用于癌症的过继性细胞疗法的组合物和方法与流程
相关申请的交叉引用
本申请要求保护2017年6月30日提交的美国临时专利申请号62/527,932的权益和优先权,其全部内容通过提述并入本文。
序列表
本申请含有序列表,该序列表已经以ascii格式以电子方式提交,且通过提述以其整体并入此文。所述ascii副本,创建于2018年7月1日,命名为115872-0384_sl.txt,大小为119,572字节。
政府资助声明
本发明是在美国国立卫生研究院授予的ai073736、ai095692、ar068118、r01ca55349、p01ca23766、gm100477和ai118224的政府资助下完成的。政府拥有本发明中的某些权利。
发明背景
过继性和工程化t细胞疗法(包括嵌合抗原受体(car)t细胞、t细胞受体(tcr)工程化t细胞和抗原过继的t细胞)最近已作为用于癌症的重要疗法出现。通过将scfv与cd3-ζ链的细胞内信号传导域融合设计了第一代car,而第二代car添加了cd28-cd80共刺激,其中cd28信号传导域被并入car构建体中(即:“28ζcar”)用于改善t细胞激活和功效。然而,对于许多种癌症类型,仍需要更大的效能和机制来击败免疫抑制性肿瘤微环境。对这些细胞活性的抗性、复发和毒性仍然是其成功的重要障碍。最近关于cart细胞疗法和tcrt细胞疗法后的显著毒性甚至死亡以及gvhd的持续发病率及其相关的短期和长期发病率和死亡率的报道,要求新的方法来加强疗效的同时更好地控制多种过继性细胞疗法。
发明概述
本文提供了用于过继性细胞疗法的组合物和方法,该过继性细胞疗法包含表达结合靶抗原的受体和第一前药转化酶,其中该第一前药转化酶在工程化免疫细胞的细胞质中(例如,细胞内地)表达,以允许细胞的选择性破坏。在一些实施方案中,工程化免疫细胞进一步包含第二前药转化酶,该第二前药转化酶在工程化免疫细胞的表面上表达或由工程化免疫细胞分泌。在一些实施方案中,受体是t细胞受体。在一些实施方案中,受体是天然天然t细胞受体。在一些实施方案中,受体是非天然t细胞受体,例如,工程化受体,诸如嵌合抗原受体(car)。在一些实施方案中,工程化免疫细胞包含嵌合抗原受体和/或编码嵌合抗原受体的核酸。
在一些实施方案中,工程化免疫细胞包含第一前药转化酶和/或编码该第一前药转化酶的核酸,其中该第一前药转化酶在该免疫细胞的细胞质中表达。在一些实施方案中,工程化免疫细胞进一步包含第二前药转化酶和/或编码该第二前药转化酶的核酸,其中该第二前药转化酶在该免疫细胞的表面上表达,或是分泌的。
在一些实施方案中,该工程化免疫细胞包含:(a)第一前药转化酶和/或编码该第一前药转化酶的核酸,其中该第一前药转化酶在该免疫细胞的细胞质中表达,且选自羧肽酶、β-内酰胺酶、葡糖苷酶、硝基还原酶和羧肽酶a;和(b)结合靶抗原的受体和/或编码该受体的核酸,诸如天然受体或嵌合抗原受体和/或编码该受体的核酸。在一些实施方案中,该工程化免疫细胞进一步包含第二前药转化酶和/或编码该第二前药转化酶的核酸,其中该第二前药转化酶在该免疫细胞的表面上表达或是分泌的。
在一些实施方案中,该工程化免疫细胞包含:(a)第一前药转化酶和/或编码该第一前药转化酶的核酸,其中该第一前药转化酶在该免疫细胞的细胞质中表达;(b)第二前药转化酶和/或编码该第二前药转化酶的核酸,其中该第二前药转化酶在该免疫细胞的表面上表达或是分泌的;和(c)结合靶抗原的受体和/或编码该受体的核酸。在一些实施方案中,该第一前药转化酶和/或第二前药转化酶选自羧肽酶、β-内酰胺酶、葡糖苷酶、硝基还原酶和羧肽酶a。
在一些实施方案中,第二前药转化酶与跨膜域融合。在一些实施方案中,前药转化酶的跨膜域包含cd8跨膜域。在一些实施方案中,第二前药转化酶通过gpi锚被附接至细胞的表面。在一些实施方案中,编码第二前药转化酶的核酸包含用于分泌前药转化酶的前导序列。
在一些实施方案中,第一前药转化酶和第二前药转化酶具有相同的酶活性和/或是相同的酶。在一些实施方案中,第一前药转化酶和第二前药转化酶具有不同的酶活性和/或是不同的酶。在一些实施方案中,第一前药转化酶是羧肽酶(例如,假单胞菌属种(pseudomonassp.)羧肽酶g2(cpg2))。在一些实施方案中,第一前药转化酶是β-内酰胺酶(例如,阴沟肠杆菌(enterobactercloacae)β-内酰胺酶)。在一些实施方案中,第二前药转化酶是羧肽酶(例如,假单胞菌属种羧肽酶g2(cpg2))。在一些实施方案中,第二前药转化酶是β-内酰胺酶(例如,阴沟肠杆菌β-内酰胺酶)。在一些实施方案中,编码第一前药转化酶和/或第二前药转化酶的核酸与启动子可操作地连接。在一些实施方案中,启动子是组成型启动子。在一些实施方案中,启动子是条件性启动子。在一些实施方案中,条件性启动子是可通过受体(例如,car)与抗原(诸如肿瘤抗原)的结合而诱导的。
在一些实施方案中,嵌合抗原受体包含(i)细胞外抗原结合域;(ii)跨膜域;和(iii)细胞内域。在一些实施方案中,细胞外抗原结合域结合肿瘤抗原。在一些实施方案中,肿瘤抗原选自cd19、wt1和prame。在一些实施方案中,细胞外抗原结合域包含单链可变片段(scfv)。在一些实施方案中,细胞外抗原结合域包含人scfv。在一些实施方案中,细胞外抗原结合域包含seqidno:19的cd19scfv。在一些实施方案中,细胞外抗原结合域包含与seqidno:19具有至少80%、85%、90%、95%、96%、97%、98%或99%序列同一性的cd19scfv。在一些实施方案中,细胞外抗原结合域包含与细胞外抗原结合域的n端共价连接的信号肽。在一些实施方案中,跨膜域包含cd8跨膜域。在一些实施方案中,细胞内域包含共刺激域。在一些实施方案中,一种或多种共刺激域选自cd28共刺激域、cd3ζ-链、4-1bbl共刺激域,或其任何组合。在一些实施方案中,免疫细胞是淋巴细胞。在一些实施方案中,淋巴细胞是t细胞、b细胞或自然杀伤(nk)细胞。在一些实施方案中,t细胞是cd4+t细胞或cd8+t细胞。在一些实施方案中,免疫细胞是肿瘤浸润淋巴细胞。在一些实施方案中,免疫细胞衍生自自体供体或同种异体供体。
还提供了编码本文公开的任何多肽的核酸。在一些实施方案中,编码多肽的核酸与启动子可操作地连接。在一些实施方案中,启动子是组成型启动子。在一些实施方案中,启动子是条件性启动子。在一些实施方案中,条件性启动子是可通过car结合抗原而诱导的。
还提供了包含本文公开的任何核酸的载体。在一些实施方案中,载体是病毒载体或质粒。在一些实施方案中,载体是逆转录病毒载体。
还提供了包含本文公开的多肽、核酸或载体的宿主细胞。
还提供了用于治疗有需要的受试者中的癌症的方法,该方法包括施用有效量的本文提供的任何工程化免疫细胞。本文还提供了用于治疗或抑制受试者中的肿瘤生长或转移的方法,该方法包括使肿瘤细胞与有效量的本文提供的任何工程化免疫细胞接触。在一些实施方案中,在施用所述工程化免疫细胞之后施用所述前药。在一些实施方案中,所述方法进一步包括向受试者施用前药,该前药被前药转化酶转化为活性药物。在一些实施方案中,所述方法进一步包括向所述受试者施用第一前药,该第一前药被所述第一前药转化酶转化为活性药物。在一些实施方案中,在施用工程化免疫细胞随后施用所述第一前药。在一些实施方案中,所述方法进一步包括向所述受试者施用第二前药,该第二前药被所述第二前药转化酶转化为活性药物。在一些实施方案中,在施用所述第一前药之前施用所述第二前药转化酶。在一些实施方案中,工程化免疫细胞是静脉内、腹腔内、皮下、肌内或瘤内施用的。在一些实施方案中,癌症或肿瘤是癌、肉瘤、黑色素瘤或造血癌症。在一些实施方案中,癌症或肿瘤选自肾上腺癌、膀胱癌、血液癌症、骨癌、脑癌、乳腺癌、癌、宫颈癌、结肠癌、结直肠癌、子宫体癌、耳鼻喉(ent)癌、子宫内膜癌、食管癌、胃肠癌、头和颈、霍奇金氏病、肠癌、肾癌、喉癌、白血病、肝癌、淋巴结癌、淋巴瘤、肺癌、黑色素瘤、间皮瘤、骨髓瘤、鼻咽癌、成神经细胞瘤、非霍奇金氏淋巴瘤、口腔癌、卵巢癌、胰腺癌、阴茎癌、喉咽癌、前列腺癌、直肠癌、肉瘤、精原细胞瘤、皮肤癌、胃癌、畸胎瘤、睾丸癌、甲状腺癌、子宫癌、阴道癌、血管瘤,及其转移。在一些实施方案中,所述方法进一步包括施用另外的癌症疗法。在一些实施方案中,另外的癌症疗法选自化学疗法、放射疗法、免疫疗法、单克隆抗体、抗癌核酸或蛋白质、抗癌病毒或微生物,及其任何组合。在一些实施方案中,所述方法进一步包括向受试者施用细胞因子。在一些实施方案中,在施用一种或多种工程化免疫细胞之前、期间或随后施用细胞因子。在一些实施方案中,细胞因子选自干扰素α、干扰素β、干扰素γ、补体c5a、il-2、tnfα、cd40l、il12、il-23、il15、il17、ccl1、ccl11、ccl12、ccl13、ccl14-1、ccl14-2、ccl14-3、ccl15-1、ccl15-2、ccl16、ccl17、ccl18、ccl19、ccl19、ccl2、ccl20、ccl21、ccl22、ccl23-1、ccl23-2、ccl24、ccl25-1、ccl25-2、ccl26、ccl27、ccl28、ccl3、ccl3l1、ccl4、ccl4l1、ccl5、ccl6、ccl7、ccl8、ccl9、ccr10、ccr2、ccr5、ccr6、ccr7、ccr8、ccrl1、ccrl2、cx3cl1、cx3cr、cxcl1、cxcl10、cxcl11、cxcl12、cxcl13、cxcl14、cxcl15、cxcl16、cxcl2、cxcl3、cxcl4、cxcl5、cxcl6、cxcl7、cxcl8、cxcl9、cxcl9、cxcr1、cxcr2、cxcr4、cxcr5、cxcr6、cxcr7和xcl2。
还提供了用于制备用于癌症疗法的免疫细胞的方法,该方法包括从供体受试者中分离免疫细胞,用下列转导免疫细胞(例如,t细胞):编码第一前药转化酶的核酸,其中所述第一前药转化酶在所述免疫细胞的细胞质中表达;和任选地编码(i)第二前药转化酶的核酸,其中所述第二前药转化酶在所述免疫细胞的表面上表达或是分泌的,和/或(ii)编码结合靶抗原的受体的核酸和/或编码所述受体的核酸。还提供了用于治疗癌症的方法,该方法包括向接受受试者施用经转导的工程化免疫细胞。在一些实施方案中,所述供体受试者和接受受试者是相同的(即,自体的)。在一些实施方案中,所述供体受试者和接受受试者是不同的(即,同种异体的)。在一些实施方案中,淋巴细胞包含t细胞、b细胞和/或自然杀伤(nk)细胞。在一些实施方案中,t细胞是cd4+t细胞或cd8+t细胞。在一些实施方案中,从供体受试者中分离的免疫细胞包含肿瘤浸润淋巴细胞(til)。
还提供了用于治疗的方法,该方法包括从供体受试者中分离t细胞,用本文提供的核酸和本文提供的载体转导免疫细胞(例如,t细胞),以及向接受受试者施用经转导的免疫细胞(例如,t细胞)。在一些实施方案中,供体受试者和接受受试者是相同的(即,自体的)。在一些实施方案中,供体受试者和接受受试者是不同的(即,同种异体的)。在一些实施方案中,从供体受试者分离的免疫细胞包含一种或多种淋巴细胞。在一些实施方案中,淋巴细胞包含t细胞、b细胞和/或自然杀伤(nk)细胞。在一些实施方案中,t细胞是cd4+t细胞或cd8+t细胞。在一些实施方案中,从供体受试者中分离的免疫细胞包含肿瘤浸润淋巴细胞(til)。
还提供了本文提供的任何工程化免疫细胞用于治疗癌症的用途。
还提供了本文提供的任何工程化免疫细胞在制备用于治疗癌症的药物中的用途。
还提供了式i的化合物:
或其两性离子、药学上可接受的盐和/或其溶剂化物,其中
x1是o或nh;
x2是n或ch;
x3是oh或h;
w1是h或nh-r3;
r1、r2和r3中的两个是h,且其余的r1、r2和r3是
x6是o、nh、ch2或c(me)2;
r4、r5、r6、r7和r8各自独立地为h、-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or13a、-(ch2)2-c(o)or13b、
r9、r10、r11、r12、r13、r13a和r13b各自独立地为烷基、芳基、杂芳基、芳烷基、杂芳烷基。在一些实施方案中,r1、r2和r3中的两个是h,且其余的r1、r2和r3是
r4、r5、r6、r7和r8各自独立地为-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or13a、-(ch2)2-c(o)or13b、
还提供了式ii的化合物:
或其两性离子、药学上可接受的盐和/或其溶剂化物,其中
x7和x8各自独立地为cl、br、i、o-s(o)2ch3或o-s(o)2-甲苯基;
x9是–c(o)-o、-c(o)-nh、o或nh;
r9是键、亚烷基、亚芳基、亚芳烷基、杂亚芳基或杂亚芳烷基;
r15是
x10和x11各自独立地为o或nh;
x6是o、nh、ch2或c(me)2;
x12是o、nh、ch2或c(me)2;
r16、r17、r18、r19和r20各自独立地为h、-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or25a、-(ch2)2-c(o)or25b、
r21、r22、r23、r24、r25、r25a和r25b各自独立地为烷基、芳基、杂芳基、芳烷基或杂芳烷基。在一些实施方案中,r15是
r16、r17、r18、r19和r20各自独立地为-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or25a、-(ch2)2-c(o)or25b、
x13和x18各自独立地为o或nh;x14、x15、x16、x17、x19、x20、x21、x22、x23和x24各自独立地为cl、br、i、o-s(o)2ch3或o-s(o)2-甲苯基;n和m各自独立地为0、1、2、3、4或5;r26、r27、r28、r29和r30各自独立地为
x25和x26各自独立地为o或nh;x27是o、nh、ch2或c(me)2;r31、r32、r33、r34和r35各自独立地为h、-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or40a、-(ch2)2-c(o)or40b、
r45、r46、r47、r48和r49各自独立地为-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or54a、-(ch2)2-c(o)or54b、
还提供了式iii的化合物或其两性离子、药学上可接受的盐和/或溶剂化物:
x35和x36各自独立地为o或nh;
x37是o、nh、ch2或c(me)2;
r56、r57、r58、r59和r60各自独立地为h、-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or65a、-(ch2)2-c(o)or65b、
r61、r62、r63、r64、r65、r65a和r65b各自独立地为烷基、芳基、杂芳基、芳烷基或杂芳烷基。在一些实施方案中,r55是
r56、r57、r58、r59和r60各自独立地为-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or65a、-(ch2)2-c(o)or65b、
附图简述
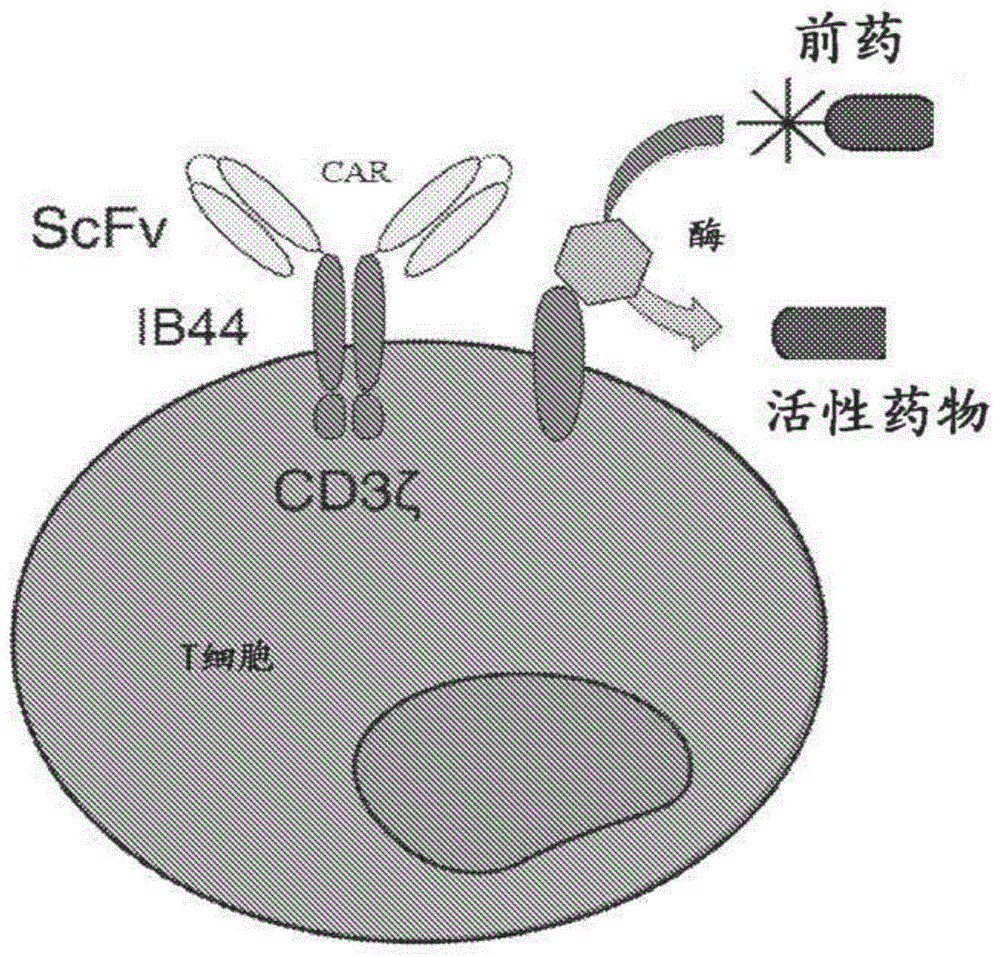
图1提供了合成性酶激活的杀伤(syntheticenzymeactivatedkiller,seaker)细胞技术的示例性概观。
图2示出了用于产生cpg2不稳定的谷氨酸盐前药的示例性方案。(a)非活性前药经历由cpg2切割谷氨酸盐(蓝色),然后自发分解接头(灰色),以揭露活性药物(红色)。(b)基于电子和立体考虑,可用于调节前药稳定性和分裂速率的多种自降解(self-immolative)接头。(c)分别地,细胞毒性氨磺酰腺苷(“ams”;1c)和氮芥zd2767(2c)的前药变体1a(“p-ams”)和2a,以及被设计用于细胞通透性的ams(1d)的前药变体。(d)合成的前体。(e)厄洛替尼的前药变体(6a),其具有烷基pab接头以避免常见的基于酰胺(参见n,n-二芳基酰胺)的接头的自发水解。(co=羰基接头;pab=对氨基苄基接头;tml=三甲基锁(lock)接头;ind=形成茚满酮的接头)。
图3提供示例性seaker构建体的示意图。在seaker细胞技术的示例性实施方案中,谷氨酸化的前药是全身性施用的,但仅在car-t细胞积聚的部位被激活为其细胞毒性形式。(a)cpg2跨膜表达通过cd8跨膜区系链在car-t细胞表面的cpg2酶。(b)cpg2分泌的表达直接被分泌到周围环境中的cpg2酶。这些示例性car-t细胞表达与4-1bb共刺激分子和cd3ζ链偶联的抗原特异性的scfv。自切割p2a肽序列允许多聚蛋白的共翻译切割。(c)示例性cpg2可诱导的car-t细胞仅在与肿瘤抗原啮合后利用syn-notch系统来激活分泌形式的cpg2的表达。
图4提供了显示由大肠杆菌表达并纯化的重组cpg2对p-ams(参见图2)的激活的示例性数据。(a)在浓度递增的纯化的cpg2存在下,将人jurkatt细胞与或不与p-ams一起孵育。在48小时处通过celltitre-glo发光测量细胞活力。(b)重组cpg2被暴露于多种浓度的谷氨酸化的ams前药。在37℃下孵育(0.5、1、2和4小时)后,将反应与
图5提供了显示膜锚定的cpg2的表面表达的示例性数据。用表达血凝素标签的膜系链的cpg2(cpg2tm)转染的hek293t细胞被暴露于使用抗ha抗体的表面染色,并与同种型对照抗体染色的细胞比较。
图6提供了显示哺乳动物细胞中cpg2的表达的示例性数据。通过总细胞裂解物(泳道1-5、11-15)的免疫印迹测定用编码分泌形式的cpg2(cpg2)和具有或无血凝素(ha)标签的膜结合的cpg2(cpg2-tm)的构建体转染的人胚肾(293t)细胞的cpg2表达,或通过细胞上清液(泳道1-10,16-20)的免疫印迹测定其cpg2(顶行)或ha(底行)。
图7提供了显示细胞分泌的cpg2激活ams前药的示例性数据。jurkatt细胞被暴露于浓度递增的ams(红色圆形)或仅具有培养基的p-ams(蓝色三角形)或来自分泌cpg2的293t细胞的上清液(黑色正方形)。在48小时通过celltitre-glo发光测量细胞活力。
图8提供了显示细胞分泌的cpg2激活氮芥前药的示例性数据。jurkatt细胞被暴露于浓度递增的具有培养基的zd2767p(蓝三角形)或来自分泌cpg2的293t细胞的上清液(黑色正方形)。在48小时通过celltitre-glo发光测量细胞活力。
图9提供了显示在稳定转染的细胞中表达的细胞分泌的cpg2和跨膜cpg2激活ams前药的示例性数据。采用无信号肽的cpg2(cpg2nosigp)的表达作为对照。将稳定转染有cpg2变体的293t细胞暴露于p-ams(1μm)48小时,并通过明视野显微镜分析(图9a)。细胞被暴露于不同浓度的p-ams,且在处理后48小时处分析细胞活力(图9b)。
图10提供示了显示在逆转录病毒转导编码与跨膜或分泌形式的cpg组合的抗cd19car的构建体后在人t细胞系、jurkat和原代人t细胞中抗cd19car的表达的示例性数据。用编码具有4-1bb共刺激元件(bbz)的抗cd19car和如上所述的无信号肽的cpg基因(cpgnosigp)或cpg跨膜基因的逆转录病毒转导人t细胞系、jurkat(顶行)和原代人t细胞(底行)。通过对细胞表面上的抗cd19scfv表达进行染色来评定转导水平,其证明了car蛋白在工程化免疫细胞的细胞表面上表达。
图11提供了显示表达具有4-1bb共刺激元件(bbz)的抗cd19car和如上所述的无信号肽的cpg基因(cpgnosigp)或cpg跨膜基因的人jurkatt细胞的活性的示例性数据。用19bbz/cpg逆转录病毒转导的人jurkatt细胞与gfp+raji(人伯基特氏淋巴瘤)癌靶细胞系共培养24小时(1:1比率),并进行流式细胞术分析。jurkat和raji细胞通过gfp表达区分(左图)。通过评定gfp阴性细胞中cd69的表达来检查jurkat细胞的激活(右图)。
图12提供示了显示非转导的原代人t细胞和表达具有4-1bb共刺激元件(bbz)的抗cd19car和如上所述的无信号肽的cpg基因(cpgnosigp)或cpg跨膜基因的转导的人原代人t细胞的活性的例性数据。原代非转导的人t细胞和转导的19bbz和19bbz/cpgcar-t细胞与gfp+raji细胞以1:1比率共培养。在48小时孵育后,通过流式细胞术分析细胞(a)且计算%gfp+细胞(b)。gfp阴性细胞(表示制备的各种cart细胞)增加至与传统cart和2种不同的seaker细胞相同的水平,表明seaker细胞的活性与传统car相当。
图13提供了显示抗cd19car和分泌的cpg2的表达的示例性数据。用编码具有4-1bb共刺激元件(bbz)的抗cd19car和如上所述的无信号肽的cpg基因(cpgnosigp)或具有信号肽的分泌形式的cpg基因的逆转录病毒转导原代人t细胞。通过免疫印迹分析来自编码19bbz或19bbz/cpg构建体的人car-t细胞的上清液的cpg2表达。
图14提供了显示人t细胞而非癌症细胞对厄洛替尼有抵抗的示例性数据。(a)用浓度递增的受体酪氨酸激酶抑制剂厄洛替尼处理人jurkatt细胞(红圆形)和人非小细胞肺癌细胞系pc9(黑色正方形)和hcc827(蓝色三角形)。(b)谷氨酸化的厄洛替尼前药(p-厄洛替尼)除非被cpg2激活,否则在pc9细胞中未表现出毒性(1μm)。在48小时处通过celltitre-
图15提供了显示表达跨膜cpg2的293t细胞的前药活性的示例性数据。gfp+293tcpgtm细胞与肺腺癌细胞(hcc827)以多种比率(293tcpgtm:hcc827)共培养,且添加厄洛替尼前药(p-厄洛替尼)。(a)在120小时处通过流式细胞术分析细胞并计算%gfp。(b)针对每种细胞比率计算293tcpgtm细胞与hcc827细胞的比率。这表明转导的细胞中的跨膜cpg2是有功能的,并将前药转化为杀伤靶癌细胞但不杀伤分泌cpg2酶的细胞的活性药物。
图16提供了示例性数据,其显示了在移植有wt1+、set2aml细胞的scid/beige小鼠的第7天开始,用2e7wt1特异性tcrm标准cart细胞(黑线)或装有il-12分泌的2e7wt1特异性tcrm标准cart细胞(黑点)处理荷瘤小鼠相对于对照改善了存活(p<0.03)。4h11是在有和没有il-12情况下的对照cart细胞。
图17提供cpg2“杀伤开关”构建体的示例性示意图。(a)在示例性实施方案中,cpg2杀伤开关car-t表达一种cpg2形式,该形式被隔离在细胞溶质内,并将转化细胞穿透性前药。(b)car-t细胞还可同时表达跨膜(或分泌的)cpg2酶用于在靶位点转化前药。
图18示出了在哺乳动物细胞中细胞溶质-隔离的cpg2的表达。通过总细胞裂解物(泳道1-3)或细胞上清液(泳道4)的免疫印迹测定用编码分泌形式的cpg2(cpg2)和细胞溶质-隔离形式的cpg2(cpg2-nosigp)的构建体转染的293t细胞。
图19提供了设计为穿透到speaker细胞中的类似物前药的示例性结构。
图20a提供了在暴露于cpg2后谷氨酸盐前药切割和接头衰变导致活性药物合成的示意图。
图20b提供用于与cpg2酶平台一起使用的合成的3类前药化合物的示意图。
图20c显示了ampexred测定法,其证明了与重组cpg2酶孵育后从前药化合物释放出游离谷氨酸盐。
图20d显示了在有或无重组cpg2的3μmams-glut中孵育的set2细胞的celltiter-glo活力测定。
图20e显示了在有或无重组cpg2的1μm厄洛替尼-glut中孵育的set2细胞的celltiter-glo活力测定。
图20f显示了在有或无重组cpg2的zd2767中孵育的jurkat细胞的细胞毒性测定。
图21a显示了纯化的重组cpg2的考马斯蓝染色。
图21b显示了纯化的重组cpg2的定量夹心elisa。
图21c显示了与ams-glut前药或ams一起孵育的lncap细胞的celltiter-glo活力测定。
图21d提供了用于与β-内酰胺酶平台一起使用的合成的前药化合物类别的示意图。
图21e显示了纯化的重组阴沟肠杆菌β-内酰胺酶的考马斯蓝染色。
图21f显示了纯化的重组β-内酰胺酶的定量elisa。
图22a提供了为了在哺乳动物细胞中最佳表达而生成的三种形式的cpg2(内部的、分泌的、膜结合的)示意图。
图22b提供了指示在hek293t细胞中转染后,所有三种cpg2同种型在细胞裂解物(左图)中或分泌到上清液(右图)中的表达的免疫印迹。
图22c显示了在来自对照hek293t细胞的上清液或来自表达分泌形式的cpg2的hek293t细胞的上清液中,与ams-glut前药一起孵育的set2细胞的celltiter-glo活力测定。
图22d证明了经转导以稳定表达cpg2基因的hek293t细胞中cpg2蛋白的表面表达分析。
图22e显示了与来自经工程化以稳定表达cpg2的hek293t细胞的ams-glut上清液一起孵育的set2细胞的celltiter-glo活力测定。
图22f显示了暴露于ams-glut前药的hek293t-cpg2细胞的celltiter-glo活力测定。
图22g提供了暴露于ams-glut前药的hek293t-cpg2细胞的显微照片。
图22h显示了来自jurkat-cpg2细胞的上清液的免疫印迹分析。
图23a显示了来自用多种形式的cpg2转导的hek293t细胞的裂解物和上清液的抗cpg2免疫印迹。
图23b显示了在来自对照hek293t细胞的上清液或来自表达cpg2-内部的、cpg2wt-分泌的或cpg2cd8-分泌的的hek293t细胞的上清液中与ams-glut前药一起孵育的set2细胞的celltiter-glo活力测定。
图23c显示了plgpw-cpg2基因盒的示意图。
图24a提供了在ams-glut前药的存在下,比率递增的hek293t-gfp+/cpg2或hek293t-gfp+细胞(效应细胞)与hek293twt(旁邻细胞)的共培养物的细胞活力分析。
图24b显示了在有或无前药情况下的共培养的gfp分析证明了酶+和酶–细胞群二者的等效耗竭。
图24c显示了在有或无前药情况下的1:1e:t共培养物中总细胞耗竭的定量。
图24d显示了在有或无前药情况下的1:1e:t共培养物中酶-阴性(旁邻细胞)耗竭的定量。
图25a提供了经工程化以在cd19-bbz基因盒上游表达cpg2或β-内酰胺酶的sfg逆转录病毒载体的示意图。
图25b显示了在car基因盒之前或之后用编码cpg2的载体转导的原代人t细胞中cpg2分泌的elisa分析。
图25c显示了与cd19+(raji)或cd19-(set2)细胞共培养的19bbz和cpg-19bbzjurkat细胞的t细胞激活分析。
图25d显示了cpg2jurkats(左图–elisa),或β-内酰胺酶jurkat(右图–头孢硝噻吩)的酶分泌分析。
图25e显示了用19bbz、cpg-19bbz和β-lac-bbzcar转导原代人t细胞。
图25f显示了在与非转导的、19bbz、cpg-bbz或β-lac-bbzcar-t细胞共培养之后,luc+/cd19+raji细胞的特异性裂解。
图25g显示了与前药和来自表达seaker构建体的人t细胞的上清液孵育的set细胞的celltiter-glo活力测定。
图25h显示了与19bbzt细胞相比,cpg2-19bbz(左图)和β-lac-19bbz(右图)t细胞的上清液中酶积累的分析。
图25i显示了与cd19+raji细胞和cd19-set2细胞共培养的cpg2-19bbz(左图)和β-lac-19bbz(右图)t细胞的上清液中酶积累的分析。
图26a提供了sfg逆转录病毒载体中19bbz-cpg2基因盒的示意图。
图26b显示了用19bbz或19bbz-cpg2基因转导原代人t细胞。
图26c显示了在与19bbz或19bbz-cpg2car-t细胞共培养后luc+/cd19+nalm6细胞的特异性裂解。
图26d显示了来自19bbz-cpg2car-t细胞的上清液的抗ha免疫沉淀的抗cpg2免疫印迹。
图26e显示了与前药和来自表达19bbz或19bbz-cpg2构建体的人t细胞的上清液孵育的set细胞的celltiter-glo活力测定。
图27a显示了为了在哺乳动物细胞中最佳表达而生成的两种形式的β-内酰胺酶的示意图。
图27b显示了指示仅截短形式的β-内酰胺酶用β-内酰胺酶表达构建体转染的hek293t细胞的上清液中表达的免疫印迹。
图27c提供来自表达β-内酰胺酶构建体的细胞的上清液中β-内酰胺酶活性的nitrocefin分析。
图28a显示了纯化的重组cpg2的定量夹心elisa(左图)和纯化的β-内酰胺酶的定量头孢硝噻吩测定(右图)。
图28b显示了与前药和不同浓度的纯化的重组cpg2(左图)或β-内酰胺酶(右图)一起孵育的set细胞的celltiter-glo活力测定。
图28c显示了用前药和来自用cpg2(左图)或β-内酰胺酶(右图)表达构建体转染的hek293t细胞的上清液一起孵育的set细胞的celltiter-glo活力测定。
图28d显示了暴露于ams-glut的hek293t-cpg2细胞(左图)或暴露于ceph-ams的hek293t-beta-lac细胞(右图)的celltiter-glo活力测定。
图28e显示了在有或无前药的情况下的hek293t-cpg2或hek293t-beta-lac共培养物的gfp分析,证明了酶+和酶-两种细胞群体的等效耗竭。
图29a显示了在零时间具有car-t细胞的中靶(raji:gfp-/cd19+)和脱靶(set2:gfp+/cd19-)共培养的代表。
图29b显示了在共培养72小时后,经ams-glut处理(底行)或未经ams-glut处理(顶行)的具有set2和raji细胞的19bbz(左列)或cpg-19bbz(右列)t细胞的流式图。
图29c显示了在各自的共培养后,脱靶(set2)细胞的定量。
图30a显示了在有或无ceph-ams前药情况下的raji/set2/beta-lac-19bbzt-细胞共培养(72hr)后细胞数目的定量。各个细胞群体用条形颜色表示。
图30b显示了在具有或不具有ceph-ams前药的情况下与β-lac19bbz共培养之后,luc+/cd19+raji细胞的特异性裂解。
图31显示了在nod小鼠中进行的与glut-ams前药有关的药代动力学和生物分布研究。
发明详述
依照本申请中描述的特定的实施方案并不限制本公开,所述特定的实施方案旨在作为本公开的各个方面的单个说明。在本文中将不描述本公开的所有各种实施方案。如对本领域技术人员将是显而易见的,在不脱离本发明的精神和范围的情况下,可以对本发明进行许多修改和变化。除了本文列举的方法和装置之外,根据前述描述,在本发明范围内的功能上等同的方法和装置对于本领域技术人员也将是显而易见的。此类修改和变化旨在落入所附权利要求的范围内。本公开仅由所附权利要求的术语连同此类权利要求所赋予的等同物的全部范围来限制。
应当理解,本公开内容不受限于特定用途、方法、试剂、化合物、组合物或生物系统,当然,它们可以变化。还应理解,本文所使用的术语仅出于描述特定实施方案的目的,而并不旨在限制。
此外,在依照马库什组描述本公开的特征或方面的情况下,本领域技术人员将认识到,由此也依照马库什组的任何单个成员或成员的子组来描述本公开。
如本领域技术人员将理解的,出于任何目的和所有目的,特别是依照提供的书面描述,本文公开的所有范围还涵盖任何和所有可能的子范围及其子范围的组合。任何列出的范围均可容易地被识别为充分描述,并且可将相同范围分解为至少相等的一半、三分之一、四分之一、五分之一、十分之几等。作为非限制性实例,可容易地将本文讨论的每个范围分解为下三分之一、中三分之一和上三分之一等。如本领域技术人员还将理解的,所有语言诸如“至多”、“至少”、“大于”、“小于”等,包括所列举的数字,并且是指可以随后被分解为以上所讨论的子范围的范围。最后,如本领域技术人员将理解的,范围包括每个单独的成员。因此,例如,具有1-3个细胞的组是指具有1、2或3个细胞的组。相似地,具有1-5个细胞的组是指具有1、2、3、4或5个细胞的组,等等。
定义
除非另有定义,否则本文所用的所有技术和科学术语具有本公开所属领域的技术人员通常理解的含义。以下参考文献为技术人员提供了此申请中使用的许多术语的一般定义:singleton等,dictionaryofmicrobiologyandmolecularbiology(第2版1994);thecambridgedictionaryofscienceandtechnology(walker编,1988);theglossaryofgenetics,第5版,r.rieger等(编),springerverlag(1991);和hale&marham,theharpercollinsdictionaryofbiology(1991)。如本文所用的,除非另有说明,否则下列术语具有以下赋予它们的含义。本文所用的术语仅出于描述特定实施方案的目的,并不旨在限制本公开。
如本文所用的,单数形式“一(a、an)”和“所述、该(the)”也旨在包括复数形式,除非上下文另外明确指示的。
如本文所用的,术语“约”或“大约”意指在如本领域普通技术人员所测定的特定值的可接受误差范围内,该误差将部分取决于如何测量或测定该值,即,测量系统的限度。例如,根据本领域的实践,“约”可意指在3个或多于3个标准偏差之内。可替换地,“约”可意指给定值的至多20%,优选地至多10%,更优选地至多5%,且更优选地至多1%的范围。可替换地,特别是关于生物系统或过程,术语可意指值的一个数量级内,优选地在值的5倍之内,且更优选地在值的2倍之内。
通常,关于诸如氢或h的某些元素意意指包括该元素的所有同位素。例如,如果将r基团定义为包括氢或h,则它也包括氘(deuterium)和氘(tritium)。因此,包含放射性同位素诸如氘(tritium)、c14、p32和s35的化合物在本技术范围内。基于本文的公开内容,用于将此类标记插入本技术的化合物中的过程对于本领域技术人员而言将是容易显而易见的。
通常,“取代的”是指如下定义的其中其含有的氢原子的一个或多个键被非氢或非碳原子的键所取代的有机基团(例如,烷基基团)。取代的基团还包括其中与一个或多个碳原子或一个或多个氢原子键合的一个或多个键被一个或多个与杂原子键合的键,包括双键或三键,所取代的基团。因此,除非另有说明,否则取代的基团被一个或多个取代基取代。在一些实施方案中,取代的基团用1、2、3、4、5或6个取代基取代。取代基基团的实例包括:卤素(即,f、cl、br和i);羟基;烷氧基、链烯氧基、芳氧基、芳烷氧基、杂环基、杂环基烷基、杂环基氧基和杂环基烷氧基;羰基(oxo);羧酸盐根;酯;氨基甲酸乙酯;肟;羟胺;烷氧基胺;芳烷氧基胺;硫醇;硫化物;亚砜;砜;磺酰;五氟磺酰基(即,sf5);磺酰胺;胺;n-氧化物;肼;酰肼;腙;叠氮化物;酰胺;尿素;脒;胍;烯胺;酰亚胺;异氰酸酯;异硫氰酸酯;氰酸盐;硫氰酸盐;亚胺;硝基;腈(即cn);等。
取代的环基团诸如取代的环烷基、芳基、杂环基和杂芳基基团还包括环和环系统,其中氢原子的键被碳原子的键替代。因此,取代的环烷基、芳基、杂环基和杂芳基也可被如下定义的取代或未取代的烷基、烯基和炔基基团取代。
烷基基团包括具有1至12个碳原子,通常为1至10个碳,或在一些实施方案中为1至8、1至6或1至4个碳原子的直链和支链烷基。烷基基团可以是取代的或未取代的。直链烷基基团的实例包括诸如甲基、乙基、正丙基、正丁基、正戊基、正己基、正庚基和正辛基的基团。支链烷基基团的实例包括但不限于异丙基、异丁基、仲丁基,叔丁基,新戊基,异戊基和2,2-二甲基丙基。代表性取代的烷基基团可被诸如上文所列的取代基取代一次或多次,且包括不限于卤代烷基(例如,三氟甲基)、羟烷基、硫烷基、氨烷基、烷基氨烷基、二烷基氨烷基、烷氧基烷基、羧基烷基等。
环烷基基团包括在一个或多个环中具有3至12个碳原子,或在一些实施方案中,具有3至10、3至8或3至4、5或6个碳原子的单、双或三环烷基基团。环烷基可以是取代的或未取代的。示例性单环环烷基基团包括,但不限于,环丙基、环丁基、环戊基、环己基、环庚基和环辛基基团。在一些实施方案中,环烷基基团具有3至8个环成员,而在其他实施方案中,环碳原子的数目为3至5、3至6或3至7。双环和三环系统包括桥联的环烷基基团和稠合的(fused)环,诸如,但不限于双环[2.1.1]己烷、金刚烷基、萘烷基等。取代的环烷基基团可被如上定义的非氢和非碳基团取代一次或多次。然而,取代的环烷基基团还包括被如上定义的直链或支链烷基基团取代的环。代表性取代的环烷基基团可以被诸如,但不限于,2,2-、2,3-、2,4-、2,5-或2,6-二取代的环己基基团(其可被诸如上面列出的取代基的取代基取代)单取代或取代一次以上。
环烷基烷基基团是如上定义的烷基,其中烷基基团的氢或碳键被对如上定义的环烷基的键取代。环烷基烷基基团可以是取代的或未取代的。在一些实施方案中,环烷基烷基基团具有4至16个碳原子、4至12个碳原子,且典型地具有4至10碳原子。取代的环烷基烷基基团可在基团的烷基、环烷基或烷基和环烷基两个部分处被取代。代表性取代的环烷基烷基基团可被单取代或取代一次以上,诸如,但不限于,被诸如以上多列的取代基单、双或三取代。
烯基基团包括如上所定义的直链和支链烷基基团,除了在两个碳原子之间存在至少一个双键外。烯基基团可以是取代的或未取代的。烯基基团具有2至12个碳原子,且典型地具有2至10个碳原子或,在一些实施方案中,具有2至8、2至6或2至4个碳原子。在一些实施方案中,烯基基团具有一个、两个或三个碳-碳双键。实例包括,但不限于尤其是乙烯基、烯丙基、-ch=ch(ch3)、-ch=c(ch3)2、-c(ch3)=ch2、-c(ch3)=ch(ch3)、-c(ch2ch3)=ch2。代表性取代的烯基基团可以被单取代或取代一次以上,例如但不限于被诸如以上所列的取代基单、双或三取代。
环烯基基团包括如上所定义的环烷基,其在两个碳原子之间具有至少一个双键。环烯基基团可以是取代的或未取代的。在一些实施方案中,环烯基基团可具有一个、两个或三个双键,但不包括芳族化合物。环烯基基团具有4至14个碳原子,或在一些实施方案中,具有5至14个碳原子、5至10个碳原子,或甚至具有5、6、7或8个碳原子。环烯基基团的实例包括环己烯基、环戊烯基、环己二烯基、环丁二烯基和环戊二烯基。
炔基基团包括如上所定义的直链和支链烷基,除了在两个碳原子之间存在至少一个三键外。炔基可以是取代的或未取代的。炔基基团具有2至12个碳原子,且典型地具有2至10个碳原子或,在一些实施方案中,具有2至8、2至6或2至4个碳原子。在一些实施方案中,炔基基团具有一个、两个或三个碳-碳三键。实例包括,但不限于尤其是–c≡ch、-c≡cch3、-ch2c≡cch3、-c≡cch2ch(ch2ch3)2。代表性取代的炔基基团可以被单取代或取代一次以上,诸如,但不限于,被诸如以上所列的取代基单、双或三取代。
芳基基团是不含有杂原子的环芳族烃。本文的芳基基团包括单环、双环和三环环系统。芳基基团可以是取代的或未取代的。因此,芳基基团包括,但不限于,苯基、薁基(azulenyl)、庚烯基、联苯基、芴基、菲基、蒽基、茚基、茚满基、并环戊二烯基和萘基基团。在一些实施方案中,芳基基团在基团的环部分中含有6-14个碳,在其它中含有6-12个或甚至6-10个碳原子。在一些实施方案中,芳基是苯基或萘基。短语“芳基基团”包括含有稠环的基团,例如稠合的芳族-脂族环系统(例如,茚满基、四氢萘基等)。代表性取代的芳基基团可被单取代(例如,甲苯基)或取代一次以上。例如,单取代的芳基基团包括,但不限于,2-、3-、4-、5-或6-取代的苯基或萘基基团,其可被诸如以上所列的取代基取代。
芳烷基基团是如上所定义的烷基,其中烷基的氢或碳键被对如上所定义的芳基基团的键替代。芳烷基可以是取代的或未取代的。在一些实施方案中,芳烷基基团含有7至16个碳原子、7至14个碳原子或7至10个碳原子。取代的芳烷基可在基团的烷基、芳基或烷基和芳基两个部分处被取代。代表性的芳烷基基团包括但不限于苄基和苯乙基基团和稠合的(环烷基芳基)烷基基团,诸如4-茚满基乙基。代表性取代的芳烷基基团可以被诸如以上所列的取代基取代一次或多次。
杂环基基团包括含有3个或更多个环成员的芳族(也称为杂芳基)和非芳族环化合物,其中环成员中的一个或多个是杂原子,诸如但不限于n、o和s。杂环基基团可以是被取代的或未取代的。在一些实施方案中,杂环基基团含有1、2、3或4个杂原子。在一些实施方案中,杂环基基团包括具有3至16个环成员的单、双或三环,而其它此类基团具有3至6、3至10、3至12或3至14个环成员。杂环基基团涵盖芳族,部分不饱和和饱和的环系统,诸如,例如,咪唑基、咪唑啉基和咪唑烷基。短语“杂环基基团”包括稠合的环种类,包括包含稠合的芳族和非芳族基团的环种类,诸如,例如,苯并三唑基、2,3-二氢苯并[1,4]二噁烷(2,3-dihydrobenzo[1,4]dioxinyl)和苯并[1,3]二噁英(benzo[1,3]dioxolyl)。该短语还包括含有杂原子(诸如,但不限于喹啉基)的桥联多环系统。该短语包括具有与环成员中的一个键合的其它基团(诸如烷基、氧代或卤素基团)的杂环基基团,其称为“取代的杂环基基团”。杂环基基团包括,但不限于,氮杂环丙烯基,氮杂环丁烷基、吡咯烷基、咪唑烷基、吡唑烷基、噻唑烷基、四氢噻吩基、四氢呋喃基、二噁英基(dioxolyl)、呋喃基、噻吩基、吡咯基、吡咯啉基、咪唑基、咪唑啉基、吡唑基、吡唑啉基、三唑基、四唑基、噁唑基、异噁唑基、噻唑基、噻唑啉基、异噻唑基、噻二唑基、恶二唑基、哌啶基、哌嗪基、吗啉基、硫代吗啉基、四氢吡喃基、四氢噻喃基、氧硫杂环己烷、二氧杂环己烷(dioxyl)、二噻烷基、吡喃基、吡啶基、嘧啶基、哒嗪基、吡嗪基、三嗪基、二氢吡啶基、二氢二硫杂环己二烯基(dihydrodithiinyl)、二氢二硫酮基(dihydrodithionyl)、高哌嗪基、奎宁环基、吲哚基、二氢吲哚基、异吲哚基、氮杂吲哚基(吡咯并吡啶基)、吲唑基、中氮茚基、苯并三唑基、苯并咪唑基、苯并呋喃基、苯并噻吩基、苯并噻唑基、苯并噁二唑基,苯并噁嗪基,苯并二硫杂环己二烯基(benzodithiinyl)、苯并氧硫杂环己二烯基(benzoxathiinyl)、benzothiazinyl、苯并噁唑基、苯并噻唑基、苯并噻二唑、苯并[1,3]二噁英基(benzo[1,3]dioxolyl)、吡唑并吡啶、咪唑并吡啶基(氮杂苯并咪唑基)、三唑并吡啶,异噁唑吡啶基(isoxazolopyridyl)、嘌呤基、黄嘌呤基、腺嘌呤基,鸟嘌呤基、喹啉基、异喹啉基、喹嗪基、喹喔啉基、喹唑啉基、噌啉基、酞嗪基、萘啶基(naphthyridinyl)、蝶啶基、硫杂萘基(thianaphthyl)、二氢苯并噻嗪基、二氢苯并呋喃基、二氢吲哚基、二氢苯并二氧杂苯基(dihydrobenzodioxinyl)、四氢吲哚基、四氢吲唑基、四氢苯并咪唑基、四氢苯并三唑基、四氢吡咯并吡啶基、四氢吡唑并吡啶基、四氢咪唑并吡啶基、四氢三唑并吡啶基和四氢喹啉基基团。代表性取代的杂环基基团可被单取代或取代一次以上,诸如,但不限于,吡啶基或吗啉基基团,其是2-、3-、4-、5-或6-取代的,或是被多种取代基诸如以上所列的取代基双取代的。
