包含转基因事件CIGBDt-Def1或CIGBIs-Def5的大豆植物的制作方法
本发明涉及植物生物技术领域,尤其涉及保护大豆对抗由真菌和卵菌引起的疾病(在该作物的商业基因型中)。特别地,本发明涉及两个新的转基因事件,其产生自两个大豆基因型的遗传转化。所述事件被称作cigbdt-def1和cigbis-def5。
现有技术
植物经常暴露于在自然界中存在的不同的病原体。因此,它们已发展出用于保护其对抗病原体攻击的复杂的防御系统。按照其表达的时刻,可以将这些防御系统分类为组成型的,如果其在植物的整个生活周期期间存在;和诱导型的,如果表明其表达在更大的比例上是面对病原体的存在。这些机制之一是植物防卫素。这些化合物为45至54个氨基酸残基的小的阳离子肽,其中在构成其的半胱氨酸中具有二硫桥(zhu等人,cellmol.lifesci.(2015)62:2257-2269)。首次在小麦和大麦的种子中描述了植物防卫素,并且其在该类型生物的防御系统中的作用已得到充分表征。按照肽的作用方式,它们与其在生物系统中所负责的作用和表达特性谱相符地分布(beer和vivier,bmcres.notes(2011)4:459)。
大豆植物被各种不同的疾病侵袭,它们对产量和种子品质产生负面影响,从而造成总的生产损失。最重要的疾病是由真菌引起的那些,这是由于其造成的损害的量级。对于改善大豆作物的常规计划,获得显示出对于侵袭该作物的主要真菌疾病的抗性的商业基因型是极其困难的。
到目前为止,化学杀真菌剂的应用是用于控制大豆的致病真菌的最有效的处理。但是,已减少了其应用,这是由于这些产品的活性成分的高的数量,这导致获得病原体对于一些杀真菌剂的抗性。此外,它们的使用增加了生产成本,因此开发抗性品种被认为是最经济的控制策略。
在侵袭大豆作物的真菌之中,刺盘孢属(colletotrichum)有着巨大的重要性,因为它是炭疽病(其是自植物发育的初始阶段起出现的热带和亚热带国家的典型疾病)的致病因子。该疾病对籽粒的品质造成巨大侵害,引起胚芽的死亡,这是由于在当感染较轻时在种子中缺乏症状。以这种方式,真菌可以造成种子中的劣化和成年植物中的全身感染,并且当出现雨、种群密度过大或收获推迟时,损害加重,这影响产量(gally等人,manejointegradodeplagasyagroecología(2006)78:86-90)。
大豆的“周期末尾”疾病是最时常发生的,并且可以引起产量的大约8%至10%,最大直至30%的损害。症状根据所存在的疾病而变化,并且在中间和晚期的生殖状态下表现出来。最突出的是健康叶面积的减少、植物的过早落叶、作物的提早成熟和种子品质的下降。在构成所谓的“周期末尾复合体”的疾病之中,尤其包括茎的黑粉病和种子的紫斑(由菊池尾孢(cercosporakikuchii)引起)、蛙眼斑(由大豆尾孢(cercosporasojina)引起)、环斑(由山扁豆生棒孢(corynesporacassiicola)引起)、棕色斑(由大豆壳针孢(septoriaglycines)引起)、豆荚和茎的黑粉病(由大豆拟茎点霉(phomopsissojae)引起)、霉病(由东北霜霉(peronosporamanshurica)引起)、由链格孢属真菌引起的叶斑(由链格孢属物种(alternariaspp.)引起)、炭疽病(由平头刺盘孢(colletotrichumtruncatum)引起)和亚洲锈病(carmona等人,phytophatologica(2011)37:134-139)。
豆薯层锈菌(phakopsorapachyrhizi)是大豆亚洲锈病的致病因子(torres等人,pesq.agropec.bras(2012)38:1053-1057)。这种疾病被认为是侵袭作物的最具破坏性的疾病,因为它在提早落叶方面和在侵害产量的主要组分方面产生影响,并且造成10-90%的生产损失(ribeiro等人,geneticsandmolecularbiology(2008)31:98-105)。该疾病在2001年在美洲大陆被检测到,并且已经给南美洲的大豆生产造成了巨大的损失。在古巴,在2010年检测到亚洲锈病的存在(pérez等人,plantpathology(2011)59:803)并且在各个大豆生产省份中传播。
已经在大豆中鉴定出了两种锈病类型。除了亚洲锈病外,还存在由山马蝗层锈菌(p.meibomiae)引起的大豆美洲锈病。用于区分这些锈病种类的结构是末孢子(telospore)。但是,这些是难以发现的,这是由于不可能在体外培养这两种病原体。为了正确鉴定,可以使用分子技术,例如使用特异于豆薯层锈菌的寡核苷酸的聚合酶链式反应(pcr)。在大田中亚洲锈病的存在可以通过使用免疫色谱法诊断条来进行测定,这些能够在感染的初始阶段中检测病原体的存在,并且被设计用于在叶组织中使用。
在最佳环境条件(最少6小时的露水和18至25℃的温度)下,豆薯层锈菌感染迅速进展,并引起提早落叶。作为结果,这导致豆荚、籽粒的数目以及其重量的减少(pandey等人,molecularplant-microbeinteractions(2010)24:194-206)。
数个实验表明,植物防卫素参与保护宿主对抗病原体的攻击。例如,分离自荠叶喜日花(heliophilacoronopifolia)这一物种的防卫素hc-afp1-4显示出针对灰葡萄孢(botrytiscinerea)和腐皮镰孢(fusariumsolani)的体外抑制活性。在被芸苔生链格孢(alternariabrassicola)感染后,防卫素rs-afp1和rs-afp2在萝卜叶中被强烈地诱导,这显示出“体外”抗真菌活性。
由portieles等人进行的研究证明,分离自特大管烟草(nicotianamegalosiphon)的防卫素基因nmdef02的组成型表达给转基因马铃薯和烟草植物提供了在大田条件下针对致病疫霉(phytophthorainfestans)、茄链格孢(alternariasolani)和尖镰孢(fusariumoxysporum)的抗性。这些作者还观察到防卫素nmdef02的强的体外抗微生物活性,针对病原体例如致病疫霉、寄生疫霉烟草变种(phytophthoraparasiticavar.nicotianae)、茄链格孢、尖镰孢古巴变种(fusariumoxysporumvar.cubense)、大丽花轮枝孢(verticilliumdahliae)(portieles等人,plantbiotechnologyjournal(2010)8:678-690)。这确证了防卫素在对抗植物病原体的保护作用中的重要作用。
