一种利用甲醇还原NAD类似物的方法与流程
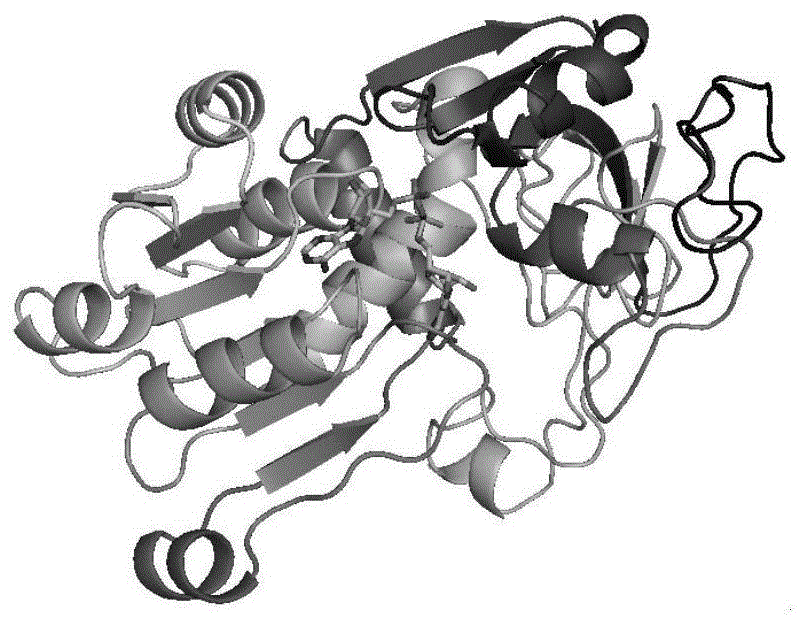
本发明属于生物技术领域,涉及辅酶烟酰胺腺嘌呤二核苷酸(nad)类似物的酶催化还原方法及其应用,具体地说,是以甲醇为还原剂,在酶催化下将nad类似物转化为其还原态,并可被其他酶用做辅酶应用于还原反应。
背景技术:
烟酰胺腺嘌呤二核苷酸(nad)及其还原态nadh是生命过程中的重要辅酶,参与生命体中的一系列氧化还原代谢及其他的重要生物化学过程。这些辅酶可用于生产手性化学品及制备同位素标记物。由于很多氧化还原酶都利用nadh或nadph作为辅酶,任何改变nad浓度及其氧化还原状态的操作都会对细胞产生全局性影响,很难在辅酶水平对生物体系中特定的氧化还原酶进行控制。由于nadh可被代谢网络中的多种途径消耗,影响了目标途径对还原力的利用效率。使用nad类似物传递还原力时,由于类似物只能被突变型氧化还原酶识别,从而在辅酶水平对目标氧化还原过程进行调控,对生物催化和合成生物学研究具有重要意义(jidb,etal.jamchemsoc,2011,133,20857-20862;wangl,etal.acscatal,2017,7,1977-1983)。
目前已经报道了几种具有良好生物相容性的nad类似物。如,烟酰胺胞嘧啶二核苷酸(ncd)、烟酰胺胸腺嘧啶二核苷酸(ntd)、烟酰胺尿嘧啶二核苷酸(nud)(jidb,etal.jamchemsoc,2011,133,20857-20862;jidb,etal.scichinachem,2013,56,296-300)。同时,也报道了一些可识别nad类似物的酶,如来自粪肠球菌enterococcusfaecalis的nadh氧化酶(nox,genbanks45681)、d-乳酸脱氢酶(dldh,genbankcaa47255)v152r突变体、苹果酸酶(me,genbankp26616)l310r/q401c突变体、苹果酸脱氢酶(mdh,genbankcaa68326)l6r突变体。
利用nad类似物及识别它们的酶,可以构建更经济有效的生物催化体系(纪德彬等.催化学报,2012,33,530-535)。通过选择合适的nad类似物及识别它们的酶,可利用粗酶液进行反应达到纯酶催化的效果,实现在辅酶水平控制复杂生物催化转化体系。目前,已经实现利用nad类似物进行胞内代谢反应的调控,通过向胞内转运ncd,dldh-v152r可利用ncd将丙酮酸还原为乳酸,实现了特异性的生物催化调控(wangl,etal.acscatal,2017,7,1977-1983)。
和使用其他氧化还原辅酶一样,nad类似物也必需再生循环。辅酶再生方式主要有酶法、电化学法、化学法和光化学法。其中酶法具有选择性高、可以与合成酶兼容及高转换数等优点。甲醇脱氢酶可以利用甲醇,将甲醇转化成甲醛,在微生物细胞中甲醛可以通过细胞内源的甲醛脱氢酶催化,转化成甲酸或者进入核糖酮单磷酸途径、四氢甲烷蝶呤途径等,产生细胞生命活动必须的代谢物和还原力,从而实现一碳资源的有效利用(müllerje,etal.metabeng,2015,28,190-201;zhangw,etal.rscadv,2017,7,4083–4091)。
虽然nad类似物再生循环对生物催化和合成生物学等领域具有重要意义,但是目前通过改造酶的结构来高效还原nad类似物的文献很少,尚没有文献报道如何改造甲醇脱氢酶来高效还原nad类似物。已报道的nad类似物还原方法(赵宗保等,一种nad类似物的还原方法.中国专利201010524767.6)利用亚磷酸脱氢酶,以亚磷酸为底物,将nad类似物还原成nadh,同时生成副产物磷酸。与亚磷酸脱氢酶催化亚磷酸化合物生成产物磷酸化合物相比,甲醇脱氢酶的底物甲醇来源丰富价格低廉,是具有较大前景的一碳化合物,利用微生物细胞,甲醇可以被甲醇脱氢酶还原最终生成高附加值化合物(whitakerwb,etal.metabeng,2017,39,49-59)。因此,通过甲醇脱氢酶还原甲醇实现nad类似物的再生是将一碳资源利用和nad类似再生相结合的新的还原方法,在实现nad类似物还原的同时,利用一碳化合物甲醇实现碳的转化和能量的传递。
基于以上背景和优势,在已有的利用甲醇及其衍生物还原nad类似物的方法基础上,本发明利用的定向进化的方法,以获得对nad类似物还原效率进一步提升的突变体。
技术实现要素:
本发明涉及一种辅酶nad类似物的酶催化还原方法,具体来说,以甲醇、氘代甲醇中的一种或者两种的任意比例组合的底物为还原剂,可利用该还原剂的酶为催化剂,将nad类似物转化为相应的还原态。这些nad类似物还原态可作为其他氧化还原酶的辅酶,用于还原反应。因此,本发明的方法可应用在生物催化和生物转化领域,具有重要价值。
本发明涉及一种利用甲醇还原nad类似物的方法,其特征在于:以甲醇、氘代甲醇中的一种或者两种的任意比例组合的底物为还原剂,以可利用甲醇的酶为催化剂,在ph4-9的缓冲体系中,反应温度15-40℃,酶的终浓度为1μg/ml-600μg/ml,nad类似物的终浓度为0.001mm-20mm、甲醇化合物的终浓度为1mm-1000mm下,反应2-120min,获得还原型nad类似物。所述的缓冲体系包括但不限于磷酸缓冲液,tris-hcl缓冲液,hepes缓冲液,mes缓冲液,pipes缓冲液,乙酸-乙酸钠缓冲体系中的一种或两种以上。
nad类似物包括ncd、烟酰胺胸腺嘧啶二核苷酸(ntd)和烟酰胺尿嘧啶二核苷酸(nud),他们具有如下的化学结构:
本发明涉及的nad类似物参照文献方法(jidb,etal.scichinachem,2013,56,296-300)制备。
本发明使用的甲醇脱氢酶为具有以甲醇为还原剂,催化还原nad类似物为相应还原态的活性蛋白质。这些酶为来源于bacillusstearothermophilus的甲醇脱氢酶bsmdh(ncbi蛋白数据库编号p42327.1,含1至339位完整氨基酸序列)的突变体(突变位点用氨基酸序号和突变前后的氨基酸名称表示,如v237t表示第237位氨基酸由v突变成t,其余位点类似),包括突变体bsmdh-v237t/n240e,bsmdh-v237t/n240e/k241a,bsmdh-y171r/v237t/n240e/k241a,bsmdh-y171r/i196v/v237t/n240e/k241a。v237t代表第237位氨基酸由v突变成t,n240e代表240位的氨基酸由n突变为e,其它类同。这些酶的表达和纯化参照在大肠杆菌中表达其他氧化还原酶的文献方法进行(jidb,etal.jamchemsoc,2011,133,20857)。