杂芳基基团是含有5个或更多个环成员的芳族环化合物,其中一个或多个环成员是杂原子,诸如,但不限于,n、o和s。杂芳基基团可以是单取代的或未取代的。杂芳基基团包括,但不限于,诸如以下的基团:吡咯基、吡唑基、三唑基、四唑基、噁唑基、异噁唑基、噻唑基、吡啶基、哒嗪基、嘧啶基、吡嗪基、噻吩基、苯并噻吩基,呋喃基,苯并呋喃基,吲哚基,氮杂吲哚基(吡咯并吡啶基)、吡唑并吡啶基、三唑并吡啶基、苯并三唑基、苯并噁唑基,苯并噻唑基,苯并噻二唑基,咪唑并吡啶基,异恶唑并吡啶基,硫杂萘基、嘌呤基、黄嘌呤基、腺嘌呤基、鸟嘌呤基、喹啉基、异喹啉基、四氢喹啉基、喹喔啉基和喹唑啉基基团。杂芳基基团包括其中所有环均是芳族的稠环化合物,诸如吲哚基,且包括其中仅一个环是芳族的稠环化合物,诸如2,3-二氢吲哚基基团。代表性取代的杂芳基基团可被多种取代基诸如以上所利的取代基取代一次或多次。
杂芳烷基基团是如上所定义的,其中烷基基团的氢或碳键被对如上所定义的杂芳基的键替代的烷基。杂芳烷基基团可以是取代的或未取代的。取代的杂芳烷基团可在基团的烷基、杂芳基或烷基和杂芳基两个部分处被取代。代表性取代的杂芳烷基团可被取代基诸如以上所列的取代基取代一次或多次。
本文所述的在本技术的化合物内具有两个或更多个附着点(即,二价、三价或多价)的基团通过使用后缀“烯”来指定。例如,二价烷基基团是亚烷基基团,二价芳基基团是亚芳基基团,二价杂芳基基团是杂亚芳基基团,等等。具有与本技术的化合物附着的单个点的取代基不使用“烯”名称来指示。因此,例如,氯乙基在本文中不被称为氯乙烯。
如本文所用的,术语向受试者“施用””药剂包括将该药剂引入或递送至受试者以进行其预期功能的任何途径。施用可通过任何适合的途径来实施,包括,但不限于,静脉内、肌内、腹腔内、皮下和如本文所述的其它适合的途径。施用包括自施用和通过另一人施用。
如本文所用的,术语“细胞群”是指表达相似或不同表型的至少两个细胞的组。在非限制性实例中,细胞群可包括表达相似或不同表型的至少约10个、至少约100个、至少约200个、至少约300个、至少约400个、至少约500个、至少约600个、至少约700个、至少约800个、至少约900个、至少约1000个细胞、至少约10,000个细胞、至少约100,000个细胞、至少约1×106个细胞、至少约1×107个细胞、至少约1×108个细胞、至少约1×109个细胞、至少约1×1010个细胞、至少约1×1011个细胞、至少约1×1012个细胞或更多个细胞。
术语“氨基酸”是指天然存在的和非天然存在的氨基酸,以及以与天然存在的氨基酸相似的方式起作用的氨基酸类似物和氨基酸模拟物。天然编码的氨基酸是20种常见的氨基酸(丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸和缬氨酸)以及吡咯赖氨酸和硒代半胱氨酸。氨基酸类似物是指是指具有与天然氨基酸相同的基本化学结构(即,与氢、羧基基团、氨基基团和r基团结合的α碳)的试剂,诸如高丝氨酸、正亮氨酸、蛋氨酸亚砜、蛋氨酸甲基锍(methioninemethylsulfonium)。此类类似物具有经修饰的r基团(诸如,正亮氨酸)或经修饰的肽骨架,但仍保留与天然存在的氨基酸相同的基本化学结构。在一些实施方案中,形成多肽的氨基酸为d型。在一些实施方案中,形成多肽的氨基酸为l型。在一些实施方案中,形成多肽的首选多个氨基酸为d型,且次要多个氨基酸为l型。
氨基酸在本文中通过其普遍知道的三个字母符号或通过iupac-iub生化命名委员会推荐的单字母符号来指代。同样地,核苷酸通过其普遍接受的单字母代码指代。
术语“多肽”、“肽”和“蛋白质”在本文中可互换地使用以指代氨基酸残基的聚合物。该术语适用于天然存在的氨基酸聚合物以及其中一个或多个氨基酸残基是非天然存在的氨基酸(例如,氨基酸类似物)的氨基酸聚合物。该术语涵盖任何长度的氨基酸链,包括全长蛋白,其中氨基酸残基通过共价肽键连接。
如本文所用的,“对照”是实验中用于比较目的的替换样品。对照可以是“阳性”或“阴性”。例如,当实验的目的是测定用于治疗特定疾病类型的治疗剂的功效的关联时,则阳性对照(已知呈现期望的治疗效果的组合物)和阴性对照(不接收疗法或接收安慰剂的受试者或样品)通常被采用。
如本文所用的,术语“有效量”或“治疗有效量”是指足以实现期望的治疗效果的药剂的量。在治疗应用的背景下,向所述受试者施用的治疗肽的量可取决于感染的类型和严重性,和个体的特征,诸如一般健康、年龄、性别、体重和对药物的耐受性。其还可取决于疾病的程度、严重性和类型。本领域技术人员将能够根据以上所述和其它因素确定适当剂量。
如本文所用的,术语“表达”是指多核苷酸被转录位mrna的过程和/或转录的mrna随后被翻译为肽、多肽或蛋白质的过程。如果多核苷酸衍生自基因组dna,表达可包括在真核细胞中剪接mrna。基因的表达水平可通过测量细胞或组织中mrna或蛋白的量来确定。在一个方面中,可以将来自一个样品的基因的表达水平直接与来自对照或参考样品的该基因的表达水平比较。在另一方面中,一个样品的基因的表达水平可直接与来自在施用本文公开的组合物后的样品的该基因的表达水平比较。术语“表达”还指以下事件中的一种或多种:(1)从细胞内的dna序列(例如,通过转录)产生rna模板;(2)在细胞内处理rna转录物(例如,通过剪接、编辑、5’帽形成和/或3’末端形成);(3)在细胞内将rna序列翻译为多肽或蛋白质;(4)在细胞内翻译后修饰多肽或蛋白质;(5)在细胞表面上呈递多肽或蛋白质;和(6)从细胞分泌或呈递或释放多肽或蛋白。
术语“接头”是指连结(connect)或连接(link)两个序列的合成序列(例如,氨基酸序列),例如,连接两个多肽域的合成序列。在一些实施方案中,接头含有1、2、3、4、5、6、7、8、9或10个氨基酸序列。
如本文所用的术语“免疫细胞”是指在免疫应答中起作用的任何细胞。免疫细胞是造血细胞,且包括淋巴细胞,诸如b细胞和t细胞;自然杀伤细胞;髓样细胞,诸如单核细胞、巨噬细胞、树突细胞、嗜曙红细胞、中性粒细胞、肥大细胞、嗜碱性粒细胞和粒细胞。
如本文所用的,术语“天然免疫细胞”是指免疫系统中天然存在的免疫细胞。
如本文所用的,术语“工程化免疫细胞”是指经遗传修饰的免疫细胞。
术语“淋巴细胞”是指所有未成熟的、成熟的、未分化的和分化的白淋巴细胞群,其包括组织特异性和专门化变种。作为非限制性实例,其涵盖b细胞、t细胞、nkt细胞和nk细胞。在一些实施方案中,淋巴细胞包括所有b细胞谱系,包括前b细胞、祖b细胞、早期前b细胞、晚期前b细胞、达前b细胞、小前b细胞、未成熟b细胞、成熟b细胞、血浆b细胞、记忆b细胞、b-1细胞、b-2细胞和无能an1/t3细胞群s。
如本文所用的,术语“t细胞”包括幼稚t细胞、cd4+t细胞、cd8+t细胞、记忆t细胞、活化的t细胞、无能t细胞、耐受性t细胞、嵌合b细胞和抗原特异性t细胞。
如本文所用的,“过继细胞治疗组合物”是指包含适于过继细胞转移的细胞的任何组合物。在示例性实施方案中,过继细胞治疗组合物包含选自肿瘤浸润淋巴细胞(til)、tcr(即,异源t细胞受体)修饰的淋巴细胞和car(即,嵌合抗原受体)修饰的淋巴细胞的细胞类型。在另一实施方案中,过继细胞治疗组合物包含选自t细胞、cd8+细胞、cd4+细胞、nk细胞、δ-γt细胞、调控t细胞和外周血单核细胞的细胞类型。在另一实施方案中,til、t细胞、cd8+细胞、cd4+细胞、nk细胞、δ-γt细胞、调控t细胞或外周血单核细胞形成过继细胞治疗组合物。在一个实施方案中,过继细胞治疗组合物包含t细胞。
如本文所用的,“肿瘤浸润性淋巴细胞”或til是指已离开血流并迁移到肿瘤中的白细胞。
如本文所用的,术语“抗体”不仅意指完整抗体分子,还意指抗体分子的片段,该片段保留免疫原结合能力。此类片段在本领域中也是熟知的,且在体外和体内均经常被采用。因此,如本文所用的,术语“抗体”不仅意指完整免疫球蛋白分子,而且是熟知的活性片段f(ab')2和fab。缺少完整抗体的fc片段的f(ab')2和fab片段,更容易从循环中清除,且可具有更少的完整抗体的非特异性组织结合(wahl等,j.nucl.med.24:316-325(1983))。本发明的抗体包含全天然抗体、单克隆抗体、人抗体、人源化抗体、骆驼化抗体、多特异性抗体、双特异性抗体、嵌合抗体、fab、fab'、单链v区片段(scfv)、单域抗体(例如,纳米抗体和单域骆驼抗体)、vnar片段、双特异性t细胞啮合(bite)抗体、小抗体、二硫键连接的fv(sdfv)和抗独特型(抗id)抗体、胞内抗体、融合多肽、非常规抗体和上述任何一种的抗原结合片段。特别地,抗体包括免疫球蛋白分子和免疫球蛋白分子的免疫活性片段,即,含有抗原-结合位点的分子。免疫球蛋白分子可以是任何类型(例如,igg、ige、igm、igd、iga和igy)、类别(例如,iggl、igg2、igg3、igg4、igal和iga2)或亚类。
在某些实施方案中,抗体是糖蛋白,其通过二硫键相互连结的包含至少两条重(h)链和两条轻(l)链。每条重链包含重链可变区(本文中简称为vh)和重链恒定(ch)区。重链恒定区包含三个域,chi、ch2和ch3。每条轻链包含轻链可变区(在本文中简称为vl)和轻链恒定cl区。轻链恒定区包含一个域,cl。vh和vl区s可进一步被细分为高可变性区,称作互补决定区(cdr),其间散置着更保守的区,称为框架区(fr)。每个vh和vl由三个cdr和以下列顺序从氨基酸至羧基端排布的四个fr:fr1、cdr1、fr2、cdr2、fr3、cdr3和fr4。重和轻链的可变区含有与抗原相互作用的结合域。抗体的恒定区可介导免疫球蛋白对宿主组织或因子(包括免疫系统的多种细胞(例如,效应细胞)和经典补体系统的第一组成(clq))的结合。如本文可互换地使用的,术语抗体的“抗原结合部分”、“抗原结合片段”或“抗原结合区”是指抗体的结合抗原且赋予对抗体抗原特异性的区或部分;抗原结合蛋白质(例如,抗体)的片段,该片段包括抗体的一种或多种保留特异性结合抗原(例如,肽/hla复合物)的能力的片段。已经显示可通过全长抗体的片段来执行抗体的抗原结合功能。术语抗体的“抗体片段”中涵盖的抗原结合部分的实例包括fab片段,其是由vl、vh、cl和chi域组成的单价片段;f(ab)2片段,其是包含通过铰链区处的二硫键桥链接的两个fab片段的二价片段;fd片段,其由vh和chi域组成;fv片段,其由抗体的单臂的vl和vh域组成;dab片段(ward等,nature341:544-546(1989)),其由vh域组成;和分离的互补决定区(cdr)。
抗体和抗体片段可全部或部分衍生自哺乳动物(例如,人、非人灵长类动物、山羊、豚鼠、仓鼠、马、小鼠、大鼠,兔和绵羊)或产生非哺乳动物抗体的动物(例如,鸡、鸭、鹅、蛇和有尾目两栖动物)。抗体和抗体片段可在动物中或在除动物外(诸如从酵母或噬菌体(例如,作为单抗体或抗体片段或作为抗体库的一部分))产生。
此外,尽管fv片段的两个结构域vl和vh由不同的基因编码,但可以使用重组方法通过合成接头将它们接合在一起,从而使它们成为单个蛋白链,其中vl和vh区配对以形成单价分子。这些被称为单链fv(scfv);参见例如,bird等,science242:423-426(1988);和huston等,proc.natl.acad.sci.85:5879-5883(1988)。使用本领域普通技术人员已知的常规技术获得这些抗体片段,且以与完整抗体相同的方式筛选片段的效用。
“分离的抗体”或“分离的抗原结合蛋白”是已经从其天然环境的组成中鉴定、分离和/或回收的一种抗体或蛋白。通常使用重组技术或使用本领域技术人员已知的肽合成技术来生成“合成抗体”或“重组抗体”。
如本文所用的,术语“单链可变片段”或“scfv”是共价连接以形成vh::vl异二聚体的免疫球蛋白(例如,小鼠或人)的重(vh)和轻链(vl)的可变区的融合蛋白。重(vh)和轻链(vl)直接接合或通过编码肽的接头(例如,约10、15、20、25个氨基酸)接合,该接头将vh的n端与vl的c端,或vh的c端与vl的n端连结。接头通常富含甘氨酸以提供柔韧性,且富含丝氨酸或苏氨酸以提供溶解度。接头可以连接细胞外抗原结合域的重链可变区和轻链可变区。在某些实施方案中,接头包含氨基酸具有如下提供的seqidno:21中所示的序列。
ggggsggggsggggs(seqidno:21)
在某些实施方案中,编码seqidno:21的氨基酸序列的核酸序列示于seqidno:22中,其提供如下:
ggcggcggcggatctggaggtggtggctcaggtggcggaggctcc(seqidno:22)
尽管去除了恒定区并引入了接头,但scfv蛋白保留了原始免疫球蛋白的特异性。单链fv多肽抗体可从如huston,等(proc.nat.acad.sci.usa,85:5879-5883(1988))所述的包含编码vh-和vl-的序列的核酸中表达。还参见,美国专利号5,091,513、5,132,405和4,956,778;和美国专利申请号20050196754和20050196754。已经描述了具有抑制活性的拮抗性scfv(参见,例如,zhao等,hybridoma(larchmt)27(6):455-51(2008);peter等,jcachexiasarcopeniamuscle(2012);shieh等,jimunol183(4):2277-85(2009);giomarelli等,thrombhaemost97(6):955-63(2007);fife等,jclininvst116(8):2252-61(2006);brocks等,immunotechnology3(3):173-84(1997);moosmayer等,therimmunol2(10):31-40(1995)。已经描述了具有刺激活性的激动性scfv(参见,例如,peter等,jbiolchem25278(38):36740-7(2003);xie等,natbiotech15(8):768-71(1997);ledbetter等,critrevimmunol17(5-6):427-55(1997);ho等,biochimbiophysacta1638(3):257-66(2003))。
如本文所用的,“f(ab)”是指抗体结构的片段,该片段结合抗原但为单价且不具有fc部分,例如,通过木瓜蛋白酶消化的抗体产生两个f(ab)片段和fc片段(例如,重(h)链恒定区;不结合抗原的fc区)。
如本文所用的,“f(ab')2”是指通过胃蛋白酶消化全igg抗体生成的抗体片段,其中此片段具有两个抗原结合(ab')(二价)区,其中每个(ab1)区包含两个不同独立的氨基酸链,通过s-s键连接的h链和轻(l)链的一部分以接合抗原,且其中其余的h链部分被连接在一起。“f(ab')2”片段可被分成两个单独的fab'片段。
如本文所用的,“cdr”被定义为抗体的互补决定区氨基酸序列,其是免疫球蛋白重和轻链的高变区。参见,例如,kabat等,sequencesofproteinsofimmunologicalinterest,4thu.s.departmentofhealthandhumanservices,nationalinstitutesofhealth(1987)。通常,抗体在可变区中包含三个重链和三个轻链cdr或cdr区。cdr提供大多数接触残基,以使抗体与抗原或表位结合。在某些实施方案中,使用kabat系统描绘cdr区(kabat,e.a.,等sequencesofproteinsofimmunologicalinterest,fifthedition,u.s.departmentofhealthandhumanservices,nihpublicationno.91-3242(1991))。
如本文所用的,术语“亲和力”意指结合强度的量度。不受理论的束缚,亲和力取决于抗体组合位点和抗原决定簇之间的立体化学拟合的紧密度,它们之间接触面积的大小以及带电和疏水基团的分布。亲和力还包括术语“亲合力”,其是指在形成可逆复合物(例如,单价或多价)之后抗原-抗体键的强度。用于计算抗体对抗原的亲和力的方法是本领域已知的,包含使用结合实验来计算亲和力。功能测定(例如,流式细胞术测定)中的抗体活性也反映了抗体亲和力。可以使用功能测定(例如,流式细胞术测定)在表型上表征和比较抗体和亲和力。在当前公开的主题中有用的核酸分子包括编码多肽或其片段的任何核酸分子。在某些实施方案中,在当前公开的主题中有用的核酸分子包括编码抗体或其抗原结合部分的核酸分子。此类核酸分子不需要与内源性核酸序列100%同一,但通常表现出基本同一性。与内源序列具有“基本同源性”或“基本同一性”的多核苷酸通常能够与双链核酸分子的至少一条链杂交。“杂交”是指在多种严格条件下在互补多核苷酸序列(例如,本文所述的基因)或其部分之间形成双链分子的对。(参见,例如,wahl,g.m.ands.l.berger,methodsenzymol.152:399(1987);kimmel,a.r.methodsenzymol.152:507(1987))。
例如,严格的盐浓度通常将会小于约约750mmnacl和75mm柠檬酸三钠,优选小于约500mmnacl和50mm柠檬酸三钠,更优选小于约约250mmnacl和25mm柠檬酸三钠。在缺失有机溶剂,例如甲酰胺的情况下可获得低严格度的杂交,而在至少约35%w/v的甲酰胺,更优选至少约50%w/v的甲酰胺的存在下可获得高严格度的杂交。严格的温度条件通常将包括至少约30℃,更优选至少约37℃,最优选至少约42℃的温度。可变的另外的参数,例如杂交时间、去污剂例如十二烷基硫酸钠(sds)的浓度以及载体dna的包含或排除,是本领域技术人员熟知的。通过根据需要组合这些多种条件来实现多种严格程度。在某些实施方案中,杂交将在30℃在750mmnacl、75mm柠檬酸三钠和1%w/vsds中发生。在某些实施方案中,杂交将在37℃在500mmnacl、50mm柠檬酸三钠、1%w/vsds、35%w/v甲酰胺和100μg/ml去饱和的鲑鱼精dna(ssdna)中发生。在某些实施方案中,杂交将在42℃在250mmnacl、25mm柠檬酸三钠、1%w/vsds、50%w/v甲酰胺和200μgssdna中发生。在这些条件下的有用的变化对于本领域技术人员将是显而易见的。
对于大多数应用,杂交后的洗涤步骤的严格性也将会变化。洗涤严格条件可通过盐浓度和通过温度来定义。如上所述,可以通过降低盐浓度或通过提高温度来提高洗涤严格性。例如,洗涤步骤的严格的盐浓度将优选小于约30mmnacl和3mm柠檬酸三钠,且最优选小于约15mmnacl和1.5mm柠檬酸三钠。洗涤步骤的严格的温度条件通常包括至少约25℃,更优选至少约42℃,甚至更优选至少约68℃的温度。在某些实施方案中,洗涤步骤将在25℃在30mmnacl、3mm柠檬酸三钠和0.1%w/vsds中发生。在某些实施方案中,洗涤步骤将在42℃在15mmnacl、1.5mm柠檬酸三钠和0.1%w/vsds中发生。在某些实施方案中,洗涤步骤将在68℃在15mmnacl、1.5mm柠檬酸三钠和0.1%w/vsds中发生。在这些条件下的另外的变化对于本领域技术人员将是显而易见的。杂交技术对于本领域技术人员是熟知的,且描述于,例如,benton和davis(science196:180(1977));grunstein和rogness(proc.natl.acad.sci.,usa72:3961(1975));ausubel等(currentprotocolsin分子的biology,wileyinterscience,newyork,2001);bergerandkimmel(currentprotocolsinmolecularbiology,wileyinterscience,newyork,2001);和sambrook等,molecularcloning:alaboratorymanual,coldspringharborlaboratorypress,newyork。
术语“基本同源”或“基本同一”意指多肽或核酸分子that与氨基酸序列(例如,本文所述的氨基酸序列中的任一个)或核酸序列(例如,本文所述的核酸序列中的任一个)参考表现出至少50%或更高的同源性或同一性。例如,此类序列在氨基酸或核酸水平上与用于比较序列(例如,野生型序列或天然序列)至少约60%、约65%、约70%、约75%、约80%、约85%、约90%、约95%或约99%同源或同一。在一些实施方案中,基本同源或基本同一的多肽相对于用于比较的序列含有一个或多个氨基酸氨基酸取代、插入或缺失。在一些实施方案中,基本同源或基本同一的多肽含有一个或多个非天然氨基酸或氨基酸类似物,包括d-氨基酸和逆转氨基(retroinversoamino),以替代同源的序列s。
通常使用序列分析软件(例如,sequenceanalysissoftwarepackageofthegeneticscomputergroup,universityofwisconsinbiotechnologycenter,1710universityavenue,madison,wis.53705,blast,bestfit,gap或pileup/prettybox程序)来测量序列同源性或序列同一性。此类软件通过将同源性程度分配给多种取代、缺失和/或其他修饰来匹配相同或相似的序列。在测定同一性程度的示例性方法中,可使用blast程序,其中e-3和e-100之间的概率得分指示密切相关的序列。
如本文所用的,两个氨基酸序列之间的同源性百分比等同于两个序列之间的同一性百分比。两个序列之间的同一性百分比是序列享有的相同位置的数目的函数(即,%同源性=相同位置数目/总位置数目x100),其中考虑了空位数目和每个空位的长度,需要引入这些空位用于两个序列的最佳比对。序列的比较和两个序列之间的同一性百分比的确定可使用数学算法来完成。
可使用e.meyers和w.miller(comput.appl.biosci.,4:11-17(1988))的算法(其已经并入align程序(2.0版本)中),使用pam120权重残留表,空位长度罚分12和空位罚分4来确定两个氨基酸序列之间的同源性百分比。此外,可使用needleman和wunsch(j.mol.biol.48:444-453(1970))算法(其已经在gcg软件安装包(可在www.gcg.com中获得)中并入至gap程序中),使用blossum62矩阵或pam250矩阵,和空位权重为16、14、12、10、8、6或4和长度权重为1、2、3、4、5或6来测定两个氨基酸序列之间的同源性百分比。
另外地或可替换地,本公开的主题的氨基酸序列可进一步被用作“查询序列”以对公共数据库进行检索以例如识别相关序列。可使用altschul,等(1990)j.mol.biol.215:403-10的xblast程序(2.0版本)进行此类检索。可用xblast程序、得分=50、字长=3进行blast蛋白检索以获得与本文公开的指定序列同源的氨基酸序列。为了获得用于比较目的的空位比对,可以如altschul等,(1997)nucleicacidsres.25(17):3389-3402中所述,利用空位blast。当利用blast和空位blast程序时,可使用相应程序(例如,xblast和nblast)的默认参数。
如本文所用的,术语“类似物”是指具有参考多肽或核酸分子的功能的结构上相关的多肽或核酸分子。
如本文所用的,术语“保守序列修饰”是指不显著影响或改变包含氨基酸序列的本公开的car的结合特性(例如,car的细胞外抗原结合域)的氨基酸修饰。保守修饰可包括氨基酸取代、添加、缺失。可以通过本领域已知的标准技术(诸如定点诱变和pcr介导的诱变)将修饰引入本公开的car的人scfv中。可根据其物理化学性质(诸如电荷和极性)将氨基酸分组。保守氨基酸取代是氨基酸残基被同一组内的氨基酸替代的一类。例如,氨基酸可以按电荷分类:带正电荷的氨基,其包括赖氨酸、精氨酸、组氨酸;带负电荷的氨基酸,其包括天冬氨酸/谷氨酸,中性电荷氨基酸,其包括丙氨酸、天冬酰胺、半胱氨酸、谷氨酰胺、甘氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸和缬氨酸。此外,氨基酸可按极性分类:极性氨基酸,其包括精氨酸(碱性极)、天冬酰胺、天冬氨酸(酸性极)、谷氨酸(酸性极)、谷氨酰胺、组氨酸(碱性极)、赖氨酸(碱性极)、丝氨酸、苏氨酸和酪氨酸;非极性氨基酸,其包括丙氨酸、半胱氨酸、甘氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、色氨酸和缬氨酸。因此,cdr区内一个或多个氨基酸残基可用来自同一组的其它氨基酸残基替代,且可以使用本文所述的功能测定法测试改变的抗体的保留功能(即,上述(1)中(c)中列出的功能)。在某些实施方案中,在指定序列或cdr区域内不超过一个、不超过两个、不超过三个、不超过四个、不超过五个残基被改变。
如本文所用的,术语“配体”是指结合受体的分子。特别地,配体结合其它细胞上的受体,以允许细胞对细胞的识别和/或相互作用。
如本文所用的,术语“共刺激信号传导域”或“共刺激域”是指包含共刺激分子的细胞内域的car的部分。共刺激分子是除抗原受体或fc受体以外的细胞表面分子,其在结合抗原后提供有效激活t淋巴细胞和使t细胞起作用所需的第二信号。此类共刺激分子的实例包括cd27、cd28、4-1bb(cd137)、ox40(cd134)、cd30、cd40、pd-1、icos(cd278)、lfa-1、cd2、cd7、light、nkd2c、b7-h2和特异性结合cd83的配体。因此,尽管本公开提供了衍生自cd28和4-1bb的示例性共刺激结构域,但可预期其它共刺激域与本文所述的car一起使用。包含一个或多个共刺激性信号传导域可增强表达car受体的t细胞的功效和扩增。细胞内信号传导和共刺激信号传导域可以以任何顺序串联连接到跨膜域的羧基端。
如本文所用的,术语“嵌合共刺激受体”或“ccr”是指结合抗原并提供共刺激信号,但不提供t细胞激活信号的嵌合受体。
如本文所用的,核酸分子的调控区意指正向或负向影响可操作连接的基因的表达的顺式作用核苷酸序列。调控区包括赋予基因可诱导(即,需要物质或刺激物以增加转录)的表达的核苷酸的序列。当诱导物存在或浓度增加时,基因表达可增加。调控区还包括赋予基因表达的抑制(即,物质或刺激物减少转录)的序列。当时阻遏物存在或浓度增加时,基因表达可降低。已知调控区影响、调节或控制许多体内生物活性,该生物活性包括细胞增殖、细胞生长和死亡、细胞分化和免疫调节。调控区通常与一种或多种反式作用蛋白结合,这导致基因转录增加或减少。
基因调节区的特定实例是启动子和增强子。启动子是位于转录或翻译起始位点周围的序列,通常位于翻译起始位点的5'端。启动子通常位于翻译起始站点的1kb之内,但也可位于更远处,例如,2kb,3kb,4kb,5kb或更多,至多且包括10kb。已知当增强子位于基因的5'或3'时或当增强子位于外显子或内含子中或是外显子或内含子的一部分时,增强子影响基因表达。增强子还可在距基因显著距离处(例如,在距约3kb、5kb、7kb、10kb、15kb或更多的距离处)起作用。
调控区还包括,但不限于,除了启动子区外的促进翻译、内含子的剪接信号、维持基因的正确阅读框以允许mrna的框内翻译的序列,以及终止密码子、前导序列和融合伴侣序列、用于创建多基因或多顺反子、信使、多腺苷酸化信号以提供基因感兴趣的和终止密码子的转录物的正确多腺苷酸化的内部核糖体结合位点(ires)元件,且可任选地包括在表达载体中。
如本文所用的,关于核酸序列、区、元件或域的“可操作地连接”抑制核酸区在功能上彼此相关。例如,编码前导肽的核酸可与编码多肽的核酸可操作地连接,凭此核酸可被转录并翻译以表达功能融合蛋白,其中前导肽影响融合多肽的分泌。在一些情况中,编码第一多肽(例如,前导肽)的核酸与编码第二多肽的核酸可操作地连接,且核酸被转录为单个mrna转录物,但mrna转录物的翻译可导致被表达的两个多肽中的一个。例如,琥珀终止密码子可位于编码第一多肽的核酸和编码第二多肽的核酸之间,使得,当导入部分琥珀抑制细胞时,所得的单个mrna转录物可被翻译以产生含有第一和第二多肽的融合蛋白,或可被翻译以仅产生第一多肽。在另一个实例中,启动子可与编码多肽的核酸可操作地连接,凭此启动子调节或介导核酸的转录。
如本文所用的,“合成的”,关于,例如,合成的核酸分子或合成的基因或合成的肽是指通过重组方法和/或通过化学合成方法产生的核酸分子或多肽分子。如本文所用的,通过使用重组dna方法的重组方式的产生意指使用分子生物学中熟知的方法来表达由dna编码的蛋白质。
如本文所用的,“表达”是指通过多核苷酸的转录和翻译来产生多肽的过程。可使用本领域已知的任何方法来评定多肽的表达水平,该方法包括,例如,测定从宿主细胞产生的多肽的量的方法。此类方法可包括,但不限于,通过elisa定量细胞裂解物中的多肽、凝胶电泳后的考马斯亮蓝染色、lowry蛋白测定和bradford蛋白测定。
如本文所用的,“宿主细胞”是用于接收、维持、复制和扩增载体的细胞。宿主细胞还可用于表达由载体编码的多肽。当宿主细胞分裂时,载体中含有的核酸被复制,由此扩增核酸。
如本文所用的,“载体”是当将载体转化到适当的宿主细胞中时,可以表达一种或多种异源蛋白质的可复制的核酸。关于载体,其典型地可通过限制性消化和连接将编码多肽的核酸或其片段引入其中的那些载体。关于载体,其还包括含有编码多肽的核酸的那些载体。载体用于将编码多肽的核酸引入宿主细胞中用于扩增核酸或用于表达/显示由核酸编码的多肽。载体典型地保持游离型,但是可被设计成实现基因或其部分整合到基因组染色体中。还预期了作为人工染色体(诸如酵母人工染色体和哺乳动物人工染色体)的载体。此类媒介物的选择和使用是本领域技术人员所熟知的。
如本文所用的,载体还包括“病毒载体”或“病毒载体”。“病毒载体”是与外源基因可操作地连接以将外源基因转移(作为媒介物或穿梭子)到细胞中的工程化病毒。
如本文所用的,“表达载体”包括能够表达与调控序列(诸如启动子区)可操作地连接的dna的载体,该调控序列能够影响此类dna片段的表达。此类另外的区段可包括启动子和终止子序列,且任选地可包括一个或多个复制起点、一个或多个可选择标记、增强子、多腺苷酸化信号等。表达载体通常衍生自质粒或病毒dna,或可含有二者的元件。因此,表达载体是指重组dna或rna构建体,诸如质粒、噬菌体、重组病毒或在引入至适当的宿主细胞中后导致克隆的dna的表达的其它载体。适当的表达载体是本领域技术人员所熟知的,且包括在真核细胞和/或原核细胞中可复制的载体和保留游离型的载体或整合到宿主细胞基因组中的载体。
如本文所用的,术语“疾病”是指损害或干扰细胞、组织或器官的正常功能的任何病况或病症。疾病的实例包括瘤形成或细胞的病原体感染。
“有效量”(或“治疗有效量”)是足以在治疗后影响有益或期望的临床结果的量。可以一个或多个剂量向受试者施用有效量。就治疗而言,有效量是足以缓和、改善、稳定、逆转疾病或减慢疾病进展(例如瘤形成)或以其他方式减轻疾病的病理后果(例如瘤形成)的量。有效量通常由医师根据具体情况确定,并且在本领域技术人员的能力范围内。当确定适当剂量以达到有效量时,通常要考虑几个因素。这些因素包括受试者的年龄、性别和体重,所治疗的病况、病况的严重性以及所施用的工程化免疫细胞的形式和有效浓度。
如本文所用的,术语“瘤形成”是指以细胞或组织的病理性增殖及其随后向其他组织或器官的迁移或侵袭为特征的疾病。瘤形成的生长通常是不受控制的和进行性的,且在不会引起正常细胞的增殖或会导致正常细胞增殖中断的条件下发生。瘤形成可影响多种细胞类型、组织或器官,其包括但不限于选自下组的器官:膀胱、结肠、骨头、脑、乳腺、软骨、神经胶质、食道、输卵管、胆囊、心脏、肠、肾脏、肝、肺、淋巴结、神经组织、卵巢、胸膜、胰腺、前列腺、骨骼肌、皮肤、脊髓、脾脏、胃、睾丸、胸腺、甲状腺、气管、泌尿生殖道、输尿管、尿道、子宫和阴道,或其组织或细胞类型。瘤形成包括癌症,诸如肉瘤、癌或浆细胞瘤(浆细胞的恶性肿瘤)。
如本文所用的,术语“异源核酸分子或多肽”是指通常在细胞或从细胞获得的样品中不存在的核酸分子(例如,cdna、dna或rna分子)或多肽。此核酸可来自另一生物体,或其可以是,例如,通常在细胞或样品中不表达的mrna分子。
如本文所用的,术语“免疫应答细胞”是指起免疫应答作用的细胞或其祖细胞或其后代。
如本文所用的,术语“调节”是指正向或负向改变。示例性调节包括约1%、约2%、约5%、约10%、约25%、约50%、约75%或约100%变化。
如本文所用的,术语“增加”是指正向改变至少约5%,其包括,但不限于,正向改变约5%、约10%、约25%、约30%、约50%、约75%或约100%。
如本文所用的,术语“降低”是指负向改变至少约5%,其包括,但不限于,负向改变约5%、约10%、约25%、约30%、约50%、约75%或约100%。
如本文所用的,术语“分离的细胞”是指从天然伴随细胞的分子组成和/或细胞组成中分离的细胞。
如本文所用的,术语“分离的”、“纯化的”或“生物学上纯净的”是指从其天然状态下通常与之伴随的组分不同程度地释放出来的材料。“分离”表示与原始来源或环境的分开程度。“纯度”表示比分离更高的分开程度。纯化的”或“生物学上纯净的”蛋白充分不含其他材料,使得任何杂质均不会实质性地影响蛋白质的生物学性质或引起其他不利后果。即,如果当通过重组dna技术产生时基本上不含细胞材料、病毒材料或培养基,或者当通过以化学方法合成时基本上不含化学前体或其他化学品,则本公开的主题中的核酸或多肽是纯化的。纯度和均质性通常使用分析化学技术测定,例如聚丙烯酰胺凝胶电泳或高效液相色谱法。术语“纯化的”可表示核酸或蛋白质在电泳凝胶中基本上产生一条带。对于可以进行修饰(例如磷酸化或糖基化)的蛋白质,不同的修饰可产生不同的分离的蛋白质,其可被单独纯化。
如本文所用的,术语“分泌的”意指通过内质网、高尔基体经由分泌途径从细胞释放的多肽,以及在细胞质膜上瞬时融合从而将蛋白质释放到细胞外的囊泡。小分子(诸如药物)也可通过膜扩散到细胞外而被分泌。
如本文所用的,术语“特异性结合”或“与……特异性结合”或“特异性靶向”意指识别并结合感兴趣的生物分子(例如,多肽)的多肽或其片段,但其实质上不识别和结合样品(例如,生物样品)中的其它分子,该样品包括或表达肿瘤抗原。
如本文所用的,术语“治疗(treating)”或“治疗(treatment)”是指试图改变所治疗的个体或细胞的疾病过程的临床干预,并且可用于预防或在临床病理过程中进行。治疗的治疗效果包括,不限于,预防疾病的发生或复发、减轻症状、减少疾病的任何直接或间接病理后果、预防转移、降低疾病进展的速度、改善(amelioration)或缓和疾病状态,以及缓解或改善(improved)预后。通过预防疾病或病症的进展,治疗可预防在受影响或经诊断的受试者或疑似患有病症的受试者中由于病症引起的恶化,而且治疗可预防出于病症或疑似患有病症的受试者中的病症的发作或病症的症状。
如本文所用的,术语“受试者”指任何动物(例如,哺乳动物),包括,但不限于,人、非人灵长类动物、啮齿动物等(例如,接受特定治疗,或从中被收集细胞的动物)。
综述
已经表明了嵌合抗原受体(car)t细胞的过继转移是针对b细胞all和其它造血系统癌症的有效疗法。然而,初次失败(primaryfailure)、不完全缓解和复发仍经由多种机制发生,该机制包括靶抗原的丢失。实体肿瘤已经对此形式的疗法更具抵抗力。因此,对于许多癌症类型,需要更大的效能和机制来击败免疫抑制性肿瘤微环境。肿瘤对cart细胞活性的抵抗力、抗原丢失变体逃逸和复发仍然是cart细胞疗法成功的重要障碍。本文提供了工程免疫细胞,其包括解决这些问题的包含工程化免疫细胞的组合物及其使用方法。如本文所述的,免疫细胞可被工程化以组成型或条件性表达在癌症细胞表面或在肿瘤微环境中从非毒性前药合成并释放有效细胞毒性化学疗法的酶(即,前药转化酶)。(图1)。在一些实施方案中,工程化免疫细胞额外地表达嵌合抗原受体用于向靶位点递送免疫细胞。这些工程化免疫细胞在本文中可互换地称为合成性酶激活的杀伤(syntheticenzymeactivatedkiller,seaker)细胞。不旨在受理论的束缚,有效的抗瘤药物在肿瘤部位的局部释放有望克服肿瘤对t细胞的微环境免疫抵抗,以及由于抗原丢失变体的逃逸,因为药物是不依赖抗原的,不受其他细胞抑制,并可局部扩散。本文提供的方法允许模块化使用大范围的药物,这取决于所期望的应用。药物还可与工程化免疫细胞(例如cart细胞)的直接基于免疫的细胞毒性作用协同作用。通过工程免疫细胞在癌细胞处选择性合成药物也可降低施用的药物的全身毒性。
本文提供了表达与很多种前药组合的抗原受体(例如,嵌合抗原受体)的工程化免疫细胞(例如,cart细胞)。在一些实施方案中,前药的活性药物是细胞毒性化学治疗药物。细胞毒性化学治疗药物可以是非靶向的或靶向的。示例性非靶向的细胞毒性癌症化学疗法以及化合物的前药形式是本文提供的或本领域已知的,其包括,例如,氮芥、甲氨蝶呤、多柔比星(doxorubicin)、核苷和ams。示例性靶向的癌症化学疗法以及化合物的前药形式是本文提供的或本领域已知的,其包括,例如,厄洛替尼(erlotinib)、达沙替尼(dasatinib)和酪氨酸激酶抑制剂。此外,如本文所述的,可采用调节性药物的前药、热休克蛋白(hsp)抑制剂和多药耐药性(mdr)抑制剂。
影响前药转化酶的局部分布的先前方法已经包括抗体指导的酶前药疗法(adept)。adept被认为是利用酶催化以在肿瘤部位生成高局部浓度的细胞毒性药物的一种手段。adept是传统抗体-药物缀合物(adc)的替代方法,adc受抗体上缀合位点的数目限制。典型地,每个抗体adc仅递送约1-4个药物分子。通过比较,adept依赖于单克隆抗体(mab)介导的以化学方法缀合物的活化酶(即,前药转化酶)向肿瘤位点的递送,其中抗体结合肿瘤特异性抗原。然后全身性施用非毒性前药,其被酶/抗体缀合物局部切割以在肿瘤位点处合成细胞毒性药物。adept方法提供了优于传统adc的几个优势,即前药的预定控制和停止、小前药和药物相对于mab的快速扩散、酶促驱动的药物浓度的局部增加以及附近的抗原丢失的癌症变体的可能杀伤。然而,由于酶/抗体缀合物在血液中的持久性,adept的毒性仍然是问题。此外,由于抗体携带的酶的肿瘤与血液比率差,难以实现良好的治疗指数。事实上,考虑到通常约0.5%或更少的注射剂量的抗体在人中的实体肿瘤的位点处发现,且其余的99.5%或更多在身体中的其他地方,所以adept和adc方法具有有限的功效。
在一些实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)或结合靶抗原(诸如肿瘤抗原和前药转化酶)的其它细胞表面配体。在一些实施方案中,t细胞受体是野生型或天然的t细胞受体。在一些实施方案中,t细胞受体是嵌合t细胞受体(car)。
在本文提供的示例性实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)(例如,car)或结合cd19肿瘤抗原的其它细胞表面配体。在一些实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)(例如,car)或结合在mhc分子的背景下呈递的cd19肿瘤抗原的其它细胞表面配体。在一些实施方案中,结合在hla-a2分子的背景下呈递的cd19肿瘤抗原。cd19是b细胞谱系特异性抗原,cd19已成为人试验中许多最有效的cart细胞的靶标。cd19由于其充分表征的活性、药理学和毒性而成为模型抗原。
在本文提供的示例性实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)(例如,car)或结合“黑色素瘤中优先表达的抗原”(prame)肿瘤抗原的其它细胞表面配体。在一些实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)(例如,car)或结合在mhc分子的背景呈递的prame肿瘤抗原的其它细胞表面配体。在一些实施方案中,在hla-a2分子的背景下呈递的prame肿瘤抗原。prame蛋白是一种目前不可成药的视黄酸受体结合蛋白,其参与分化、增殖阻滞和细胞凋亡。prame是一种癌症-睾丸抗原,其在限于睾丸、卵巢和子宫内膜的健康成人组织中有限表达。然而,prame在多种癌症(包括乳腺癌、结肠癌、急性白血病(50%)、黑色素瘤(90%)、淋巴瘤、肉瘤等)中过表达,这使其成为极具吸引力的治疗靶标。蛋白酶体加工后,在hla-1单倍型hla*a02:01(hla-a2)的背景下,在细胞表面上呈递prame300-309肽(alyvdslffl(seqidno:23)。
在本文提供的示例性实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)(例如,car)或结合肾母细胞瘤(wilm’stumor)蛋白1(wt1)肿瘤抗原的其它细胞表面配体。在一些实施方案中,本文提供的工程化免疫细胞表达t细胞受体(tcr)(例如,car)或结合在mhc分子的背景下呈递的wt1肿瘤抗原的其它细胞表面配体。在一些实施方案中,结合在hla-a2分子的背景下呈递的wt1肿瘤抗原。wt1是一种重要的、经验证且nci排名最高的癌症靶抗原。wt1是锌指转录因子,其对泌尿生殖系统的胚胎发育至关重要。wt1在大多数白血病(包括aml、cml、all和mds)以及骨髓瘤和几种实体瘤(尤其是卵巢癌和间皮瘤)中高表达。wt1疫苗已进入针对具有多种癌症患者的临床试验。wt1以其对形成无性系的白血病细胞存活的重要性以及在异种移植的nod/scid小鼠中用对wt1肽具有特异性的t细胞治疗肿瘤的能力而不会不利影响正常造血而著称。wt1肽疫苗已与疾病的完全或部分缓解和延长的存活相关联。
cart细胞疗法和tcrt细胞疗法后显著的毒性甚至死亡,以及gvhd的高发生率及其相关的短期和长期发病率和死亡率,强烈需要更好地控制各种t细胞疗法的毒性的机制。在本文提供的一些实施方案中,本文提供的工程化免疫细胞在细胞内(例如,在工程化免疫细胞的细胞质内)表达前药转化酶,使得当施用穿透细胞的前药时,工程化免疫细胞将死亡,其中穿透细胞的前药被细胞内前药转化酶转化为活性药物。在一些实施方案中,穿透细胞的前药对工程化免疫细胞的细胞类型具有特异性或穿透特定的细胞类型。在一些实施方案中,穿透细胞的前药对任何细胞类型不具有特异性。不表达细胞内前药转化酶的其他正常细胞将不受影响。因此,本文所述的技术可以用作“杀伤开关”或自杀载体,以控制引起毒性或需要被终止的细胞疗法。本方法的一个优点是传统的化学治疗药物可用于实现杀伤开关作用。
