本发明属于基因工程和酶工程技术领域,具体内容涉及耐热性提高的羰基还原酶突变体。
背景技术:
作为氧化还原酶家族的一员,羰基还原酶(carbonylreductase,ec1.1.1.184)能够催化羰基的不对称还原生成相应的手性醇。手性醇是一类重要的有机合成砌块,在医药、农业、食品和化工等行业都有很高的应用价值。利用羰基还原酶生物催化生产手性醇越来越广泛地应用于工业生产中。
为了适应极端工业环境下的应用,如极端温度、强酸碱环境等,对羰基还原酶的酶学性能提出了更高的要求,理想条件是酶能够在这些环境下保持良好的活力和稳定性。其中酶的热稳定性是工业应用的重要指标。高温条件下,酶的催化效率高,反应周期短,可以有效地提高时空产率,节约成本,且便于操作,也有利于避免反应过程中被其他微生物污染。
分子改造技术为研究酶的底物特异性和催化活性位点、改善酶的热稳定性以及提高酶作用温度等提供了有效手段。通过分子改造,可以快速定向地获得符合工业需求的酶。定向进化和理性/半理性设计是酶分子改造的主要技术手段,也是当前酶工程领域研究的热点。目前,已有许多学者运用这项技术成功的改造了各种各样的酶,取得了令人瞩目的进展(c.a.denard,h.renandh.zhao,curropinchembiol,2015,25,55-64.)。其中,定点突变技术已成为酶分子改造中常用的手段,大大加速了蛋白质的进化过程(h.yang,j.li,h.d.shin,g.du,l.liuandj.chen,applmicrobiolbiotechnol,2014,98,23-29.)。
技术实现要素:
本发明以理性设计的方案,利用consensus方法以及fireport在线网站预测来源于金黄杆菌ca49(chryseobacteriumsp.ca49)的羰基还原酶chkred03(氨基酸序列为seqidno.2)热稳定性相关的氨基酸位点,并进行分子改造,利用定点突变技术进行单点突变和组合突变,从而获得热稳定性提高的突变体。
所述羰基还原酶突变体是以seqidno.2为出发序列,将第183位的苏氨酸突变为缬氨酸,或者将第183位的苏氨酸突变为异亮氨酸,或者将第188位的天冬酰胺突变为亮氨酸,或者将第211位的丙氨酸突变为脯氨酸,或者将第224位的丝氨酸突变为辅氨酸,或者以上四个位点的任意组合得到的突变体。
根据本领域公共知识,构建的能表达上述突变体的载体、基因工程菌等也属于本发明的保护范围。
为了达到以上目的,本发明根据理性设计预测得到的氨基酸位点,以定点突变技术对母本羰基还原酶chkred03基因进行突变,获得5个热稳定性提高的单点突变体。以定点突变技术将单点突变体进行多位点整合,获得3个耐热性进一步提高的组合突变体。
本发明的具体技术方案为:
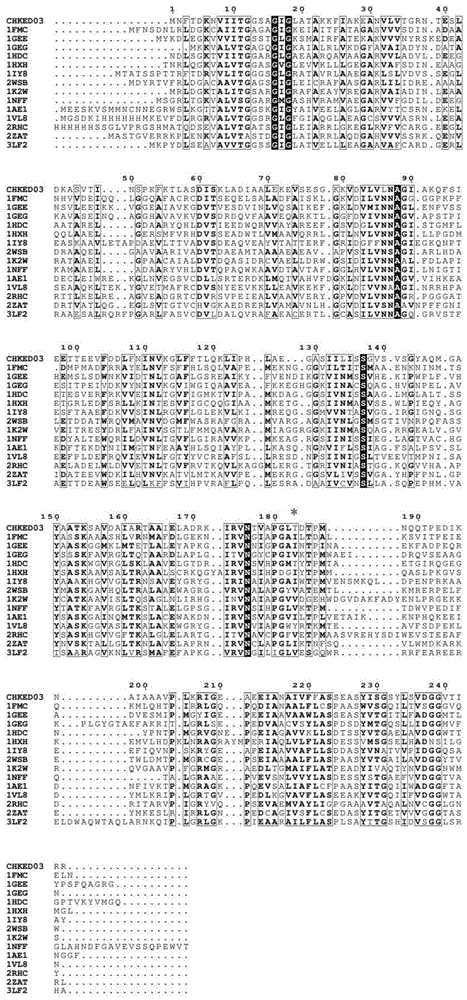
(1)羰基还原酶chkred03热稳定性相关位点的预测:我们已经公开了从金黄杆菌ca49(chryseobacteriumsp.ca49,2012年11月27日在中国典型培养物保藏中心保藏,保藏编号为no:cctccm2012484)中克隆的编码244个氨基酸残基的羰基还原酶chkred03的基因序列(ncbi登录号:kc342003,氨基酸序列见seqidno.2)。consensus方法预测热稳定性位点:将羰基还原酶chkred03的氨基酸序列与pdb数据库中已知晶体结构的14种同源性的羰基还原酶的氨基酸序列进行一级序列比对,序列相似性在45%-30%,序列比对软件为clustalx和在线网站espript3.0(http://espript.ibcp.fr/espript/cgi-bin/espript.cgi),见说明书附图1。比对结果发现,羰基还原酶chkred03的氨基酸序列中第183位的苏氨酸在其同源耐热性羰基还原酶氨基酸序列相应位点占多数的氨基酸为缬氨酸和异亮氨酸,拟决定将羰基还原酶chkred03的氨基酸序列中第183位的苏氨酸替换成缬氨酸或异亮氨酸,构建突变体t183v和t183i。