本发明涉及的nad类似物和nad一样含有烟酰胺单核苷酸单元,该单元的还原态为1,4-二氢烟酰胺单核苷酸。因此,nad类似物还原态产物在340nm附近的紫外光谱区有较强吸收,摩尔消光系数ε340约为6220m-1·cm-1(jidb,etal.creationofbioorthogonalredoxsystemsdependingonnicotinamideflucytosinedinucleotide.jamchemsoc.2011,133,20857-20862)。本发明利用该性质分析nad类似物还原过程。利用液相色谱定量nad类似物及其还原态产物的条件为:液相色谱仪为agilent1100,分析柱为zorbax150mm×3.0mm(3.5μm),流动相为5mm硫酸四丁胺,流速为0.5ml/min。每个样品检测20min。检测波长为260nm(辅因子及其还原态在260nm有较强的光吸收)和340nm(还原态辅酶在340nm有较强的光吸收)。
制备的nad类似物还原态产物,可被其他酶用作辅酶,应用于还原反应。因此,本发明可视为一种再生循环nad类似物还原态的技术。通过本发明的技术,将甲醇的还原力转移并贮存在nad类似物还原态中,以便于对其他底物进行选择性还原。
所用缓冲体系中甲醇脱氢酶的终浓度为1μg/ml-600μg/ml(优选100μg/ml-500μg/ml,再优选100μg/ml-300μg/ml)、nad类似物的终浓度为0.001mm-20mm(优选0.01mm-20mm,再优选0.1mm-10mm)、甲醇的终浓度为1mm-1000mm(优选10mm-1000mm,再优选100mm-900mm),所述的缓冲体系包括但不限于磷酸缓冲液,tris-hcl缓冲液,hepes缓冲液,mes缓冲液,pipes缓冲液中的一种或两种以上。
利用所述甲醇脱氢酶还原nad类似物,为包括但不局限于催化丙酮酸还原的苹果酸酶me(ncbino.np_415996.1)突变体me-l310r/q401c;催化丙酮酸还原的乳酸脱氢酶dldh(genebankno.caa47255.1,pdbid:2dld)突变体dldh-v152r、v152r/i177r/a212g或v152r/i177s/a212d或酿酒酵母醇脱氢酶提供还原型辅酶时,采用ph4-9的缓冲体系,反应温度15-40℃。
所述可利用甲醇的酶表达在微生物的细胞内,nad类似物可通过nad转运蛋白atndt2(accessionno.nc_003070)或ntt4(haferkampi,etal.nature,2004,432,622-625.)转入胞内;甲醇渗透进入细胞,nad类似物还原反应在细胞内进行。
所述表达甲醇脱氢酶并用于胞内还原nad类似物的微生物细胞包括但不局限于原核微生物,如大肠杆菌、乳酸乳球菌等或真核微生物,如酿酒酵母、红酵母或里氏木霉等。
本发明的优点和有益效果:作为还原剂的甲醇价格低廉,储量丰富,是具有较大前景的一碳资源。酶催化甲醇的反应可用于生物反应体系,以微生物发酵等手段,将甲醇转化成高附加值的化学品,实现还原力的选择性传递和碳的有效利用。此外,还可以通过使用氘代甲醇,可获得氘代nad类似物还原态,用于制备高纯度氘取代生物催化产物。
附图说明:
图1为bsmdh-y171r/i196e/v237t/n240e/k241a复合物晶体结构图;
图2为bsmdh-y171r/v237t/n240e/k241a复合物晶体结构图。
具体实施方式
以下实施例将有助于本领域普通技术人员进一步理解本发明,但不以任何形式限制本发明。
对比例1:无酶条件下甲醇与nad类似物的反应
nad类似物(ncd、ntd和nud)参照文献方法(jidb,etal.scichinachem,2013,56,296-300)制备。nad类似物用水制成浓度为20mm的溶液,备用。
将1mm的nad类似物底物和8mm的甲醇溶解在1ml浓度为50mm,ph7.5的tris-hcl缓冲液中,混匀,30℃反应2h,取20μl分析。
用hplc检测nad类似物底物及其还原态产物。液相色谱仪为agilent1100,分析柱为zorbax150mm×3.0mm(3.5μm),流动相为5mm硫酸四丁胺,流速为0.5ml/min。每个样品检测20min。检测波长为260nm(辅因子及其还原态在260nm有较强吸收)和340nm(还原态辅酶在340nm有较强的光吸收)。
分析发现,所有反应的样品在340nm没有特征峰,在260nm仅检测到与nad类似物保留时间相同的特征峰。说明在无酶条件下甲醇不能直接还原nad类似物。
对比例2:酶失活条件下甲醇与nad类似物的反应
将来源于bacillusstearothermophilus的甲醇脱氢酶bsmdh(ncbi蛋白数据库编号p42327.1,含1至339位完整氨基酸序列)在98℃水浴加热60min,备用。参考文献测定nadh的方法(guoq,etal.biochemistry,2016,55,2760-2771),以nad为底物,检测表明样品失去了催化还原nad为nadh的活性。
将nad类似物ncd、ntd和nud,逐一按照下述方法进行反应:将1mm的nad类似物、8mm甲醇和80μg灭活的甲醇脱氢酶bsmdh溶解在1ml浓度50mm,ph7.5的tris-hcl缓冲液中混匀,30℃反应2h,取20μl分析。
按对比例1的方法分析发现,所有反应的样品在340nm没有特征峰,在260nm仅检测到与nad类似物保留时间相同的特征峰。说明加热灭活的酶不能催化甲醇还原nad类似物。
实施例1:以甲醇为还原剂,甲醇脱氢酶催化还原nad类似物