与现有的adept和adc技术相比,本文提供的表达与在细胞表面上表达或分泌的前药转化酶结合的嵌合抗原受体的工程化免疫细胞(例如,cart细胞)提供了许多优点。这些优势的非详细列表包括,例如:1)工程化免疫细胞(例如,cart细胞)显著增加肿瘤位点处的活化酶的量(例如,100倍或更多倍)的能力。这是因为工程化免疫细胞含有编码前药转化酶的核酸,以通过细胞来表达许多拷贝的酶。此外,当工程化免疫细胞在肿瘤位点遇到肿瘤特异性抗原时,其将会广泛增殖(例如,100倍或更多),从而显著增加酶的产生。2)可在工程化免疫细胞(例如,cart细胞)在靶位点达到峰值后施用前药,这可以降低药物的全身性暴露并确保前药的最大局部转化。3)通过用编码嵌合抗原和前药转化酶的核酸体外转导免疫细胞,可容易地生成工程化免疫细胞(例如,cart细胞)。因此,与adept或adc形成对比,没有难于制造的化学缀合物。4)工程化免疫细胞(例如,cart细胞)也可以具有自身的累加或协同抗肿瘤活性。进一步地,可通过选择嵌合抗原受体中包括的共刺激分子来调整工程化免疫细胞(例如,cart细胞)的活性。5)可采用前药转化酶的门控条件表达以允许更好地控制毒性。6)如果仅需要药物介导的癌症杀伤和/或不需要cart介导的杀伤,则可进一步修饰工程免疫细胞(例如,cart细胞),以去除(engineerout)t细胞介导的炎症应答(例如,细胞因子释放),t细胞介导的炎症应答是在人类中所见的大量毒性的原因。7)进一步地,“杀伤开关”经由前药转化酶的细胞内表达可允许工程化免疫细胞的选择性破坏。
前药和前药转化酶
本文提供的工程化免疫细胞(例如,cart细胞)表达至少一种将前药转化为活性药物的前药转化酶。在一些实施方案中,前药转化酶在工程化免疫细胞的表面上表达。在一些实施方案中,前药转化酶是由工程化免疫细胞分泌的。该酶可以是能够将前药转化为活性药物且通常不在细胞(例如,哺乳动物细胞,诸如人细胞)的表面上表达或不被释放到循环中的任何酶。在一些实施方案中,本文提供的工程化免疫细胞表达细胞内前药转化酶(例如在工程化免疫细胞的细胞质中)。在一些实施方案中,细胞内前药转化酶充当“杀伤开关”,允许经由穿透细胞的前药的细胞内转化来选择性破坏工程免疫化细胞。
在一些实施方案中,本文提供的工程化免疫细胞(例如,cart细胞)表达在工程化免疫细胞的表面上表达的前药转化酶和细胞内前药转化酶。在一些实施方案中,本文提供的工程化免疫细胞(例如,cart细胞)表达由工程化免疫细胞分泌的前药转化酶和细胞内前药转化酶。在一些实施方案中,细胞表面或分泌的前药转化酶和细胞内前药转化酶具有相同的酶活性(例如,细胞表面或分泌的前药转化酶与细胞内前药转化酶是相同的酶)。在一些实施方案中,细胞表面或分泌的前药转化酶和细胞内前药转化酶具有不同的酶活性(例如,细胞表面或分泌的前药转化酶和细胞内的前药转化酶是不同的酶)。除非特别区分,否则本文提及的前药转化酶是指细胞表面或分泌的前药转化酶、细胞内前药转化酶或两者。
在一些实施方案中,前药转化酶将会通过从前药中去除保护基将前药转化为活性药物。在一些实施方案中,保护基将作为整体从前药被切割。在一些实施方案中,酶切割或改变保护基的一部分,得到不稳定的部分切割或改变的保护基,导致自发除去该基团的其余部分。
在一些实施方案中,前药转化酶是非哺乳动物酶。适合的非哺乳动物酶包括细菌酶。细菌酶包括羧肽酶,诸如羧肽酶g2(cpg2),其是从假单胞菌属种分离rs-16的细菌水解酶,其基于其从叶酸衍生物中切割n-连接的谷氨酸盐的能力(假单胞菌属γ-谷酰基水解酶ec3.4.22.12,如在wo88/07378以及levy和goldstein,j.biol.chem.242:2933(1967)中公开的)。cpg2对芳香族n-酰基部分和谷氨酸盐之间的切割具有特异性,并已广泛用于adept策略,包括一些已进入针对癌症的人类临床试验的系统。在一些实施方案中,通过在谷氨酸盐水解后自发分解为惰性副产物的多种自降解连接子(图2b)偶联至药物中的胺或醇部分(图2a),可将活性药物掩盖为谷氨酸盐前药。在某些实施方案中,前药转化酶是cpg2。在一些实施方案中,cpg2包含seqidno:1中所示的序列,其是cpg2的分泌形式,包括用于分泌的前导序列。在一些实施方案中,cpg2包含seqidno:3中所示的序列,其是cpg2多肽,其无seqidno:1的前导序列,且针对人表达而优化。在一些实施方案中,cpg2包含seqidno:5中所示的序列,其是cpg2的示例性分泌形式,包括用于分泌的前导序列且针对人表达而优化。在一些实施方案中,cpg2包含seqidno:7或40中所示的序列,其是针对人表达而优化的cpg2的示例性跨膜形式,其具有cd8前导序列(例如,seqidno:11)、跨膜部分和具有破坏的lyk结合域的cd8细胞内部分(例如,seqidno:15)。
在一些实施方案中,前药转化酶是β-内酰胺酶,诸如阴沟肠杆菌β-内酰胺酶。β-内酰胺酶(ec3.5.2.6)是由多种细菌产生的丝氨酸蛋白酶;它催化青霉素和其它类似的β-内酰胺抗生素中的β-内酰胺部分水解为β-氨基酸。β-内酰胺酶对含有β-内酰胺的化合物具有高度选择性。头孢菌素由于其能够被β-内酰胺酶切割β-内酰胺环后从c3'位置释放药物,而被广泛用作β-内酰胺酶激活的前药的核心。在一些实施方案中,β-内酰胺酶包含seqidno:43中所示的序列,其是β-内酰胺酶的分泌形式,包括用于分泌的前导序列。在一些实施方案中,β-内酰胺酶包含seqidno:44中所示的序列,其是不具有seqidno:43的前导序列且针对人表达而优化的β-内酰胺酶多肽。在一些实施方案中,β-内酰胺酶包含seqidno:45中所示的序列,其是β-内酰胺酶的示例性分泌形式,包括用于分泌的前导序列且针对人表达而优化。在一些实施方案中,β-内酰胺酶包含seqidno:46或47中所示的序列,其是β-内酰胺酶的示例性跨膜形式,其针对人表达而优化,具有cd8前导序列(例如,seqidno:11)、跨膜部分和任选地具有破坏的lyk结合域的cd8细胞内部分(例如,seqidno:15)。
其它适合的非哺乳动物酶的实例包括硝基还原酶,诸如如wo93/08288中公开的大肠杆菌(e.coli)硝基还原酶、胸苷激酶(tk)(其包括病毒tk,诸如vzv或hsvtk)、β-葡萄糖醛酸苷酶、青霉素v酰胺酶、青霉素g酰胺酶和胞嘧啶脱氨酶。
在一些实施方案中,酶是不在人类中天然存在的哺乳动物酶。在一些实施方案中,酶是人类酶,其正常不可接近前药,在细胞外以有限量表达,在前药不到达的区室中表达,由可通过前药激活而被杀伤并且对患者来说非至关重要的细胞表达,和/或由对活性药物具有抗性或不被活性药物杀伤的细胞表达。这包括,例如,来自其他物种的酶以及哺乳动物酶,其以对前药有选择性的方式被改变。在一些实施方案中,改变是指通过天然酶将前药转化为活性药物的速率比经改变的酶起作用的速率小一个或多个数量级。可通过标准重组dna技术(例如,通过克隆酶,确定其基因序列并通过诸如定点诱变的方法来改变基因序列)来制备经改变的酶。在一些实施方案中,前药转化酶是羧肽酶a,或其突变体,例如羧肽酶a的t268g突变体。
对于分泌的前药转化酶、跨膜前药转化酶或细胞内前药转化酶的表达,采用基于真核生物的表达系统。对于分泌或跨膜表达,在蛋白质的n端处包括信号肽。信号序列或前导序列可以是存在于新合成的蛋白质的n端的肽序列(约5、约10、约15、约20、约25或约30个氨基酸长),其指导所述蛋白质进入分泌途径。在某些实施方案中,信号肽与前药转化酶(例如,cpg2酶或β-内酰胺酶)的n端共价连接。在某些实施方案中,信号肽包含cpg2信号序列或β-内酰胺酶信号序列。在某些实施方案中,信号肽包含cpg2信号序列,该cpg2信号序列包含具有如下提供的seqidno:9中所示的序列的氨基酸。
mrpsihrtaiaavlatafvagt(seqidno:9)。
编码seqidno:9的氨基酸序列的核苷酸序列示于seqidno:10中,seqidno:10提供如下:
atgcgaccgagtatccacagaacagcaatagctgctgtgcttgcaacagcgtttgtagcgggcacg(seqidno:10)
在某些实施方案中,信号肽包含cd8信号多肽,该cd8信号多肽包含具有如下提供的seqidno:11中所示的序列的氨基酸。
malpvtalllplalllhaarp(seqidno:11)
编码seqidno:11的氨基酸序列的核苷酸序列示于seqidno:12中,seqidno:12提供如下:
atggccctgccagtaacggctctgctgctgccacttgctctgctcctccatgcagccaggcct(seqidno:12)。
对于细胞内表达,表达的前药转化酶不包含信号肽。因此,该酶在翻译时不被定位于内质网,而是保留在细胞质中。
对于前药转化酶的细胞表面表达,经表达的蛋白被锚定至细胞膜。在一些实施方案中,前药转化酶包含跨膜域。在一些实施方案中,跨膜域被共价附接至cpg2酶或β-内酰胺酶的c端。在一些实施方案中,跨膜域被共价附接至cpg2酶、β-内酰胺酶的n端。在一些实施方案中,跨膜域是cd8跨膜域。在一些实施方案中,跨膜域包含跨膜跨越部分。在一些实施方案中,膜跨膜部分包含以下序列:
iyiwaplagtcgvlllslvit(seqidno:38)
在一些实施方案中,cd8跨膜域还包含细胞内部分,该细胞内部分具有氨基酸序列:lycnhrnrrrvckcprpvvksgdkpslsaryv(seqidno:13)。
在一些实施方案中,cd8跨膜域包含细胞内部分,在细胞内部分中天然lyk结合域被破坏以使cd8的内源功能与cpg2或β-内酰胺酶分子解偶联。例如,在一些实施方案中,具有破坏的lyk结合域的cd8跨膜域部分具有下列序列:
lycnhrnrrrvgggrpvvksgdkpslsaryv(seqidno:15)
在一些实施方案中,跨膜cpg2酶具有下列氨基酸序列,其包括在内质网中的加工期间被去除的信号肽:
malpvtalllplalllhaarpalaqkrdnvlfqaatdeqpaviktleklvnietgtgdaegiaaagnfleaelknlgftvtrsksaglvvgdnivgkikgrggknlllmshmdtvylkgilakapfrvegdkaygpgiaddkggnavilhtlkllkeygvrdygtitvlfntdeekgsfgsrdliqeeakladyvlsfeptsagdeklslgtsgiayvqvqitgkashagaapelgvnalveasdlvlrtmniddkaknlrfqwtiakagqvsniipasatlnadvryarnedfdaamktleeraqqkklpeadvkvivtrgrpafnageggkklvdkavayykeaggtlgveertgggtdaayaalsgkpvieslglpgfgyhsdkaeyvdisaiprrlymaarlimdlgagkypydvpdyagggiyiwaplagtcgvlllslvitlycnhrnrrrvgggrpvvksgdkpslsaryv(seqidno:40)。
在一些可替换的实施方案中,用于表达cpg2酶的构建体包含不具有信号肽的cpg2。
在一些实施方案中,跨膜β-内酰胺酶具有下列氨基酸序列,其包括在内质网中的加工期间被去除的信号肽:
malpvtalllplalllhaarptpvsekqlaevvantitplmaaqsvpgmavaviyqgkphyytfgkadiaankpvtpqtlfelgsisktftgvlggdaiargeislddavtrywpqltgkqwqgirmldlatytagglplqvpdevtdnasllrfyqnwqpqwkpgttrlyanasiglfgalavkpsgmpyeqamttrvlkplkldhtwinvpkaeeahyawgyrdgkavrvspgmldaqaygvktnvqdmanwvmanmapenvadaslkqgialaqsrywrigsmyqglgwemlnwpveantvvegsdskvalaplpvaevnppappvkaswvhktgstggfgayvafipekqigivmlantsypnparveaayhilealqypydvpdyaggglycnhrnrrrvckcprpvvksgdkpslsaryv(seqidno:47)。
在一些可替换的实施方案中,用于表达β-内酰胺酶的构建体包含不具有信号肽的β-内酰胺酶。
在一些实施方案中,cpg2或β-内酰胺酶酶经由糖基磷脂酰肌醇(gpi)-接头被附接至细胞表面。glypiated(gpi-连接的)蛋白含有将蛋白质靶向至内质网(er)的内腔的可切割的、疏水性氨基末端信号序列和指导gpi锚定的可切割的、羧基端信号序列。gpi-锚定信号由疏水区组成,该疏水区通过亲水性间隔区与gpi附接位点(ω-位点)隔开。(参见例如galian等(2012)jbiolchem.11;287(20):16399–16409)。
因为经表达的分泌的或跨膜的前药转化酶是通过高尔基体和内质网加工的,所以它们可被糖基化,与其非糖基化形式相比,糖基化可导致酶活性降低。因此,在一些实施方案中,前药转化酶通过在一个或多个(例如,两个、三个或四个)糖基化位点处的取代、缺失或插入而从其天然序列改变。例如,在cpg2的一级氨基酸序列中,存在三个此类共有糖基化基序,其位于seqidno:1的残基asn222、asn264和asn272。在一些实施方案中,改变这些糖基化位点中的一个或多个以去除糖基化位点。在一些实施方案中,去除asn222、asn264和asn272中的一个或多个。在一些实施方案中,用亮氨酸或谷氨酰胺取代asn222、asn264和asn272中的一个或多个(参见,例如,seqidno:3、5、7和40,其代表用于表达的示例性cpg2多肽)。
在一些实施方案中,前药转化酶中的至少1、2、3、4、5、6、7、8、9、10或更多个糖基化位点被取代。在一些实施方案中,在共有糖基化位点处或附近缺失或插入1、2、3、4、5或更多个氨基酸。典型地,此类改变将使得该酶保持其以与未变化的非糖基化酶基本相同的速率将前药转化为活性药物的能力。在此背景中,“基本未变化的”是在1个数量级内,诸如从少约2倍活性至2、3、4、5、6、7、8、9或10倍或更多活性。
在一些实施方案中,相对于其天然形式,酶可通过截短、取代、缺失或插入而改变,只要酶的活性是如上所定义的基本未变化的。例如,在一些实施方案中,采用相对于天然全长序列的n端和/或c端序列中的小的截短(例如,约1至约20个氨基酸)。在一些实施方案中,需要此类截短以将多肽连接至本文所述的多种其它信号序列或肽。可以在适合的模型系统中测量改变的酶的活性,该模型系统可以以本领域已知的常规方式制备。
选择用于在系统中使用的前药以与前药转化酶相容,即,使得该酶能够将前药转化为活性药物。在一些实施方案中,前药对被治疗的患者的毒性将比活性药物对该患者低至少一个数量级。在一些实施方案中,活性药物的毒性要多几个数量级,例如,2、3、4、5或更多个数量级。适合的前药包括细胞毒性癌症化学治疗药物的前药,该细胞毒性癌症化学治疗药物包括,但不限于,氮芥、甲氨蝶呤、多柔比星、核苷、ams、靶向的细胞毒性癌症疗法,诸如厄洛替尼、达沙替尼、tk抑制剂和其它化合物,诸如在通过提述并入本文的wo88/07378、wo89/10140、wo90/02729、wo91/03460、ep-a-540263、wo94/02450、wo95/02420或wo95/03830中所述的那些。此外,前药形式可以为以下而制备:影响细胞功能(例如,hdac和甲基化的调节剂)以上调蛋白质和抗原呈递机制(例如,hla、β-2微球蛋白、tap、肽表位、蛋白酶体)或以允许免疫介导的药物以改善癌症细胞杀伤(例如,免疫检查点阻断药物、cart细胞或其它过继施用的细胞或抗体)的调节性药物,通过其它细胞毒性药物改善癌症细胞杀伤的hsp抑制剂,和使有抗性的癌细胞对癌症治疗(癌细胞对该癌症治疗有抗性)敏感的mdr抑制剂。在一些实施方案中,当前药转化酶是cpg2时,前药是cpg2不稳定的谷氨酸盐前药。在一些实施方案中,当前药转化酶是β-内酰胺酶时,前药是β-内酰胺酶不稳定的β-内酰胺前药。在某些实施方案中,当前药转化酶在细胞内表达(例如,“杀灭开关”前药转化酶)时,该前药能够被细胞吸收(例如,是能透过膜的)。因此,在一些实施方案中,前药在细胞内被转化为细胞毒性药物。
示例性前药包括式i的化合物或其两性离子、药学上可接受的盐和/或溶剂化物,
[1]
其中x1是o或nh;x2是n或ch;x3是oh或h;w1是h或nh-r3;r1、r2和r3中的两个是h且其余的r1、r2和r3是
x4和x5各自独立地为o或nh;x6是o、nh、ch2或c(me)2;r4、r5、r6、r7和r8各自独立地为h、-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or13a、-(ch2)2-c(o)or13b、
在本技术中有用的示例性前药还包括式ii的化合物或其两性离子、药学上可接受的盐和/或溶剂化物,
其中x7和x8各自独立地为cl、br、i、o-s(o)2ch3或o-s(o)2-甲苯基;x9是–c(o)-o、-c(o)-nh、o或nh;r9是键、亚烷基、亚芳基、亚芳烷基、杂亚芳基或杂亚芳烷基;r15是
x10和x11各自独立地为o或nh;x6是o、nh、ch2或c(me)2;x12是o、nh、ch2或c(me)2;r16、r17、r18、r19和r20各自独立地为h、-(ch2)3-nh(nh)-nh2、-(ch2)3-nh2、-(ch2)4-nh2、-ch2-c(o)or25a、-(ch2)2-c(o)or25b、
式ii的前药化合物可以是式iia-iie中的任一个的化合物或其两性离子、药学上可接受的盐和/或溶剂化物,
其中x13和x18各自独立地为o或nh;x14、x15、x16、x17、x19、x20、x21、x22、x23和x24各自独立地为cl、br、i、o-s(o)2ch3或o-s(o)2-甲苯基;n和m各自独立地为0、1、2、3、4或5;r26、r27、r28、r29和r30各自独立地为
式ii的前药化合物可以是式iif或式iig的化合物或其两性离子、药学上可接受的盐和/或溶剂化物,
其中;x28、x29、x30和x31各自独立地为cl、br、i、o-s(o)2ch3或o-s(o)2-甲苯基;r41和r42中的一个是h且r43和r44中的一个是h且其余的r41和r42(对于式iif)以及r43和r44(对于式iig)各自独立地为
本技术中有用的示例性前药进一步包括厄洛替尼的前药,诸如式iii的化合物或其两性离子、药学上可接受的盐和/或溶剂化物,
其中r55是
本文所述的化合物的药学上可接受的盐包括保留期望的药理活性并且是在生物学上非不希望(例如,该盐没有过度毒性、变应性或刺激性,且是可生物利用的)的酸或碱加成盐。当时本技术的化合物具有碱性基团(诸如,例如,氨基)时,可以使用无机酸(诸如盐酸、氢硼酸(hydroboricacid)、硝酸、硫酸和磷酸)、有机酸(例如,海藻酸、甲酸、乙酸、苯甲酸、葡萄糖酸、富马酸、草酸、酒石酸、乳酸、马来酸、柠檬酸、琥珀酸、苹果酸、甲磺酸、苯磺酸、萘磺酸和对甲苯磺酸)或酸性氨基酸(诸如天冬氨酸和谷氨酸)形成药学上可接受的盐。当本技术的化合物具有酸性基团(诸如例如,羧酸基团)时,其可与金属一起形成盐,诸如碱金属和碱土金属(例如,na+、li+、k+、ca2+、mg2+、zn2+)、氨或有机胺(例如,二环己胺、三甲胺、三乙胺、吡啶、甲基吡啶、乙醇胺、二乙醇胺、三乙醇胺)或碱性氨基酸(例如,精氨酸、赖氨酸和鸟氨酸)。此类盐可以在化合物的分离和纯化期间原位制备,或者通过使纯化的化合物以其游离碱或游离酸形式分别与适合的酸或碱分开反应,并分离由此形成的盐来制备。
本领域技术人员将理解,本技术的化合物可表现出互变异构、构象异构、几何异构和/或立体异构现象。由于说明书和权利要求书中的式图仅可代表可能的互变异构、构象异构、立体化学或几何异构形式中的一种,因此应理解,本技术涵盖具有本文所述的一个或多个公用程式(utilities)的化合物的任何互变异构、构象异构、立体化学和/或几何异构形式以及这些多种不同形式的混合物。
“互变异构体”是指彼此平衡的化合物的异构形式。异构形式的存在和浓度将取决于发现该化合物的环境,并且可能根据,例如,该化合物是否是固体或者在有机溶液或水溶液中而不同。例如,在水溶液中,喹唑酮可表现出下列异构形式,其被称为彼此的互变异构体:
作为另一实例,胍在质子有机溶液中可表现出下列异构形式,也被称为彼此的互变异构体:
由于通过结构式表示化合物的限制,应理解本文所述化合物的所有化学式表示化合物的所有互变异构形式,并且在本技术的范围内。
化合物的立体异构体(也称为光学异构体)包括结构的所有手性、非对映异构和外消旋形式,除非明确指出具体的立体化学。因此,如从描述中显而易见的,用于本技术的化合物包括在任何或所有不对称原子处的富集或拆分的光学异构体。外消旋和非对映异构混合物二者以及单独的光学异构体都可以被分离或合成,以便基本不含它们的对映异构或非对映异构伴侣,并且这些立体异构体均在本技术的范围内。
本技术的化合物可作为溶剂化物,特别是水合物而存在。水合物可在制造化合物或包含化合物的组合物期间形成,或水合物由于化合物的吸湿性性质而随时间形成。本技术的化合物也可作为有机溶剂化物而存在,包括dmf、醚和醇溶剂化物等。任何特定溶剂化物的鉴定和制备均在合成有机或药物化学的普通技术人员的技能范围内。
可基于表达的前药转化酶的选择来修饰本文所教导的前药化合物。可与本技术组合使用的其它前药系统的非限制性实例在以下领域中描述:羧肽酶a(例如,利用苯基丙氨酸掩蔽的前药,其例如,vitols等cancerres.55(3):478–81(1995)和deckert等intjoncol.24(5):1289–95(2004)中所述的);葡糖苷酶(例如,利用糖苷掩蔽的前药,其例如,在tietze等jmedchem.52(2):537–43(2009)和tietze等chemeurj.17(6):1922–9(2001)中描述);β-内酰胺酶(例如,利用头孢菌素掩蔽的前药,其例如,在kerr等bioconjugatechem.9(2):255–9(1998)和zhou等oncotargetsther.7:535–41(2014)中描述);和硝基还原酶(例如,利用硝基→胺掩蔽的前药,其例如,在biochemj.471(2):131–53(2015)中描述)。
本文所教导的前药化合物可用于需要前药化合物的任何方法或用途中。例如,采用可将本文提供的前药切割成活性药物的酶的任何临床应用都可以使用提供的前药化合物。此类方法和用途不受限于本文所述的方法和用途。例如,本文所教导的前药化合物不限于与本文提供的工程化免疫细胞(例如,seaker细胞)和相关技术一起使用。在一些实施方案中,本文提供的前药化合物可在细胞内(例如,适合的前药转化酶被转导至细胞中或通过细胞被表达)被任何适合的前药转化酶转化为活性药物。在一些实施方案中,本文提供的前药化合物可在细胞外被任何适合的前药转化酶转化为活性药物。在一些实施方案中,通过与配体缀合的任何适合的前药转化酶将前药化合物转化为活性药物。在一些实施方案中,配体是抗体。在一些实施方案中,前药化合物与定向酶前药疗法(dept)组合使用。在一些实施方案中,定向酶前药疗法是抗体定向的酶前药疗法(adept),基因定向的酶前药疗法(gdept)、病毒定向的酶前药疗法(vdept)、凝集素定向的酶激活的前药疗法(leapt)、聚合物定向的酶前药疗法(pdept)、梭菌定向的酶前药疗法(cdept)或其任何组合。
靶向配体和靶抗原
在一些实施方案中,本文提供的工程化的免疫细胞表达t细胞受体(tcr)或结合靶抗原(诸如肿瘤抗原)的其它细胞表面配体。细胞-表面配体可以是将免疫细胞定向至靶位点(例如,肿瘤位点)的任何分子。示例性细胞表面配体包括,例如内源受体、工程化受体或实现将免疫细胞靶向至靶位点的其它特异性配体。在一些实施方案中,受体是t细胞受体。在一些实施方案中,t细胞受体是结合靶抗原的野生型的或天然的t细胞受体。在一些实施方案中,受体(例如t细胞受体)是非天然受体(例如,对于免疫细胞而言不是内源的)。在一些实施方案中,受体是结合靶抗原的嵌合抗原受体(car),例如,t细胞car。
在一些实施方案中,靶抗原是由肿瘤细胞表达的。在一些实施方案中,靶抗原在肿瘤细胞的表面上表达。在一些实施方案中,靶抗原是细胞表面受体。在一些实施方案中,靶抗原是细胞表面糖蛋白。在一些实施方案中,靶抗原是由肿瘤细胞分泌的。在一些实施方案中,靶抗原定位于肿瘤微环境。在一些实施方案中,靶抗原定位于肿瘤微环境的细胞外基质(matrix)或基质(stroma)。在一些实施方案中,靶抗原是由位于肿瘤微环境的细胞外基质(matrix)或基质(stroma)内的一个或多个细胞表达的。
在一些实施方案中,靶抗原是肿瘤抗原,该肿瘤抗原选自5t4、α5β1-整合素、707-ap、a33、afp、art-4、b7h4、bage、bcl-2、β-连环蛋白、bcr-abl、mn/cix抗体、ca125、ca19-9、camel、cap-1、casp-8、cd4、cd5、cd19、cd20、cd21、cd22、cd25、cdc27/m、cd33、cd37、cd45、cd52、cd56、cd80、cd123、cdk4/m、cea、c-met、cs-1、ct、cyp-b、cyclinb1、dage、dam、ebna、egfr、erbb3、elf2m、emmprin、epcam、ephrinb2、雌激素受体、etv6-aml1、fap、铁蛋白、叶酸结合蛋白、gage、g250、gd-2、gm2、gnt-v、gp75、gp100(pmel17)、hage、her-2/neu、hla-a*0201-r170i、hpve6、hpve7、ki-67、hsp70-2m、hst-2、htert(或htrt)、ice、igf-1r、il-2r、il-5、kiaa0205、lage、ldlr/fut、lrp、mage、mart、mart-1/黑色素a(melan-a)、mart-2/ski、mc1r、间皮素、muc、mum-1–b、myc、mum-2、mum-3、na88-a、nyeso-1、ny-eso-b、p53、蛋白酶-3、p190小bcr-abl、pml/rarα、prame、孕酮受体、psa、psm、psma、ras、rage、ru1或ru2、rori、sart-1或sart-3、生存素、tel/aml1、tgfβ、tpi/m、trp-1、trp-2、trp-2/int2、肌腱蛋白、tsta酪氨酸酶、vegf和wt1。在某些实施方案中,靶抗原是选自cd19、wt1、prame的肿瘤抗原。
在不限制前述的情况下,可以通过靶向相关提供的抗原来治疗的示例性癌症包括:白血病/淋巴瘤(cd19、cd20、cd22、ror1、cd33);多发性骨髓瘤(b细胞成熟抗原(bcma));前列腺癌(psma、wt1、前列腺干细胞抗原(psca)、sv40t);乳腺癌(her2、erbb2);干细胞癌症(cd133);卵巢癌(l1–cam、muc16的细胞外域(muc-cd)、叶酸结合蛋白(叶酸受体)、lewisy);肾细胞癌(羧基-脱水酶-ix(caix);黑色素瘤(gd2);和胰腺癌(间皮素、cea、cd24)。
典型的治疗性抗癌mab(如结合cd19的mab)识别细胞表面蛋白,该细胞表面蛋白仅组成细胞蛋白成分的很少部分。大多数突变或致癌的肿瘤相关蛋白通常是核或细胞质的。在某些情况中,这些细胞内蛋白可在蛋白酶体中被降解、处理并通过mhci类分子呈递在细胞表面上,作为由t细胞受体(tcr)识别的t细胞表位。模拟tcr功能的mab(“tcr模拟物(tcrm)”或“类tcr”,即在细胞表面上mhc的背景下识别关键细胞内蛋白的肽抗原)的开发极大扩展了可通过有效mab定位的肿瘤靶标的潜在库。已经开发了tcrmfab或scfv,和对黑色素瘤ag特异的小鼠igg、ny-eso-1、htert、mart1、gp100和pr1等。此类抗体的抗原结合部分可被并入到本文提供的car中。hla-a2是美国和欧盟中最常见(约40%的人口)的hla单倍型。因此,在hla-a2的背景下提出的针对肿瘤抗原的有效tcrmmab和天然tcr可有用于治疗大量人群。
因此,在一些实施方案中,靶抗原是在mhc分子的背景下呈递的肿瘤抗原。在一些实施方案中,mhc蛋白是mhci类蛋白。在一些实施方案中,mhci类蛋白是hla-a、hla-b或hla-c分子。在一些实施方案中,靶抗原是在hla-a2分子的背景下呈递的肿瘤抗原。先前已经开发了针对在hla-a2分子的背景下呈递的细胞内wt1和prame抗原的mab。已经制备了igg1、岩藻糖基化fc形式、双特异性、bite和cart细胞形式,其在多种临床前动物模型中均表现出有力的治疗活性。可采用本文所述的此类抗体或其部分用于识别在mhc的背景下在靶细胞(例如,肿瘤细胞)的表面上呈递的靶抗原。嵌合抗原受体
在一些实施方案中,本文提供的工程化免疫细胞表达至少一种嵌合抗原受体(car)。car是工程化受体,其将感兴趣的特异性移植或赋予到免疫效应细胞上。例如,car可用于将单克隆抗体的特异性移植到免疫细胞(诸如t细胞)上。在一些实施方案中,car的编码序列的转移是通过核酸载体(诸如逆转录病毒载体)促进的。
目前存在三代car。在一些实施方案中,本文提供的工程化免疫细胞表达“第一代”car。“第一代”car典型地包含与跨膜域融合的细胞外抗原结合域(例如,单链可变片段(scfv)),该跨膜域与t细胞受体(tcr)链的细胞质域/细胞内域融合。“第一代”car典型地具有来自cd3ζ链的细胞内域,其是来自内源tcr的信号的初级发送介质。“第一代”car可提供从新抗原识别并导致在单个融合分子中通过它们的cd3ζ链信号传导域激活cd4+和cd8+t细胞二者,这不依赖于hla介导的抗原呈递。
在一些实施方案中,本文提供的工程化免疫细胞表达“第二代”car。“第二代”car将来自多种共刺激分子(例如,cd28、4-1bb、icos、ox40)的细胞内域添加至car的细胞质尾区,以向t细胞提供另外的信号。“第二代”car包含提供共刺激(例如,cd28或4-ibb)和激活(例如,cd3ζ)的分子。临床前研究已经表明“第二代”car可改善t细胞的抗肿瘤活性。例如,在患有慢性成淋巴细胞白血病(cll)和急性成淋巴细胞白血病(all)的患者中靶向cd19分子的临床试验证明了“第二代”car修饰的t细胞的强大功效。
在一些实施方案中,本文提供的工程化免疫细胞表达“第三代”car。“第三代”car包含提供多种共刺激(例如,cd28和4-1bb)和激活(例如,cd3ζ)的分子。
根据当前公开的主题,本文提供的工程免疫细胞的car包含细胞外抗原结合域、跨膜域和细胞内域。
car的细胞外抗原结合域
在某些实施方案中,car的细胞外抗原结合域特异性结合肿瘤抗原。在某些实施方案中,细胞外抗原结合域衍生自结合肿瘤抗原的单克隆抗体(mab)。在一些实施方案中,细胞外抗原结合域包含scfv。在一些实施方案中,细胞外抗原结合域包含fab,fab是任选交联的。在一些实施方案中,细胞外结合域包含f(ab)2。在一些实施方案中,任何前述分子包含在具有异源序列的融合蛋白中以形成细胞外抗原结合域。在某些实施方案中,细胞外抗原结合域包含特异性结合肿瘤抗原的人scfv。在某些实施方案中,scfv是通过用肿瘤抗原-fc融合蛋白筛选scfv噬菌体文库来鉴定的。
在某些实施方案中,本公开的car的细胞外抗原结合域对肿瘤抗原(例如,哺乳动物肿瘤抗原,诸如人肿瘤抗原)具有高结合特异性和高结合亲和力。例如,在一些实施方案中,car的细胞外抗原结合域(体现在例如,人scfv或其类似物中)以约1x10-5m或更少的解离常数(kd)结合特定肿瘤抗原。在某些实施方案中,kd为约5x10-6m或更少、约1x10-6m或更少、约5x10-7m或更少、约1x10-7m或更少、约5x10-8m或更少、约1x10-8m或更少、约5x10-9或更少、约4x10-9或更少、约3x10-9或更少、约2x10-9或更少或约1x10-9m或更少。在某些非限制性实施方案中,kd为约3x10-9m或更少。在某些非限制性实施方案中,kd为约3x10-9至约2x10-7。
本公开的靶向肿瘤抗原的car的细胞外抗原结合域(体现在例如,人scfv或其类似物中)的结合可通过,例如,酶联免疫吸附测定(elisa)、放射免疫测定(ria)、facs分析、生物测定(例如,生长抑制)或western印记测定来证实。这些测定法中的每一种通常通过采用对感兴趣的复合物具有特异性的标记试剂(例如,抗体或scfv)来检测特别感兴趣的蛋白质-抗体复合物的存在。例如,scfv可以是放射性标记的,且可用于放射免疫测定(ria)(参见,例如,weintraub,b.,principlesofradioimmunoassays,seventhtrainingcourseonradioligandassaytechniques,theendocrinesociety,march,1986,其通过提述并入本文)。放射性同位素可以通过使用γ计数器或闪烁计数器的这样方式或通过放射自显影来检测。在某些实施方案中,靶向肿瘤抗原的car的细胞外抗原结合域用荧光标记物来标记。荧光标记物的非限制性实例包括绿色荧光蛋白(gfp)、蓝色荧光蛋白(例如,ebfp、ebfp2、azurite和mkalamal),青色荧光蛋白(例如,ecfp、cerulean和cypet),和黄色荧光蛋白(例如,yfp、citrine、venus和ypet)。在某些实施方案中,本公开的靶向肿瘤抗原的car的人scfv是用gfp标记的。
在一些实施方案中,表达的car的细胞外抗原结合域结合有肿瘤细胞表达的肿瘤抗原。在一些实施方案中,表达的car的细胞外抗原结合域结合在肿瘤细胞的表面上表达的肿瘤抗原。在一些实施方案中,表达的car的细胞外抗原结合域结合在肿瘤细胞的表面上与mhc蛋白组合表达的肿瘤抗原。在一些实施方案中,mhc蛋白是mhci类蛋白。在一些实施方案中,mhci类蛋白是hla-a、hla-b或hla-c分子。在一些实施方案中,表达的car的细胞外抗原结合域结合不与mhc蛋白组合的在肿瘤细胞的表面上表达的肿瘤抗原。
在一些实施方案中,表达的car的细胞外抗原结合域结合肿瘤抗原,该肿瘤抗原选自5t4、α5β1-整合素、707-ap、a33、afp、art-4、b7h4、bage、bcl-2、β-连环蛋白、bcr-abl、mn/cix抗体、ca125、ca19-9、camel、cap-1、casp-8、cd4、cd5、cd19、cd20、cd21、cd22、cd25、cdc27/m、cd33、cd37、cd45、cd52、cd56、cd80、cd123、cdk4/m、cea、c-met、cs-1、ct、cyp-b、细胞周期蛋白b1、dage、dam、ebna、egfr、erbb3、elf2m、emmprin、epcam、ephrinb2、雌激素受体、etv6-aml1、fap、铁蛋白、叶酸结合蛋白、gage、g250、gd-2、gm2、gnt-v、gp75、gp100(pmel17)、hage、her-2/neu、hla-a*0201-r170i、hpve6、hpve7、ki-67、hsp70-2m、hst-2、htert(或htrt)、ice、igf-1r、il-2r、il-5、kiaa0205、lage、ldlr/fut、lrp、mage、mart、mart-1/黑色素a、mart-2/ski、mc1r、间皮素、muc、mum-1–b、myc、mum-2、mum-3、na88-a、nyeso-1、ny-eso-b、p53、蛋白酶-3、p190小bcr-abl、pml/rarα,prame、孕酮受体、psa、psm、psma、ras、rage、ru1或ru2、rori、sart-1或sart-3、存活素、tel/aml1、tgfβ、tpi/m、trp-1、trp-2、trp-2/int2、肌腱蛋白、tsta酪氨酸酶、vegf和wt1。在某些实施方案中,表达的car的细胞外抗原结合域结合肿瘤抗原,该肿瘤抗原选自cd19、wt1、prame。示例性细胞外抗原结合域以及生成此类域和相关car的方法描述于例如,wo2016/191246、wo2017/023859、wo2015/188141、wo2015/070061、wo2012/135854、wo2014/055668中,其(包括其中提供的序列表)通过提述以其整体并入。
在一些实施方案中,表达的car的细胞外抗原结合域结合cd19肿瘤抗原。在一些实施方案中,表达的car的细胞外抗原结合域结合在mhc分子的背景下呈递的cd19肿瘤抗原。在一些实施方案中,结合在hla-a2分子的背景下呈递的cd19肿瘤抗原。
在一些实施方案中,表达的car的细胞外抗原结合域结合“黑色素瘤中优先表达的抗原”(prame)肿瘤抗原。在一些实施方案中,表达的car的细胞外抗原结合域结合在mhc分子的背景下呈递的prame肿瘤抗原。在一些实施方案中,结合在hla-a2分子的背景下呈递的prame肿瘤抗原。
在一些实施方案中,表达的car的细胞外抗原结合域结合wt1(wilm’s肿瘤蛋白1)肿瘤抗原。在一些实施方案中,表达的car的细胞外抗原结合结构域与在mhc分子的背景下呈递的wt1肿瘤抗原结合。在一些实施方案中,表达的car的细胞外抗原结合域结合在hla-a2分子的背景下呈递的wt1肿瘤抗原。
在某些实施方案中,细胞外抗原结合域(例如,人scfv)包含重链可变区和轻链可变区,其任选地使用在重链可变区和轻链可变区之间的接头序列(例如接头肽(例如,seqno:21))连接。在某些实施方案中,细胞外抗原结合域是具有vh和vl区的人scfv-fc融合蛋白或全长人igg。
在某些实施方案中,细胞外抗原结合域包含结合cd19抗原的人scfv。在一些实施方案中,scfv包含具有seqidno:19的氨基酸序列的多肽。
malpvtalllplalllhaevklqqsgaelvrpgssvkisckasgyafssywmnwvkqrpgqglewigqiypgdgdtnyngkfkgqatltadkssstaymqlsgltsedsavyfcarktissvvdfyfdywgqgttvtvssggggsggggsggggsdieltqspkfmstsvgdrvsvtckasqnvgtnvawyqqkpgqspkpliysatyrnsgvpdrftgsgsgtdftltitnvqskdladyfcqqynrypytsgggtkleikr(seqidno:19)
在一些实施方案中,scfv包含具有与seqidno:19至少80%、至少85%、至少90%或至少95%同一的氨基酸序列的多肽。例如,scfv包含具有与seqidno:19约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一的氨基酸序列的多肽。
在一些实施方案中,scfv是由具有seqidno:20的核酸序列的核酸编码的。
atggctctcccagtgactgccctactgcttcccctagcgcttctcctgcatgcagaggtgaagctgcagcagtctggggctgagctggtgaggcctgggtcctcagtgaagatttcctgcaaggcttctggctatgcattcagtagctactggatgaactgggtgaagcagaggcctggacagggtcttgagtggattggacagatttatcctggagatggtgatactaactacaatggaaagttcaagggtcaagccacactgactgcagacaaatcctccagcacagcctacatgcagctcagcggcctaacatctgaggactctgcggtctatttctgtgcaagaaagaccattagttcggtagtagatttctactttgactactggggccaagggaccacggtcaccgtctcctcaggtggaggtggatcaggtggaggtggatctggtggaggtggatctgacattgagctcacccagtctccaaaattcatgtccacatcagtaggagacagggtcagcgtcacctgcaaggccagtcagaatgtgggtactaatgtagcctggtatcaacagaaaccaggacaatctcctaaaccactgatttactcggcaacctaccggaacagtggagtccctgatcgcttcacaggcagtggatctgggacagatttcactctcaccatcactaacgtgcagtctaaagacttggcagactatttctgtcaacaatataacaggtatccgtacacgtccggaggggggaccaagctggagatcaaacgg(seqidno:20)
在一些实施方案中,scfv是由具有与seqidno:20至少80%、至少85%、至少90%或至少95%同一的核酸序列的核酸编码的。在一些实施方案中,scfv是由具有seqidno:20的核酸序列的核酸编码的。在一些实施方案中,scfv是由具有与seqidno:20约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一的核酸序列的核酸编码的。
在某些非限制性实施方案中,本公开的car的细胞外抗原结合域可包含连接细胞外抗原结合域的重链可变区和轻链可变区的接头。如本文所用的,术语“接头”是指共价连接两个或更多个多肽或核酸以使它们彼此连接的官能团(例如,化学品或多肽)。如本文所用的,“肽接头”是指用于将两种蛋白质偶联在一起(例如,使vh和vl域偶联)的一个或多个氨基酸。在某些实施方案中,接头包含具有seqidno:21中所示的序列的氨基酸。在某些实施方案中,编码seqidno:21的氨基酸序列的核苷酸序列示于seqidno:22中。
此外,细胞外抗原结合域可包含指导新生蛋白进入内质网的前导序列或信号肽。如果要对car进行糖基化并将其锚定在细胞膜中,信号肽或前导序列能够是至关重要的。信号序列或前导序列可以是存在于新合成的蛋白质的n端处的肽序列(约5、约10、约15、约20、约25或约30个氨基酸长),其指导新合成的蛋白质进入分泌途径。在某些实施方案中,信号肽与细胞外抗原结合域的n端共价连接。在某些实施方案中,信号肽包含cd8信号多肽,该cd8信号多肽包含具有如下提供的seqidno:11中所示的序列的氨基酸。
malpvtalllplalllhaarp(seqidno:11)
编码seqidno:11的氨基酸序列的核苷酸序列示于以下提供的seqidno:12中:
atggccctgccagtaacggctctgctgctgccacttgctctgctcctccatgcagccaggcct(seqidno:12)
car的跨膜域
在某些非限制性实施方案中,car的跨膜域包含跨越膜的至少一部分的疏水性α螺旋。不同跨膜域导致不同受体稳定性。在抗原识别后,受体聚集并且信号被传递到细胞。依照本公开的主题,car的跨膜域可包含cd8多肽、cd28多肽、cd3ζ多肽、cd4多肽、4-ibb多肽、ox40多肽、icos多肽、ctla-4多肽、pd-1多肽、lag-3多肽、2b4多肽、btla多肽、合成肽(例如,不基于与免疫应答相关联的蛋白质的跨膜肽),或其组合。
在某些实施方案中,本公开的car的跨膜域包含cd28多肽。cd28多肽可具有与具有ncbi参考号:pi0747或np006130(seqidno:24)的序列或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或100%同源的氨基酸序列,和/或可任选地包含最高达一个或最高达两个或最高达三个保守氨基酸取代。在某些实施方案中,cd28多肽可具有作为seqidno:24的连续部分的氨基酸序列,seqidno:24的连续部分的长度为至少20、或至少30、或至少40、或至少50、且多达220个氨基酸。可替换地或此外,在非限制性的多个实施方案中,cd28多肽具有seqidno:24的氨基酸1至220、1至50、50至100、100至150、114至220、150至200,或200至220的氨基酸序列。在某些实施方案中,本公开的car包含包含cd28多肽的跨膜域,以及包含共刺激信号传导区的细胞内域,该共刺激信号传导区包含cd28多肽。在某些实施方案中,跨膜域和细胞内域中包含的cd28多肽具有seqidno:24的氨基酸114至220的氨基酸序列。
seqidno:24提供如下:
mlrlllalnlfpsiqvtgnkilvkqspmlvaydnalsckysynlfsrefraslhkgldsavevcwygnysqqlqvysktgfncdgklgnesvtfylqnlyqtdiyfckievmypppyldneksngtiihvkgkhlcpsplfpgpskpfwvlvwggvlacysllvtvafiifwvrskrsrllhsdymnmtprrpgptrkhyqpyapprdfaayrs(seqidno:24)
依照本公开的主题,“cd28核酸分子”是指编码cd28多肽的多核苷酸。在某些实施方案中,编码本公开的car的跨膜域和细胞内域(例如,共刺激信号传导区)中包含的cd28多肽(seqidno:24的氨基酸114至220)的cd28核酸分子包含具有如下提供的seqidno:25中所示的序列的核酸。
attgaagttatgtatcctcctccttacctagacaatgagaagagcaatggaaccattatccatgtgaaagggaaacacctttgtccaagtcccctatttcccggaccttctaagcccttttgggtgctggtggtggttggtggagtcctggcttgctatagcttgctagtaacagtggcctttattattttctgggtgaggagtaagaggagcaggctcctgcacagtgactacatgaacatgactccccgccgccccgggcccacccgcaagcattaccagccctatgccccaccacgcgacttcgcagcctatcgctcc(seqidno:25)
在某些实施方案中,跨膜域包含cd8多肽。cd8多肽可具有与如下提供的seqidno:26至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%)同源(本文中的同源性可通过使用标准软件诸如blast或fasta来测定)的氨基酸序列或其片段,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。在某些实施方案中,cd8多肽可具有作为seqidno:26的连续部分的氨基酸序列,seqidno:26的连续部分的长度为至少20、或至少30、或至少40、或至少50,且最高达235个氨基酸。可替换地或此外,在非限制性的多个实施方案中,cd8多肽具有seqidno:26的氨基酸1至235、1至50、50至100、100至150、150至200或200至235的氨基酸序列。
malpvtalllplalllhaarpsqfrvspldrtwnlgetvelkcqvllsnptsgcswlfqprgaaasptfllylsqnkpkaaegldtqrfsgkrlgdtfvltlsdfrrenegyyfcsalsnsimyfshfvpvflpakptttpaprpptpaptiasqplslrpeacrpaaggavhtrgldfacdiyiwaplagtcgvlllslvitlycnhrnrrrvckcprpwksgdkpslsaryv(seqidno:26)
依照本公开的主题,“cd8核酸分子”是指编码cd8多肽的多核苷酸。
在某些非限制性实施方案中,car还可包含使细胞外抗原结合域与跨膜域连接的间隔区。间隔区可以足够柔性以允许抗原结合结构域在不同方向上定向以促进抗原识别的同时保留car的激活活性。在某些非限制性实施方案中,间隔区可以是来自iggl的铰链区、免疫球蛋白的ch2ch3区和cd3的一部分、cd28多肽的一部分(例如,seqidno:24)、cd8多肽的一部分(例如,seqidno:26)、至少约80%、至少约85%、至少约90%或至少约95%与前述任一同源的前述任一变化、或合成间隔序列。在某些非限制性实施方案中,间隔区可具有约1-50(例如,5-25、10-30或30-50)个氨基酸的长度。
car的细胞内域
在某些非限制性实施方案中,car的细胞内域可包含cd3ζ多肽,其可激活或刺激细胞(例如,淋巴样谱系的细胞,例如,t细胞)。cd3ζ包含3个itam,并在结合抗原后向细胞(例如,淋巴样谱系的细胞,例如,t细胞)传递激活信号。cd3ζ多肽可具有与具有ncbi参考号:np_932170(seqidno:48)的序列或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。在非限制性的某些实施方案中,cd3ζ多肽可具有作为seqidno:27的连续部分的氨基酸序列,seqidno:27的连续部分的长度为至少20、或至少30、或至少40、或至少50,且最高达164个氨基酸。可替换地或此外地,在非限制性的多个实施方案中,cd3ζ多肽具有seqidno:27的氨基酸1至164、1至50、50至100、100至150或150至164的氨基酸序列。在某些实施方案中,cd3ζ多肽具有seqidno:27的氨基酸52至164的氨基酸序列。
seqidno:27提供如下:
mkwkalftaailqaqlpiteaqsfglldpklcylldgilfiygviltalflrvkfsrsadapayqqgqnqlynelnlgrreeydvldkrrgrdpemggkpqrrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr(seqidno:27)
在某些实施方案中,cd3ζ多肽具有seqidno:28中所示的氨基酸序列,seqidno:28提供如下:
rvkfsrsaeppayqqgqnqlynelnlgrreeydvldkrrgrdpemggkprrknpqeglynelqkdkmaeayseigmkgerrrgkghdglyqglstatkdtydalhmqalppr(seqidno:28)
依照本公开的主题,“cd3ζ核酸分子”是指编码cd3ζ多肽的多核苷酸。在某些实施方案中,编码本公开的car的细胞内域中包含的cd3ζ多肽(seqidno:28)的cd3ζ核酸分子包含如下提供的seqidno:29中所示的核苷酸序列。
agagtgaagttcagcaggagcgcagagccccccgcgtaccagcagggccagaaccagctctataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggccgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaatgaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgccggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacctacgacgcccttcacatgcaggccctgccccctcgcg(seqidno:29)
在某些非限制性实施方案中,car的细胞内域进一步包含至少一种信号传导区。该至少一种信号传导区可包括cd28多肽、4-ibb多肽、ox40多肽、icos多肽、dap-10多肽、pd-1多肽、ctla-4多肽、lag-3多肽、2b4多肽、btla多肽、合成肽(不基于与免疫应答相关联的蛋白质),或其组合。
在某些实施方案中,信号传导区是共刺激信号传导区。
在某些实施方案中,共刺激信号传导区包含至少一种共刺激分子,其可提供最佳的淋巴细胞激活。如本文所用的,“共刺激分子”是指淋巴细胞对抗原的有效应答所需的除抗原受体或其配体外的细胞表面分子。该至少一种共刺激信号传导区可包括cd28多肽、4-ibb多肽、ox40多肽、icos多肽、dap-10多肽,或其组合。共刺激分子可结合共刺激配体,其是在细胞表面上表达的蛋白质,该蛋白质一旦与其受体结合产生共刺激应答,即,影响当抗原结合其car分子时提供的影响该刺激的细胞内应答。共刺激配体,包括,但不限于cd80、cd86、cd70、ox40l、4-1bbl、cd48、tnfrsf14和pd-ll。作为一个实例,4-1bb配体(即,4-1bbl)可结合4-1bb(也称作“cd137”)以提供细胞内信号,该细胞内信号与car信号组合诱导car+t细胞的效应细胞功能。包含包含共刺激信号传导区的细胞内域的car在美国7,446,190(其通过提述以其整体并入本文)中公开,该共刺激信号传导区包含4-1bb、icos或dap-10。在某些实施方案中,car的细胞内域包含共刺激信号传导区,该共刺激信号传导区包含cd28多肽。在某些实施方案中,car的细胞内域包含共刺激信号传导区,该共刺激信号传导区包含两种共刺激分子:cd28和4-1bb或cd28和ox40。
4-ibb可充当肿瘤坏死因子(tnf)配体并具有刺激活性。4-ibb多肽可具有与具有ncbi参考号:p41273或np_001552(seqidno:30)的序列或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:30提供如下:
mgnscynivatlllvlnfertrslqdpcsncpagtfcdnnrnqicspcppnsfssaggqrtcdicrqckgvfrtrkecsstsnaecdctpgfhclgagcsmceqdckqgqeltkkgckdccfgtfndqkrgicrpwtncsldgksvlgtkerdwcgpspadlspgassvtppaparepghspqiisfflaltstallfllffltlrfswkrgrkkllyifkqpfmrpvqttqeedgcscrfpeeeeggcel(seqidno:30)
依照本公开的主题,“4-ibb核酸分子”是指编码4-ibb多肽的多核苷酸。
ox40多肽可具有与具有ncbi参考号:p43489或np003318(seqidno:31)的序列或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:31提供如下:
mcvgarrlgrgpcaallllglglstvtglhcvgdtypsndrcchecrpgngmvsrcsrsqntvcrpcgpgfyndwsskpckpctwcnlrsgserkqlctatqdtvcrcragtqpldsykpgvdcapcppghfspgdnqackpwtnctlagkhtlqpasnssdaicedrdppatqpqetqgpparpitvqpteawprtsqgpstrpvevpggravaailglglvlgllgplaillalyllrrdqrlppdahkppgggsfrtpiqeeqadahstlaki(seqidno:31)
依照本公开的主题,“ox40核酸分子”是指编码ox40多肽的多核苷酸。
icos多肽可具有与具有ncbi参考号:np_036224(seqidno:32)的序列或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:32提供如下:
mksglwyfflfclrikvltgeingsanyemfifhnggvqilckypdivqqfkmqllkggqilcdltktkgsgntvsikslkfchsqlsnnsvsfflynldhshanyyfcnlsifdpppfkvtltggylhiyesqlccqlkfwlpigcaafvwcilgcilicwltkkkysssvhdpngeymfmratakksrltdvtl(seqidno:32)
依照本公开的主题,“icos核酸分子”是指编码icos多肽的多核苷酸。
ctla-4是由激活的t细胞表达的抑制性受体,当通过其相应配体(分别为cd80和cd86;b7-1和b7-2)啮合ctla-4时,ctla-4介导激活的t细胞抑制或无能。在临床前和临床研究中,尽管通过全身性抗体输注的ctla-4阻断会增强内源性抗肿瘤应答,但在临床环境中具有显著的不可预见的毒性。
ctla-4含有细胞外v域、跨膜域和胞质尾区。已经表征了编码不同同种型的替代剪接变体。膜结合的同种型作为通过二硫键互连的同型二聚体起作用,而可溶性同种型作为单体起作用。细胞内域与cd28的细胞内域相似之处在于,其不具有固有的催化活性且含有一个能够结合pi3k、pp2a和shp-2的yvkm基序以及一个能够结合含有sh3的蛋白质的富含脯氨酸的基序。ctla-4在抑制t细胞应答中的一个作用似乎直接是经由tcr近端信号传导蛋白(诸如cd3和lat)的shp-2和pp2a脱磷酸化。ctla-4也可经由与cd28竞争cd80/86结合而间接影响信号传导。还显示出ctla-4与pi3k、cd80、ap2m1和ppp2r5a结合和/或相互作用。
依照本公开的主题,ctla-4多肽可具有与uniprotkb/swiss-prot参考号:p16410.3(seqidno:33)或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%同源的氨基酸序列(本文中的同源性可使用标准软件诸如blast或fasta测定),和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:33提供如下:
maclgfqrhkaqlnlatrtwpctllffllfipvfckamhvaqpawlassrgiasfvceyaspgkatevrvtvlrqadsqvtevcaatymmgneltflddsictgtssgnqltiqglramdtglyickvelmypppyylgigngtqiyvidpepcpdsdfllwilaavssglffysflltavslskmlkkrsplttgvyvkmpptepecekqfqpyfipin(seqidno:33)
依照本公开的主题,“ctla-4核酸分子”是指编码ctla-4多肽的多核苷酸。
pd-1是与其相应的在内源巨噬细胞和树突细胞上表达的配体pd-l1和pd-l2啮合后,激活的t细胞的负免疫调节物。pd-1是268个氨基酸的i型膜蛋白。pd-1具有两个配体,pd-l1和pd-l2,其是b7家族的成员。蛋白质的结构包含细胞外igv域,然后是跨膜区和细胞内尾。细胞内尾含有位于基于免疫受体酪氨酸的抑制性基序和基于免疫受体酪氨酸的开关基序中的两个磷酸化位点,pd-1负调节tcr信号。在配体结合后,shp-i和shp-2磷酸酶结合pd-1的胞质尾区。pd-l1的上调是肿瘤细胞可逃避宿主免疫系统的一种机制。在临床前和临床试验中,通过拮抗性抗体的pd-1阻断会诱导通过宿主内源性免疫系统介导的抗肿瘤应答。依照本公开的主题,pd-1多肽可具有与ncbi参考号:np_005009.2(seqidno:34)或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:34提供如下:
mqipqapwpvvwavlqlgwrpgwfldspdrpwnpptfspallwtegdnatftcsfsntsesfvlnwyrmspsnqtdklaafpedrsqpgqdcrfrvtqlpngrdfhmsvvrarrndsgtylcgaislapkaqikeslraelrvterraevptahpspsprpagqfqtlvvgwggllgslvllvwvlavicsraargtigarrtgqplkedpsavpvfsvdygeldfqwrektpeppvpcvpeqteyativfpsgmgtssparrgsadgprsaqplrpedghcswpl(seqidno:34)
依照本公开的主题,“pd-1核酸分子”是指编码pd-1多肽的多核苷酸。
淋巴细胞-激活蛋白3(lag-3)是免疫细胞的负免疫调节物。lag-3属于免疫球蛋白(ig)超家族,且含有4个细胞外ig样结构域。lag3基因含有8个外显子。序列数据、外显子/内含子组织、和染色体定位均表明lag3与cd4的密切关系。lag3也被指定为cd223(分化簇223)。
依照本公开的主题,lag-3多肽可具有与uniprotkb/swiss-prot参考号:p18627.5(seqidno:35)或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:35提供如下:
mweaqflgllflqplwvapvkplqpgaevpwwaqegapaqlpcsptiplqdlsllrragvtwqhqpdsgppaaapghplapgphpaapsswgprprrytvlsvgpgglrsgrlplqprvqldergrqrgdfslwlrparradageyraavhlrdralscrlrlrlgqasmtasppgslrasdwvilncsfsrpdrpasvhwfrnrgqgrvpvresphhhlaesflflpqvspmdsgpwgciltyrdgfnvsimynltvlglepptpltvyagagsrvglpcrlpagvgtrsfltakwtppgggpdllvtgdngdftlrledvsqaqagtytchihlqeqqlnatvtlaiitvtpksfgspgslgkllcevtpvsgqerfvwssldtpsqrsfsgpwleaqeaqllsqpwqcqlyqgerllgaavyftelsspgaqrsgrapgalpaghlllflilgvlsllllvtgafgfhlwrrqwrprrfsaleqgihppqaqskieeleqepepepepepepepepepeql(seqidno:35)
依照本公开的主题,“lag-3核酸分子”是指编码lag-3多肽的多核苷酸。自然杀伤细胞受体2b4(2b4)介导对nk细胞和t细胞亚组的非mhc限制的细胞杀伤。迄今为止,2b4的功能仍在研究中,2b4-s同种型被认为是激活受体,且2b4-l同种型被认为是免疫细胞的负免疫调节物。2b4通过结合其高亲和力配体cd48而啮合。2b4含有基于酪氨酸的开关基序,这是一种分子开关,允许蛋白质与多种磷酸酶相关联。2b4还被指定为cd244(分化簇244)。
依照本公开的主题,2b4多肽可具有与uniprotkb/swiss-prot参考号:q9bzw8.2(seqidno:36)或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%同源的氨基酸序列,和/或可任选地包含最高达一个、或最高达两个、或最高达三个保守氨基酸取代。
seqidno:36提供如下:
mlgqwtlilllllkvyqgkgcqgsadhwsisgvplqlqpnsiqtkvdsiawkkllpsqngfhhilkwengslpsntsndrfsfivknlsllikaaqqqdsglyclevtsisgkvqtatfqvfvfesllpdkvekprlqgqgkildrgrcqvalsclvsrdgnvsyawyrgskliqtagnltyldeevdingthtytcnvsnpvsweshtlnltqdcqnahqefrfwpflviivilsalflgtlacfcvwrrkrkekqsetspkefltiyedvkdlktrrnheqeqtfpgggstiysmiqsqssaptsqepaytlysliqpsrksgsrkrnhspsfnstiyevigksqpkaqnparlsrkelenfdvys(seqidno:36)
依照本公开的主题,“2b4核酸分子”是指编码2b4多肽的多核苷酸。
在t细胞激活期间,b-和t-淋巴细胞弱化子(btla)表达被诱导,且btla保持在thl细胞而非th2细胞上表达。像pd1和ctla4一样,btla与b7同源物b7h4相互作用。然而,不同于pd-1和ctla-4,btla显示出经由与肿瘤坏死家族受体(tnf-r)(而不仅仅是细胞表面受体的b7家族)相互作用的t细胞抑制。btla是肿瘤坏死因子(受体)超家族成员14(tnfrsf14)(也被称为疱疹病毒进入介体(hvem))的配体。btla-hvem复合物负调节t细胞免疫应答。已经显示btla激活会抑制人cd8+癌症特异性t细胞的功能。btla还被指定为cd272(分化簇272)。
依照本公开的主题,btla多肽可具有与uniprotkb/swiss-prot参考号:q7z6a9.3(seqidno:37)或其片段至少约85%、约90%、约95%、约96%、约97%、约98%、约99%或约100%同源的氨基酸序列,和/或可任选地包含最多达一个、或最多达两个、或最多达三个保守氨基酸取代。
seqidno:37提供如下:
mktlpamlgtgklfwvfflipyldiwnihgkescdvqlyikrqsehsilagdpfelecpvkycanrphvtwcklngttcvkledrqtswkeeknisffilhfepvlpndngsyrcsanfqsnlieshsttlyvtdvksaserpskdemasrpwllyrllplgglpllittcfclfcclrrhqgkqnelsdtagreinlvdahlkseqteastrqnsqvllsetgiydndpdlcfrmqegsevysnpcleenkpgivyaslnhsvigpnsrlarnvkeapteyasicvrs(seqidno:37)
依照本公开的主题,“btla核酸分子”是指编码btla多肽的多核苷酸。
示例性car和前药转化酶构建体
在某些实施方案中,car和前药转化酶作为通过自切割的接头(诸如p2a接头)连接的单一多肽被表达。在某些实施方案中,car和前药转化酶作为两个分开的多肽被表达。
在某些实施方案中,car包含细胞外抗原结合区、跨膜域和细胞内域,该细胞外抗原结合区包含特异性结合人肿瘤抗原的人scfv,该跨膜域包含cd28多肽,该细胞内域包含cd3ζ多肽和包含4-1bb多肽的共刺激信号传导区,如图3中所示的。如图3中所示的,car还包含与细胞外抗原结合域的n端共价连接的信号肽或前导序列。信号肽包含具有seqidno:11中所示的序列的氨基酸。在某些实施方案中,人scfv选自抗cd19scfv和抗wt1scfv和抗pramescfv。
在一些实施方案中,编码car和前药转化酶(例如,cpg2或β-内酰胺酶)的核酸可操作地连接诱导型启动子。在一些实施方案中,编码car和前药转化酶(例如,cpg2或β-内酰胺酶)的核酸可操作地连接组成型启动子。在一些实施方案中,编码car的核酸和编码前药转化酶(例如,cpg2或β-内酰胺酶)的核酸与两个分开的启动子可操作地连接。在一些实施方案中,编码car的核酸可操作地连接组成型启动子,且编码前药转化酶(例如,cpg2orβ-内酰胺酶)的核酸可操作地连接组成型启动子。在一些实施方案中,编码car的核酸可操作地连接组成型启动子,且编码前药转化酶(例如,cpg2或β-内酰胺酶)的核酸可操作地连接诱导型启动子。
在一些实施方案中,诱导型启动子是合成性notch启动子,其在cart细胞中是可激活的,其中当car与肿瘤抗原的啮合诱导膜内蛋白水解时,car的细胞内域含有从膜释放的转录调节物(参见,例如morsut等,细胞164(4):780–791(2016))。因此,在工程化免疫细胞与肿瘤抗原结合后,诱导前药转化酶的转录。
本公开的主题还提供分离的核酸分子,其编码本文所述的car/前药转化酶构建体或其功能性部分。在某些实施方案中,分离的核酸分子编码抗cd19-靶向的car,该抗cd19-靶向的car包含特异性结合人cd19多肽的人scfv、包含cd8多肽的跨膜域,和包含cd3ζ多肽和共刺激信号传导区(包含4-1bb多肽)的细胞内域、p2a自切割肽以及与信号肽和包含cd8多肽的跨膜域融合的cpg2或β-内酰胺酶多肽(参见,例如,图3a和seqidno:48-51中所示的示例性构建体)。
在某些实施方案中,分离的核酸分子编码抗cd19-靶向的car,该抗cd19-靶向的car包含特异性结合人cd19多肽的人scfv、包含cd8多肽的跨膜域、和包含cd3ζ多肽和共刺激信号传导区(包含4-1bb多肽)的细胞内域、p2a自切割肽、以及与信号肽融合的cpg2或β-内酰胺酶多肽(参见,例如,图3b)。
在某些实施方案中,分离的核酸分子编码抗cd19-靶向的car,该抗cd19-靶向的car包含特异性结合人cd19多肽的人scfv,该人scfv与合成性notch跨膜域和细胞内可切割的转录因子融合(参见,例如,图3c)。在某些实施方案中,分离的核酸分子编码通过释放合成性notch系统的转录因子可诱导的cpg2或β-内酰胺酶。
在某些实施方案中,分离的核酸分子编码本公开的car构建体的功能性部分。如本文所用的,术语“功能性部分”是指car的任何部分(portion)、部分(part)或片段,该部分(portion)、部分(part)或片段保留靶向的car(亲本car)的生物活性。例如,功能性部分涵盖靶向肿瘤抗原的car的部分(portion)、部分(part)或片段,其保留与亲本car相似、相同或甚至更高程度的识别靶细胞以治疗疾病(例如,实体肿瘤)的能力。在某些实施方案中,编码靶向肿瘤抗原的car的功能性部分的分离的核酸分子可编码包含例如亲本car的约10%、约20%、约25%、约30%、约35%、约40%、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、约85%、约90%和约95%或更多的蛋白质。
免疫细胞
本公开的主题提供表达前药转化酶和t细胞受体(例如,car)或其它配体的工程化免疫细胞,该t细胞受体或其它配体包含细胞外抗原结合域、跨膜域和细胞内域,其中细胞外抗原结合域特异性结合肿瘤抗原,包括肿瘤受体或配体,如上所述的。在某些实施方案中,可用本公开的car/前药转化酶构建体转导免疫细胞,使得细胞表达car和前药转化酶。本公开的主题还提供使用此类细胞来治疗肿瘤的方法。本公开的主题的工程化免疫细胞可以是淋巴样谱系或髓样谱系的细胞。淋巴样谱系,其包含b、t和自然杀伤(nk)细胞,提供抗体的产生、细胞免疫系统的调节、血液中外源剂的检测、宿主外源细胞的检测等。淋巴样谱系的免疫细胞的非限制性实例包括t细胞、自然杀伤(nk)细胞、胚胎干细胞和多能干细胞(例如,可分化出淋巴样细胞的多能干细胞)。t细胞可以是在胸腺中成熟且主要负责细胞介导的免疫的淋巴细胞。t细胞参与适应性免疫系统。本公开的主题的t细胞可以是任何类型的t细胞,包括,但不限于,t辅助细胞、细胞毒性t细胞、记忆t细胞(包括中枢记忆t细胞、干细胞样记忆t细胞(或干样记忆t细胞),和两种类型的效应记忆t细胞:例如,tem细胞和temra细胞)、调控t细胞(也被称为抑制t细胞)、自然杀伤t细胞、粘膜相关性不变t细胞和γδt细胞。细胞毒性t细胞(ctl或杀伤t细胞)是能够诱导感染的体细胞或肿瘤细胞死亡的t淋巴细胞的亚组。在某些实施方案中,表达car的t细胞表达foxp3以实现并维持t调控表型。
自然杀伤(nk)细胞可以是作为细胞介导的免疫力的一部分且在固有性免疫应答期间起作用的淋巴细胞。nk细胞不需要在先激活就能够实施其对靶细胞的细胞毒性作用。
本公开的主题的工程化免疫细胞可表达特异性结合肿瘤抗原的细胞外抗原结合域(例如,人scfv,任选地交联的fab,或f(ab)2),用于治疗癌症,例如,用于治疗实体肿瘤。此类工程化免疫细胞可被施用至有需要的受试者(例如,人受试者),用于治疗癌症。在一些实施方案中,免疫细胞是淋巴细胞,诸如t细胞、b细胞或自然杀伤(nk)细胞。在某些实施方案中,工程化免疫细胞是t细胞。t细胞可以是cd4+t细胞或cd8+t细胞。在某些实施方案中,t细胞是cd4+t细胞。在某些实施方案中,t细胞是cd8+t细胞。
本公开的工程化免疫细胞可进一步包括至少一种重组或外源共刺激配体。例如,可进一步用至少一种共刺激配体转导本公开的工程化免疫细胞,使得工程化免疫细胞共表达或经诱导以共表达靶向肿瘤抗原的car和至少一种共刺激配体。靶向肿瘤抗原的car和至少一种共刺激配体之间的相互作用提供了非抗原特异性的信号,该信号对于免疫细胞(例如,t细胞)的完全激活是重要的。共刺激配体包括,但不限于,肿瘤坏死因子(tnf)超家族的成员,和免疫球蛋白(ig)超家族配体。tnf是参与全身性炎症且刺激急性期反应的细胞因子。其主要作用在于调节免疫细胞。tnf超家族的成员享有许多共同特征。大多数tnf超家族成员被合成为ii型跨膜蛋白(细胞外c端),其含有短细胞质区段和相对长的细胞外区。tnf超家族成员包括,不限于,神经生长因子(ngf)、cd40l(cd40l)/cd154、cd137l/4-1bbl、tnf-a、cd134l/ox40l/cd252、cd27l/cd70、fas配体(fasl)、cd30l/cd153、肿瘤坏死因子β(tnfp)/淋巴毒素-α(lta)、淋巴毒素-βο-τβ)、激活cd257/b细胞的因子(baff)/blys/thank/tall-1、糖皮质激素诱导的tnf受体配体(gitrl)和tf-相关的诱导细胞凋亡的配体(trail)、light(tnfsf14)。免疫球蛋白(ig)超家族是一大组参与细胞的识别、结合或粘附过程的细胞表面蛋白和可溶性蛋白。这些蛋白与免疫球蛋白共享结构特征—它们具备免疫球蛋白结构域(折叠)。免疫球蛋白超家族配体包括,但不限于,cd80和cd86(cd28的两个配体)、pd-l1/(b7-h1)(pd-1的配体)。在某些实施方案中,至少一种共刺激配体选自4-1bbl、cd80、cd86、cd70、ox40l、cd48、tnfrsf14、pd-l1,和其组合。在某些实施方案中,工程化免疫细胞包含一种重组共刺激配体,其是4-1bbl。在某些实施方案中,工程化免疫细胞包含两种重组共刺激配体,其是4-1bbl和cd80。包含至少一种共刺激配体的car描述于美国专利号8,389,282(其通过提述以其整体并入)中。
此外,本公开的工程化免疫细胞可进一步包含至少一种外源细胞因子。例如,可进一步用至少一种细胞因子转导本公开的工程化免疫细胞,使得工程化免疫细胞分泌该至少一种细胞因子以及表达靶向肿瘤抗原的car。在某些实施方案中,至少一种细胞因子选自il-2、il-3、il-6、il-7、il-11、il-12、il-15、il-17和il-21。在某些实施方案中,细胞因子是il-12。
工程化免疫细胞可从外周供体淋巴细胞生成,该外周供体淋巴细胞是例如,sadelain,m.,等,natrevcancer3:35-45(2003)(公开了经遗传修饰以表达car的外周供体淋巴细胞)中,morgan,r.a.等,science314:126-129(2006)(公开了经遗传修饰以表达识别全长肿瘤抗原的t细胞受体复合物(包含α和β异二聚体)的外周供体淋巴细胞)中,panelli等,jimmunol164:495-504(2000);panelli等jimmunol164:4382-4392(2000)(公开了衍生自肿瘤活检中的肿瘤浸润性淋巴细胞(til)的淋巴细胞培养物)中,和dupont等cancerres65:5417-5427(2005);papanicolaou等blood102:2498-2505(2003)(选择性地公开了采用人工抗原呈递细胞(aapc)或脉冲的树突状细胞的v/yro-扩增的抗原特异性外周血淋巴细胞)中公开的外周血供体淋巴细胞。工程化免疫细胞(例如,t细胞)可以是自体的、非自体的(例如,同种异体的)或在体外衍生自工程化祖细胞或干细胞。
在某些实施方案中,本公开的工程化免疫细胞(例如,t细胞s)表达每个具有本公开的靶向肿瘤抗原的car和/或前药转化酶的细胞约1至约5、约1至约4、约2至约5、约2至约4、约3至约5、约3至约4、约4至约5、约1至约2、约2至约3、约3至约4或约4至约5载体拷贝数。
例如,工程化免疫细胞中car表达水平越高,工程化免疫细胞表现出更大的细胞毒性和细胞因子产生。具有高的靶向肿瘤抗原的car表达水平的工程化免疫细胞(例如,t细胞)可诱导抗原特异性细胞因子产生或分泌和/或对具有低表达水平的靶向肿瘤抗原的car的组织或细胞(例如,约2,000或更少、约1,000或更少、约900或更少、约800或更少、约700或更少、约600或更少、约500或更少、约400或更少、约300或更少、约200或更少、约100或更少的肿瘤抗原结合位点/细胞)表现出细胞毒性。此外或可替换地,本公开的工程化免疫细胞(例如,t细胞)的细胞毒性和细胞因子产生与靶组织或靶细胞中肿瘤抗原的表达水平成比例。例如,靶标中人肿瘤抗原的表达水平越高,则工程化免疫细胞表现出越大的细胞毒性和细胞因子产生。
如本文所述的,前药转化酶的共表达通过在靶位点将前药转化为活性药物来增加cart细胞的细胞毒性作用。在某些实施方案中,本公开的工程化免疫细胞表现出针对表达肿瘤抗原的细胞的细胞毒性作用,其是在缺少前药转化酶的情况下的细胞毒性作用的至少约2倍、约3倍、约4倍、约5倍、约6倍、约7倍、约8倍、约9倍、约10倍、约20倍、约30倍、约40倍、约50倍、约60倍、约70倍、约80倍、约90倍或约100倍。
未纯化的免疫细胞来源可以是本领域已知的任何来源,例如骨髓、胎儿、新生儿或成年或其他造血细胞来源,例如胎儿肝、外周血或脐带血。可采用多种技术以使细胞分开。例如,阴性选择方法可最初去除非免疫细胞。单克隆抗体对于鉴定与特定细胞谱系和/或分化阶段用于阳性和阴性选择有关的标记物特别有用。
最初可以通过相对粗略的分离去除大部分末期分化的细胞。例如,磁珠分离最初可用于去除大量不相关细胞。优选地,至少约80%,通常为至少70%的总造血细胞将在细胞分离之前被去除。
分离的程序包括,但不限于,密度梯度离心;重置;与修改细胞密度的颗粒偶联;使用抗体包被的磁珠的磁分离;亲和色谱法;连接mab或与mab缀合使用的细胞毒性剂,其包括但不限于补体和细胞毒素;和使用附接于固体基质(例如板、芯片)的抗体的淘选(panning)、淘析或任何其他方便的技术。
用于分离和分析的技术包括但不限于流式细胞术,其可以具有不同的复杂程度,例如,多个颜色通道、低角度和钝角光散射检测通道、阻抗通道。
通过采用与死细胞相关的染料(诸如碘化丙啶(pi)),可以针对死细胞选择细胞。优选地,将细胞收集在包含2%胎牛血清(fcs)或0.2%牛血清白蛋白(bsa)的培养基或任何其他合适的,优选无菌的等渗培养基中。
在一些实施方案中,工程化免疫细胞包含一种或多种另外的修饰。例如,在一些实施方案中,工程化免疫细胞包含并表达(经转导以表达)识别受体的抗原,该受体结合不同于所选择的肿瘤抗原的第二抗原。除了在工程化免疫细胞上的本公开的car之外还纳入识别抗原的受体,可增加car或包含car的工程化免疫细胞对靶向的细胞的亲合力,尤其是,car是对特定肿瘤抗原具有低结合亲和力(例如,kd为约2x10-8m或更多、约5x10-8m或更多、约8x10-8m或更多、约9x10-8m或更多、约1x10-7m或更多、约2x10-7m或更多或约5x10-7m或更多)的car。
在某些实施方案中,识别抗原的受体是嵌合共刺激受体(ccr)。在krause,等,j.exp.med.188(4):619-626(1998)和us20020018783(其内容通过提述以其整体并入)中描述了ccr。ccr模拟共刺激信号,但与car不同,不提供t细胞激活信号,例如,ccr缺少cd3ζ多肽。在不存在抗原呈递细胞上的天然共刺激配体的情况下,ccr提供共刺激,例如,cd28样信号。组合性抗原识别,即,ccr与car组合使用,可增大t细胞针对表达双抗原的t细胞的反应性,从而改善选择性肿瘤靶向。kloss等,描述了一种策略,该策略整合了组合性抗原识别、分裂信号传导以及(极为重要地)t细胞激活和共刺激的平衡强度以生成t细胞,该t细胞消除表达抗原组合的靶细胞的同时保留单独表达每种抗原的细胞的(kloss等,naturebiotechnology31(l):71-75(2013))。在这种方法情况下,t细胞激活需要car介导的一种抗原的识别,而共刺激是由对第二种抗原特异的ccr独立介导的。为了实现肿瘤选择性,组合性抗原识别方法将t细胞激活的效率降低到无法通过同时ccr识别第二种抗原提供挽救的情况下无效的水平。在某些实施方案中,ccr包含细胞外抗原结合域、跨膜域和共刺激信号传导区,该细胞外抗原结合域结合不同于所选择的肿瘤抗原的抗原,该共刺激信号传导区包含至少一种共刺激分子,该共刺激分子包括,但不限于,cd28、4-1bb、ox40、icos、pd-1、ctla-4、lag-3、2b4和btla。在某些实施方案中,ccr的共刺激信号传导区包含一种共刺激信号传导分子。在某些实施方案中,该一种共刺激信号传导分子是cd28。在某些实施方案中,该一种共刺激信号传导分子是4-ibb。在某些实施方案中,ccr的共刺激信号传导区包含两种共刺激信号传导分子。在某些实施方案中,该两种共刺激信号传导分子是cd28和4-ibb。选择第二抗原,使得所选择的肿瘤抗原和第二抗原二者的表达被限制在靶向的细胞(例如,癌组织或癌细胞)。类似于car,细胞外抗原结合域可以是scfv、fab、f(ab)2;或具有异源序列以形成细胞外抗原结合域的融合蛋白。在某些实施方案中,ccr包含结合cd138的scfv、包含cd28多肽的跨膜域和包含两种共刺激信号传导分子的共刺激信号传导区,该两种共刺激信号传导分子是cd28和4-ibb。
[2]在某些实施方案中,识别抗原的受体是截短的car。“截短的car”不同于缺少细胞内信号传导域的car。例如,截短的car包含细胞外抗原结合域和跨膜域,且缺少细胞内信号传导域。依照本公开的主题,截短的car具有对表达在靶向的细胞(例如骨髓瘤细胞)上的第二抗原的高结合亲和力。截短的car作为粘附分子起作用,其增强本公开的car尤其是对肿瘤抗原具有低结合亲和力的car的亲合力,从而改善本公开的car或包含本公开的car的工程化免疫细胞(例如,t细胞)的功效。在某些实施方案中,截短的car包含结合cd138的细胞外抗原结合域、包含cd8多肽的跨膜域。本公开的t细胞包含或经转导以表达靶向肿瘤抗原的本公开的car和靶向cd138的截短的car。在某些实施方案中,靶向的细胞是实体肿瘤细胞。
在一些实施方案中,工程化免疫细胞经进一步修饰以抑制一种或多种基因的表达。在一些实施方案中,工程化免疫细胞经由基因组编辑被进一步修饰。已经描述了用于靶向的切割基因组dna的多种方法和组合物。此类靶向的切割事件可用于,例如,诱导靶向的诱变、诱导靶向的细胞dna序列缺失以及促进靶向的预定染色体基因座处的重组。参见,例如,美国专利号7,888,121;7,972,854;7,914,796;7,951,925;8,110,379;8,409,861;8,586,526;美国专利公开20030232410;20050208489;20050026157;20050064474;20060063231;201000218264;20120017290;20110265198;20130137104;20130122591;20130177983和20130177960,其公开的内容通过提述以其整体并入。这些方法通常涉及使用工程化切割系统以在靶dna序列中诱导双链断裂(dsb)或切口,使得通过错误出现过程(例如非同源末端连接(nhej))的断裂的修复或使用修复模板(同源重组修复或hdr)的修复可导致基因敲除或感兴趣的序列(靶向整合)的插入。切割可通过使用特定的核酸酶(诸如工程化锌指核酸酶(zfn)、转录激活子样效应核酸酶(talen)),或使用具有工程化crrna/tracrrna(‘单链指导rna’)以指导具体的切割的crispr/cas系统来发生。在一些实施方案中,工程化免疫细胞经修饰以破坏或减少内源t细胞受体基因的表达(参见,例如wo2014153470,其通过提述以其整体并入)。在一些实施方案中,工程化免疫细胞经修饰以导致破坏或抑制pd1、pdl-1或ctla-4(参见,例如美国专利公开20140120622),或本领域已知的其它免疫抑制因子(wu等(2015)oncoimmunology4(7):e1016700,mahoney等(2015)naturereviewsdrugdiscovery14,561–584)。
载体
许多表达载体是可获得的并且是本领域技术人员已知的,且可以用于表达本文提供的多肽,例如,用于表达嵌合抗原受体、第一前药转化酶和/或第二前药转化酶。表达载体的选择将受到宿主表达系统选择的影响。此类选择完全在技术人员的技术水平内。通常,表达载体可包括转录启动子和任选地增强子、翻译信号以及转录和翻译终止信号。用于稳定转化的表达载体典型地具有可选择的标记物,该标记物允许选择和维持转化的细胞。在一些情况下,复制的起点可用于扩增细胞中载体的拷贝数目。
载体还可含有与连接的核酸分子可操作地连接的另外的核苷酸序列,诸如,例如,表位标签(诸如用于定位),例如六-his标签或myc标签、血凝素标签或用于纯化的标签(例如gst融合蛋白),以及用于指导蛋白质分泌和/或膜缔合的序列。