fireport预测热稳定性位点:将羰基还原酶chkred03的氨基酸序列提交至在线网站i-tasser(https://zhanglab.ccmb.med.umich.edu/i-tasser/)进行同源建模,获得羰基还原酶chkred03的三维模型,再将三维模型提交至fireport(https://loschmidt.chemi.muni.cz/fireprot/)进行热稳定性相关氨基酸位点预测。从预测的突变子中经过各参数的比较分析,选择最有潜力的3个突变子(n188l、a211p和s224p,见说明书附图2),构建热稳定性突变体。
(2)突变体构建和热稳定性验证:利用定点突变技术,以pet-28a-chkred03为模板,构建单点突变质粒。本发明也可以根据大肠杆菌密码子偏好性优化后的基因序列为模板(基因序列见seqidno.1,氨基酸序列不变),实施例中以优化后的序列seqidno.1为模板来详细说明本发明的具体方案。
将突变体质粒化学法转入大肠杆菌dh5α,送于上海生工生物工程股份有限公司测序,测序正确后,提取质粒转入表达菌株大肠杆菌bl2(de3),挑选单克隆进行诱导表达蛋白,菌体冷冻离心,加入磷酸钾缓冲液(0.1m、ph7.0)超声波破碎细胞,冷冻离心,取适量上清粗酶液用于反应,以n-叔丁氧羰基-3-哌啶酮为底物,反应适当时间测定酶活。同时,另取等量的上清液于不同温度热处理一定时间,以相同的方法测定残余活性。再以未经过高温处理的粗酶液的酶活为参比,得到残余酶活百分比。详细方案见实例1和2。
经上述consensus方法和fireport预测和筛选,获得了5个热稳定性提高的突变体,分别为t183v、t183i、n188l、a211p、s224p,其特征如下:
t183v:第183位的苏氨酸突变为缬氨酸(dna序列由acc变为gtt);
t183i:第183位的苏氨酸突变为异亮氨酸(dna序列由acc变为att);
n188l:第188位的天冬酰胺突变为亮氨酸(dna序列由aat变为ctg);
a211p:第211位的丙氨酸突变为脯氨酸(dna序列由gcg变为ccg);
s224p:第224位的丝氨酸突变为脯氨酸(dna序列由agc变为ccg)。
以上5个突变体(t183v、t183i、n188l、a211p、s224p)热稳定性相比母本均有不同程度的提高。其中,t183v和n188l热稳定性提高最大,在40℃热处理1.5h,t183v和n188l仍保留90%以上的相对酶活性,而野生型相同温度下热处理1.5h,仅残余8%的相对酶活性。
(3)4个位点的整合:
通过上述(1)突变筛选,获得5有益突变位点,由于t183i的热稳定性比t183v稍差。因此,利用t183v、n188l、a211p、s224p进行整合。以热稳定性最好的n188l为模板,利用定点突变技术,构建三个或以上位点组合突变体。优选构建以下组合突变体:m814、m834、m8314,其特征如下:
m814:第188位的天冬酰胺突变为亮氨酸(dna序列由aat变为ctg)、第211位的丙氨酸突变为脯氨酸(dna序列由gcg变为ccg)、第224位的丝氨酸突变为脯氨酸(dna序列由agc变为ccg)。
m834:第188位的天冬酰胺突变为亮氨酸(dna序列由aat变为ctg)、第183位的苏氨酸突变为缬氨酸(dna序列由acc变为gtt)、第224位的丝氨酸突变为脯氨酸(dna序列由agc变为ccg)。
m8314:第188位的天冬酰胺突变为亮氨酸(dna序列由aat变为ctg)、第183位的苏氨酸突变为缬氨酸(dna序列由acc变为gtt)、第211位的丙氨酸突变为脯氨酸(dna序列由gcg变为ccg)、第224位的丝氨酸突变为脯氨酸(dna序列由agc变为ccg)。
这3种组合突变体的热稳定性与单点突变体相比又有进一步提高。45℃热处理3h,m814、m834和m8314的酶活力基本未发生变化,而野生型在40℃热处理1.5h,仅剩余8%的相对酶活性。m8314稳定性提升幅度最大,55℃热处理2h,仍残余高达80%的相对酶活性。
本发明有益效果:上述所有突变体与母本相比,热稳定性均有大幅度提高,且酶的活性基本未受到影响,这些突变体能够在较高温度下进行工业生产,利于生产工艺的灵活性,具有良好的工业应用前景。
附图说明
图1chkred03与同源性的羰基还原酶氨基酸序列比对,*号为chkred03的第183位点所在位置。