另一方面,对于除草剂具有抗性的转基因大豆品系的获得是在现代农业中生物技术的应用之一。在植物中表达与莽草酸的代谢途径相关联的土壤杆菌属(agrobacterium)基因在经修饰的作物中赋予对于以大田剂量的除草剂草甘膦(n-膦酰基甲基甘氨酸)的抗性。草甘膦通过竞争性地抑制5-烯醇丙酮酸莽草酸-3-磷酸合成酶而打断了植物中芳香族氨基酸的合成(funke等人,pnas(2006)103(35):13010-5)。在植物的分生组织中该除草剂的存在抑制生长,并且引起植物的死亡。但是,包含基因cp4epsps(其序列以登录号ab209952.1保存在ncbi的genbank数据库中)的转基因大豆植物在该除草剂存在下继续芳香族氨基酸的合成。
具有商业价值的大豆基因型的遗传转化经证明是方便的,以获得转基因材料,其具有针对一系列的影响大豆作物的真菌和卵菌的保护作用,并且其中可以利用除草剂草甘膦的使用来控制影响所述作物的杂草。
发明描述
通过提供以包含转基因事件cigbdt-def1或转基因事件cigbis-def5为特征的大豆植物或所述植物的部分,本发明解决了上面提出的问题。这两个事件的代表性大豆种子保藏于英国国家工业、食品和和海洋微生物保藏中心(ncimb;nationalcollectionsofindustrial,foodandmarinebacteria),其是被承认作为国际保藏单位的机构。事件cigbdt-def1的代表性种子以登录号ncimb42724进行保藏,和事件cigbis-def5的代表性种子以登录号ncimb42725进行保藏。所述转基因事件通过遗传工程来获得,并且以独一无二的方式赋予包含它们的植物以自身杀真菌剂活性。以这种方式,转基因植物能够避免真菌和卵菌病原体在大豆作物中引起的负面效应。本发明的这两个转基因事件表达防卫素基因nmdef02和基因cp4epsps(其负责对于除草剂草甘膦的抗性),并且通过土壤杆菌方法或通过生物射弹来产生。
为了本发明的目的,“大豆植物”是指大豆(glycinemax)这一物种的所有植物,并且包括具有商业价值的所有大豆品种,还包括所述植物的所有部分。
在本发明中,被命名为cigbdt-def1的大豆事件也被称为cigb-dtdef1、dtdef1、dtdef1和dtdef1。在其方面,被命名为cigbis-def5的大豆事件也被称为cigb-isdef5、isdef5和isdef5。除了这些事件中的每一个的特征性序列外,本发明的大豆植物还可以在所述特征性序列侧翼包含另外的脱氧核糖核酸(dna)序列。也获得了的并且在本发明的实施例中提及的其他转基因事件通过采用dt(或dt)和is(或is)和相应事件的编号的组合来命名,例如dt12、is20等。
事件cigbdt-def1和cigbis-def5是从两个常规大豆品种dt84和incasoy36(is36)开始来获得的。由基因nmdef02所编码的防卫素的表达对于对抗与作物相关的植物病原体真菌的保护作用来说是重要的,所述植物病原体真菌限制生产水平以及籽粒品质。包含这些转基因事件的植物耐受直至五倍大田有效剂量的非选择性除草剂草甘膦,这允许在生产条件下使用这种除草剂来控制杂草。
在一个实施方案中,本发明公开了大豆植物,其特征在于,其为包含事件cigbdt-def1或事件cigbis-def5的大豆植物的任何世代的子代。在另一个实施方案中,本发明提供了大豆植物,其特征在于,其产生自包含转基因事件cigbdt-def1或cigbis-def5的大豆植物与另一非转基因大豆植物的杂交。
术语“转基因(的)”包括任何已通过异源核酸的存在而被修饰的品系、植株、细胞或事件,其包括最初经修饰的转基因的那些,以及通过初始再生出的事件的有性杂交或无性繁殖而产生的那些。
在本发明中,术语“子代”是指通过在包含事件cigbdt-def1和cigbis-def5的植物与另一商业大豆基因型之间的有性杂交(例如,回交、自交或互交)而产生的植物。
如在本文中所使用的,术语“植物的部分”包括植物的细胞、植物的器官、植物原生质体、植物细胞组织培养物(从其开始可以再产生出所述植物)、植物的愈伤组织、胚芽和植物中的完整植物细胞。术语“植物的部分”还包括胚、花粉、胚珠、种子、种子豆荚、叶、花、枝、茎、根、根尖、花药、子叶、下胚轴等。术语“籽粒”意指由商业栽培者以与所述物种的生长和繁殖不同的目的而产生的成熟种子。
在一个实施方案中,本发明涉及包含事件cigbdt-def1或事件cigbis-def5的植物的部分,其中所述部分可以为根、芽、叶、花粉、胚珠、花或细胞。本发明还提供了包含事件cigbdt-def1或cigbis-def5的大豆植物的种子。
为了本发明的目的,术语“转基因品系”是指这样的植物,其基因组已通过遗传工程进行了修饰,以引入不相关的其他植物或不同物种的一个或几个新基因;或者修饰本身基因的功能。作为基因的插入或修饰的结果,转基因品系显示出新的特性,其从确定的克隆世代开始被有规律地传递给后代。
在本发明中,除了用本发明的事件而已经拥有的表型外,大豆植物还可以获取新的表型,并且为此可以采用对源自包含所述事件的植物的植物材料或外植体进行转化。除了所述及的事件外,这些特征也可以通过可得的遗传转化方法中的任一种来获取。术语“转化”是指将核酸片段转移到宿主生物的基因组中,作为结果导致在遗传学上稳定的遗传。
包含事件cigbdt-def1或cigbis-def5的植物可以在转化方法中用作植物材料的来源,以将异源核酸分子引入至新的大豆事件。可以制备转化载体以引入其他目的基因,并因此取得具有被引入到相同植物中的不同特征的其他品系。
“转基因事件”通过下述方式来产生:用包含异源dna的遗传构建体(包括包含目的转基因的核酸表达盒)来转化植物细胞,使由于将转基因插入到植物的基因组中而得到的植物群体再生,并且选择特定的再生出的植物。所述事件以插入物在特定基因组中的定位为特征,和在表型上以所述转基因的表达为特征。在基因水平上,事件是构成植物的遗传组成的一部分的dna区段。在本发明的情况下,所述转基因事件包含两个转基因。