抗体或其抗原结合片段的表达可通过本领域已知的任何启动子/增强子来控制。适合的细菌启动子是本领域熟知的,且在下文中描述。用于哺乳动物细胞、酵母细胞和昆虫细胞的其它适合的启动子是本领域熟知的,且以下示例了一些。用于指导异源核酸表达的启动子的选择取决于特定应用,且在本领域技术人员的水平内。可使用的启动子包括但不限于含有以下启动子的真核表达载体:sv40早期启动子(bernoist和chambon,nature290:304-310(1981))、劳斯肉瘤病毒的3'长末端重复序列中含有的启动子(yamamoto等,细胞22:787-797(1980)),疱疹胸苷激酶启动子(wagner等,proc.natl.acad.sci.usa75:1441-1445(1981)),金属硫蛋白基因的调控序列(brinster等,nature296:39-42(1982));原核细胞表达载体诸如β-内酰胺酶启动子(jay等,proc.natl.acad.sci.usa75:5543(1981))或tac启动子(deboer等,proc.natl.acad.sci.usa50:21-25(1983));还参见“usefulproteinsfromrecombinantbacteria”:inscientificamerican242:79-94(1980));含有以下启动子的植物表达载体:胭脂碱合成酶启动子(herrera-estrella等,nature505:209-213(1984))或花椰菜花叶病毒35srna启动子(gardner等,nucleicacidsres.9:2871(1981)),和光合酶核酮糖双磷酸羧化酶的启动子(herrera-estrella等,nature510:115-120(1984));来自酵母和其他真菌的启动子元件,诸如gal4启动子、醇脱氢酶启动子、磷酸甘油激酶启动子、碱性磷酸酶启动子,以及以下表现出组织特异性并已用于转基因动物的动物转录控制区:在胰腺腺泡细胞中有活性的弹性蛋白酶i基因控制区(swift等,cell55:639-646(1984);ornitz等,coldspringharborsymp.quant.biol.50:399-409(1986);macdonald,hepatology7:425-515(1987));在胰腺β细胞中有活性的胰岛素基因控制区(hanahan等,nature515:115-122(1985)),在淋巴样细胞中有活性的免疫球蛋白基因控制区(grosschedl等,cell55:647-658(1984);adams等,nature515:533-538(1985);alexander等,mol.cellbiol.7:1436-1444(1987)),在睾丸、乳腺、淋巴样和肥大细胞中有活性的小鼠乳腺肿瘤病毒控制区(leder等,cell15:485-495(1986)),在肝脏中有活性的白蛋白基因控制区(pinckert等,genesanddevel.1:268-276(1987)),在肝脏中有活性的α-甲胎蛋白基因控制区(krumlauf等,mol.cell.biol.5:1639-403(1985));hammer等,science255:53-58(1987)),在肝脏中有活性的α-1抗胰蛋白酶基因控制区(kelsey等,genesanddevel.7:161-171(1987)),在髓样细胞中有活性的β珠蛋白基因控制区(magram等,nature515:338-340(1985));kollias等,cell5:89-94(1986)),在脑的少突胶质细胞中有活性的髓磷脂碱性蛋白基因控制区(readhead等,cell15:703-712(1987)),在骨骼肌中有活性的肌球蛋白轻链-2基因控制区(shani,nature514:283-286(1985)),和在下丘脑的促性腺激素细胞中有活性的促性腺释放激素基因控制区(mason等,science254:1372-1378(1986))。
除了启动子,表达载体典型地含有转录单元或表达盒,其含有在宿主细胞中表达抗体或其部分所需的所有其他元件。典型的表达盒含有与编码抗体链可操作地连接的启动子、以及转录物的有效聚腺苷酸化、核糖体结合位点和翻译终止所需的信号。盒的另外的元件可包括增强子。此外,盒典型地含有结构基因下游的的转录终止区以提供有效终止。终止区可以从与启动子序列相同的基因获得,或者可从不同的基因获得。
一些表达系统具有提供基因扩增的标记,诸如胸苷激酶和二氢叶酸还原酶。可替换地,不涉及基因扩增的高产量表达系统也是合适的,诸如在昆虫细胞中使用杆状病毒载体,其具有在多面体启动子或其他强杆状病毒启动子的指导下编码种系抗体链的核酸序列。
本领域技术人员已知的用于将dna片段插入载体的任何方法均可用于构建含有编码本文提供的任何多肽的核酸的表达载体。这些方法可包括体外重组dna和合成技术,以及体内重组(遗传重组)。插入到克隆载体中可以,例如,通过将dna片段连接到具有互补粘性末端的克隆载体中来完成。如果克隆载体中不存在用于片段化dna的互补限制位点,则可酶促修饰dna分子的末端。可替换地,通过将核苷酸序列(接头)连接到dna末端可产生期望的任何位点;这些连接的接头可含有编码限制性核酸内切酶识别序列的特定的化学合成的核酸。
有用于产生本文提供的多肽的示例性质粒载体含有强启动子(诸如hcmv即时早期增强子/启动子或mhci类启动子),增强转录物加工的内含子(诸如hcmv即时早期基因内含子a),和聚腺苷酸化(聚a)信号(诸如晚期sv40聚a信号)。
工程化免疫细胞(例如,t细胞、nk细胞)的遗传修饰可通过用重组dna或rna构建体转导基本上均质的细胞组合物来完成。载体可以是逆转录病毒载体(例如,γ逆转录病毒),其用于将dna或rna构建体引入宿主细胞基因组。例如,编码靶向肿瘤抗原的car和前药转化酶的多核苷酸可被克隆至逆转录病毒载体中,且表达可衍生自其内源性启动子,衍生自逆转录病毒长末端重复序列,或衍生自替换的天然内部启动子。
同样可使用非病毒载体或rna。可使用随机染色体整合、或靶向整合(例如,使用核酸酶、转录激活子样效应核酸酶(talen)、锌指核酸酶(zfn),和/或成簇的规则间隔的短回文重复序列(crispr))、或转基因表达(例如,使用天然或化学修饰的rna)。
对于细胞的初始遗传修饰以提供表达靶向肿瘤抗原的car和前药转化酶的细胞,一般采用逆转录病毒载体用于转导,然而可使用任何其他适合的病毒载体或非病毒递送系统。对于随后的细胞遗传修饰以提供包含包含至少两种共同刺激性配体的抗原呈递复合物的细胞,逆转录病毒基因转移(转导)同样证明是有效的。逆转录病毒载体和适当的包装系的组合也是适合的,其中衣壳蛋白将具有感染人细胞的功能。已知多种产生双嗜性病毒的细胞系,包括但不限于pa12(miller,等mol.cell.biol.5:431-437(1985));pa317(miller,等mol.cell.biol.6:2895-2902(1986));和crip(danos,等proc.natl.acad.sci.usa85:6460-6464(1988))。非双嗜性颗粒也是适合的,例如,用vsvg、rd114或galv包膜假型化的颗粒和本领域已知的任何其它。
可能的转导方法还包括将细胞与生产细胞直接共培养(例如,通过bregni,等blood80:1418-1422(1992)的方法),或仅用病毒上清液或含有或不含适当生长因子和聚阳离子的浓缩载体储备物进行培养(例如,通过xu,等exp.hemat.22:223-230(1994);和hughes,等j.clin.invest.89:1817(1992)的方法)。
转导病毒载体可用于在工程化免疫细胞中表达共刺激配体和/或分泌细胞因子(例如,4-1bbl和/或il-12)。优选地,选择的载体表现出高感染效率以及稳定的整合和表达(参见,例如,cayouette等,humangenetherapy8:423-430(1997);kido等,currenteyeresearch15:833-844(1996);bloomer等,journalofvirology71:6641-6649,1997;naldini等,science272:263267(1996);和miyoshi等,proc.natl.acad.sci.u.s.a.94:10319,(1997))。可以使用的其他病毒载体包括,例如,腺病毒、慢病毒和腺相关病毒载体、牛痘病毒、牛乳头瘤病毒或疱疹病毒,诸如爱泼斯坦-巴尔病毒(epstein-barrvirus)(还参见,例如,miller,humangenetherapy15-14,(1990);friedman,science244:1275-1281(1989);eglitis等,biotechniques6:608-614,(1988);tolstoshev等,currentopinioninbiotechnology1:55-61(1990);sharp,thelancet337:1277-1278(1991);cornetta等,nucleicacidresearchandmolecularbiology36:311-322(1987);anderson,science226:401-409(1984);moen,bloodcells17:407-416(1991);miller等,biotechnology7:980-990(1989);legallasalle等,science259:988-990(1993);和johnson,chest107:77s-83s(1995))的载体)。逆转录病毒载体被开发得特别好,且已被用于临床环境(rosenberg等,n.engl.j.med323:370(1990);anderson等,美国专利号5,399,346)。
在某些非限制性实施方案中,表达本公开的靶向肿瘤抗原的car的载体是逆转录病毒载体,例如,肿瘤逆转录病毒(oncoretroviral)载体。
也可采用非病毒方法用于在细胞中表达蛋白质。例如,可以通过在脂质转染的存在下施用核酸(feigner等,proc.nat'l.acad.sci.u.s.a.84:7413,(1987);ono等,neuroscienceletters17:259(1990);brigham等,am.j.med.sci.298:278.(1989);staubinger等,methodsinenzymology101:512(1983)),脱唾液酸黏蛋白-多聚赖氨酸缀合(wu等,journalofbiologicalchemistry263:14621(1988);wu等,journalofbiologicalchemistry264:16985(1989)),或通过在手术条件下的微注射(wolff等,science247:1465(1990))将核酸分子引入细胞。其他非病毒的基因转移方式包括使用磷酸钙、deae葡聚糖、电穿孔和原生质体融合进行体外转染。脂质体也可能对将dna递送到细胞中有益。也可以通过将正常核酸离体转移到可培养的细胞类型(例如自体或异源原代细胞或其后代)中来完成将正常基因移植到受试者的受影响组织中,之后该细胞(或其后代)被注射到靶向的组织中或被全身性注射。也可以使用转座酶或靶向的核酸酶(例如,锌指核酸酶、大范围核酸酶或tale核酸酶)衍生出或获得重组受体。可通过rna电穿孔获得瞬时表达。
可从任何适合的启动子(例如,人巨细胞病毒(cmv)、猿猴病毒40(sv40)或金属硫蛋白启动子)来指导,且通过任何适当的哺乳动物调控元件或内含子(例如,延伸因子la增强子/启动子/内含子结构)来调控供在多核苷酸疗法方法中使用的cdna表达。例如,如果需要,可以使用已知优先指导特定细胞类型中的基因表达的增强子来指导核酸的表达。所使用的增强子可包括,但不限于,被表征为组织或细胞特异性增强子的那些。可替换地,如果将基因组克隆用作治疗性构建体,则可以通过同源调控序列或如果需要通过衍生自异源来源的调控序列(包括上述的任何启动子或调控元件)来介导调控。
所得的细胞可以在与未修饰的细胞相似的条件下生长,由此可以扩增修饰的细胞并将其用于多种目的。
vi.多肽和类似物和多核苷酸
本公开的主题还包括特异性结合肿瘤抗原(例如,人肿瘤抗原)的细胞外抗原结合域(例如,scfv(例如,人scfv)、fab或a(fab)2)、cd3ζ、cd8、cd28等、多肽或其片段,以及当在工程化免疫细胞中表达时,以增强其抗肿瘤活性的方式修饰的编码其的多核苷酸。本公开的主题提供了通过产生序列改变来优化氨基酸序列或核酸序列的方法。此类改变可包含某些突变、缺失、插入或翻译后修饰。本公开的主题进一步包括本公开的主题的任何天然存在的多肽的类似物。类似物可以通过氨基酸序列差异、通过翻译后修饰或通过两者与本公开的主题的天然存在的多肽有差异。本公开的主题的类似物一般可与本公开的主题的天然存在的氨基酸序列的全部或部分展现出至少约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%)、约98%、约99%或更多同一性或同源性。序列比对的长度为至少约5、约10、约15、约20、约25、约50、约75、约100个或更多个氨基酸残基。此外,在测定同一性程度的示例性方法中,可使用blast程序,其中介于e-3和e-100之间的概率得分,用于指示密切相关的序列。修饰包含多肽的体内和体外化学衍生,例如,乙酰化、羧化、磷酸化或糖基化;此类修饰可在多肽合成或加工期间或在用分离的修饰酶处理之后发生。类似物还可以通过一级序列的改变而不同于本公开的主题的天然存在的多肽。这些包括遗传变体,天然的和诱导的(例如,源自通过辐射或暴露于乙烷甲基硫酸盐中的随机诱变或通过位点特异性诱变,如上述sambrook,fritsch和maniatis,molecularcloning:alaboratorymanual(第2版),csh出版社,1989,或ausubel等中所述的)。还包括环化的肽、分子和类似物,其含有除l-氨基酸外的残基,例如d-氨基酸或非天然存在的或合成的氨基酸,例如β或γ氨基酸。
除全长多肽外,本公开的主题还提供本公开的主题的任何一种多肽或肽域的片段。片段可以是至少约5、约10、约13或约15个氨基酸。在一些实施方案中,片段为至少约20个连续氨基酸,至少约30个连续氨基酸,或至少约50个连续氨基酸。在一些实施方案中,片段为至少约60至约80、约100、约200、约300或更多个连续氨基酸。本公开的主题的片段可通过本领域普通技术人员已知的方法生成,或者可由正常蛋白质加工产生(例如,从新生多肽中去除对于生物活性不需要的氨基酸,或通过替代的mrna剪接或替代的蛋白质加工事件去除氨基酸)。
非蛋白质类似物具有经设计以模拟本发明蛋白质的功能活性的化学结构。根据本公开的主题的方法施用此类类似物。此类类似物可超过原始多肽的生理活性。类似物设计的方法在本领域中是熟知的,并且类似物的合成可根据此类方法通过修饰化学结构来进行,使得所得的类似物当在工程化免疫细胞中表达时增加原始多肽的抗肿瘤活性。这些化学修饰包括,但不限于,取代替代的r基团和改变参考多肽的特定碳原子处的饱和度。蛋白质类似物可以对体内降解有抵抗,从而在施用后导致更延长的治疗效果。用于测量功能活性的测定包括但不限于以下实例中描述的那些。
依照本公开的主题,编码特异性结合肿瘤抗原(例如,人肿瘤抗原)的细胞外抗原-结合域(例如,scfv(例如,人scfv)、fab或(fab)2)、cd3、cd8、cd28)的多核苷酸可通过密码子优化来修饰。密码子优化可改变天然存在的基因序列和重组基因序列,以在任何给定的表达系统中实现最高的可能的生产力水平。蛋白质表达的不同阶段涉及的因素包括密码子适应性、mrna结构以及转录和翻译中的多种顺式元件。本领域技术人员已知的任何适合的密码子优化方法或技术可用于修饰本公开的主题的多核苷酸,包括,但不限于,optimumgenetm、encoroptimization和blueheron。
施用
表达本公开的主题的靶向肿瘤抗原的car和前药转化酶的工程化免疫细胞可以全身地或直接地提供给受试者用于治疗或预防瘤形成。在某些实施方案中,将工程化免疫细胞直接注射到感兴趣的器官(例如,受瘤形成影响的器官)中。可替换地或另外地,例如,通过施用至循环系统(例如,肿瘤脉管系统)中,将工程化免疫细胞间接提供至感兴趣的器官。可以在施用细胞和组合物之前、期间或之后提供扩增和分化剂,以增加t细胞在体外或体内的产生。
可以在任何生理上可接受的媒介物中全身或局部地,通常在血管内、腹膜内、鞘内或胸膜内施用本公开的主题的工程化免疫细胞,尽管也可将它们引入骨骼或其他方便的位点,在这些位点中细胞可以找到适于再生和分化的位点(例如胸腺)。在某些实施方案中,可施用至少1x105个细胞,最终达到1x1010或更多个。在某些实施方案中,可施用至少1x106个细胞。包含工程化免疫细胞的细胞群可包含纯化的细胞群。,本领域技术人员使用多种公知的方法,诸如荧光激活细胞分选(facs)可容易地测定细胞群中工程化免疫细胞的百分比。包含工程化免疫细胞的细胞群的纯度范围可以是约50%至约55%、约55%至约60%、约65%至约70%、约70%至约75%、约75%至约80%、约80%至约85%;约85%至约90%、约90%至约95%或约95至约100%。剂量可以由本领域技术人员容易地调整(例如,纯度的降低可能需要剂量的增加)。可以通过注射、导管等引入工程化免疫细胞。如果需要,还可以包括多种因子,包括,但不限于白介素例如,il-2、il-3、il6、il-11、il-7、il-12、il-15、il-21以及其它白介素,集落刺激因子诸如g-、m-和gm-csf,干扰素例如,γ-干扰素。
在某些实施方案中,本公开的主题的组合物包含药物组合物,该药物组合物包含表达靶向肿瘤抗原的car和前药转化酶的工程化免疫细胞以及药学上可接受的载剂。施用可以是自体的或非自体的。例如,表达靶向肿瘤抗原的car和前药转化酶的工程化免疫细胞和包含其的组合物可以从一个受试者中获得,且被施用至同一受试者或不同的、相容的受试者。可通过局部注射,包括导管施用,全身性注射,局部注射,静脉内注射或肠胃外施用,来施用本公开的主题的外周血衍生的t细胞或其后代(例如,体内、离体或体外衍生的)。当施用本公开的主题的药物组合物(例如,包含表达靶向肿瘤抗原的car的工程化免疫细胞的药物组合物)时,它可配制成单位剂量可注射形式(溶液、混悬液、乳剂)。
配制剂
表达靶向肿瘤抗原的car和前药转化酶的工程化免疫细胞和包含其的组合物可以方便地作为无菌液体制剂的形式提供,无菌液体制剂例如等渗水溶液、悬浮液、乳剂、分散剂或粘性组合物,其可以被缓冲至所选的ph。液体制剂通常比凝胶、其他粘性组合物和固体组合物更容易制备。此外,液体组合物在某种程度上更方便施用,尤其是通过注射。在另一方面,可在适当的粘度范围内配制粘性组合物以提供与具体组织的更长的接触时间。液体或粘性组合物可包含载剂,所述载剂可以是溶剂或分散介质,其含有,例如,水、盐水、磷酸盐缓冲盐水、多元醇(例如,甘油、丙二醇、液体聚乙二醇等)及其适合的混合物。
可以通过在所需量的适当溶剂中掺入本公开的主题的组合物,例如包含工程化免疫细胞的组合物,来制备无菌注射溶液,根据需要,该溶剂中包含多种量的其它成分。此类组合物可以与适合的载剂、稀释剂或赋形剂诸如无菌水、生理盐水、葡萄糖、右旋糖等混合。组合物也可被冻干。组合物可含有辅助物质,诸如润湿剂、分散剂或乳化剂(例如,甲基纤维素)、ph缓冲剂、胶凝剂或增粘剂、防腐剂、调味剂、色素等,这取决于施用途径和期望的制剂。可查阅通过提述并入本文的标准文本,诸如“remington'spharmaceuticalscience”,第17版,1985年,以制备适合的制剂,而无需过度的实验。
可添加增强组合物的稳定性和无菌性的多种添加剂,包括抗微生物防腐剂、抗氧化剂、螯合剂和缓冲剂。可通过多种抗细菌和抗真菌剂,例如对羟基苯甲酸酯、三氯叔丁醇、苯酚、山梨酸等,来确保防止微生物的作用。可通过使用延迟吸收的试剂,例如单硬脂酸铝和明胶,来产生可注射药物形式的延长吸收。然而,依照本公开的主题,所使用的任何媒介物、稀释剂或添加剂将必须与本公开的主题的工程化免疫细胞相容。
该组合物可以是等渗的,即它们可以具有与血液和泪液相同的渗透压。本公开的主题的组合物的期望的等渗性可以使用氯化钠或其它药学上可接受的试剂(诸如葡萄糖、硼酸、酒石酸钠、丙二醇或其他无机或有机溶质)来完成。特别优选氯化钠用于含有钠离子的缓冲液。
如果需要,可使用药学上可接受的增稠剂将组合物的粘度维持在选择的水平。可以使用甲基纤维素,因为它容易且经济上可获得并且易于起作用。其他适合的增稠剂包括,例如,黄原胶、羧甲基纤维素、羟丙基纤维素、卡波姆等。增稠剂的浓度可取决于选择的试剂。重点是要使用能够达到选择的粘度的量。明显地,适合的载体和其它添加剂的选择将取决于准确的施用途径和特定剂型的性质,例如液体剂型(例如,是否将组合物配制成溶液、悬浮液、凝胶或其它液体形式,诸如定时释放形式或液体填充的形式)。
本领域技术人员将认识到,应该选择组合物的组成以使其具有化学惰性,并且不会影响如本公开的主题中所述的工程化免疫细胞的生存力或功效。对于本领域的化学和制药学原理的技术人员而言,这不会存在问题,或存在通过参考标准文本或通过简单的实验(不涉及过度的实验),可容易避免的问题,所述标准文本或简单的实验来自本公开和本文引用的文献。
关于本公开的主题的工程化免疫细胞的治疗用途的一个考虑是达到最佳效果所需的细胞数量。对于所治疗的受试者,待施用的细胞数量将有所不同。在某些实施方案中,向受试者施用约102至约1012、约103至约1011、约104至约1010、约105至约109或约106至约108个本公开的主题的工程化免疫细胞。可以以甚至更少的数目施用更有效的细胞。在一些实施方案中,向人受试者施用至少约1x108、约2x108、约3x108、约4x108、约5x108、约1x109、约5x109、约1x1010、约5x1010、约1x1011、约5x1011、约1x1012或更多个本公开的主题的工程化免疫细胞。可基于每个受试者的个体因素,包括其尺寸、年龄、性别、体重和特定受试者的情况,来精确测定所认为的有效剂量。本领域技术人员从本公开和本领域知识中可容易地确定剂量。通常,以对患者无毒或可耐受的剂量施用前药。在一些实施方案中,以比活性药物可能的用量高约10至约1000倍的量施用前药。前药的适合剂量可以是每位患者每天约0.1至200mg/kg,诸如约10至100mg/kg,或5至2000mg/m2(例如,200mg/m2)。
本领域技术人员可容易地确定组合物中的细胞和任选的添加剂、媒介物和/或载剂的量,并且可以在本发明公开的主题的方法中施用。典型地,(除一种或多种活性细胞和/或一种或多种剂以外的)任何添加剂溶液以按重量计约0.001%至约50%的量存在于磷酸盐缓冲盐水中,且活性成分以微克至毫克为单位存在,诸如约0.0001wt%至约5wt%、约0.0001wt%至约1wt%、约0.0001wt%至约0.05wt%、约0.001wt%至约20wt%、约0.01wt%至约10wt%或约0.05wt%至约5wt%。对于待施用于动物或人类的任何组合物,以及对于任何特定的施用方法,应确定毒性,诸如通过在适合的动物模型(例如啮齿动物诸如小鼠)中确定致死剂量(ld)和ld50;和引起适合应答的一种或多种组合物的剂量、其中的组分浓度和施用一种或多种组合物的时间。根据技术人员的知识、本公开和本文引用的文献,此类测定不需要过度的实验。而且,无需过度实验即可确定顺序施用的时间。
治疗方法
对于治疗,本文提供的工程化免疫细胞的施用的量是有效产生期望效果的量,例如,治疗癌症或一种或多种癌症症状。可以在本文提供的工程免疫细胞的一次或一系列施用中提供有效量。可以在推注中或通过连续灌注提供有效量。对于使用抗原特异性的t细胞的过继免疫疗法,典型地输注范围为约106至约1010的细胞剂量。如本文公开的前药转化酶的共表达可以允许较低剂量的待施用的工程化免疫细胞,例如,约104至约108。在将工程免疫细胞施用至受试者中后,诱导了针对一种肿瘤抗原的工程化免疫细胞。t细胞的“诱导”可包括,诸如通过缺失或无能性,使抗原特异性t细胞失活。失活对于建立或重建耐受性(诸如自身免疫性病症中的耐受性)特别有用。可通过本领域已知的任何方法施用本公开的主题的工程化免疫细胞,所述方法包括,但不限于,胸膜施用、静脉内施用、皮下施用、结内施用、肿瘤内施用、鞘内施用、胸膜内施用、腹膜内施用和对胸腺的直接施用。在某些实施方案中,工程化免疫细胞和包含其组合物被静脉内待施用至有需要的受试者。用于施用用于过继性细胞疗法的细胞的方法(包括,例如,供体淋巴细胞输注和cart细胞疗法)和施用方案是本领域已知的,且可被用于施用本文提供的工程化免疫细胞。
本公开的主题提供提使用本文提供的工程化免疫细胞(例如,t细胞),表达靶向肿瘤抗原的受体(例如,car)和前药转化酶的多种方法。例如,本公开的主题提供降低受试者中肿瘤符合的方法。在一个非限制性实例中,降低肿瘤符合的方法包括向受试者施用有效量的本公开的工程化免疫细胞,和施用适合的前药以通过表达的前药转化酶进行转化,从而在受试者中诱导肿瘤细胞死亡。在一些实施方案中,在不同时间施用工程化免疫细胞和前药。例如,在一些实施方案中,施用工程化的免疫细胞,然后施用前药。在一些实施方案中,在施用工程化免疫细胞后1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、18小时、24小时、30小时、26小时、48小时或更长施用前药。
单独的本公开的工程化免疫细胞或与前药组合的本公开的工程化免疫细胞可降低肿瘤细胞的数目,降低肿瘤大小,和/或根除受试者中的肿瘤。在某些实施方案中,降低肿瘤负荷的方法包括向受试者施用有效量的工程化免疫细胞,从而在受试者中诱导肿瘤细胞死亡。适合的肿瘤的非限制性实例包括肾上腺癌、膀胱癌、血癌、骨癌、脑癌、乳腺癌、癌(carcinoma)、宫颈癌、结肠癌、结直肠癌、子宫体癌、耳鼻喉(ent)癌、子宫内膜癌、食管癌、胃肠癌、头颈癌、霍奇金氏病、肠癌、肾癌、喉癌、急性和慢性白血病、肝癌、淋巴结癌、淋巴瘤、肺癌、黑色素瘤、间皮瘤、骨髓瘤、鼻咽癌、成神经细胞瘤、非霍奇金氏淋巴瘤、口腔癌、卵巢癌、胰腺癌、阴茎癌、喉咽癌、前列腺癌、直肠癌、肉瘤、精原细胞瘤、皮肤癌、胃癌、畸胎瘤、睾丸癌、甲状腺癌、子宫癌、阴道癌、血管瘤及其转移。在一些实施方案中,癌症是复发性或难治性癌症。在一些实施方案中,癌症对一种或多种癌症疗法,例如一种或多种化学治疗药物具有抗性。
本公开的主题还提供增加或延长具有瘤形成(例如,肿瘤)的受试者的存活的方法。在一个非限制性实例中,增加或延长具有瘤形成(例如,肿瘤)的受试者的存活的方法包括向受试者施用有效量的本公开的工程化免疫细胞,从而增加或延长受试者的存活。本公开的主题进一步提供用于治疗或防止受试者中的瘤形成(例如,肿瘤)的方法,其包含向受试者施用本公开的工程化免疫细胞。
使用本公开的主题的工程化免疫细胞可以抑制其生长的癌症包括通常对免疫疗法有应答的癌症。治疗的癌症的非限制性实例包括多发性骨髓瘤、成神经细胞瘤、神经胶质瘤、急性髓样白血病、结肠癌、胰腺癌、甲状腺癌、小细胞肺癌和nk细胞淋巴瘤。在某些实施方案中,癌症是多发性骨髓瘤。
此外,本公开的主题提供对受试者中的癌细胞应答而增加免疫激活细胞因子产生的方法。在一个非限制性实例中,该方法包括向受试者施用本公开的工程化免疫细胞。免疫激活细胞因子可以是粒细胞巨噬细胞集落刺激因子(gm-csf)、ifnα、ifn-β、ifn-γ、tnf-a、il-2、il-3、il-6、il-11、il-7、il-12、il-15、il-21、干扰素调节因子7(irf7),及其组合。在某些实施方案中,包括本公开的主题的肿瘤抗原特异性的car的工程化免疫细胞增加gm-csf、ifn-γ和/或tnf-a的产生。
适合治疗的人受试者通常包括可通过临床标准区分的两个治疗组。具有“晚期疾病”或“高肿瘤负荷”的受试者是带有临床上可测量的肿瘤(例如,多发性骨髓瘤)的受试者。临床上可测量的肿瘤是一种可以基于肿瘤块(例如,通过触诊、cat扫描、超声波扫描、乳房x线照片或x射线;阳性生化或组织病理学标记本身不足以识别群体)检测的肿瘤。向这些受试者施用本公开的主题中体现的药物组合物,以引发抗肿瘤应答,达到减轻其病况的目标。理想地,肿瘤块的减少作为结果出现,但是任何临床改善均构成益处。临床改善包含进展的风险或速率降低或肿瘤(例如多发性骨髓瘤)的病理结果减少。
第二组适合的受试者在本领域称为“佐剂组”。其是已经有瘤形成史(例如,多发性骨髓瘤),但对另一种治疗方式有应答的个体。先前的疗法可以包括但不限于手术切除、放射疗法和传统化学疗法。结果,这些个体没有临床上可测量的肿瘤。然而,他们被怀疑有在原发肿瘤位点附近或通过转移的疾病进展风险。此组可以进一步细分为高风险和低风险的个体。根据在初始治疗之前或之后观察到的特征进行细分。这些特征在临床领域中是已知的,并且针对每种不同的瘤形成适当地定义。高危亚组的典型特征是肿瘤(例如,多发性骨髓瘤)已经侵犯邻近组织或显示淋巴结的参与的特征。另一组具有肿瘤形成(例如,多发性骨髓瘤)的遗传易感性,但尚未证实肿瘤形成(例如,多发性骨髓瘤)的临床体征。例如,对与乳腺癌相关联的基因突变测试呈阳性但仍处于育龄的女性,希望可在预防性治疗中接受本文所述的一种或多种抗原结合片段,以预防瘤形成的发生,直到其适合进行预防性手术为止。
受试者可具有疾病(例如,多发性骨髓瘤)的晚期形式,在这种情况下,治疗目标可包括减轻或逆转疾病进展和/或改善副作用。受试者可具有已经经治疗的病史,在这种情况下,治疗目标典型地包括降低或延迟复发风险。
可将进一步修饰引入至表达靶向肿瘤抗原的car的工程化免疫细胞(例如,t细胞)以避免或最小化免疫并发症(也称为“恶性t细胞转化”,例如,移植物抗宿主疾病(gvhd))的风险,或当健康组织表达与肿瘤细胞相同靶抗原时,导致与gvfid相似的结果。
在本文提供的一些实施方案中,工程化免疫细胞在细胞内表达前药转化酶。在一些实施方案中,细胞内前药转化酶是组成型表达的。在一些实施方案中,细胞内前药转化酶是条件性表达的。在一些实施方案中,患者被施用前药,该前药是细胞穿透性前药,其被工程化免疫细胞的细胞质内的细胞内前药转化酶转化为活性药物。在一些实施方案中,转化的活性药物导致工程化免疫细胞的死亡。可以在施用工程化免疫细胞后的时间点施用前药。在一些实施方案中,时间点取决于一种或多种因素,诸如一种或多种毒性迹象的出现(例如,发烧、低血压、水肿、肿胀、发红、皮疹、瘙痒、神经系统改变、昏迷、精神错乱、恶心、腹泻、咯血、血尿、便血)、肿瘤部位处免疫细胞的浓度、疾病的进展和/或由医生确定的受试者终止治疗的愿望。
工程化免疫细胞的进一步修饰还可包括将自杀基因改造至表达靶向肿瘤抗原的car的t细胞中。适合的自杀基因包括,但不限于,单纯疱疹病毒胸苷激酶激酶(hsv-tk)、可诱导的半胱天冬酶9自杀基因(icasp-9),和截短的人表皮生长因子受体(egfrt)多肽。在某些实施方案中,自杀基因是egfrt多肽。egfrt多肽可通过施用抗egfr单克隆抗体(例如,西妥昔单抗)使t细胞消除。egfrt可以共价连接到靶向肿瘤抗原的car的细胞内域的c端。可在包含编码本公开的靶向肿瘤抗原的car的核酸内的载体包括自杀基因。在方式中,在恶性t细胞转化(例如,gvhd)期间施用经设计以激活自杀基因的前药(例如,前药(例如,可激活icasp-9的api903)触发表达自杀基因激活的car的t细胞的凋亡。将自杀基因掺入到本公开的靶向肿瘤抗原的car中产生增加的安全性水平,具有在非常短的时间内消除大部分cart细胞的能力。掺入了自杀基因的本公开的工程化免疫细胞(例如,t细胞)可以在cart细胞输注后的给定时间点被抢先消除,或在毒性的最早迹象时被根除。
制品和试剂盒
本公开的主题提供用于治疗或防止瘤形成(例如,实体肿瘤)的试剂盒。在某些实施方案中,试剂盒包含单一剂量形式的含有有效量的工程化免疫细胞的治疗性或预防性组合物,所述工程化免疫细胞表达靶向肿瘤抗原的受体(例如car)、细胞内表达第一前药转化酶和在工程化免疫细胞表面的或分泌的第二前药转化酶。在一些的实施方案中,工程化免疫细胞进一步表达至少一种共刺激配体。在一些实施方案中,试剂盒包含无菌容器,其含有治疗性或预防性疫苗;此类容器含有盒、安瓿、瓶子、小瓶、管、袋子、小袋、泡罩包装或本领域已知的其他适合的容器形式。此类容器由塑料、玻璃、层压纸、金属箔或其它适合容纳药物的材料制成。
如果需要,可将工程化免疫细胞与用于向具有瘤形成(例如,实体肿瘤)或处于发展瘤形成(例如,实体肿瘤)的风险的受试者施用工程化免疫细胞的说明书一起提供。通常将包括关于组合物用于治疗或防止瘤形成(例如,实体肿瘤)的用途的信息。在其它实施方案中,说明书包括以下至少一项:治疗剂的描述;治疗或预防瘤形成(例如,实体肿瘤)或其症状的剂量计划表和施用;预防措施;警告;适应症;禁忌;过量信息;不良反应;动物药理学;临床研究;和/或参考文献。说明书可直接印在容器上(当存在时),也可以作为标签应用在在容器上,也可作为容器中或连同容器的独立的纸页、小册子、卡片或文件夹。
实施例
除非另有说明,否则本发明的实践采用分子生物学(包括重组技术)、微生物学、细胞生物学、生物化学和免疫学的常规技术,其完全在技术人员的能力范围内。此类技术已在文献中得到充分解释,诸如,“molecularcloning:alaboratorymanual”,第二版(sambrook,1989);“oligonucleotidesynthesis”(gait,1984);“animalcellculture”(freshney,1987);“methodsinenzymology”“handbookofexperimentalimmunology”(weir,1996);“genetransfervectorsformammaliancells”(miller和calos,1987);“currentprotocolsinmolecularbiology”(ausubel,1987);“pcr:thepolymerasechainreaction”,(mullis,1994);“currentprotocolsinimmunology”(coligan,1991)。这些技术适用于本发明的多核苷酸和多肽的生成,并且因此可以在进行和实施本发明时考虑。在随后的部分中将讨论用于特定实施方案的特别有用的技术。
提出以下实施例以向本领域普通技术人员提供如何制备和使用本发明的组合物以及测定、筛选和治疗方法的完整公开和描述,并且不旨在限制发明人认为其发明的范围。
实施例1.供与seaker细胞一起使用的细胞毒性和靶向的小分子的前药的合成。
此实施例描述了基于氨磺酰腺苷(ams)(结构1c)(图2c)的示例性前药的合成,以由细菌水解酶cpg2切割。ams是核杀菌素和有效的细胞毒性分子的密切类似物(hepg2ic50=9nm)。进行了详细的结构-活性关系(sar)研究,该研究指示,通过6-氨基(me2,>500μm)c2(ph,≈500μm),c8(n-pr,>250μm)或氨基磺酸盐位置(水杨基,>500μm)处的取代,ams的细胞毒性显著降低或被消除。基于这些sar数据,通过使用cdi-激活的双-o-t-bu-谷氨酸盐(30%)的6-n-酰化、脱甲硅基(tbaf)、氨磺酰化(h2nso2cl,dma)和丙酮化合物水解(tfa,h2o,15%经3个步骤后)从2′,3′-o-异亚丙基-5′-o-tbs-腺苷(结构3,图2d)合成ams前药1a(“p-ams”;图2c)。合成了经由co-pab-co和tml-co接头附接的两种可替代的n-氨基磺酸盐连接的前药(未显示)(图2b)。在此实施例中的以下提供了对p-ams合成和经由tml-co接头(“三甲基锁(trimethyllock)-ams”)附接的n-氨基磺酸盐连接的前药的合成的更详细的讨论。作为使用已知抗癌治疗剂的可替代细胞毒剂,氮芥前药zd2767p(结构2a,图2c)也如先前报道(springer等,jmedchem.37(15):2361–70(1994)和niculescu-duvaz等,tetrahedronlett.46(40):6919–22(2005))的合成。
作为靶向的治疗方法的实例,还设计了厄洛替尼前药(结构6a,图2e)。为了避免n,n-二芳基酰胺(参见co或co-pab-co接头)的潜在水解不稳定性,合成了在谷氨酸盐切割后经历自发消除的n-苄基接头(pab-co)。已证明对氨基苯甲酸季铵盐的相似的1,6-消除(staben等natchem.8(12):1112–9(2016)。通过将已知的苯甲醇(结构4,图2d)转化为相应的甲磺酸盐(mscl)(结构5,图.2d),厄洛替尼的n-烷基化(25%经2个步骤后),以及叔丁基酯的脱保护(tfa,ch2cl2,3%,未优化)来实现合成。在此实施例中的以下提供了此厄洛替尼前药合成的更详细的讨论和表征。
使用celltitre
还评估了ams相关结构的体外细胞毒性,以确定改变细胞毒性的ams修饰。使用celltitre
表1
出乎意料的是,尽管大多数修饰导致细胞毒性的显著降低,但是几种化合物持续具有足够的细胞毒性活性以用于本技术。例如,amsn、2’-脱氧-ams、6-desn-ams、2-nh2-ams和杀结核菌素-ms各自可以很好地用于生成前药和在本技术中使用。
式i的化合物的代表性合成和实例
p-ams的合成和表征
前药化合物p-ams的合成总结在下面的方案1中(其中p-ams也称为s5),且此后进一步详述。
方案1.
((9-((3ar,4r,6r,6ar)–6-(((叔丁基二甲基甲硅烷基)氧基)甲基)-2,2-二甲基四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-基)氨基甲酰基)-l-谷氨酸二叔丁酯)(s2)
如somu,r.v.;boshof,h.;qiao,c.;bennett,e.m.;barry,c.e.;aldrich,c.c.j.med.chem.2006,49(1),31-34中所述的制备s1。在25℃,在25ml圆底烧瓶中,将(s)-谷氨酸二叔丁酯(300mg,1.16mmol,1.0当量)悬浮于8ml无水dmf中。在一份中添加1,1’-羰基二咪唑(cdi,187mg,1.16mmol,1当量),且搅拌混合物1h。分别地,在矿物油中的60%nah(117.8mg,2.95mmol,2.6当量)添加在无水dmf(4ml)中的s1(828mg,1.96mmol,1.7当量)溶液,持续15分钟。在25℃下,将该分离的腺苷混合物逐滴加入到cdi激活的谷氨酸酯混合物中。将反应在25℃下搅拌过夜,持续另外16-20h,直到通过lc-ms判断已经发生完全转化。然后将混合物用饱和的nacl水溶液淬灭。用乙酸乙酯(320ml)萃取含水混合物,并将合并的有机萃取物经mgso4干燥,过滤,并通过旋转蒸发浓缩,以提供粗产物,其为黄色油。通过二氧化硅快速色谱法的纯化(3:1己烷/etoac至1:1己烷/etoac)产生产物s2(286mg,35%),其为浅黄色油。tlc:rf0.35(1:1己烷/etoac)。1h-nmr(500mhz,cdcl3):9.91(d,1h,j=8.0),8.61(s,1h),8.19(s,1h),7.94(s,1h),6.20(s,1h),5.23(dd,1h,j=10,5.2),4.94(dd,1h,j=10,4.9),4.61(q,1h,4.6),3.89(dd,1h,j=8.8,4.0),3.77(dd,1h,j=8.7,3.9),3.33(dd,1h,j=7.9,5.2),2.42(m,2h),2.26(m,1h),2.09(m,1h),1.64(s,3h),1.50(s,9h),1.43(s,9h),1.41(s,3h),0.83(s,9h),0.01(dd,6h,j=1.6)。13c-nmr(126mhz,cdcl3):206.8,172.0,171.0,153.3,151.3,150.0,120.8,114.1,91.9,87.5,85.1,82.1,81.5,80.5,77.2,63.6,60.3,53.4,31.6,30.9,28.3,28.0,27.2,25.8,25.3,21.0,18.3,14.2,-5.5。esi-msm/z(relint):(pos)707.4([m+h]+,100)。
((9-((3ar,4r,6r,6ar)-6-(羟甲基)-2,2-二甲基四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-yl)氨基甲酰基)-l-谷氨酸二叔丁酯(s3)。
在25℃,在50ml圆底烧瓶中,将s2(286mg,404μmol,1.0当量)悬浮于15mlthf中。在一份中添加thf中的1mtbaf(0.88ml,0.88mmol,2.2当量),且在25℃搅拌混合物1h。然后用饱和的nacl水溶液淬灭混合物。用乙酸乙酯(320ml)萃取含水混合物,并经mgso4干燥,过滤,并通过旋转蒸发浓缩,以提供粗产物(274mg),其为黄色油。粗产物无需进一步纯化即可用于下一步骤。粗1h-nmr(500mhz,cdcl3)δ8.57(s,1h),8.41(s,1h),6.01(d,j=4.0hz,1h),5.43(s,1h),5.18(dd,j=5.9,4.1hz,1h),5.05(d,j=5.9hz,1h),4.56(dt,j=7.9,3.9hz,1h),4.53(s,1h),3.95(d,j=1.8hz,0h),3.94–3.91(m,1h),3.77(d,j=12.6hz,1h),3.42(s,2h),3.34–3.27(m,1h),2.43–2.27(m,2h),2.27–2.17(m,1h),2.10–2.00(m,1h),1.61(s,3h),1.60(s,3h),1.48(s,9h),1.38(s,9h)。esi-msm/z(relint):(pos)593.3([m+h]+,100)。
((9-((3ar,4r,6r,6ar)-2,2-二甲基–6-氨磺酰基氧基)甲基)-四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-yl)氨基甲酰基)-l-谷氨酸二叔丁酯(s4)。
在0℃,在25ml圆底烧瓶中,将s3(274.4mg,462μmol,1.0当量)悬浮于10ml无水dma中。在一份中添加氨磺酰氯(133.4mg,1.156mmol,2.5当量),且加热至25℃搅拌混合物1h。然后用饱和的nahco3水溶液淬灭混合物。用乙酸乙酯(315ml)萃取含水混合物,并经mgso4干燥,过滤,并通过旋转蒸发浓缩,以提供粗产物(310mg),其为黄色油。粗产物无需进一步纯化即可用于下一步骤。粗1h-nmr(500mhz,cdcl3)δ8.24(s,1h),8.20(s,1h),5.94(d,j=2.8hz,1h),5.04(dd,j=6.1,2.8hz,1h),4.77(dd,j=6.2,2.6hz,1h),4.29–4.18(m,2h),4.02(dd,j=10.9,4.4hz,1h),3.97(dd,j=10.8,4.6hz,1h),3.72(q,j=7.1hz,2h),2.11–1.84(m,3h),1.65(s,6h),1.14(s,9h),1.05(s,9h)。esi-msm/z(relint):(pos)672.3([m+h]+,100)。
((9-((2r,3r,4s,5r)-3,4-二羟基-5-((氨磺酰基氧基)甲基)四氢呋喃-2-基)-9h-嘌呤-6-基)氨基甲酰基)-l-谷氨酸(s5)。
在25℃,在25ml圆底烧瓶中,将s4(310.3mg,462μmol,1.0当量)悬浮于22.5ml80%tfa水溶液中。在25℃下搅拌反应,持续另外的过夜,持续5-6小时,直到通过lc-ms判断已经发生完全转化。然后用10mlmeoh淬灭混合物,共沸共3次。然后通过制备型hplc纯化粗混合物,并冻干过夜,以产生s4(36.7mg,70.6mol,15%经三个步骤),其为白色粉末。1h-nmr(500mhz,甲醇-d4):δ8.67(s,1h),8.56(s,1h),6.20(d,j=4.9hz,1h),4.75(t,j=5.0hz,1h),4.63(dd,j=7.9,5.2hz,1h),4.46(dd,j=8.0,3.0hz,2h),4.43–4.35(m,2h),2.59–2.44(m,2h),2.37(dq,j=13.6,7.5,7.0hz,1h),2.16(dq,j=14.4,7.6hz,1h)。未观察到7个可交换的h。13c-nmr(126mhz,甲醇-d4):176.3,174.6,173.5,155.5,151.9,150.3,144.0,121.3,90.5,83.8,75.7,71.8,69.6,54.0,49.5,31.0,28.4。esi-msm/z(relint):(pos)520.2([m+h]+,100)。
三甲基锁-ams的合成和表征
前药化合物三甲基锁-ams的合成总结在下面的方案2中(其中三甲基锁-ams也称为s13),且此后进一步详述。
方案2.
