激动性CD40抗体的制作方法
本发明涉及与人cd40受体特异性结合并且能够独立于fcγ介导的cd40受体交联而诱导cd40信号传导的人源化单克隆激动性抗体或其抗原结合片段。本发明还涉及所述抗体和包含它们的药物组合物的用途。
背景技术:
最近在癌症免疫治疗方面取得的成功恢复了以下假设:免疫系统可以控制许多(即使不是大多数)癌症,在一些情况下以许多小分子药物所不具备的方式产生持久的反应。激动性cd40单克隆抗体(mab)提供了一种新的治疗选择,它有可能通过各种机制产生抗癌免疫力。
cd40是细胞表面分子,是肿瘤坏死因子(tnf)受体超家族的成员。它在诸如树突细胞、b细胞和单核细胞以及许多非免疫细胞等抗原呈递细胞(apc)上广泛表达,并在一系列肿瘤中表达。
cd40的天然配体是cd154,它主要在活化t淋巴细胞的表面上表达,并为免疫应答提供t细胞“帮助”的主要成分:通过cd40在apc上的信号传导在很大程度上介导了辅助t细胞授权apc的能力。例如,cd40在dc上的连接诱导增加共刺激分子和mhc分子的表面表达、促炎性细胞因子的产生以及增强的t细胞触发作用。在静止的b细胞上的cd40连接增加了抗原呈递功能和增殖。
cd40信号传导的影响是多方面的,并取决于表达cd40的细胞类型和提供cd40信号的微环境。像tnf受体家族的其他成员一样,cd40信号传导由衔接子分子介导,而不是由cd40胞质尾区的固有信号转导活性介导。受体组装后,下游激酶被激活,多组分信号传导复合体从cd40移位至细胞溶胶中,并激活了几个特征明确的信号转导途径。
拮抗人cd40抗体是本领域已知的。相应的拮抗抗体可以是沉默的fc变体,显示出减少的fcγ介导的cd40受体交联。人igg1fc区的相应的突变描述于例如us2018/0118843中。
在最近设计的免疫调节方法中,靶向cd40的激动剂单克隆抗体(mab)用于增强免疫系统识别和破坏癌细胞的能力。相应的临床前研究表明,激动性cd40mab可以激活apc并促进抗肿瘤t细胞应答,并在缺乏t细胞免疫力的情况下培养具有控制癌症的潜能的细胞毒性髓样细胞。因此,激动性cd40mab根本不同于mab,后者通过阻断如ctla-4或pd-1等的阴性检查点分子来实现免疫激活。
cp-870,893是首个完全人igg2mab,其作为cd40的有效和选择性激动剂。有趣的是,cp-870,893的结合不会与cd154与cd40的结合竞争。在临床前研究中,cp-870,893已显示出介导对肿瘤细胞存活的免疫系统依赖性和非依赖性影响。在首次人类研究中,观察到有希望的抗肿瘤活性,尤其是在黑色素瘤患者中。在药效学上,施用cp-870,893导致外周血b细胞的短暂减少,并导致apc上激活标志物的上调。
因此,激动性cd40mab代表了新型癌症疗法的有希望的策略。然而,关于其潜在的细胞毒性副作用也已经引起了关注。激动性单克隆cd40抗体有望触发细胞因子释放综合征、自身免疫反应、血栓栓塞综合征(由于血小板和内皮细胞表达cd40)、过度免疫刺激导致激活诱导的细胞死亡或耐受性、以及肿瘤血管生成。这些作用可导致不良的毒性或促进肿瘤生长。从机理上讲,在动物研究中,激动性cd40和其他靶向tnf受体家族的抗体与fcγ受体相互作用的能力已经与毒性的发生有关(li&ravetch2012,xu等人2003,byrne等人2016)。
对于检测的最强的激动剂cp-870,893,已报道的最常见的副作用是细胞因子释放综合征,在输注后不久表现为发冷、发烧、僵硬和其他症状。同样,使用cp-870,893还观察到了几例血栓栓塞事件。使用达西妥珠单抗(dacetuzumab),已经观察到非传染性炎症性眼炎。
因此,需要提供激动性cd40mab,其表现出降低的细胞毒性,导致较少的临床副作用,同时保持其效力和临床有效性。本发明的激动性cd40mab可以满足该需求,允许利用激动性cd40抗体的全部免疫调节潜力。
技术实现要素:
本发明提供与人cd40受体特异性结合并独立于fcγ介导的cd40受体交联而诱导cd40信号传导的单克隆抗体或其抗原结合片段。更具体地,本发明的抗体结合与cd40配体的表位重叠的cd40表位,并能够激活人apc。本发明还提供包含所述抗体的组合物,以及该抗体和组合物在治疗期望刺激免疫系统的病症或疾病中的用途,例如,在患有癌症的患者的治疗中。
定义
术语“抗体”涵盖抗体结构的各种形式,包括但不限于整个抗体和抗体片段,只要其显示出根据本发明的特性即可。
“抗体片段”是指除完整抗体以外的分子,其包含完整抗体的一部分,结合完整抗体所结合的抗原。抗体片段的实例包括但不限于fv、fab、fab'、fab'-sh、f(ab')2;双抗体;线性抗体;单链抗体分子(例如scfv);和由抗体片段形成的多特异性抗体。
如本文所用,术语“单克隆抗体”或“单克隆抗体组合物”是指单一氨基酸组合物的抗体分子的制剂。
术语“人源化抗体”或“抗体的人源化形式”是指由于抗体工程化而使重链和轻链均被人源化的抗体。人源化链通常是这样的链,其中v区氨基酸序列已改变,因此从整体上分析,其与人种系序列的同源性比与来源物种的种系序列的同源性更近。人源化评估是基于所得的氨基酸序列,而不是基于方法本身。
如本文所用,术语“针对靶标或抗靶标抗体特异性结合”是指通过elisa测定的、抗体与相应抗原(靶标)或抗原表达细胞的结合,其中所述elisa优选包括以下:包被相应的抗原至固相支持物,在允许与相应的抗原或蛋白质形成免疫复合体的条件下添加所述抗体,使用与本发明所述的抗体结合的二抗通过测定光密度值(od)来检测所述免疫复合体,并使用过氧化物酶介导的显色。
根据本发明的术语“抗原”是指用于免疫的抗原或包含所述抗原作为其蛋白质序列的一部分的蛋白质。例如,为了免疫,可以使用蛋白质的胞外结构域的片段(例如前20个氨基酸),并且对于检测/测定等,可以使用蛋白质或全长蛋白质的胞外结构域。
本文中的术语“特异性结合”或“特异性识别”是指抗体对抗原表现出明显的亲和力,并且优选地,不表现出明显的交叉反应性。
“不表现出明显的交叉反应性”的抗体是不会明显结合至不希望的其他蛋白质的抗体。可以根据本领域公知的用于确定这种结合的任何方法来确定特异性结合,例如,通过竞争性结合测定如elisa。
“与相同表位结合的抗体”作为参考抗体是指在竞争测定中将参考抗体与其抗原的结合阻断50%以上的抗体,相反,在竞争测定中参考抗体阻断抗体与其抗原的结合达50%以上。
如本文所用,“根据本发明的抗体的可变区(或结构域)”(轻链的可变区(vl),重链的可变区(vh))表示直接参与抗体与抗原结合的该对轻链和重链区中的每一个。可变轻链区和重链区具有相同的一般结构,并且每个区均包含四个框架(fr)区,它们的序列是广泛保守的,通过三个互补决定区cdr连接。
当在本文中使用时,术语“抗体的抗原结合部分”是指负责抗原结合的抗体的氨基酸残基。抗体的抗原结合部分优选包含来自“互补决定区”或“cdr”的氨基酸残基。cdr序列是根据kabat等人,sequencesofproteinsofimmunologicalinterest,第5版publichealthservice,nationalinstitutesofhealth,bethesda,md.(1991)定义的。使用该编号系统,实际的线性氨基酸序列可以包含较少的或额外的氨基酸,其对应于可变区的fr或cdr的缩短或插入。例如,重链可变区可以包括在h2的残基52之后单个氨基酸插入(根据kabat的残基52a)和在重链fr残基82之后的插入的残基(例如,根据kabat的残基82、,82b和82c等)。对于给定抗体,残基的kabat编号可以通过在抗体序列与“标准”kabat编号序列的同源性区域进行比对来确定。
“恒定结构域(恒定部分)”不直接参与抗体与抗原的结合,但是表现出例如还有效应子功能。对应于人igg1的重链恒定区基因片段称为γ1链。对应于人igg3的重链恒定区基因片段称为γ3链。人恒定的γ重链在kabat,e.a等人,sequencesofproteinsofimmunologicalinterest,第5版,publichealthservice,nationalinstitutesofhealth,bethesda,md.(1991)和brueggemann,m.等人,j.exp.med.166(1987)1351-1361;love,t.w.等人,methodsenzymol.178(1989)515-527中详细描述。
本文中的术语“fc区”用于定义包含至少一部分恒定区的免疫球蛋白重链的c末端区域。该术语包括天然序列fc区和变体fc区。
除非本文另有说明,否则fc区或恒定区中氨基酸残基的编号是根据eu编号系统(也称为eu索引)进行的,如kabat等人,sequencesofproteinsofimmunologicalinterest,第5版.publichealthservice,nationalinstitutesofhealth,bethesda,md(1991)所述。
“变体fc区”包含由于本文定义的至少一个“氨基酸修饰”而不同于“天然”或“野生型”序列fc区的氨基酸序列。
如本文所用,术语“fc变体”是指在fc结构域中包含修饰的多肽。该修饰可以是添加、缺失或取代。取代可以包括天然存在的氨基酸和非天然存在的氨基酸。变体可以包含非天然氨基酸。
术语“含fc区的多肽”是指包含fc区的多肽,例如抗体。
术语“fc受体”或“fcr”用于描述与抗体的fc区结合的受体。结合igg抗体的fcr(γ受体)包括fcyri、fcyrii和fcyriii亚类的受体,包括这些受体的等位基因变体和可选择地剪接形式。fcyrii受体包括fcyriia(“激活受体”)和fcyriib(“抑制受体”),其具有相似的氨基酸序列,其主要区别在于其胞质结构域。活化受体fcyriia在其胞质结构域中含有基于免疫受体酪氨酸的活化基序(itam)。抑制受体fcyriib在其胞质结构域中含有基于免疫受体酪氨酸的抑制基序(itim)(参见daeron,m.,annu.rev.immunol.15(1997)203-234中的综述)。fcr在ravetch和kinet,annu.rev.immunol9(1991)457-492;capel等人,immunomethods4(1994)25-34;和dehaas等人,j.lab.clin.med.126(1995)330-41中进行综述。其他fcr,包括将来要鉴定的那些,由本文中的术语“fcr”所涵盖。该术语还包括新生儿受体fcrn,它负责将母体igg转移至胎儿(guyer等人,j.immunol.117(1976)587和kim等人,j.immunol.24(1994)249)。
如本文所用,“iggfc配体”是指来自任何生物的分子,优选多肽,其结合至igg抗体的fc区以形成fc/fc配体复合体。fc配体包括但不限于fcyr、fcrn、clq、c3、甘露聚糖结合凝集素、甘露糖受体、葡萄球菌蛋白a、链球菌蛋白g和病毒fcyr。fc配体还包括fc受体同源物(fcrh),其是与fcyr同源的fc受体家族(davis等人,immunologicalreviews190(2002)123-136,通过引用整体并入)。fc配体可包括未发现的结合fc的分子。特定的iggfc配体是fcrn和fcγ受体。如本文所用,“fc配体”是指来自任何生物的分子,优选多肽,其结合至抗体的fc区以形成fc/fc配体复合体。
如本文所用,“fcγ受体”、“fcyr”或“fcγr”是指结合igg抗体fc区并由fcyr基因编码的蛋白质家族的任何成员。在人中,该家族包括但不限于fcyri(cd64),包括同工型fcyria、fcyrib和fcyric;fcyrii(cd32),包括同工型fcyriia(包括同种异型h131和r131)、fcyriib(包括fcyriib-1和fcyriib-2)、和fcyriic;和fcyriii(cd16),包括同工型fcyriiia(包括同种异型v158和f158)和fcyriiib(包括同种异型fcyriib-na1和fcyriib-na2)(jefferis等人,immunollett82(2002)),以及任何未发现的人fcyr或fcyr同种型或同种异型。fcyr可以来自任何生物,包括但不限于人类、小鼠、大鼠、兔子和猴子。小鼠fcyr包括但不限于fcyri(cd64)、fcyrii(cd32)、fcyriii(cd16)和fcyriii-2(cd16-2),以及任何未发现的小鼠fcyr或fcyr同种型或同种异型。
如本文所用,“fcrn”或“新生儿fc受体”是指结合igg抗体fc区并且至少部分由fcrn基因编码的蛋白质。fcrn可以来自任何生物,包括但不限于人、小鼠、大鼠、兔和猴子。如本领域已知的,功能性fcrn蛋白包含两个多肽,通常称为重链和轻链。轻链是β-2-微球蛋白,重链由fcrn基因编码。除非本文另有说明,否则fcrn或fcrn蛋白是指fcrn重链与β-2-微球蛋白的复合体。
在比对序列并引入空位后(如有必要),将相对于肽或多肽序列的“氨基酸序列同一性百分比(%)”定义为候选序列中与特定肽或多肽序列中的氨基酸残基相同的氨基酸残基的百分比,以实现最大的序列同一性百分比,并且不考虑将任何保守取代作为序列同一性的一部分。为了确定氨基酸序列同一性百分比的目的,比对可以以本领域技术范围内的多种方式,例如,使用公开可用的计算机软件,例如blast、blast-2、align或megalign(dnastar)软件来实现。
“抗体依赖性细胞介导的细胞毒性”和“adcc”是指细胞介导的反应,其中表达fcr的非特异性细胞毒性细胞(例如天然杀伤(nk)细胞、嗜中性粒细胞和巨噬细胞)识别靶细胞上结合的抗体,随后引起靶细胞的裂解。用于介导adcc的主要细胞nk细胞仅表达fcyriii,而单核细胞表达fcyri、fcyrii和fcyriii。造血细胞上的fcr表达总结于ravetch和kinet,annu.rev.immunol9(1991)457-492的第464页的表3中。
术语“抗体依赖性细胞吞噬作用”和“adcp”是指将抗体包被的细胞通过结合至免疫球蛋白fc区的吞噬性免疫细胞(例如巨噬细胞、嗜中性粒细胞和树突细胞)进行全部或部分内化的过程。
