一种β-半乳糖苷酶galRBM20_1及其制备方法和应用与流程
本发明属于基因工程技术领域,具体涉及一种β-半乳糖苷酶galrbm20_1及其制备方法和应用。
背景技术:
β-半乳糖苷酶(ec3.2.1.238)学名为β-d-半乳糖苷半乳糖水解酶,常用名为乳糖酶,该酶能催化半乳糖苷键水解,水解乳糖生成葡萄糖和半乳糖。乳糖是哺乳动物乳汁中特有的糖类,牛奶中绝大多数糖类是乳糖,但乳糖只有被β-半乳糖苷酶水解成葡萄糖和半乳糖后才能被机体小肠粘膜细胞吸收。大多数哺乳动物刚出生后体内β-半乳糖苷酶活性较高,但之后随着年龄增长活性逐渐下降,最终乳糖酶缺乏并导致乳糖不耐症。亚洲人和非洲人是世界上最易发生乳糖不耐症的人群,这类人群摄入乳糖后由于乳糖不能被小肠吸收而直接进入大肠,刺激大肠蠕动加快,造成腹鸣、腹泻等症状。
β-半乳糖苷酶广泛应用于乳制品行业。在奶类和乳制品加工过程中,加入安全的β-半乳糖苷酶,使其消化为葡萄糖和半乳糖,这样的奶类被称作低乳糖奶。低乳糖奶不仅可以满足正常消费者的需要,更可以满足乳糖不耐症者、先天性β-半乳糖苷酶缺乏者食用乳制品、充分吸收乳制品营养的需要。目前,已获得的β-半乳糖苷酶主要来源于纯培养的微生物菌株。从特殊环境中筛选特定微生物分离得到纯培养的方法较为传统,步骤繁琐。这种方法减小了实验的可靠性,很大程度上降低了实验的成功率。由于自然界中仅有约0.1-1%的微生物能通过传统纯培养分离方法获得,大大限制了β-半乳糖苷酶的开发利用。宏基因组学技术避开了微生物分离培养的问题,使得从环境中获得大量未培养微生物基因资源成为现实,极大地扩展了微生物酶及基因资源的利用空间,是发现新型生物催化剂的有效方法。
耐盐酶的研究是酶学性质板块中的重点部分,β-半乳糖苷酶虽来源广泛,但具有实际运用价值的较少。在食品工业领域,β-半乳糖苷酶被广泛运用于乳糖的水解,提高乳制品的消化率、甜度、溶解度和风味等。除此之外,β-半乳糖苷酶也被运用于生物技术和医药等领域,具有良好性质的β-半乳糖苷酶能为实际生产生活带来较好的经济效益,故其耐盐与否与其运用前景息息相关。据文献报道,具有良好耐盐性的β-半乳糖苷酶较少,而本发明的β-半乳糖苷酶galrbm20_1除了具备优良的水解性能外,还具备较强的耐盐性。
技术实现要素:
本发明的目的旨在提供一种具有耐盐性的β-半乳糖苷酶galrbm20_1及其制备方法。且其基因序列经blastn比对后,与收录于ncbi中第35家族的β-半乳糖苷酶基因序列相似性最高,具有转糖基产gos的活性。
本发明具体通过一下技术方案实现:
一种β-半乳糖苷酶galrbm20_1,所述的β-半乳糖苷酶galrbm20_1的氨基酸序列如seqidno.1所示。
本发明所述的β-半乳糖苷酶galrbm20_1的编码基因,所述的编码基因如seqidno.2所示。
一种重组载体,所述的重组载体为包含seqidno.2所示核苷酸序列的载体。
优选的,所述的载体为peasy-e2。
一种重组菌,所述的重组菌为包含seqidno.2所示核苷酸序列的菌株。
优选的,所述的菌株为大肠杆菌bl-21(de3)。
本发明所述的β-半乳糖苷酶galrbm20_1制备方法,包括以下步骤:
1)设计引物f1和r1,以滇金丝猴粪便微生物宏基因组fosmid文库混合质粒为模板,进行pcr扩增得到β-半乳糖苷酶galrbm20_1基因;
2)设计引物引物f2和r2,以β-半乳糖苷酶基因galrbm20_1为摸板,进行pcr扩增,得到含有内切酶ndeⅰ和xhoⅰ识别位点的基因片段galrbm20_1/ndeⅰ/xhoⅰ;
3)用限制性内切酶ndeⅰ与xhoⅰ切割表达载体peasy-e2,并与基因片段galrbm20_1/ndeⅰ/xhoⅰ连接;
4)将得到的表达质粒转化大肠杆菌bl-21(de3)获得重组大肠杆菌菌株bl-21(de3)/galrbm20_1的阳性克隆子,命名为bl-21(de3)/galrbm20_1;
5)将菌株bl-21(de3)/galrbm20_1接种于lb液体培养基培养,纯化得到β-半乳糖苷酶galrbm20_1。
所述的引物f1和r1的核苷酸序列如seqidno.3和seqidno.4所示。
