一种利用FLD1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株及构建方法与流程
本发明属于分子生物学技术领域,尤其是一种利用fld1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株及构建方法,更具体地,为重组质粒pfld9k-tgz的构建和通过启动子pfld1调控黏玉米谷氨酰胺转氨酶在巴斯德毕赤酵母中的表达。
背景技术:
谷氨酰胺转氨酶(transglutaminase,tgase,ec2.3.2.13)是一种通过催化谷氨酰胺残基和赖氨酸残基之间的酰基转移反应,形成赖氨酸异肽键,从而实现蛋白质分子内、分子间交联的酶。tgase在不改变产品风味和颜色的前提下可以改善食品的凝胶性、溶解性、乳化性、发泡性和持水力。同时这种交联反应可以将必需氨基酸转移到蛋白质,从而提高蛋白质的营养价值。
tgase在食品、生物医学、纺织等方面有着广阔的应用前景,而且tgase是被食品药品监督管理局和其他科学界认可的人类摄入安全的物质。酪蛋白,大豆球蛋白,谷蛋白,肌原纤维蛋白等食物蛋白质都是tgase的良好底物,因此其在食品行业的应用最受关注,这也进一步增加了对廉价、有效和安全tgase的需求。
植物来源tgase主要分布于植物的细胞质、细胞壁、叶绿体和线粒体等亚细胞区室,可能与植物组织中的能量转移、细胞骨架形成有关。植物来源tgase的分离纯化工艺复杂,同时数据库中缺少相应的氨基酸和基因序列信息,这些共同阻碍了植物来源tgase的研究。目前已获得拟南芥、黏玉米、菊芋、稻子、梨及苹果tgase的基因序列,研究者也已陆续将拟南芥和黏玉米tgase进行异源表达。但是构建的基因工程菌株的产酶量较低,限制了植物来源tgase的应用研究。因此,进一步提高表达水平,获得高活性谷氨酰胺转氨酶重组菌株,是目前亟待解决的问题。
通过检索,尚未发现与本发明专利申请相关的专利公开文献。
技术实现要素:
本发明的目的在于克服现有技术的不足之处,提供一种利用fld1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株,该重组菌株易于培养、分泌能力强,利于谷氨酰胺转氨酶的大量表达,本发明利用毕赤酵母为宿主发酵得到的谷氨酰胺转氨酶具有安全可靠的特点,且具有重要的应用前景。
本发明解决其技术问题是采取以下技术方案实现的:
一种利用fld1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株,所述重组毕赤酵母菌株为将来源于黏玉米(zeamays)的谷氨酰胺转氨酶(transglutaminase)基因转入巴斯德毕赤酵母(pichiapastoris)中得到的基因工程菌株,该重组毕赤酵母菌株通过将ppic9k质粒中的醇氧化酶启动子paox1替换成甲醛脱氢酶启动子pfld1,并与谷氨酰胺转氨酶基因连接成环,构成重组表达载体的方式得到的。
而且,所述谷氨酰胺转氨酶的氨基酸序列如seqidno.1所示。
而且,所述pfld1的核苷酸序列如seqidno.2所示。
而且,所述重组毕赤酵母的宿主为p.pastorisgs115。
如上所述的重组毕赤酵母菌株的构建方法,步骤如下:
⑴获得来源于黏玉米的谷氨酰胺转氨酶基因与启动子pfld1序列;
⑵将ppic9k载体与谷氨酰胺转氨酶基因连接,获得含有黏玉米谷氨酰胺转氨酶基因的重组质粒paox9k-tgz;
⑶启动子pfld1进行saci和bamhi双酶切并定向插入同样经saci和bamhi双酶切的重组质粒paox9k-tgz中,获得重组载体pfld9k-tgz;
⑷将步骤⑶得到的重组载体pfld9k-tgz转化到宿主中,获得重组毕赤酵母菌株。
如上所述的重组毕赤酵母菌株生产黏玉米来源谷氨酰胺转氨酶的方法,步骤如下:
将重组毕赤酵母菌株接种于ypd培养基中,25-30℃,210-260r/min活化培养20-27h后,得活化的培养液,再将活化的培养液转接至发酵体系中发酵生产谷氨酰胺转氨酶。
而且,按10%的接种量将活化的培养液转接到发酵培养基中,每24h加入甲胺,使其质量终浓度为0.5%,发酵时间为96h,即得谷氨酰胺转氨酶。
