CLE25基因在调控植物韧皮部起始及发育过程中的应用的制作方法
本发明属于农业生物技术领域,具体涉及cle25基因在调控植物韧皮部起始及发育过程中的应用。
背景技术:
植物的维管组织主要由韧皮部、原形成层和木质部组成。韧皮部主要负责光合产物,如糖类、激素及其他一些信号分子,木质部主要负责水分和无机盐等的运输。原形成层可分化为韧皮部和木质部。韧皮部位于维管系统内,是整个植物体内营养物质和信号分子传递的关键。尽管现有技术报道了韧皮部分化的形态学过程和几种调控因子,但其发生的分子机制仍不清楚。
小分子多肽激素通常作为信号分子发挥作用,其编码基因一般翻译成前体蛋白,经过剪切后的加工修饰,最后形成成熟的多肽。成熟小分子多肽通常由几个到几十个氨基酸组成。小分子多肽一般分泌到细胞外发挥作用,通常可以移动几个细胞层的距离,与定位在细胞质膜的受体类激酶结合,激活下游信号通路。小分子多肽激素也可以通过维管束进行长距离运输。
现有技术对小肽编码基因clavata3/embryosurroundingregion25(cle25)功能的研究如下:cle25基因在维管组织、根维管束的韧皮部区域表达,且在根中的表达受干旱诱导;拟南芥cle25多肽激素通过维管组织从根到叶片转运缺水信号,通过受体bams影响叶片aba合成和气孔开闭;假激酶crn及lrr受体蛋白clv2的突变体对cle25p体外处理不敏感,lrr受体激酶clerk的突变体对cle25p体外处理不敏感。
技术实现要素:
本发明的目的在于提供cle25基因的应用。
根据本发明的具体实施方式,cle25基因在调控植物的韧皮部起始及发育过程的应用,其中,cle25基因的基因组序列如seqidno.1所示:
attctttctttatttctaatttaatttactttggtgcataacacttcagtagagcatgtaacttgacacctctagagtgcaatactggcatggctttgctcacacaatcactgtgagaatcaatgctataaggacactatcctgaccatgtgtaagattctaaatgatataggagaaactgatatatagtaatgaaattgtggggccagatctgttcaaactttggctttactatgttagaaggcaagcttttctttcccaccaaggccaaaaggaagtttgcttgtgtcatggaaacaaactggctcaatcttccctagctgtcaatcagatatagattctttttttttcacagtttagatatgtttcttaatgtttttgctttagtttatcgtttaaaggttatgaagaacaagcatacatttcttgctgctaattggcttttggtagggagggaatacttaacaaggaagaaaaattgtaggctctagctccttttcacaggtttggttatcttacttgtgtattgttacattcttaattgattttgttattgtttaaaggaaacatccaagtgatttaagatagtggtaaggaaagtagtaggcttgctaatacactcagtcacaaaaggtacactgtcctctgcttttaaaatggcttaaaatatatgtagttaagggagcttgtgaggtccccaatggtttgctgcatgaaaattcacaaatcaaagcaaagaaaaaggtactcaacgggcccatatgttacataaagatatgttttttcttaactttcacttgctcttgcctcctctctctctctctacgtggacacctacgtatgtatattgttgctttgtgttgcttcatctttaaatagggccaatcacggatttgtcttccaaggtatataaaaacaacctaaatgcttgttttttgcttcccatttcgctttccccttttttgagcctcttctgtccaaagatatctctctctatttatgtgacagtcacttcaccaacatcatggatgttctgctcagtttattcttgggtttggttggtcagttgtttatgttaaacaggaagctgtagggacataggtttcagtatgggtggaaatggcattagagctttggttggagtgattgcatctttgggtttgattgtgtttcttcttgtcggtatcttagcaaactctgcaccaagtgttccatcatcagaaaatgtcaagactttgcggtttagtggtaaggatgtgaatctgtttcatgtaagcaagcgaaaagttcctaatggacctgatcctatccacaacaggttcttatcccttctctctcgaattttcaatcttttacttttgcttctttgagtttgctaccagagatcaagcgttttgagaactttgtcatgaagtttgtttaaagttcgtttatacaaaaactaatcatccaatattagtgtttcctctgttcaagtttttttgttgtcactgtatacacagataccgataatcactggtttctaaaaagattattagcatgcagaattcttgttctttccttcttttcagtgtcactatacattttgtgtacattaaagagctcgtcttttgaaagcagacaatgttatcatgttttagtaaaagcttcttctttatctaattttaaagccttagagaagaattaaaggcacttgtgaagaactttgaccactttagttaaagtttattactgcagaaagaactctactttaataagcttttgtttcaaatctatttcttcaggaaagcagaaacttcgagacggccaccaagagtatgatattgactgtaatgtttatgtcatgcgggcttcgttcattttcgggtttttctaccgtctcaagacacagtgaagtattcaacctctcaattacgtttccacgtcaagaagctactattttttctgaggcaatcattaacttcagatgttagagatatgctatgaactcatagctttgtgaagtcttgctctgtagaattttggaaggtgaagtctctgctctctctttcgttttggaacattgtgtctaaagattgaaaatcattcagaagattgttgctaagatattaaagacttcttaagagtttttgtaataacatttgtatttagttattataggcttatgtctttatctaactctctgtttcctgttttcctcatctttctttatttgccttttgttttagaatctgttaactttaaagtttgcttttttatgtttcctctggctaaagcaaaaaaccattaaactaaagttgctccattttcaaaaatcattctcccactccaatgatatttgcatcttaatacaaggcttgtccttgcacaactttagaatcacatacattttcatttttgggttcttgtgaactttgggaatcttatattctctgtgtaattttgatagcatcttacaaagtgaaattcaaaaggagtgagtgaccatagatgctaaaaacttgtctttctctttctttcttgccttctttagcttcttttctgggattgattttgactgtatcgagaagcagacccctaaagtaagattctgcagagacatcttctttatc
cle25基因的cdna序列如seqidno.2所示:
atgggtggaaatggcattagagctttggttggagtgattgcatctttgggtttgattgtgtttcttcttgtcggtatcttagcaaactctgcaccaagtgttccatcatcagaaaatgtcaagactttgcggtttagtggtaaggatgtgaatctgtttcatgtaagcaagcgaaaagttcctaatggacctgatcctatccacaacaggaaagcagaaacttcgagacggccaccaagagtatg