所述的引物f2和r2的核苷酸序列如seqidno.3和seqidno.4所示。
所述的引物f2和r2分别含有限制性内切酶ndeⅰ和xhoⅰ的识别位点。
所述的lb液体培养基含有100mg/lamp。
本发明所述的β-半乳糖苷酶galrbm20_1在乳糖水解中的应用。
所述的β-半乳糖苷酶galrbm20_1酶学性质:该酶能较好的将乳糖水解为半乳糖和葡萄糖,其最适作用温度为45℃,在37℃和45℃下耐受一个小时后酶活未改变,在50℃下耐受10min具有50%的剩余酶活。其最适ph为5.0,在ph4-7之间仍能保持70%及以上的活性。
本发明的有益效果为:
本发明通过从滇金丝猴粪便微生物中获得多种酶类基因,并构建滇金丝猴粪便微生物宏基因组fosmid文库,从滇金丝猴粪便微生物宏基因组中克隆得到β-半乳糖苷酶基因galrbm20_1,重组酶具有较高的ph稳定性和热稳定性,同时还具有良好的耐盐性。本发明为β-半乳糖苷酶基因galrbm20_1的获得提供了新的来源,并通过高效重组表达为工业生产提供大量的β-半乳糖苷酶基因galrbm20_1,可广泛应用于乳制品行业。
附图说明
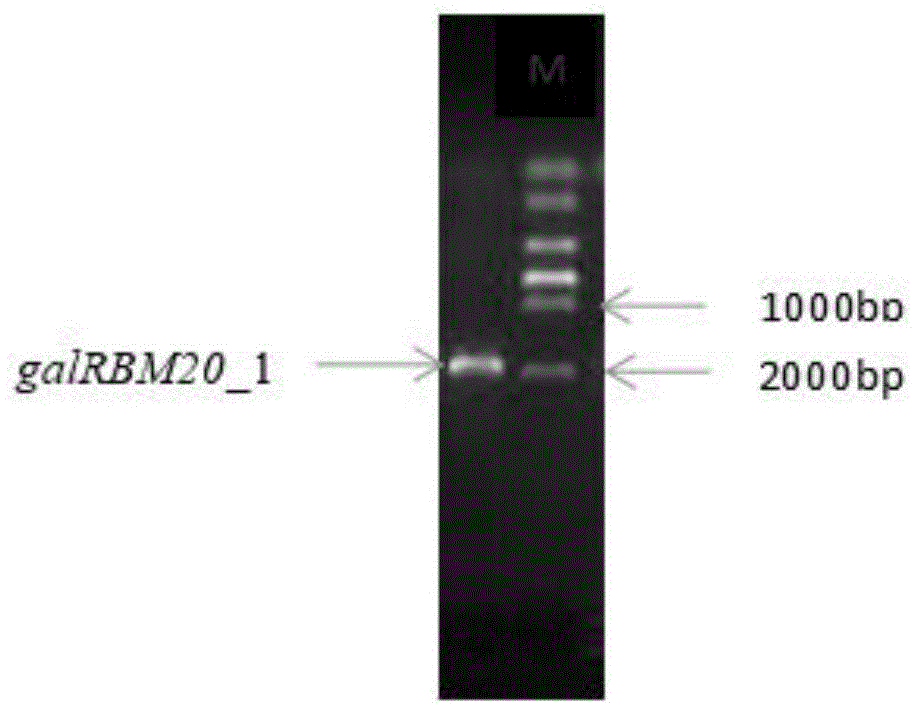
图1是基因galrbm20_1的pcr扩增;
图2是β-半乳糖苷酶galrbm20_1的sds-page分析;
图3是β-半乳糖苷酶galrbm20_1的最适温度;
图4是β-半乳糖苷酶galrbm20_1的温度稳定性;
图5是β-半乳糖苷酶galrbm20_1的最适ph;
图6是β-半乳糖苷酶galrbm20_1的ph稳定性;
图7是nacl对β-半乳糖苷酶galrbm20_1的影响;
图8是β-半乳糖苷酶galrbm20_1的nacl稳定性;
图9是β-半乳糖苷酶galrbm20_1降解乳糖的tlc分析;
图10是β-半乳糖苷酶galrbm20_1与25%(w/v)的乳糖反应24h后的产物tlc分析。
具体实施方式
下面将结合本发明具体的实施例,对本发明技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
实施例1β-半乳糖苷酶基因galrbm20_1的克隆
设计引物galrbm20_1f1(5′-atggcctgccggcgtaggggcgtt-3′)和galrbm20_1r1(5′-ccactggttgtccaggtttttcgta-3′),以滇金丝猴粪便微生物宏基因组fosmid文库混合质粒为模板,进行touch-downpcr扩增β-半乳糖苷酶基因galrbm20_1。
pcr反应体系:2×gcbufferⅰ25.0μl,2.5mmol/ldntpmix4.0μl,la-taqdnapolymerase1u,10μmol/lgalrbm20_1dnaf11.0μl,10μmol/lgalrbm20_1dnar11.0μl,宏基因组文库的fosmid混合质粒0.5μl,ddh2o补足50.0μl。
pcr反应程序为:94℃5min;94℃30s;63℃30s(每个循环降0.5℃);72℃1min,20个循环;94℃30s,53℃30s,72℃1min,10个循环;72℃10min。
pcr反应产物用1%的琼脂糖凝胶电泳分析结果(如图1)。pcr获得大量目的基因后,凝结电泳分离pcr产物,利用promega公司的dna回收试剂盒从凝胶中回收β-半乳糖苷酶基因galrbm20_1。
β-半乳糖苷酶基因galrbm20_1与线性化载体peasy-e2连接得到的原核表达质粒peasy-e2/galrbm20_1,经转化大肠杆菌bl-21(de3)后获得重组大肠杆菌菌株bl-21(de3)/galrbm20_1的阳性克隆子,命名为bl-21(de3)/galrbm20_1,送上海生工测序。
实施例2大肠杆菌重组表达系统的构建
根据β-半乳糖苷酶基因galrbm20_1的全长序列以及一步快速克隆试剂盒使用说明设计表达引物:galrbm20_1ndef2(5′-taagaaggagatatacatatggaattggctgccggcgtaggggcg-3′)与galrbm20_1xhor2(5′-gtggtggtggtggtgctcgagacaaattgcaaaggaatat-3′),其中galrbm20_1ndef和galrbm20_1xhor分别含有限制性内切酶ndeⅰ(catatg)和xhoⅰ(ctcgag)的识别位点。以β-半乳糖苷酶基因galrbm20_1为摸板,使用galrbm20_1。
ndef和galrbm20_1xhor两种引物pcr,得到含有内切酶ndeⅰ和xhoⅰ识别位点的基因片段galrbm20_1/ndeⅰ/xhoⅰ。使用限制性内切酶ndeⅰ与xhoⅰ切割表达载体peasy-e2使其线性化。采用一步快速克隆试剂盒将基因片段galrbm20_1/ndeⅰ/xhoⅰ克隆到线性化的表达载体peasy-e2上,连接得到原核表达质粒peasy-e2/galrbm20_1,转化大肠杆菌bl-21(de3)获得重组大肠杆菌菌株bl-21(de3)/galrbm20_1的阳性克隆子,命名为bl-21(de3)/galrbm20_1。
取大肠杆菌菌株bl-21(de3)/galrbm20_1以0.1%的量接种于lb(含100mg/lamp)的液体培养基中,37℃、180r/min摇床培养约3-4h(od600到0.6-1.0)后,加入iptg(终浓度为0.7mmol/l)在20℃和160r/min摇床下培养10h以诱导重组蛋白产生。8000r/min离心6min,收集菌体,用tris-hcl(ph7.0)缓冲液重悬菌体,之后再冰水浴中超声破碎细胞(300w,超声开5s,超声关7s)。超声破碎的细胞液体经冷冻离心机4℃,12000r/min离心10min,重复2-3次(无杂细胞碎片)收集上清。将上清通过亲和层析柱ni-ntaagarose进行蛋白纯化,纯化后的蛋白即为含有6×his-tag的β-半乳糖苷酶galrbm20_1。以空载体peasy-e2转化bl-21(de3)作为阴性对照检测β-半乳糖苷酶galrbm20_1的活性和通过sds-page检测纯化后酶的纯度(图2)。
实施例3最适温度和最适ph的测定
将2-硝基苯基β-d-吡喃半乳糖苷(onpg)溶于0.1mol/l的柠檬酸-磷酸氢二钠缓冲液(ph5.0),配制成浓度为2mmol/l的底物溶液。取450μl的底物于45℃水浴预热5min,加入50μl纯化并适当稀释的酶液,反应10min。之后加入1ml1mol/lnaco3终止反应,测定波长在405nm处的吸光值。
酶活单位定义:一个酶活单位(u)即在ph5.0,45℃条件下每分钟水解底物onpg生成1μmolonp所需的酶量。
在ph7.0的缓冲液中,于不同温度(0℃、10℃、20℃、30℃、40℃、45℃、50℃、55℃、60℃、65℃、70℃、80℃)条件下测定β-半乳糖苷酶galrbm20_1的酶活力。
分别配制ph2.2-10.0的不同缓冲液(ph2.2-8.0:0.1mol/l柠檬酸-磷酸氢二钠缓冲液;ph8.0-10.0:甘氨酸-氢氧化钠缓冲液),将底物onpg溶于上述配制的不同ph缓冲液中,并将β-半乳糖苷酶用相应的ph缓冲液稀释。在45℃的条件下测定各ph下的酶活力。
最适温度和温度稳定性:重组β-半乳糖苷酶galrbm20_1在45℃条件下相对酶活力最高(图3),比商业β-半乳糖苷酶(最适温度40℃)的最适温度略高。在37-50℃之间,该酶的相对酶活均在60%以上。经37℃、45℃和50℃分别处理0-1h后发现,在37℃和45°下仍能保留100%的活性,经50℃处理10min,剩余酶活为50%,处理20min,剩余酶活为20%(图4)。
热稳定性和ph稳定性:galrbm20_1在ph5.0时,相对酶活力最高,且在ph4-7之间仍能保持70%及其以上的活性(图5)。表明在天然牛乳的ph值(ph6.5)范围内,该酶具有良好的反应潜能。在ph4-8的范围内处理1h后,酶活均在60%以上,其中在ph5-8的处理范围内,酶活均在100%左右(图6)。
底物特异性和动力学参数:β-半乳糖苷酶galrbm20_1对底物onpg具有较高的比活性,其次是pnpg和乳糖,对其它硝基苯衍生物没有检测到活性(数据未显示)。其水解onpg、pnpg和乳糖的动力学参数如下(表1)。
表1β-半乳糖苷酶galrbm20_1水解底物onpg、pnpg和乳糖的动力学参数