将nad及其类似物ncd、ntd或nud,与甲醇脱氢酶bsmdh、bsmdh-v237t,bsmdh-v237t/n240e,bsmdh-v237t/n240e/k241a,bsmdh-y171r/v237t/n240e/k241a,bsmdh-y171r/i196v/v237t/n240e/k241a、逐一进行nad类似物-甲醇脱氢酶组合,按下述方法进行反应:将1mm的nad或类似物、8mm的甲醇和80μg的甲醇脱氢酶溶解在1ml浓度50mm,ph7.5的hepes缓冲液中混匀,30℃反应20min,取20μl分析。
按对比例1的分析方法分析发现,所用样品在340nm均出现了特征吸收峰,但不同组合得到的吸收峰强度有明显差别,说明甲醇脱氢酶可以催化甲醇还原nad类似物。由于nad类似物还原态产物的摩尔消光系数ε340约为6220m-1·cm-1,与nadh相同,用nadh标准品绘制曲线,得到定量结果(表1)。可见,bsmdh的催化活性整体较低,其他几种甲醇脱氢酶具有较好的活性。可依据不同的nad类似物,选择合适的甲醇脱氢酶。突变体bsmdh-y171r/i196e/v237t/n240e/k241a和bsmdh-y171r/v237t/n240e/k241a还原ncd的效率分别为96%和93%,比之前专利中获得的突变体bsmdh-i196q、bsmdh-k242y/a243m和bsmdh-d195e/a238v催化效率(分别为89%、46%和83%,专利申请号201811154784.8)有所提高。突变体bsmdh-v237t/n240e/k241a还原nud的效率为91%,高于之前专利中所获突变体对nud的活性(突变体bsmdh-i196q对nud转化率最高为89%,专利申请号201811154784.8)。突变体bsmdh-v237t/n240e对ntd还原效率为85%,高于之前专利中所获突变体对ntd的活性(bsmdh-d195e/a238v对ntd转化率最高,为69%,专利申请号201811154784.8)此外,本发明涉及的甲醇脱氢酶突变体对ntd和nud的催化效率整体上高于已有突变体(见上述专利,中国专利申请号201811154784.8)。
实施例1的结果表明,甲醇脱氢酶可以有效催化甲醇还原本发明涉及的nad类似物,制备相应的还原态产物。综合实施例1、对比例1和对比例2的结果说明,以甲醇为还原剂,还原nad类似物,活性甲醇脱氢酶起到了不可替代的作用。
表1甲醇脱氢酶催化甲醇还原nad及其类似物的实验结果
实施例2:还原态nad类似物的制备
将实施例1的反应体系放大,可用于制备还原态nad类似物。以nudh为例,说明制备过程。将20mm的nud、20mm的甲醇和5mg的甲醇脱氢酶bsmdh-v237t溶解在10ml浓度为50mm,ph为5.7的磷酸钠缓冲液中混匀,30℃反应80min。反应结束后直接冷冻干燥,浓缩至总体积约为4ml,用甲酸型阴离子交换树脂柱分离,在紫外波长340nm跟踪收集产物,冷冻干燥,得白色粉末5.6mg,产率约44%。
将上述白色粉末样品进行高分辨质谱分析,测得精确分子量(m+h)+为643.1026,与nudh的理论分子量(c20h29n4o16p2+,643.1054)一致,表明得到了还原态产物nudh。
按照实施例2的方法用相同用量的nud、亚磷酸钠和亚磷酸脱氢酶rspdh-i151r生产nad类似物,得到了还原态的nudh,产率约35%。说明甲醇脱氢酶bsmdh-v237t与亚磷酸脱氢酶rspdh-i151r均能催化相应的底物生产nad类似物,且产率接近。
实施例3:以甲醇为还原剂,甲醇脱氢酶催化还原nad类似物
将0.1mm的nud、0.8mm的甲醇和80μg甲醇脱氢酶bsmdh-v237t/n240e溶解在1ml浓度为50mm,ph8.0的pipes缓冲液中混匀,40℃反应3min,取20μl分析。
按对比例1的方法分析发现,样品在340nm出现了特征吸收峰。生成的nbrcdh浓度达到60μm,即产率达到60%。
综合实施例1和实施例3的结果表明,甲醇脱氢酶催化还原nad类似物的反应中,采用甲醇为还原剂,可使nad类似物还原。
按照实施例3的方法用相同用量的nud、亚磷酸钠和亚磷酸脱氢酶rspdh-i151r生产nad类似物,生成的nudh浓度达到43μm,即产率达到40%。说明甲醇脱氢酶bsmdh-v237t/n240e与亚磷酸脱氢酶rspdh-i151r均能催化相应的底物生产nad类似物,甲醇脱氢酶bsmdh-v237t催化的反应产率高于亚磷酸脱氢酶rspdh-i151r。
实施例4:以氘代甲醇为还原剂,甲醇脱氢酶催化还原nad类似物
将1mm的ncd、4mm氘代甲醇和40μg甲醇脱氢酶bsmdh-v237t/n240e/k241a溶解在1ml浓度50mm,ph5.0的mes缓冲液中混匀,10℃反应120min,取20μl分析。
按对比例1的分析方法分析发现,样品在340nm出现了特征吸收峰。产物ncdh浓度达到0.61mm,即产率为60%。
将样品进行高分辨质谱分析,测得精确分子量(m-h)-为641.1118,与ncd2h的理论分子量(c20h272hn5o15p2-,641.1125)一致,表明得到了氘代ncd还原态产物。
实施例4的结果表明,甲醇脱氢酶可用氘代甲醇为还原剂,催化还原nad类似物生成相应的氘代还原态产物。
按照实施例4的方法用相同用量的ncd、亚磷酸钠和亚磷酸脱氢酶rspdh-i151r/e213c生产氘代nad类似物,产物ncdh浓度达到0.38mm,即产率为38%。说明甲醇脱氢酶bsmdh-v237t/n240e/k241a与rspdh-i151r/e213c均能催化相应的底物生产nad类似物,甲醇脱氢酶rspdh-i151r/e213c催化的反应产率高于亚磷酸脱氢酶rspdh-i151r/e213c。
实施例5:甲醇脱氢酶、苹果酸酶me-l310r/q401c和nad类似物体系催化丙酮酸还原羧化制备苹果酸
参照文献(jidb,etal.jamchemsoc,2011,133,20857-20862)方法纯化苹果酸酶(ncbino.np_415996.1)突变体me-l310r/q401c,备用。me-l310r/q401c偏好类似物ncdh而对nadh活性低,需要以ncdh为辅因子。
