内皮细胞接触。在一些实施方案中,内皮细胞是bbb内皮细胞。
可以使用本领域熟知的方法(例如免疫测定法或spr)测量候选adbp和tfr之间的相互作用。在一些实施方案中,通过elisa、biacoretm系统或共免疫沉淀法测量候选adbp与嵌合tfr的结合。
筛选可以穿过bbb的adbp
如上所述的表达嵌合tfr的非人转基因动物可被用于表征adbp结合嵌合tfr的顶端结构域的能力以及最终穿过bbb的能力。
通常,为了评估adbp穿过bbb的能力,向携带本文所公开的嵌合tfr的转基因动物施用adbp,优选地通过静脉内注射。在一段时间(例如,至少10min、至少20min、至少30min、至少60min、至少90min、至少120min、至少180min或至少240min)后,处死转基因动物并分析脑组织以确定adbp的存在。可以通过测定adbp和/或与其连接的效应分子的存在来确定adbp的存在。在一些实施方案中,用盐水(例如pbs)灌注脑组织,并在检测前固定。可以使用标准成像方法(例如,免疫组织化学或免疫荧光方法)检测切片中效应分子的存在。脑组织中效应分子的阳性检测表明效应分子可以穿过bbb。在一些情况下,确定脑中adbp的存在包括进行定量免疫测定法。使用嵌合tfr转基因小鼠测量穿过bbb的转运的测定是稳健的并且可以测量adbp摄取中大于10倍、20倍、30倍、40倍或50倍的提高。
在一些实施方案中,除了使用成像方法或免疫测定法来检测脑中adbp的存在之外,检测效应分子的底物变化的方法也可被用于评估效应分子的脑摄取。在一个示例性实施方案中,adbp包含能够抑制脑中酶的酶活性的效应分子。在一些实施方案中,可以通过评估由adbp或与其连接的效应分子调节的酶的酶活性来测量adbp的脑摄取(即反映其bbb转运的能力)。
在一些实施方案中,测量脑中候选adbp的脑摄取。还可以监测血浆并评估药代动力学简况。在施用候选效应分子后,与非bbb穿透分子相比,脑-血浆比率的增加表明候选adbp可以穿过bbb。
在一些情况下,包含编码嵌合tfr的多核苷酸的非人转基因动物可以与经过工程化以显示某种疾病表型的非人转基因动物杂交。在一些情况下,非人转基因动物是可以与各种小鼠模型杂交的转基因小鼠,例如,诸如美国专利第8,476,485号中所描述的als小鼠模型;诸如美国专利第5,898,094和美国专利第6,175,057号中所描述的ad小鼠模型;诸如美国专利公开第2016/0050895号中所描述的tspo小鼠模型;诸如美国专利公开第2014/0041062号中所描述的自闭症谱系障碍(asd)小鼠模型。这些上述专利和专利申请的全部内容在此通过引用并入。在某些情况下,通过此类杂交产生的杂交小鼠可被用于评估包含效应分子的adbp在脑中的分布以及adbp或效应分子在治疗脑疾病中的功效。
试剂盒
在一些实施方案中,提供了包含如本文所述的嵌合转铁蛋白受体多核苷酸或多肽,或表达此类多肽的细胞的试剂盒。在一些实施方案中,试剂盒用于筛选如上所述的adbp。
在一些实施方案中,试剂盒还包含可被用于测定法中的缓冲液和容器,以检测嵌合tfr多肽和候选adbp之间的结合。在一些实施方案中,试剂盒还包含含有用于实施本文所述方法的指导(即方案)的说明材料(例如,使用所述试剂盒跨越血脑屏障施用组合物的说明书)。虽然说明材料通常包括书面或印刷材料,但它们不限于此。本发明考虑了能够存储此类说明书并将它们传递给最终用户的任何介质。此类介质包括但不限于电子存储介质(例如,磁盘、磁带、卡匣(cartridge)、芯片)、光学介质(例如cd-rom)等。此类介质可以包括提供此类说明材料的因特网站点的地址。
实施例
以下实施例仅用于说明目的,不应解释为对所要求保护的发明的限制。存在多种本领域技术人员可以使用的替代技术和程序,它们类似地允许本领域技术人员成功进行预期的发明。
实施例1:hutfr小鼠的产生和表征
用于产生敲入/敲除小鼠的方法已经公开在文献中并且是本领域技术人员众所周知的。简而言之,c57b16小鼠被用于经由原核显微注射到单细胞胚胎中,然后将胚胎转移到假孕雌性动物,来产生人顶端tfr小鼠系的敲入。具体地,将cas9、seqidno:10-11的sgrna和seqidno:4的供体dna引入胚胎中。供体dna包含已为在小鼠中表达而密码子优化的人顶端结构域编码序列seqidno:2。顶端结构域编码序列侧翼为左同源臂(seqidno:4的第1-817位核苷酸)和右同源臂(seqidno:4的第1523-2329位核苷酸)。以这种方式设计供体序列,使得在第四小鼠外显子后插入顶端结构域,并且就在其3'末端侧翼为第九小鼠外显子。将来自接受胚胎的雌性动物的后代的建群雄性动物与野生型雌性动物交配以产生f1杂合的小鼠。随后从f1代杂合的小鼠的交配中产生纯合的小鼠。
实施例2:用于监测抗体脑摄取的工具抗体的产生
通过用含有编码重链和轻链的dna的表达质粒转化expi293或expicho细胞并使用本领域技术人员熟悉的方案产生靶向人tfr或人/小鼠bace1的工具抗体。使用“杵臼(knobs-into-holes)”技术产生双特异性抗体;分别表达杵和臼半抗体,然后使用公开的方法连接。首先用蛋白a纯化抗体,然后通过尺寸排阻色谱纯化。为这些研究产生的抗体如下:
抗tfr:能与人tfr顶端结构域结合的人igg1抗体。
