团头鲂耐低氧关联SNP位点的单倍型在育种中的应用的制作方法
本发明涉及育种领域,更具体地,涉及团头鲂耐低氧关联snp位点的单倍型在育种中的应用。
背景技术:
团头鲂(megalobramaamblycephala),隶属于鲂属、鲤科、鲤形目,俗称武昌鱼,系草食性经济鱼类,具有肉质鲜美、含肉率高、食性广、生长快、抗病力强等特点,是中国主要的淡水养殖对象之一,2018年总产量约83万t。然而,团头鲂不耐低氧,易浮头,养殖生产的进一步发展迫切需要耐低氧的良种。
技术实现要素:
本发明的目的是提供团头鲂耐低氧关联snp位点的单倍型在育种中的应用,可以用于团头鲂耐低氧新品系分子辅助育种。
本发明提供了一种团头鲂耐低氧关联snp位点的单倍型在育种中的应用。
在上述团头鲂耐低氧关联snp位点的单倍型在育种中的应用中,其中,团头鲂耐低氧关联snp位点的单倍型为基因egln2的t397t715。
在本申请中,双倍ⅱ型个体表现出较高的耐低氧能力,单倍ⅱ型可用于团头鲂耐低氧新品系分子辅助育种。
附图说明
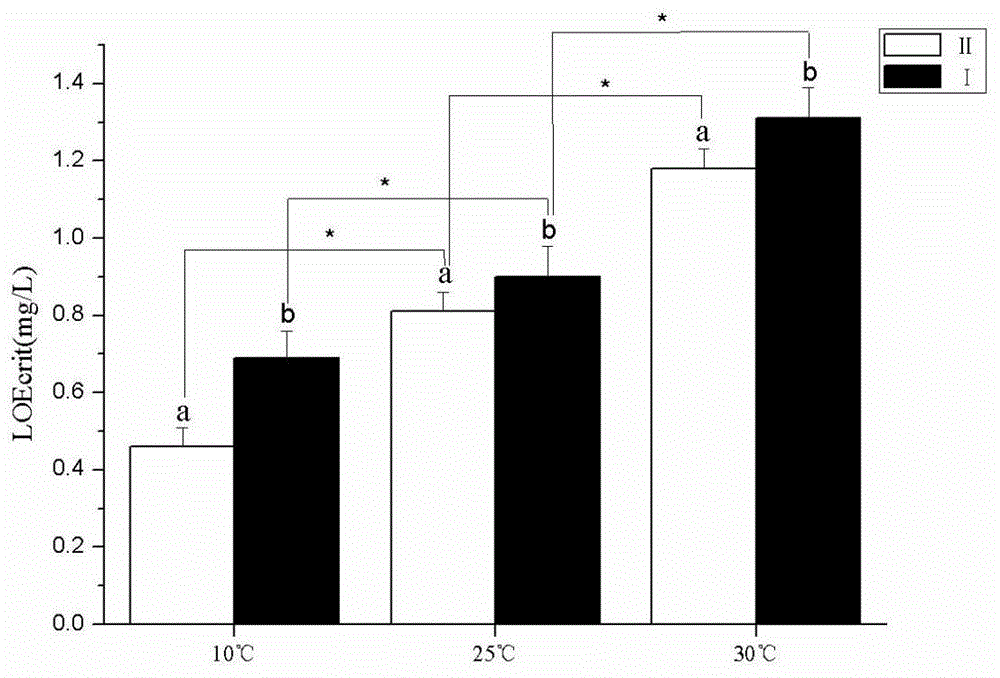
图1示出了双倍ⅰ型和ⅱ型个体的体型失衡的关键溶氧值,其中,不同字母显示在10℃,25℃,30℃的差异有统计学意义(p<0.01)。*p<0.01。ⅰ:双倍ⅰ型个体;ⅱ:双倍ⅱ型个体。
图2为双倍ⅰ型(a–d)和ⅱ型(e–h)个体鳃小片的光学显微结构,其中,t=10±1.0℃,do=2.0mg·l-1。双倍ⅰ型个体(a-d),双倍ⅱ型个体(e-h);常氧(a,e),低氧4天(b,f),低氧7天(c,g),常氧恢复7天(d,h)。标尺=50μm。
图3示出了双倍ⅰ型和ⅱ型个体鳃小片表面积(a)和ilcm体积(b)的变化,其中,t=10±1.0℃,do=2.0mg·l-1。0d-h:常氧,4d-h:低氧4天,7d-h:低氧7天。不同字母显示差异有统计学意义(p<0.01)。*p<0.01。ⅰ:双倍ⅰ型个体。ⅱ:双倍ⅱ型个体。
图4示出了双倍ⅰ型和ⅱ型个体红细胞的数目(a)和血红蛋白浓度(b)的变化,其中,t=10℃,do=2.0mg·l-1。不同字母显示差异有统计学意义(p<0.01)。*p<0.01。0d-h:常氧,4d-h:低氧4天,7d-h:低氧7天。ⅰ:双倍ⅰ型个体,ⅱ:单倍ⅱ型个体。
具体实施方式
下面的实施例可以使本领域技术人员更全面地理解本发明,但不以任何方式限制本发明。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的试验材料,如无特殊说明,均为常规生化试剂公司购买所得。
脯氨酸羟化酶(prolylhydroxylase,phd)作为低氧调控过程中重要的氧感受器,有三种亚基,分别为phdi,phd2和phd3,其中,phd1基因又名egln2(egglayingdeficientnine-likeprotein2),它们都属于依赖氧、α-酮戊二酸和fe2+催化的非血红素、铁依赖性加双氧化酶家族,而且都可以催化hif1a的羟基化反应,从而调控着血管的发生、红细胞的生成等生命过程。目前,在人类、鼠等哺乳类以及其他动物中都有相关的研究报道。有研究学者报道称egln1基因突变与人类适应高原低氧环境进化有重要的关系。egln基因在鱼类低氧应答hif信号通路中,通过修饰泛素化后的hif1a使其降解而调节体内氧平衡。