如本文所用,术语“一种或多种抗体效应子功能”或“效应子功能”是指由igg的一个或多个fc效应子结构域(例如,免疫球蛋白的fc区)贡献的功能。此类功能可通过例如将一个或多个fc效应子结构域与具有吞噬或裂解活性的免疫细胞上的fc受体结合或通过将一个或多个fc效应子结构域与补体系统的组分结合来影响。典型的效应子功能是adcc、adcp和cdc。
“c1q”是包括免疫球蛋白fc区结合位点的多肽。c1q与两个丝氨酸蛋白酶c1r和c1s一起形成复合体c1,这是补体依赖性细胞毒性(cdc)途径的第一组分。
抗体的“类别”是指其重链所具有的恒定结构域或恒定区的类型。抗体主要分为五类:iga、igd、ige、igg和igm,其中一些可以进一步分为亚类(同种型),例如igg1、igg2、igg3、igg4、iga1和iga2。对应于不同类别的免疫球蛋白的重链恒定结构域分别称为α、δ、ε、γ和μ。
试剂例如药物制剂的“有效量”是指在所需剂量和时间段内有效达到所需治疗或预防效果的量。
如本文所用,术语“癌症”可以是例如肺癌、非小细胞肺癌(nscl)癌、细支气管肺泡细胞肺癌、骨癌、胰腺癌、晚期胰腺癌皮肤癌、头或颈癌、皮肤或眼内黑色素瘤、子宫癌、卵巢癌、直肠癌、肛区癌、肠胃癌(stomachcancer)、胃癌(gastriccancer)、结肠癌、乳腺癌、子宫癌、输卵管癌、子宫内膜癌、子宫颈癌、阴道癌、外阴癌、霍奇金病、食道癌、小肠癌、内分泌系统癌、甲状腺癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、阴茎癌、前列腺癌、膀胱癌、肾脏或输尿管癌、肾细胞癌、肾盂癌、间皮瘤、肝细胞癌、胆管癌、中枢神经系统肿瘤(cns)、脊柱肿瘤、脑干神经胶质瘤、多形胶质母细胞瘤、星形细胞瘤、神经鞘瘤、室管膜瘤、髓母细胞瘤、脑膜瘤、鳞状细胞癌、垂体腺瘤、淋巴瘤、淋巴细胞性白血病、包括上述任何一种癌症的难治性形式(refractoryversion),或一种或多种上述癌症的组合。
具体实施方式
与现有技术的激动性cd40抗体相比,本发明满足了提供表现出降低的细胞毒性、导致更少的临床副作用,同时至少保持(如果没有增加的话)它们的信号传导能力和临床有效性的激动性的cd40mab的需求。
本发明的抗体或抗原结合片段提供了这些有利的特性,因为它们能够与人cd40受体特异性结合并且能够独立于fcγ介导的cd40受体交联而诱导cd40信号传导。
在优选的实施方案中,根据本发明的抗体是人源化igg1lala抗体、人源化igg1型抗体,其在人fc1区的234和235位具有至少两个丙氨酸氨基酸。因此,根据优选的实施方案,igg1lala包含人fc1区的突变l234a和l235a。
进一步优选的是,根据本发明的抗体是重组分子。
本发明提供一种激动性单克隆抗体或其抗原结合片段,其能够与人cd40受体结合并独立于fcγ介导的cd40受体交联而诱导cd40信号传导(也可参见实施例5、图6和下面的文字)。此外,当与野生型iggfcγ受体信号传导或与现有技术的抗体的fcγ信号传导相比时,根据本发明的抗体可以通过人fcγ受体表现出降低的或耗尽的信号传导能力。
在某些实施方案中,与野生型iggfcγ相比,本发明的激动性单克隆cd40抗体或其抗原结合片段可表现出对人fcγ受体的减少或耗尽的亲和力。根据优选的实施方案,本发明的抗体不结合fcγ受体-相应地,本发明的抗体不触发fcγ介导的cd40受体交联。
在优选的实施方案中,本发明的抗体至少包含在人igg1fc区的l234a和l235a或人igg4fc区的s228p和l235e处的氨基酸取代。
对于本发明进一步优选的是,抗体结合与cd40l结合位点重叠的cd40表位。图3和图16显示了所检测的人源化抗cd40igg1-lala抗体的这种表位重叠。还优选的是,cd40抗体与cd40l竞争结合cd40受体。因此,在表位竞争测定中,本发明的抗体与cd40l竞争。这种测定描述于实施例3和11中,并且本发明的抗体的实验结果描述于图3和16中。因此,根据进一步优选的实施方案,本发明的抗体抑制cd40l与cd40受体的结合。图19显示本发明的抗体不与cp-870,893竞争结合cd40。
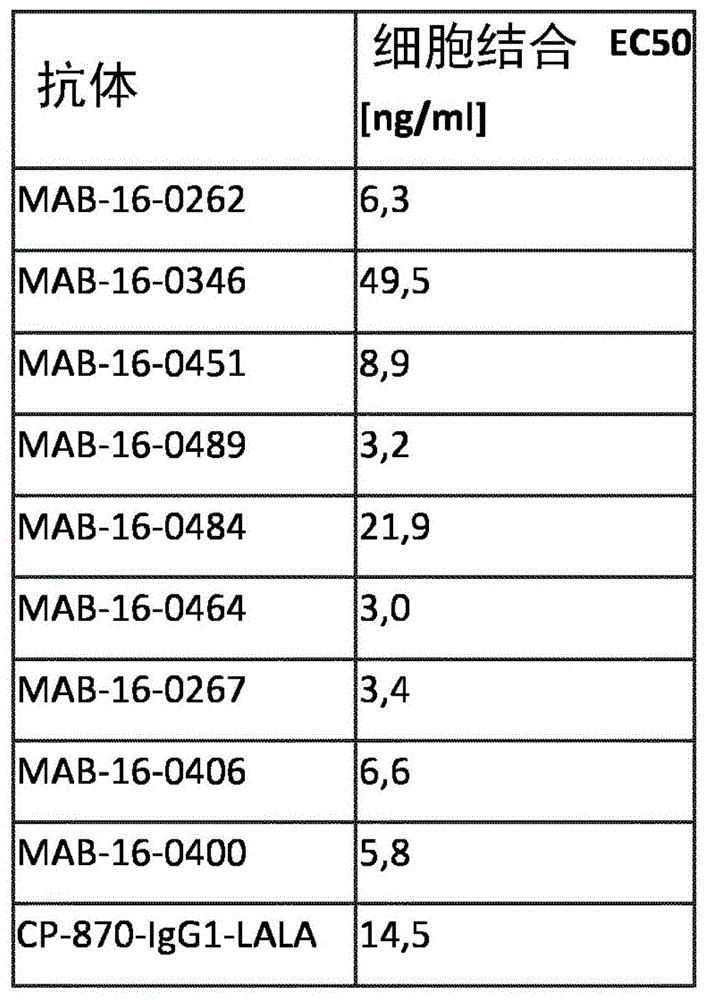
根据本发明的抗体具有与cd40受体非常高的结合活性。因此,在如实施例1中概述的细胞结合测定中,根据本发明的抗体表现出结合活性,ec50至多为49.5ng/ml。优选地,ec50小于25ng/ml,更优选小于15ng/ml、小于9ng/ml、7ng/ml、6ng/ml、5ng/ml、4ng/ml。最优选地,如实施例1所述和图1所示,在细胞结合测定中,ec50为3ng/ml。
根据本发明的人源化激动性抗cd40抗体可以表征为对可溶性人或食蟹猴cd40三聚体蛋白的生化亲和力(参见实施例13;图18)。本发明的抗体对于人cd40可以显示等于或小于15.7nm的kd值。本发明的抗体可以与食蟹猕猴cd40蛋白具有kd值等于或小于10.3nm的交叉反应。
此外,本发明的抗体能够高效诱导细胞nf-κb信号传导。实验总结(参见实施例2)在图2中示出,显示ec50结合值的范围为1127至6243ng/ml。ec50值显示了抗体诱导nf-kb信号传导的强大效能。
还应理解,本发明的抗体可以结合食蟹猴cd40。人源化抗cd40igg1-lala单克隆抗体与食蟹猴(macacafascicularis)的结合活性显示于使用重组食蟹猴cd40重组蛋白的elisa实验中(参见实施例4)。图4所示的ec50值表明抗体的有效结合。
本发明的抗体的另一个特征是它们可以激活人apc。例如,抗体可以激活选自由以下组成的组的细胞:树突细胞(dc)、b细胞、单核细胞和骨髓细胞。优选地,抗体激活dc。
在激活apc中这种有效的cd40激动活性不是由fcγ-受体介导的cd40蛋白的交联引起的(参见图5、6和7所示以及实施例5中所述的实验结果)。
这样,在一个实施方案中,在如实施例5中所述的树突细胞成熟测定中,本发明的抗体诱导il-12p40的释放。在这种测定中用本发明的抗体进行的实验结果显示于图5至7中。
根据另一个优选的实施方案,本发明的抗体诱导抗原呈递细胞的成熟,其通过至少等于在用抗体cp-870,893-igg2刺激时诱导的释放的il12p70释放、ec50值等于或小于208ng/ml来确定(图12和14)。此外,本发明的抗体诱导抗原呈递细胞的成熟,其通过诱导cd86至少7.5倍和ec50等于或小于148ng/ml来确定(图11和13)。
优选地,抗体诱导il12p40从单核细胞来源的dc中的释放,其至少等于在用抗体cp-870,893-igg2刺激时诱导的释放(参见图6)。如前所述,这种诱导dc成熟的潜力不是由通过fc受体的信号传导引起的。本发明的人源化抗cd40igg1-lala单克隆抗体以不依赖fcγ受体的方式有效诱导单核细胞来源的树突细胞活化(参见实施例5和图6)。图6说明由cp-870,893变体诱导的il12p40释放的水平与变体结合fc受体的能力相关(igg1-lala<igg2<igg1<igg1-v11)。惊人地,本发明的人源化抗cd40igg1-lala单克隆抗体的刺激导致不依赖fc的il12p40分泌水平,其覆盖甚至超过了cp-870,893变体产生的范围。因此,本发明的抗cd40抗体为原代单核细胞来源的树突细胞提供有效的激动活性,而无需fcγ-受体介导的cd40蛋白交联。
此外,本发明的抗体在其活化方面是非常特异性的。它们不诱导炎症性细胞因子例如tnf-α的一般释放(参见实施例6和图8)。
本发明的抗体的另一个特征是细胞表面上抗体清除的减少。已知cp-870,893与细胞上的cd40受体结合后会内化。临床研究表明,cp-870,893可迅速从患者体内清除,估计半衰期少于6小时,这反映了患者体内大的cd40受体库(sink),这可能是由于细胞内化所致(rüter等人2010)。
本发明的抗体在允许内吞和内化的条件下保留在细胞表面上(参见实施例7和图9)。相反,在允许内吞和内化的条件下,cp-870,893变体没有保留在细胞表面上(图9)。
应理解,根据本发明的抗体对肿瘤细胞死亡具有间接的(免疫介导的)作用。因此,抗体对肿瘤细胞表现出间接的免疫细胞介导的细胞毒性作用。
在一个具体的实施方案中,根据本发明的抗体不通过adcc、adcp或cdc的机制导致表达cd40的免疫细胞的耗尽。
因此,总而言之,本发明的抗体可以通过以下方式来进一步表征
(a)不与fcγ受体结合;
(b)具有等于或小于约49.5ng/ml的ec50值的cd40细胞结合亲和力;
(c)具有等于或小于约15.7nm的kd值;
(d)与食蟹猴cd40具有kd值等于或小于约10.3nm的交叉反应性;
(e)通过结合cd40抑制cd40l;
(f)防止cd40l介导的功能的协同作用和累加作用;
(g)诱导抗原呈递细胞的成熟,其通过至少等于用抗体cp-870,893-igg2刺激时诱导的释放的il12p70释放、ec50值等于或小于约208ng/ml来确定,
和/或通过在树突细胞上诱导cd86至少7.5倍且ec50等于或小于约148ng/ml确定;和/或
(h)将细胞表面上cd40的水平降低至比cp-870,893更少的程度。
根据优选的实施方案,本发明的抗体的特征在于具有至少一种、两种、三种、四种、五种、六种、七种或八种或全部上述特性(a至h)。
由于本发明的抗体的有利性质,它们能够抑制人肿瘤的生长。
在某些实施方案中,根据本发明的抗体可包含选自包含cdr区的vh区的组的vh区,所述cdr区选自由以下组成的组:seqidno:29+n的cdr1h区、seqidno:43+n的cdr2h区和seqidno:57+n的cdr3h区,其中n是选自由0至13组成的组的数字,以及选自包含cdr区的vl区的组的vl区,所述cdr区选自由以下组成的组:seqidno:71+m的cdr1l区、seqidno:85+m的cdr2l区和seqidno:99+m的cdr3l区,其中m是选自由0至13组成的组的数字,并且其中cdr可包含根据本发明不减弱其活性的任何一种或多种氨基酸突变。
优选地,抗体包含选自包含cdr区的vh区的组的vh区,所述cdr区选自由以下组成的组:seqidno:29+n的cdr1h区、seqidno:43+n的cdr2h区和seqidno:57+n的cdr3h区,其中n是选自由0至13组成的组的数字,和选自包含cdr区的vl区的组的vl区,所述cdr区选自由以下组成的组:seqidno:71+m的cdr1l区、seqidno:85+m的cdr2l区和seqidno:99+m的cdr3l区,其中m是选自由0至13组成的组的数字。
在某些实施方案中,根据本发明的抗体可包含选自包含cdr区的vh区的组的vh区,所述cdr区选自由以下组成的组:seqidno:29+n的cdr1h区、seqidno:43+n的cdr2h和seqidno:57+n的cdr3h区,和选自包含cdr区的vl区的组的vl区,所述cdr区选自由以下组成的组:seqidno:71+n的cdr1l区、seqidno:85+n的cdr2l区和seqidno:99+n的cdr3l区,其中n是选自由0至13组成的组的数字,并且其中cdr可包含根据本发明不减弱其活性的任何一种或多种氨基酸突变。
优选地,cdr与它们各自的seqidno具有至少91%,优选92%、93%、94%、95%、96%、97%、98%或99%的序列同一性。
在另一个实施方案中,根据本发明的抗体或抗原结合片段包含重链可变(vh)区,其与选自由seqidno:1至14的vh区组成的组的vh区具有至少60%同一性、优选至少70%同一性、更优选至少80%同一性、更优选至少85%同一性。
优选地,所述抗体包含重链可变区(vh)序列,其与选自seqidno:1至14的vh序列的组的氨基酸序列具有至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列同一性。
在某些实施方案中,相对于参考序列具有至少60%、70%、80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性的vh序列包含取代(例如,保守取代)、插入或缺失,从而抗体保留了根据本发明与各自的抗原特异性结合的能力。
本发明还涵盖包含重链可变区(vh)的抗体,所述重链可变区包含选自seqidno:1至14的组的氨基酸序列。
优选地,重链可变区(vh)序列是seqidno:1,可选地seqidno:2,或seqidno:3、seqidno:4、seqidno:5、seqidno:6、seqidno:7、seqidno:8、seqidno:9、seqidno:10、seqidno:11、seqidno:12、seqidno:13、seqidno:14。
本发明还涉及一种抗体,其包含轻链可变(vl)区,所述轻链可变区与选自由seqidno:15至28的vl区组成的组的vl区具有至少60%同一性、优选至少70%同一性、更优选至少80%同一性、更优选至少85%同一性。
优选地,所述抗体包含vl序列,所述vl序列与选自seqidno:15至28的vl序列的组的氨基酸序列具有至少86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列同一性。
在某些实施方案中,相对于参考序列具有至少60%、70%、80%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性的vl序列包含取代(例如,保守取代)、插入或缺失,从而抗体保留了根据本发明与各自的抗原特异性结合的能力。
本发明还涵盖包含轻链可变区(vl)的抗体,所述轻链可变区包含选自seqidno:15至28的组的氨基酸序列。
优选地,轻链可变区(vl)序列是seqidno:15,可选地seqidno:16,或seqidno:17、seqidno:18、seqidno:19、seqidno:20、seqidno:21、seqidno:22、seqidno:23、seqidno:24、seqidno:25、seqidno:26、seqidno:27或seqidno:28。