抗bace1:人igg1抗体,其能与人bace1结合并与小鼠bace1交叉反应。该抗体抑制bace1的酶活性。
抗tfr/bace1:人igg1杵臼双特异性抗体,其能与人tfr顶端结构域以及人和小鼠bace1结合。杵半抗体具有来自抗bace1抗体的可变结构域;臼半抗体具有来自抗tfr抗体的可变结构域。
实施例3:hutfr顶端的+/-和hutfr顶端的+/+小鼠的血液分析
从野生型c57b16、hutfr顶端的+/-和hutfr顶端的+/+小鼠(n=3/组)收集血液并进行标准全血细胞计数(cbc)分析。在任何红血细胞参数(包括总红细胞、血红蛋白和血细胞比容水平)中均未观察到基因型特异性差异(图1)。
实施例4:tfr靶向抗体在hutfr顶端的+/-和hutfr顶端的+/+小鼠中的脑定位
在该实施例中,产生抗tfr抗体以评估tfr靶向治疗剂在hutfr顶端的+/-小鼠中的脑摄取。用5mg/kg抗tfr抗体静脉内注射hutfr顶端的+/-小鼠或野生型c57b16。1小时后,处死小鼠并用pbs灌注。将半脑在4%pfa中微量固定过夜,然后30%蔗糖保存。使用切片机切割矢状脑切片(35μm),在5%bsa+0.3%tritonx-100中封闭,然后用alexa488抗huigg1(1:500)进行荧光次级染色。使用具有20x物镜的zeiss宽场显微镜拍摄脑图像。在hutfr顶端的+/-小鼠中观察到显著的血管染色,表明人顶端特异性抗tfr与bbb处tfr高度表达的脑内皮细胞的强烈结合(图2)。相反,在野生型小鼠中观察到非常少的染色。
为了证实tfr特异性bbb转运,使用如上所述的类似方法测试抗bace1抗体和抗tfr/bace1双特异性抗体。用50mg/kg的任一抗体静脉内注射hutfr顶端的+/+小鼠。24小时后,用pbs灌注小鼠,并如上所述对hutfr顶端的+/-小鼠处理半脑并染色。观察到抗tfr/bace1广泛的脑实质染色,而未观察到抗bace1的染色,表明在这些小鼠中bbb转胞吞作用需要tfr顶端结构域结合多肽(图3)。
实施例5:抗体在hutfr顶端的+/+小鼠中的脑和血浆pk/pd
在该实施例中,用50mg/kg的抗bace1抗体或抗tfr/bace1双特异性抗体静脉内注射hutfr顶端的+/+小鼠。24小时后,经由心脏穿刺收集血液,并用pbs灌注小鼠。将脑组织在10×组织重量的裂解缓冲液(含有1%np-40的pbs)中匀浆。在edta管中收集血液以防止凝结并以14000rpm旋转7分钟以分离血浆。按照制造商的说明书,使用通用人igg测定法(msd人igg试剂盒#k150jld)定量小鼠血浆和脑裂解物中的抗体浓度。简而言之,用msd阻断剂a将预包被的板封闭30min。使用hamiltonnimbus液体处理器以1:10,000稀释血浆样品,并一式两份加入到封闭的板中。将脑样品在1%np40裂解缓冲液中匀浆,并将裂解物1:10稀释以用于pk分析。还在同一板上分析给药溶液以确认正确的剂量。使用四参数逻辑回归拟合0.78-200ng/mligg的标准曲线。
24小时后,抗tfr/bace1的血浆水平低于抗bace1的水平,可能是由于该抗体经由与外周表达的hutfr顶端的结合而清除(图4a)。在脑中,观察到与抗bace1相比,抗tfr/bace1的浓度的约28倍的增加(图4b)。抗tfr/bace1的显著积聚是由于bbb处的tfr介导的转胞吞作用,该结果验证了hutfr顶端的+/+小鼠作为测量人tfr顶端结构域结合多肽的bbb摄取的工具。
使用淀粉样蛋白前体蛋白(app)剪切的bace1抑制作为血浆和脑中抗体活性的药效动力学读数。将脑组织在10×组织重量的5m盐酸胍中匀浆,然后在含0.25%酪蛋白缓冲液的pbs中以1:10稀释。使用夹心elisa测量血浆和脑裂解物中的小鼠aβ40水平。将384孔maxisorp板用特异性针对aβ40肽的c末端的多克隆捕获抗体(millipore#abn240)包被过夜。将酪蛋白稀释的胍脑裂解物在elisa板上进一步1:2稀释,并与检测抗体,即生物素化的m3.2同时加入。以1:5稀释度分析血浆。在加入链霉抗生物素蛋白-hrp,随后加入tmb底物之前将样品在4℃孵育过夜。使用四参数逻辑回归拟合0.78-50pg/mlmsaβ40的标准曲线。
与抗bace1相比,抗tfr/bace1处理导致hutfr顶端的+/+小鼠中a-β的减少增加,表明用抗tfr/bace1实现了脑中的bace1靶标参与(图4c)。与未处理的野生型小鼠相比,抗tfr/bace1和抗bace1的血浆a-β降低至相似程度(图4d)。这些数据支持hutfr顶端的+/+小鼠在需要人tfr介导的脑摄取的靶标参与研究中,特别是用于评估人tfr顶端结构域结合多肽的用途。
实施例6:tfr在hutfr顶端的+/+小鼠中的表达
从野生型和hutfr顶端的+/+小鼠中分离脑和各种外周组织,以确定hutfr顶端的+/+小鼠中的tfr表达水平是否改变。在用pbs灌注后从小鼠中取出脑、肝脏、肺和肾脏。将组织在10×组织重量的裂解缓冲液(含有1%np-40的pbs)中匀浆。样品在蛋白质印迹上运行,并且使用识别tfr的细胞内部分并因此与野生型和hutfra顶端的+/+均交叉反应的tfr抗体(1:2000;thermofisher#13-6800)确定tfr表达水平。tfr表达的定量表示为与肌动蛋白(1:5000;abcam8227)的比率。图5a-5d显示在脑(图5a)、肝脏(图5b)、肾脏(图5c)和肺(图5d)中,hutfr顶端的+/+小鼠中的tfr表达与野生型小鼠非常相似。