苹果酸酶me-l310r/q401c可催化的反应为:丙酮酸+co2+ncdh→苹果酸+ncd。甲醇脱氢酶催化的反应为:甲醇+ncd→甲醛+ncdh。将两反应合并,净反应为:甲醇+丙酮酸+co2→苹果酸+甲醛。因此甲醇脱氢酶和苹果酸酶构成的体系可以实现以甲醇为还原剂,还原羧化丙酮酸生成苹果酸。该体系中,nad类似物被再生循环,而且达到了固定co2的效果,具有一定的潜力。以甲醇为底物的反应体系,在再生还原型nad类似物的同时,将来自甲醇的碳引入到细胞代谢体系中,同时实现底物的物质代谢和能量代谢。其中具有代表性的实验过程如下:
采用50mm,ph5.0的tris-hcl缓冲体系,100μl的反应体系组成为:800mm甲醇、50mm丙酮酸、0.1mmncd、1.0mmmncl2、10mm碳酸氢钠、0.05mg/mlbsmdh-y171r/v237t/n240e/k241a和0.06mg/mlme-l310r/q401c。10℃反应120min,加900μl乙腈水混合液(体积比,乙腈:水=4:1)终止反应(rabinowitzjd,etal.analchem,2007,79,6167-6173)。
利用美国戴安公司ics-2500离子色谱系统在ed50脉冲电化学检测模式下分析测定反应溶液中苹果酸,丙酮酸和甲醇的含量。使用ionpacas11-hc阴离子交换分析柱(200mm×4mm),ionpacag11-hc阴离子交换保护住(50mm×4mm)。分析条件:流动相为24mmnaoh,流速1ml/min,柱温:30℃,进样量25μl。检测结果表明,反应液含0.1mm甲醇、46.1mm丙酮酸和3.6mm苹果酸。
在进行上述反应时,设置了另外4组对照实验体系,分别缺少甲醇、ncd、bsmdh-y171r/v237t/n240e/k241a或me-l310r/q401c中的一种,分析发现这些反应没有产生苹果酸。根据反应的化学计量关系可知,ncd再生循环利用360次。
在进行上述反应时,还设置了用氘代甲醇代替甲醇,其他成分和条件相同,分析发现反应液丙酮酸浓度降低为46.3mm同时产生3.5mm苹果酸,说明该体系可用氘代甲醇为还原剂,达到甲醇为还原剂相当的效率。
按照实施例5的方法,用亚磷酸脱氢酶rspdh-i151r/e213c、苹果酸酶me-l310r/q401c和nad类似物体系催化丙酮酸还原羧化制备苹果酸。用相同的反应体系和分析方法,检测结果表明,反应液含有0.2mm亚磷酸、47.1mm丙酮酸和2.6mm苹果酸。说明甲醇脱氢酶、苹果酸酶me-l310r/q401c和nad类似物体系催化丙酮酸还原羧化制备苹果酸要高于亚磷酸脱氢酶rspdh-i151r/e213c、苹果酸酶me-l310r/q401c和nad类似物体系的催化活性。
实施例6:甲醇脱氢酶、d-乳酸脱氢酶dldh-v152r和nad类似物体系催化丙酮酸还原制备乳酸
参照文献(jidb,etal.jamchemsoc,2011,133,20857-20862)方法纯化d-乳酸脱氢酶(genebankno.caa47255.1,pdbid:2dld)突变体dldh-v152r,备用。d-乳酸脱氢酶dldh-v152r偏好nad类似物,需要以还原型类似物为辅因子。
乳酸脱氢酶催化的反应为:丙酮酸+ntcdh→d-乳酸+ntcd。甲醇脱氢酶催化的反应为:甲醇+ntcd→甲醛+ntcdh。将两反应合并,总反应为:甲醇+丙酮酸→d-乳酸+甲醛。因此,甲醇脱氢酶和d-乳酸脱氢酶构成的体系可实现以甲醇为还原剂,还原丙酮酸生成d-乳酸。该体系中,nad类似物被循环再生,具有一定的应用潜力。其中具有代表性的实验过程如下:
采用50mm,ph8.0的mes缓冲液体系,100μl的反应体系组成为:8mm甲醇、4.0mm丙酮酸、0.1mmntcd、0.05mg/mlbsmdh-y171r/i196v/v237t/n240e/k241a和0.06mg/mldldh-v152r。40℃反应10min,加900μl乙腈水混合液(体积比,乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统分析,结果表明反应液含有0.6mm甲醇、3.3mmd-乳酸、0.6mm丙酮酸。
实验结果表明,该体系利用甲醇为还原剂,近定量地将丙酮酸还原为乳酸,取得了很高的原料利用效率。根据反应的化学计量关系可知,ntcd再生循环利用33次。
按照实施例6的方法,用甲醇脱氢酶突变体bsmdh-i196q(见之前专利,中国专利申请号201811154784.8)、d-乳酸脱氢酶dldh-v152r和nad类似物体系催化丙酮酸还原制备乳酸。用相同的反应体系和分析方法,检测结果表明,反应液含有0.6mm甲醇、3.0mmd-乳酸、0.8mm丙酮酸。说明甲醇脱氢酶突变体bsmdh-y171r/i196v/v237tn240e/k241a、d-乳酸脱氢酶dldh-v152r和nad类似物体系催化丙酮酸还原制备乳酸要高于甲醇脱氢酶突变体bsmdh-i196q、d-乳酸脱氢酶dldh-v152r和nad类似物类似物体系的催化活性。
实施例7:甲醇脱氢酶、d-乳酸脱氢酶dldh-v152r和nad类似物体系催化丙酮酸还原制备乳酸
按照实施例6的方法,将反应的缓冲体系改为ph9.0的tris-hcl缓冲液,按照相同的反应条件和分析方法,结果表明反应终止后的反应液含有8mm甲醇、2.9mmd-乳酸、0.9mm丙酮酸。说明在ph9.0的条件下,含有突变体bsmdh-y171r/i196e/v237t/n240e/k241a的体系,可以以甲醇为还原剂,近定量地将丙酮酸还原为乳酸,取得了很高的原料利用效率。但转化效率低于ph8.0的反应体系。
实施例8:甲醇脱氢酶、d-乳酸脱氢酶dldh-v152r和nad类似物体系催化丙酮酸还原制备乳酸
按照实施例6的方法,将反应的缓冲体系改为ph4.0的乙酸-乙酸钠缓冲液,按照相同的反应条件和分析方法,结果表明反应终止后的反应液含有6mm甲醇、1.7mmd-乳酸、2.2mm丙酮酸。说明在ph4.0的条件下,含有bsmdh-y171r/i196e/v237t/n240e/k241a的体系,可以以甲醇为还原剂,近定量地将丙酮酸还原为乳酸,取得了相应的原料利用效率。但转化效率低于ph8.0和ph9.0的反应体系。
实施例9:醇脱氢酶利用还原态nad类似物催化醛还原反应
参照实施例2的方法制备还原态类似物ntdh,备用。