cle25多肽激素的氨基酸序列如seqidno.3所示:
mggngiralvgviaslglivfllvgilansapsvpssenvktlrfsgkdvnlfhvskrkvpngpdpihnrkaetsrrpprv
其中,rkvpngpdpihn为cle25多肽激素的结构域。
根据本发明的具体实施方式,cle25基因在植物的根干细胞区域表达以发挥调控作用。
根据本发明的具体实施方式,cle25基因编码的多肽激素通过与clerk受体激酶和/或clv2受体激酶结合发挥调控作用。
根据本发明的具体实施方式,cle25基因作为marker指示植物胚胎发育过程中韧皮部发育的应用。
根据本发明的具体实施方式,cle25基因在调控植物光合产物运输中的应用。
本发明的有益效果:
本发明将cle25基因敲除导致原韧皮部发育滞后,在cle25g6t植株中,拮抗性cle25g6t肽的原位表达损害了根中韧皮部的连续性,茎中韧皮部的形成也受到影响。遗传和生化分析表明,由富含亮氨酸重复序列(lrr)受体激酶(clerk)和clv2lrr受体蛋白组成的复合体参与了cle25多肽激素信号转导过程。
本发明发现cle25基因在胚胎发育的64细胞期即开始在原维管束起始细胞表达,并在胚胎后根发育过程中继续在筛管元件原形成层干细胞和韧皮部细胞系中表达,促进拟南芥韧皮部的起始。cle25是目前发现在胚胎发育最早期表达的韧皮部相关基因,可用于调控根的韧皮部起始过程及连续性发育,并可作为marker指示胚胎发育过程中的韧皮部发育。
附图说明
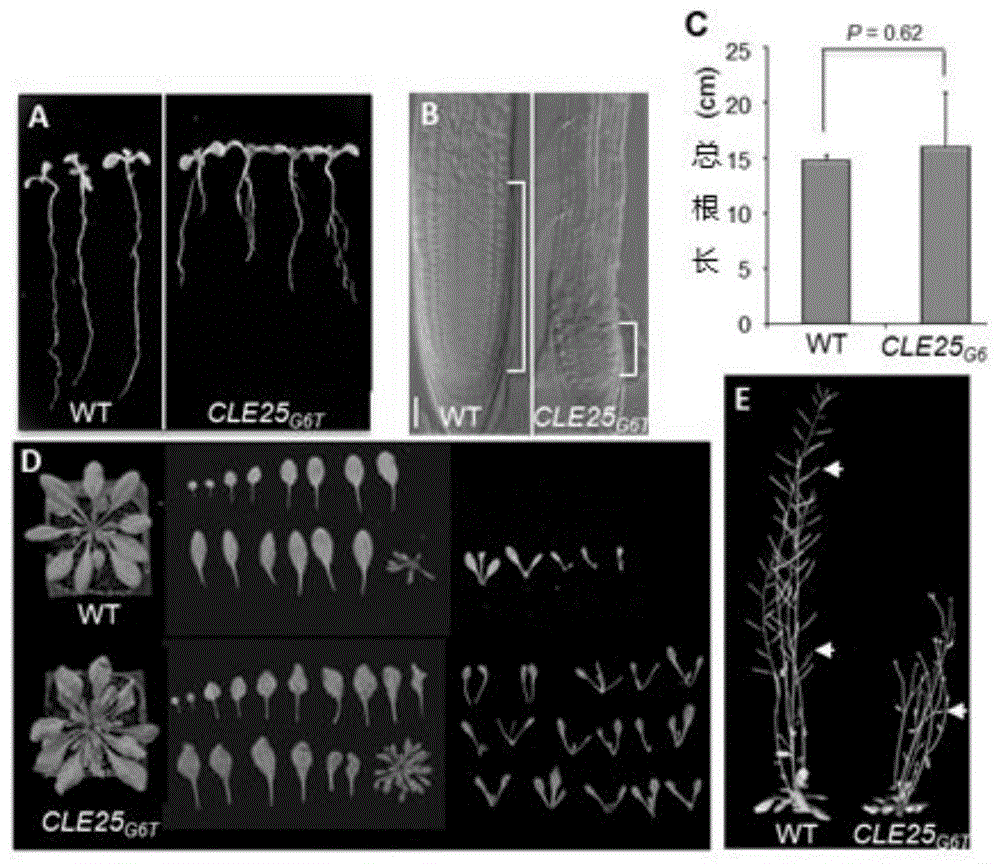
图1显示转基因cle25g6t植物的表型;其中,a显示cle25g6t植物与野生型的幼苗根生长情况;b显示cle25g6t植物与野生型的根分生区生长情况;c显示cle25g6t植物与野生型的根总长;d显示cle25g6t植物与野生型的叶腋数量对比情况;e显示cle25g6t植物也野生型的结实率对比情况;
图2显示转基因cle25g6t植物的韧皮部运输功能受阻,其中,a显示cle25g6t植物莲座叶光合作用产物不能有效运出;b:显示cle25g6t植物花序与野生型的蔗糖含量情况;c显示cfda染料在cle25g6t植物韧皮部与野生型韧皮部的运输情况;
图3显示细胞水平转基因cle25g6t植物的韧皮部发育异常,其中,a显示cle25g6t植物与野生型的根分生区韧皮部发育对比情况;b显示cle25g6t植物与野生型的根成熟区韧皮部发育对比情况;c显示cle25g6t植物与野生型的茎韧皮部元件对比情况;d显示cle25g6t植物与野生型的茎韧皮部元件对比情况;
图4显示转基因cle25g6t植物的韧皮部发育中断,相关基因表达受抑制;其中,a显示韧皮部markerpsuc2:gfpspor在cle25g6t植物根尖区的表达明显滞后于野生型;b显示cle25g6t植物根韧皮部有不表达markerpsuc2:gfpspor的细胞,称为gap细胞;c显示cle25g6t植物根缺少韧皮部-原形成层起始细胞的平周分裂;d显示韧皮部markerpcvp2:nls-venus在cle25g6t植物根尖区的表达明显弱于野生型;
图5显示cle25-1突变体的韧皮部分化滞后;其中,a表示cle25-1的突变形式;b显示原生韧皮部细胞层,从紧邻静止中心的细胞开始,每个细胞距静止中心距离的总长度,cle25-1要明显短于野生型;
图6显示cle25在胚胎和根尖的表达模式;其中,a、b显示cle25在胚胎64细胞时期和心形期的维管束起始细胞表达;c显示cle25主要在根尖的原生韧皮部和后生韧皮部表达,在中柱鞘和原形成层也有较弱表达;d显示根尖横切显示cle25在韧皮部及紧邻韧皮部的中柱鞘和原形成层细胞表达;
图7显示不同启动子驱动cle25g6t转基因植物的短根统计结果;其中,a显示部分维管束相关基因的不同表达部位模式图;b显示不同维管束相关基因的启动子驱动cle25g6t转基因植物的根长统计;