应理解,本文描述的实施例和实施方案仅用于说明目的,并且鉴于其的各种修改或改变均将推荐给本领域技术人员,并且被包括在本申请的精神和范围以及所附权利要求的范围内。出于所有目的,本文引用的所有出版物、序列登录号、专利和专利申请均通过引用整体并入本文。
示例性序列表
seqidno:1:人顶端结构域插入物的蛋白质序列
aqnsviivdkngrlvylvenpggyvayskaatvtgklvhanfgtkkdfedlytpvngsivivragkitfaekvanaeslnaigvliymdqtkfpivnaelsffghahlgtgdpytpgfpsfnhtqfppsrssglpnipvqtisraaaeklfgnmegdcpsdwktdstcrmvtsesknvkltvsn
seqidno:2:人顶端结构域插入物的dna序列
gctcagaactccgtgatcatcgtggataagaacggccggctggtgtacctggtggagaaccctggcggatacgtggcttactctaaggccgctaccgtgacaggcaagctggtgcacgccaacttcggaaccaagaaggactttgaggatctgtacacaccagtgaacggctctatcgtgatcgtgcgcgctggaaagatcaccttcgccgagaaggtggctaacgccgagagcctgaacgccatcggcgtgctgatctacatggatcagacaaagtttcccatcgtgaacgctgagctgtctttctttggacacgctcacctgggcaccggagacccatacacacccggattccctagctttaaccacacccagttccccccttccaggtctagcggactgccaaacatccccgtgcagacaatcagcagagccgctgccgagaagctgtttggcaacatggagggagactgcccctccgattggaagaccgactctacatgtaggatggtgacctccgagtcaaaaaatgtcaaactcaccgtgtccaat
seqidno:3:在转基因小鼠中表达的嵌合tfr序列(斜体部分代表细胞质结构域,粗体部分代表跨膜结构域,灰色部分代表细胞外结构域,以及粗体和下划线部分代表顶端结构域)
seqidno:4:完整供体dna的序列(左同源臂:1-817;右同源臂:1523-2329;人apical结构域:941-1492;密码子优化序列:821-1522)
ctatacagatatataaggatggggctttttttttttaatttttaaaaaagatttgtttattattatatgtaagtacactgtagctgtcttcagacactccagaagagggcatcagatctcattacagatggttgtgagctaccatgtggtcactgggatttgaactcaggaccttcagaagagcagtcagtgctcttaactgataagttaataataagttaactgataaggtaataaaggtcccctatgaaaagggttcagacccaaagagtcagagatccacaggttgagaacctcctgccctaaatcttgttgctctccttattcaagaccactcctgttgcagttgctcttaagcatgagtatgctcccttctgaaagtctccatagcagccatctctccagccccagagtgaggcttttaaaggaatcttcatgataaatagaatttttaaaaaagtaactgaagttacttaaggtgttaaggtacattttattccctcagtaactggttaatctagcagttttgagtcatacttcatttatcttgactttgaagagtaagatattaaaacaatttgcttgatccttgaagtaagtatttaaatagacattttaatgcagactttttttagttgactggtggtgttgcacgtggtcaatccaagtactcatgggaggcagaggcaggaggatctctctctagaccagcctggtctatagagcaagttccaggacagccagggctacacagaaaccttgtttcaaacaagacttttatccttccaggcagctgagccagaatacatacactcctagggaagctggttcacagaaggacgaatccctggcatactacatcgagaatcagtttcacgagttcaagtttagcaaagtctggagagatgagcactacgtgaagatccaggtgaagagctccgctcagaactccgtgatcatcgtggataagaacggccggctggtgtacctggtggagaaccctggcggatacgtggcttactctaaggccgctaccgtgacaggcaagctggtgcacgccaacttcggaaccaagaaggactttgaggatctgtacacaccagtgaacggctctatcgtgatcgtgcgcgctggaaagatcaccttcgccgagaaggtggctaacgccgagagcctgaacgccatcggcgtgctgatctacatggatcagacaaagtttcccatcgtgaacgctgagctgtctttctttggacacgctcacctgggcaccggagacccatacacacccggattccctagctttaaccacacccagttccccccttccaggtctagcggactgccaaacatccccgtgcagacaatcagcagagccgctgccgagaagctgtttggcaacatggagggagactgcccctccgattggaagaccgactctacatgtaggatggtgacctccgagtcaaaaaatgtcaaactcaccgtgtccaatgtgctgaaagaacgacgcatcctgaatatctttggagttattaaaggttatgaggaaccaggtaaagacctgctttgtactttttcactttactgttttgcttactgtagataggtctagtgcaggaaggagaaggatgctagcttggcatgaactgctatatcttgtttgtcctaatgtgaactttgtaatatatgtgtatataacacataatatggccatgtaagtgtatggagaggccagagttaagtattaaatatctttctgtaatcatttaaaattttacatatgaaggtcagtgaacagattgaaggagttttgtccaggtgggacttggatctaaattttttacaatgcctggcagcaaacacctttttaatcaactgagctgtctccccaaataaagtgaatgtgatatcagcttgtggataatttttttttgttgctttgataagtggttttcttacaggatcacataccagttctgtccatagcattaaacaaacataactgtcatgcagtagattaatgtgcagggcacatccaacagtcacatttattaataggacaaaaagttggaccttatatgtagcacacctataattccagtgctaggaagatccgggtaggagatccttagttcggtgctacttagtgagggtttgtttcaaaaaacaaaagctatgatggtgtgttgccttttttcttttagaccgttatgttgtagtaggagcccagagagacgctttgggtgctggtgttgcggcgaagtccagtgtgggaacaggtcttctgttgaaacttgcccaagtattctcagatatgatttcaaaaggt
seqidno:5:小鼠tfr蛋白质序列(uniprotq62351)(斜体部分代表细胞质结构域,粗体部分代表跨膜结构域,灰色部分代表细胞外结构域,以及粗体和下划线部分代表顶端结构域)
seqidno:6:人tfr蛋白质序列(uniprotp02786)(斜体部分代表细胞质结构域,粗体部分代表跨膜结构域,灰色部分代表细胞外结构域,以及粗体和下划线部分代表顶端结构域)
seqidno:7:恒河猴(猕猴)tfr的顶端结构域(ncbi参考序列np_001244232.1);它与天然人tfr的顶端结构域具有95%的同一性
aqnsviivdkngglvylvenpggyvayskaatvtgklvhanfgtkkdfedldspvnsfnhtqfppsqssglpnipvqtisraaaeklfgnmegdcpsdwktdstckmvtsenksvkltvsn
seqidno:8:黑猩猩tfr的顶端结构域(ncbi参考序列xp_003310238.1);它与天然人tfr的顶端结构域具有98%的同一性
aqnsviivdkngslvylvenpggyvayskaatvtgklvhanfgtkkdfedlhtpvngsivivragkitfaekvanaeslnaigvliymdqtk1fpivnaelsffgh1ahlgtgdpytpg1fpsfnhtqfppsr1ssglpdstcrmvtsesknvkltvsn
seqidno:9:食蟹猴(macacafascicularis)(食蟹猴(cynomolgousmonkey))tfr的顶端结构域(ncbi参考序列xp_005545315);它与天然人tfr的顶端结构域具有96%的同一性
aqnsviivdkngglvylvenpggyvayskaatvtgklvhanfgtkkdfedldspvngsivivragkitfaekvanaeslnaigvliymdqtkfpivkadlsffghahlgtgdpytpgfpsfnhtqfppsqssglpnipvqtisraaaeklfgnmegdcpsdwktdstckmvtsenksvkltvsn
seqidno:10
gaatacatacactcctcgtgagg
seqidno:11
agaagaatacttaacatctttgg
序列表
<110>戴纳立制药公司(denalitherapeuticsinc.)
马克·s·丹尼斯(dennis,marks.)
亚当·p·西尔弗曼(silverman,adamp.)