本发明取得的优点和积极效果是:
本发明重组毕赤酵母菌株所用的重组载体是在ppic9k载体基础上进行改造,替换了启动子后转入毕赤酵母,高效生产谷氨酰胺转氨酶。利用本发明提供的重组毕赤酵母发酵得到的谷氨酰胺转氨酶,酶活可达635mu/ml,高于整合paox9k-tgz载体的重组毕赤酵母的酶活,即586mu/ml(已在文章heterologousexpressionandpurificationofzeamaystransglutaminaseinpichiapastoris.foodscienceandbiotechnology.2014,23:1507-1513中公开发表)。重组菌株易于培养、分泌能力强,利于谷氨酰胺转氨酶的大量表达,本发明利用毕赤酵母为宿主发酵得到的谷氨酰胺转氨酶具有安全可靠的特点,且具有重要的应用前景。
附图说明
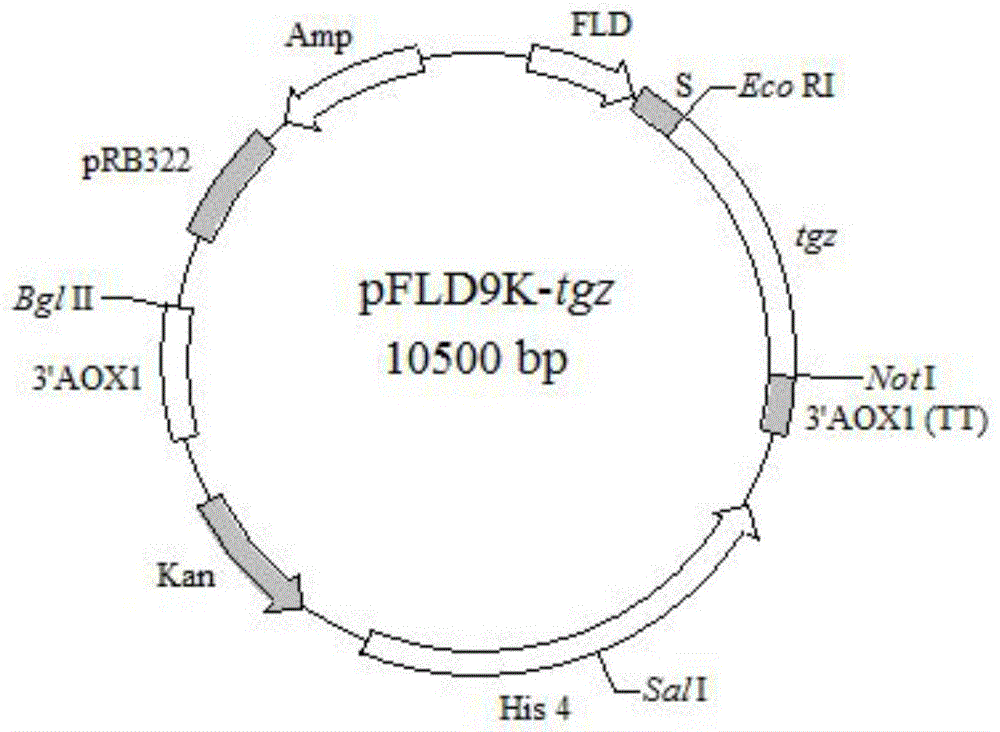
图1为本发明中整合表达质粒pfld9k-tgz图谱;
图2为本发明中重组质粒pfld9k-tgz的双酶切鉴定图;
图3为本发明中阳性重组菌株的pcr鉴定图;
图4为本发明中活性表达黏玉米谷氨酰胺转氨酶重组菌株发酵上清sds-page图。
具体实施方式
下面结合通过具体实施例对本发明作进一步详述,以下实施例只是描述性的,不是限定性的,不能以此限定本发明的保护范围。
本发明中所使用的原料,如无特殊说明,均为常规的市售产品;本发明中所使用的方法,如无特殊说明,均为本领域的常规方法。
本发明中使用到的培养基可以如下:
lb培养基:胰蛋白胨10g/l,酵母提取物5g/l,氯化钠10g/l。
ypd培养基:蛋白胨20g/l,酵母提取物10g/l,葡萄糖20g/l。
固体培养基为液体培养基中添加琼脂,添加量为20g/l。
发酵培养基:蛋白胨20g/l,酵母提取物10g/l,无氨基酸的酵母氮源13.4g/l,山梨醇5.4g/l,生物素0.4mg/l,100ml1mol/l(ph6.0)磷酸盐缓冲液。
本发明中谷氨酰胺转氨酶活力测定可以如下:
采用比色法测定谷氨酰胺转氨酶酶活。1个单位酶活的定义为:37℃,ph8.0的条件下,每分钟催化产生1μmol单羟肟酸所需的酶量(u/ml)。酶活测定条件:50μl酶液与50μl试剂a(100mgn-cbz-gln-gly溶解于2ml0.2mol/lnaoh,加入4ml0.2mol/ltris-hcl缓冲液(ph8.0),2ml0.01mol/l还原型谷胱甘肽和2ml0.1mol/l羟胺,并调节ph至8.0)混合,37℃反应30min后加50μl试剂b(50g/lfecl3,120g/l三氯乙酸及3mol/l盐酸按体积比为1:1:1的比例混合)终止反应,用酶标仪于525nm波长下比色。通过l-谷氨酸-γ-单羟肟酸绘制标准曲线,根据曲线计算酶活。
一种利用fld1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株,所述重组毕赤酵母菌株为将来源于黏玉米(zeamays)的谷氨酰胺转氨酶(transglutaminase)基因转入巴斯德毕赤酵母(pichiapastoris)中得到的基因工程菌株,该重组毕赤酵母菌株通过将ppic9k质粒中的醇氧化酶启动子paox1替换成甲醛脱氢酶启动子pfld1,并与谷氨酰胺转氨酶基因连接成环,构成重组表达载体的方式得到的。
较优地,所述谷氨酰胺转氨酶的氨基酸序列如seqidno.1所示。
较优地,所述pfld1的核苷酸序列如seqidno.2所示。
较优地,所述重组毕赤酵母的宿主为p.pastorisgs115。
如上所述的重组毕赤酵母菌株的构建方法,步骤如下:
⑴获得来源于黏玉米的谷氨酰胺转氨酶基因与启动子pfld1序列;
⑵将ppic9k载体与谷氨酰胺转氨酶基因连接,获得含有黏玉米谷氨酰胺转氨酶基因的重组质粒paox9k-tgz;
⑶启动子pfld1进行saci和bamhi双酶切并定向插入同样经saci和bamhi双酶切的重组质粒paox9k-tgz中,获得重组载体pfld9k-tgz;