在某些实施方案中,在所述vl序列中共有1至10个氨基酸已经被取代、插入和/或缺失。在其他实施方案中,在所述vh序列中共有1至10个氨基酸已经被取代、插入和/或缺失。在某些实施方案中,在每个所述vh或vl序列中,共有1至10个氨基酸已经被取代、插入和/或缺失。所述取代、插入或缺失可发生在cdr之外的区域(即,在fr中)。
本发明还包含可以根据本领域已知方法产生的亲和成熟抗体。marks等人bio/technology10:779-783(1992)描述了通过vh和vl结构域改组的亲和成熟。cdr和/或框架残基的随机诱变由以下描述:barbas等人,procnat.acad.sci,usa91:3809-3813(1994);schier等人,gene169:147-155(1995);yelton等人,j.immunol.155:1994-2004(1995);jackson等人,j.immunol.154(7):3310-9(1995);和hawkins等人,j.mol.biol.226:889-896(1992)和wo2010/108127。
本发明还涵盖一种抗体,其包含vh区和vl区,所述vl区和vl区包含选自图10中所列抗体的,即包含抗体mab-16-0283、mab-16-0377、mab-16-0267、mab-16-0386、mab-16-0451、mab-16-0346、mab-16-0325、mab-16-0388、mab-16-0464、mab-16-0262、mab-16-0406、mab-16-0484、mab-16-0400、mab-16-0489的组的抗体的,分别的cdr1、cdr2和cdr3区。
本发明还涵盖包含seqidno:1和15、或seqidno:2和16的抗体。根据本发明的抗体也可以包含seqidno:3和17、或seqidno:4和18、或seqidno:5和19、或seqidno:6和20、或seqidno:7和21、或seqidno:8和22、或seqidno:9和23、或seqidno:10和24、或seqidno:11和25、或seqidno:12和26。可选地,根据本发明的抗体包含seqidno:13和27、或seqidno:14和28。
在另一方面,本发明的抗体用于治疗患有癌症的患者。
所述癌症可以是选自包含以下癌症的组的一种或多种类型的癌症:胰腺癌、晚期胰腺癌肺癌、非小细胞肺癌(nscl)癌症、细支气管肺泡细胞肺癌、骨癌、胰腺癌、皮肤癌、头或颈癌、皮肤或眼内黑色素瘤、卵巢癌、直肠癌、肛区癌、肠胃癌、胃癌、结肠癌、乳腺癌、肾癌、霍奇金淋巴瘤、肝癌、胆囊癌、膀胱癌、前列腺癌、甲状腺癌、唾液腺癌或子宫癌。
在某些实施方案中,癌症是实体瘤。
在一些实施方案中癌症是表达cd40的癌症也是可能的。然而,这对于本发明的抗体的有效功能不是必需的。
将进一步理解,本发明的抗体可以用作患者中癌症的单独治疗或作为联合治疗的一部分(进一步的治疗可以是药物细胞毒性或细胞生长抑制性剂(pharmaceuticalcytotoxicorcytostaticagent)、放射疗法、靶向疗法和/或手术)。
因此,患者还可以接受针对癌症的一种或多种其他治疗方法,例如药剂(例如细胞毒性或细胞生长抑制性剂、靶向治疗)、放射治疗和/或手术。
因此,在一些实施方案中,根据本发明的抗体与细胞毒性或细胞生长抑制性剂、放射疗法、靶向疗法和/或免疫疗法联合用于治疗癌症。
本发明的抗体也可以用于治疗对细胞毒性或细胞生长抑制剂、放射疗法、靶向疗法和/或免疫疗法反应不足和/或抗药性不足的患者。
所述放射疗法可以选自包含以下疗法的组:外部束放射疗法、接触x射线近距离放射疗法、近距离放射疗法、全身放射性同位素疗法或术中放射疗法。
根据本发明的细胞毒性或细胞生长抑制性抗癌剂可以选自包含以下的组:紫杉烷类、蒽环类、烷基化剂、组蛋白脱乙酰基酶抑制剂、拓扑异构酶抑制剂、激酶抑制剂、核苷酸类似物、肽抗生素和铂基试剂。
优选地,靶向的抗癌剂用于靶向治疗,并且选自以下之一或其组合:抗egfr化合物,例如西妥昔单抗(cetuximab)、吉非替尼(gefitinib)、埃罗替尼(erlotinib)、拉帕替尼(lapatinib)、帕尼单抗(panitumumab)、抗her2化合物例如曲妥珠单抗(trastuzumab)、曲妥珠单抗-美坦新偶联物(ado-trastuzumabemtansine)、帕妥珠单抗(pertuzumab)、靶向vegf的化合物例如贝伐单抗(bevacizumab)、阿柏西普(aflibercept)和哌加他尼(pegaptanib)和酪氨酸激酶抑制剂例如舒尼替尼(sunitinib)、帕唑帕尼(pazopanib)、阿昔替尼(axitinib)、凡德他尼(vandetatanib)、卡博替尼(cabozantinib)和瑞戈非尼(regorafinib)。
如果患者正在接受免疫治疗,则这可以是免疫检查点抑制,也可以使用一种或多种免疫检查点抑制剂。一种或多种免疫检查点抑制剂可选自包含以下的组:抗pd-l1、抗pd-1、抗ctla-4、抗cd137、抗lag-3、抗tim-3、抗ox40和/或抗gitr。
本发明的抗体也可以用于与特异性结合人pd-l1、ctla-4、lag-3、tim-3、cd137、ox40、gitr的抗体联合使用,和/或与药物纳武单抗(nivolumab)、派姆单抗(pembrolizumab)、urelumab,utomilumab、阿特朱单抗(atezolizumab)、阿维鲁单抗(avelumab)、杜鲁伐单抗(durvalumab)、曲美单抗(tremelimumab)、伊匹单抗(ipilimumab)联合使用。
在根据本发明的一些实施方案中,抗体以每周至每月的给药方案用于癌症的治疗。
本发明的特殊优点之一是,由于它们在fc区中的突变,与现有技术的抗体相比,该抗体表现出更少的剂量或治疗限制性毒性。如果有的话,这些抗体仅会在非常有限的范围内引起cd40抗体的典型副作用。此类副作用是选自包含以下的组的病症:细胞因子释放综合征、血栓形成、脑栓塞、转氨酶升高、淋巴细胞减少、疲劳、周围神经病变、脱发、便秘、恶心和中性粒细胞减少症。
在另一方面,本发明涉及药物组合物,所述药物组合物包含药学上可接受的载体和治疗有效量的根据本发明的抗体。
根据本发明的药物组合物可以用于治疗患有癌症的患者。这样的癌症可以是实体瘤。癌症还可以选自由以下组成的组:胰腺癌、晚期胰腺癌肺癌、非小细胞肺癌(nscl)癌、细支气管肺泡细胞肺癌、骨癌、胰腺癌、皮肤癌、头或颈癌、皮肤或眼内黑色素瘤、卵巢癌、直肠癌、肛区癌、肠胃癌、胃癌、结肠癌、乳腺癌、肾癌、霍奇金淋巴瘤、肝癌、胆囊癌、膀胱癌、前列腺癌、甲状腺癌、唾液腺癌或子宫癌。
组合物还可以与化学疗法、放射疗法、靶向疗法和/或免疫疗法联合用于癌症的治疗。所述免疫疗法可以是免疫检查点抑制。
用所述组合物治疗的患者可对化学疗法、放射疗法、靶向疗法和/或免疫疗法没有足够的反应和/或抗性。
根据本发明的药物组合物还可与一种或多种细胞毒性化合物、细胞生长抑制性化合物或靶向的抗癌化合物联合用于癌症的治疗。
它可以与一种或多种免疫检查点抑制剂联合用于癌症的治疗,其中所述免疫检查点抑制剂可以选自由以下组成的组:抗pd-l1、抗pd-1、抗ctla-4、抗-cd137、抗lag-3、抗tim-3、抗ox40和/或抗gitr。
组合物还可以与特异性结合人pd-l1、ctla-4、lag-3、tim-3、cd137、ox40、gitr的抗体联合使用和/或与药物纳武单抗、派姆单抗、urelumab,utomilumab、阿特朱单抗、阿维鲁单抗、杜鲁伐单抗、曲美单抗、伊匹单抗联合使用。
它也可以以每周一次至每月一次的给药方案用于治疗癌症。
患者将特别理解,如果有的话,根据本发明的抗体和组合物仅会在非常有限的范围内表现任何剂量或治疗限制毒性,并且重要的是,其扩展程度小于现有技术的抗体和组合物。
在另一方面,本发明还涉及治疗方法,其包含将有效量的根据本发明的抗体施用于有需要的个体。这样的人可以是患有癌症的患者。因此,本发明还涉及治疗癌症的方法,其中所述癌症可以是实体瘤。
施用于有需要的个体的步骤可包含局部施用,例如局部施用至患者的肿瘤(例如,肿瘤内或肿瘤周围)。
由于本发明的基于抗体的试剂适用于治疗任何类型的癌症,其中cd40活化可为其提供治疗益处,因此包含施用所述抗体的方法也适用于治疗任何类型的癌症,其中cd40活化可为其提供治疗益处。
例如,癌症可以选自由以下组成的组:前列腺癌;乳腺癌;大肠癌;胰腺癌;卵巢癌;肺癌;宫颈癌;横纹肌肉瘤;神经母细胞瘤;多发性骨髓瘤;白血病、急性淋巴细胞白血病、黑素瘤、膀胱癌和胶质母细胞瘤。
将进一步理解,根据本发明的治疗方法可以包含向患者单独施用本发明的基于抗体的试剂或作为联合治疗的一部分(其中进一步治疗可以是药物细胞毒性或细胞生长抑制性剂或放射疗法、靶向疗法和/或手术)。
实际上,如上所述的本发明的抗体的所有特征和有利特性也反映并包含在根据本发明的抗体的治疗方法和用途中。
实施例
下列实施例结合附图和表格用于说明本发明。
实施例1:抗cd40抗体的细胞结合
为了确定人源化抗cd40igg1-lala单克隆抗体与表达cd40的细胞的结合的效力,将hek-blue-cd40ltm(invivogen)细胞在细胞培养处理的透明底384孔平板中以1000个细胞/孔的细胞密度接种在25μl含10%fbs的dmem中。在5μl培养基中,将抗体添加至最终浓度范围为1.25μg/ml至0.01ng/ml。24小时后,将细胞用25μl洗涤缓冲液(pbs,0.05%tween)洗涤3次,然后在20μl培养基中加入0.8μg/ml浓度的alexa-fluor-488缀合的山羊抗人igg(jacksonlaboratories)。4小时后,加入5μlhoechst染料至培养基中使终浓度为5μg/ml。荧光细胞结合信号通过使用cellinsight自动化高内涵成像仪(thermofisherscientific)进行测定。拟合曲线和ec50计算通过使用excel(microsoft)和xlfit(idbs)获得。图1总结了ec50结合值范围为3至49.5ng/ml。
实施例2:抗cd40激动抗体诱导细胞nf-κb信号传导
人源化抗cd40igg1-lala单克隆抗体的激动活性通过刺激hek-blue-cd40ltm(invivogen)细胞来进行检测,所述细胞带有nf-κb诱导型分泌性胚胎碱性磷酸酶(seap)基因构建体。在含有10%fbs的20μldmem中将25000个细胞/孔接种至细胞培养处理的透明底384孔板中,并培养过夜。然后将抗体添加到5μl体积的培养基中,最终浓度范围为20至0.013μg/ml。在37℃和5%co2下培养6小时后,将每个孔的5μl培养基上清液转移至含有20μl的2xquanti-bluetm试剂(invivogen)的白色、透明底384孔板中。在37℃和5%co2下培养1小时后,进行620波长处的光密度测量,反映了nf-κb依赖性磷酸酶分泌的激活。拟合曲线和ec50计算通过使用excel(microsoft)和xlfit(idbs)获得。图2中的ec50值表明抗cd40抗体在hek-blue-cd40ltm细胞系中诱导nf-kb信号传导的能力。
实施例3:与cd40l结合的竞争
人源化抗cd40igg1-lala单克隆抗体与cd40l结合cd40的竞争通过使用elisa测定法进行测定。在25μlpbs的体积中将cd40l涂覆至384孔nunctmmaxisorptm板的表面上,并且在室温下以1μg/ml的浓度放置一小时。在室温下于elisa缓冲液(pbs,0.5%bsa,0.05%tween)中将抗体以5μg/ml的浓度与1.7μg/ml浓度的重组cd40蛋白在40μl的总体积中进行预培养1.5小时。将nunctmmaxisorptm板用洗涤缓冲液(pbs,0.1%tween)洗涤3次,并在室温下用pbs、2%bsa、0.05%tween封闭1小时。在洗涤缓冲液中洗涤3次后,将25μl抗体-cd40复合体添加至nunctmmaxisorptm板孔中,并在室温下培养1小时。在洗涤缓冲液中洗涤3次后,将孔与25μl的1:2000稀释的抗人过氧化物酶连接的、来自山羊的物种特异性f(ab)2片段(abdserotec)在elisa缓冲液中于室温培养1小时。用洗涤缓冲液洗涤孔六次,并添加30μl/孔的tmb底物溶液(invitrogen)。在室温下放置10分钟后,每孔添加30μl终止液(1mhcl),并使用tecanm1000微孔板读数器测定450和620nm波长的吸光度。用cp-870,893培养的样品的elisa信号表明缺乏与cd40l的竞争,而根据本发明的人源化抗cd40igg1-lala单克隆抗体则与cd40l竞争结合cd40(见图3)。
实施例4:食蟹猴cd40结合活性
人源化的抗cd40igg1-lala单克隆抗体与食蟹猴cd40蛋白的结合在生化elisa中进行检测。重组cyno-cd40蛋白(acrobiosystems)在384孔nunctmmaxisorptm板中以在pbs中0.5μg/ml的浓度在室温下培养一小时。用洗涤缓冲液(pbs,0.1%tween)洗涤3次后,在室温下用pbs、2%bsa、0.05%tween封闭板一小时。再用洗涤缓冲液洗涤板3次,并将在pbs、0.5%bsa、0.05%tween中的浓度范围为500至0.03ng/ml的抗体在室温下培养1小时。在洗涤缓冲液中洗涤3次后,在室温下将12.5μl的1:3000稀释的抗人过氧化物酶连接的、来自山羊的物种特异性f(ab)2片段(abdserotec)在elisa缓冲液中培养1小时。用洗涤缓冲液洗涤孔六次,并加入15μl/孔的tmb底物溶液(invitrogen)。在室温下放置10分钟后,每孔加入15μl终止溶液(1mhcl),并使用tecanm1000酶标仪测量450和620nm波长的吸光度。拟合曲线和ec50计算通过使用excel(microsoft)和xlfit(idbs)获得。如图4所示,大多数抗体以8至31.8ng/ml的ec50值与cyno-cd40结合。
实施例5:诱导树突细胞成熟