乔伊·余·祖切罗(zuchero,joyyu)
<120>转铁蛋白受体转基因模型
<130>1076191
<150>us62/460,692
<151>2017-02-17
<150>us62/543,559
<151>2017-08-10
<150>us62/543,658
<151>2017-08-10
<150>us62/583,314
<151>2017-11-08
<160>11
<170>siposequencelisting1.0
<210>1
<211>184
<212>prt
<213>智人(homosapiens)
<400>1
alaglnasnservalileilevalasplysasnglyargleuvaltyr
151015
leuvalgluasnproglyglytyrvalalatyrserlysalaalathr
202530
valthrglylysleuvalhisalaasnpheglythrlyslysaspphe
354045
gluaspleutyrthrprovalasnglyserilevalilevalargala
505560
glylysilethrphealaglulysvalalaasnalagluserleuasn
65707580
alaileglyvalleuiletyrmetaspglnthrlyspheproileval
859095
asnalagluleuserphepheglyhisalahisleuglythrglyasp
100105110
protyrthrproglypheproserpheasnhisthrglnphepropro
115120125
serargserserglyleuproasnileprovalglnthrileserarg
130135140
alaalaalaglulysleupheglyasnmetgluglyaspcysproser
145150155160
asptrplysthraspserthrcysargmetvalthrsergluserlys
165170175
asnvallysleuthrvalserasn
180
<210>2
<211>552
<212>dna
<213>智人(homosapiens)
<400>2
gctcagaactccgtgatcatcgtggataagaacggccggctggtgtacctggtggagaac60
cctggcggatacgtggcttactctaaggccgctaccgtgacaggcaagctggtgcacgcc120
aacttcggaaccaagaaggactttgaggatctgtacacaccagtgaacggctctatcgtg180
atcgtgcgcgctggaaagatcaccttcgccgagaaggtggctaacgccgagagcctgaac240
gccatcggcgtgctgatctacatggatcagacaaagtttcccatcgtgaacgctgagctg300
tctttctttggacacgctcacctgggcaccggagacccatacacacccggattccctagc360
tttaaccacacccagttccccccttccaggtctagcggactgccaaacatccccgtgcag420
acaatcagcagagccgctgccgagaagctgtttggcaacatggagggagactgcccctcc480
gattggaagaccgactctacatgtaggatggtgacctccgagtcaaaaaatgtcaaactc540
accgtgtccaat552
<210>3
<211>763
<212>prt
<213>人工序列(artificialsequence)
<220>
<223>合成的嵌合转铁蛋白受体(tfr)多肽序列
<220>
<221>domain
<222>(1)..(67)
<223>细胞质结构域
<220>
<221>domain
<222>(68)..(88)
<223>跨膜结构域
<220>
<221>domain
<222>(89)..(763)
<223>细胞外结构域
<220>
<221>domain
<222>(198)..(381)
<223>顶端结构域
<400>3
metmetaspglnalaargseralapheserasnleupheglyglyglu
151015
proleusertyrthrargpheserleualaargglnvalaspglyasp
202530
asnserhisvalglumetlysleualaalaaspgluglugluasnala
354045
aspasnasnmetlysalaservalarglysprolysargpheasngly
505560
argleucysphealaalailealaleuvalilephepheleuilegly
65707580
phemetserglytyrleuglytyrcyslysargvalgluglnlysglu
859095
glucysvallysleualagluthrglugluthrasplyssergluthr
100105110
metgluthrgluaspvalprothrserserargleutyrtrpalaasp
115120125
leulysthrleuleuserglulysleuasnserilegluphealaasp
130135140
thrilelysglnleuserglnasnthrtyrthrproargglualagly
145150155160
serglnlysaspgluserleualatyrtyrilegluasnglnphehis
165170175
gluphelyspheserlysvaltrpargaspgluhistyrvallysile
180185190
glnvallysserseralaglnasnservalileilevalasplysasn
195200205
glyargleuvaltyrleuvalgluasnproglyglytyrvalalatyr
210215220
serlysalaalathrvalthrglylysleuvalhisalaasnphegly
225230235240
thrlyslysaspphegluaspleutyrthrprovalasnglyserile
245250255
valilevalargalaglylysilethrphealaglulysvalalaasn
260265270
alagluserleuasnalaileglyvalleuiletyrmetaspglnthr
275280285
lyspheproilevalasnalagluleuserphepheglyhisalahis
290295300
leuglythrglyaspprotyrthrproglypheproserpheasnhis
305310315320
thrglnpheproproserargserserglyleuproasnileproval
325330335
glnthrileserargalaalaalaglulysleupheglyasnmetglu
340345350
glyaspcysproserasptrplysthraspserthrcysargmetval
355360365
thrsergluserlysasnvallysleuthrvalserasnvalleulys
370375380
gluargargileleuasnilepheglyvalilelysglytyrgluglu
385390395400
proaspargtyrvalvalvalglyalaglnargaspalaleuglyala
405410415
glyvalalaalalysserservalglythrglyleuleuleulysleu
420425430
alaglnvalpheseraspmetileserlysaspglypheargproser
435440445
argserileilephealasertrpthralaglyasppheglyalaval
450455460
glyalathrglutrpleugluglytyrleuserserleuhisleulys
465470475480
alaphethrtyrileasnleuasplysvalvalleuglythrserasn
485490495
phelysvalseralaserproleuleutyrthrleumetglylysile
500505510
metglnaspvallyshisprovalaspglylysserleutyrargasp
515520525
serasntrpileserlysvalglulysleuserpheaspasnalaala
530535540
tyrpropheleualatyrserglyileproalavalserphecysphe
545550555560
cysgluaspalaasptyrprotyrleuglythrargleuaspthrtyr
565570575