图8显示clv2-6和clerk-5突变基因模式图及clerk在根尖的表达模式;其中,a为clv2-6和clerk-5的蛋白模式图;b显示clerk在根尖的原生韧皮部、后生韧皮部的表达情况;
图9显示clerk和clv2在生化和遗传水平的相互作用。
具体实施方式
实施例1
利用拮抗多肽技术构建pcle25:cle25g6t:cle25ter转基因植物。
设计引物如下:
cle25sr:ggggaccactttgtacaagaaagctgggtatatggactcggtg,
cle25sf:ggggacaagtttgtacaaaaaagcaggctactgatagaaggtt;
cle25mtf:tcctaatacacctgatcctatccacaac,
cle25mtr:atcaggtgtattaggaacttttcgcttg。
以拟南芥col-0野生型植物基因组dna为模板,利用引物对cle25sr与cle25mtf及cle25sf与cle25mtr进行扩增,再以所得pcr产物作为模板,利用引物对cle25sr与cle25sf进行二次扩增,所得产物即为cle25g6t的基因全长;将cle25g6t基因全长克隆到pbgwfs7双元载体,并转化入农杆菌gv3101;利用花序侵染的方法进行植物转基因,最终得到pcle25:cle25g6t:cle25ter转基因植物。
cle25g6t为cle25基因功能缺失的植物材料,对cle25g6t植物的表型进行分析。如图1所示,cle25g6t植物的根长变短、分生区变短,但总的根长(主根和侧根)没有受到影响;地上器官如莲座叶和叶腋增多,但结实率大大降低。
在16hr光周期结束和8hr暗周期结束时,利用lugol染色法分别对野生型和cle25g6t植物莲座叶进行染色分析。
结果如图2所示,在cle25基因功能缺失的植物材料中,叶片的光合产物不能有效转运出去。通过检测花序组织的糖含量发现,花序的蔗糖含量降低。利用一种只能在韧皮部运输的荧光分子cfda对cle25g6t植物韧皮部的运输功能进行检测,将cfda溶液滴在子叶的背面,40分钟后在荧光显微镜下观察根尖是否有cfda荧光信号,结果发现:cle25g6t植物根尖检测不到cfda荧光信号,说明其韧皮部运输受到影响。
通过半薄切片技术进行细胞学水平观察。结果如图3所示,cle25g6t植物根和茎的维管束发育均出现异常,根中韧皮部的筛管原件不能有效形成,茎中韧皮部筛管元件减少,组织混乱,同时形成层区域增加。
将一些分子标记通过遗传杂交的方式导入cle25g6t植物。结果如图4所示,cle25g6t植物根韧皮部发育的连续性被破坏,可以看到有不表达韧皮部marker基因的gap细胞;韧皮部的起始分化与野生型的两次平周分裂相比,缺失一次平周分裂,导致最终缺少一层韧皮部细胞;一些调控韧皮部发育的基因的表达在cle25g6t植物中均受到抑制。
利用crispr技术创建cle25-1突变体。
设计带有接头的靶向rna引物(上游引物:gtcagcaagcgaaaagttcctaa;下游引物:aaacttaggaacttttcgcttgc)。将该靶向序列克隆到pyl-u3-grna载体上。经过两轮pcr。
第一轮pcr引物:
u-f:5-ctccgttttacctgtggaatcg-3;
grna-r:5-cggaggaaaattccatccac-3;
第二轮pcr引物:
uctcg-b1’:ttcagaggtctctctcgcactggaatcggcagcaaagg-3;
grctga-b2:agcgtgggtctcgtcagggtccatccactccaagctc-3。
将第二轮pcr产物回收后连接到带有cas9的双元载体pylcrispr/cas9-mtmono中并进行测序鉴定。最后,将构建好的带有靶向rna序列的pylcrispr/cas9-mtmono构建转化进农杆菌gv3101中,随后通过农杆菌介导的遗传转化将上述构建导入col-0野生型拟南芥基因组中。
如图5所示,该突变体表现出韧皮部发育滞后的表型,原生韧皮部细胞层从紧邻静止中心的细胞开始,每个细胞距静止中心距离的总长度,cle25-1要明显短于野生型,验证了cle25g6t植物是一个cle25基因功能缺失的材料。