在另一个方面,本发明提供了产生自包含事件cigbdt-def1或cigbis-def5的大豆植物或大豆种子的大豆产品。在本发明的一个实施方案中,所述产品在于磨粉、薄片、油或用于人或动物膳食的产品。
本发明的另一个方面为用于产生对于除草剂草甘膦和由真菌或卵菌引起的疾病具有抗性的大豆植物的方法,其中在所述植物的基因组中引入事件cigbdt-def1或cigbis-def5。在本发明的一个实施方案中,所述大豆疾病为由豆薯层锈菌引起的亚洲锈病。为了本发明的目的,对亚洲锈病具有抗性的植物被认为是其中在天然感染条件下该疾病的症状的存在不超过50%的植物的品系的那些植物。
在本发明的一个实施方案中,所述方法包括使包含事件cigbdt-def1或事件cigbis-def5的大豆植物与另一大豆植物杂交,并且选择包含这些事件之一的子代。
用于在大豆基因组核酸样品中检测相应于事件cigbdt-def1或cigbis-def5的核酸的试剂套盒也是本发明的目标,其中所述套盒包含至少一个用于扩增相应于在大豆植物的核酸与相应于事件cigbdt-def1或cigbis-def5的核酸之间的连接区域的核酸片段的寡核苷酸对。
可能的是,设计和使用有用的寡核苷酸或引物对,包括与在插入片段的dna和侧翼dna的5’末端之间或者在插入片段的dna和侧翼dna的3’末端之间的连接点相重叠的引物。用于设计、获得和使用在本发明中的dna探针或引物的方法是分子生物学中已知的。为此,pcr引物对可以源自已知序列,例如通过使用旨在该目的的计算机程序,例如在vectornti的第11版中的pcr引物分析工具(thermofisherscientific)。
在另一个方面,本发明提供了用于在大豆样品中诊断事件cigbdt-def1或事件cigbis-def5的方法,其包括分析所述样品以检测相应于事件cigbdt-def1或cigbis-def5的核酸或蛋白质的存在。
如在本文件中所使用的,“样品”包括任何这样的级分,其包含核酸或多肽,并且获得自植物、植物材料或产品,例如用于动物消耗的饲料或者源自植物材料的新鲜或经加工的产品。
根据所选择的用于诊断事件cigbdt-def1或cigbis-def5的方法,可能的是例如分析获得自大豆植物(包括子代,除了所述植物的衍生物外)的样品。此类方法可应用于在任何生物学材料中鉴定或检测所述及的事件的核酸区域或分子。此类方法包括,例如用于确证纯度的方法,和用于在一批种子中筛选包含转基因事件cigbis-def5或cigbdt-def1的特征性核酸分子的种子的方法。在实施例3中显示了用于在生物学样品中鉴定本发明的事件的核酸的方法。该方法包括制备生物学样品和与所述核酸互补的引物对(其能够扩增对于事件cigbis-def5或cigbdt-def1的核酸分子来说特异的区域)的混合物。
在诊断或检测本发明的事件的方法之中,包括任何用于通过pcr来扩增事件cigbis-def5或cigbdt-def1的核酸分子的方法。此外,所述事件可以通过检测在所述大豆植物中表达的防卫素的多肽或肽来进行检测或鉴定。在检测所述多肽或肽中可以使用任何方法。例如,可以采用针对所述防卫素而产生的抗体,和利用任何已知的免疫化学方法,例如elisa、免疫印迹、斑点印迹。
在本发明的一个特别的实施方案中,在大豆样品中诊断事件cigbdt-def1或事件cigbis-def5以使用这样的试剂套盒为特征,所述试剂套盒包含至少一个用于扩增相应于在大豆植物的核酸与相应于事件cigbdt-def1或cigbis-def5的核酸之间的连接区域的核酸片段的寡核苷酸对。
本发明还提供了用于在大田中增加大豆作物的产量的方法,其包括:播种包含事件cigbdt-def1或cigbis-def5的大豆植物的种子,将大豆作物定植在该大田中,和用有效量的除草剂草甘膦处理该大田以控制杂草。在本发明的一个实施方案中,所述方法的特征在于,在作物期v1和r4之间处理该大田。将大豆作物定植和施加除草剂的方法是该技术领域中的专业人员所熟知的。
可以在改善计划中使用包含事件cigbdt-def1或cigbis-def5的大豆植物,其中使用杂交或杂合方法,以使这些事件在其他基因型的大豆植物中再现。此类改善方法可以用于产生大豆植物,例如为了将其用于在不同地理区域中的商业生产,或者为了产生另外的大豆繁殖群体。
此外,可以在繁殖计划中使用携带本发明的事件的植物,其中使用改善方法,以产生具有与对于除草剂草甘膦的耐受性和对于真菌的抗性相组合的另外的令人感兴趣的特征(也称为“堆叠性状”)的大豆植物。例如,另外的特征可以为对于另外的除草剂例如草铵膦的抗性,和与其他在农艺学上重要的性状(包括对于其他疾病和病原体的抗性)(例如,bt基因)的组合,以及涉及在大豆籽粒中存在的油和蛋白质的数量和品质的性状的改善。
所希望的特征(堆叠的)的这些组合可以通过任何方法来产生,包括但不限于通过任何已知的方法学或通过遗传转化来改善所述植物。如果通过植物的遗传转化来堆叠所述序列,那么可以在任何时刻和以任何顺序来组合事件cigbdt-def1或cigbis-def5的典型的核酸序列。可以同时地引入所述性状,通过与由转化盒的任何组合所提供的目的核酸分子的共转化。例如,如果引入两个另外的序列,那么可以将所述两个序列包含在分开的转化盒中(反式)或者包含在相同的转化盒中(顺式)。所述序列的表达可以由相同的启动子或不同的启动子来调控。
在某些情况下,可能希望的是,引入阻抑目的核酸分子的表达的转化盒。这可以用其他阻抑盒或过表达盒的任何组合来进行调整,以在植物中产生所希望的特征组合。此外,公认的是,可以在所希望的基因组位置处堆叠所述核酸分子,通过使用位点特异性重组系统。
附图简述
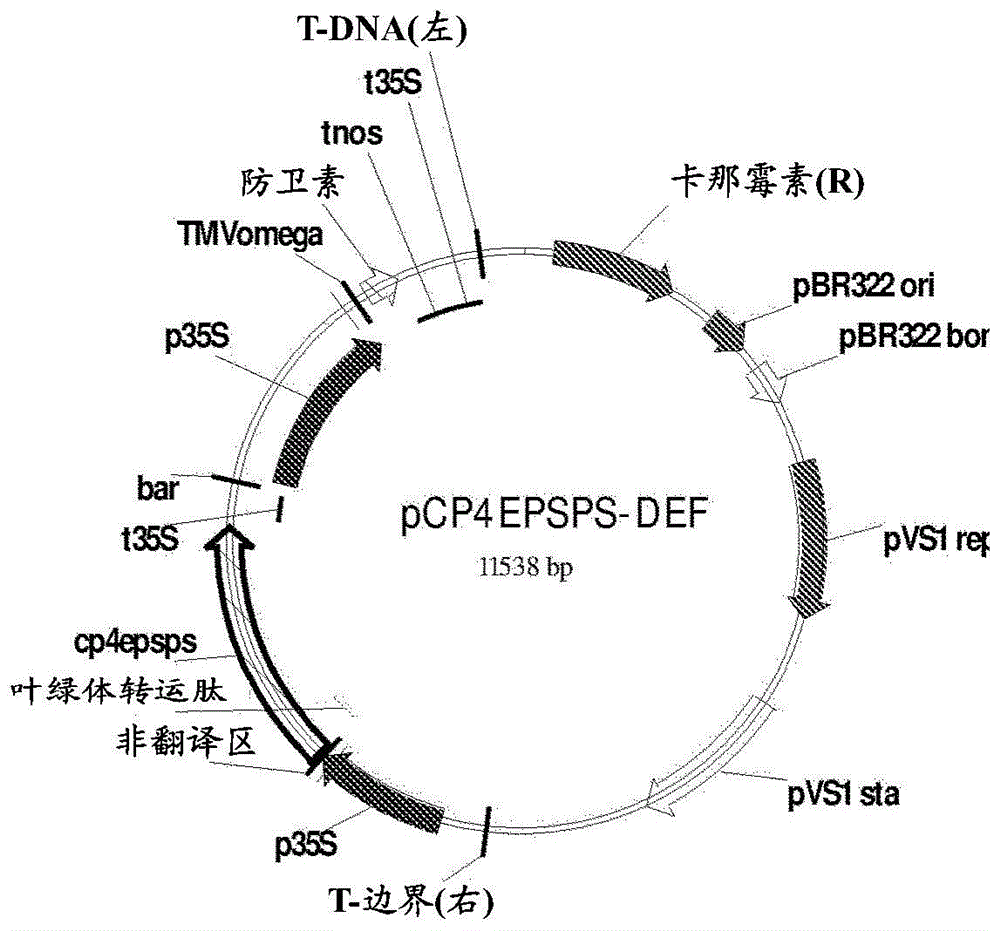
图1.用于通过土壤杆菌和生物射弹来转化大豆分生组织的质粒pcp4epsps-def的图谱。
图2.蛋白质cp4epsps的免疫检测,roundup
图3.从用质粒pcp4epsps-def进行转化的大豆植物和非转基因对照的基因组dna开始的pcr反应的产物。扩增相应于基因nmdef02的140bp的序列。c+:质粒pcp4epsps-def。m:分子量标准参照物(promega)。泳道dtdef1、dt3、dt4、dt5、dt16、dt17、dt18和isdef5:转基因大豆品系。c-:未转化的植物。