a.产生单核细胞来源的树突细胞以检测人源化抗cd40igg1-lala单克隆抗体刺激树突细胞成熟的能力,通过分泌il12p40细胞因子来测量。将来自不同供体的人血沉棕黄层制剂用于体外区分树突细胞与单核细胞。将来自巴伐利亚红十字会的血沉棕黄层用dpbs以1:4稀释,并在ficoll-paque(gehealthcare)密度梯度上铺层。离心后,用dpbs将相间的外周血单核细胞(pbmc)洗涤3次,并根据制造商的说明使用磁性cd14microbeads(miltenyibiotec)分离单核细胞。在t-175细胞培养瓶中,将单核细胞以1.2x106个细胞/ml的细胞密度在含有10%fcs、1xpen/strep、1xl-谷氨酰胺、50ng/ml重组人gm-csf(r&dsytems)和10ng/ml重组人il-4(r&dsystems)的rpmi-1640中培养。每48小时,将90%的培养基更换为新鲜的含细胞因子的培养基。在第5天,收集体外分化的未成熟树突细胞(idc),并在100μl的相同培养基中以106个细胞/ml的细胞密度分布至细胞培养96孔板中。
b.在一项实验中,通过添加浓度为5μg/ml的抗cd40抗体刺激idc。刺激后48h,使用商购可得的elisa试剂盒(r&dsystems)根据制造商的说明对培养基上清液中分泌的il12p40细胞因子进行定量。图5显示人源化抗cd40igg1-lala单克隆抗体在不同程度上刺激单核细胞来源的树突细胞释放il12p40,范围从小于1ng/ml到大于24ng/ml,而非cd40结合对照igg1-lala抗体不会导致可检测水平的il12p40的刺激。
c.本发明的人源化抗cd40igg1-lala单克隆抗体不能结合在单核细胞来源的树突细胞上表达的fcγ受体。为了将这些抗体与具有不同的fcγ受体结合活性的抗体进行比较,我们构建了含有人igg1、igg1-lala、igg2和igg1-v11fc部分的参照cp-870,893抗cd40抗体,以及刺激具有不同浓度的这些抗体的单核细胞来源的树突细胞。igg1-v11形式在重链fc部分包含四个突变(g237d、h268d、p271g、a330r)。已经描述了这些突变以选择性增加对fcγ-受体riib的亲和力(mimoto等人,2013)。图6说明cp-870,893变体以fc依赖性方式刺激il12p40释放(igg1-lala<igg2<igg1<igg1-v11)。惊人地,本发明的人源化抗cd40igg1-lala单克隆抗体的刺激导致不依赖fc的il12p40分泌水平,其覆盖并且甚至超过cp-870,893变体产生的范围。因此,本发明的抗cd40抗体对原代单核细胞来源的树突细胞提供有效的激动活性,而无需fcγ受体介导的cd40蛋白交联。
d.在另一个实验中,人源化抗cd40igg1-lala单克隆抗体诱导单核细胞来源的树突细胞分泌il12p40的能力是通过以0.005到10μg/ml的抗体浓度刺激来确定的。图7显示,不同抗体的ec50值介于380至743ng/ml之间。
实施例6:高密度pbmc测定中的tnf-α释放
为了确定人源化抗cd40igg1-lala单克隆抗体是否诱导血细胞中细胞因子的一般释放,根据
实施例7:细胞脉冲追踪测定
使用hek-blue-cd40ltm细胞(invivogen)在脉冲追踪测定中分析抗cd40抗体的细胞结合和内化动力学。在含有10%fcs的dmem培养基中将2000个细胞/孔接种在两个具有透明底的黑色384孔板中。过夜培养后,将本发明的抗cd40抗体和如上所述的cp-870,893fc变体抗体以0.8μg/ml的浓度添加至一个板中,并在37℃和5%co2下培养15分钟。随后,将两个板用细胞洗涤缓冲液(pbs0.05%tween)洗涤3次,并在培养基中培养1小时。在最后15分钟内,将抗体以0.8μg/ml的浓度添加到平板2中。然后将两块板用细胞洗涤缓冲液洗涤三次,置于冰上,并与0.8μg/ml的二次抗人alexa-fluor-488偶联抗体(jacksonlaboratories)和5μg/mlhoechst染色剂(invitrogen)一起在冰上培养30分钟。使用cellinsight高内涵成像仪(thermofisherscientific)对细胞表面荧光信号进行定量。图9显示,在允许内化的条件下培养一小时后,cp-870,893变体抗体的细胞表面信号大大降低。相反,在相同条件下许多根据本发明的抗cd40抗体的培养仅导致细胞表面信号的轻微降低,其表明有限的内化速率。
实施例8:人源化抗cd40抗体的基因报道基因诱导与树突细胞成熟活性的相关性
a.分别如实施例2和5所述进行细胞基因报告基因(hek-blue-cd40ltm)和树突细胞(dc)成熟测定。对于dc测定,以5μg/ml使用抗体,在hek-blue-cd40ltm基因报告基因测定中的浓度范围为13至20000ng/ml。图10比较了在用5μg/ml抗体刺激后在基因报告基因测定中观察到的最大诱导与dc释放的il12p40细胞因子。虽然所有88种人源化的抗cd40-igg1-lala抗体均以相似的程度诱导基因报告基因表达,但一些抗体显示dc释放的il12p40的非常高的刺激。hek-blue-cd40ltm细胞不表达fcγ受体。该测定捕获与fcγ受体结合无关的cd40抗体的基本激动活性。dc表达fcγ受体,并且抗cd40抗体例如cp-870,893的激动活性取决于fcγ受体结合(实施例5)。然而,尽管88种igg1-lala抗体中的大多数缺乏dc的强激活,但一小部分可以诱导非常强的dc激活,而无需fcγ受体介导的交联。因此,其对原代树突细胞的活性与fcγ受体交联无关的高激动性抗cd40抗体是一种罕见的情况,并且鉴定需要筛选大量具有基本激动活性的候选抗体。
实施例9:激动性抗cd40抗体刺激树突状细胞上的共刺激受体和细胞因子释放
a.为了检测人源化、激动性抗cd40igg1-lala抗体在刺激共刺激受体表达和dc释放的炎性细胞因子的活性,如实施例5所述,由三个独立的供体产生未成熟的单核细胞来源的dc(idc)。用浓度为2μg/ml的抗体或浓度为20μg/ml的cd40l(r&dsystems6245-cl-050)处理idc48小时。收获刺激的成熟dc,使用针对hla-dr、cd86、cd80、cd83、cd54和cd95(均来自miltenyibiotech)的荧光团标记抗体染色,并在bdfacsverse设备上通过流式细胞术进行分析。图11显示受体表达刺激,其诱导程度是同种型抗体对照治疗的诱导倍数。数据显示共刺激受体,特别是cd86的强烈诱导。与本发明的抗体mab-16-0262、mab-16-0451、mab-16-0464和mab-16-0406相比,cp-870,893显示总体较低的活性。当使用含有igg1-lala恒定部分的cp-870,893变体时,观察到活性进一步降低,证实了该抗体的fcγ受体结合依赖性。
b.图12显示根据制造商的说明,使用bd人炎性细胞计数珠阵列试剂盒(bd#551811)在dc培养物上清液中进行细胞因子测定的结果。本发明的抗体mab-16-0262、mab-16-0451、mab-16-0464和mab-16-0406显示非常高的il-12-p40和il-12p70释放水平,而其他细胞因子例如tnf-α、il-1β、il-10和il-6产生和分泌的程度要小得多。与用本发明的抗体观察到的释放相比,用cp-870,893igg2和igg1变体处理的dc的il-12p40和il12p70释放显著更低。在类似的实验中,将来自三个独立供体的体外分化的idc用激动性抗cd40抗体以10000至5ng/ml范围内的浓度处理48小时。受体表达和细胞因子释放按如上所述分析。拟合曲线和ec50计算通过使用excel(microsoft)和xlfit(idbs)获得。图13和14所示的数据例证了在一个供体中观察到的剂量依赖性效果。另外两个供体的结果显示出定性相似的效果。总之,通过ec50值确定的本发明人源化抗cd40抗体的效力类似于cp-870,893,而最大的诱导作用,特别是对il-12细胞因子释放和共受体表达的最大诱导作用显著大于cp-870,893的最大诱导作用。
实施例10:激动性抗cd40抗体对b细胞上的共刺激受体的刺激
a.人源化抗cd40igg1-lala抗体对共刺激受体的刺激作用也已在b细胞上进行检测。通过ficoll密度梯度离心从人血沉棕黄层中分离出三个不同供体的pbmc,并根据制造商的说明,使用b细胞分离试剂盒ii(miltenyibiotech)通过负磁富集纯化未接触的b细胞。将100μl的rpmi-1640+10%人ab血清中的2x105个b细胞用浓度范围从500到0.2ng/ml的抗体刺激48小时。收集刺激的b细胞,使用针对hla-dr、cd86和cd80的荧光团标记抗体染色(均来自miltenyibiotech),并在bdfacsverse设备上通过流式细胞仪进行分析。图15显示一个供体的b细胞中受体表达的剂量依赖性刺激,其诱导程度是同种型抗体对照治疗的诱导倍数。拟合曲线和ec50计算通过使用excel(microsoft)和xlfit(idbs)获得。结果表明,本发明的抗体还刺激b细胞上的共刺激受体,然而,上调水平低于dc中观察到的水平。
实施例11:结合至细胞上cd40的cd40l与抗cd40抗体竞争
a.对存在cd40l的情况下的本发明的抗cd40抗体与hek-blue-cd40ltm细胞的结合进行检测,以验证抗体是否结合至细胞表面上cd40的cd40l结合位点。将hek-blue-cd40ltm细胞与它们的ec90结合浓度的抗体在40℃下预培养30分钟。加入浓度为10000至9.8ng/ml范围内的含有小鼠igg2afc-tag(abbiosciences)的cd40l,并将细胞在4℃下培养60分钟。使用二次dylight405缀合的抗小鼠igg和alexafluor488缀合的抗人igg(jacksonlaboratories)检测与细胞表达的cd40结合的抗cd40抗体和cd40l,并使用facsverse仪器(bd)分析。图16a显示抗cd40浓度的稳定结合,除了cp-870,893,其抗体结合信号在较高的cd40l浓度下略有降低。cd40l以剂量依赖性方式与细胞结合,并且cp-870,893不会显著干扰cd40l结合(图16b)。相反,本发明的抗体强烈阻止cd40l与细胞表达的cd40结合,表明这些抗体与cd40的cd40l结合位点结合并阻止cd40l结合cd40(图16b)。
实施例12:激动性cd40抗体与cd40l组合诱导fasr(cd95)死亡受体表达
a.为了检测cd40l对抗cd40抗体介导的对细胞的作用的干扰,将ramos细胞单独用cd40l处理或用cd40l与激动性抗cd40抗体联合处理。将ramos细胞以1.25x106个细胞/ml的细胞密度接种在含有10%fcs的rpmi中的96孔板中。将抗体以10μg/ml的浓度添加至孔中,并将板在37℃、5%co2、95%湿度下培养10分钟。然后将cd40l(r&dsystems)添加至一些孔中,使其最终浓度为10μg/ml,并将板在37℃、5%co2、95%湿度下培养过夜。用dpbs洗涤细胞,并用针对cd95的fitc标记的抗体(miltenyibiotech)染色。图17显示,通过添加cd40l,cp-870,893对cd95的诱导大大增加,而本发明所有检测的抗cd40抗体的共同处理降低了cd40l的作用。数据表明,结合与cd40结合的cd40l的本发明的激动性抗cd40抗体防止cd40l的协同作用和累加作用,因此允许控制的和安全的药理学。
实施例13:人源化、激动性抗cd40igg1-lala抗体的亲和力
a.本发明抗体的生化亲和力是通过表面等离子共振测量来确定的。将抗体通过抗人fc抗体可逆地固定至cm5传感器芯片表面上。在biacoret200spr仪器上分析固定化抗体与可溶性人或食蟹猴cd40单体蛋白(acrobiosystems)相互作用的动力学。动力学数据采用langmuir1:1结合模型测定。图18显示抗体mab-16-0451和mab-16-0464的kd分别为1.2和2.6,而mab-16-0262和cp-870,893的kd值分别为15.7或8.9。使用食蟹猴cd40蛋白产生的kd值显示出相似的亲和力。
实施例14:抗cd40抗体与cd40的竞争性结合
a.为了检测本发明的人源化、激动性抗cd40抗体是否结合至cd40分子上的重叠区域,进行竞争性结合elisa。在pbs中将抗体以625ng/ml的浓度涂覆于384孔maxisorp板上60分钟,然后用pbs、2%bsa、0.05%tween封闭70分钟。在elisa缓冲液(pbs,0.5%bsa,0.05%tween)中将所有抗体分别以10μg/ml的浓度与330ng/ml带有his标签的cd40重组蛋白(acrobiosystems)和4μg/ml过氧化物酶偶联的抗his检测抗体(sigma-aldrich)一起在试管中培养60分钟。用pbs、0.1%tween洗涤板3次,然后将抗体/his-cd40/抗-his-过氧化物酶混合物添加至板的孔中。将板培养60分钟。用pbs洗涤孔六次,并加入0.1%tween和15μl/孔的tmb底物溶液(invitrogen)。用15μl/孔的终止液(1mhcl)终止反应,并使用tecanm1000酶标仪测定450和620nm波长的吸光度。图19显示本发明的抗体不与cp-870,893竞争结合cd40。然而,每种抗体与本发明的任何其他抗体竞争,这表明这些抗体结合至cd40上的相同区域。重要的是,由于本发明抗体的激动活性涵盖广泛的范围(实施例9和10),这表明激动性抗cd40抗体的互补位主要决定抗cd40抗体的激动活性。
实施例15:igg1-lala抗cd40抗体的抗体介导的效应子功能
a.为了检测igg1抗体的恒定部分中的lala突变在减弱例如adcc等抗体介导的效应子功能方面的潜力,根据制造商的说明,已经采用例如基于jurkat效应细胞报告细胞系的检测方法(promegaadccbioassay,#g701a),并使用hek-blue-cd40ltm细胞作为靶细胞。在白色平底384孔试验板的每一孔中、在25μl的dmem+10%fcs中接种5000个hek-blue-cd40ltm细胞,并在37℃、5%co2下培养20h。用含4%低iggfcs的8μlrpmi培养基更换培养基,然后将每孔4000个效应细胞添加至8μl相同的培养基中。最后,将含有igg1或igg1-lalafc部分两者之一的cp-870,893抗cd40抗体以10000至0.002ng/ml范围内的浓度添加至8μl培养基中。将板在37℃、5%co2下培养6小时。根据制造商的说明,使用bioglo萤光素酶测定试剂(promega)测定效应细胞的萤光素酶活性。使用tecanm1000酶标仪读取发光。用公式rlu(抗体处理-背景)/rlu(载体-背景)计算诱导倍数。通过使用excel(microsoft)和xlfit(idbs)获得拟合曲线。图20表明,igg1中的lala突变消除了效应细胞中fc受体介导的信号传导。