((2-(4-((叔丁基二甲基甲硅烷基)氧基)-2-甲基丁烷-2-基)-3,5-二甲基苯氧基)羰基)-l-谷氨酸二叔丁酯(s7)。
如wang,b.;gangwar,s.;pauletti,g.m.;siahaan,t.j.;borchardt,r.t.;j.org.chem.1997,62,1363-1367中所述的合成s6。在25℃,在50ml圆底烧瓶中,将l-谷氨酸二叔丁酯(1.12g,4.32mmol,1.0当量)悬浮在10.79ml无水dmf中。在一份中添加1,1’-羰基二咪唑(cdi,770mg,4.75mmol,1.1当量)和三乙胺(662l,4.75mmol,1.1当量),且搅拌混合物1h。分别地,将s6(1.04g,3.24mmol,0.75当量)悬浮在无水dmf(10.8ml)中,并添加矿物油中的60%nah(173mg,4.32mmol,1.0当量)达15分钟。将此单独的s6混合物在25℃下逐滴添加至cdi激活的谷氨酸酯合物中。在25℃下搅拌反应过夜,持续另外17-20h,直至通过tlc判断已经发生完全转化。然后用饱和的nacl水溶液淬灭混合物,用50ml乙酸乙酯稀释。然后萃取有机层,依次用饱和的nacl水溶液(130ml),然后用水(230ml)洗涤。然后合并水层,并用乙酸乙酯(230ml)萃取。然后合并所有有机层,经mgso4干燥,过滤,并通过旋转蒸发浓缩,以提供粗产物,其为黄色油。通过二氧化硅快速色谱法纯化(9:1己烷/etoac至3:1己烷/etoac),产生产物s7(1.16g,60%)其为黄色油。tlc:rf0.46(3:1己烷/etoac)。1h-nmr(500mhz,cdcl3):δ6.77(s,1h),6.63(s,1h),5.67(d,j=8.2hz,1h),3.50(t,j=7.5hz,2h),2.50(s,3h),2.46–2.25(m,2h),2.21(s,3h),2.04(t,j=7.5hz,2h),2.01-1.92(m,1h),1.49(s,13h),1.45(s,9h),0.84(s,9h),-0.03(s,6h)。13cnmr(126mhz,cdcl3)δ155.6,137.5,135.7,128.9,126.6,116.5,61.8,45.0,39.4,32.0,28.0,25.5,20.2,18.3,-5.3。esi-msm/z(relint):(pos)608.5([m+h]+,100)。
((2-(4-羟基-2-甲基丁烷-2-基)-3,5-二甲基苯氧基)羰基)-l-谷氨酸二叔丁酯(s8)。
在25℃,在50ml圆底烧瓶中,将s7(1.159g,1.906mmol,1.0当量)悬浮在12ml1:1v:vthf:h2o溶液中。添加12ml乙酸,并在25℃下搅拌反应持续另外3h,直到通过lc-ms判断已经发生完全转化。然后通过旋转蒸发仪浓缩混合物,并将甲苯(10ml)添加到混合物中并通过旋转蒸发仪浓缩。重复进行与甲苯共沸共三次。粗产物s8(941mg)无需进一步纯化即可用于下一步骤。粗1h-nmr(500mhz,cdcl3)δ6.73(s,1h),6.59(s,1h),6.23(d,j=8.1hz,1h),4.23(m,1h),3.47(td,j=7.3,6.4,3.3hz,2h),2.45(s,3h),2.38–2.27(m,2h),2.16(s,3h),2.07–2.00(m,1h),1.99(s,3h),1.96–1.91(m,1h),1.90(s,3h),1.44(s,9h),1.42(s,9h)。粗13cnmr(126mhz,cdcl3)δ172.5,171.2,155.6,137.6,135.9,129.0,128.5,126.6,125.2,116.2,82.7,81.2,61.3,54.3,44.9,39.4,31.8,28.0,25.9,25.5,20.2。esi-msm/z(relint):(pos)494.4([m+h]+,100)。
((2-(4-羟基-2-甲基丁烷-2-基)-3,5-二甲基苯氧基)羰基)-l-谷氨酸二叔丁酯(s9)。
在0℃,在50ml圆底烧瓶中,将粗s8(941mg,1.906mmol,1.0当量)悬浮在19mlch2cl2中。添加戴斯-马丁过碘烷(dess-martinperiodinane)(1.78g,4.19mmol,2.2当量),并加热至25℃搅拌17h。然后通过旋转蒸发仪浓缩混合物,并通过s直接纯化,并将甲苯(10ml)添加到混合物中并通过旋转蒸发仪浓缩。通过二氧化硅快速色谱法的纯化(3:1己烷/etoac)产生产物s10(513mg,55%经两个步骤)其为浅黄色油。tlc:rf0.27(3:1己烷/etoac)。1h-nmr(500mhz,cdcl3):δ9.52(t,j=2.6hz,1h),6.78(s,1h),6.64(s,1h),5.85(d,j=8.1hz,1h),4.27(dt,j=8.1,4.0hz,2h),2.80(s,2h),2.48(s,3h),2.42–2.27(m,2h),2.19(s,3h),2.17–2.08(m,1h),1.98–1.88(m,1h),1.55(s,6h),1.54(s,3h),1.45(s,9h),1.42(s,9h)。13c-nmr(126mhz,cdcl3):δ203.7,172.2,170.9,154.4,153.2,152.8,149.3,137.7,136.8,135.0,133.0,132.5,94.3,82.6,81.0,68.4,64.7,56.9,54.2,49.2,38.9,38.8,38.2,31.6,28.1,27.5,27.2,25.7,25.5,22.6,20.8,20.3,15.2。esi-msm/z(relint):(pos)493.2([m+h]+,100)。
(s)-3-(2-(((1,5-二叔丁氧基-1,5-二氧代戊烷-2-基)氨基甲酰基)氧基)-4,6-二甲基苯基)-3-甲基丁酸(s10)。
在0℃,在50ml圆底烧瓶中,将粗s9(513mg,1.04mmol,1.0当量)悬浮在12ml1:1h2o:2-甲基-2-丁烯中。顺序地添加nah2po4(264mg,2.2mmol,2.1当量)、tbuoh(48.7mg,0.657mmol,0.63当量)和naclo2(416mg,3.685mmol,3.5当量),且加温至25℃搅拌22h。然后用饱和nh4cl水溶液淬灭混合物,然后用1mhcl水溶液将混合物的ph调节至~3。然后用乙酸乙酯(3
10ml)洗涤混合物,然后用盐水(brine)(220ml)洗涤合并的有机层。然后通过旋转蒸发仪浓缩有机层。通过二氧化硅快速色谱法的纯化(3:1己烷/etoac至1:1己烷/etoac)产生产物s10(384mg,72%),其为浅黄色油。tlc:rf0.15(1:1己烷/etoac)。1h-nmr(500mhz,cdcl3)δ6.76(s,1h),6.63(s,1h),5.96(d,j=8.2hz,1h),4.28(td,j=8.3,4.9hz,2h),2.90–2.74(m,2h),2.49(s,3h),2.45–2.26(m,2h),2.19(s,3h),2.15(m1h),1.99–1.91(m,1h),1.57(s,3h),1.56(s,3h),1.46(s,9h),1.43(s,9h)。13c-nmr(126mhz,cdcl3)δ176.4,172.0,170.9,154.6,149.3,137.7,136.0,133.6,132.1,123.0,82.3,80.7,60.3,54.1,48.0,38.6,31.4,31.2,27.9,27.4,25.1,20.9,20.1,14.0。esi-msm/z(relint):(pos)508.3([m+h]+,100)。
((2-(4-(((((3ar,4r,6r,6ar)–6-(6–氨基-9h-嘌呤-9-基)-2,2-二甲基四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)甲氧基)磺酰基)氨基)-2-甲基-4-氧代丁烷-2-基)-3,5-二甲基苯氧基)羰基)-l-谷氨酸二叔丁酯(s12)。
如moreau,c.;等j.med.chem.2013,56,10079-10102中所讨论的合成s11。在25℃,在25ml圆底烧瓶中,将s10(307mg,0.605mmol,1.0当量)悬浮在6ml无水dmf中。然后向溶液中添加edci(140mg,0.904mmol,3.2当量)和dmap(110mg,0.904mmol,3.1当量)持续20分钟。分别地,然后将s11(111mg,0.287mmol,1当量)悬浮在6ml无水dmf中,且然后逐滴被添加至s10混合物中。然后在25℃下搅拌反应过夜持续18h。然后用9ml乙酸乙酯稀释混合物,用饱和nh4cl水溶液(310ml)和饱和nahco3水溶液(310ml)洗涤,然后用mgso4干燥有机层。通过二氧化硅快速色谱法的纯化(ch2cl2中的5%meoh至ch2cl2中的10%meoh)产生半纯产物s12(83mg,33%),其为透明油。tlc:rf0.44(ch2cl2中的10%meoh)。粗1h-nmr(500mhz,cdcl3):7.38–7.28(m,5h),5.60(m,2h),4.63(m,1h),4.58(s,2h),4.36(t,1h,j=5.6),3.49(dd,1h,j=9.5,3.7),3.44(dd,1h,j=9.6,7.9),3.34(s,3h),3.32(s,3h),2.68(d,1h,j=2.5),2.52(m,1h),2.38(m,1h)。粗13c-nmr(125mhz,cdcl3):137.9,131.0,128.5–127.5(ar),103.8,73.6,73.4,66.7,53.5,53.2,31.8。esi-msm/z(relint):(pos)876.1([m+h]+,100)。
((2-(4-(((((2r,3s,4r,5r)-5-(6-氨基-9h-嘌呤-9-基)-3,4-二羟基四氢呋喃-2-基)甲氧基)磺酰基)氨基)-2-甲基-4-氧代丁烷-2-基)-3,5-二甲基苯氧基)羰基)-l-谷氨酸(s13)。
在25℃,在10ml玻璃小瓶中,将s12(20mg,22.8mol,1.0当量)悬浮在456l80%tfa水溶液中。然后在25℃下搅拌反应持续2h。然后用5mlh2o稀释混合物,通过旋转蒸发仪浓缩使其与具有甲苯的tfa(35ml)共沸。通过制备型hplc纯化和冻干,产生产物s13(6.95mg,42%),其为白色粉末。1h-nmr(500mhz,甲醇-d4)δ8.41(s,1h),8.29(s,1h),6.73(d,j=2.0hz,1h),6.63(s,1h),6.07(s,1h),4.57(t,j=5.1hz,1h),4.31(dd,j=9.4,4.9hz,1h),4.22(dt,j=9.7,3.7hz,4h),2.85(d,j=15.3hz,1h),2.77(d,j=15.3hz,1h),2.52(t,j=7.4hz,3h),2.48(s,3h),2.37–2.24(m,1h),2.15(s,3h),2.09–1.97(m,1h),1.60(d,j=3.0hz,6h),未观察到7个可交换的h。esi-msm/z(relint):(pos)724.3([m+h]+,100)。
现有技术的ams相关的前-前药的合成和表征
代表性的ams相关的前-前药化合物(s23)的合成总结在下面的方案3中,且其后进一步详述。
方案3
(s)-2-异氰酸酯戊二酸二苄酯(s18)
在0℃,在25ml圆底烧瓶中,将谷氨酸二苄酯(693mg,2.1mmol,1.0当量)悬浮在7mlch2cl2中,添加吡啶(681μl,8.46mmol),并搅拌15min。加入三光气(313mg,1.06mmol,0.5当量),并将所得黄色溶液在0℃下搅拌2h。用10ml冷的0.5mhcl水溶液和10ml碎冰萃取反应混合物两次。用10mlch2cl2再萃取每个水层。合并的有机相用10ml冷的饱和nacl水溶液和10ml碎冰的混合物萃取,经mgso4干燥,过滤,并通过旋转蒸发浓缩,以得到粗异氰酸酯(分离出733mg,99%),产物为浅棕色油(在处理过程中,异氰酸酯仅在水中暴露总共5-10min)。化合物s18的表征与如kozikowski,a.p.;etal.j.med.chem.,2004,47,1729–1738中先前所报道的一致。粗产物无需进一步纯化即可使用。
9-((3ar,4r,6r,6ar)-6-(((叔丁基二甲基甲硅烷基)氧基)甲基)-2,2-二甲基四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-胺(s19)。
在50ml的圆底烧瓶中,将s18(747mg,2.11mmol,1.75当量)悬浮在12ml的无水thf中,并加入三乙胺(186μl,1.33mmol,1.1当量)。分别地,将s1(509mg,1.21mmol,1.0当量)悬浮于12ml无水thf中,然后逐滴添加至s18混合物中。在回流下搅拌反应另外18小时,直到添加另外的s18(426mg,1.21mmol,1当量)和三乙胺(186μl,1.33mmol,1.1当量)并搅拌另外6小时。然后用40mlch2cl2稀释反应物,然后用饱和nacl水溶液(15ml)和水(15ml)洗涤有机层。然后通过旋转蒸发仪浓缩有机层,并通过二氧化硅快速色谱纯化(9:1己烷:etoac至3:1己烷:etoac至3:1etoac:己烷),产生产物s19(618mg,66%),其为浅黄色油。1h-nmr(500mhz,cdcl3)δ10.05(d,j=7.9hz,1h),8.53(s,1h),8.22(s,1h),8.07(s,1h),7.33(m,10h),6.21(d,j=2.5hz,1h),5.21(s,3h),5.18(s,1h),5.13(s,1h),5.09(s,3h),4.80(td,j=7.8,5.2hz,1h),4.59(td,j=8.1,4.8hz,1h),4.47(d,j=2.9hz,1h),3.89(dd,j=11.3,3.6hz,1h),3.77(dd,j=11.3,3.9hz,1h),2.45–2.37(m,2h),2.27–2.15(m,1h),2.04(m,1h),1.65(s,3h),1.42(s,3h),0.82(s,9h),0.01(d,j=2.2hz,6h)。13c-nmr(126mhz,cdcl3):172.3,171.5,153.4,151.3,149.9,141.2,135.8,135.3,128.2,120.8,114.2,91.9,87.5,85.2,81.4,67.2,66.4,63.6,52.8,41.2,30.4,27.8,27.2,25.8,18.3,13.8,-5.5.esi-msm/z(relint):(pos)775.6([m+h]+,100)。
((9-((3ar,4r,6r,6ar)-6-(((叔丁基二甲基甲硅烷基)氧基)甲基)-2,2-二甲基四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-yl)氨基甲酰基)-l-谷氨酸(s20)
在100ml圆底烧瓶中,将s19(618mg,0.797mmol,1.0当量)悬浮在40ml无水etoh中,加入10%pd/c(170mg,0.159mmol,0.2当量)和10%pd(oh2)/c(112mg,0.159mmol,0.2当量),并密封。然后将所得混合物真空抽空,并经历氢气气氛总共进行三次。然后设置氢气球,且反应进行2小时,并通过lc/ms判断完成。然后用硅藻土垫过滤所得混合物,然后用meoh(2×60ml)洗涤硅藻土垫。然后将所得滤液通过旋转蒸发仪浓缩并经受高真空,以产生产物s20(430mg,91%),其为透明油。该产物无需进一步纯化即可使用。粗1h-nmr(500mhz,methanol-d4):δ8.62(s,1h),8.45(s,1h),6.25(s,1h),5.42(d,j=6.1hz,1h),5.02(d,j=6.1hz,1h),4.50(d,j=6.7hz,1h),4.39(s,1h),4.27(s,1h),3.85(d,j=4.2hz,1h),3.82–3.72(m,1h),2.59–2.30(m,2h),2.15-2.10(m,2h),1.60(s,3h),1.39(s,3h),1.15(s,2h),0.82(s,9h),-0.01(s,6h)。粗13c-nmr(126mhz,methanol-d4)δ177.2,155.7,152.7,151.8,151.4,143.6,121.8,115.2,92.9,89.3,86.1,83.2,64.9,42.4,32.1,31.7,29.6,27.6,26.4,25.7,19.3,14.2,-5.2.esi-msm/z(relint):(pos)595.4([m+h]+,100)。
((9-((3ar,4r,6r,6ar)-6-(((叔丁基二甲基甲硅烷基)氧基)甲基)-2,2-二甲基四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-基)氨基甲酰基)-l-谷氨酸双((新戊酰氧基)甲基)酯(s21)
在25℃,在25ml圆底烧瓶中,将s20(260mg,0.437mmol,1.0当量)悬浮在2.1ml无水dmf中。加入三乙胺(184μl,1.31mmol,3当量)、新戊酸氯甲酯(630μl,4.37mmol,10当量)和四丁基碘化铵(484mg,1.31mmol,3当量),并搅拌5h。然后将混合物升至45℃,且搅拌另外23小时。然后将所得混合物用乙酸乙酯(10ml)稀释,用饱和nh4cl水溶液(2×10ml)和饱和nacl水溶液(2×10ml)洗涤,并经mgso4干燥。然后通过旋转蒸发仪浓缩有机层,随后通过二氧化硅快速色谱纯化(5%meoh的ch2cl2溶液),得到半粗产物s20(152mg,42%粗产率),其为黄色油。在tlc上观察到三个不能通过色谱分离的共斑点。tlc:rf0.45(5%meoh的ch2cl2溶液)。粗1h-nmr(500mhz,cdcl3):δ7.38–7.28(m,5h),5.60(m,2h),4.63(m,1h),4.58(s,2h),4.36(t,1h,j=5.6),3.49(dd,1h,j=9.5,3.7),3.44(dd,1h,j=9.6,7.9),3.34(s,3h),3.32(s,3h),2.68(d,1h,j=2.5),2.52(m,1h),2.38(m,1h)。粗13c-nmr(125mhz,cdcl3):δ137.9,131.0,128.5–127.5(ar),103.8,73.6,73.4,66.7,53.5,53.2,31.8.esi-msm/z(relint):(pos)823.7([m+h]+,100)。
((9-((3ar,4r,6r,6ar)-2,2–二甲基–6-((氨磺酰基氧基)甲基)四氢呋喃[3,4-d][1,3]二氧杂环戊烷-4-基)-9h-嘌呤-6-基)氨基甲酰基)-l-谷氨酸双((新戊酰氧基)甲基)酯(s22)
在25℃,在25ml塑料离心管中,将s21(76mg,0.128mmol,1.0当量)悬浮在1.3ml无水thf中。逐滴加入氟化氢吡啶(609μl,4.65mmol,50当量)并搅拌25分钟。然后通过加入冷的饱和nahco3水溶液至ph为7淬灭反应。然后用ch2cl2(2x10ml)萃取含水性混合物,并用水(2x5ml)洗涤合并的有机萃取液,经mgso4干燥,并通过旋转蒸发仪浓缩,得到粗中间体。然后在25℃,在15ml圆底烧瓶中,将粗中间体与苯(5ml)共沸,并将其悬浮在1.9ml无水dma中。在一份中加入氨磺酰氯(54mg,465μmol,5当量),搅拌30分钟。然后用乙酸乙酯(5ml)稀释反应混合物,并用饱和nacl水溶液(2×5ml)和水(2×5ml)洗涤有机层。然后通过旋转蒸发仪浓缩合并的有机萃取物,随后通过制备型hplc纯化并冻干以产生s22(55mg,75%,经两步),其为白色粉末。tlc:rf0.45(5%meohinch2cl2)。1hnmr(500mhz,氯仿-d)δ8.72(s,1h),8.63(s,1h),6.30(s,1h),5.87(d,j=5.7hz,1h),5.80(d,j=5.5hz,1h),5.72(s,1h),5.49–5.38(m,1h),5.01(d,j=6.0hz,1h),4.71(s,1h),4.32(q,j=10.8hz,2h),2.52(t,j=7.1hz,2h),2.33(dd,j=14.2,6.9hz,1h),2.18–2.06(m,1h),1.62(s,3h),1.40(s,3h),1.20(s,9h),1.19(s,9h)。esi-msm/z(relint):(pos)788.6([m+h]+,100)。
((9-((2r,3r,4s,5r)-3,4-二羟基-5-((氨磺酰基氧基)甲基)四氢呋喃-2-基)-9h-嘌呤-6-基)氨基甲酰基)-l-谷氨酸双((新戊酰氧基)甲基)酯
在25℃,在10ml玻璃小瓶中,将s22(12.5mg,15.9μmol,1.0当量)悬浮在1.58mltfa中。搅拌反应混合物6小时。用1mlmeoh淬灭反应,然后通过旋转蒸发仪浓缩。然后将粗混合物与meoh(3×1ml)共沸。然后通过制备型hplc纯化提取物,且经冻干以产生s23(3.91mg,33%),其为呈白色粉末。tlc:rf0.45(5%meohinch2cl2)。1hnmr(500mhz,甲醇-d4)δ8.67(d,j=1.6hz,1h),8.53(s,1h),8.47(s,1h),6.20(d,j=4.9hz,1h),5.94(d,j=5.6hz,1h),5.82(d,j=5.7hz,1h),5.76(t,j=4.7hz,2h),4.77(t,j=5.0hz,1h),4.69(td,j=5.5,2.7hz,1h),4.48(dd,j=9.6,4.4hz,1h),4.39(ddd,j=11.3,9.3,3.9hz,1h),2.61(h,j=9.9hz,2h),2.35(dt,j=11.8,6.0hz,1h),2.26–2.13(m,1h),1.23(d,j=3.1hz,9h),1.21(s,9h)。esi-msm/z(relint):(pos)748.5([m+h]+,55);537.3([m-ribose+h]+,100)。
式ii的化合物的代表性合成和实例
如上所注释的,氮芥前药zd2767p(结构2a,图2c)是如先前报道的(springer等,jmedchem.37(15):2361–70(1994)和niculescu-duvaz等,tetrahedronlett.46(40):6919–22(2005))合成的。基于本公开,包括关于式i和iii的前药化合物和式i的前-前药化合物的代表性合成,本领域普通技术人员能够很好地生成式ii的化合物。可在niculescu-duvaz等,j.med.chem.47(10):2651-58(2004),niculescu-duvaz等,j.med.chem.46(9):1690-705(2003),springer等,j.med.chem.38(26):5051-65,和springer等,eurjcancer27(11):1361-66(1991)(其中每个通过提述并入本文)以及其中每个引用的参考文献中发现对式ii的化合物的合成的进一步指导。
式iii的化合物的代表性合成和实例
glu-paba-厄洛替尼的合成和表征
厄洛替尼,glu-paba-厄洛替尼(s17),的代表性前药的合成总结于以下方案4中,且此后进一步详述。
方案4.
((4-(((甲磺酰基)氧基)甲基)苯基)氨基甲酰基)-l-谷氨酸二叔丁酯(s15)。
如niculescu-duvaz,d.;等j.med.chem.1998,41,5297–5309中所讨论的合成s14。在0℃,在15ml圆底烧瓶中,将s14(90mg,220μmol,1.0当量)悬浮在2.2ml无水ch2cl2中。添加三乙胺(61μl,441μmol,2当量)和甲磺酰氯(34μl,441μmol,2当量),并在0℃下搅拌反应持续另外2小时,直到通过tlc判断完成。然后用5mlch2cl2稀释混合物,并用水(110ml)洗涤有机层。然后用ch2cl2(310ml)洗涤水层,然后通过旋转蒸发仪浓缩合并的有机萃取物。将粗产物共沸(35ml)并置于高真空下1小时。粗产物s15(107mg)是不稳定的,且无需进一步纯化即可用于下一步骤。tlc:rf0.80(1:1己烷/etoac)。粗1hnmr:(500mhz,cdcl3)δ7.34(m,hz,2h),7.24–6.85(m,2h),3.66(s,3h),2.42–2.27(m,2h),2.13–2.03(m,1h),1.89(m,1h),1.40(s,9h)。
((4-(((6,7-双(2-甲氧基乙氧基)喹唑啉-4-yl基)(3-乙炔基苯基)氨基)甲基)苯基)氨基甲酰基)-l-谷氨酸二叔丁酯(s16)。
在0℃,在15ml圆底烧瓶中,将厄洛替尼(72mg,184μmol,1.0当量)悬浮在1.8ml无水dmf中,且添加矿物油中的60%nah(11mg,275μmol,1.5当量)持续5min。分别地,将s15(107mg,220μmol,1.2当量,假设100%纯)悬浮在1.8ml无水dmf中,冷却至0℃,然后在0℃逐滴添加至厄洛替尼混合物。在0℃下搅拌所得物持续另外30min,直到通过lc-ms判断已经发生完全转化。然后用1ml水淬灭混合物,然后用乙酸乙酯(35ml)洗涤水层。然后通过旋转蒸发仪浓缩合并的有机萃取物,置于高真空下,且无需进一步纯化即可作为粗产物s16(11.1mg)用于下一步骤。esi-msm/z(relint):(pos)784.7([m+h]+,100)。
((4-(((6,7-双(2-甲氧基乙氧基)喹唑啉-4-基)(3-乙炔基苯基)氨基)甲基)苯基)氨基甲酰基)-l-谷氨酸(s17)
在25℃,在10ml玻璃小瓶中,将粗化合物s16(11.1mg,14.1μmol,1.0当量,假设100%纯)悬浮在950μl1:1tfa:ch2cl2溶液中。将反应搅拌1小时,直到通过lc-ms判断已经发生完全转化。然后将混合物与meoh(33ml)共沸,通过旋转蒸发仪浓缩,并通过制备型hplc纯化,以产生产物s17(0.43mg,经两个步骤3.5%产量),其为白色粉末。1hnmr(500mhz,甲醇-d4)δ8.79(s,1h),7.58–7.40(m,4h),7.32–7.21(m,8h),7.17(s,1h),6.42(s,1h),5.52(s,2h),4.37(dd,j=8.4,5.1hz,1h),4.32–4.27(m,3h),3.84–3.77(m,3h),3.65(s,1h),3.52(d,j=5.2hz,8h),3.41(s,3h),3.35(s,3h),2.42(m,6.7hz,2h),2.25–2.13(m,1h),2.02–1.89(m,1h)。esi-msm/z(relint):(pos)672.4([m+h]+,100)。
基于本公开,包括关于式i的前-前药化合物的代表性合成,本领域普通技术人员能够很好地产生式iii的化合物。
实施例2.测试前药候选物的方法
用于与选择的前药转化酶一起使用的候选前药可以通过体外测定法测试,并在每个步骤进行过滤以实现目标产品概况(表2)。
表2
这些结果可用于设计改善的前药。使用重组的、纯化的cpg2确定cpg2介导的前药切割的动力学(km,kcat)(jeyaharan等proteinexprpurif.127:44-52(2016))。如先前针对zd2767p(springeretal.,jmedchem.37(15):2361–70(1994)和springer等,jmedchem.38(26):5051–65(1995))和甲氨蝶呤(sherwood等,eurjbiochem.148(3):447–53(1985))所述的,基于降低的uv吸光度来监测反应。在生理条件下确认前药的稳定性。发展了kcat/km≥1s–1μm–1(理想的kcat/km≥10)的前药。
可在抗原阳性的肿瘤细胞系中测试每个前药/药物对的细胞毒性,所述抗原阳性的肿瘤细胞系包括hl60(白血病)、jurkat(t细胞)、mda231(乳腺)、ovcar(卵巢)、sw480(结肠)、set2(aml),以及mdr变体和原代癌细胞。发展了药物活性≤1μm(例如,≤100nm)且si≥10(例如,≥100)的对。
还测试了在纯化的cpg2和来自分泌cpg2的hek293t细胞的上清液的存在下的前药细胞毒性。通过lcms/ms定量前药和揭露的从属药物。发展了ic50在游离药物的20倍以内(例如,5倍)的前药。
还测试了表达分泌的、表面缔合的和细胞内的cpg2的hek293t细胞中的药物和前药活性。发展了ic50在游离药物的20倍以内(例如,5倍)的前药。
还测试了前药的体外adme(吸收、分布、代谢和排泄)特性:例如,ph7.4时的溶解度、小鼠/人血浆/全血稳定性、小鼠/人微粒体稳定性(±nadph、±udpga)、小鼠/人血浆蛋白结合性。发展了具有可接受的体外特性的前药以用于体内评估。
实施例3.表达嵌合抗原受体的细胞的构建
此实施例描述了包含car(例如,具有对cd19、wt1或prame特异的抗原结合域)的细胞的构建体。在此示例性实施方案中,cpg2经工程化以具有跨膜域且定位到细胞膜的细胞外侧,在那里它可水解并激活与细胞接触的前药为活性药物(例如,实施例1的前药,其被水解为活性细胞毒性药物)。药物的激活在经由抗原结合结构域靶向cart细胞的位点处的肿瘤内局部增强溶细胞作用。
已经制得cd19cart细胞和wt1cart细胞,并描述了有关产生它们和表征其活性的方法的最初出版物(brentjens等,sci.trans.med.5(177):177ra38(2013),pegram等leukemia29(2):415-22(2015))。将会类似地制得和表征prame反应性cart细胞。
利用哺乳动物优化的cd19、wt1和pr20(prame300-309)scfv序列以生成car的抗原结合域。经由(gly4ser)3接头连接scfv的可变重和轻链。向scfv的n端添加前导肽(例如,cd8信号序列)用于定位。在替换的实施方案中,可添加c-myc标签,以允许通过流式细胞术检测car。cd8跨膜域跟随抗原结合结构域。在car的细胞质侧,由于4-1bbcart细胞的持久性增加,因此4-1bb用作共刺激元件(oka等pnas.101:13885-90(2004))。如果需要,可优化car,以在cd8跨膜域的上游包括间隔区域。将编码car的核酸克隆至含有4-1bb信号传导和cd3ζ激活域的sfg逆转录病毒载体中,这形成第二代car(brentjens等clincancerres.13(18pt1):5426-35(2007))。也可生成没有c-myc标签的临床级构建体,为此可生成允许检测car的抗独特型mab。
如前所述,生成了产生稳定hek293t病毒的细胞系,经亚克隆且用于转导原代人t细胞(curran等americansocietyofgenetherapy23(4):769-78(2015))。转导后,使用抗独特型抗体检测抗cd19scfv,通过流式细胞术验证car表达。
为了cpg2酶的共表达,如实施例5中所述的,将编码羧肽酶cpg2酶的核酸克隆到car载体中。
实施例4.跨膜和分泌的cpg2构建体的生成和表达的cpg2的表征
此实施例描述了跨膜和分泌形式的cpg在细菌或哺乳动物细胞中表达时的生成及其表征。跨膜形式的cpg2在编码cpg2的序列的5′端和在其3′端编码cd8信号序列(malpvtalllplalllhaarp(seqidno:11),该序列衍生自cd8的跨膜域的序列,以允许酶的膜锚定和细胞外暴露(图3a)。
首先使用pet151directional
用于cpg2的氨基酸序列(无起始met和前导信号序列)如下:
alaqkrdnvlfqaatdeqpaviktleklvnietgtgdaegiaaagnfleaelknlgftvtrsksaglvvgdnivgkikgrggknlllmshmdtvylkgilakapfrvegdkaygpgiaddkggnavilhtlkllkeygvrdygtitvlfntdeekgsfgsrdliqeeakladyvlsfeptsagdeklslgtsgiayvqvqitgkashagaapelgvnalveasdlvlrtmniddkaknlrfqwtiakagqvsniipasatlnadvryarnedfdaamktleeraqqkklpeadvkvivtrgrpafnageggkklvdkavayykeaggtlgveertgggtdaayaalsgkpvieslglpgfgyhsdkaeyvdisaiprrlymaarlimdlgagk(seqidno:3,无前导序列且经优化用于人表达的cpg2)
生成跨膜cpg2构建体的两个变体。一种构建体包括cd8信号序列、cpg2序列、可选的ha标签、cd8跨膜部分和cd8细胞内部分。
cd8信号序列:
malpvtalllplalllhaarp(seqidno:11)
cd8跨膜部分:
iyiwaplagtcgvlllslvit(seqidno:38)
cd8细胞内域氨基酸序列:
lycnhrnrrrvckcprpvvksgdkpslsaryv(seqidno:13)
完整cpg-cd8tm序列如下:
malpvtalllplalllhaarpalaqkrdnvlfqaatdeqpaviktleklvnietgtgdaegiaaagnfleaelknlgftvtrsksaglvvgdnivgkikgrggknlllmshmdtvylkgilakapfrvegdkaygpgiaddkggnavilhtlkllkeygvrdygtitvlfntdeekgsfgsrdliqeeakladyvlsfeptsagdeklslgtsgiayvqvqitgkashagaapelgvnalveasdlvlrtmniddkaknlrfqwtiakagqvsniipasatlnadvryarnedfdaamktleeraqqkklpeadvkvivtrgrpafnageggkklvdkavayykeaggtlgveertgggtdaayaalsgkpvieslglpgfgyhsdkaeyvdisaiprrlymaarlimdlgagkypydvpdyagggiyiwaplagtcgvlllslvitlycnhrnrrrvgggrpvvksgdkpslsaryv(seqidno:40)。
第二构建体与上述序列相同,除了天然lyk结合域被破坏以使cd8的内源性功能与cpg分子分开。具有经破坏的lyk结合域的cd8跨膜域部分:lycnhrnrrrvgggrpvvksgdkpslsaryv(seqidno:15)。
对于分泌形式的cpg2,cpg2信号序列(mrpsihrtaiaavlatafvagt(seqidno:9)被插入在编码cpg2的序列的5′端(图3b),以促进分泌。
将cpg2-pet151载体转化到bl21明星大肠杆菌细胞中,并用iptg诱导。在frenchpress中使细菌匀浆,并使用标准规程通过在ni++珠上的亲和色谱法纯化。经由pbs透析交换咪唑,经由脱盐旋转柱纯化蛋白质。测定经表达和纯化的cpg2以证实活性酶是谷氨酸化前药的有效转化体。图4a显示了初始细菌产生的cpg2是高活性的。在体外通过重组cpg2激活pro-ams。在浓度递增的纯化的cpg2的存在下,将人jurkatt细胞与p-ams或不与p-ams孵育。通过celltitre-
通过将cpg2变体克隆至pcdna3.1+载体(invitrogen),实施在真核细胞中的瞬时表达研究,该pcdna3.1+载体利用用于真核表达的cmv启动子。使用脂质体以在hek293t细胞中转染进行表达研究。
然后将cpg2表达载体转导至hek293t细胞中,以表征跨膜形式的适当表达和活性。图4证明了膜锚定的cpg2的表面表达。用表达血凝素(ha)标签的膜系链的cpg2(cpg2tm)转染hek293t细胞。使细胞暴露于用抗ha抗体的表面染色,并与用同种型对照抗体染色的细胞比较(图5)。
图6通过比较用于检测跨膜形式的hek293t总细胞裂解物(图6,左)与用于分泌形式的上清液(图6,右),证明了多种cpg构建体(跨膜或分泌形式的)的定位。
图7证明在hek293t细胞中表达的分泌形式的cpg2在体外具有酶促活性。使jurkatt细胞暴露于递增浓度的ams(圆形)或仅具有培养基的pro-ams(三角形)或具有来自分泌cpg2的hek293t细胞的上清液(正方形)的pro-ams。在48小时通过celltitre-
图8进一步明了在hek293t细胞中表达的分泌形式的cpg2在体外具有酶促活性。使jurkatt细胞暴露于递增浓度的仅具有培养基的zd2767p(三角形)或具有来自分泌cpg2的hek293t细胞的上清液的zd2767p(正方形)。在48小时处通过celltitre-
还使用逆转录病毒plgpw载体(一种衍生自plhcx(addgene)的经修饰的载体),其驱动从5’ltr启动子的gfp表达和从cmv启动子的cpg2表达。生成稳定表达整合的cpg2变体的hek293t细胞系。通过转染逆转录病毒载体和vsvg糖蛋白生成逆转录病毒,用于在gp293包装细胞(clontech)中进行假分型。
图9证明了在稳定转染的hek293t细胞中表达的细胞分泌的和跨膜的cpg2激活ams前药。无信号肽的cpg2(cpg2nosigp)的表达用作对照。用cpg2变体稳定转染的hek293t细胞被暴露于p-ams(1μm)48小时,并通过明视野显微镜分析(图9a)。使细胞暴露于不同浓度的p-ams,并在处理后48小时分析细胞活力(图9b)。细胞分泌的和跨膜的cpg2各自诱导显著的细胞死亡,而对照构建体则没有。
实施例5.包含cd19、wt1或pramecart以及cpg2酶的seaker细胞的构建
为了生成实施例3中所述的cart细胞的分泌cpg2的和cpg2跨膜seaker变体,将分别编码分泌形式或跨膜形式的cpg2酶的核酸克隆并插入car的cd3-ζ链的下游,通过生成独立cpg2蛋白的自切割p2a肽序列而分离(图3a、b)。
使用gibsonassembly策略(neb)以将cpg构建体插入到表达cd19scfv的现有sfg逆转录病毒载体中来生成构建体。简而言之,将抗cd19sfg载体(19-1bbz和19-28z)线性化到cd3ζ序列的下游,并插入含有p2a序列且随后是cpg变体的片段(参见,例如seqidno:48和49)。
将car-cpg2构建体转导至hek293t细胞中,以表征细胞上清液中分泌形式的表达和分泌以及体外酶促活性。如实施例4所证明的,该分泌形式在激活p-ams和zd2767p前药的细胞毒性药物中显示出显著活性。跨膜形式也是有活性的。
除了跨膜和分泌形式的cpg2,还使用合成的notch可诱导的系统来构建cpg酶的条件性表达的形式(图3c)(参见例如,kloss等,naturebiotechnology31(1):71-5(2013);lim等cell168(4):724-40(2017);和roybal等cell164(4):770-9(2016))。此格式仅在啮合原代car的靶标时才门控cpg2(分泌的或在膜中)的表达。因此,仅在与靶细胞接触时,cpg2酶选择性激活前药,这进一步增强特异性,并预期降低局部和全身性的潜在毒性。评定了从前药形成药物的动力学、seeker构建体的效力和对抗原阴性细胞的脱靶作用。
实施例6.用编码抗cd19car和cpg2的seaker细胞逆转录病毒构建体转导的原代t细胞和t细胞系的表征
然后,将编码上述抗cd19car和cpg2酶的逆转录病毒构建体引入t细胞系(例如,jurkat)和原代t细胞中,以测量嵌合抗原受体(car)和cpg2的表达并表征它们的活性。用编码仅具有4-1bb共刺激元件的抗cd19car(19bbz)或具有cpgnosigp(无信号肽对照)或cpgtm(跨膜)基因的抗cd19car的逆转录病毒转导的jurkat细胞和原代人t细胞。通过对在细胞表面上抗cd19scfv表达进行染色和流式细胞术评定转导水平。图10证明了car蛋白在表达cpg2酶的工程化免疫细胞的细胞表面上表达。
还评定了表达具有4-1bb共刺激元件(bbz)的抗cd19car和无信号肽的cpg基因(cpgnosigp)或cpg跨膜基因的人jurkatt细胞的活性。将用19bbz/cpg逆转录病毒转导的人jurkatt细胞与gfp+raji(人伯基特淋巴瘤)癌靶细胞系共培养24小时(1:1比率),并进行流式细胞术分析。通过gfp表达区分jurkat和raji细胞(左图)。通过评定gfp阴性细胞中cd69的表达来检查jurkat细胞的激活(右图)。图11显示如通过增加的cd69表达所证明的,cart细胞识别靶癌细胞并被靶癌细胞激活,这证明了该途径在seaker细胞中起作用。
在单独的实验中,还评定了表达具有4-1bb共刺激元件(bbz)的抗cd19car和无信号肽(cpgnosigp)的cpg基因或cpg跨膜基因的原代人t细胞的活性。原代人t细胞与人19bbz和19bbz/cpgcar-t细胞与gfp+raji细胞以1:1比率共培养。48小时孵育后,通过流式细胞术分析细胞(图12a),并计算%gfp+细胞(图12b)。gfp阴性细胞(表示准备的多种cart细胞)增加至与传统cart和2种不同的seaker细胞相同的水平,显示seaker细胞具有与传统car相当的活性。
还分析了用逆转录病毒构建体转导的用于分泌cpg2的人t细胞。通过免疫印迹分析来自编码19bbz、19bbz/cpg或对照19bbz/cpgnosigp构建体的人car-t细胞的细胞上清液的cpg2表达。如图13中所示的,分泌cpg2的seaker细胞免疫印迹显示在上清液中发现了该酶,这表明该细胞活跃地分泌cpg2酶。具有无信号序列的cpg2(cpg3nosigp)的seaker细胞不分泌该酶。
实施例7.人t细胞对厄洛替尼的抵抗和通过跨膜cpg2的谷氨酸化厄洛替尼前药(p-厄洛替尼)的转化
可通过采用前药来改善工程化免疫细胞的功效,该前药被工程化免疫细胞转化为细胞毒性药物,该细胞毒性药物导致靶位点处的细胞(例如,肿瘤细胞)的细胞死亡但不导致工程化细胞本身死亡,因此提供了工程化免疫细胞在患者中保持活力和活性。对t细胞无毒性的药物的一个实例是厄洛替尼。如实施例1中所述的,生成厄洛替尼的前药形式。
图14提供了显示人t细胞而非癌细胞对厄洛替尼有抵抗的示例性数据。用递增浓度的受体酪氨酸激酶抑制剂厄洛替尼处理人jurkatt细胞、原代t细胞和人非小细胞肺癌细胞pc9和hcc827。如图中所示的,相较于显示出在药物的存在下的显著细胞死亡的pc9和hcc827,原代t细胞和t细胞系表现出对递增浓度的厄洛替尼有抵抗(图14a)。然而,除非由cpg2激活,否则谷氨酸化的厄洛替尼前药(p-厄洛替尼)在pc9细胞(1μm)中未表现出毒性(图14b)。在厄洛替尼的存在下、在p-厄洛替尼的存在下,在p--厄洛替尼和经纯化的cpg2的存在下,孵育pc9细胞。在48小时处通过celltitre-
将以上所述的编码抗cd19car和跨膜cpg2酶的逆转录病毒构建体转导至293t细胞中以测试跨膜cpg2对厄洛替尼前药的转化活性。gfp+293tcpgtm细胞与肺腺癌细胞(hcc827)以多种比率(293tcpgtm:hcc827)共培养,且添加厄洛替尼前药(p-厄洛替尼)。在120小时时通过流失细胞术分析细胞(图15a),并计算每种细胞比率的gfp细胞的百分比和293tcpgtm细胞与hcc827细胞的比率(图15b)。图15显示了经转导的细胞中的跨膜cpg2是有功能的,并且将前药转化为杀伤靶癌细胞但不杀伤分泌该酶的细胞的活性药物。
实施例8.每种seaker细胞在体外对多种癌症靶标的功效。
一旦产生了每种seaker格式,且每种seaker格式是稳定的,则使用先前公开的方法和细胞,生成使用不同供体的cart细胞,并测试其抗癌细胞系(阳性和阴性、新鲜癌细胞和正常pbmc)的活性(brentjens等,scitransmed5(177):177ra38(2013);dao等,scitransmed13(5):ra33(2013))。如先前对cart细胞所描述的,通过检测seaker细胞的癌细胞系特异性细胞毒性、细胞因子分泌和增殖功能而在体外分析多种seaker重定向t细胞功能的能力(curran等,moleculartherapy:jamsocgenether.23(4):769-78(2015))。
使用标准测定法测量经转化的药物介导的细胞毒性(参见,例如,图5-7)。使用标准的铬释放测定法测量针对一组cd19+、wt1+、prame+或阴性肿瘤细胞的特异性t细胞细胞毒性(参见,例如,lasek等cancerimmunology,immunotherapy:cii63(5):419-35(2014);dao等,scitransmed13(5):ra33(2013);pinilla-ibarz等,leukemia20(11):2025-33(2006))。
通过收集来自seaker细胞和prame+或prame-肿瘤细胞的24小时共培养物的上清液来测量特异性细胞因子分泌。使用luminex技术分析细胞因子的存在。
如先前出版的(koneru等,oncoimmunology4(3):e994446(2015)),通过将转导的t细胞与抗原+或-肿瘤细胞共培养,并使用计数珠用流式细胞术监测t细胞扩增,来分析cd3-28zcar刺激t细胞增殖的能力。经转导以表达靶向无关抗原的car的t细胞用作对照。
还测定了cart细胞,以确定相对于单独的cart,药物作用加上cart细胞毒性是否具有累加或协同杀伤。将抗原阳性和阴性靶细胞混合以测量差异杀伤。创建了通过缺失4-1bb和cd3信号传导而在结合靶标(“无效”或较弱激活性第一代car)后并不激活的cart。在体外测定和比较构建体,其中需要增殖以评定癌细胞处信号的扩增。还通过使用以下方法从药物作用中分离出t细胞细胞毒性:反式孔实验(transwellexperiments)并且分离向其中转移了来自seaker细胞的培养基的所述孔或多个孔中的抗原阳性和抗原阴性细胞靶标的“顺式相对反式”杀伤,目的是评定单独的药物在多种构建体中的作用以及例如以比较膜结合形式相对分泌形式。
实施例9.seaker细胞系统在动物模型中的功效和pk。
在体外评定seaker细胞活性后,评定了seaker细胞在多种癌症的动物模型中的体内功效。还评定了前药和药物在正常和瘤组织中的药代动力学(pk)。在相关动物模型中评定了每种前药/药物组合的功效、毒性、pk和生物分布。还评定了模型中每种seaker细胞类型和药物系统的功效、毒性、pk、治疗指数和生物分布。
使用临床前异种鼠模型评定了seaker细胞在体内根除肿瘤的能力。例如,将scid-beige或nsg小鼠接种肿瘤细胞(iv用于血液系统恶性肿瘤,或ip或sc用于实体瘤),所述肿瘤细胞被修饰以表达萤火虫萤光素酶以允许生物发光成像(图16)(参见,例如,koneru等oncoimmunology4(3):e994446(2015);lasek等,cancerimmunology,immunotherapy:cii.63(5):419-35(2014);和pegram等,blood119(18):4133-41(2012))。待测定的示例性抗原阳性肿瘤细胞系(取决于seaker细胞特异性)包括set2aml、bv173all、aml14,mac1t淋巴瘤、乳腺癌mda-231或卵巢癌ovcar3系、结肠sw480。这些细胞已经在小鼠中增殖。
在肿瘤细胞接种后,随后用seaker或对照细胞(例如,表达靶向无关抗原的car)的全身输注处理小鼠。如先前所述(santos等,naturemedicine15(3):338-44(2009)),通过临床和生物发光成像监测疾病进展。通过收集来自经处理的小鼠的外周血和检测seaker细胞的流式细胞术来确定seaker细胞的持久性。通过使用luminex技术检测经处理的小鼠血清中的细胞因子来确定seaker随时间的功能。通过lc-ms/ms测定法测量血清、靶标和脱靶正常组织中的药物和前药水平。还比较了中靶和脱靶标组织中药物水平与每个seaker的比较。
正常小鼠和荷瘤小鼠中seaker细胞和药物的pk分析和生物分布。当seaker细胞在肿瘤细胞部位达到峰值浓度时,预期前药的施用时间是最佳的。用发光探针标记seaker细胞,以允许对其动力学和小鼠中的生物分布进行实时成像。最初在施用后0.08、0.25、0.5、1、2、4、8和24小时通过lcms/ms对血浆中单独的前药进行定量,以确定合适的剂量和方案(基于先前的前药血浆蛋白结合[c游离]和药物的体外细胞毒性[ic50]的确定)。随后,定量seaker预处理的小鼠中的血浆和肿瘤中前药和药物浓度,以将药物释放和清除与观察到的抗肿瘤功效相关联。
小鼠中的毒性。每天跟踪健康和异种移植的小鼠,对5种临床毒性症状(按照iacuc方案)评分,每周对体重增加或减轻评分。在选择的小鼠中评定外周血细胞计数,并在处死时分析骨髓、脾脏、肾脏和肝脏的病理。进行抗癌活性与临床毒性的比较。
实施例10.供在seaker细胞技术中使用的“杀伤开关”前药系统的设计和分析
此实施例描述了与工程化细胞质cpg2酶组合的car的seaker细胞的构建,该细胞质cpg2酶起“杀伤开关”的作用,以激活穿透细胞的前药,该前药选择性破坏所述工程化免疫细胞,但不破坏不表达细胞质cpg2酶的细胞。(图17)
过继性t细胞方法(包括t细胞和cart细胞)的强大效能,外加它们在体内快速增殖的能力,伴随着在患者中的严重毒性和死亡。本文所述的策略允许在毒性出现或需要体内减少细胞的情况下安全关闭施用的工程免疫细胞。cpg2酶在工程化免疫细胞的细胞质中表达。在输注细胞后,可以将无毒的细胞穿透性前药施用于患者,其中仅在工程化免疫细胞中该前药被转化为细胞毒性药物,而在受试者中的其他位置则不被转化。此外,因为可以以各种剂量和时间表施用前药,所以可以根据需要调节关闭作用的调节和效应细胞的重新出现。在一些实例中,工程化免疫细胞在细胞内(例如,在细胞质中)表达cpg2,并且还分泌或在表面上表达cpg2。在将这些工程化免疫细胞施用于受试者后,细胞将经由表达肿瘤抗原特异性受体(诸如嵌合抗原受体)而定位于肿瘤部位(参见图17b)。然后施用非穿透性前药以激活癌症靶标的杀伤。然后,在需要时施用细胞穿透性前药来杀伤工程化免疫细胞。
制备了初始构建体(参见图17a),其表达嵌合抗原受体和细胞内形式的cpg2酶。如图18所示,从此构建体表达的cpg2在细胞内表达而不是分泌的。
测定了对于杀伤来自多个供体的293t细胞和seaker或cart细胞的数个对数的剂量反应关系。最初的研究是使用ams前-前药1d(图1)进行的,其中将谷氨酸盐部分掩盖为相应的双-pom酯,以使其能够穿透细胞,然后去除细胞酯酶,使pom基团露出谷氨酸盐部分(1a)用于被细胞内cpg2切割,在细胞中选择性释放活性药物(1a)。通过lc-ms/ms定量上清液和细胞裂解液中的前-前药、前药和药物浓度,以评定细胞穿透和前药切割率。用几种前药的每一种杀伤工程化免疫细胞的剂量范围预期在体内模型可达到的范围内。还在体外评估了1-4天的杀伤程度和时间。
首先测试没有肿瘤的小鼠模型,以确定细胞是否被根除或减少以及在什么时间范围内。实时成像用于对施用前药的作用进行定量。
然后评定荷瘤模型的工程化免疫细胞生长的改变(例如,由于增殖)和前药的pk。监测和测定对动物的毒性,以优化细胞穿透性前药的施用时间。
实施例11
此实施例描述了跨膜和分泌形式的cpg和/或β-内酰胺酶的生成及其在哺乳动物细胞中表达时的特征。在此示例性实施方案中,合成了几种细胞毒性药物,其携带通过自我杀灭部分连接的末端谷氨酸盐掩蔽残基,该残基在谷氨酸盐裂解后衰变,导致活性细胞毒素的形成(图20a)。这些前药包括掩蔽形式的核苷类似物ams(glut-ams)、烷基化氮芥(zdp275)和被靶向的激酶的抑制剂厄洛替尼(glut-厄洛替尼)(图20b)。生成针对一组癌细胞系和原代细胞系的每对药物的ic50值,以确定可用新的前药平台实现的有效治疗指数。
最初合成、纯化重组cpg2蛋白(图21a和21b),并在谷氨酸盐释放测定中与递增浓度的前药一起孵育(图20c)。质谱分析法用于确定谷氨酸盐部分从所有谷氨酸盐前药中释放。使用celltitre
已确定ams前药对于所有测试的细胞类型均表现出高(>100)选择指数,并且在人前列腺细胞中表达的内源性谷氨酸羧肽酶的存在下保持稳定,这确认了其对cpg2羧肽酶的特异性(图21c)。选择此前药/药物组合用于进一步开发seaker系统。
此实施例描述了跨膜和分泌形式的cpg的生成及其在细菌或哺乳动物细胞中表达时的特征。接下来,证明了cpg2可以在保持酶功能且不损害宿主细胞的同时在真核细胞中被遗传编码和表达。hek293t细胞的瞬时转染证明了cpg2耐受各种结构修饰,包括添加ha表位标签、限制至细胞质、系链至细胞表面或通过分泌系统规定路线以允许分泌到细胞外环境中(图22a和22b)。免疫印迹分析确定将cd8胞质尾巴添加到cpg2的c端增加了分泌(图23a)。
图22c和23b-23c证明了在hek293t细胞中表达的分泌形式的cpg2在体外具有酶活性。将set2细胞暴露于浓度递增的ams(正方形)或仅具有培养基的glut-ams(-对照,三角形)或具有来自分泌cpg2的hek293t细胞的上清液的glut-ams(圆形)。通过celltitre-
图23a证明了在用逆转录病毒转导的hek293t细胞中cpg2和β-内酰胺酶的稳定表达的免疫印迹分析,随后对其进行酶表达和功能分析。ha表位标签允许对表达分泌或跨膜形式的酶的细胞中的表面水平cpg2进行染色(图22d)。在细胞上清液以及在来自293tcpg2-分泌的的上清液的转移物中检测到酶,但是在表达其他形式的cpg2或gfp的对照细胞系中在前药的存在下没有检测到酶,该酶导致set2细胞死亡(图22e)。在48小时处通过celltitre-
然后将编码上述cpg2酶的逆转录病毒构建体引入t细胞系(例如,jurkat),以测量cpg2的表达并表征其活性。图22h显示免疫印迹分析,其证明cpg2酶经由逆转录病毒转导而稳定表达。
基于阴沟肠杆菌衍生的β-内酰胺酶(b-lac)以及由重组b-lac激活的一系列内酰胺前药(图21d),开发了第二种适应真核生物的前药激活酶。与cpg2一样,从稳定的293t细胞(293tβ-lac)和jurkat细胞(图21e)中生成并分泌b-lac。分泌形式的β内酰胺酶在稳定的细胞系中显示酶活性,并成功激活了多种内酰胺前药(图28c),这些内酰胺前药在多种细胞类型中具有高选择性指数(表3)。
表3.ams前药的细胞选择性数据
在人上皮caco-2细胞中的细胞通透性研究表明,glut-ams前药未表现出细胞通透性,因此在酶释放之前不会扩散到表达cpg2的细胞中(表4),然而,活性药物仍然可以在暴露后杀伤分泌性细胞。为了进一步研究此作用,进行了旁观者毒性测定,在该测定中在前药的存在下,将酶阳性细胞(效应细胞)与酶阴性细胞(靶细胞)共培养。前药处理后的细胞计数表明,增加的效应子/靶标比率导致更高的总细胞死亡,当将293tcpg2-sec/gfp或293tβ-lac/gfp与wt293t细胞以1:1比率共培养时,耗竭~80%的细胞(图24a)。细胞荧光分析揭示了gfp+(效应子)和gfp-(靶标)细胞二者的等效耗竭,其指示这两种细胞群(效应细胞和靶细胞)均被激活的前药破坏(图24b-24d)。
表4.前药在人上皮caco-2细胞中的细胞通透性研究。
为了将酶前药系统与现有的car-t细胞平台整合,创建了将cpg2或b-lac置于19bbzcar盒上游的构建体(图25a)。还开发了定量elisa以确定每种构建体的各自酶输出(图21f)。作为比较,还通过在先前公开的sfg逆转录病毒载体骨架中,在含有4-1bb共刺激元件(19bbz)的原型抗cd-19car的下游直接引入p2a自切割肽序列随后是cpg2,来克隆分泌的cpg2构建体(图26a)。从19bbz-cpg2car-t细胞分离的上清液不能有效地激活前药,因为,尽管在原代人t细胞中cpg2-sec-19bbz的转导水平高,与标准19bbzcar相比的有效的t细胞杀伤活性以及检测到细胞上清液中分泌的cpg2(图26),但用前药进行的上清液转移实验并未导致set2细胞死亡(图26)。出乎意料地,表达cpg-19bbz构建体的原代t细胞表达的cpg2水平比用19bbz-cpg2构建体转导的细胞高近20倍(图25b)。这些数据表明基因序列对于产生的酶的水平至关重要,并且如果需要,可以对系统的幅度进行第二水平的控制。针对所有随后的car-t细胞研究(包括beta-lac-19bbzcar),选择更高幅度配置。
在多种t细胞供体中以高效率转导了cpg-19bbz和β-lac-19bbz构建体(图25c),其维持了在wt19bbz细胞中观察到的等效的抗原依赖性细胞毒性作用(图25d)并分泌了高水平的每种单独酶(图25e)。进一步地,来自seakercar-t细胞(19bbz、cpg-19bbz和(β-lac-19bbz))的上清液在反式毒性测定中转化前药并杀伤set2细胞,这指示seaker细胞产生并分泌足够高水平的功能酶以达到目的(图25f)。通过cpg-19bbz和β-lac-19bbzt细胞的celltitre
为了确定经功能化以生成小分子药物的car-t细胞是否将消除抗原低或抗原阴性的细胞(抗原损失变体)以及cart细胞不直接啮合的细胞,在共培养实验中检查了seakert表达细胞cpg2或β-内酰胺酶以消除抗原阴性细胞的能力(图25h)。seakert细胞与cd19+raji细胞和cd19-set2细胞共培养。在共培养24小时后,向孔中添加glut-ams,且在另外的24小时孵育时期后测量总细胞活力(图25i)。尽管19bbzcar-t细胞仅消除cd19+raji细胞,但seaker细胞能够消除抗原+和抗原-细胞群二者。数据证明了功能化car-t细胞可消除抗原低或抗原阴性细胞。
此示例性实施方案将描述能够在体内消除抗原损失变体肿瘤模型中的肿瘤群体的seakercar-t细胞的生成。将开发一种“中靶(ontarget)”skov-3人卵巢癌细胞系,其将表达人cd19胞外域和mcherry报告分子(skov-3cd19-ecto/mcherry),且“脱靶”系表达萤火虫萤光素酶和gfp报告分子(skov-3luc/gfp)。细胞活力研究将证明skov-3细胞将对活性ams药物而非glut-ams前药的杀伤敏感。skov-3细胞系与seakert细胞的共培养实验应证明,尽管标准19bbzcart细胞或seakert细胞在不存在前药的情况下将仅耗竭skov-3cd19-ecto/mcherry细胞,但将消除具有含有glut-ams前药的seakert细胞的共培养物中的两种skov-3细胞类型。
然后将skov-3细胞适应于体内混合的肿瘤模型,其中将以1:1比率的skov-3cd19-ecto/mcherry和skov-3luc/gfpip植入nsg小鼠的ip腔。植入后5天,将通过ip注射施用19bbzcart细胞或seakercart细胞。先前在nod小鼠中进行的药代动力学和生物分布研究表明,glut-ams前药是无毒的,且被快速清除,半衰期为30-40分钟(图31)。不希望受到理论的束缚,认为高前药浓度的每日ip给药将被充分转化为ams。t细胞注射后三天,将连续3天每天经由ip注射来施用glut-ams前药。seakercar-t+glut-ams部门中的小鼠应证明明显提高的存活率。
生物发光成像研究应证明在接受seakercar-t细胞加前药的小鼠中luc+脱靶肿瘤的消除,但在不接受前药或标准19bbzcar-t细胞以及前药的小鼠中不消除luc+脱靶肿瘤。从小鼠收获的肿瘤的流式细胞术分析可证明在接受seaker-t细胞+前药的小鼠中两种肿瘤类型的耗竭,而来自其他实验组的小鼠中则没有。elisa分析将用于确认来自接受seakercart细胞的小鼠ip腔中液体中的高水平cpg2。串联质谱将用于鉴定来自seaker-t细胞+前药小鼠的ip液体中活性ams的存在,这将证实分泌的cpg2成功激活了前药。此数据证明,通过消除抗原逃逸的实体肿瘤模型中的中靶肿瘤细胞和脱靶肿瘤细胞,seakert细胞具有增强的功能。
实施例12
此实施例描述了跨膜和分泌形式的β-内酰胺酶的生成及其在哺乳动物细胞中表达时的特征。在此示例性实施方案中,hek293t细胞的瞬时转染证明了β-内酰胺酶耐受多种结构修饰,包括指定酶通过分泌系统的路线以允许分泌到细胞外环境中(图27a和27b)。表达多种构建体(cd8-β-lac或β-lac-trunc)的hek293t细胞的上清液的免疫印迹分析指示仅截短形式的β-内酰胺酶(β-lac-trunc)的表达,而不表达膜连接形式的β-内酰胺酶(cd8-β-lac)(图27b)。来自表达多种构建体(cd8-β-lac或β-lac-trunc)的hek293t细胞的上清液中的β-内酰胺酶活性的头孢硝噻吩分析确认了,相较于来自对照hek293t细胞的上清液或经纯化的重组β-内酰胺酶,来自hek293t细胞的β-内酰胺酶的表达和分泌(图27c)。
为了确定相对于先前cpg2构建体的β-内酰胺酶构建体的各自的酶输出,定量夹心elisa用于确定纯化的重组cpg2的浓度(ng/ml),定量头孢硝噻吩测定法用于确定纯化的重组β-内酰胺酶的浓度(ng/ml)(图28a)。在前药(glut-ams或ceph-ams)和不同浓度的纯化的重组cpg2或β-内酰胺酶的存在下,通过celltitre-
为了评定表达cpg2和β-内酰胺酶的多种构建体的hek293t的顺式毒性,实施了celltitre
使用将cpg2或b-lac置于19bbzcar盒上游的构建体,将酶前药系统并入现有的car-t细胞平台中。
为了确定功能化以产生小分子药物的car-t细胞是否会消除抗原低或抗原阴性的细胞(抗原损失变体)以及cart细胞不直接啮合的细胞,在共培养实验中检查了seakert表达细胞表达cpg2以消除抗原阴性细胞的能力(图29a-29c)。seakert细胞(cpg-19bbz)与(gfp-/cd19+)raji细胞和(gfp+/cd19-)set2细胞共培养。如图29b中所示的,在72的共培养后,用或不用glut-ams前药处理共培养物。利用流式细胞术分析脱靶的命中(set2),其在图29c中定量。图30a证明了β-lac-19bbzt细胞与raji(cd19+)或set2(cd19-)细胞在具有或无ceph-ams前药的情况下共培养72小时后的raji(cd19+)、set2(cd19-)和β-lac-19bbzt细胞的总细胞活力。图30b证明了在ceph-ams前药的存在下,β-lac-19bbzt细胞能够在萤光素酶测定中以约0.1:1的比率诱导表达萤光素酶的raji细胞(raji/luc;cd19+/luc+)的细胞裂解。
实施例13
方案5.碘化的头孢噻吩(cephalothin)衍生物的生成
方案6.本技术的示例性β-内酰胺酶前药前体的未优化生成
然后可以将方案6的示例性β-内酰胺酶前药前体脱保护(诸如通过hf·吡啶和适当的烯丙基脱保护条件诸如pd(0)-催化的脱保护)以提供以下方案7中所示的示例性β-内酰胺酶前药。greene等(1991)protectivegroupsinorganicsynthesis,第3版.(johnwiley&sons,inc.,newyork),其通过提述并入本文,其提供示例性脱保护步骤。
方案7
方案8.本技术的示例性的含有氨基甲酸酯的β-内酰胺酶前药前体的未优化生成
还生成了以下所示的亚砜变体,其中可以利用方案8中描述的类似合成来生成此亚砜变体。
然后可将方案7的示例性β-内酰胺酶前药前体和上述亚砜变体脱保护(诸如通过hf·吡啶和适当的烯丙基脱保护条件诸如pd(0)-催化的脱保护)以提供以下方案9中所示的示例性β-内酰胺酶前药。
方案9
方案10.本技术的示例性的含有paba的β-内酰胺酶前药前体的未优化生成
方案10中所示的前体可经由mitsunobu型反应与,例如
方案11.
序列表
<110>memorialsloankettering
<120>用于癌症的过继性细胞疗法的组合物和方法
<130>115872-0384
<150>62/527,932
<151>2017-06-30
<160>53
<170>patentin版本3.5
<210>1
<211>415
<212>prt
<213>智人
<400>1
metargproserilehisargthralailealaalavalleualathr
151015
alaphevalalaglythralaleualaglnlysargaspasnvalleu
202530
pheglnalaalathraspgluglnproalavalilelysthrleuglu
354045
lysleuvalasnilegluthrglythrglyaspalagluglyileala
505560
alaalaglyasnpheleuglualagluleulysasnleuglyphethr
65707580
valthrargserlysseralaglyleuvalvalglyaspasnileval