采用20mm、ph7.5的磷酸钠缓冲液体系,500μl反应体系组成为:3.0mm乙醛、2.0mmntdh、0.1mg/ml来源于酿酒酵母的醇脱氢酶(购自sigma公司,货号:a3263)。30℃反应,用分光光度计在紫外波长340nm处跟踪反应。反应30min后,体系中ntdh降低为0.8mm。同时,体系中产生了1.1mm乙醇。
在没有添加酿酒酵母醇脱氢酶的对照实验体系中,反应30min后ntdh浓度未见明显变化。实施例7的结果说明还原态nad类似物可被氧化还原酶用作辅酶,催化还原反应。
实施例10:甲醇脱氢酶介导的胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶和nad类似物转运蛋白同时在宿主中表达,形成一种依赖nad类似物的生物催化体系。当培养基中的甲醇和nad类似物进入宿主细胞后,启动该生物催化体系。因此,应用甲醇脱氢酶介导的胞内还原nad类似物的技术,可将胞外还原力选择性传递到胞内目标氧化还原反应。下面以改造的大肠杆菌escherichiacolixz654(zhangx,etal.applenvironmicrobiol,2011,77,427-434),构建生产苹果酸的工程菌株为例予以说明。
nad转运蛋白atndt2(accessionno.nc_003070)具有较宽泛的底物谱(palmierif,etal.jbiolchem,2009,284,31249-31259),可以转运ncd。将表达转运蛋白的atndt2的基因由gapap1启动子(charpentierb,etal.jbacteriol,1994,176,830-839)控制表达。将编码bsmdh-y171r/i196e的基因和编码me-l310r/q401c的基因,由异丙基硫代半乳糖(iptg)诱导的lac启动子控制,将上述三种表达盒通过替换puc18的lacz基因克隆到同一质粒获得工程质粒。
将上述工程质粒导入e.colibl21(de3)获得工程菌株e.coliwjt007。在lb培养基中诱导工程菌株e.coliwjt007表达上述三种功能蛋白,培养基中添加50μg/ml氨苄青霉素、0.1mm的iptg,在25℃、200rpm的摇床中培养48h至菌体密度od600nm为4.5,2000×g离心6min收集菌体。
用ph7.5的mops培养基洗涤重悬菌体,将菌体密度od600nm调整到9。上述工程菌悬液中加入10mm碳酸氢钠、10mm丙酮酸、8mm甲醇、0.1mm的ncd,分别在16℃、30℃、42℃的200rpm摇床中厌氧反应4h,取100μl加900μl乙腈水混合液(乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统分析,结果表明16℃反应液含2mm甲醇、2.8mm苹果酸、7.1mm丙酮酸。30℃反应液含0.2mm甲醇、4.3mm苹果酸、5.4mm丙酮酸。42℃反应液含0.5mm甲醇、3.8mm苹果酸、5.6mm丙酮酸。
在只添加甲醇和ncd中的一种及不添加甲醇和ncd的对照实验中,苹果酸浓度分别为2.2mm、1.9mm和1.9mm。
实验结果表明在全细胞催化过程中,甲醇脱氢酶bsmdh-y171r/i196e通过氧化甲醇为me-l310r/q401c提供ncdh,催化丙酮酸还原羧化成苹果酸,使苹果酸产量由1.9mm提高到4mm。只添加甲醇时苹果酸产量没有明显增加,只添加ncd时苹果酸不增加。
实施例8说明,在全细胞催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态nad类似物,被me-l310r/q401c用作辅酶应用于还原反应,可作为一种通过提供氧化还原力调控微生物体内苹果酸的代谢强度的方式。
按照实施例8的方法,将表达rspdh-i151r/e213c的基因和me-l310r/q410c的基因由异丙基硫代半乳糖(iptg)诱导的lac启动子控制,构建相应的工程菌株,用同样的实验和分析方法检测各成分的含量。结果表明16℃反应液含1.8mm亚磷酸、2.8mm苹果酸、7.1mm丙酮酸。30℃反应液含0.1mm亚磷酸、4.2mm苹果酸、5.6mm丙酮酸。42℃反应液含0.5mm亚磷酸、3.8mm苹果酸、5.6mm丙酮酸。说明甲醇脱氢酶bsmdh-y171r/i196e参与的上述催化体系与亚磷酸脱氢酶rspdh-i151r/e213c参与的类似的催化体系比,前者催化效率在30℃时稍高,在16℃和42℃时二者效率相当。
实施例11:甲醇脱氢酶介导的胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶和nad类似物转运蛋白同时在宿主中表达,形成一种依赖nad类似物的生物催化体系。下面以改造大肠杆菌escherichiacolixz654(zhangx,etal.applenvironmicrobiol,2011,77,427-434),构建生产乳酸的工程菌株为例予以说明。
nad转运蛋白ntt4(haferkampi,etal.nature,2004,432,622-625.)可以转运ngd。将上述三种表达转运蛋白ntt4的基因由gapap1启动子控制表达。将编码bsmdh-y171r/v237t的基因和编码dldh-v152r的基因,由异丙基硫代半乳糖苷(iptg)诱导的lac启动子控制,将上述三种表达盒通过替换puc18的lacz基因克隆到同一质粒上获得工程质粒。
将上述工程质粒导入e.colibl21(de3)获得工程菌株e.coliwjt008。在lb培养基中诱导工程菌e.coli008表达上述三种功能蛋白,培养基中添加50μg/ml氨苄青霉素、0.1mm的iptg,在25℃的200rpm摇床中培养48h至菌体密度od600nm为4.5,2000×g离心6min收集菌体。
用ph8.0的m9培养基洗涤重悬菌体,将密度od600nm调整到9。上述工程菌悬液中加入10mm丙酮酸、5mm甲醇、0.1mmnud,分别在30℃的200rpm摇床中厌氧反应3h,取100μl加900μl乙腈水混合液(体积比,乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统进行分析,结果表明30℃反应液含0.1mm甲醇、4.9mm乳酸、4.7mm丙酮酸。