实施例16:激动性抗cd40抗体治疗在人源化小鼠模型中的安全性
为了评估安全性,采用干细胞人源化小鼠模型。nod/scid/γ(c)(null)fcrg-/-小鼠缺乏激活小鼠的fc受体。因此,在该模型中,人免疫细胞对治疗性抗体的fc受体结合不会受到小鼠fc受体结合的损害。在出生后的头24小时内,对小鼠进行1,4gy亚致死剂量的辐射。4-6小时后,用从脐带血中分离出的20000–50000个人造血干细胞对小鼠静脉内移植。移植后12周,通过外周血细胞的流式细胞术验证了人免疫细胞的存在。用3μg/gcp-870,893、mab-16-0451或同种型对照抗体对成功的人源化小鼠进行一次静脉注射。在处理前和抗体注射后的不同时间点测定体重和温度(图21)。数据显示,用cp-870,893处理的6只小鼠中有3只的体温显著降低,这些小鼠由于身体状况的严重损害而不得不处死。相反,用mab-16-0451处理的小鼠对体温没有显著的影响,也没有其他任何明显的身体状况受损迹象。这表明,缺乏fc受体结合活性的高激动性抗cd40-igg1-lala抗体可在无明显毒性迹象的情况下应用于治疗,而活性较低的激动性cp-870893-igg2抗体可在该模型中在体内表现出毒性作用。
附图说明
图1:细胞结合
检测了抗cd40igg1-lala单克隆抗体与hek-blue-cd40ltm细胞(invivogen)上细胞表达的cd40抗原的结合。ec50值表明检测抗体的有效结合。
图2:8点分析中的hek-blueec50测定
在基于细胞的nf-kb基因报告基因测定中检测了人源化抗cd40igg1-lala单克隆抗体的激动活性。将hek-blue-cd40ltm细胞(invivogen)与不同浓度的抗体培养24小时。ec50值表明了抗体诱导nf-kb信号传导的能力。
图3:cd40-配体表位竞争
为了检测人源化的抗cd40igg1-lala单克隆抗体是否结合与cd40l结合位点重叠的表位,进行了cd40l竞争elisa。将不同浓度的抗cd40抗体与cd40重组蛋白预培养以形成结合复合体。随后,将复合体添加至包被有重组cd40l的微量滴定板中。洗涤后,使用过氧化物酶连接的抗人f(ab)2抗体检测结合的cd40-抗cd40复合体。参照cp-870,893抗体的elisa信号表明与cd40l无竞争,因此与不同于cd40l结合位点的表位结合。数据表明,所检测的人源化抗cd40igg1-lala单克隆抗体结合与cd40l结合位点重叠的表位。
图4:食蟹猴cd40结合活性
使用重组食蟹猴cd40重组蛋白(acrobiosystems),在elisa中检测人源化抗cd40igg1-lala单克隆抗体与食蟹猴(macacafascicularis)的结合活性。ec50值指示抗体的有效结合。n.d.=在检测浓度范围内不可检测
图5a)b):树突细胞成熟的诱导
为了检测人源化的抗cd40igg1-lala单克隆抗体对原靶细胞的激动活性,分析了单核细胞来源的、未成熟树突细胞的成熟。将从单核细胞体外分化的未成熟树突状细胞以5μg/ml的浓度与激动性抗c40抗体培养48小时。随后通过生化elisa在培养基上清液中对源自树突细胞的分泌的il12p40进行定量。
图6:树突状细胞成熟
确定了人源化抗cd40igg1-lala单克隆抗体在不同抗体浓度下刺激树突细胞分泌il12p40的活性,并将该活性与带有不同fc部分(igg1、igg1-lala、igg2和igg1-v11)的cp-870,893抗体进行比较。将抗体与体外分化的未成熟树突细胞培养48小时。通过elisa测定il12p40释放。
图7:树突细胞成熟过程中抗cd40抗体的ec50测定
通过检测10–0.005μg/ml范围内的抗体浓度来测定树突细胞介导的il12p40分泌中人源化抗cd40igg1-lala单克隆抗体的ec50值。将抗体与体外分化的未成熟树突细胞培养48小时。通过elisa测定il12p40释放。
图8:细胞因子释放测定
将人源化抗cd40igg1-lala单克隆抗体以10μg/ml在高密度pbmc细胞因子释放测定中进行检测,以确定如tnf-α等炎性细胞因子的一般诱导作用。数据表明,与抗cd3(okt3)抗体相反,抗cd40抗体不会诱导显著的tnf-α分泌。
图9:抗体脉冲追踪细胞测定
在脉冲追踪细胞测定中检测抗体结合动力学和内化。将抗体以0.8μg/ml的浓度与hek-blue-cd40ltm细胞培养物培养15分钟。洗涤后,使抗体内化60分钟,然后再次洗涤细胞,并用alexa-488标记的二抗检测剩余的细胞表面定位的抗cd0抗体。在不允许内化的条件下,对细胞进行相似的处理,但是抗体仅培养15分钟,然后进行洗涤和二抗培养。数据显示,在允许内化的条件下,对于检测的人源化抗cd40igg1-lala单克隆抗体,来自表面定位的抗体的信号会降低至不同程度。对于所有cp-870,893抗体同工型均观察到强烈的信号减弱。
图10:人源化抗cd40抗体的基因报告基因诱导与树突细胞成熟活性的相关性
在hek-blue基因报告基因和树突细胞成熟测定中检测了88种人源化抗cd40igg1-lala抗体的活性。hek-blue基因报告基因活性通过与诱导的seap分泌相关的od@655定量,树突细胞成熟通过il12p40释放(elisa)定量。
图11:激动性抗cd40抗体刺激树突状细胞上的共刺激受体
用激动性cd40抗体、同种型对照抗体或cd40l刺激体外分化的未成熟的idc48小时。通过流式细胞术测定共刺激受体分子的表达。将平均荧光强度归一化至同种型对照处理、或将cd40l归一化至未处理样品。表达的诱导表示为相对于对照处理的诱导倍数(foi)。
图12:抗cd40处理的树突细胞释放细胞因子
用激动性cd40抗体、同种型对照抗体或cd40l刺激体外分化的未成熟的idc48小时。通过elisa(il-12p40)或通过使用bd细胞计数珠阵列的流式细胞术测定细胞因子的释放。
图13:激动性抗cd40抗体对树突细胞上的共刺激受体的剂量依赖性刺激
用激动性cd40抗体或同种型对照抗体以10000至5ng/ml范围内的浓度刺激体外分化的未成熟的idc48小时。通过流式细胞术测定共刺激受体分子的表达。将平均荧光强度归一化至同种型对照处理。表达的诱导表示为相对于对照处理的诱导倍数(foi)。表中显示计算的ec50值。
图14:抗cd40处理的树突细胞的剂量依赖性细胞因子释放
用激动性cd40抗体和同种型对照抗体刺激体外分化的未成熟的idc48小时。使用bd细胞计数珠阵列通过流式细胞术测定细胞因子释放。
图15:激动性抗cd40抗体对b细胞上的共刺激受体的剂量依赖性刺激
用激动性cd40抗体或同种型对照抗体以500至0.2ng/ml范围内的浓度刺激b细胞48小时。通过流式细胞术测定共刺激受体分子的表达。将平均荧光强度归一化至同种型对照处理。表达的诱导表示为相对于对照处理的诱导倍数(foi)。表中显示了计算的ec50值。
图16:结合至细胞表达的cd40的抗cd40抗体与cd40l的竞争
将hek-blue-cd40ltm细胞以ec90浓度与抗cd40-igg1-lala抗体预培养,然后以10000至9.8ng/ml范围内的浓度添加cd40l。使用不同的二抗荧光团偶联抗体检测结合至细胞表面上表达的cd40的抗cd40抗体和cd40l。
图17:激动性cd40抗体与cd40l联合诱导fasr(cd95)死亡受体表达
单独用cd40l或与激动性抗cd40抗体或同种型对照抗体联合过夜刺激ramosb淋巴瘤细胞。通过流式细胞术定量cd95表达。表达的上调表示为相对于对照处理的诱导倍数(foi)。
图18:人源化、激动性抗cd40igg1-lala抗体的亲和力
生化亲和力由biacoret200spr仪器上的表面等离子体共振测定。动力学数据采用langmuir1:1结合模型测定。
图19:cd40结合的抗体竞争
将带有his标签cd40蛋白和不同的抗cd40抗体的预培养混合物与涂覆有不同的抗cd40抗体的板结合。两种抗cd40抗体的连续结合由抗hispod标记的抗体检测。
图20:igg1-lala抗cd40抗体的抗体介导的效应子功能
使用jurkat效应荧光素酶基因报告基因细胞系和hek-blue-cd40ltm作为靶细胞,分析抗cd40抗体介导的效应子功能。将igg1-lala或igg1抗cd40抗体以10000至0.002ng/ml范围内的剂量与靶细胞和效应子细胞培养6小时。所测量的荧光素酶活性的诱导倍数表明抗cd40抗体介导的效应子细胞活化。
图21:激动性抗cd40抗体治疗在人源化小鼠模型中的安全性
在第0天向nod/scid/γ(c)(null)fcrg-/-小鼠注射3μg/gmab-16-0451或cp-870,893抗cd40抗体。在注射之前和之后的不同时间点测量温度。用cp-870893处理的显示出明显的体温下降的3只小鼠在3天后不得不处死。
图22:序列(单字母代码表示的氨基酸)
可变区(vr)的完整序列:
重链:vh完整:seqidno:1-14
轻链:vl完整:seqidno:15-28
互补决定区(cdr):
重链:cdr-h1:seqidno:29-42
cdr-h2:seqidno:43-56
cdr-h3:seqidno:57-70
轻链:cdr-l1:seqidno:71-84
cdr-l2:seqidno:85-98
cdr-l3:seqidno:99-112
序列表
<110>mab发现股份有限公司(mabdiscoverygmbh)
<120>激动性cd40抗体
<130>b198-0013ep1
<160>112
<170>patentin版本3.5
<210>1
<211>122
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>1
gluvalglnleuglugluserglyglyaspleuvalglnproglyala
151015
serleuargleusercysalaalaserglypheserpheserpheser
202530
tyrtrpilecystrpvalargglnalaproglylysglyleugluleu
354045
valsercysiletyrthrthrserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserileaspasnserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargserserglyvalsertyrprosertyrphehisleutrp
100105110
glyglnglythrleuvalthrvalserser
115120
<210>2
<211>125
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>2
gluvalglnleuglugluserglyglyglyleuvalglnproglyala
151015
serleuargleusercysalaalaserglypheserpheserglytyr
202530
trpmetcystrpvalargglnalaproglylysglyleuglutrpval
354045
glycysiletyrthrasnserglyvalthrtyrtyralaasntrpala
505560
lysglyargphethrileserlysaspthrserlysthrthrleutyr
65707580
leuglnmetasnserleuargalagluaspthralathrtyrphecys
859095
alaargglyglyalailetyrasnasptyrasptyralaphetyrtyr
100105110
serleutrpglyglnglythrleuvalthrvalserser
115120125
<210>3
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>3
gluvalglnleuglugluserglyglyaspleuvalglnproglyala
151015
serleuargleusercysalaalaserglypheasppheasnserasn
202530
alametsertrpvalargglnalaproglylysglyleuglutrpval
354045
alaseriletyralaglyglyserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyilethrargleuproleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>4
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>4
gluvalglnleuglugluserglyglyaspleuvalglnproglyala
151015
serleuargleusercysalavalserglypheasppheserserasn
202530
alametsertrpvalargglnalaproglylysglyleuglutrpval
354045
serseriletyralaglyserserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspalaserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyvalthrargleuproleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>5