本实验利用已得到的团头鲂耐低氧转录组信息,从epo、hif1α、hif2α、vhi、hif1an、vegfaa、egln1a、egln1b、egln2、egln3等10个与耐低氧性状密切相关的基因中发现egln2基因的cdna上存在2个snp位点。通过将团头鲂egln2基因的2个snp位点与团头鲂耐低氧性状进行关联分析,发现该2个snp位点与耐低氧性状显著相关。进一步对2种双倍型个体进行低氧胁迫处理,发现团头鲂血红细胞数和血红蛋白(hb)浓度、鳃小片的伸出长度、鳃小片呼吸面积、鳃小片层间基质(ilcm)的厚度和体积在耐低氧基因型个体中存在显著差异,可作为耐低氧性状相关的分子标记,以加快团头鲂耐低氧新品系的选育进程。
1材料与方法
1.1实验材料
实验材料为团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”,体质量(40~50)g,均购自上海海洋大学农业部团头鲂遗传育种中心。暂养在室内塑料水族箱中,水温25±1.0℃,溶解氧7.0±0.5mg·l-1,ph7.5±0.3,使用经曝气的自来水,每日吸污换水1/3,每日投喂2次(8:00,17:00),保持自然光照。
1.2酶活力测定及基因组dna的提取
t=25±1.0℃,do=2.0mg·l-1条件下,对团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各100尾,进行4h的急性低氧胁迫,取每条鱼的鳃和鳍条,一一对应。鳃保存于-80℃冰箱,鳍条保存于无水乙醇中。
鳃组织按重量(g):体积(ml)=1:9的比例加入9倍体积的0.86%的生理盐水于离心管中,制备匀浆液,采用南京建成生物公司的检测试剂盒测定团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”鳃组织中过氧化氢酶(cat)、超氧化物歧化酶(sod)及na+/k+-atp酶的活力。
基因组dna的提取使用天根海洋动物dna提取试剂盒,分别用1.5%的琼脂糖凝胶电泳和分光光度计检测基因组dna的质量和浓度,然后将样品置于–20℃冰箱中备用。
1.3egln2基因snp位点筛选
利用已得到的团头鲂耐低氧转录组信息,选取epo、hif1α、hif2α、vhi、hif1an、vegfaa、egln1a、egln1b、egln2、egln3这10个与耐低氧性状相关的基因,将编码序列与团头鲂基因组序列进行对比,得到基因的内含子和外显子。然后利用primerpremier软件在基因的内含子区域设计引物,扩增外显子区域。实验发现基因egln2的397位和715位存在差异性snp位点,扩大样本量进行实验。设计引物序列及扩增长度见表1,覆盖基因全部编码区域。
表1
利用设计的6对引物,分别用团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各20个样品进行pcr扩增。pcr反应体系共20μl∶10μl2×taqpcrmastermix,模板dna(50ng·μl-1)1μl,上下游引物(10μmol·l-1)各1μl,ddh2o为7.0μl。扩增程序:94℃预变性3min;94℃变性30s,55℃退火30s,72℃延伸45s,共35个循环;72℃延伸7min。扩增产物用1.5%的琼脂糖凝胶电泳检测,检测合格的样品由上海迈浦生物科技有限公司负责测序,用sequencher5对测序结果进行比对寻找snp位点。然后对存在snp位点的引物用团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各100尾进行扩增验证。本申请所用试剂均购于天根生化科技有限公司,引物合成由上海迈浦生物科技有限公司完成。