glualaleuthrglnlysvalproglnleuasnglnmetvalargthr
580585590
alaalagluvalalaglyglnleuileilelysleuthrhisaspval
595600605
gluleuasnleuasptyrglumettyrasnserlysleuleuserphe
610615620
metlysaspleuasnglnphelysthraspileargaspmetglyleu
625630635640
serleuglntrpleutyrseralaargglyasptyrpheargalathr
645650655
serargleuthrthraspphehisasnalaglulysthrasnargphe
660665670
valmetarggluileasnaspargilemetlysvalglutyrhisphe
675680685
leuserprotyrvalserproarggluserprophearghisilephe
690695700
trpglyserglyserhisthrleuseralaleuvalgluasnleulys
705710715720
leuargglnlysasnilethralapheasngluthrleupheargasn
725730735
glnleualaleualathrtrpthrileglnglyvalalaasnalaleu
740745750
serglyaspiletrpasnileaspasngluphe
755760
<210>4
<211>2329
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的供体dna序列
<220>
<221>misc_feature
<222>(1)..(817)
<223>左同源臂
<220>
<221>misc_feature
<222>(821)..(1522)
<223>密码子优化的序列
<220>
<221>misc_feature
<222>(941)..(1492)
<223>人顶端结构域
<220>
<221>misc_feature
<222>(1523)..(2329)
<223>右同源臂
<400>4
ctatacagatatataaggatggggctttttttttttaatttttaaaaaagatttgtttat60
tattatatgtaagtacactgtagctgtcttcagacactccagaagagggcatcagatctc120
attacagatggttgtgagctaccatgtggtcactgggatttgaactcaggaccttcagaa180
gagcagtcagtgctcttaactgataagttaataataagttaactgataaggtaataaagg240
tcccctatgaaaagggttcagacccaaagagtcagagatccacaggttgagaacctcctg300
ccctaaatcttgttgctctccttattcaagaccactcctgttgcagttgctcttaagcat360
gagtatgctcccttctgaaagtctccatagcagccatctctccagccccagagtgaggct420
tttaaaggaatcttcatgataaatagaatttttaaaaaagtaactgaagttacttaaggt480
gttaaggtacattttattccctcagtaactggttaatctagcagttttgagtcatacttc540
atttatcttgactttgaagagtaagatattaaaacaatttgcttgatccttgaagtaagt600
atttaaatagacattttaatgcagactttttttagttgactggtggtgttgcacgtggtc660
aatccaagtactcatgggaggcagaggcaggaggatctctctctagaccagcctggtcta720
tagagcaagttccaggacagccagggctacacagaaaccttgtttcaaacaagactttta780
tccttccaggcagctgagccagaatacatacactcctagggaagctggttcacagaagga840
cgaatccctggcatactacatcgagaatcagtttcacgagttcaagtttagcaaagtctg900
gagagatgagcactacgtgaagatccaggtgaagagctccgctcagaactccgtgatcat960
cgtggataagaacggccggctggtgtacctggtggagaaccctggcggatacgtggctta1020
ctctaaggccgctaccgtgacaggcaagctggtgcacgccaacttcggaaccaagaagga1080
ctttgaggatctgtacacaccagtgaacggctctatcgtgatcgtgcgcgctggaaagat1140
caccttcgccgagaaggtggctaacgccgagagcctgaacgccatcggcgtgctgatcta1200
catggatcagacaaagtttcccatcgtgaacgctgagctgtctttctttggacacgctca1260
cctgggcaccggagacccatacacacccggattccctagctttaaccacacccagttccc1320
cccttccaggtctagcggactgccaaacatccccgtgcagacaatcagcagagccgctgc1380
cgagaagctgtttggcaacatggagggagactgcccctccgattggaagaccgactctac1440
atgtaggatggtgacctccgagtcaaaaaatgtcaaactcaccgtgtccaatgtgctgaa1500
agaacgacgcatcctgaatatctttggagttattaaaggttatgaggaaccaggtaaaga1560
cctgctttgtactttttcactttactgttttgcttactgtagataggtctagtgcaggaa1620
ggagaaggatgctagcttggcatgaactgctatatcttgtttgtcctaatgtgaactttg1680
taatatatgtgtatataacacataatatggccatgtaagtgtatggagaggccagagtta1740
agtattaaatatctttctgtaatcatttaaaattttacatatgaaggtcagtgaacagat1800
tgaaggagttttgtccaggtgggacttggatctaaattttttacaatgcctggcagcaaa1860
cacctttttaatcaactgagctgtctccccaaataaagtgaatgtgatatcagcttgtgg1920
ataatttttttttgttgctttgataagtggttttcttacaggatcacataccagttctgt1980
ccatagcattaaacaaacataactgtcatgcagtagattaatgtgcagggcacatccaac2040
agtcacatttattaataggacaaaaagttggaccttatatgtagcacacctataattcca2100
gtgctaggaagatccgggtaggagatccttagttcggtgctacttagtgagggtttgttt2160
caaaaaacaaaagctatgatggtgtgttgccttttttcttttagaccgttatgttgtagt2220
aggagcccagagagacgctttgggtgctggtgttgcggcgaagtccagtgtgggaacagg2280
tcttctgttgaaacttgcccaagtattctcagatatgatttcaaaaggt2329
<210>5
<211>763
<212>prt
<213>小家鼠(musmusculus)
<220>
<221>domain
<222>(1)..(67)
<223>细胞质结构域
<220>
<221>domain
<222>(68)..(88)
<223>跨膜结构域
<220>
<221>domain
<222>(89)..(763)
<223>细胞外结构域
<220>
<221>domain
<222>(199)..(381)
<223>顶端结构域
<400>5
metmetaspglnalaargseralapheserasnleupheglyglyglu
151015
proleusertyrthrargpheserleualaargglnvalaspglyasp
202530
asnserhisvalglumetlysleualaalaaspgluglugluasnala
354045
aspasnasnmetlysalaservalarglysprolysargpheasngly
505560
argleucysphealaalailealaleuvalilephepheleuilegly