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>5
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheasppheserserasn
202530
thrmetcystrpvalargglnalaproglylysglyleuglutrpval
354045
alacysiletyralaglyserserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspileserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyleuserargpheserleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>6
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>6
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheasppheserthrasn
202530
alavalsertrpvalargglnalaproglylysglyleuglutrpval
354045
glyserileseralaglyserserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglytyrthrtyrleuthrleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>7
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>7
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheserpheserserasn
202530
alametsertrpvalargglnalaproglylysglyproglutrpval
354045
valthriletyralaglyserserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyalathrtyrleuthrleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>8
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>8
gluvalglnleuvalgluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheasppheserserasn
202530
alametsertrpvalargglnalaproglylysglyleuglutrpval
354045
glyileiletyralaglyserserglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyalathrtyrilethrleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>9
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>9
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheasppheserserasn
202530
alametsertrpvalargglnalaproglylysglyleuglutrpval
354045
glythriletyralaglyserasnglyasnthrasptyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyalasertyrphethrleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>10
<211>117
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>10
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglypheasppheserthrasn
202530
alametcystrpvalargglnalaproglylysglyleuglutrpval
354045
alacysilealaalaglyserserileilethrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralavaltyrphe
859095
cysalaargglyleuserargphealaleutrpglyglnglythrleu
100105110
valthrvalserser
115
<210>11
<211>123
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>11
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglyileasppheserargtyr
202530
tyrtyrmetcystrpvalargglnalaproglylysglyproglutrp
354045
valalacystyrserasnglyaspglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyalaasptyrseralaglyalaalaalapheasnleu
100105110
trpglyglnglythrleuvalthrvalserser
115120
<210>12
<211>123
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>12
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglyileasppheserargtyr
202530
tyrtyrilecystrpvalargglnalaproglylysglyproglutrp
354045
valalacysphealaasnglyaspglyserthrtyrtyralasertrp
505560
alalysglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyalaasptyrserglyglyalaalaalapheasnleu
100105110
trpglyglnglythrleuvalthrvalserser
115120
<210>13
<211>126
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>13
gluvalglnleuglugluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglyileasppheserargtyr
202530
phetyrmetcystrpvalargglnalaproglylysglyleuglutrp
354045
valalacysileglyproglyvalserglyaspthrtyrtyralaser
505560
trpalalysglyargphethrileserglyaspthrserlysthrthr
65707580
leutyrleuglnmetasnserleuargalagluaspthralathrtyr
859095
phecysalaargglyvalasptyrthrtyrglyaspalaglyalaala
100105110
pheasnleutrpglyglnglythrleuvalthrvalserser
115120125
<210>14
<211>125
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>14
gluvalglnleuglugluserglyglyglyleuvalglnproglyala
151015
serleuargleusercysalaalaserglyileasppheserargtyr
202530
phetyrvalcystrpvalargglnalaproglylysglyleuglutrp
354045
valglycysphealaasnhisaspaspseriletyrtyralaglytrp
505560
metasnglyargphethrileserlysaspthrserlysthrthrleu
65707580
tyrleuglnmetasnserleuargalagluaspthralathrtyrphe
859095
cysalaargglyvalasptyrthrvalglytyrglyglyalaalaphe
100105110
asnleutrpglyglnglythrleuvalthrvalserser
115120125
<210>15
<211>112
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>15
aspileglnmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserilesersertyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrseralaserlysleuproserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrhiscysglnthrtyrtyrtyrserserser
859095
sersertyrasptyrglypheglyglnglythrlysvalvalilelys
100105110
<210>16
<211>111
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>16
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserilesersertyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrlysalaserthrleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnsertyrtyrglyserserser
859095
ilesertyrasnalapheglyglnglythrlysvalvalilelys
100105110
<210>17
<211>110
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>17
aspileglnmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserilesersertyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyraspalaserlysleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglythrtyrtyrglyserthr
859095
thrileseralapheglyglnglythrlysvalvalilelys
100105110
<210>18
<211>110
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>18
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserilesersertyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyraspalaserthrleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglythrvaltyrglyserser
859095
thrileseralapheglyglnglythrlysvalvalilelys
100105110
<210>19
<211>111
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>19
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserileserasntyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyraspalaserlysleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglyglyasptyrtyrglyser
859095
sertyrvalvalalapheglyglyglythrlysvalvalilelys
100105110
<210>20
<211>111
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>20
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserhisserileserserthr
202530