⑷将步骤⑶得到的重组载体pfld9k-tgz转化到宿主中,获得重组毕赤酵母菌株。
如上所述的重组毕赤酵母菌株生产黏玉米来源谷氨酰胺转氨酶的方法,步骤如下:
将重组毕赤酵母菌株接种于ypd培养基中,25-30℃,210-260r/min活化培养20-27h后,得活化的培养液,再将活化的培养液转接至发酵体系中发酵生产谷氨酰胺转氨酶。
较优地,按10%的接种量将活化的培养液转接到发酵培养基中,每24h加入甲胺,使其质量终浓度为0.5%,发酵时间为96h,即得谷氨酰胺转氨酶。
更具体地,如上所述的重组毕赤酵母菌株的构建方法,步骤如下:
(1)表达载体pfld9k-tgz的构建
以黏玉米谷氨酰胺转氨酶基因为模板,参照genbank上黏玉米谷胺酰转氨酶基因序列(序列号为aj421525)设计引物tgz-f(atcgaattcatggctcatcgtggacatctaga,下划线为ecori酶切位点)和tgz-r(attagcggccgcatttcaccatatttgtct,下划线为noti酶切位点),进行pcr扩增,扩增条件为94℃预变性2min;94℃变性15s,60℃退火30s,68℃延伸50s,反应30个循环;68℃延伸5min。pcr扩增产物经胶回收后连接到pta2载体,构建pta2-tgz重组载体,转化到e.colidh5α感受态细胞,进行序列测定。将验证正确的重组载体pta2-tgz和表达载体ppic9k分别进行ecori和noti双酶切,使用胶回收试剂盒回收后用ligationhigh进行连接以构建重组表达载体paox9k-tgz。将重组载体转化e.colidh5α感受态细胞,进行菌落pcr和ecori和noti双酶切验证。随后,将验证正确的重组载体paox9k-tgz转化毕赤酵母感受态细胞,构建重组菌株gs115/paox9k-tgz。
根据酵母基因组提取试剂盒说明书提取毕赤酵母gs115基因组,并以其为模板,参照genbank上fld1基因序列(序列号为af066054)设计引物fld1(ctagagctcgcatgcaggaatctctg,下划线为saci酶切位点)和fld2(gcgaggatcctgtgaatatcaagaattg,下划线为bamhi酶切位点),通过pcr扩增启动子pfld1。扩增条件为94℃预变性2min;94℃变性15s,52℃退火30s,68℃延伸30s,反应30个循环;68℃延伸5min。pcr扩增产物经胶回收后连接到pta2载体,构建重组载体pta2-fld,转化到e.colidh5α感受态细胞,进行序列测定。将验证正确的pta2-fld和重组质粒paox9k-tgz分别进行saci和bamhi双酶切,使用胶回收试剂盒回收后用ligationhigh进行连接以构建重组表达载体pfld9k-tgz,如图1所示。将重组载体转化e.colidh5α感受态细胞,进行菌落pcr和saci和bamhi双酶切验证。结果如图2所示。分别得到大小约为11000bp和600bp的载体片段和插入片段,与阳性对照大小一致。结果表明外源基因以正确的方向和阅读框插入,说明成功构建了表达载体pfld9k-tgz。
(2)表达载体pfld9k-tgz转化毕赤酵母
将步骤(1)得到的表达载体pfld9k-tgz进行sali单酶切线性化,用2倍体积无水乙醇进行质粒浓缩,离心后倒去上清,用100μl70%(v/v)乙醇洗涤沉淀3-4次,离心去上清,风干后用20-30μl无菌ddh2o溶解。将0.5-1μg线性化质粒与80μl毕赤酵母gs115感受态细胞混合后进行电击转化,经rdb平板筛选后获得重组菌株gs115/pfld9k-tgz,挑取阳性克隆进行pcr鉴定。结果如图3所示,在约600bp(图3a)和1600bp(图3b)的位置各有一条清晰条带,大小与预计的片段大小相符,表明线性化重组质粒成功整合到p.pastorisgs115基因组染色体上。
(3)p.pastoris转化子gs115/pfld9k-tgz发酵产酶