1.4体型失衡的关键溶解氧值(loecrit)测定
为了对筛选出的snp位点和构建的优势双倍型进行验证,利用双倍ⅰ型和ⅱ型个体各15尾(40~50g),分别放养在10℃(do=10±0.5mg·l-1),25℃(do=8.5±0.5mg·l-1)和30℃(do=7.5±0.5mg·l-1)的20l玻璃缸中适应12h。在玻璃缸水面下5cm处放置一块同水面大小的纱网,防止团头鲂上浮呼吸。向玻璃缸中同时充氮气和空气,使用溶氧仪监测水体中溶解氧含量,在1h内,将溶解氧含量从常氧(≥10mg·l-1)降到5mg·l-1,维持1h;然后在30min内,将溶解氧含量从5mg·l-1降到2.5mg·l-1,维持30min;然后在30min内,将溶解氧含量从2.5mg·l-1降到1mg·l-1,维持30min;然后在30min内,将溶解氧含量从1mg·l-1降到0.5mg·l-1,维持30min。直至溶解氧降到0mg·l-1。同一温度的实验重复进行3次。当实验鱼出现侧翻失去平衡时,记下此时的溶解氧值和所用的时间。loecrit值计算按照brett’s的方法:
式(1)中:[o2]2i表示低氧处理倒数第二溶解氧含量(mg·l-1);[o2]2ii表示每一次溶解氧降低的数量(mg·l-1);ti表示实验鱼失去平衡所用时间;tii表示每一个稳定阶段的时间0.5h。
1.5低氧胁迫
将双倍ⅰ型和ⅱ型个体各20尾(40~50g),放在20l液浴恒温循环槽中(t=10℃),充入氮气和空气,用溶氧仪监测,将溶解氧含量保持在2.0mg·l-1以下,进行7d的低氧胁迫。
1.6组织切片
在低氧胁迫0d、4d、7d和恢复7d时,双倍ⅰ型和ⅱ型个体各取5尾,用0.5g·l-1的ms-222麻醉后活体冰上解剖,从右侧第二鳃弓上取下鳃片作为光学显微镜观察样本,在0.75%的生理盐水中反复润洗,以清除取样时粘附在鳃丝上的血细胞和粘液,放入boiun氏液中固定24h,常规脱水,二甲苯透明,石蜡包埋,连续切片(厚度5μm),经h.e染色后在显微镜下观察并拍照。
1.7形态学测量
为了评估双倍ⅰ型和ⅱ型个体鳃小片呼吸面积和层间基质(ilcm:interlamellarcellmass)体积的变化,进行以下形态测量:(1)伸出鳃小片基部的长度(lm);(2)鳃小片厚度(lm);(3)伸出鳃小片的长度(lm)。每个参数在同一低氧处理时间进行30次测量。鳃小片面积以近似为半椭圆计算。
公式为:
a=pl(2)
v=dhil(5)
式(2)中,a是鳃小片呼吸面积,l代表鳃小片基部的长度,p是椭圆的周长。
式(3)中,h为鳃小片伸出长度。
式(4)中,t是鳃小片厚度。
式(5)中,d是相邻两鳃小片间距离,hi是鳃小片层间基质厚度。
1.8血液分析
在低氧处理时,记录失去平衡个体的体质量和长度,用肝素冲洗过的1ml塑料注射器从鱼尾静脉抽取血样。血红蛋白[hb]浓度、红细胞(rbc)计数按标准方法测定。
1.9数据分析
数据均用平均数±标准差表示,采用双向方差分析(anova)(tukey’spair-wisemethod),p<0.05为存在显著性差异,p<0.01为存在极显著性差异。采用popgene软件分析遗传参数。利用jmp8.0软件的卡方检验来分析egln2基因片段的不同snp位点的基因型与团头鲂耐低氧性状之间的相关性。用shesis在线分析进行连锁不平衡分析和单倍型构建。
2结果