tyrleusertrptyrglnglnlysproglyglnalaprolysleuleu
354045
iletyrargalaserthrleualaserglyvalproserargpheser
505560
glyserglyserglythraspphethrleuthrileserserleugln
65707580
progluaspphealathrtyrtyrcysglntyrthrasptyrglyser
859095
sertyrvalserthrpheglyglnglythrlysvalvalilelys
100105110
<210>21
<211>112
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>21
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserileserasntyr
202530
leusertrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrargalaserthrleuproserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglytyrtyrtyrserglythr
859095
thrtyraspserthralapheglyglnglythrlysvalvalilelys
100105110
<210>22
<211>114
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>22
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserileglysertyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrargalaserthrleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglytyrtyrtyrserthrthr
859095
thrthrthrtyraspserseralapheglyglnglythrlysvalval
100105110
ilelys
<210>23
<211>112
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>23
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalasergluservalvalserasn
202530
asnargleualatrptyrglnglnlysproglyglnalaprolysleu
354045
leuiletyrleualaserthrleuproserglyvalproserargphe
505560
serglyserglyserglythraspphethrleuthrileserserleu
65707580
glnprogluaspphealathrtyrtyrcysalaglytyrlysserser
859095
serthraspglythralapheglyglnglythrlysvalvalilelys
100105110
<210>24
<211>111
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>24
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserilesersertyr
202530
leusertrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrleuthrserthrleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglytyrtyrserserserser
859095
tyrvalserasnglypheglyglnglythrlysvalvalilelys
100105110
<210>25
<211>110
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>25
aspileglnmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalasergluserileglyasnala
202530
leuvaltrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrargalaserileleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnasptyrtyrglyserserthr
859095
glutyrasnthrpheglyglnglythrlysvalvalilelys
100105110
<210>26
<211>110
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>26
aspileglnmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserileserserarg
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrargalaserthrleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnasptyrtyrglyserserthr
859095
glutyrasnalapheglyglnglythrlysvalvalilelys
100105110
<210>27
<211>114
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>27
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserilesersertyr
202530
leualatrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrargalaserileleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnthrtyrtyrserserargthr
859095
tyrsertyrglyserproasnalapheglyglnglythrlysvalval
100105110
ilelys
<210>28
<211>109
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>28
aspilevalmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysglnalaserglnserileglysertyr
202530
leusertrptyrglnglnlysproglyglnalaprolysleuleuile
354045
tyrargalathrthrleualaserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnsertyrtyrargaspserser
859095
serseralapheglyglnglythrlysvalvalilelys
100105
<210>29
<211>6
<212>prt
<213>人工序列
<220>
<223>cdr
<400>29
phesertyrtrpilecys
15
<210>30
<211>5
<212>prt
<213>人工序列
<220>
<223>cdr
<400>30
glytyrtrpmetcys
15
<210>31
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>31
serasnalametser
15
<210>32
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>32
serasnalametser
15
<210>33
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>33
serasnthrmetcys
15
<210>34
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>34
thrasnalavalser
15
<210>35
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>35
serasnalametser
15
<210>36
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>36
serasnalametser
15
<210>37
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>37
serasnalametser
15
<210>38
<211>5
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>38
thrasnalametcys
15
<210>39
<211>6
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>39
argtyrtyrtyrmetcys
15
<210>40
<211>6
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>40
argtyrtyrtyrilecys
15
<210>41
<211>6
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>41
argtyrphetyrmetcys
15
<210>42
<211>6
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>42
argtyrphetyrvalcys
15
<210>43
<211>17
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>43
cysiletyrthrthrserglyserthrtyrtyralasertrpalalys
151015
gly
<210>44
<211>17
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>44
cysiletyrthrasnserglyvalthrtyrtyralaasntrpalalys
151015
gly
<210>45
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>45
seriletyralaglyglyserglyserthrtyrtyralasertrpala
151015
lysgly
<210>46
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>46
seriletyralaglyserserglyserthrtyrtyralasertrpala
151015
lysgly
<210>47
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>47
cysiletyralaglyserserglyserthrtyrtyralasertrpala
151015
lysgly
<210>48
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>48
serileseralaglyserserglyserthrtyrtyralasertrpala
151015
lysgly
<210>49
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>49
thriletyralaglyserserglyserthrtyrtyralasertrpala
151015
lysgly
<210>50
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>50
ileiletyralaglyserserglyserthrtyrtyralasertrpala
151015
lysgly
<210>51
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>51
thriletyralaglyserasnglyasnthrasptyralasertrpala
151015
lysgly
<210>52
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>52
cysilealaalaglyserserileilethrtyrtyralasertrpala
151015
lysgly
<210>53
<211>17
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>53
cystyrserasnglyaspglyserthrtyrtyralasertrpalalys
151015
gly
<210>54
<211>17
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>54
cysphealaasnglyaspglyserthrtyrtyralasertrpalalys
151015
gly
<210>55
<211>18