将步骤(2)中构建的重组菌株gs115/pfld9k-tgz和对照菌株gs115/ppic9k分别接种于ypd培养基中,并在28℃以250rpm的转速震荡培养24h,获得种子液。按10%接种量转接于发酵培养基中,在28℃,每24h加入甲胺至其质量终浓度为0.5%,250rpm培养96h。将诱导表达后的菌液离心收集上清,进行sds-page分析和酶活测定。结果如图4所示,阴性对照菌gs115/ppic9k在74kda处没有特异条带,而阳性重组菌gs115/pfld9k-tgz在74kda处出现特异条带。该结果表明启动子pfld1成功的调控了黏玉米谷氨酰胺转氨酶在p.pastorisgs115中的表达。酶活测定结果表明,上清液的酶活为635mu/ml。
尽管为说明目的公开了本发明的实施例,但是本领域的技术人员可以理解:在不脱离本发明及所附权利要求的精神和范围内,各种替换、变化和修改都是可能的,因此,本发明的范围不局限于实施例所公开的内容。
序列表
<110>天津科技大学
<120>一种利用fld1启动子调控黏玉米谷氨酰胺转氨酶表达的重组毕赤酵母菌株及构建方法
<160>6
<170>siposequencelisting1.0
<210>1
<211>534
<212>prt
<213>谷氨酰胺转氨酶的氨基酸序列(unknown)
<400>1
metalahisalaglyhisleualaglyleuthrglyglyalaproala
151015
leumetalahisglyserproalaalaglyserleuserseralaser
202530
proleuglyserserserthrleuglymetleuglyalaleuleuala
354045
metglythrthrglyvalglyleuleuilethrglyalaglyalaleu
505560
alaserserhisvalvalleualaglyalailevalalathrglyleu
65707580
glymetglymetilealathrhisleuglyglyvalglythrglythr
859095
alaleuglyilealaalaleuleuglyalailealaleumetglyval
100105110
alailehisserglyalavalvalalaleuglyleuhisglymethis
115120125
metglyalaleualaleuilethrglyalaglymetleuthrleugly
130135140
ileglyalavalthrleuglyleuglyleuleuseralaserglyala
145150155160
alaleuserleuproglyleuleuserglyleualaalaleualaleu
165170175
glyhishisalaleualaserglyproglyproglyleualathrala
180185190
valleuglyvalglyglymetalathrmetglymetalaleuilethr
195200205
metthrleuglyalaglyleuleualavalalavalalaalaalagly
210215220
alaalaalaglyalaalaalaalaglyalaalaalahisalaalagly
225230235240
alaglyvalthralaserglyproglyglyleuleuleuproalapro
245250255
glyglyglyglyproglythrhismetglyvalhisileproalathr
260265270
proleuhisileseralaglyproalaleuglyhisileseralaval
275280285
leualaleuglythrileseralaglyproalaleuglyhisileser
290295300
alaglyproalaleuglyhisileseralaglyalaalametglyhis
305310315320
ileseralaglyleualaleuglyhisileseralaglyproalaleu
325330335
glyhisileseralaglyproalaleuglyhisileseralavalleu