859095
glylysilelysglyargglyglylysasnleuleuleumetserhis
100105110
metaspthrvaltyrleulysglyileleualalysalaprophearg
115120125
valgluglyasplysalatyrglyproglyilealaaspasplysgly
130135140
glyasnalavalileleuhisthrleulysleuleulysglutyrgly
145150155160
valargasptyrglythrilethrvalleupheasnthraspgluglu
165170175
lysglyserpheglyserargaspleuileglngluglualalysleu
180185190
alaasptyrvalleuserphegluprothrseralaglyaspglulys
195200205
leuserleuglythrserglyilealatyrvalglnvalasnilethr
210215220
glylysalaserhisalaglyalaalaprogluleuglyvalasnala
225230235240
leuvalglualaseraspleuvalleuargthrmetasnileaspasp
245250255
lysalalysasnleuargpheasntrpthrilealalysalaglyasn
260265270
valserasnileileproalaseralathrleuasnalaaspvalarg
275280285
tyralaargasngluasppheaspalaalametlysthrleugluglu
290295300
argalaglnglnlyslysleuproglualaaspvallysvalileval
305310315320
thrargglyargproalapheasnalaglygluglyglylyslysleu
325330335
valasplysalavalalatyrtyrlysglualaglyglythrleugly
340345350
valglugluargthrglyglyglythraspalaalatyralaalaleu
355360365
serglylysprovalilegluserleuglyleuproglypheglytyr
370375380
hisserasplysalaglutyrvalaspileseralaileproargarg
385390395400
leutyrmetalaalaargleuilemetaspleuglyalaglylys
405410415
<210>2
<211>1248
<212>dna
<213>智人
<400>2
atgcgtccctctatccatcgtacggccattgcggctgtattagccacagcgtttgtagcg60
ggcacggccttggctcagaaacgtgacaacgtactgttccaagcagctactgacgagcaa120
cccgccgttattaaaacattagagaaattggtaaatattgaaactgggacgggcgacgct180
gaaggcattgccgcggctgggaacttcttggaagccgaattaaaaaatcttgggtttaca240
gtcactcgctcgaaatcagcggggctggtcgttggtgataacatcgtcggcaaaattaaa300
gggcgcgggggtaaaaatttattgttgatgagccacatggacaccgtttacctgaaaggg360
atccttgcgaaagccccctttcgcgtcgagggagacaaggcttacggtcccggcatcgca420
gatgataaaggtggaaatgcagtgattttacacacactgaaattgcttaaagagtacgga480
gttcgcgattacggcacgatcacagtattgttcaatacggacgaggaaaagggatctttc540
ggcagccgtgatcttatccaggaagaagcgaaactggcggattatgtgttaagttttgaa600
cccacaagcgccggagatgagaagctttcattaggtacatcaggtattgcctacgtacag660
gtaaatatcacaggtaaagcaagccacgcaggggcggcaccagagttaggcgttaatgca720
ctggtagaagctagtgacctggtccttcgtactatgaatatcgatgataaagcgaaaaac780
ttgcgctttaactggacgatcgccaaggcgggtaatgtatcgaacattattccagccagt840
gcgacattaaatgctgatgttcgttacgcgcgcaatgaagacttcgacgctgctatgaaa900
acacttgaggagcgcgctcaacagaaaaaacttccagaggccgacgtaaaagttatcgtt960
acacgtggacgtccggcctttaacgcgggtgaaggaggcaagaagctggtagacaaagct1020
gttgcttattataaagaagcggggggaaccttgggagtggaggaacgtaccggtggaggg1080
actgatgcggcttacgccgcactttctggaaagcctgtaatcgagagcttaggtttaccc1140
ggttttgggtatcacagtgataaagcagagtacgtcgatatttcagcgatcccacgccgt1200
ctttatatggctgcccgcttgattatggatttgggggcggggaagtag1248
<210>3
<211>393
<212>prt
<213>智人
<400>3
alaleualaglnlysargaspasnvalleupheglnalaalathrasp
151015
gluglnproalavalilelysthrleuglulysleuvalasnileglu
202530
thrglythrglyaspalagluglyilealaalaalaglyasnpheleu
354045
glualagluleulysasnleuglyphethrvalthrargserlysser
505560
alaglyleuvalvalglyaspasnilevalglylysilelysglyarg
65707580
glyglylysasnleuleuleumetserhismetaspthrvaltyrleu
859095
lysglyileleualalysalapropheargvalgluglyasplysala
100105110
tyrglyproglyilealaaspasplysglyglyasnalavalileleu
115120125
histhrleulysleuleulysglutyrglyvalargasptyrglythr
130135140
ilethrvalleupheasnthraspgluglulysglyserpheglyser
145150155160
argaspleuileglngluglualalysleualaasptyrvalleuser
165170175
phegluprothrseralaglyaspglulysleuserleuglythrser
180185190
glyilealatyrvalglnvalglnilethrglylysalaserhisala
195200205
glyalaalaprogluleuglyvalasnalaleuvalglualaserasp
210215220
leuvalleuargthrmetasnileaspasplysalalysasnleuarg
225230235240
pheglntrpthrilealalysalaglyglnvalserasnileilepro
245250255
alaseralathrleuasnalaaspvalargtyralaargasngluasp
260265270
pheaspalaalametlysthrleuglugluargalaglnglnlyslys
275280285
leuproglualaaspvallysvalilevalthrargglyargproala
290295300
pheasnalaglygluglyglylyslysleuvalasplysalavalala
305310315320
tyrtyrlysglualaglyglythrleuglyvalglugluargthrgly
325330335
glyglythraspalaalatyralaalaleuserglylysprovalile
340345350
gluserleuglyleuproglypheglytyrhisserasplysalaglu
355360365
tyrvalaspileseralaileproargargleutyrmetalaalaarg
370375380
leuilemetaspleuglyalaglylys
385390
<210>4
<211>1182
<212>dna
<213>智人
<400>4
gccctcgcgcaaaaacgggacaatgtattgttccaagcggctactgatgaacagccagca60
gtaattaaaacgcttgaaaaacttgtcaacatcgagaccgggacaggggatgctgaaggg120
attgcagccgcaggaaactttcttgaagccgaactcaaaaaccttgggtttaccgtaacg180
cgctctaagtcagccggtttggtcgtaggtgacaacattgtgggaaagatcaagggacgg240
ggcggtaaaaacttgcttttgatgtctcatatggatactgtatatcttaaggggatcctc300
gcgaaggcgccctttcgggtggaaggcgataaggcatatggaccagggatagctgacgac360
aaggggggtaacgcggtcattctccacacactgaaacttcttaaagaatacggtgtgaga420
gattatgggacgatcactgtactgttcaacaccgatgaggaaaagggctcttttggctct480
agagatttgattcaagaagaagcgaaactggcagactacgtgttgtccttcgaacccaca540
tctgctggcgatgagaagctcagtcttggaacctctgggattgcgtacgtccaggtccaa600
ataaccggaaaggcaagccatgctggagcagccccagaacttggcgtaaatgcgctggtg660
gaggcgagtgatctggtgctccggaccatgaacatagatgacaaagccaaaaatctgagg720
tttcaatggaccatcgctaaagcagggcaggtctctaatatcatcccggcttcagccact780
ttgaacgccgatgttaggtacgcccggaatgaagattttgatgccgctatgaaaacattg840
gaggaacgagcgcagcaaaaaaagctgccagaagccgatgttaaagtaatagtaactaga900
ggacgacccgcgttcaatgcgggcgagggtggaaagaagttggtcgataaggctgttgcg960
tactataaggaagcggggggtacgcttggagttgaggagaggaccggagggggcactgac1020
gcggcgtacgccgccctctcaggcaagcccgtaattgagtcacttgggcttcctggtttt1080
ggttatcactcagacaaggccgagtacgtggatatatcagctatccctcggcgattgtac1140
atggcggcacgcttgataatggatttgggggctggaaagtaa1182
<210>5
<211>415
<212>prt
<213>智人
<400>5
metargproserilehisargthralailealaalavalleualathr
151015
alaphevalalaglythralaleualaglnlysargaspasnvalleu
202530
pheglnalaalathraspgluglnproalavalilelysthrleuglu
354045
lysleuvalasnilegluthrglythrglyaspalagluglyileala
505560
alaalaglyasnpheleuglualagluleulysasnleuglyphethr
65707580
valthrargserlysseralaglyleuvalvalglyaspasnileval
859095
glylysilelysglyargglyglylysasnleuleuleumetserhis
100105110
metaspthrvaltyrleulysglyileleualalysalaprophearg
115120125
valgluglyasplysalatyrglyproglyilealaaspasplysgly
130135140
glyasnalavalileleuhisthrleulysleuleulysglutyrgly
145150155160
valargasptyrglythrilethrvalleupheasnthraspgluglu
165170175
lysglyserpheglyserargaspleuileglngluglualalysleu
180185190
alaasptyrvalleuserphegluprothrseralaglyaspglulys
195200205
leuserleuglythrserglyilealatyrvalglnvalglnilethr
210215220
glylysalaserhisalaglyalaalaprogluleuglyvalasnala
225230235240
leuvalglualaseraspleuvalleuargthrmetasnileaspasp
245250255
lysalalysasnleuargpheglntrpthrilealalysalaglygln
260265270
valserasnileileproalaseralathrleuasnalaaspvalarg
275280285
tyralaargasngluasppheaspalaalametlysthrleugluglu
290295300
argalaglnglnlyslysleuproglualaaspvallysvalileval
305310315320
thrargglyargproalapheasnalaglygluglyglylyslysleu
325330335
valasplysalavalalatyrtyrlysglualaglyglythrleugly
340345350
valglugluargthrglyglyglythraspalaalatyralaalaleu
355360365
serglylysprovalilegluserleuglyleuproglypheglytyr
370375380
hisserasplysalaglutyrvalaspileseralaileproargarg
385390395400
leutyrmetalaalaargleuilemetaspleuglyalaglylys
405410415
<210>6
<211>1248
<212>dna
<213>智人
<400>6
atgcgaccgagtatccacagaacagcaatagctgctgtgcttgcaacagcgtttgtagcg60
ggcacggccctcgcgcaaaaacgggacaatgtattgttccaagcggctactgatgaacag120
ccagcagtaattaaaacgcttgaaaaacttgtcaacatcgagaccgggacaggggatgct180
gaagggattgcagccgcaggaaactttcttgaagccgaactcaaaaaccttgggtttacc240
gtaacgcgctctaagtcagccggtttggtcgtaggtgacaacattgtgggaaagatcaag300
ggacggggcggtaaaaacttgcttttgatgtctcatatggatactgtatatcttaagggg360
atcctcgcgaaggcgccctttcgggtggaaggcgataaggcatatggaccagggatagct420
gacgacaaggggggtaacgcggtcattctccacacactgaaacttcttaaagaatacggt480
gtgagagattatgggacgatcactgtactgttcaacaccgatgaggaaaagggctctttt540
ggctctagagatttgattcaagaagaagcgaaactggcagactacgtgttgtccttcgaa600
cccacatctgctggcgatgagaagctcagtcttggaacctctgggattgcgtacgtccag660
gtccaaataaccggaaaggcaagccatgctggagcagccccagaacttggcgtaaatgcg720
ctggtggaggcgagtgatctggtgctccggaccatgaacatagatgacaaagccaaaaat780
ctgaggtttcaatggaccatcgctaaagcagggcaggtctctaatatcatcccggcttca840
gccactttgaacgccgatgttaggtacgcccggaatgaagattttgatgccgctatgaaa900
acattggaggaacgagcgcagcaaaaaaagctgccagaagccgatgttaaagtaatagta960
actagaggacgacccgcgttcaatgcgggcgagggtggaaagaagttggtcgataaggct1020
gttgcgtactataaggaagcggggggtacgcttggagttgaggagaggaccggagggggc1080
actgacgcggcgtacgccgccctctcaggcaagcccgtaattgagtcacttgggcttcct1140
ggttttggttatcactcagacaaggccgagtacgtggatatatcagctatccctcggcga1200
ttgtacatggcggcacgcttgataatggatttgggggctggaaagtaa1248
<210>7
<211>458
<212>prt
<213>智人
<400>7
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargproalaleualaglnlysargaspasnvalleuphe
202530
glnalaalathraspgluglnproalavalilelysthrleuglulys
354045
leuvalasnilegluthrglythrglyaspalagluglyilealaala
505560
alaglyasnpheleuglualagluleulysasnleuglyphethrval
65707580
thrargserlysseralaglyleuvalvalglyaspasnilevalgly
859095
lysilelysglyargglyglylysasnleuleuleumetserhismet
100105110
aspthrvaltyrleulysglyileleualalysalapropheargval
115120125
gluglyasplysalatyrglyproglyilealaaspasplysglygly
130135140
asnalavalileleuhisthrleulysleuleulysglutyrglyval
145150155160
argasptyrglythrilethrvalleupheasnthraspgluglulys
165170175
glyserpheglyserargaspleuileglngluglualalysleuala
180185190
asptyrvalleuserphegluprothrseralaglyaspglulysleu
195200205
serleuglythrserglyilealatyrvalglnvalglnilethrgly
210215220
lysalaserhisalaglyalaalaprogluleuglyvalasnalaleu
225230235240
valglualaseraspleuvalleuargthrmetasnileaspasplys
245250255
alalysasnleuargpheglntrpthrilealalysalaglyglnval
260265270
serasnileileproalaseralathrleuasnalaaspvalargtyr
275280285
alaargasngluasppheaspalaalametlysthrleuglugluarg
290295300
alaglnglnlyslysleuproglualaaspvallysvalilevalthr
305310315320
argglyargproalapheasnalaglygluglyglylyslysleuval
325330335
asplysalavalalatyrtyrlysglualaglyglythrleuglyval
340345350
glugluargthrglyglyglythraspalaalatyralaalaleuser
355360365
glylysprovalilegluserleuglyleuproglypheglytyrhis
370375380
serasplysalaglutyrvalaspileseralaileproargargleu
385390395400
tyrmetalaalaargleuilemetaspleuglyalaglylystyrpro
405410415
tyraspvalproasptyralaglyglyglyleutyrcysasnhisarg
420425430
asnargargargvalcyslyscysproargprovalvallyssergly
435440445
asplysproserleuseralaargtyrval
450455
<210>8
<211>1377
<212>dna
<213>智人
<400>8
atggccttgcccgttaccgcactcctgctgccgctggcgctgttgctgcacgcagctcga60
ccggccctcgcgcaaaaacgggacaatgtattgttccaagcggctactgatgaacagcca120
gcagtaattaaaacgcttgaaaaacttgtcaacatcgagaccgggacaggggatgctgaa180
gggattgcagccgcaggaaactttcttgaagccgaactcaaaaaccttgggtttaccgta240
acgcgctctaagtcagccggtttggtcgtaggtgacaacattgtgggaaagatcaaggga300
cggggcggtaaaaacttgcttttgatgtctcatatggatactgtatatcttaaggggatc360
ctcgcgaaggcgccctttcgggtggaaggcgataaggcatatggaccagggatagctgac420
gacaaggggggtaacgcggtcattctccacacactgaaacttcttaaagaatacggtgtg480
agagattatgggacgatcactgtactgttcaacaccgatgaggaaaagggctcttttggc540
tctagagatttgattcaagaagaagcgaaactggcagactacgtgttgtccttcgaaccc600
acatctgctggcgatgagaagctcagtcttggaacctctgggattgcgtacgtccaggtc660
caaataaccggaaaggcaagccatgctggagcagccccagaacttggcgtaaatgcgctg720
gtggaggcgagtgatctggtgctccggaccatgaacatagatgacaaagccaaaaatctg780
aggtttcaatggaccatcgctaaagcagggcaggtctctaatatcatcccggcttcagcc840
actttgaacgccgatgttaggtacgcccggaatgaagattttgatgccgctatgaaaaca900
ttggaggaacgagcgcagcaaaaaaagctgccagaagccgatgttaaagtaatagtaact960
agaggacgacccgcgttcaatgcgggcgagggtggaaagaagttggtcgataaggctgtt1020
gcgtactataaggaagcggggggtacgcttggagttgaggagaggaccggagggggcact1080
gacgcggcgtacgccgccctctcaggcaagcccgtaattgagtcacttgggcttcctggt1140
tttggttatcactcagacaaggccgagtacgtggatatatcagctatccctcggcgattg1200
tacatggcggcacgcttgataatggatttgggggctggaaagtatccttatgatgttcca1260
gactacgcagggggcggactgtattgtaaccaccgaaatcgacggcgagtatgcaagtgc1320
cctcgaccagttgttaaatctggggataaaccttcactgtccgctagatacgtgtaa1377
<210>9
<211>22
<212>prt
<213>智人
<400>9
metargproserilehisargthralailealaalavalleualathr
151015
alaphevalalaglythr
20
<210>10
<211>66
<212>dna
<213>智人
<400>10
atgcgaccgagtatccacagaacagcaatagctgctgtgcttgcaacagcgtttgtagcg60
ggcacg66
<210>11
<211>21
<212>prt
<213>智人
<400>11
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargpro
20
<210>12
<211>63
<212>dna
<213>智人
<400>12
atggccttgcccgttaccgcactcctgctgccgctggcgctgttgctgcacgcagctcga60
ccg63
<210>13
<211>32
<212>prt
<213>智人
<400>13
leutyrcysasnhisargasnargargargvalcyslyscysproarg
151015
provalvallysserglyasplysproserleuseralaargtyrval
202530
<210>14
<211>99
<212>dna
<213>智人
<400>14
ctgtattgtaaccaccgaaatcgacggcgagtatgcaagtgccctcgaccagttgttaaa60
tctggggataaaccttcactgtccgctagatacgtgtaa99
<210>15
<211>31
<212>prt
<213>智人
<400>15
leutyrcysasnhisargasnargargargvalglyglyglyargpro
151015
valvallysserglyasplysproserleuseralaargtyrval
202530
<210>16
<211>96
<212>dna
<213>智人
<400>16
ctgtattgtaaccaccgaaatcgacggcgagtaggaggaggacgaccagttgttaaatct60
ggggataaaccttcactgtccgctagatacgtgtaa96
<210>17
<211>22
<212>prt
<213>智人
<400>17
glyserglyalathrasnpheserleuleulysglnalaglyaspval
151015
glugluasnproglypro
20
<210>18
<211>66
<212>dna
<213>智人
<400>18
ggaagcggagctactaacttcagcctgctgaagcaggctggagacgtggaggagaaccct60
ggacct66
<210>19
<211>245
<212>prt
<213>智人
<400>19
gluvallysleuglnglnserglyalagluleuvalargproglyser
151015
servallysilesercyslysalaserglytyralaphesersertyr
202530
trpmetasntrpvallysglnargproglyglnglyleuglutrpile
354045
glyglniletyrproglyaspglyaspthrasntyrasnglylysphe
505560
lysglyglnalathrleuthralaasplysserserserthralatyr
65707580
metglnleuserglyleuthrsergluaspseralavaltyrphecys
859095
alaarglysthrileserservalvalaspphetyrpheasptyrtrp
100105110
glyglnglythrthrvalthrvalserserglyglyglyglysergly
115120125
glyglyglyserglyglyglyglyseraspilegluleuthrglnser
130135140
prolysphemetserthrservalglyaspargvalservalthrcys
145150155160
lysalaserglnasnvalglythrasnvalalatrptyrglnglnlys
165170175
proglyglnserprolysproleuiletyrseralathrtyrargasn
180185190
serglyvalproaspargphethrglyserglyserglythraspphe
195200205
thrleuthrilethrasnvalglnserlysaspleualaasptyrphe
210215220
cysglnglntyrasnargtyrprotyrthrserglyglyglythrlys
225230235240
leugluilelysarg
245
<210>20
<211>735
<212>dna
<213>智人
<400>20
gaggtgaagctgcagcagtctggggctgagctggtgaggcctgggtcctcagtgaagatt60
tcctgcaaggcttctggctatgcattcagtagctactggatgaactgggtgaagcagagg120
cctggacagggtcttgagtggattggacagatttatcctggagatggtgatactaactac180
aatggaaagttcaagggtcaagccacactgactgcagacaaatcctccagcacagcctac240
atgcagctcagcggcctaacatctgaggactctgcggtctatttctgtgcaagaaagacc300
attagttcggtagtagatttctactttgactactggggccaagggaccacggtcaccgtc360
tcctcaggtggaggtggatcaggtggaggtggatctggtggaggtggatctgacattgag420
ctcacccagtctccaaaattcatgtccacatcagtaggagacagggtcagcgtcacctgc480
aaggccagtcagaatgtgggtactaatgtagcctggtatcaacagaaaccaggacaatct540
cctaaaccactgatttactcggcaacctaccggaacagtggagtccctgatcgcttcaca600
ggcagtggatctgggacagatttcactctcaccatcactaacgtgcagtctaaagacttg660
gcagactatttctgtcaacaatataacaggtatccgtacacgtccggaggggggaccaag720
ctggagatcaaacgg735
<210>21
<211>15
<212>prt
<213>智人
<400>21
glyglyglyglyserglyglyglyglyserglyglyglyglyser
151015
<210>22
<211>45
<212>dna
<213>智人
<400>22
ggcggcggcggatctggaggtggtggctcaggtggcggaggctcc45
<210>23
<211>10
<212>prt
<213>智人
<400>23
alaleutyrvalaspserleuphepheleu
1510
<210>24
<211>214
<212>prt
<213>智人
<400>24
metleuargleuleuleualaleuasnleupheproserileglnval
151015
thrglyasnlysileleuvallysglnserprometleuvalalatyr
202530
aspasnalaleusercyslystyrsertyrasnleupheserargglu
354045
pheargalaserleuhislysglyleuaspseralavalgluvalcys
505560
trptyrglyasntyrserglnglnleuglnvaltyrserlysthrgly
65707580
pheasncysaspglylysleuglyasngluservalthrphetyrleu
859095
glnasnleutyrglnthraspiletyrphecyslysilegluvalmet
100105110
tyrproproprotyrleuaspasnglulysserasnglythrileile
115120125
hisvallysglylyshisleucysproserproleupheproglypro
130135140
serlysprophetrpvalleuvaltrpglyglyvalleualacystyr
145150155160
serleuleuvalthrvalalapheileilephetrpvalargserlys
165170175
argserargleuleuhisserasptyrmetasnmetthrproargarg
180185190
proglyprothrarglyshistyrglnprotyralaproproargasp
195200205
phealaalatyrargser
210
<210>25
<211>321
<212>dna
<213>智人
<400>25
attgaagttatgtatcctcctccttacctagacaatgagaagagcaatggaaccattatc60
catgtgaaagggaaacacctttgtccaagtcccctatttcccggaccttctaagcccttt120
tgggtgctggtggtggttggtggagtcctggcttgctatagcttgctagtaacagtggcc180
tttattattttctgggtgaggagtaagaggagcaggctcctgcacagtgactacatgaac240
atgactccccgccgccccgggcccacccgcaagcattaccagccctatgccccaccacgc300
gacttcgcagcctatcgctcc321
<210>26
<211>234
<212>prt
<213>智人
<400>26
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargproserglnpheargvalserproleuaspargthr
202530
trpasnleuglygluthrvalgluleulyscysglnvalleuleuser
354045
asnprothrserglycyssertrpleupheglnproargglyalaala
505560
alaserprothrpheleuleutyrleuserglnasnlysprolysala
65707580
alagluglyleuaspthrglnargpheserglylysargleuglyasp
859095
thrphevalleuthrleuserasppheargarggluasngluglytyr
100105110
tyrphecysseralaleuserasnserilemettyrpheserhisphe
115120125
valprovalpheleuproalalysprothrthrthrproalaproarg
130135140
proprothrproalaprothrilealaserglnproleuserleuarg
145150155160
proglualacysargproalaalaglyglyalavalhisthrarggly
165170175
leuaspphealacysaspiletyriletrpalaproleualaglythr
180185190
cysglyvalleuleuleuserleuvalilethrleutyrcysasnhis
195200205
argasnargargargvalcyslyscysproargprotrplyssergly
210215220
asplysproserleuseralaargtyrval
225230
<210>27
<211>164
<212>prt
<213>智人
<400>27
metlystrplysalaleuphethralaalaileleuglnalaglnleu
151015
proilethrglualaglnserpheglyleuleuaspprolysleucys
202530
tyrleuleuaspglyileleupheiletyrglyvalileleuthrala
354045
leupheleuargvallyspheserargseralaaspalaproalatyr
505560
glnglnglyglnasnglnleutyrasngluleuasnleuglyargarg
65707580
gluglutyraspvalleuasplysargargglyargaspproglumet
859095
glyglylysproglnargarglysasnproglngluglyleutyrasn
100105110
gluleuglnlysasplysmetalaglualatyrsergluileglymet
115120125
lysglygluargargargglylysglyhisaspglyleutyrglngly
130135140
leuserthralathrlysaspthrtyraspalaleuhismetglnala
145150155160
leuproproarg
<210>28
<211>164
<212>prt
<213>智人
<400>28
metlystrplysalaleuphethralaalaileleuglnalaglnleu
151015
proilethrglualaglnserpheglyleuleuaspprolysleucys
202530
tyrleuleuaspglyileleupheiletyrglyvalileleuthrala
354045
leupheleuargvallyspheserargseralaaspalaproalatyr
505560
glnglnglyglnasnglnleutyrasngluleuasnleuglyargarg
65707580
gluglutyraspvalleuasplysargargglyargaspproglumet
859095
glyglylysproglnargarglysasnproglngluglyleutyrasn
100105110
gluleuglnlysasplysmetalaglualatyrsergluileglymet
115120125
lysglygluargargargglylysglyhisaspglyleutyrglngly
130135140
leuserthralathrlysaspthrtyraspalaleuhismetglnala
145150155160
leuproproarg
<210>29
<211>337
<212>dna
<213>智人
<400>29
agagtgaagttcagcaggagcgcagagccccccgcgtaccagcagggccagaaccagctc60
tataacgagctcaatctaggacgaagagaggagtacgatgttttggacaagagacgtggc120
cgggaccctgagatggggggaaagccgagaaggaagaaccctcaggaaggcctgtacaat180
gaactgcagaaagataagatggcggaggcctacagtgagattgggatgaaaggcgagcgc240
cggaggggcaaggggcacgatggcctttaccagggtctcagtacagccaccaaggacacc300
tacgacgcccttcacatgcaggccctgccccctcgcg337
<210>30
<211>251
<212>prt
<213>智人
<400>30
metglyasnsercystyrasnilevalalathrleuleuleuvalleu
151015
asnphegluargthrargserleuglnaspprocysserasncyspro
202530
alaglythrphecysaspasnasnargasnglnilecysserprocys
354045
proproasnserpheserseralaglyglyglnargthrcysaspile
505560
cysargglncyslysglyvalpheargthrarglysglucysserser
65707580
thrserasnalaglucysaspcysthrproglyphehiscysleugly
859095
alaglycyssermetcysgluglnaspcyslysglnglyglngluleu
100105110
thrlyslysglycyslysaspcyscyspheglythrpheasnaspgln
115120125
lysargglyilecysargprotrpthrasncysserleuaspglylys
130135140
servalleuglythrlysgluargasptrpcysglyproserproala
145150155160
aspleuserproglyalaserservalthrproproalaproalaarg
165170175
gluproglyhisserproglnileileserphepheleualaleuthr
180185190
serthralaleuleupheleuleuphepheleuthrleuargpheser
195200205
trplysargglyarglyslysleuleutyrilephelysglnprophe
210215220
metargprovalglnthrthrglnglugluaspglycyssercysarg
225230235240
pheproglugluglugluglyglycysgluleu
245250
<210>31
<211>276
<212>prt
<213>智人
<400>31
metcysvalglyalaargargleuglyargglyprocysalaalaleu
151015
leuleuleuglyleuglyleuserthrvalthrglyleuhiscysval
202530
glyaspthrtyrproserasnaspargcyscyshisglucysargpro
354045
glyasnglymetvalserargcysserargserglnasnthrvalcys
505560
argprocysglyproglyphetyrasnasptrpserserlysprocys
65707580
lysprocysthrtrpcysasnleuargserglysergluarglysgln
859095
leucysthralathrglnaspthrvalcysargcysargalaglythr
100105110
glnproleuaspsertyrlysproglyvalaspcysalaprocyspro
115120125
proglyhispheserproglyaspasnglnalacyslysprotrpthr
130135140
asncysthrleualaglylyshisthrleuglnproalaserasnser
145150155160
seraspalailecysgluaspargaspproproalathrglnprogln
165170175
gluthrglnglyproproalaargproilethrvalglnprothrglu
180185190
alatrpproargthrserglnglyproserthrargprovalgluval
195200205
proglyglyargalavalalaalaileleuglyleuglyleuvalleu
210215220
glyleuleuglyproleualaileleuleualaleutyrleuleuarg
225230235240
argaspglnargleuproproaspalahislysproproglyglygly
245250255
serpheargthrproileglnglugluglnalaaspalahisserthr
260265270
leualalysile
275
<210>32
<211>196
<212>prt
<213>智人
<400>32
metlysserglyleutrptyrphepheleuphecysleuargilelys
151015
valleuthrglygluileasnglyseralaasntyrglumetpheile
202530
phehisasnglyglyvalglnileleucyslystyrproaspileval
354045
glnglnphelysmetglnleuleulysglyglyglnileleucysasp
505560
leuthrlysthrlysglyserglyasnthrvalserilelysserleu
65707580
lysphecyshisserglnleuserasnasnservalserphepheleu
859095
tyrasnleuasphisserhisalaasntyrtyrphecysasnleuser
100105110
ilepheaspproproprophelysvalthrleuthrglyglytyrleu
115120125
hisiletyrgluserglnleucyscysglnleulysphetrpleupro
130135140
ileglycysalaalaphevaltrpcysileleuglycysileleuile
145150155160
cystrpleuthrlyslyslystyrserserservalhisaspproasn
165170175
glyglutyrmetphemetargalathralalyslysserargleuthr
180185190
aspvalthrleu
195
<210>33
<211>220
<212>prt
<213>智人
<400>33
metalacysleuglypheglnarghislysalaglnleuasnleuala
151015
thrargthrtrpprocysthrleuleuphepheleuleupheilepro
202530
valphecyslysalamethisvalalaglnproalatrpleualaser
354045
serargglyilealaserphevalcysglutyralaserproglylys
505560
alathrgluvalargvalthrvalleuargglnalaaspserglnval
65707580
thrgluvalcysalaalathrtyrmetmetglyasngluleuthrphe
859095
leuaspaspserilecysthrglythrserserglyasnglnleuthr
100105110
ileglnglyleuargalametaspthrglyleutyrilecyslysval
115120125
gluleumettyrproproprotyrtyrleuglyileglyasnglythr
130135140
glniletyrvalileaspprogluprocysproaspserasppheleu
145150155160
leutrpileleualaalavalserserglyleuphephetyrserphe
165170175
leuleuthralavalserleuserlysmetleulyslysargserpro
180185190
leuthrthrglyvaltyrvallysmetproprothrgluproglucys
195200205
glulysglnpheglnprotyrpheileproileasn
210215220
<210>34
<211>286
<212>prt
<213>智人
<400>34
metglnileproglnalaprotrpprovalvaltrpalavalleugln
151015
leuglytrpargproglytrppheleuaspserproaspargprotrp
202530
asnproprothrpheserproalaleuleutrpthrgluglyaspasn
354045
alathrphethrcysserpheserasnthrsergluserphevalleu
505560
asntrptyrargmetserproserasnglnthrasplysleualaala
65707580
pheprogluaspargserglnproglyglnaspcysargpheargval
859095
thrglnleuproasnglyargaspphehismetservalvalargala
100105110
argargasnaspserglythrtyrleucysglyalaileserleuala
115120125
prolysalaglnilelysgluserleuargalagluleuargvalthr
130135140
gluargargalagluvalprothralahisproserproserproarg
145150155160
proalaglyglnpheglnthrleuvalvalglytrpglyglyleuleu
165170175
glyserleuvalleuleuvaltrpvalleualavalilecysserarg
180185190
alaalaargglythrileglyalaargargthrglyglnproleulys
195200205
gluaspproseralavalprovalpheservalasptyrglygluleu
210215220
asppheglntrpargglulysthrprogluproprovalprocysval
225230235240
progluglnthrglutyralathrilevalpheproserglymetgly
245250255
thrserserproalaargargglyseralaaspglyproargserala
260265270
glnproleuargprogluaspglyhiscyssertrpproleu
275280285
<210>35
<211>524
<212>prt
<213>智人
<400>35
mettrpglualaglnpheleuglyleuleupheleuglnproleutrp
151015
valalaprovallysproleuglnproglyalagluvalprotrptrp
202530
alaglngluglyalaproalaglnleuprocysserprothrilepro
354045
leuglnaspleuserleuleuargargalaglyvalthrtrpglnhis
505560
glnproaspserglyproproalaalaalaproglyhisproleuala
65707580
proglyprohisproalaalaprosersertrpglyproargproarg
859095
argtyrthrvalleuservalglyproglyglyleuargserglyarg
100105110
leuproleuglnproargvalglnleuaspgluargglyargglnarg
115120125
glyasppheserleutrpleuargproalaargargalaaspalagly
130135140
glutyrargalaalavalhisleuargaspargalaleusercysarg
145150155160
leuargleuargleuglyglnalasermetthralaserproprogly
165170175
serleuargalaserasptrpvalileleuasncysserpheserarg
180185190
proaspargproalaservalhistrppheargasnargglyglngly
195200205
argvalprovalarggluserprohishishisleualagluserphe
210215220
leupheleuproglnvalserprometaspserglyprotrpglycys
225230235240
ileleuthrtyrargaspglypheasnvalserilemettyrasnleu
245250255
thrvalleuglyleugluproprothrproleuthrvaltyralagly
260265270
alaglyserargvalglyleuprocysargleuproalaglyvalgly
275280285
thrargserpheleuthralalystrpthrproproglyglyglypro
290295300
aspleuleuvalthrglyaspasnglyaspphethrleuargleuglu
305310315320
aspvalserglnalaglnalaglythrtyrthrcyshisilehisleu
325330335
glngluglnglnleuasnalathrvalthrleualaileilethrval
340345350
thrprolysserpheglyserproglyserleuglylysleuleucys
355360365
gluvalthrprovalserglyglngluargphevaltrpserserleu
370375380
aspthrproserglnargserpheserglyprotrpleuglualagln
385390395400
glualaglnleuleuserglnprotrpglncysglnleutyrglngly
405410415
gluargleuleuglyalaalavaltyrphethrgluleuserserpro
420425430
glyalaglnargserglyargalaproglyalaleuproalaglyhis
435440445
leuleuleupheleuileleuglyvalleuserleuleuleuleuval
450455460
thrglyalapheglyphehisleutrpargargglntrpargproarg
465470475480
argpheseralaleugluglnglyilehisproproglnalaglnser
485490495
lysileglugluleugluglngluprogluprogluprogluproglu
500505510
progluprogluprogluprogluprogluglnleu
515520
<210>36
<211>368
<212>prt
<213>智人
<400>36
metleuglyglntrpthrleuileleuleuleuleuleulysvaltyr
151015
glnglylysglycysglnglyseralaasphistrpserilesergly
202530
valproleuglnleuglnproasnserileglnthrlysvalaspser
354045
ilealatrplyslysleuleuproserglnasnglyphehishisile
505560
leulystrpgluasnglyserleuproserasnthrserasnasparg
65707580
pheserpheilevallysasnleuserleuleuilelysalaalagln
859095
glnglnaspserglyleutyrcysleugluvalthrserilesergly
100105110
lysvalglnthralathrpheglnvalphevalphegluserleuleu
115120125
proasplysvalglulysproargleuglnglyglnglylysileleu
130135140
aspargglyargcysglnvalalaleusercysleuvalserargasp
145150155160
glyasnvalsertyralatrptyrargglyserlysleuileglnthr
165170175
alaglyasnleuthrtyrleuaspglugluvalaspileasnglythr
180185190
histhrtyrthrcysasnvalserasnprovalsertrpgluserhis
195200205
thrleuasnleuthrglnaspcysglnasnalahisglngluphearg
210215220
phetrppropheleuvalileilevalileleuseralaleupheleu
225230235240
glythrleualacysphecysvaltrpargarglysarglysglulys
245250255
glnsergluthrserprolysglupheleuthriletyrgluaspval
260265270
lysaspleulysthrargargasnhisgluglngluglnthrphepro
275280285
glyglyglyserthriletyrsermetileglnserglnserserala
290295300
prothrserglngluproalatyrthrleutyrserleuileglnpro
305310315320
serarglysserglyserarglysargasnhisserproserpheasn
325330335
serthriletyrgluvalileglylysserglnprolysalaglnasn
340345350
proalaargleuserarglysgluleugluasnpheaspvaltyrser
355360365
<210>37
<211>289
<212>prt
<213>智人
<400>37
metlysthrleuproalametleuglythrglylysleuphetrpval
151015
phepheleuileprotyrleuaspiletrpasnilehisglylysglu
202530
sercysaspvalglnleutyrilelysargglnsergluhisserile
354045
leualaglyaspprophegluleuglucysprovallystyrcysala
505560
asnargprohisvalthrtrpcyslysleuasnglythrthrcysval
65707580
lysleugluaspargglnthrsertrplysgluglulysasnileser
859095
phepheileleuhisphegluprovalleuproasnaspasnglyser
100105110
tyrargcysseralaasnpheglnserasnleuilegluserhisser
115120125
thrthrleutyrvalthraspvallysseralasergluargproser
130135140
lysaspglumetalaserargprotrpleuleutyrargleuleupro
145150155160
leuglyglyleuproleuleuilethrthrcysphecysleuphecys
165170175
cysleuargarghisglnglylysglnasngluleuseraspthrala
180185190
glyarggluileasnleuvalaspalahisleulyssergluglnthr
195200205
glualaserthrargglnasnserglnvalleuleusergluthrgly
210215220
iletyraspasnaspproaspleucyspheargmetglngluglyser
225230235240
gluvaltyrserasnprocysleuglugluasnlysproglyileval
245250255
tyralaserleuasnhisservalileglyproasnserargleuala
260265270
argasnvallysglualaprothrglutyralaserilecysvalarg
275280285
ser
<210>38
<211>21
<212>prt
<213>智人
<400>38
iletyriletrpalaproleualaglythrcysglyvalleuleuleu
151015
serleuvalilethr
20
<210>39
<211>63
<212>dna
<213>智人
<400>39
atctacatctgggcgcccctggccgggacttgtggggtccttctcctgtcactggttatc60
acc63
<210>40
<211>478
<212>prt
<213>智人
<400>40