65707580
phemetserglytyrleuglytyrcyslysargvalgluglnlysglu
859095
glucysvallysleualagluthrglugluthrasplyssergluthr
100105110
metgluthrgluaspvalprothrserserargleutyrtrpalaasp
115120125
leulysthrleuleuserglulysleuasnserilegluphealaasp
130135140
thrilelysglnleuserglnasnthrtyrthrproargglualagly
145150155160
serglnlysaspgluserleualatyrtyrilegluasnglnphehis
165170175
gluphelyspheserlysvaltrpargaspgluhistyrvallysile
180185190
glnvallysserserileglyglnasnmetvalthrilevalglnser
195200205
asnglyasnleuaspprovalgluserprogluglytyrvalalaphe
210215220
serlysprothrgluvalserglylysleuvalhisalaasnphegly
225230235240
thrlyslysasppheglugluleusertyrservalasnglyserleu
245250255
valilevalargalaglygluilethrphealaglulysvalalaasn
260265270
alaglnserpheasnalaileglyvalleuiletyrmetasplysasn
275280285
lyspheprovalvalglualaaspleualaleupheglyhisalahis
290295300
leuglythrglyaspprotyrthrproglypheproserpheasnhis
305310315320
thrglnpheproproserglnserserglyleuproasnileproval
325330335
glnthrileserargalaalaalaglulysleupheglylysmetglu
340345350
glysercysproalaargtrpasnileaspsersercyslysleuglu
355360365
leuserglnasnglnasnvallysleuilevallysasnvalleulys
370375380
gluargargileleuasnilepheglyvalilelysglytyrgluglu
385390395400
proaspargtyrvalvalvalglyalaglnargaspalaleuglyala
405410415
glyvalalaalalysserservalglythrglyleuleuleulysleu
420425430
alaglnvalpheseraspmetileserlysaspglypheargproser
435440445
argserileilephealasertrpthralaglyasppheglyalaval
450455460
glyalathrglutrpleugluglytyrleuserserleuhisleulys
465470475480
alaphethrtyrileasnleuasplysvalvalleuglythrserasn
485490495
phelysvalseralaserproleuleutyrthrleumetglylysile
500505510
metglnaspvallyshisprovalaspglylysserleutyrargasp
515520525
serasntrpileserlysvalglulysleuserpheaspasnalaala
530535540
tyrpropheleualatyrserglyileproalavalserphecysphe
545550555560
cysgluaspalaasptyrprotyrleuglythrargleuaspthrtyr
565570575
glualaleuthrglnlysvalproglnleuasnglnmetvalargthr
580585590
alaalagluvalalaglyglnleuileilelysleuthrhisaspval
595600605
gluleuasnleuasptyrglumettyrasnserlysleuleuserphe
610615620
metlysaspleuasnglnphelysthraspileargaspmetglyleu
625630635640
serleuglntrpleutyrseralaargglyasptyrpheargalathr
645650655
serargleuthrthraspphehisasnalaglulysthrasnargphe
660665670
valmetarggluileasnaspargilemetlysvalglutyrhisphe
675680685
leuserprotyrvalserproarggluserprophearghisilephe
690695700
trpglyserglyserhisthrleuseralaleuvalgluasnleulys
705710715720
leuargglnlysasnilethralapheasngluthrleupheargasn
725730735
glnleualaleualathrtrpthrileglnglyvalalaasnalaleu
740745750
serglyaspiletrpasnileaspasngluphe
755760
<210>6
<211>760
<212>prt
<213>智人(homosapiens)
<220>
<221>domain
<222>(1)..(67)
<223>细胞质结构域
<220>
<221>domain
<222>(68)..(89)
<223>跨膜结构域
<220>
<221>domain
<222>(90)..(760)
<223>细胞外结构域
<220>
<221>domain
<222>(196)..(379)
<223>顶端结构域
<400>6
metmetaspglnalaargseralapheserasnleupheglyglyglu
151015
proleusertyrthrargpheserleualaargglnvalaspglyasp
202530
asnserhisvalglumetlysleualavalaspgluglugluasnala
354045
aspasnasnthrlysalaasnvalthrlysprolysargcyssergly
505560
serilecystyrglythrilealavalilevalphepheleuilegly
65707580
phemetileglytyrleuglytyrcyslysglyvalgluprolysthr
859095
glucysgluargleualaglythrgluserprovalarggluglupro
100105110
glygluasppheproalaalaargargleutyrtrpaspaspleulys
115120125
arglysleuserglulysleuaspserthraspphethrglythrile
130135140
lysleuleuasngluasnsertyrvalproargglualaglysergln
145150155160
lysaspgluasnleualaleutyrvalgluasnglnphearggluphe
165170175
lysleuserlysvaltrpargaspglnhisphevallysileglnval
180185190
lysaspseralaglnasnservalileilevalasplysasnglyarg
195200205
leuvaltyrleuvalgluasnproglyglytyrvalalatyrserlys
210215220
alaalathrvalthrglylysleuvalhisalaasnpheglythrlys
225230235240
lysaspphegluaspleutyrthrprovalasnglyserilevalile
245250255
valargalaglylysilethrphealaglulysvalalaasnalaglu
260265270
serleuasnalaileglyvalleuiletyrmetaspglnthrlysphe
275280285
proilevalasnalagluleuserphepheglyhisalahisleugly
290295300
thrglyaspprotyrthrproglypheproserpheasnhisthrgln
305310315320
pheproproserargserserglyleuproasnileprovalglnthr