340345350
alaleuglyhisileseralaglyproalaleuglyhisileserala
355360365
glyproserleuglyhisileseralaglyproalaleuglyhisile
370375380
seralaglyproalametglyhisileseralaglyproalametgly
385390395400
hisileseralavalleualaleuglyhisthrthrmetleumetmet
405410415
leualaalaleumethismetglyvalthrleualaileglyleugly
420425430
alathralaleuvalglycysprothrileprometleuhisleuala
435440445
serglyglyalaalavalglyleualathrproglyglyalasermet
450455460
glyglyleuvalvalleualaileleuleuglyleuproseralaala
465470475480
valalaleuglymetglyalaleuhisleuleuleuhishishisala
485490495
glyhishisileproproalahismetthrleuproglyglyproala
500505510
alaleuilethralavalalaglymetalavalcyshisalahisleu
515520525
leuseralaglyilethr
530
<210>2
<211>597
<212>dna/rna
<213>pfld1的核苷酸序列(unknown)
<400>2
gcatgcaggaatctctggcacggtgctaatggtagttatccaacggagctgaggtagtcg60
atatatctggatatgccgcctataggataaaaacaggagagggtgaaccttgcttatggc120
tactagattgttcttgtactctgaattctcattatgggaaactaaactaatctcatctgt180
gtgttgcagtactattgaatcgttgtagtatctacctggagggcattccatgaattagtg240
agataacagagttgggtaactagagagaataatagacgtatgcatgattactacacaacg300
gatgtcgcactctttccttagttaaaactatcatccaatcacaagatgcgggctggaaag360
acttgctcccgaaggataatcttctgcttctatctcccttcctcatatggtttcgcaggg420
ctcatgccccttcttccttcgaactgcccgatgaggaagtccttagcctatcaaagaatt480
cgggaccatcatcgatttttagagccttacctgatcgcaatcaggatttcactactcata540
taaatacatcgctcaaagctccaactttgcttgttcatacaattcttgatattcaca597
<210>3
<211>32
<212>dna/rna
<213>引物tgz-f(unknown)
<400>3
atcgaattcatggctcatcgtggacatctaga32
<210>4
<211>30
<212>dna/rna
<213>引物tgz-r(unknown)
<400>4
attagcggccgcatttcaccatatttgtct30
<210>5
<211>26
<212>dna/rna
<213>引物fld1(unknown)
<400>5
ctagagctcgcatgcaggaatctctg26
<210>6
<211>28
<212>dna/rna
<213>引物fld2(unknown)
<400>6
gcgaggatcctgtgaatatcaagaattg28
- 还没有人留言评论。精彩留言会获得点赞!