图4.包含两个关于酶ecorv的限制位点的质粒pcp4epsps-def的片段(5.3kb)和用作探针的基因cp4epsps的片段(886bp)的图示。
图5.用于检测基因cp4epsps和nmdef02整合入大豆事件中(t3)的southern印迹。dtdef1、dt12、dt15、dt16、dt17、dt18和isdef-5:转基因大豆品系。c-is36和c-dt84:未转化的植物。
图6.在用质粒pcp4epsps-def进行转化的转基因大豆植物中基因nmdef02的相对表达。将采用逆转录酶的定量聚合酶链式反应(qrt-pcr)用于测量防卫素基因nmdef02的转录物水平,相比于未转化的对照dt84的内源性肌动蛋白的组成型表达而言。转基因品系:dtdef1、dt3、dt4、dt5、dt6和isdef5。未转化的对照:dt84(nt)和is36(nt)。
图7.在t3代的转基因大豆植物中基因nmdef02的相对表达。条棒表示从三次重复获得的结果的平均值,对于p<0.0001。将qrt-pcr用于测量防卫素基因nmdef02的转录物水平,相比于未转化的对照的内源性肌动蛋白的组成型表达而言。转基因品系:dtdef1、dt3、dt4、dt5、dt6和isdef5。未转化的对照:dt84(nt)和is36(nt)。
图8.在转基因大豆品系dtdef1、dt3、dt4、dt5和dt6中和在未转化的对照dt84中存在的真菌豆薯层锈菌的生物量的定量。所述定量通过qrt-pcr来进行。
图9.在播种在实验地块中的大豆植物中亚洲锈病的发生率,其被表示为受侵袭的植物的百分比。转基因品系:dtdef1、dt3、dt6和isdef5。未转化的对照:dt84(nt)和is36(nt)。
图10.在转基因大豆植物中和在未转化的对照dt84(nt)和is36(nt)中亚洲锈病的发生率。显示了在几个转基因品系中被真菌豆薯层锈菌侵袭的植物的百分比。
图11.在播种在大田中的表达基因nmdef02和cp4epsps的转基因大豆植物和未转化的对照(dt84)中亚洲锈病的严重度的评价。
图12.在检测到该疾病后36和60天,在转基因品系中和在未转化的对照dt84(nt)中由于亚洲锈病的发生而引起的落叶(没有叶的植物)的百分比。
图13.在不同转基因品系和未转化的对照dt84(nt)的大豆植物中豆薯层锈菌的生物量的定量。所述定量通过qrt-pcr来进行。
图14.从大豆植物(品系dtdef1)和非转基因对照的dna开始的pcr反应的产物,其中使用在与在生物信息学分析中鉴定出的位点邻近的大豆基因组区域中所设计的引物。扩增相应于位点8002360的344bp的序列和相应于位点8334726的273bp的序列(在未转化的大豆植物中)。泳道:1kb:分子量标准参照物(promega);dtdef1:转基因大豆品系dtdef1;dt84:未转化的大豆植物;b:空白。
图15.从转基因品系cigbdtdef1和非转基因对照的植物的dna开始的pcr反应的产物,其是用大豆的引物与在载体pcp4epsps-def(图1)中的基因cp4epsps的引物的组合而获得的。对于转基因事件cigbdtdef1,扩增在位点8002360处的1000bp的序列(a)和在位点8334726处的900bp的序列(b)。泳道:1kb:分子量标准参照物(promega);dtdef1:转基因大豆品系;dt84:未转化的大豆植物;b:空白。
图16.从转基因品系cigbdtdef1和非转基因对照的dna开始的pcr反应的产物,其是用大豆的引物与在载体pcp4epsps-def(图1)中的基因nmdef02的引物的组合而获得的。对于转基因事件cigbdtdef1,扩增在位点8002360处的2000bp的序列(a)和在位点8334726处的300bp的序列(b)。泳道:1kb:分子量标准参照物(promega);dtdef1:转基因大豆品系;dt84:未转化的大豆植物;b:空白。
实施方式的详细描述/实施例实施例1.从基因型dt84和is36的胚性尖端再生出植物
从来自国家农业科学研究所(institutonacionaldecienciasagrícolas)的品种dt84和is36的经消毒的种子开始来获得胚轴。将种子通过下述方式来进行消毒:在70%乙醇中温育一分钟,然后在用无菌蒸馏水进行连续漂洗后在10%(v/v)过氧化氢溶液中7小时。让经消毒的种子静置,浸没在无菌蒸馏水中,持续28小时,在环境温度下和在黑暗中。着手在培养皿上去除种子的皮,并且分开子叶以取出胚,其在大豆转化和再生实验中用作外植体。后来,着手借助于体视镜来去除初生叶以使顶端分生组织暴露,并且将15个胚以顶端分生组织区域朝上的方式放置在包含ms培养基(murshige和skoog,plantphysiol(1962)15:473-479)的平皿中。
在10-15天后,在三种所测试的培养基组合中,进行从所述外植体的顶端分生组织区域开始的芽再生。在采用0.2mg/l的苄基氨基嘌呤(bap)和0.2mg吲哚丁酸(iba)/l的变化形式a中,在相对短的时间内(20天)和以1.6个芽/外植体的再生效率获得了高的再生频率(83%)。使经确定的芽开始在再生培养基中进行生根,并且在15天后观察到白而细的根的出现。对于在15天时还未生根的芽,切除基部,并将其转至新鲜培养基,其中90%令人满意地发育出了根。
在培养基中的bap/iba组合有利于从分生组织轴的顶端区开始的芽再生,但是它引起在胚根区中愈伤组织的大量形成。为此,评价了在没有生长素的再生培养基(变化形式b)中外植体的应答。在芽再生方面获得的结果在变化形式a和b中是相似的,但是在没有iba的变化形式中取得了更高的再生频率和更大的芽数目。
另一方面,在没有激素的培养基(变化形式c)中芽的再生高于具有激素的变化形式。在5mg/l的bap中诱导芽48小时足以在msb5培养基(补充有维生素b5的ms培养基)中取得有效的再生和更大的芽数目/外植体。用5mgbap/l进行预处理有利于在msb5培养基中在25天后形成多个原芽,其在当被转移至新鲜培养基时开始被确定。这些原芽再生出正常的芽、叶和不生长的小芽。