利用cle25启动子融合gus或gfp,通过透明或半薄切片或共聚焦显微镜观察cle25的表达情况。
结果如图6所示,cle25在胚胎发育的64细胞期即开始在原维管束起始细胞表达,且其维管束表达的模式一直延续到胚胎后发育阶段。在根中,cle25在紧挨静止中心的筛管元件-原形成层干细胞及后面分化的原生韧皮部、后生韧皮部及紧挨韧皮部的中柱鞘和原形成层细胞表达。因此,cle25在胚胎发育的64细胞期即开始表达,是目前发现最早表达的韧皮部发育调控基因。
利用arr5和tmo5等基因启动子,进行启动子置换,即利用维管束不同细胞类型特异表达基因的启动子驱动cle25g6t,并转化野生型植物。
结果如图7所示,类似cle25启动子驱动的cle25g6t植物,arr5和tmo5启动子驱动的cle25g6t植物也出现短根表型。通过比较这些基因的表达模式发现,arr5和tmo5启动子所驱动cle25g6t均在根干细胞区域表达,说明cle25g6t在根干细胞区域表达对其行使功能是必须的,而在哪个细胞层表达并不重要。
实施例2
通过筛选ems诱变群体及t-dna插入的lrr受体激酶突变群体,鉴定到cle25的类受体蛋白clv2及受体激酶clerk。具体操作如下:
通过ems诱变的方法建立了一个包括8000粒cle25g6t种子的诱变群体t1代,t1代单株收种得到t2代,每个t2代种8棵,8棵植物的种子混收得到t3代种子。对t3代种子铺1/2ms平板,从中筛选根长比cle25g6t长的回复突变。结果,从该ems诱变群体中筛选到一个根长回复的突变体,由于该突变体表现出类似clv系列突变体的多心皮表型,因此对该突变体基因组的clv1、clv2及clv3分别进行pcr扩增和测序分析,结果显示clv2第456位的苏氨酸突变为丙氨酸,是clv2的一个新突变类型,命名为clv2-6,clv2编码一个类受体蛋白。利用cle25p体外处理拟南芥223个lrr受体激酶的t-dna插入突变体,发现clerk-5突变体对cle25p的处理不敏感,进而鉴定到cle25p的受体clerk受体激酶。
以col-0野生型基因组dna为模板,通过pcr将clerk基因启动子扩增出来,克隆到带有报告基因gus的双元载体,并转化col-0野生型拟南芥。通过抗性筛选的方法得到阳性转基因植物,利用gus染色的方法对clerk的组织表达模式进行分析发现,clerk在根尖的原生韧皮部、后生韧皮部有较强表达,其在胚胎和根尖的组织表达模式与cle25非常相似。
将clerk和clv2分别克隆到bifc载体,然后转化农杆菌eha105,利用注射器注射入烟草叶片,24-48小时后在共聚焦显微镜下观察。
结果如图8所示,可以观察到yfp荧光信号,说明clerk和clv2之间存在相互作用。
将上述烟草叶片收集起来,提取总蛋白,进行免疫共沉淀实验,然后利用western进行检测。
结果如图9所示,clv2可以被clerk免疫共沉淀出来,图9中bifc(a)和co-ip(b)结果显示clerk和clv2之间存在相互作用,clerk也能和自身发生相互作用,而clv2不能和自身相互作用;图9中的c显示,clerk-5不能回复brx-2的短根表型,而clv2-101能够部分回复brx-2的短根表型。
鉴于ops也是参与调控原生韧皮部发育的关键基因,利用遗传杂交的手段,即将clerk-5与clv2-101的花分别去雄,用ops-1突变体的花粉分别对clerk-5和clv2-101进行授粉,所结种子为f1代,f1代自交后得到f2代种子。在f3代进行双突变体clerk-5ops-1和clv2-101ops-1的鉴定,对鉴定到的纯合双突变体进行表型分析。
如图9中d所示,clerk-5ops-1和clv2-101ops-1的根均较ops-1长,说明clerk-5和clv2-101均能部分回复ops-1的短根表型,对照为bam3-2能够完全回复brx-2和ops-1的短根表型,从遗传上证明clerk和clv2在同一途径参与cle25多肽激素信号转导过程。
序列表
<110>中国科学院植物研究所
<120>cle25基因在调控植物韧皮部起始及发育过程中的应用
<160>3
<170>siposequencelisting1.0