图2fireport热稳定性突变子预测结果,方框部分为本发明所选择的突变子。
图340℃条件下,单点突变体与野生型热稳定性比较,其中“○”表示chkred03的热稳定性曲线,“■”表示t183v的热稳定性曲线,“▲”表示n188l的热稳定性曲线,表示a211p的热稳定性曲线,表示s224p的热稳定性曲线。
图455℃条件下,多位点突变体稳定性比较,“●”表示m8314的热稳定性曲线,“■”表示m834的热稳定性曲线。
具体实施方法
以下结合实施实例对本发明做进一步说明,需要指出的是,本实施例仅用于解释本发明,而非对本发明范围的限制。
实施例15个单点突变体的构建
理性设计方法对潜在耐热性位点预测的结果见说明书附图1和2。单点突变体t183v、t183i、n188l、a211p和s224p均以羰基还原酶chkred03密码子优化后的基因seqidno.1为模板进行构建,所用引物:
t183v-f:5′–ggctccgggcctggttgataccccgatgaatc–3′
t183v-r:5′–gattcatcggggtatcaaccaggcccggagcc–3′
t183i-f:5′–ggctccgggcctgattgataccccgatgaatc–3′
t183i-r:5′–gattcatcggggtatcaatcaggcccggagcc–3′
n188l-f:5′–accgataccccgatgctgcagcagaccccgg–3′
n188l-r:5′–ccggggtctgctgcagcatcggggtatcggt–3′
a211p-f:5′–gaaacggataggcgaaccggaagagattgccaatg–3′
a211p-r:5′–cattggcaatctcttccggttcgcctatccgtttc–3′
s224p-f:5′–cgtgttttttgcgagcccggaggcgagctatatat–3′
s224p-r:5′–atatatagctcgcctccgggctcgcaaaaaacacg–3′
pcr条件为:5×hfbuffer10μl,mgcl2(1mm)1μl,引物(50ng/μl)各1.5μl,dntp(2.5mm)4μl,phu(1u)1μl,质粒50ng,超纯水补足50μl,条件:98℃预变性3min,98℃变性10s,55℃退火45s,72℃延伸2min,共25个循环,再72℃延伸10min。pcr产物用1μldpni,37℃处理1h。取dpni消化的pcr产物10μl,化学法转入大肠杆菌dh5α。送于上海生工生物工程股份有限公司测序,测序正确后,提取质粒转入表达菌株大肠杆菌bl21(de3)。
实施例2粗酶液制备及酶活力的测定
2.1粗酶液的制备
将实施例1中各个突变体质粒,化学法转入大肠杆菌表达菌株bl21-de3,涂布含有卡那霉素(50μg/ml)的lb平板,37℃培养14h,挑取单克隆于3ml含有卡那霉素(50μg/ml)的lb液体培养基中,37℃,180rpm,过夜培养。1%的接种量接种于200ml含有卡那霉素(50μg/ml)的tb培养基中,37℃培养3h后,加入终浓度0.5mmiptg,30℃诱导18h后,5000rpm、4℃离心10min,弃上清,菌体用生理盐水清洗两次,加入15ml0.1m的磷酸钾缓冲液(ph7.0),混匀,超声波破碎细胞(工作条件:工作时间3s,间歇时间3s,工作次数99,功率200w),12000rpm,4℃离心,20min,取上清粗酶液用于反应。
2.2粗酶活力的测定
粗酶活力测定反应条件见表1,以葡萄糖脱氢酶gdh进行辅酶循环。底物溶解于甲醇中配制成1m的贮存液。反应体系于30℃温浴5min,最后加入终浓度为5mg/ml的粗酶液,30℃、150rpm反应15min。反应结束后,加入1ml的乙酸乙酯萃取,无水硫酸钠干燥,而后旋蒸去除溶剂,再以异丙醇复溶(hplc级),用于检测。
检测条件:色谱柱(chiralpakic,4.6mm×250mm,daicel,japan),柱温30℃,流动相(正己烷/异丙醇=90/10,v/v),流速0.8ml/min,检测波长210nm,保留时间(底物30.4min;(s)-产物14.9min;(r)-产物,16.0min)。