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>55
cysileglyproglyvalserglyaspthrtyrtyralasertrpala
151015
lysgly
<210>56
<211>17
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>56
cysphealaasnhisaspaspseriletyrtyralaglytrpmetasn
151015
gly
<210>57
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>57
serserglyvalsertyrprosertyrphehisleu
1510
<210>58
<211>16
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>58
glyglyalailetyrasnasptyrasptyralaphetyrtyrserleu
151015
<210>59
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>59
glyilethrargleuproleu
15
<210>60
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>60
glyvalthrargleuproleu
15
<210>61
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>61
glyleuserargpheserleu
15
<210>62
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>62
glytyrthrtyrleuthrleu
15
<210>63
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>63
glyalathrtyrleuthrleu
15
<210>64
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>64
glyalathrtyrilethrleu
15
<210>65
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>65
glyalasertyrphethrleu
15
<210>66
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>66
glyleuserargphealaleu
15
<210>67
<211>13
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>67
glyalaasptyrseralaglyalaalaalapheasnleu
1510
<210>68
<211>13
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>68
glyalaasptyrserglyglyalaalaalapheasnleu
1510
<210>69
<211>15
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>69
glyvalasptyrthrtyrglyaspalaglyalaalapheasnleu
151015
<210>70
<211>15
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>70
glyvalasptyrthrvalglytyrglyglyalaalapheasnleu
151015
<210>71
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>71
glnalaserglnserilesersertyrleuala
1510
<210>72
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>72
glnalaserglnserilesersertyrleuala
1510
<210>73
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>73
glnalaserglnserilesersertyrleuala
1510
<210>74
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>74
glnalaserglnserilesersertyrleuala
1510
<210>75
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>75
glnalaserglnserileserasntyrleuala
1510
<210>76
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>76
glnalaserhisserileserserthrtyrleuser
1510
<210>77
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>77
glnalaserglnserileserasntyrleuser
1510
<210>78
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>78
glnalaserglnserileglysertyrleuala
1510
<210>79
<211>13
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>79
glnalasergluservalvalserasnasnargleuala
1510
<210>80
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>80
glnalaserglnserilesersertyrleuser
1510
<210>81
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>81
glnalasergluserileglyasnalaleuval
1510
<210>82
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>82
glnalaserglnserileserserargleuala
1510
<210>83
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>83
glnalaserglnserilesersertyrleuala
1510
<210>84
<211>11
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>84
glnalaserglnserileglysertyrleuser
1510
<210>85
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>85
seralaserlysleuproser
15
<210>86
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>86
lysalaserthrleualaser
15
<210>87
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>87
aspalaserlysleualaser
15
<210>88
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>88
aspalaserthrleualaser
15
<210>89
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>89
aspalaserlysleualaser
15
<210>90
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>90
argalaserthrleualaser
15
<210>91
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>91
argalaserthrleuproser
15
<210>92
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>92
argalaserthrleualaser
15
<210>93
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>93
leualaserthrleuproser
15
<210>94
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>94
leuthrserthrleualaser
15
<210>95
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>95
argalaserileleualaser
15
<210>96
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>96
argalaserthrleualaser
15
<210>97
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>97
argalaserileleualaser
15
<210>98
<211>7
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>98
argalathrthrleualaser
15
<210>99
<211>14
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>99
glnthrtyrtyrtyrsersersersersertyrasptyrgly
1510
<210>100
<211>13
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>100
glnsertyrtyrglyserserserilesertyrasnala
1510
<210>101
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>101
glnglythrtyrtyrglyserthrthrileserala
1510
<210>102
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>102
glnglythrvaltyrglyserserthrileserala
1510
<210>103
<211>13
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>103
glnglyglyasptyrtyrglysersertyrvalvalala
1510
<210>104
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>104
glntyrthrasptyrglysersertyrvalserthr
1510
<210>105
<211>14
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>105
glnglytyrtyrtyrserglythrthrtyraspserthrala
1510
<210>106
<211>16
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>106
glnglytyrtyrtyrserthrthrthrthrthrtyraspserserala
151015
<210>107
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>107
alaglytyrlysserserserthraspglythrala
1510
<210>108
<211>13
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>108
glnglytyrtyrsersersersertyrvalserasngly
1510
<210>109
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>109
glnasptyrtyrglyserserthrglutyrasnthr
1510
<210>110
<211>12
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>110
glnasptyrtyrglyserserthrglutyrasnala
1510
<210>111
<211>16
<212>prt
<213>人工序列
<220>
<223>人源化抗体序列
<400>111
glnthrtyrtyrserserargthrtyrsertyrglyserproasnala
151015
<210>112
<211>11
<212>prt
<213>人工序列
<220>
<223>cdr
<400>112
glnsertyrtyrargaspserserserserala
1510
- 还没有人留言评论。精彩留言会获得点赞!