后来,在品种is36和dt84中采用以5mgbap/l中进行预处理来重复该再生实验。在优化了该再生系统后,结果是更好的。所有外植体都再生出了芽,并且分别取得了2.6和3.7个芽/外植体的再生效率,如在表1中所显示的。数据为两次实验的平均值。
表1.在5mgbap/l中诱导48小时的时间段后,从成熟种子的胚轴开始的芽再生。
实施例2.转基因大豆品系的开发
为了建立在大豆种子的分生组织中的转化实验方案,用质粒pcambia2301(cambia,canberra,australia)轰击100个外植体。β-葡糖醛酸糖苷酶(gus)的组织化学测定法确证了在具有5mgbap/l的芽诱导培养基中,在48小时后,在所述100个外植体中的56个之中酶促活性的存在。gus的瞬时表达在以强的分裂能力为特征的区域(例如,胚轴的顶端区和胚根的尖端)中是明显的。
为了测定用于选择经转化的芽的最佳的最小致死浓度,首先评价了大豆胚轴面对不同浓度的草甘膦的应答。用作再生对照的在没有选择的msb5培养基中的外植体显示出100%的芽再生。在该实验中,对于草甘膦的敏感性在15mg/l的浓度下变得明显,其中在35天后,仅16%的胚轴显示出芽和叶的发育。在20mg草甘膦/l下观察到相似的结果,其中在50天后具有6%的再生。所有外植体保持其绿色,尽管在胚轴的顶端区中显示出其对于该除草剂的敏感性。仅在25mg草甘膦/l下在60天后实现形态发生的完全抑制。在该浓度下,一些外植体在顶端区域中展示出坏死。最后,确定在msb5培养基中的25mg草甘膦/l足以抑制未转化的芽的正常分化。为此,在用基因cp4epsps的遗传转化实验中使用该浓度,以选择耐受草甘膦的芽。
一旦建立了用于选择的条件,就用质粒pcp4epsps轰击胚轴的顶端区域(soto等人,plantcelltissue&organculture(2017)128:187-196),并且获得携带基因cp4epsps的转基因大豆植物。芽的诱导在5mgbap/l下在48小时后出现,并且将外植体传代培养至具有25mg草甘膦/l的msb5培养基。在具有选择的msb5中的阴性对照没有显示出任何形态发生,并且外植体保持其绿色着色。在阳性对照中,在没有选择的msb5中在10天后观察到经充分确定的芽的发育(2cm)。
在这些实验中,胚轴的分生组织仅被轰击一次,这足以得到耐受除草剂的芽而没有在外植体中的损伤。数据显示,在705个经轰击的外植体中的32个(4.5%)在该除草剂存在下发育出芽,其代表了从5.0至8.5%变动的转化效率(其被定义为耐受草甘膦的植物的总数目除以经轰击的外植体的总数目)的平均值。在选择下15至20天后,在外植体中出现初生芽,通过直接的器官发生。将所有达到3至4cm的大小的再生出的芽转移至没有选择的生根培养基。在15天后,46株植物形成根,并且显示出与非转基因植物相似的表型。所述植物发育出主根(具有有着根毛的次生根),这有利于在短时间(10-15天)内新叶的生长和形成。
用质粒pcp4epsps-def进行遗传转化
如前面所提出的,对于大豆的遗传转化,确立了25mg草甘膦/l为用于体外选择耐受性芽的该除草剂的最小致死浓度。用在图1中所呈现的质粒pcp4epsps-def来轰击胚轴的顶端区域。接下来,描述在所述质粒中所掺入的遗传元件:t-边界(右):右边界;p35s:相应于花椰菜花叶病毒(camv35s)的启动子;cp4epsps:对于除草剂草甘膦的抗性基因;t35s:相应于camv35s的终止子;防卫素:关于对于真菌的抗性的防卫素基因nmdef02;tmvomega:烟草花叶病毒(tmv)的前导序列;tnos:胭脂碱合成酶的终止子;t-dna(左):左边界;卡那霉素(r):关于对于卡那霉素的抗性的基因nptii;pbr322ori:质粒pmb1的复制起点;pvs1:源自铜绿假单胞菌(pseudomonasaeruginosa)的复制起点。
在以bap进行诱导下48小时后,经轰击的外植体快速生长。在具有草甘膦选择的msb5培养基中并在光照和黑暗的光周期下7天后,所有胚轴都从白色变为深绿色。在阳性对照中,对于基因型dt84和is36,在没有选择试剂的msb5培养基中,在10天后观察到经确定的芽的发育(2cm)。另一方面,用作阴性对照的在具有25mg草甘膦/l的msb5培养基中进行培养的外植体未显示出芽的再生,尽管它们保持绿色着色。
在这些实验中,总共轰击了品种dt84的177个外植体和品种is36的219个外植体,其分别显示出10.7%和6.8%的再生频率;在25mg除草剂/l的选择压力下。在具有选择的msb5培养基中在15天后开始观察到芽再生,其中取得13.5%(dt84)和10.5%(is36)的转化效率。当在草甘膦下再生出的芽达到3-4cm的大小时,将它们转至在没有选择的生根培养基中进行生根。在10天后,所述植物生根并且具有正常的发育,与未转化的对照相似。
将栽培种is36的胚轴用于通过根癌土壤杆菌(agrobacteriumtumefaciens)进行的转化,为此用质粒pcp4epsps-def(图1)转化菌株lba4404。该程序在于,培养该细菌至0.8的在620nm下的光密度;和然后在200μμ的乙酰丁香酮存在下感染外植体30分钟。将外植体在黑暗条件下共培养48小时,并且转移至具有25mg草甘膦/l的msb5培养基。在没有选择的培养基中培养从经感染的外植体中出现的经确定的芽,以实现其生根。使生根的芽在天然条件下进行适应,以用于其分子和生物学表征。
在用免疫检测试剂盒roundup
实施例3.用质粒pcp4epsps-def进行转化的转基因大豆品系的分子表征
为了进行表征,从转基因植物(t2代)和未转化的对照的幼叶中分离基因组dna。通过pcr来分析所有耐受除草剂草甘膦(并且对于蛋白质cp4epsps的免疫检测试剂盒来说阳性的)的转基因品系,以确证基因nmdef02的存在。在该分析中,在七个源自基因型dt的品系中以及在源自基因型is36的品系中,均获得了140bp的信号,其与用于确证所述基因存在的所预期的大小相符,其中选择品系isdef5。该分析的结果显示在图3中。在用作阴性对照的未转化的植物中,未检测到该防卫素基因的扩增反应。