实施例4金属离子和化学试剂对重组β-半乳糖苷酶galrbm20_1的影响测定
将各种金属离子和化学试剂(na+、li+、ca2+、k+、fe2+、zn2+、mg2+、na+、mn2+、fe3+、pb2+、i2+、co2+、ag+、cu2+、hg2+、al3+、edta、sds、ctab、β-mercaptoethanol、tritonx-100、tween-80)加入到酶促反应体系中,使其终浓度为1mmol/l(其中tritonx-100、tween-80终浓度为0.5%(体积比)),在45℃,ph5.0条件下测定β-半乳糖苷酶活力。
金属离子和化学试剂的影响:1mmol/l的hg2+、ag2+、sds完全抑制β-半乳糖苷酶galrbm20_1的活性,ag2+对该酶的活性具有很强的抑制作用,tritonx-100对该酶具有很强激活作用(表2)。与大多数报道一致,大部分二价金属离子对该β-半乳糖苷酶都具有激活作用,mg2+是生理离子,与β-半乳糖苷酶的效率增加有关(kcat上升),因此增加了对底物的亲和力(km下降)。
表2金属离子和化学试剂对重组酶活性的影响
实施例5nacl对重组β-半乳糖苷酶galrbm20_1的影响和稳定性的测定
用ph5.0(0.1mol/lph5.0柠檬酸-磷酸氢二钠)的缓冲液配制不同浓度的nacl溶液(0.5-5mol/l)。在45℃下,向不同浓度的nacl溶液加入底物和酶液进行酶促反应,以反应体系中不含nacl的反应酶活力为100%,每种不同浓度的nacl下设1个对照和3个重复测定nacl对β-半乳糖苷酶galrbm20_1的影响。将适当稀释的酶液加入不同浓度的nacl溶液中,使nacl终浓度为0.5-5mon/l,45℃恒温水浴中放置1h和24h后,于45℃、ph5.0(0.1mol/lph5.0柠檬酸-磷酸氢二钠)条件下测定β-半乳糖苷酶的剩余酶活力。
对β-半乳糖苷酶galrbm20_1进行耐盐性研究发现,反应环境中nacl的浓度为2mol/l时,该酶具有130%的相对酶活,比来自小麦秸秆降解物微生物宏基因组中的β-半乳糖苷酶高出40%。当nacl的浓度为4mol/l时,该酶仍然具有40%以上的酶活,比来自于南极嗜冷平板球菌(planococcus)的耐盐β-半乳糖苷酶高出两倍以上。经5mol/l的nacl处理1h后的β-半乳糖苷酶galrbm20_1的活性较原始酶活(100%)提高到135%,经1mol/l的nacl处理24h后,仍然剩余45%的酶活。相比于其他微生物及宏基因组来源的β-半乳糖苷酶,本实验研究的重组β-半乳糖苷酶galrbm20_1具有良好的耐盐性,在食品工业行业、生物技术领域报告基因[等方面具有良好的运用前景。
nacl浓度的影响:当体系中nacl的浓度为1mol/l时,相对酶活达153%;且nacl的浓度在0~2mol/l时,相对酶活均在100%以上;5m的nacl的反应体系中,β-半乳糖苷酶galrbm20_1剩余20%酶活(图7)。
nacl稳定性:β-半乳糖苷酶galrbm20_1经浓度为1~5mol/l的nacl在45℃处理1h后,其相对酶活均能保持在100%以上,当nacl的作用浓度为4mol/l时,相对酶活达到146%的最高值,当nacl的作用浓度为5mol/l时,仍有135%的相对酶活(图8)。经浓度为1~5mol/l的nacl在45℃处理24h后发现,当nacl的作用浓度为1mol/l时,仍具有45%的活性(图8),这些结果表明该酶具有良好的nacl稳定性。
实施例6重组β-半乳糖苷酶galrbm20_1的水解性能测定
将纯化后的β-半乳糖苷酶分别加入到1ml5%(w/v)的乳糖溶液和牛奶中,在45℃,ph5.0条件下反应,间隔不同时间取样(30min、2h、8h、12h、),煮沸5min使变性失活,用正丁醇:乙醇:水(5:3:2,v:v:v)作为展开剂,进行薄层层析(tlc)分析。
水解性能测定:当β-半乳糖苷酶galrbm20_1与5%(w/v)的乳糖反应30min后,部分乳糖已被水解。当反应时长达12h时,乳糖几乎完全被水解(图9)。β-半乳糖苷酶主要用于乳制品行业,用于生产无乳糖牛奶。经过薄层层析分析后,在12h后,β-半乳糖苷酶galrbm20_1能完全水解5%(w/v)的乳糖,说明该酶能有效的水解乳糖,因此,其在无糖牛奶的生产中具有巨大的潜力。