在只添加甲醇和nud中的一种及不添加甲醇和nud的对照实验中,乳酸浓度分别为0.9mm、0.9mm和0.6mm。
实验结果表明在全细胞催化过程中甲醇脱氢酶bsmdh-y171r/v237t通过氧化甲醇为dldh-v152r提供nudh,催化丙酮酸还原成乳酸,使乳酸产量由0.6mm提高到4.9mm。只添加甲醇或nud时乳酸产量没有明显增加。
实施例9说明,在全细胞催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态的nad类似物,被dldh-v152r用作辅酶应用于还原反应,可作为一种通过提供氧化还原力调控微生物体内乳酸的代谢强度的方式。
按照实施例9的方法,将表达rspdh-i151r/i218f的基因和dldh-v152r的基因由异丙基硫代半乳糖(iptg)诱导的lac启动子控制,构建相应的工程菌株,用同样的实验和分析方法检测各成分的含量。结果表明30℃反应液含0.1mm亚磷酸、4.8mm乳酸、4.6mm丙酮酸。说明甲醇脱氢酶bsmdh-y171r/v237t参与的上述催化体系与亚磷酸脱氢酶rspdh-i151r/i218f参与的类似的催化体系效率接近。
实施例12:甲醇脱氢酶介导的透性化细胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶同时在宿主细胞中表达,形成一种依赖nad类似物的生物催化体系。当培养基中的甲醇和nad类似物进入宿主细胞后,启动该生物催化体系。
将编码bsmdh-v237t/n240v/k241a的基因和编码dldh-v152r的基因,由异丙基硫代半乳糖苷(iptg)诱导的lac启动子控制,将上述两种表达盒通过替换puc18的lacz基因克隆到同一质粒上获得工程质粒。
将上述工程质粒导入e.colixz654获得工程菌株e.coliwjt009。在lb培养基中诱导工程菌e.coliwjt009表达上述两种功能蛋白,培养基中添加100μg/ml氨苄青霉素、1mm的iptg,在25℃、200rpm的摇床中培养48h至菌体密度od600nm为4.5,2000×g离心6min收集菌体,用浓度50mm,ph7.5的tris-cl洗涤重悬菌体,将菌体密度od600nm调整到9,按文献方法使细胞透性化(zhangw,etal.applenvironmicrobiol,2009,75,687-694),制备方式为,取5ml的冻存细胞室温下水浴解冻,加入5mm的edta和体积比1%的甲苯,30℃、200rpm摇床温浴30min,然后再在4℃放置1h。2000g离心6min去除含有edta和甲苯的上清,用浓度50mm、ph7.5的tris-cl洗两遍后重悬于5ml的浓度50mm、ph5.0的tris-cl,获得透性化细胞。
上述用浓度50mm、ph5.0的tris-cl重悬的透性化工程菌悬液中加入10mm丙酮酸、5mm甲醇、0.1mm的ncd,在30℃、200rpm的摇床中厌氧反应0.5h。取100μl样品,加入900μl乙腈水混合液(乙腈:水=4:1)终止反应。
参照实施例5的方法用离子色谱系统分析,结果表明反应液含2mm甲醇、2.6mm乳酸和7.1mm丙酮酸。
在只添加甲醇和ncd的一种及不添加甲醇和ncd的对照实验中,乳酸浓度分别为0.6mm、0.4mm和0.3mm。
实验结果表明在全细胞催化过程中甲醇脱氢酶bsmdh-v237t/n240v/k241a通过氧化甲醇为dldh-v152r提供ncdh,催化丙酮酸还原生成乳酸,使乳酸产量由0.3mm提高到2.6mm。只添加甲醇或ncd时苹果酸产量没有明显增加。
实施例10说明,在全细胞催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态的nad类似物,被dldh-v152r用作辅酶应用于还原反应,可作为一种通过提供氧化还原力调控微生物体内乳酸的代谢强度的方式。
按照实施例10的方法,将表达rspdh-i151r的基因和me-l310r/q401c的基因由异丙基硫代半乳糖(iptg)诱导的lac启动子控制,构建相应的工程菌株,用同样的实验和分析方法检测各成分的含量。结果表明30℃反应液含2.4mm亚磷酸、2.5mm乳酸和7.8mm丙酮酸。说明甲醇脱氢酶bsmdh-v237t/n240v/k241a参与的上述催化体系效率高于亚磷酸脱氢酶rspdh-i151r参与的类似的催化体系。
实施例13:甲醇脱氢酶介导的透性化乳酸乳球菌(lactococcuslactis)as1.2829细胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶同时在乳酸乳球菌中表达,形成一种依赖于nad类似物的生物催化体系。当培养基中的甲醇和nad类似物进入宿主细胞后,启动该生物催化体系。
将编码bsmdh-y171r的基因和编码dldh-v152r的基因,由组成型表达启动子p32控制,将上述两种表达盒通过替换pmg36e的p32表达盒(guchtemv,etal.applenvironmicrobiol,1989,55,224-228.)获得工程质粒。
将上述工程质粒导入乳酸乳球菌获得工程菌株l.lactiswjt010。用ph6.8的含有10g/l蔗糖、10g/l酵母提取物、10g/l蛋白胨、10g/l的kh2po4、2g/l的mgso4和5mg/l红霉素的培养基诱导工程菌l.lactiswjt010表达上述两种功能蛋白,在25℃、200rpm的摇床中培养48h至菌体密度为4.5,2000×g离心6min收集菌体,用浓度50mm、ph7.5的tris-cl洗涤重悬菌体,将菌体密度od600nm调整至9。参照实施例10的方法使细胞透性化,制备方式为:取5ml的冻存细胞在室温下水浴解冻,加入5mm的edta和体积比1%的甲苯,30℃、200rpm摇床中温浴30min,然后再在4℃放置1h。2000g离心6min去除含有edta和甲苯的上清,用浓度50mm、ph7.5的tris-cl洗两遍后重悬于5ml的浓度为50mm、ph7.5的tris-cl中,获得透性化细胞。