为了评价在较晚世代(t3)中所述基因的稳定性,用酶ecorv消化该世代的转基因植物和未转化的对照的基因组dna。该酶在质粒pcp4epsps-def中具有两个限制位点,如在图4中所显示的。通过southern印迹来分析经消化的基因组dna,以确定该质粒整合在转基因大豆事件中。
southern印迹的结果显示在图5中,其中显现出了相应于包含在两个由酶ecorv所识别的位点之间的质粒区域的信号。这些信号证明了包含基因cp4epsps和nmdef02的质粒区段整合在植物dtdef1、dt12、dt15、dt16、dt17、dt18和isdef5的基因组中,它们呈现出大小未5.3kb的杂交信号。用作阴性对照的未转化的大豆植物的dna未显示出任何杂交信号。
通过qrt-pcr对t2代的转基因品系(dtdef1、dt3、dt4、dt5、dt6和isdef5)进行防卫素基因nmdef02的相对表达的分析,以选择具有较高表达的那些用于大田测试。所评价的品系显示出不同的防卫素基因的表达水平,如在图6中所显示的。证明了基因nmdef02的转录物积累水平在dtdef1和isdef5系中较高,接着是dt3和dt6,因此选择它们用于针对真菌病原体在大田中进行评价。在该分析中证实,这些品系结果显著地不同于其余的品系和非转基因对照(p<0.0001)。
在相同的品系中(但在t3代中)再次进行通过qrt-pcr的分析。结果显示,所有品系都显示出不同于其各自对照的nmdef02的表达水平,如在图7中所看出的,这与在前一代的评价中所观察到的相一致。品系dtdef1和isdef5再次显示出最高的相对表达水平,其中相关于其余的品系和非转基因对照而言具有非常显著的差异(p<0.0001)。
实施例4.在转基因大豆品系中对于除草剂草甘膦的耐受性的评价
在温室条件下和在大田中评价转基因植物对于草甘膦的应答。在其适应15天时在经转化的植物中进行在温室条件下的除草剂草甘膦的施加。该除草剂以3.5l/ha的最终剂量进行使用,并且通过容量为25l的手动喷雾器来进行施加。在施加后两或三天,所有对该除草剂敏感的植物显示出缺绿症的症状,并且在7天时死亡。然而,耐受性植物保持其绿色着色。经转化的植物对于该除草剂的表现在所述两种大豆基因型中是相似的。所有未转化的对照植物对于该除草剂是敏感的。对于该除草剂具有抗性的转基因植物具有与未经历该除草剂施加的未转化的对照相似的生长和发育。在所评价的三个世代(t1、t2和t3)中对于该除草剂的耐受性百分比的数据显示在表2中。
表2.在温室条件下,在转基因大豆事件中和在未转化的对照dt84和is36中对于除草剂草甘膦的耐受性的评价
ne:未评价
在温室条件下对于该除草剂的耐受性的评价显示出了在t1代中具有80%或更多的对于该除草剂具有耐受性的植物的一组转基因品系。相反地,品系is19和is20是具有更多数目的对于草甘膦敏感的植物的品系,因此在随后的世代中未进行评价。从t2代开始,对于dtdef1和dt5,观察到100%的对于该除草剂的耐受性,其中关于基因cp4epsps达到了纯合。其余所评价的事件在t3代中显示出对于该除草剂的耐受性,并且在品系isdef5中在t3代中达到了100%的对于该除草剂具有抗性的植物。
实施例5.在转基因大豆品系中由镰孢属(fusarium)引起的感染的评价
对于该测试,使用源自基因型dt84的品系的在温室中生长的t0代转基因植物的健康幼叶。将真菌在28℃的温度下在马铃薯-葡萄糖-琼脂培养基中培养7天。将所述叶以背面朝上放置在具有1ml的无菌蒸馏水的培养皿中。然后,在叶的中央放置具有镰孢属物种真菌的马铃薯-葡萄糖-琼脂培养基的圆片(直径5mm)。将所述叶在28℃下进行温育,以在接种后7、10、15和25天时进行观察。布置两个未转化的植物对照:阳性对照(具有真菌圆片的叶)和阴性对照(没有真菌圆片的叶)。计算感染指数,通过借助于由ntui等人(ntui等人,plantcellreport(2010)29:943-54)所提出的4级量表来测量被由于真菌的存在而引起的疾病所侵袭的叶的面积。在接种后7天时,在未转化的对照的叶中在真菌周围出现以缺绿症形式的初步症状,并且观察到真菌的菌丝体在这些叶上生长。在10天时,所述叶在褪绿晕圈的中央呈现出坏死,包括大约50%的叶面积。在15天后,该坏死包括75%的叶。在25天时,100%的叶是坏死的,并且在对照中变成半透明的。相反地,在经遗传转化的植物的叶中,没有以该如此晚期的侵袭水平观察到这些症状。在接种后10天时,事件dt3和dt4显示出在叶中的叶脉缺绿症,和在中央叶脉中的轻微坏死。这些损伤包括在这两个品系中少于12%的总叶面积。相反,在品系dtdef1中,不具有镰孢属物种真菌的侵袭,并且所有叶都保持其强烈绿色着色。在一些在该实验中所评价的品系中所观察到的感染指数显示在表3中。
表3.用镰孢属物种进行接种的转基因大豆植物和未转化的对照的叶的感染指数
考虑到在用t0代进行的生物测定法中所获得的结果,将所评价的三个品系归类为抗性的,这是由于其低的感染指数(0.025-1.20)。相反,将未转化的对照归类为高度易感的,这是由于其高的感染指数(33.1),如在表3中所显示的。
实施例6.在网室条件下转基因品系针对由豆薯层锈菌引起的天然感染的应答
该实验用转基因品系dtdef1、dt3、dt4、dt5、dt6和非转基因对照(dt84)的t1代来进行。播种在网室的地块中的植物不暴露于任何化学产品施加。在网室附近的区域中亚洲锈病的存在使得在患病植物中所产生的夏孢子被空气流所卷带,并且散布至很远的距离。为此,每3天观察植物,以就真菌豆薯层锈菌的发生率来评价它们。在所有播种在网室中的非转基因对照植物中观察到与在文献中对于亚洲锈病所描述的那些相似的症状。对照的一些叶呈现出在植物的下三分之一的叶的背面处的棕红色疣突,达到覆盖整个叶的缺绿症,并且一些植物显示出落叶,剩下附着在茎上的叶柄。在转基因品系的叶中也观察到这些症状中的一些,例如在接近地面的基部区中的叶的背面处的缺绿症和疣突,但是这些不侵袭植物的上三分之一。
用免疫检测试剂盒envirologix
然后,通过qrt-pcr来分析被真菌豆薯层锈菌侵袭或未侵袭的经转化的植物和对照dt84的dna,其中使用特异于亚洲锈病的寡核苷酸(sconyers等人,plantdisease(2006)90:972-972)。以这种方式来定量在受侵袭的植物中存在的该真菌的生物量,并且将所获得的数据显示在图8中。在植物中该真菌的生物量的定量证明,在对照和经转化的品系之间存在显著差异,因为后者显示出非常低水平的该真菌的存在。这确证了在经转化的植物的叶中所观察到的针对引起亚洲锈病的活体营养真菌的抗性,主要在dtdef1和dt5中。