实施例7重组β-半乳糖苷酶galrbm20_1转糖基产gos性能测定
将纯化后的β-半乳糖苷酶加入到25%(w/w)的乳糖溶液中,在45℃,ph5.0条件下反应24h,煮沸5min使酶变性失活,以灭活后的酶液做阴性对照进行tlc分析。
转糖基产gos性能测定:β-半乳糖苷酶galrbm20_1与25%(w/v)的乳糖反应24h后,经薄层层析(tlc)分析见部分低聚半乳糖(gos)的产生(图10),表明该酶具有转糖基产gos的活性。
尽管已经示出和描述了本发明的实施例,对于本领域的普通技术人员而言,可以理解在不脱离本发明的原理和精神的情况下可以对这些实例进行多种变化、修改、替换和变型,本发明的范围由所附权利要求及其等同物限定。
序列表
<110>云南师范大学
<120>一种β-半乳糖苷酶galrbm20_1及其制备方法和应用
<160>6
<170>siposequencelisting1.0
<210>1
<211>630
<212>prt
<213>β-半乳糖苷酶(galrbm20_1)
<400>1
metalaalaglyvalglyalaleualatrpalaglyproglnalaile
151015
gluasnvalglnargproalaprothrargglyvalprovalthrphe
202530
serpheglyglygluglyasnglngluphemetleuasnglylyspro
354045
pheglnileargglyalaglumethisproglnargileproargglu
505560
tyrtrparghisargileglnthralalysalametglyleuasnthr
65707580
ilealaphetyrvalphetrpasnasphisgluglnproaspglyser
859095
pheaspphelysthraspargargasnleuserglupheleulysile
100105110
cysglnglugluglymettrpvalleupheargproglyprotyrthr
115120125
cysglyglutrpasppheglyglyleuprohistyrleuleulysasp
130135140
proasnalalysvalargthrthrgluaspalalysphemetlysala
145150155160
glnthrargtyrleuglualavalalaargvalalaglupropheleu
165170175
alalysasnglyglyproileleumetthrglnleugluasnglutyr
180185190
glyserpheglnarglysasparglystyrmetglutrpleuglnala
195200205
phetrpthrlyslysglypheglyprophetyrthrseraspglyala
210215220
glygluhispheleulysglyvalthrleuproglyvalalailegly
225230235240
leuaspproglyleuasnaspglyalatrpasnvalalaasnlyscys
245250255
asnproglyvalprovalphesersergluthrtyrproglytrpleu
260265270
arghistrpglygluglyasntrpalaprothrproglnilevalasn
275280285
hisvalargtrpphemetasplysglyargserpheserleupheval
290295300
phehisglyglythrasnpheglyphethralaglyalaasnasngly
305310315320
glyproglyasptyrglnproaspleuthrsertyrasptyrglyser
325330335
provalaspgluglnglyargmetaspglntyrtyralaglnmetarg