2.1egln2基因的序列扩增
利用已得到的团头鲂耐低氧转录组信息,从10个耐低氧性状相关的基因中筛选出差异性snp位点,结果发现在基因egln2的397位和715位存在差异性snp位点。根据在egln2基因上设计的6对引物,分别用团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各20个样品扩增相应的基因片段。对扩增片段进行测序和比对,在p1和p2两对引物扩增的基因片段上各发现1个snp位点:t397c397和t715g715(表2)。而在其他4对引物扩增片段上没有发现snp位点。
表2
2.2egln2基因多态性分析
用团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各100尾对egln2基因上筛选得到的snp位点进行扩增测序,分析测序结果发现egln2基因上的2个snp位点均发生错义突变。其中,t397c397位点密码子改变类型为tac-cac,分别编码酪氨酸和组氨酸;t715g715位点密码子改变类型为ttg-gtg,分别编码亮氨酸和缬氨酸。
egln2基因上的snp位点的多态性遗传参数见表3。根据pic>0.5为高度多态,0.25<pic<0.5为中度多态,pic<0.25为低度多态的原则,发现egln2上的2个snp位点均为高度多态位点。
表3
2.3egln2基因snp位点与低氧性状关联分析
将egln2基因上检测得到的snp位点基因型与团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各100尾的低氧性状关联酶(cat酶、sod酶及na+/k+-atp酶)活力进行相关性分析。分析结果发现,在egln2基因上的2个snp位点不同基因型均在低氧性状关联酶(cat酶、sod酶及na+/k+-atp酶)活力上存在显著差异(p<0.05)(表4)。
表4
注:同一位点同列中不同大写字母表示差异显著(p<0.05)
2.4egln2基因snp位点连锁不平衡和单倍型分析
用shesis软件分析差异位点的连锁不平衡性。结果显示egln2基因上的t397c397和t715g715这2个位点为一组完全连锁不平衡(d'=1,r2=1)。这也验证了表2和表3中这两组完全连锁位点组内的基因型频率、基因频率、群体遗传参数完全相同的结果。用这2个snp位点做单倍型构建和分析,剔除频率小于1%的单倍型,共得到了2种单倍型,发现单倍型ⅰ(c397g715)在对照团头鲂“浦江1号”群体中出现的频率为83.5%,在团头鲂耐低氧选育f5代群体中出现的频率为27.5%,差异极显著(p<0.01)。而单倍型ⅱ(t397t715)在对照团头鲂“浦江1号”群体中出现的频率为16.5%,在团头鲂耐低氧选育f5代群体中出现的频率为72.5%,差异极显著(p<0.01)(表5)。
表5
注:**表示差异极显著(p<0.01)
2.5egln2基因snp位点双倍型分析
随机将2种单倍型两两组合成双倍型,剔除个体数小于样本数5%的组合,可组成3种双倍型,即双倍ⅰ型(c397c397g715g715)、双倍ⅱ型(t397t397t715t715)和双倍iii型(t397c397t715g715),分析其与耐低氧性状的相关性。分析结果发现,双倍ⅱ型组合在低氧性状关联酶(na+/k+-atp酶、cat酶和sod酶)活力上显著高于其他组合(p<0.05)(表6)。
表6
注:同一列内数字上标小写字母不同表示差异显著(p<0.05).