收获所评价的品系,并且对它们进行农艺学评价,其中考虑不同的与产量相关的参数。在农艺学评价中确证,转基因植物在任何其器官中都没有呈现出侵袭,并且其表型特征与对照相似。所评价的参数在五个转基因品系之间是相似的,除了种子的数目和重量外,其在品系dtdef1、dt4和dt5中较大。存在对照dt84的产量的减少,当与转基因品系中的三个进行比较时,如在表4中所看出的。
表4.在网室条件下收获的转基因和非转基因植物的形态-农艺学评价
在该表格的相同列中具有不同字母的平均值表明了显著的统计学差异(关于p<0.05),根据tukey检验。数据相应于每个品系30株植物的平均值。
实施例7.在大田条件下转基因品系针对由豆薯层锈菌引起的天然感染的应答
在十一月至二月期间,当气候条件有利于真菌性疾病的发展时,进行在表达防卫素基因和负责草甘膦耐受性的基因的转基因大豆品系中豆薯层锈菌的发生率的评价。评价了转基因品系dtdef1、dt3和dt6和未转化的对照(dt84),以及品系isdef5及其各自的对照(is36)。当植物处于籽粒充填期时,在非转基因对照中观察到被亚洲锈病侵袭的初步症状。采用用于检测亚洲锈病的免疫检测试剂盒envirologix
在大田评价中观察到,品系dt3和dt6被锈病侵袭,但是侵袭低于在非转基因对照dt84中所观察到的,如在图9中所显示的。另一方面,克隆dtdef1显示出低的真菌发生率,并且仅在一些植物的基部的叶中观察到小的坏死点,不同于对照dt84,其具有100%的亚洲锈病侵袭。在该对照组中,在所有植物中都观察到普遍的缺绿症、叶中的疣突和提早落叶。而与之同时,品系isdef5显示出小于5%的低的在植物中的锈病侵袭,相比于在用作该实验的未转化的对照的基因型is36中大于80%的受侵袭的植物而言。在周期结束时,收获植物并且评价不同的与产量相关的参数。这些在品系dtdef1中是更优异的,具有统计学显著性,相关于未转化的对照和其余的在该实验中所评价的转基因品系而言。
在第二个实验中,在十一月至二月期间(与最有利于该疾病发展的季节相一致),在实验地块中进行针对大豆被亚洲锈病感染的转基因品系(基因型dt84)的评价。锈病的初步症状非常早地出现,在当植物处于开花和籽粒形成期时。在未转化的对照中和在一些转基因品系中观察到这些。该疾病的存在通过免疫检测试剂盒envirologix
在该相同的实验中,还以受侵袭的植物的百分比定量了该疾病的发生率,其中转基因品系具有一定程度的亚洲锈病侵袭。这些植物显示出比未转化的对照低的发生率值,如在图10中所看出的。品系dtdef1、dt12、dt16、dt17和isdef5为具有较低发生率百分比的品系。
使用由ploper等人(ploper等人,avanceagroindustrial-estaciónexperimentalagro-industrialobispocolombres(2006)27:35-37)在2006年提出的量表来分析该疾病的严重度(被锈病侵袭的叶的面积的百分比)。为此,对植物的基部区、中间区和顶端区的小叶进行取样。在图11中所显示的结果表明,以“侵袭的%”表示的严重度值在基部区中比在中间区中高,而中间区又比顶端区高。所有转基因品系都显示出比对照dt84低的疾病严重度,后者在基部区的叶中具有大于40%的锈病侵袭。品系dtdef1、dt17和dt18的植物显示出较少的亚洲锈病侵袭;其中品系dtdef1在基部区中具有少于8%的受侵袭的叶,在中间区和顶端区中具有低于5%的值。
在该相同的测试中,用作阴性对照(基因型dt84)的植物具有100%的该疾病的发生率,这证明了该基因型对于亚洲锈病的高的易感性。所评价的易感品系的转基因植物具有在58-97%之间波动的锈病发生率。然而,抗性转基因品系的植物具有16-35%的发生率。在所述评价中证实,转基因植物的叶保持其绿色着色(尽管该病原体的发病率),不同于非转基因对照,其在其叶中显示出缺绿症,随后为普遍化的落叶。
所评价的其他参数为在检测到该疾病的症状后36和60天,被豆薯层锈菌侵袭的植物的落叶(图12)。结果显示,经转化的植物具有低于未转化的对照dt84的落叶百分比。在该评价中观察到,在36天时,品系dt12和dt16未落叶。在该相同的时间,品系dt1、dt5、dt17和dt18具有低于10%的落叶百分比。此外,在评价这些品系的60天时,落叶值不超过10%。品系dtdef1显示出对于所述病原体的抗性,并且落叶低于5%。
在该相同的实验中,还进行了在受侵袭的植物中存在的真菌豆薯层锈菌的生物量的定量。该分析通过采用特异于该真菌的寡核苷酸的qrt-pcr来进行,这使得能够确证该病原体物种。相比于未转化的对照而言,所有转基因品系都具有更低量的该病原体的生物量。品系dtdef1、dt17和dt18显示出较低的在植物中存在的该真菌的生物量(图13),并且与非转基因对照具有显著差异(p<0.0001)。这些结果证明了在大田条件下在大豆中表达的防卫素nmdef02针对亚洲锈病的抗真菌效应。
在被真菌豆薯层锈菌侵袭的叶的分析中,观察到在非转基因植物的疣突(或夏孢子堆)中的大量的孢子形成,这与在这些对照植物中高的该真菌的拷贝数相一致。在转基因植物中,夏孢子的产生在一些品系(dtdef1、dt12、dt17和dt18)中大大减少,这与较低量的该真菌的生物量相一致。在这些品系的一些植物中,尽管形成了夏孢子堆,但是不存在孢子的产生,这证明存在对于豆薯层锈菌的抗性应答。
在周期结束时,收获植物并且评价不同的与产量相关的参数。在表5中,显示了评价植物收获物的一些结果,在转基因品系和对照之间存在大的差异。在接近籽粒充填的阶段(r3至r5)中高于20%的落叶造成在未转化的对照中产量受到影响。
表5.在实验地块条件下收获的转基因和非转基因植物的形态-农艺学评价
在相同列中具有不同字母的平均值(30株植物/品系)表明了显著的统计学差异(关于p<0.05),根据tukey检验。
大多数的转基因品系与未遗传转化的对照具有在统计学上显著的差异。在作物的早期发育阶段中被该病原体侵袭影响到在对照中所观察到的低的产量。被该病原体的这种侵袭使枝、豆荚和种子的数目以及每株植物的种子重量减少。