340345350
gluileileleuserlysleuproalaglyalaalavalproglupro
355360365
proalaaspileproalametgluileprogluphethrprolysmet
370375380
hisalaglyleutrpgluasnmetprolysproleuargasnlysphe
385390395400
proglnalaprotyrphegluglntrpglyglnasnglnglymetala
405410415
valtyrserthralavalproalaglyproalaglulysleugluphe
420425430
thrasnvalhisasptyralahisvaltyrleuasnglygluproval
435440445
glythrvalaspargargleuglyglnlysservalserleuproglu
450455460
argalalysalaglylysleuaspileleuvalglualametglyhis
465470475480
ileasnphehisilesermetgluserasparglysglyiletyrgly
485490495
provallysleuglyserarggluleulysglytrpmetvalargpro
500505510
leuproleulysalaserserilevalglnalaprolysglylysgly
515520525
valserasnlysarggluglyalahispheargalavalvalasnile
530535540
gluasnproglnaspthrpheleuaspmetalaargtyrthrlysgly
545550555560
tyrvaltrpvalasnglyileasnvalglyargtyrtrpasnvalgly
565570575
proglnleuargleutyrvalproalapropheleulyslysglyglu
580585590
asnvalileaspvalleuaspleuhismetthrserprolysproile
595600605
argglymetlysgluargasnlysgluproglylysileasnthrlys
610615620
asnleuaspasnglntrp
625630
<210>2
<211>1893
<212>dna
<213>β-半乳糖苷酶(galrbm20_1)
<400>2
atggctgccggcgtaggggcgttggcttgggccggccctcaggctattgaaaatgtgcag60
aggcctgctcctacgcggggtgtgcctgtgacgttttcttttggtggggaagggaatcag120
gagtttatgctgaatggtaagcctttccagatccgcggtgcggaaatgcaccctcagcgg180
attcccagagagtattggcgtcaccgtattcaaaccgccaaggctatgggtctgaatacc240
attgcgttttatgttttctggaatgatcatgaacagccggacggtagcttcgattttaag300
acggatcgccgtaatttgagcgagttcctcaaaatctgtcaggaagaaggaatgtgggtt360
ttattccgtcccggtccttatacttgcggagaatgggactttggcgggctgccacattat420
ttgctgaaggatcccaatgctaaggttagaacgaccgaagatgccaagttcatgaaggct480
cagactcggtatctggaagccgtggctcgtgtcgcagaaccgtttttagcgaagaatggc540
gggccgatcctgatgactcagctggaaaatgaatacggaagtttccagcgcaaggatcgt600