325330335
ileserargalaalaalaglulysleupheglyasnmetgluglyasp
340345350
cysproserasptrplysthraspserthrcysargmetvalthrser
355360365
gluserlysasnvallysleuthrvalserasnvalleulysgluile
370375380
lysileleuasnilepheglyvalilelysglyphevalgluproasp
385390395400
histyrvalvalvalglyalaglnargaspalatrpglyproglyala
405410415
alalysserglyvalglythralaleuleuleulysleualaglnmet
420425430
pheseraspmetvalleulysaspglypheglnproserargserile
435440445
ilephealasertrpseralaglyasppheglyservalglyalathr
450455460
glutrpleugluglytyrleuserserleuhisleulysalaphethr
465470475480
tyrileasnleuasplysalavalleuglythrserasnphelysval
485490495
seralaserproleuleutyrthrleuileglulysthrmetglnasn
500505510
vallyshisprovalthrglyglnpheleutyrglnaspserasntrp
515520525
alaserlysvalglulysleuthrleuaspasnalaalapheprophe
530535540
leualatyrserglyileproalavalserphecysphecysgluasp
545550555560
thrasptyrprotyrleuglythrthrmetaspthrtyrlysgluleu
565570575
ilegluargileprogluleuasnlysvalalaargalaalaalaglu
580585590
valalaglyglnphevalilelysleuthrhisaspvalgluleuasn
595600605
leuasptyrgluargtyrasnserglnleuleuserphevalargasp
610615620
leuasnglntyrargalaaspilelysglumetglyleuserleugln
625630635640
trpleutyrseralaargglyaspphepheargalathrserargleu
645650655
thrthrasppheglyasnalaglulysthraspargphevalmetlys
660665670
lysleuasnaspargvalmetargvalglutyrhispheleuserpro
675680685
tyrvalserprolysgluserprophearghisvalphetrpglyser
690695700
glyserhisthrleuproalaleuleugluasnleulysleuarglys
705710715720
glnasnasnglyalapheasngluthrleupheargasnglnleuala
725730735
leualathrtrpthrileglnglyalaalaasnalaleuserglyasp
740745750
valtrpaspileaspasngluphe
755760
<210>7
<211>184
<212>prt
<213>恒河猴(macacamulatta)
<400>7
alaglnasnservalileilevalasplysasnglyglyleuvaltyr
151015
leuvalgluasnproglyglytyrvalalatyrserlysalaalathr
202530
valthrglylysleuvalhisalaasnpheglythrlyslysaspphe
354045
gluaspleuaspserprovalasnglyserilevalilevalargala
505560
glylysilethrphealaglulysvalalaasnalagluserleuasn
65707580
alaileglyvalleuiletyrmetaspglnthrlyspheproileval
859095
lysalaaspleuserphepheglyhisalahisleuglythrglyasp
100105110
protyrthrproglypheproserpheasnhisthrglnphepropro
115120125
serglnserserglyleuproasnileprovalglnthrileserarg
130135140
alaalaalaglulysleupheglyasnmetgluglyaspcysproser
145150155160
asptrplysthraspserthrcyslysmetvalthrsergluasnlys
165170175
servallysleuthrvalserasn
180
<210>8
<211>184
<212>prt
<213>黑猩猩(pantroglodytes)
<400>8
alaglnasnservalileilevalasplysasnglyserleuvaltyr
151015
leuvalgluasnproglyglytyrvalalatyrserlysalaalathr
202530
valthrglylysleuvalhisalaasnpheglythrlyslysaspphe
354045
gluaspleuhisthrprovalasnglyserilevalilevalargala
505560
glylysilethrphealaglulysvalalaasnalagluserleuasn
65707580
alaileglyvalleuiletyrmetaspglnthrlyspheproileval
859095
asnalagluleuserphepheglyhisalahisleuglythrglyasp
100105110
protyrthrproglypheproserpheasnhisthrglnphepropro
115120125
serargserserglyleuproasnileprovalglnthrvalserarg
130135140
alaalaalaglulysleupheglyasnmetgluglyaspcysproser
145150155160
asptrplysthraspserthrcysargmetvalthrsergluserlys
165170175
asnvallysleuthrvalserasn
180
<210>9
<211>184
<212>prt
<213>食蟹猴(macacafascicularis)
<400>9
alaglnasnservalileilevalasplysasnglyglyleuvaltyr
151015
leuvalgluasnproglyglytyrvalalatyrserlysalaalathr
202530
valthrglylysleuvalhisalaasnpheglythrlyslysaspphe
354045
gluaspleuaspserprovalasnglyserilevalilevalargala
505560
glylysilethrphealaglulysvalalaasnalagluserleuasn
65707580
alaileglyvalleuiletyrmetaspglnthrlyspheproileval
859095
lysalaaspleuserphepheglyhisalahisleuglythrglyasp
100105110
protyrthrproglypheproserpheasnhisthrglnphepropro
115120125
serglnserserglyleuproasnileprovalglnthrileserarg
130135140
alaalaalaglulysleupheglyasnmetgluglyaspcysproser
145150155160
asptrplysthraspserthrcyslysmetvalthrsergluasnlys
165170175
servallysleuthrvalserasn
180
<210>10
<211>23
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的sgrna序列
<400>10
gaatacatacactcctcgtgagg23
<210>11
<211>23
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>合成的sgrna序列
<400>11
agaagaatacttaacatctttgg23
- 还没有人留言评论。精彩留言会获得点赞!