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargproalaleualaglnlysargaspasnvalleuphe
202530
glnalaalathraspgluglnproalavalilelysthrleuglulys
354045
leuvalasnilegluthrglythrglyaspalagluglyilealaala
505560
alaglyasnpheleuglualagluleulysasnleuglyphethrval
65707580
thrargserlysseralaglyleuvalvalglyaspasnilevalgly
859095
lysilelysglyargglyglylysasnleuleuleumetserhismet
100105110
aspthrvaltyrleulysglyileleualalysalapropheargval
115120125
gluglyasplysalatyrglyproglyilealaaspasplysglygly
130135140
asnalavalileleuhisthrleulysleuleulysglutyrglyval
145150155160
argasptyrglythrilethrvalleupheasnthraspgluglulys
165170175
glyserpheglyserargaspleuileglngluglualalysleuala
180185190
asptyrvalleuserphegluprothrseralaglyaspglulysleu
195200205
serleuglythrserglyilealatyrvalglnvalglnilethrgly
210215220
lysalaserhisalaglyalaalaprogluleuglyvalasnalaleu
225230235240
valglualaseraspleuvalleuargthrmetasnileaspasplys
245250255
alalysasnleuargpheglntrpthrilealalysalaglyglnval
260265270
serasnileileproalaseralathrleuasnalaaspvalargtyr
275280285
alaargasngluasppheaspalaalametlysthrleuglugluarg
290295300
alaglnglnlyslysleuproglualaaspvallysvalilevalthr
305310315320
argglyargproalapheasnalaglygluglyglylyslysleuval
325330335
asplysalavalalatyrtyrlysglualaglyglythrleuglyval
340345350
glugluargthrglyglyglythraspalaalatyralaalaleuser
355360365
glylysprovalilegluserleuglyleuproglypheglytyrhis
370375380
serasplysalaglutyrvalaspileseralaileproargargleu
385390395400
tyrmetalaalaargleuilemetaspleuglyalaglylystyrpro
405410415
tyraspvalproasptyralaglyglyglyiletyriletrpalapro
420425430
leualaglythrcysglyvalleuleuleuserleuvalilethrleu
435440445
tyrcysasnhisargasnargargargvalglyglyglyargproval
450455460
vallysserglyasplysproserleuseralaargtyrval
465470475
<210>41
<211>1437
<212>dna
<213>智人
<400>41
atggccttgcccgttaccgcactcctgctgccgctggcgctgttgctgcacgcagctcga60
ccggccctcgcgcaaaaacgggacaatgtattgttccaagcggctactgatgaacagcca120
gcagtaattaaaacgcttgaaaaacttgtcaacatcgagaccgggacaggggatgctgaa180
gggattgcagccgcaggaaactttcttgaagccgaactcaaaaaccttgggtttaccgta240
acgcgctctaagtcagccggtttggtcgtaggtgacaacattgtgggaaagatcaaggga300
cggggcggtaaaaacttgcttttgatgtctcatatggatactgtatatcttaaggggatc360
ctcgcgaaggcgccctttcgggtggaaggcgataaggcatatggaccagggatagctgac420
gacaaggggggtaacgcggtcattctccacacactgaaacttcttaaagaatacggtgtg480
agagattatgggacgatcactgtactgttcaacaccgatgaggaaaagggctcttttggc540
tctagagatttgattcaagaagaagcgaaactggcagactacgtgttgtccttcgaaccc600
acatctgctggcgatgagaagctcagtcttggaacctctgggattgcgtacgtccaggtc660
caaataaccggaaaggcaagccatgctggagcagccccagaacttggcgtaaatgcgctg720
gtggaggcgagtgatctggtgctccggaccatgaacatagatgacaaagccaaaaatctg780
aggtttcaatggaccatcgctaaagcagggcaggtctctaatatcatcccggcttcagcc840
actttgaacgccgatgttaggtacgcccggaatgaagattttgatgccgctatgaaaaca900
ttggaggaacgagcgcagcaaaaaaagctgccagaagccgatgttaaagtaatagtaact960
agaggacgacccgcgttcaatgcgggcgagggtggaaagaagttggtcgataaggctgtt1020
gcgtactataaggaagcggggggtacgcttggagttgaggagaggaccggagggggcact1080
gacgcggcgtacgccgccctctcaggcaagcccgtaattgagtcacttgggcttcctggt1140
tttggttatcactcagacaaggccgagtacgtggatatatcagctatccctcggcgattg1200
tacatggcggcacgcttgataatggatttgggggctggaaagtatccttatgatgttcca1260
gactacgcagggggcggaatctacatctgggcgcccctggccgggacttgtggggtcctt1320
ctcctgtcactggttatcaccctgtattgtaaccaccgaaatcgacggcgagtaggagga1380
ggacgaccagttgttaaatctggggataaaccttcactgtccgctagatacgtgtaa1437
<210>42
<211>491
<212>prt
<213>智人
<400>42
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalagluvallysleuglnglnserglyalagluleuvalargpro
202530
glyserservallysilesercyslysalaserglytyralapheser
354045
sertyrtrpmetasntrpvallysglnargproglyglnglyleuglu
505560
trpileglyglniletyrproglyaspglyaspthrasntyrasngly
65707580
lysphelysglyglnalathrleuthralaasplysserserserthr
859095
alatyrmetglnleuserglyleuthrsergluaspseralavaltyr
100105110
phecysalaarglysthrileserservalvalaspphetyrpheasp
115120125
tyrtrpglyglnglythrthrvalthrvalserserglyglyglygly
130135140
serglyglyglyglyserglyglyglyglyseraspilegluleuthr
145150155160
glnserprolysphemetserthrservalglyaspargvalserval
165170175
thrcyslysalaserglnasnvalglythrasnvalalatrptyrgln
180185190
glnlysproglyglnserprolysproleuiletyrseralathrtyr
195200205
argasnserglyvalproaspargphethrglyserglyserglythr
210215220
aspphethrleuthrilethrasnvalglnserlysaspleualaasp
225230235240
tyrphecysglnglntyrasnargtyrprotyrthrserglyglygly
245250255
thrlysleugluilelysargalaalaalaprothrthrthrproala
260265270
proargproprothrproalaprothrilealaserglnproleuser
275280285
leuargproglualacysargproalaalaglyglyalavalhisthr
290295300
argglyleuaspphealacysaspiletyriletrpalaproleuala
305310315320
glythrcysglyvalleuleuleuserleuvalilethrleutyrcys
325330335
asnlysargglyarglyslysleuleutyrilephelysglnprophe
340345350
metargprovalglnthrthrglnglugluaspglycyssercysarg
355360365
pheproglugluglugluglyglycysgluleuargvallyspheser
370375380
argseralaaspalaproalatyrglnglnglyglnasnglnleutyr
385390395400
asngluleuasnleuglyargarggluglutyraspvalleuasplys
405410415
argargglyargaspproglumetglyglylysproargarglysasn
420425430
proglngluglyleutyrasngluleuglnlysasplysmetalaglu
435440445
alatyrsergluileglymetlysglygluargargargglylysgly
450455460
hisaspglyleutyrglnglyleuserthralathrlysaspthrtyr
465470475480
aspalaleuhismetglnalaleuproproarg
485490
<210>43
<211>381
<212>prt
<213>智人
<400>43
metmetarglysserleucyscysalaleuleuleuglyilesercys
151015
seralaleualathrprovalserglulysglnleualagluvalval
202530
alaasnthrilethrproleumetlysalaglnservalproglymet
354045
alavalalavaliletyrglnglylysprohistyrtyrthrphegly
505560
lysalaaspilealaalaasnlysprovalthrproglnthrleuphe
65707580
gluleuglyserileserlysthrphethrglyvalleuglyglyasp
859095
alailealaargglygluileserleuaspaspalavalthrargtyr
100105110
trpproglnleuthrglylysglntrpglnglyileargmetleuasp
115120125
leualathrtyrthralaglyglyleuproleuglnvalproaspglu
130135140
valthraspasnalaserleuleuargphetyrglnasntrpglnpro
145150155160
glntrplysproglythrthrargleutyralaasnalaserilegly
165170175
leupheglyalaleualavallysproserglymetprotyrglugln
180185190
alametthrthrargvalleulysproleulysleuasphisthrtrp
195200205
ileasnvalprolysalagluglualahistyralatrpglytyrarg
210215220
aspglylysalavalargvalserproglymetleuaspalaglnala
225230235240
tyrglyvallysthrasnvalglnaspmetalaasntrpvalmetala
245250255
asnmetalaprogluasnvalalaaspalaserleulysglnglyile
260265270
alaleualaglnserargtyrtrpargileglysermettyrglngly
275280285
leuglytrpglumetleuasntrpprovalglualaasnthrvalval
290295300
gluglyseraspserlysvalalaleualaproleuprovalalaglu
305310315320
valasnproproalaproprovallysalasertrpvalhislysthr
325330335
glyserthrglyglypheglysertyrvalalapheileproglulys
340345350
glnileglyilevalmetleualaasnthrsertyrproasnproala
355360365
argvalglualaalatyrhisileleuglualaleugln
370375380
<210>44
<211>361
<212>prt
<213>智人
<400>44
thrprovalserglulysglnleualagluvalvalalaasnthrile
151015
thrproleumetalaalaglnservalproglymetalavalalaval
202530
iletyrglnglylysprohistyrtyrthrpheglylysalaaspile
354045
alaalaasnlysprovalthrproglnthrleuphegluleuglyser
505560
ileserlysthrphethrglyvalleuglyglyaspalailealaarg
65707580
glygluileserleuaspaspalavalthrargtyrtrpproglnleu
859095
thrglylysglntrpglnglyileargmetleuaspleualathrtyr
100105110
thralaglyglyleuproleuglnvalproaspgluvalthraspasn
115120125
alaserleuleuargphetyrglnasntrpglnproglntrplyspro
130135140
glythrthrargleutyralaasnalaserileglyleupheglyala
145150155160
leualavallysproserglymetprotyrgluglnalametthrthr
165170175
argvalleulysproleulysleuasphisthrtrpileasnvalpro
180185190
lysalagluglualahistyralatrpglytyrargaspglylysala
195200205
valargvalserproglymetleuaspalaglnalatyrglyvallys
210215220
thrasnvalglnaspmetalaasntrpvalmetalaasnmetalapro
225230235240
gluasnvalalaaspalaserleulysglnglyilealaleualagln
245250255
serargtyrtrpargileglysermettyrglnglyleuglytrpglu
260265270
metleuasntrpprovalglualaasnthrvalvalgluglyserasp
275280285
serlysvalalaleualaproleuprovalalagluvalasnpropro
290295300
alaproprovallysalasertrpvalhislysthrglyserthrgly
305310315320
glypheglyalatyrvalalapheileproglulysglnileglyile
325330335
valmetleualaasnthrsertyrproasnproalaargvalgluala
340345350
alatyrhisileleuglualaleugln
355360
<210>45
<211>381
<212>prt
<213>智人
<400>45
metmetarglysserleucyscysalaleuleuleuglyilesercys
151015
seralaleualathrprovalserglulysglnleualagluvalval
202530
alaasnthrilethrproleumetalaalaglnservalproglymet
354045
alavalalavaliletyrglnglylysprohistyrtyrthrphegly
505560
lysalaaspilealaalaasnlysprovalthrproglnthrleuphe
65707580
gluleuglyserileserlysthrphethrglyvalleuglyglyasp
859095
alailealaargglygluileserleuaspaspalavalthrargtyr
100105110
trpproglnleuthrglylysglntrpglnglyileargmetleuasp
115120125
leualathrtyrthralaglyglyleuproleuglnvalproaspglu
130135140
valthraspasnalaserleuleuargphetyrglnasntrpglnpro
145150155160
glntrplysproglythrthrargleutyralaasnalaserilegly
165170175
leupheglyalaleualavallysproserglymetprotyrglugln
180185190
alametthrthrargvalleulysproleulysleuasphisthrtrp
195200205
ileasnvalprolysalagluglualahistyralatrpglytyrarg
210215220
aspglylysalavalargvalserproglymetleuaspalaglnala
225230235240
tyrglyvallysthrasnvalglnaspmetalaasntrpvalmetala
245250255
asnmetalaprogluasnvalalaaspalaserleulysglnglyile
260265270
alaleualaglnserargtyrtrpargileglysermettyrglngly
275280285
leuglytrpglumetleuasntrpprovalglualaasnthrvalval
290295300
gluglyseraspserlysvalalaleualaproleuprovalalaglu
305310315320
valasnproproalaproprovallysalasertrpvalhislysthr
325330335
glyserthrglyglypheglyalatyrvalalapheileproglulys
340345350
glnileglyilevalmetleualaasnthrsertyrproasnproala
355360365
argvalglualaalatyrhisileleuglualaleugln
370375380
<210>46
<211>391
<212>prt
<213>智人
<400>46
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargprothrprovalserglulysglnleualagluval
202530
valalaasnthrilethrproleumetalaalaglnservalprogly
354045
metalavalalavaliletyrglnglylysprohistyrtyrthrphe
505560
glylysalaaspilealaalaasnlysprovalthrproglnthrleu
65707580
phegluleuglyserileserlysthrphethrglyvalleuglygly
859095
aspalailealaargglygluileserleuaspaspalavalthrarg
100105110
tyrtrpproglnleuthrglylysglntrpglnglyileargmetleu
115120125
aspleualathrtyrthralaglyglyleuproleuglnvalproasp
130135140
gluvalthraspasnalaserleuleuargphetyrglnasntrpgln
145150155160
proglntrplysproglythrthrargleutyralaasnalaserile
165170175
glyleupheglyalaleualavallysproserglymetprotyrglu
180185190
glnalametthrthrargvalleulysproleulysleuasphisthr
195200205
trpileasnvalprolysalagluglualahistyralatrpglytyr
210215220
argaspglylysalavalargvalserproglymetleuaspalagln
225230235240
alatyrglyvallysthrasnvalglnaspmetalaasntrpvalmet
245250255
alaasnmetalaprogluasnvalalaaspalaserleulysglngly
260265270
ilealaleualaglnserargtyrtrpargileglysermettyrgln
275280285
glyleuglytrpglumetleuasntrpprovalglualaasnthrval
290295300
valgluglyseraspserlysvalalaleualaproleuprovalala
305310315320
gluvalasnproproalaproprovallysalasertrpvalhislys
325330335
thrglyserthrglyglypheglyalatyrvalalapheileproglu
340345350
lysglnileglyilevalmetleualaasnthrsertyrproasnpro
355360365
alaargvalglualaalatyrhisileleuglualaleuglntyrpro
370375380
tyraspvalproasptyrala
385390
<210>47
<211>426
<212>prt
<213>智人
<400>47
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargprothrprovalserglulysglnleualagluval
202530
valalaasnthrilethrproleumetalaalaglnservalprogly
354045
metalavalalavaliletyrglnglylysprohistyrtyrthrphe
505560
glylysalaaspilealaalaasnlysprovalthrproglnthrleu
65707580
phegluleuglyserileserlysthrphethrglyvalleuglygly
859095
aspalailealaargglygluileserleuaspaspalavalthrarg
100105110
tyrtrpproglnleuthrglylysglntrpglnglyileargmetleu
115120125
aspleualathrtyrthralaglyglyleuproleuglnvalproasp
130135140
gluvalthraspasnalaserleuleuargphetyrglnasntrpgln
145150155160
proglntrplysproglythrthrargleutyralaasnalaserile
165170175
glyleupheglyalaleualavallysproserglymetprotyrglu
180185190
glnalametthrthrargvalleulysproleulysleuasphisthr
195200205
trpileasnvalprolysalagluglualahistyralatrpglytyr
210215220
argaspglylysalavalargvalserproglymetleuaspalagln
225230235240
alatyrglyvallysthrasnvalglnaspmetalaasntrpvalmet
245250255
alaasnmetalaprogluasnvalalaaspalaserleulysglngly
260265270
ilealaleualaglnserargtyrtrpargileglysermettyrgln
275280285
glyleuglytrpglumetleuasntrpprovalglualaasnthrval
290295300
valgluglyseraspserlysvalalaleualaproleuprovalala
305310315320
gluvalasnproproalaproprovallysalasertrpvalhislys
325330335
thrglyserthrglyglypheglyalatyrvalalapheileproglu
340345350
lysglnileglyilevalmetleualaasnthrsertyrproasnpro
355360365
alaargvalglualaalatyrhisileleuglualaleuglntyrpro
370375380
tyraspvalproasptyralaglyglyglyleutyrcysasnhisarg
385390395400
asnargargargvalcyslyscysproargprovalvallyssergly
405410415
asplysproserleuseralaargtyrval
420425
<210>48
<211>970
<212>prt
<213>智人
<400>48
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalaalaargproalaleualaglnlysargaspasnvalleuphe
202530
glnalaalathraspgluglnproalavalilelysthrleuglulys
354045
leuvalasnilegluthrglythrglyaspalagluglyilealaala
505560
alaglyasnpheleuglualagluleulysasnleuglyphethrval
65707580
thrargserlysseralaglyleuvalvalglyaspasnilevalgly
859095
lysilelysglyargglyglylysasnleuleuleumetserhismet
100105110
aspthrvaltyrleulysglyileleualalysalapropheargval
115120125
gluglyasplysalatyrglyproglyilealaaspasplysglygly
130135140
asnalavalileleuhisthrleulysleuleulysglutyrglyval
145150155160
argasptyrglythrilethrvalleupheasnthraspgluglulys
165170175
glyserpheglyserargaspleuileglngluglualalysleuala
180185190
asptyrvalleuserphegluprothrseralaglyaspglulysleu
195200205
serleuglythrserglyilealatyrvalglnvalglnilethrgly
210215220
lysalaserhisalaglyalaalaprogluleuglyvalasnalaleu
225230235240
valglualaseraspleuvalleuargthrmetasnileaspasplys
245250255
alalysasnleuargpheglntrpthrilealalysalaglyglnval
260265270
serasnileileproalaseralathrleuasnalaaspvalargtyr
275280285
alaargasngluasppheaspalaalametlysthrleuglugluarg
290295300
alaglnglnlyslysleuproglualaaspvallysvalilevalthr
305310315320
argglyargproalapheasnalaglygluglyglylyslysleuval
325330335
asplysalavalalatyrtyrlysglualaglyglythrleuglyval
340345350
glugluargthrglyglyglythraspalaalatyralaalaleuser
355360365
glylysprovalilegluserleuglyleuproglypheglytyrhis
370375380
serasplysalaglutyrvalaspileseralaileproargargleu
385390395400
tyrmetalaalaargleuilemetaspleuglyalaglylystyrpro
405410415
tyraspvalproasptyralaglyglyglyleutyrcysasnhisarg
420425430
asnargargargvalcyslyscysproargprovalvallyssergly
435440445
asplysproserleuseralaargtyrvalglyserglyalathrasn
450455460
pheserleuleulysglnalaglyaspvalglugluasnproglypro
465470475480
alaleuprovalthralaleuleuleuproleualaleuleuleuhis
485490495
alagluvallysleuglnglnserglyalagluleuvalargprogly
500505510
serservallysilesercyslysalaserglytyralapheserser
515520525
tyrtrpmetasntrpvallysglnargproglyglnglyleuglutrp
530535540
ileglyglniletyrproglyaspglyaspthrasntyrasnglylys
545550555560
phelysglyglnalathrleuthralaasplysserserserthrala
565570575
tyrmetglnleuserglyleuthrsergluaspseralavaltyrphe
580585590
cysalaarglysthrileserservalvalaspphetyrpheasptyr
595600605
trpglyglnglythrthrvalthrvalserserglyglyglyglyser
610615620
glyglyglyglyserglyglyglyglyseraspilegluleuthrgln
625630635640
serprolysphemetserthrservalglyaspargvalservalthr
645650655
cyslysalaserglnasnvalglythrasnvalalatrptyrglngln
660665670
lysproglyglnserprolysproleuiletyrseralathrtyrarg
675680685
asnserglyvalproaspargphethrglyserglyserglythrasp
690695700
phethrleuthrilethrasnvalglnserlysaspleualaasptyr
705710715720
phecysglnglntyrasnargtyrprotyrthrserglyglyglythr
725730735
lysleugluilelysargalaalaalaprothrthrthrproalapro
740745750
argproprothrproalaprothrilealaserglnproleuserleu
755760765
argproglualacysargproalaalaglyglyalavalhisthrarg
770775780
glyleuaspphealacysaspiletyriletrpalaproleualagly
785790795800
thrcysglyvalleuleuleuserleuvalilethrleutyrcysasn
805810815
lysargglyarglyslysleuleutyrilephelysglnprophemet
820825830
argprovalglnthrthrglnglugluaspglycyssercysargphe
835840845
proglugluglugluglyglycysgluleuargvallyspheserarg
850855860
seralaaspalaproalatyrglnglnglyglnasnglnleutyrasn
865870875880
gluleuasnleuglyargarggluglutyraspvalleuasplysarg
885890895
argglyargaspproglumetglyglylysproargarglysasnpro
900905910
glngluglyleutyrasngluleuglnlysasplysmetalagluala
915920925
tyrsergluileglymetlysglygluargargargglylysglyhis
930935940
aspglyleutyrglnglyleuserthralathrlysaspthrtyrasp
945950955960
alaleuhismetglnalaleuproproarg
965970
<210>49
<211>911
<212>prt
<213>智人
<400>49
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalagluvallysleuglnglnserglyalagluleuvalargpro
202530
glyserservallysilesercyslysalaserglytyralapheser
354045
sertyrtrpmetasntrpvallysglnargproglyglnglyleuglu
505560
trpileglyglniletyrproglyaspglyaspthrasntyrasngly
65707580
lysphelysglyglnalathrleuthralaasplysserserserthr
859095
alatyrmetglnleuserglyleuthrsergluaspseralavaltyr
100105110
phecysalaarglysthrileserservalvalaspphetyrpheasp
115120125
tyrtrpglyglnglythrthrvalthrvalserserglyglyglygly
130135140
serglyglyglyglyserglyglyglyglyseraspilegluleuthr
145150155160
glnserprolysphemetserthrservalglyaspargvalserval
165170175
thrcyslysalaserglnasnvalglythrasnvalalatrptyrgln
180185190
glnlysproglyglnserprolysproleuiletyrseralathrtyr
195200205
argasnserglyvalproaspargphethrglyserglyserglythr
210215220
aspphethrleuthrilethrasnvalglnserlysaspleualaasp
225230235240
tyrphecysglnglntyrasnargtyrprotyrthrserglyglygly
245250255
thrlysleugluilelysargalaalaalaprothrthrthrproala
260265270
proargproprothrproalaprothrilealaserglnproleuser
275280285
leuargproglualacysargproalaalaglyglyalavalhisthr
290295300
argglyleuaspphealacysaspiletyriletrpalaproleuala
305310315320
glythrcysglyvalleuleuleuserleuvalilethrleutyrcys
325330335
asnlysargglyarglyslysleuleutyrilephelysglnprophe
340345350
metargprovalglnthrthrglnglugluaspglycyssercysarg
355360365
pheproglugluglugluglyglycysgluleuargvallyspheser
370375380
argseralaaspalaproalatyrglnglnglyglnasnglnleutyr
385390395400
asngluleuasnleuglyargarggluglutyraspvalleuasplys
405410415
argargglyargaspproglumetglyglylysproargarglysasn
420425430
proglngluglyleutyrasngluleuglnlysasplysmetalaglu
435440445
alatyrsergluileglymetlysglygluargargargglylysgly
450455460
hisaspglyleutyrglnglyleuserthralathrlysaspthrtyr
465470475480
aspalaleuhismetglnalaleuproproargglnproleuglugly
485490495
serglyalathrasnpheserleuleulysglnalaglyaspvalglu
500505510
gluasnproglyprometalaleualaglnlysargaspasnvalleu
515520525
pheglnalaalathraspgluglnproalavalilelysthrleuglu
530535540
lysleuvalasnilegluthrglythrglyaspalagluglyileala
545550555560
alaalaglyasnpheleuglualagluleulysasnleuglyphethr
565570575
valthrargserlysseralaglyleuvalvalglyaspasnileval
580585590
glylysilelysglyargglyglylysasnleuleuleumetserhis
595600605
metaspthrvaltyrleulysglyileleualalysalaprophearg
610615620
valgluglyasplysalatyrglyproglyilealaaspasplysgly
625630635640
glyasnalavalileleuhisthrleulysleuleulysglutyrgly
645650655
valargasptyrglythrilethrvalleupheasnthraspgluglu
660665670
lysglyserpheglyserargaspleuileglngluglualalysleu
675680685
alaasptyrvalleuserphegluprothrseralaglyaspglulys
690695700
leuserleuglythrserglyilealatyrvalglnvalglnilethr
705710715720
glylysalaserhisalaglyalaalaprogluleuglyvalasnala
725730735
leuvalglualaseraspleuvalleuargthrmetasnileaspasp
740745750
lysalalysasnleuargpheglntrpthrilealalysalaglygln
755760765
valserasnileileproalaseralathrleuasnalaaspvalarg
770775780
tyralaargasngluasppheaspalaalametlysthrleugluglu
785790795800
argalaglnglnlyslysleuproglualaaspvallysvalileval
805810815
thrargglyargproalapheasnalaglygluglyglylyslysleu
820825830
valasplysalavalalatyrtyrlysglualaglyglythrleugly
835840845
valglugluargthrglyglyglythraspalaalatyralaalaleu
850855860
serglylysprovalilegluserleuglyleuproglypheglytyr
865870875880
hisserasplysalaglutyrvalaspileseralaileproargarg
885890895
leutyrmetalaalaargleuilemetaspleuglyalaglylys
900905910
<210>50
<211>880
<212>prt
<213>智人
<400>50
metthrprovalserglulysglnleualagluvalvalalaasnthr
151015
ilethrproleumetalaalaglnservalproglymetalavalala
202530
valiletyrglnglylysprohistyrtyrthrpheglylysalaasp
354045
ilealaalaasnlysprovalthrproglnthrleuphegluleugly
505560
serileserlysthrphethrglyvalleuglyglyaspalaileala
65707580
argglygluileserleuaspaspalavalthrargtyrtrpprogln
859095
leuthrglylysglntrpglnglyileargmetleuaspleualathr
100105110
tyrthralaglyglyleuproleuglnvalproaspgluvalthrasp
115120125
asnalaserleuleuargphetyrglnasntrpglnproglntrplys
130135140
proglythrthrargleutyralaasnalaserileglyleuphegly
145150155160
alaleualavallysproserglymetprotyrgluglnalametthr
165170175
thrargvalleulysproleulysleuasphisthrtrpileasnval
180185190
prolysalagluglualahistyralatrpglytyrargaspglylys
195200205
alavalargvalserproglymetleuaspalaglnalatyrglyval
210215220
lysthrasnvalglnaspmetalaasntrpvalmetalaasnmetala
225230235240
progluasnvalalaaspalaserleulysglnglyilealaleuala
245250255
glnserargtyrtrpargileglysermettyrglnglyleuglytrp
260265270
glumetleuasntrpprovalglualaasnthrvalvalgluglyser
275280285
aspserlysvalalaleualaproleuprovalalagluvalasnpro
290295300
proalaproprovallysalasertrpvalhislysthrglyserthr
305310315320
glyglypheglyalatyrvalalapheileproglulysglnilegly
325330335
ilevalmetleualaasnthrsertyrproasnproalaargvalglu
340345350
alaalatyrhisileleuglualaleuglntyrprotyraspvalpro
355360365
asptyralaalathrasnpheserleuleulysglnalaglyaspval
370375380
glugluasnproglyproalaleuprovalthralaleuleuleupro
385390395400
leualaleuleuleuhisalagluvallysleuglnglnserglyala
405410415
gluleuvalargproglyserservallysilesercyslysalaser
420425430
glytyralaphesersertyrtrpmetasntrpvallysglnargpro
435440445
glyglnglyleuglutrpileglyglniletyrproglyaspglyasp
450455460
thrasntyrasnglylysphelysglyglnalathrleuthralaasp
465470475480
lysserserserthralatyrmetglnleuserglyleuthrserglu
485490495
aspseralavaltyrphecysalaarglysthrileserservalval
500505510
aspphetyrpheasptyrtrpglyglnglythrthrvalthrvalser
515520525
serglyglyglyglyserglyglyglyglyserglyglyglyglyser
530535540
aspilegluleuthrglnserprolysphemetserthrservalgly
545550555560
aspargvalservalthrcyslysalaserglnasnvalglythrasn
565570575
valalatrptyrglnglnlysproglyglnserprolysproleuile
580585590
tyrseralathrtyrargasnserglyvalproaspargphethrgly
595600605
serglyserglythraspphethrleuthrilethrasnvalglnser
610615620
lysaspleualaasptyrphecysglnglntyrasnargtyrprotyr
625630635640
thrserglyglyglythrlysleugluilelysargalaalaalapro
645650655
thrthrthrproalaproargproprothrproalaprothrileala
660665670
serglnproleuserleuargproglualacysargproalaalagly
675680685
glyalavalhisthrargglyleuaspphealacysaspiletyrile
690695700
trpalaproleualaglythrcysglyvalleuleuleuserleuval
705710715720
ilethrleutyrcysasnlysargglyarglyslysleuleutyrile
725730735
phelysglnprophemetargprovalglnthrthrglnglugluasp
740745750
glycyssercysargpheproglugluglugluglyglycysgluleu
755760765
argvallyspheserargseralaaspalaproalatyrglnglngly
770775780
glnasnglnleutyrasngluleuasnleuglyargarggluglutyr
785790795800
aspvalleuasplysargargglyargaspproglumetglyglylys
805810815
proargarglysasnproglngluglyleutyrasngluleuglnlys
820825830
asplysmetalaglualatyrsergluileglymetlysglygluarg
835840845
argargglylysglyhisaspglyleutyrglnglyleuserthrala
850855860
thrlysaspthrtyraspalaleuhismetglnalaleuproproarg
865870875880
<210>51
<211>887
<212>prt
<213>智人
<400>51
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalagluvallysleuglnglnserglyalagluleuvalargpro
202530
glyserservallysilesercyslysalaserglytyralapheser
354045
sertyrtrpmetasntrpvallysglnargproglyglnglyleuglu
505560
trpileglyglniletyrproglyaspglyaspthrasntyrasngly
65707580
lysphelysglyglnalathrleuthralaasplysserserserthr
859095
alatyrmetglnleuserglyleuthrsergluaspseralavaltyr
100105110
phecysalaarglysthrileserservalvalaspphetyrpheasp
115120125
tyrtrpglyglnglythrthrvalthrvalserserglyglyglygly
130135140
serglyglyglyglyserglyglyglyglyseraspilegluleuthr
145150155160
glnserprolysphemetserthrservalglyaspargvalserval
165170175
thrcyslysalaserglnasnvalglythrasnvalalatrptyrgln
180185190
glnlysproglyglnserprolysproleuiletyrseralathrtyr
195200205
argasnserglyvalproaspargphethrglyserglyserglythr
210215220
aspphethrleuthrilethrasnvalglnserlysaspleualaasp
225230235240
tyrphecysglnglntyrasnargtyrprotyrthrserglyglygly
245250255
thrlysleugluilelysargalaalaalaprothrthrthrproala
260265270
proargproprothrproalaprothrilealaserglnproleuser
275280285
leuargproglualacysargproalaalaglyglyalavalhisthr
290295300
argglyleuaspphealacysaspiletyriletrpalaproleuala
305310315320
glythrcysglyvalleuleuleuserleuvalilethrleutyrcys
325330335
asnlysargglyarglyslysleuleutyrilephelysglnprophe
340345350
metargprovalglnthrthrglnglugluaspglycyssercysarg
355360365
pheproglugluglugluglyglycysgluleuargvallyspheser
370375380
argseralaaspalaproalatyrglnglnglyglnasnglnleutyr
385390395400
asngluleuasnleuglyargarggluglutyraspvalleuasplys
405410415
argargglyargaspproglumetglyglylysproargarglysasn
420425430
proglngluglyleutyrasngluleuglnlysasplysmetalaglu
435440445
alatyrsergluileglymetlysglygluargargargglylysgly
450455460
hisaspglyleutyrglnglyleuserthralathrlysaspthrtyr
465470475480
aspalaleuhismetglnalaleuproproargglnproleuglugly
485490495
serglyalathrasnpheserleuleulysglnalaglyaspvalglu
500505510
gluasnproglyprothrprovalserglulysglnleualagluval
515520525
valalaasnthrilethrproleumetalaalaglnservalprogly
530535540
metalavalalavaliletyrglnglylysprohistyrtyrthrphe
545550555560
glylysalaaspilealaalaasnlysprovalthrproglnthrleu
565570575
phegluleuglyserileserlysthrphethrglyvalleuglygly
580585590
aspalailealaargglygluileserleuaspaspalavalthrarg
595600605
tyrtrpproglnleuthrglylysglntrpglnglyileargmetleu
610615620
aspleualathrtyrthralaglyglyleuproleuglnvalproasp
625630635640
gluvalthraspasnalaserleuleuargphetyrglnasntrpgln
645650655
proglntrplysproglythrthrargleutyralaasnalaserile
660665670
glyleupheglyalaleualavallysproserglymetprotyrglu
675680685
glnalametthrthrargvalleulysproleulysleuasphisthr
690695700
trpileasnvalprolysalagluglualahistyralatrpglytyr
705710715720
argaspglylysalavalargvalserproglymetleuaspalagln
725730735
alatyrglyvallysthrasnvalglnaspmetalaasntrpvalmet
740745750
alaasnmetalaprogluasnvalalaaspalaserleulysglngly
755760765
ilealaleualaglnserargtyrtrpargileglysermettyrgln
770775780
glyleuglytrpglumetleuasntrpprovalglualaasnthrval
785790795800
valgluglyseraspserlysvalalaleualaproleuprovalala
805810815
gluvalasnproproalaproprovallysalasertrpvalhislys
820825830
thrglyserthrglyglypheglyalatyrvalalapheileproglu
835840845
lysglnileglyilevalmetleualaasnthrsertyrproasnpro
850855860
alaargvalglualaalatyrhisileleuglualaleuglntyrpro
865870875880
tyraspvalproasptyrala
885
<210>52
<211>263
<212>prt
<213>智人
<400>52
metalaleuprovalthralaleuleuleuproleualaleuleuleu
151015
hisalagluvallysleuglnglnserglyalagluleuvalargpro
202530
glyserservallysilesercyslysalaserglytyralapheser
354045
sertyrtrpmetasntrpvallysglnargproglyglnglyleuglu
505560
trpileglyglniletyrproglyaspglyaspthrasntyrasngly
65707580
lysphelysglyglnalathrleuthralaasplysserserserthr
859095
alatyrmetglnleuserglyleuthrsergluaspseralavaltyr
100105110
phecysalaarglysthrileserservalvalaspphetyrpheasp
115120125
tyrtrpglyglnglythrthrvalthrvalserserglyglyglygly
130135140
serglyglyglyglyserglyglyglyglyseraspilegluleuthr
145150155160
glnserprolysphemetserthrservalglyaspargvalserval
165170175
thrcyslysalaserglnasnvalglythrasnvalalatrptyrgln
180185190
glnlysproglyglnserprolysproleuiletyrseralathrtyr
195200205
argasnserglyvalproaspargphethrglyserglyserglythr
210215220
aspphethrleuthrilethrasnvalglnserlysaspleualaasp
225230235240
tyrphecysglnglntyrasnargtyrprotyrthrserglyglygly
245250255
thrlysleugluilelysarg
260
<210>53
<211>789
<212>dna
<213>智人
<400>53
atggctctcccagtgactgccctactgcttcccctagcgcttctcctgcatgcagaggtg60
aagctgcagcagtctggggctgagctggtgaggcctgggtcctcagtgaagatttcctgc120
aaggcttctggctatgcattcagtagctactggatgaactgggtgaagcagaggcctgga180
cagggtcttgagtggattggacagatttatcctggagatggtgatactaactacaatgga240
aagttcaagggtcaagccacactgactgcagacaaatcctccagcacagcctacatgcag300
ctcagcggcctaacatctgaggactctgcggtctatttctgtgcaagaaagaccattagt360
tcggtagtagatttctactttgactactggggccaagggaccacggtcaccgtctcctca420
ggtggaggtggatcaggtggaggtggatctggtggaggtggatctgacattgagctcacc480
cagtctccaaaattcatgtccacatcagtaggagacagggtcagcgtcacctgcaaggcc540
agtcagaatgtgggtactaatgtagcctggtatcaacagaaaccaggacaatctcctaaa600
ccactgatttactcggcaacctaccggaacagtggagtccctgatcgcttcacaggcagt660
ggatctgggacagatttcactctcaccatcactaacgtgcagtctaaagacttggcagac720
tatttctgtcaacaatataacaggtatccgtacacgtccggaggggggaccaagctggag780
atcaaacgg789
- 还没有人留言评论。精彩留言会获得点赞!