2.6体型失衡的关键溶氧值
鱼类体型失衡的关键溶氧值(loecrit)的高低受温度的影响。双倍ⅰ型个体在10℃时loecrit值为0.69mg·l-1,在25℃和30℃时分别为0.90mg·l-1和1.31mg·l-1,耐低氧能力显著降低(p<0.01);双倍ⅱ型个体在10℃时loecrit值为0.46mg·l-1,在25℃和30℃时分别为0.81mg·l-1和1.18mg·l-1,差异显著(p<0.01)。
在同一温度下,双倍ⅰ型个体loecrit值均高于双倍ⅱ型个体。10℃时,双倍ⅰ型个体loecrit值为0.69mg·l-1,双倍ⅱ型个体为0.46mg·l-1,耐低氧能力提高了50%,差异显著(p<0.01)(图1);25℃时,双倍ⅰ型个体loecrit值为0.90mg·l-1,双倍ⅱ型个体为0.81mg·l-1,耐低氧能力提高了11.1%,差异显著(p<0.01)(图1)。30℃时,双倍ⅰ型个体loecrit值为1.31mg·l-1,双倍ⅱ型个体为1.18mg·l-1,耐低氧能力提高11.0%,差异显著(p<0.01)(图1)。
2.7鳃形态变化
低氧条件下(t=10±1.0℃,do=2.0mg·l-1),双倍ⅰ型和ⅱ个体呼吸面积均不断增加,低氧7d时,双倍ⅰ型个体呼吸面积(4.11×104μm2)显著大于双倍ⅱ型(3.12×104μm2)(p<0.01)(图2,图3)。
低氧条件下(t=10±1.0℃,do=2.0mg·l-1),低氧处理4和7d,双倍ⅱ型个体鳃小片层间基质体积分别是3.1×105μm3和2.8×105μm3,而双倍ⅰ型个体分别是2.35×105μm3和1.89×105μm3,双倍ⅰ型和ⅱ个体鳃小片层间基质体积均快速减少。但,双倍ⅰ型个体的鳃小片层间基质体积明显少于双倍ⅱ型个体(p<0.01)。从这方面也说明双倍ⅱ型个体对低氧胁迫有更大的潜力(图2,图3)。
2.8红细胞计数和血红蛋白浓度对低氧的反应
在常氧时,双倍ⅰ型个体红细胞平均计数为1.75×1012l-1,低氧4和7d后分别为1.85×1012l-1和2.05×1012l-1,差异显著(p<0.01)(图4中的a)。双倍ⅰ型个体常氧时血红蛋白(hb)浓度为49.6±2.1g·l-1,低氧4和7d后分别为56.3±0.9g·l-1和61.3±1.0g·l-1,差异显著(p<0.01)(图4中的b)。
双倍ⅱ型个体在低氧4和7d后红细胞平均计数分别为1.96×1012l-1和2.17×1012l-1,显著高于双倍ⅰ型个体(p<0.01)(图4中的a)。此外,双倍ⅱ型个体在低氧4和7d后,血红蛋白浓度从常氧时的52.2±0.9g·l-1升高到62.1±0.9g·l-1和69.5±0.8g·l-1,差异性显著(p<0.01)(图4中的b)。低氧4和7d时,双倍ⅱ型个体的血红蛋白浓度与双倍ⅰ型个体相比,差异显著(p<0.01)(图4中的b)。
鱼类耐低氧性状作为一个可遗传性状,属于多基因控制的数量性状。脯氨酸羟化酶作为生物体与低氧相关的限速酶,在低氧应答过程中起到重要作用。本实验根据已分离的团头鲂egln2基因的cdna序列设计引物,经引物扩增、序列对比后,在egln2基因cdna序列上发现2个snp位点:t397c397和t715g715。多态性分析发现在egln2基因中,2个位点表现出高度多态性。经连锁不平衡分析发现,t397c397和t715g715为一组完全连锁位点。多个位点的连锁可能蕴含着更大的信息量,在分子辅助育种中更具应用价值。