上述用50mm、ph7.5的tris-cl重悬的透性化工程菌悬液中加入10mm丙酮酸、5mm甲醇。0.1mm的nfcd,在30℃、200rpm的摇床中厌氧反应1h。取100μl反应液,加入900μl乙腈水混合液(乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统分析,结果表明反应液中含有1.9mm甲醇、2.7mm乳酸和7.1mm丙酮酸。
在只添加甲醇和nfcd的一种及不添加甲醇和nfcd的对照实验中,乳酸浓度分别为0.4mm、0.4mm和0.2mm。
实施例11说明,在乳酸乳球菌全细胞催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态的nad类似物,被dldh-v152r用作辅酶应用于还原反应,乳酸的积累量与不添加甲醇和nfcd的对照实验相比提高了13.5倍,因此可作为一种通过提供氧化还原力调控微生物体内乳酸代谢强度的方式。
实施例14:甲醇脱氢酶介导的透性化酿酒酵母(saccharomycescerevisiae)by4741细胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶同时在酿酒酵母细胞中表达,形成一种依赖于nad类似物的生物催化体系。当培养基中的甲醇和nad类似物进入宿主细胞后,启动该生物催化体系。
将编码bsmdh-y171r/i196r/v237t/n240e的基因和编码dldh-v152r的基因,由tef组成型启动子、cyc1终止子控制,将上述两种表达盒整合到p416酵母游离型穿梭表达载体获得工程质粒。
将上述工程质粒导入酿酒酵母获得工程菌株s.cerevisiaewjt011。用ph6.0的含有20g/l葡萄糖、10g/l酵母提取物、20g/l蛋白胨的yepd培养基培养工程菌s.cerevisiaewjt011表达上述两种功能蛋白,在25℃、200rpm的摇床中培养48h至菌体密度od600nm为4.5,2000×g离心6min收集菌体,用浓度50mm、ph7.5的tris-cl洗涤重悬菌体,将菌体密度od600nm调整到9。参照实施例11的方法使细胞透性化,获得透性化细胞。
上述用50mm、ph7.5的tris-cl重悬的透性化工程菌悬液中加入10mm丙酮酸、5mm甲醇、0.1mmnud,在30℃、200rpm的摇床中厌氧反应1h。取100μl加入900μl乙腈水混合液(体积比,乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统分析,结果表明反应液含有0.6mm甲醇、3.7mm乳酸和6.1mm丙酮酸。
在只添加甲醇和nud中的一种及不添加甲醇和nud的对照实验中,乳酸的浓度分别为0.4mm、0.6mm和0.4mm。
实施例12说明,在酿酒酵母全细胞催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态的nad类似物,被dldh-v152r用做辅酶应用于还原反应,乳酸的积累量与不添加甲醇和nud的对照实验相比提高了9.3倍,因此可作为一种通过提供氧化还原力调控微生物体内乳酸的代谢强度的方式。
实施例15:甲醇脱氢酶介导的里氏木霉(trichodermareesei)胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶同时在里氏木霉细胞中表达,形成一种依赖nad类似物的生物催化体系。当培养基中的甲醇和nad类似物进入宿主细胞后,启动该生物催化体系。
将编码bsmdh-y171r/i196v/v237t/n240e/k241a的基因和编码dldh-v152r的基因,由启动子pcbh1和终止子tcbh1控制,将上述两种表达盒整合到pcambia1300载体上获得工程质粒。
将上述工程质粒导入里氏木霉获得工程菌株t.reeseiwjt012,用ph4.8的含有15g/l乳糖、10g/l酵母提取物、1g/l(nh4)2so4、3g/l的kh2po4、0.5g/l的mgso4、0.6g/l的cacl2、0.05g/l的feso4·7h2o、0.0016g/l的mnso4·h2o、0.0014g/l的znso4·7h2o、0.0037g/l的cocl2·6h2o的培养基诱导工程菌t.reeseiwjt012表达上述两种功能蛋白,在25℃、200rpm的摇床中培养48h,2000×g离心6min收集菌体,用浓度50mm、ph7.5的tris-cl洗涤重悬菌体,将菌体密度调整到3g菌体干重/l。参照实施例11方法使细胞透性化,获得透性化细胞。
上述浓度50mm、ph7.5的tris-cl重悬的透性化工程菌悬液中加入10mm丙酮酸、5mm甲醇、0.1mm的ncd,在30℃、200rpm的摇床中厌氧反应2h。取100μl样品,加入900μl乙腈水混合液(体积比,乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统分析,结果表明反应液含1.5mm甲醇、3.3mm乳酸、6.4mm丙酮酸。
在只添加甲醇和ncd中的一种及不添加甲醇和ncd的对照实验中,乳酸浓度分别为1.2mm、0.9mm和0.6mm。
实施例13说明,在里氏木霉全细胞催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态的nad类似物,被dldh-v152r用作辅酶应用于还原反应,乳酸的积累量与不添加甲醇和ncd的对照实验相比提高了5.5倍,因此可作为一种通过提供氧化还原力调控微生物体内乳酸代谢强度的方式。
实施例16:甲醇脱氢酶介导的圆红冬胞酵母(rhodosporidiumtoruloides)胞内还原nad类似物及其应用
可以将所识别nad类似物的甲醇脱氢酶、偏好nad类似物的氧化还原酶同时在圆红冬胞酵母中表达,形成一种依赖nad类似物的生物催化体系。当培养基中的甲醇和nad类似物进入宿主细胞后,启动该生物催化体系。
将编码bsmdh-y171r/n240e的基因和编码dldh-v152r的基因,分别由启动子gpd、pgk和终止子hspt、tnos控制,将上述两种表达盒整合到pzpk载体上获得工程质粒。