aagtatatggaatggttgcaggctttctggacgaagaagggctttgggccgttctataca660
tctgatggagccggggaacatttcctgaagggtgtaacgctgcccggcgtggctataggg720
ctggatcccggtttgaatgacggtgcttggaatgtggcgaacaagtgcaatcccggagtt780
cccgtattttcttcagaaacgtatccgggttggttacggcactggggtgaagggaactgg840
gctcctaccccacaaattgtgaaccatgtccgttggtttatggataaggggcgttctttc900
agtttgtttgttttccacggcggaaccaactttggctttactgcaggagccaataatggt960
ggtccgggggattatcaaccggacttgacaagttatgattatggttctcctgtggatgaa1020
cagggacgcatggatcaatattatgcccaaatgcgggaaattatcttgtctaaactccct1080
gccggagcagccgttccggaaccgccggcagatattccggcgatggaaatcccggaattt1140
acgcctaaaatgcatgccggtctttgggaaaatatgcccaagcccctacgtaataagttc1200
ccgcaagctccttattttgaacagtgggggcagaatcagggaatggctgtttacagcacc1260
gccgtgcctgccggccctgccgaaaaactggagtttaccaatgtacatgattacgcccat1320
gtgtatttgaatggtgaaccggtgggtaccgtggatcgccgactggggcagaagagtgtg1380
tcattgcctgaacgagccaaggccgggaagttggacatccttgtggaggccatgggccat1440
atcaacttccatatcagcatggaaagtgaccgtaaggggatttatggacctgtgaagttg1500
ggctcccgtgagctgaagggatggatggtcagaccgcttccgctgaaggcttcttccatt1560
gttcaggctcctaagggtaagggcgtttccaataagagggaaggcgctcatttccgcgct1620
gtcgtgaatattgaaaatccgcaggatacgttcttggatatggctcgttacaccaagggg1680
tatgtgtgggtaaacggtatcaatgtgggtcgttactggaatgtggggccgcagttgaga1740
ctgtacgttccggctcccttcctgaagaagggcgagaatgtgattgatgtgctggatttg1800
cacatgacctctcccaagcccattcgtgggatgaaggaacgtaataaggaaccgggtaag1860
attaatacgaaaaacctggacaaccagtggtaa1893
<210>3
<211>24
<212>dna
<213>人工序列(artificialsequence)
<400>3
atggcctgccggcgtaggggcgtt24
<210>4
<211>25
<212>dna
<213>人工序列(artificialsequence)
<400>4
ccactggttgtccaggtttttcgta25
<210>5
<211>45
<212>dna
<213>人工序列(artificialsequence)
<400>5
taagaaggagatatacatatggaattggctgccggcgtaggggcg45
<210>6
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>6
gtggtggtggtggtgctcgagacaaattgcaaaggaatat40
- 还没有人留言评论。精彩留言会获得点赞!