表1粗酶反应体系
2.3热稳定性的测定
将总蛋白浓度50mg/ml的粗酶液100μl置于容量250μl的pcr管,在40℃条件下热处理1.5h,间隔一定时间取样,迅速放置冰上冷却,取热处理后的粗酶液测定残余活性,测定方法参照实施例2.2粗酶活力测定。再以未经过高温处理的粗酶液为参比,得到残余相对酶活性。
本轮突变获得5个突变体t183v、t183i、n188l、a211p和s224p。野生型与突变体对底物的转化率以及相对酶活性见表2和说明书附图3。
40℃热处理1.5h,野生型对底物的转化率为6%,而突变体t183v、n188l、a211p和s224p对底物仍保留较高的转化率。
表2野生型和突变体的转化率及相对酶活力
a所有数值均由粗酶液测定
实施例3构建多位点组合突变体
3.1突变体m814的构建
定点突变方法将突变体n188l的第211位的丙氨酸突变为脯氨酸、第224位的丝氨酸突变为脯氨酸,构建突变体m814,所用的引物如下:
a211p-f:5′–gaaacggataggcgaaccggaagagattgccaatg–3′
a211p-r:5′–cattggcaatctcttccggttcgcctatccgtttc–3′
s224p-f:5′–cgtgttttttgcgagcccggaggcgagctatatat–3′
s224p-r:5′–atatatagctcgcctccgggctcgcaaaaaacacg–3′
pcr条件以及操作同实施例1,得到新突变体m814。
3.2突变体m834的构建
定点突变方法将突变体n188l的第183位的苏氨酸突变为缬氨酸、第224位的丝氨酸突变为脯氨酸,构建突变体m834,所用的引物如下:
n188l-t183v-f:5′–ggctccgggcctggttgataccccgatgctgc–3′
n188l-t183v-r:5′–gcagcatcggggtatcaaccaggcccggagcc–3′
s224p-f:5′–cgtgttttttgcgagcccggaggcgagctatatat–3′
s224p-r:5′–atatatagctcgcctccgggctcgcaaaaaacacg–3′
pcr条件以及操作同实施例1,得到新突变体m834。
3.3突变体m8314的构建
定点突变方法将突变体n188l的第183位的苏氨酸突变为缬氨酸、第211位的丙氨酸突变为脯氨酸、第224位的丝氨酸突变为脯氨酸,构建突变体m8314,所用的引物如下:
n188l-t183v-f:5′–ggctccgggcctggttgataccccgatgctgc–3′
n188l-t183v-r:5′–gcagcatcggggtatcaaccaggcccggagcc–3′
a211p-f:5′–gaaacggataggcgaaccggaagagattgccaatg–3′
a211p-r:5′–cattggcaatctcttccggttcgcctatccgtttc–3′
s224p-f:5′–cgtgttttttgcgagcccggaggcgagctatatat–3′
s224p-r:5′–atatatagctcgcctccgggctcgcaaaaaacacg–3′
pcr条件以及操作同实施例1,得到新突变体m8314。
3.4组合突变体热稳定性测定
将总蛋白浓度50mg/ml的粗酶液100μl置于容量250μl的pcr管,在55℃水浴处理2h,然后将样品管放置于冰上冷却,测定方法见2.2。以未经过高温处理的酶液为参比,得到酶相对活力。
与热稳定性最高的单点突变体n188l相比,组合突变体m814、m834和m8314热稳定性有不同程度提高。其中,突变体m8314的热稳定性提高幅度最大。组合突变体对底物的转化率以及相对酶活性见表3和说明书附图4。55℃温度下热处理2h后,野生型和突变体n188l几乎没有活性,m8314仍保留81%的相对酶活性。
表3组合突变体和母本的相对酶活力及转化率
a所有数值均由粗酶液测定。
序列表
<110>中国科学院成都生物研究所
<120>耐热性提高的羰基还原酶突变体
<130>基因工程和酶工程技术领域
<160>2
<170>siposequencelisting1.0