基于所获得的应答(关于由豆薯层锈菌所产生的症状的严重度、大豆植物的落叶百分比和在大田中所评价的转基因品系的产量),确定转基因事件cigbdt-def1和cigbis-def5对于其在具有商业价值的大豆作物中的使用来说具有最好的表现。
实施例8.在事件cigbdtdef1的基因组中转基因的插入位点的鉴定
通过经改进的由doyle和doyle(1987)所报道的ctab实验方案来进行转基因品系的总dna的提取,并且着手由macrogen公司(韩国)通过illuminahiseq2500来进行其大规模测序。将序列用fastqc程序(第0.11.4版)进行检查,并且证实它们遵循对于所使用的平台(illuminahiseq2500)所预期的质量模式。为了将测序出的读数映射至大豆基因组和载体以及鉴定转基因的可能插入位点,使用程序itis(第1版)。itis是为了从平行大规模测序(mps;massiveparallelsequencing)数据开始鉴定转座子而设计的程序,其是与在基因组中鉴定载体的那一个相似的应用。itis利用了ilumina平台的成对末端技术的特征。在这之中,文库的每个片段是2次读数的目标。一个从5'至3'末端,而另一个以相反方向,从3'至5'末端。这两次读数被认为是一对。itis利用bwa程序(第0.7.7版)来比对关于基因组和关于载体的读数。映射至载体的序列的分析表明,它们中的大部分具有很少的覆盖,除了位于2号染色体中的那些。由于这个原因,决定继续在映射在所述染色体中和在载体(其描绘在图1中)中的读数之中的分析。作为结果,在大豆(品系dtdef1)的基因组的2号染色体中,在位置8002360和8334726处鉴定出了两个潜在的转基因插入位点。这些位置是指在大豆品种willian82的基因组中的核苷酸的编号。
用在与在生物信息学分析中鉴定出的位点相邻的大豆基因组区域中所设计的引物,通过pcr来验证在位点8002360和8334726处的转基因插入。对于位点8002360,引物的序列为:3'-aagcggcaagtcaatcgtgtcg-5'(seqidno.1)和5'-ctgaatccctacattgcgattctcg-3'(seqidno.2),其组合产生具有没有遗传修饰的大豆dna的大小为344bp的条带,如在图14中所显示的。对于位点8334726,引物的序列为:3'-atgtgcaataattccttcttcg-5'(seqidno.3)和5'-cgaaacacgaatcacgaagc-3'(seqidno.4),该组合产生具有没有遗传修饰的大豆dna的大小为273bp的条带(图14)。在转基因品系中,当用这两个寡核苷酸对进行该程序时,均没有扩增出任何条带。所获得的结果确证了在位点8002360和8334726(两者都在大豆的2号染色体中)处的转基因插入。
然后,将从大豆基因组的序列开始设计的引物与在基因cp4epsps和nmdef02(两者都在载体pcp4epsps-def(其描绘在图1中)中)中的引物相组合。在大豆中的引物(正向和反向)与在基因cp4epsps中的引物(5'-ggatttcagcatcagtggctacagc-3'(seqidno.5))和(3'-gcgggttgatgacttcgatgtcg-5'(seqidno.6))的组合确证了在位点8002360(图15a)和8334726(图15b)处转基因的存在,对于转基因事件cigbdtdef1。在图15a中,观察到在位点8002360处的1000bp的序列的扩增;和在图16b中,观察到在位点8334726处的900bp的序列的扩增,对于转基因事件cigbdtdef1。这些所扩增出的区域相应于包含植物的核苷酸以及所引入的转基因的核苷酸序列。
当使用从大豆基因组的序列开始设计的引物与从基因nmdef02设计的引物(3'-gctggcttatgcttcctcttcttg-5'(seqidno.7)和5'-tcacagacttggacgcagttcg-3'(seqidno.8))的组合时,发生了相同的事情,对于位点8002360(图16a)和8334726(图16b)。在图16a中,观察到在位点8002360处的2000bp的序列的扩增;和在图16b中,观察到在位点8334726处的300bp的序列的扩增,对于转基因事件cigbdtdef1。这些所扩增出的区域相应于包含植物的核苷酸以及所引入的转基因的核苷酸序列。以这种方式,证明了在事件dtdef1中转基因的插入。
序列表
<110>centrodeingenieriageneticaybiotecnologia
<120>包含转基因事件cigbdt-def1或cigbis-def5的大豆植物
<130>eventossoyatransgenica
<140>
<141>
<160>8
<170>patentinver.2.1
<210>1
<211>22
<212>adn
<213>大豆
<400>1
aagcggcaagtcaatcgtgtcg22
<210>2
<211>25
<212>adn
<213>大豆
<400>2
ctgaatccctacattgcgattctcg25
<210>3
<211>22
<212>adn
<213>大豆
<400>3
atgtgcaataattccttcttcg22
<210>4
<211>20
<212>adn
<213>大豆
<400>4
cgaaacacgaatcacgaagc20
<210>5
<211>25
<212>adn
<213>根癌土壤杆菌
<400>5
ggatttcagcatcagtggctacagc25
<210>6
<211>23
<212>adn
<213>根癌土壤杆菌
<400>6
gcgggttgatgacttcgatgtcg23
<210>7
<211>24
<212>adn
<213>特大管烟草
<400>7
gctggcttatgcttcctcttcttg24
<210>8
<211>22
<212>adn
<213>特大管烟草
<400>8
tcacagacttggacgcagttcg22
pct/ro/134表
- 还没有人留言评论。精彩留言会获得点赞!