<210>1
<211>2612
<212>dna
<213>拟南芥(arabidopsisthaliana)
<400>1
attctttctttatttctaatttaatttactttggtgcataacacttcagtagagcatgta60
acttgacacctctagagtgcaatactggcatggctttgctcacacaatcactgtgagaat120
caatgctataaggacactatcctgaccatgtgtaagattctaaatgatataggagaaact180
gatatatagtaatgaaattgtggggccagatctgttcaaactttggctttactatgttag240
aaggcaagcttttctttcccaccaaggccaaaaggaagtttgcttgtgtcatggaaacaa300
actggctcaatcttccctagctgtcaatcagatatagattctttttttttcacagtttag360
atatgtttcttaatgtttttgctttagtttatcgtttaaaggttatgaagaacaagcata420
catttcttgctgctaattggcttttggtagggagggaatacttaacaaggaagaaaaatt480
gtaggctctagctccttttcacaggtttggttatcttacttgtgtattgttacattctta540
attgattttgttattgtttaaaggaaacatccaagtgatttaagatagtggtaaggaaag600
tagtaggcttgctaatacactcagtcacaaaaggtacactgtcctctgcttttaaaatgg660
cttaaaatatatgtagttaagggagcttgtgaggtccccaatggtttgctgcatgaaaat720
tcacaaatcaaagcaaagaaaaaggtactcaacgggcccatatgttacataaagatatgt780
tttttcttaactttcacttgctcttgcctcctctctctctctctacgtggacacctacgt840
atgtatattgttgctttgtgttgcttcatctttaaatagggccaatcacggatttgtctt900
ccaaggtatataaaaacaacctaaatgcttgttttttgcttcccatttcgctttcccctt960
ttttgagcctcttctgtccaaagatatctctctctatttatgtgacagtcacttcaccaa1020
catcatggatgttctgctcagtttattcttgggtttggttggtcagttgtttatgttaaa1080
caggaagctgtagggacataggtttcagtatgggtggaaatggcattagagctttggttg1140
gagtgattgcatctttgggtttgattgtgtttcttcttgtcggtatcttagcaaactctg1200
caccaagtgttccatcatcagaaaatgtcaagactttgcggtttagtggtaaggatgtga1260
atctgtttcatgtaagcaagcgaaaagttcctaatggacctgatcctatccacaacaggt1320
tcttatcccttctctctcgaattttcaatcttttacttttgcttctttgagtttgctacc1380
agagatcaagcgttttgagaactttgtcatgaagtttgtttaaagttcgtttatacaaaa1440
actaatcatccaatattagtgtttcctctgttcaagtttttttgttgtcactgtatacac1500
agataccgataatcactggtttctaaaaagattattagcatgcagaattcttgttctttc1560
cttcttttcagtgtcactatacattttgtgtacattaaagagctcgtcttttgaaagcag1620