在测定的耐低氧性状参数中,超氧化物歧化酶(sod)和过氧化氢酶(cat)是生物抗氧化体系中的关键酶,na+/k+-atp酶则是参与能量代谢的关键酶,都可以作为评价鱼类低氧耐受性能的指标。本申请在用团头鲂耐低氧选育f5代和对照团头鲂“浦江1号”各100尾检测egln2基因上的2个snp位点时发现,这2个位点不同基因型均在低氧性状关联酶(cat酶、sod酶及na+/k+-atp酶)活力上存在显著差异(p<0.05)。这说明egln2基因对团头鲂耐低氧能力有重要影响。通过对egln2基因上不同位点基因型进行个体选育,不但可以改良团头鲂耐低氧性状,而且将有可能推进团头鲂耐低氧新品种的选育。在egln2基因上的2个snp位点发生了错义突变,而且这2个位点均位于该基因的编码区内并与低氧性状关联酶(cat酶、sod酶及na+/k+-atp酶)活力显著相关,这有可能是错义突变导致编码氨基酸的改变,进而改变了脯氨酸羟化酶功能活性,对耐低氧性状产生影响。
之前用传统方法分析单个snp位点与目标性状之间的相关性,总会有一些位点信息模糊和不完整等问题出现,而单倍型的构建和分析则可以有效解决这些问题。本申请在egln2基因上挑选与低氧性状关联酶(cat酶、sod酶及na+/k+-atp酶)活力均存在显著差异的snp位点,并进一步做了单倍型的构建和分析。在egln2基因构建的2种单倍型中,团头鲂“浦江1号”群体中存在ⅰ这种主要优势单倍型,团头鲂耐低氧选育f5代群体中存在ⅱ这种主要优势单倍型。单倍型ⅰ在对照团头鲂“浦江1号”群体中出现的频率极显著高于团头鲂耐低氧选育f5代群体,而单倍型ⅱ在团头鲂耐低氧选育f5代群体中出现的频率极显著高于团头鲂“浦江1号”群体。双倍ⅱ型在低氧性状关联酶(cat酶、sod酶和na+/k+-atp酶)活力上显著高于其他组合(p<0.05)。可以推断这两个位点的纯合突变为耐低氧的有利突变,因此,单倍型ⅰ和ⅱ可分别作为对照团头鲂“浦江1号”群体和团头鲂耐低氧选育f5代群体的优势单倍型。在今后的团头鲂耐低氧新品种选育工作中,可以针对耐低氧性状选择具有优势基因型的个体,从而达到加快育种的目的。
loecrit值是用来测定鱼类耐低氧性能的指标。loecrit值较低的鱼能够适应更低的溶解氧水平。在本申请中,双倍ⅱ型个体在25℃时的loecrit值为0.81±0.05mg·l-1,远低于一些海水鱼。在10℃时,双倍ⅰ型和ⅱ个体loecrit值分别为0.72mg·l-1和0.54mg·l-1。同时,在25℃和30℃时,双倍ⅰ型个体loecrit值分别为1.03mg·l-1和1.41mg·l-1。在相同温度条件下,双倍ⅰ型个体loecrit值大于双倍ⅱ型个体。这些结果表明双倍ⅱ型个体耐低氧性高于双倍ⅰ型个体。