将上述工程质粒通过atmt转化导入圆红冬胞酵母获得工程菌株r.toruloideswjt013,用ph6.0的含有20g/l葡萄糖、10g/l酵母提取物、20g/l蛋白胨的yepd培养基培养工程菌wjt013表达上述两种功能蛋白,在28℃、200rpm的摇床中培养48h,2000×g离心6min收集菌体,用浓度50mm、ph7.5的tris-cl洗涤重悬菌体,将菌体密度调整到3g菌体干重/l。参照实施例11方法使细胞透性化,获得透性化细胞。
上述浓度50mm、ph7.5的tris-cl重悬的透性化工程菌悬液中加入10mm丙酮酸、5mm甲醇、0.1mm的ntd,在30℃、200rpm的摇床中厌氧反应2h。取100μl样品,加入900μl乙腈水混合液(体积比,乙腈:水=4:1)终止反应。
参照实施例5的方法利用离子色谱系统分析,结果表明反应液含1.2mm甲醇、3.0mm乳酸、6.0mm丙酮酸。
在只添加甲醇和ntd中的一种及不添加甲醇和ntd的对照实验中,乳酸浓度分别为1.2mm、0.7mm和0.6mm。
实施例14说明,在圆红冬胞酵母催化过程中胞内甲醇脱氢酶可以通过氧化甲醇提供还原态的nad类似物,被dldh-v152r用作辅酶应用于还原反应,乳酸的积累量与不添加甲醇和ntd的对照实验相比提高了5倍,因此可作为一种通过提供氧化还原力调控微生物体内乳酸代谢强度的方式。
实施例17:甲醇脱氢酶突变体晶体解析
分别将甲醇脱氢酶突变体bsmdh-y171r/i196e/v237t/n240e/k241a和bsmdh-y171r/v237t/n240e/k241a与ncd进行晶体解析,以获得含有配体ncd的晶体结构。
纯化后的蛋白,采用座滴法,选取hamptonresearchandwizard公司的结晶筛选试剂盒进行筛选。用于衍射的晶体培养条件为0.1m硫酸铵,0.1mtris(ph7.0),10%聚乙二醇单乙醚5000,蛋白浓度6mg/ml,温度30℃。晶体浸母液中,添加5mmncd,然后进行低温处理、数据收集。所获得的晶体结构如图1或图2所示。
来源于的甲醇脱氢酶,ncbi蛋白数据库编号p42327.1,含1至339位完整氨基酸序列,如下:
mkaavvnefkkaleikeverpkleegevlvkieacgvchtdlhaahgdwpikpklplipghegvgivvevakgvksikvgdrvgipwlysacgeceycltgqetlcphqlnggysvdggyaeyckapadyvakipdnldpvevapilcagvttykalkvsgarpgewvaiygigglghialqyakamglnvvavdisdeksklakdlgadiainglkedpvkaihdqvggvhaaisvavnkkafeqayqsvkrggtlvvvglpnadlpipifdtvlngvsvkgsivgtrkdmqealdfaargkvrpivetaeleeinevfermekgkingrivlklked。
序列表
<110>中国科学院大连化学物理研究所
<120>一种利用甲醇还原nad类似物的方法
<160>1
<170>siposequencelisting1.0
<210>1
<211>339
<212>prt
<213>甲醇脱氢酶bsmdh(bacillusstearothermophilus)
<400>1
metlysalaalavalvalasngluphelyslysalaleugluilelys
151015
gluvalgluargprolysleuglugluglygluvalleuvallysile
202530
glualacysglyvalcyshisthraspleuhisalaalahisglyasp
354045
trpproilelysprolysleuproleuileproglyhisgluglyval
505560
glyilevalvalgluvalalalysglyvallysserilelysvalgly
65707580
aspargvalglyileprotrpleutyrseralacysglyglucysglu
859095
tyrcysleuthrglyglngluthrleucysprohisglnleuasngly
100105110
glytyrservalaspglyglytyralaglutyrcyslysalaproala
115120125
asptyrvalalalysileproaspasnleuaspprovalgluvalala
130135140
proileleucysalaglyvalthrthrtyrlysalaleulysvalser
145150155160
glyalaargproglyglutrpvalalailetyrglyileglyglyleu
165170175
glyhisilealaleuglntyralalysalametglyleuasnvalval
180185190
alavalaspileseraspglulysserlysleualalysaspleugly
195200205
alaaspilealaileasnglyleulysgluaspprovallysalaile
210215220
hisaspglnvalglyglyvalhisalaalaileservalalavalasn
225230235240
lyslysalaphegluglnalatyrglnservallysargglyglythr
245250255
leuvalvalvalglyleuproasnalaaspleuproileproilephe
260265270
aspthrvalleuasnglyvalservallysglyserilevalglythr
275280285
arglysaspmetglnglualaleuaspphealaalaargglylysval
290295300
argproilevalgluthralagluleuglugluileasngluvalphe
305310315320
gluargmetglulysglylysileasnglyargilevalleulysleu
325330335
lysgluasp
- 还没有人留言评论。精彩留言会获得点赞!