acaatgttatcatgttttagtaaaagcttcttctttatctaattttaaagccttagagaa1680
gaattaaaggcacttgtgaagaactttgaccactttagttaaagtttattactgcagaaa1740
gaactctactttaataagcttttgtttcaaatctatttcttcaggaaagcagaaacttcg1800
agacggccaccaagagtatgatattgactgtaatgtttatgtcatgcgggcttcgttcat1860
tttcgggtttttctaccgtctcaagacacagtgaagtattcaacctctcaattacgtttc1920
cacgtcaagaagctactattttttctgaggcaatcattaacttcagatgttagagatatg1980
ctatgaactcatagctttgtgaagtcttgctctgtagaattttggaaggtgaagtctctg2040
ctctctctttcgttttggaacattgtgtctaaagattgaaaatcattcagaagattgttg2100
ctaagatattaaagacttcttaagagtttttgtaataacatttgtatttagttattatag2160
gcttatgtctttatctaactctctgtttcctgttttcctcatctttctttatttgccttt2220
tgttttagaatctgttaactttaaagtttgcttttttatgtttcctctggctaaagcaaa2280
aaaccattaaactaaagttgctccattttcaaaaatcattctcccactccaatgatattt2340
gcatcttaatacaaggcttgtccttgcacaactttagaatcacatacattttcatttttg2400
ggttcttgtgaactttgggaatcttatattctctgtgtaattttgatagcatcttacaaa2460
gtgaaattcaaaaggagtgagtgaccatagatgctaaaaacttgtctttctctttctttc2520
ttgccttctttagcttcttttctgggattgattttgactgtatcgagaagcagaccccta2580
aagtaagattctgcagagacatcttctttatc2612
<210>2
<211>245
<212>dna
<213>拟南芥(arabidopsisthaliana)
<400>2
atgggtggaaatggcattagagctttggttggagtgattgcatctttgggtttgattgtg60
tttcttcttgtcggtatcttagcaaactctgcaccaagtgttccatcatcagaaaatgtc120
aagactttgcggtttagtggtaaggatgtgaatctgtttcatgtaagcaagcgaaaagtt180
cctaatggacctgatcctatccacaacaggaaagcagaaacttcgagacggccaccaaga240
gtatg245
<210>3
<211>81
<212>prt
<213>拟南芥(arabidopsisthaliana)
<400>3
metglyglyasnglyileargalaleuvalglyvalilealaserleu
151015
glyleuilevalpheleuleuvalglyileleualaasnseralapro
202530
servalprosersergluasnvallysthrleuargpheserglylys
354045
aspvalasnleuphehisvalserlysarglysvalproasnglypro
505560
aspproilehisasnarglysalagluthrserargargproproarg
65707580
val
- 还没有人留言评论。精彩留言会获得点赞!