水体中的溶解氧随着环境的改变而改变,鱼类为了适应溶解氧的变化,或是增加鳃呼吸表面积,或是增加血液中氧的承载能力。在此之前,我们研究发现团头鲂在低氧水环境中具有鳃重构能力。在本申请中,双倍ⅰ型和ⅱ个体在低氧的情况下,出现了鳃小片的呼吸面积增加和层间基质体积减少的现象。在自然水体中,由于鳃小片层间基质细胞的增殖,鳃呼吸面积减少,这种现象不利于鱼类呼吸。本申请中,双倍ⅱ型个体在10℃低氧胁迫7d,鳃小片伸出高度和鳃小片呼吸面积显著低于双倍ⅰ型(p<0.01)。这些数据表明,双倍ⅱ型个体在阻止有毒物质流入血液和应对进一步的低氧胁迫方面具有更大的潜力。
之前有报道对团头鲂进行转录组分析,结果发现团头鲂在低氧条件下,促红细胞生成素(epo)mrna明显上调,这可以导致团头鲂红细胞数和血红蛋白浓度升高,提高血液携氧能力。本申请中,在低氧4和7d,双倍ⅱ型个体红细胞数和血红蛋白浓度比双倍ⅰ型个体显著升高(p<0.01)。这种极高的血红蛋白氧亲和力,即使是在鳃小片完全被细胞团包围,血液中也会有足够的氧气。这可能就是双倍ⅱ型个体在低氧4d和7d时鳃ilcm体积小于双倍ⅰ型个体的原因。
因此,双倍ⅱ型个体表现出较高的耐低氧能力,单倍ⅱ型可用于团头鲂耐低氧新品系分子辅助育种。
本领域技术人员应理解,以上实施例仅是示例性实施例,在不背离本发明的精神和范围的情况下,可以进行多种变化、替换以及改变。
序列表
<110>上海海洋大学
<120>团头鲂耐低氧关联snp位点的单倍型在育种中的应用
<130>hp191211lz
<141>2019-06-11
<160>12
<170>siposequencelisting1.0
<210>1
<211>22
<212>dna
<213>未知()
<400>1
gtttactgtcgagcttcgtgat22
<210>2
<211>21
<212>dna
<213>未知()
<400>2
aaggggagcgacagcctgtgt21
<210>3
<211>19
<212>dna
<213>未知()
<400>3
cggaaatgccccatggact19
<210>4
<211>23
<212>dna
<213>未知()
<400>4
tttagtgcaatggaacagtttca23
<210>5
<211>21
<212>dna
<213>未知()
<400>5
atttaaccgttcatcaccctt21
<210>6
<211>21
<212>dna
<213>未知()
<400>6
gacgcttttaagcctgccgtt21
<210>7
<211>24
<212>dna
<213>未知()
<400>7
tcagtcaagatgagagtcttctct24
<210>8
<211>26
<212>dna
<213>未知()
<400>8
caattgtagactttgagtcactgaat26
<210>9
<211>22
<212>dna
<213>未知()
<400>9
gcacatctcaaactccatatca22
<210>10
<211>21
<212>dna
<213>未知()
<400>10
gaaaatgtgttcccaccaacc21
<210>11
<211>22
<212>dna
<213>未知()
<400>11
aaaggtcaggtcatttttgcta22
<210>12
<211>21
<212>dna
<213>未知()
<400>12
atgttgggcttctcgcatctt21
- 还没有人留言评论。精彩留言会获得点赞!