<210>1
<211>735
<212>dna
<213>chryseobacteriumsp.
<400>1
atgaacttcaccgacaaaaatgtgattattaccggaggctctgcgggcattggactggcg60
accgcgaaaaagtttattgcgaaagaagcgaatgtactggtgaccggccgtaataccgaa120
agcctggataaagcgagcgtgaccattaacagccccaaattcaaaacgctggcgagcgat180
attagcaagctggctgatatagcggcgctggaaaaagaagtgagcgaaagcggcaaaaag240
gtggatgtgctggtgcttaacgcaggtatcgcgaaacagtttagcattgaagaaaccacc300
gaagaagtatttgatgatctgttcaacattaacgtgaaaggcctgtttttcaccctgcag360
aaactgattccgcatctggcggaaggcgcgagcattattctgattagctcaggcgtgagc420
gtaagcggctatgcgcaaatgggagcgtatgcggcgaccaaaagcgcggtggacgctatt480
gcccgtaccgcagcgattgaactggcagaccgtaagattcgtgtgaataccgtggctccg540
ggcctgaccgataccccgatgaatcagcagaccccggaagatataaagaacgcgatagca600
gcggcggtgccgctgaaacggataggcgaagcggaagagattgccaatgcgatcgtgttt660
tttgcgagcagcgaggcgagctatatatcgggttcgtatctgagcgtggatggcggcgtc720
accattcgtcgttaa735
<210>2
<211>244
<212>prt
<213>chryseobacteriumsp.
<400>2
metasnphethrasplysasnvalileilethrglyglyseralagly
151015
ileglyleualathralalyslyspheilealalysglualaasnval
202530
leuvalthrglyargasnthrgluserleuasplysalaservalthr
354045
ileasnserprolysphelysthrleualaseraspileserlysleu
505560
alaaspilealaalaleuglulysgluvalsergluserglylyslys
65707580
valaspvalleuvalleuasnalaglyilealalysglnpheserile
859095
glugluthrthrglugluvalpheaspaspleupheasnileasnval
100105110
lysglyleuphephethrleuglnlysleuileprohisleualaglu
115120125
glyalaserileileleuileserserglyvalservalserglytyr
130135140
alaglnmetglyalatyralaalathrlysseralavalaspalaile
145150155160
alaargthralaalailegluleualaasparglysileargvalasn
165170175
thrvalalaproglyleuthraspthrprometasnglnglnthrpro
180185190
gluaspilelysasnalailealaalaalavalproleulysargile
195200205
glyglualaglugluilealaasnalailevalphephealaserser
210215220
glualasertyrileserglysertyrleuservalaspglyglyval
225230235240
thrileargarg