一种快速构建3型鸭甲肝病毒反向遗传株的方法与流程
本发明属于兽医病毒分子生物学技术领域,具体涉及一种快速构建3型鸭甲肝病毒反向遗传株的方法。
背景技术:
鸭病毒性肝炎(duckviralhepatitis,dvh)是由鸭肝炎病毒(duckhepatitisvirus,dhv)感染雏鸭引起的一种急性、高度接触性传染病。目前世界上主要养鸭地区均有本病的存在,具有间歇爆发、地方流行性等特点,是危害养鸭业的主要疾病之一。该病主要侵害四周龄以内的雏鸭,具有发病急,传播迅速,病程短和死亡率高等特点;临床主要表现为雏鸭死前发生痉挛,头向背部后仰,呈“角弓反张”,病理变化主要为剖检可见肝脏肿大发炎和大量的出血性斑点。该病主要由属于小rna病毒科禽肝病毒属的鸭甲型肝炎病毒(duckhepatitisavirus,dhav)引起。dhav具有三种血清型,即1型、2型和3型。近年来,我国流行的dhav主要是1型鸭甲肝病毒(dhav-1)和3型鸭甲肝病毒(dhav-3)。
反向遗传操作技术是开展病毒分子生物学研究的一种重要平台,可以在体外对rna病毒基因组进行基因敲除、定点诱变等人工操作,在阐明病毒致病机制和疫苗研制中能够发挥重要作用并具比自然诱变周期短的优点。传统rna病毒感染性克隆方法的关键是获得病毒基因组cdna全长片段克隆,并且病毒基因组转换为cdna后需要克隆到合适的载体中,为了避免病毒序列在细菌中的不稳定问题,研究者通常采取分段克隆方法,通过小片段连接成大片段,最后将大片段通过酶切连接的方法得到全长cdna克隆。但是该方法酶切位点选取限制较多,体外将多个大片段进行连接效率较低,另外一些复制酶基因cdna克隆在细菌中具有不稳定性,因此获得病毒基因组全长cdna的整个过程不仅操作步骤麻烦,而且耗时较长,成功率不高,同时,有些病毒的全长cdna无法克隆或是虽然能克隆进载体但在宿主菌中易发生变异而导致不能成功拯救病毒。目前,一种被称为“感染性亚基因组复制子(infectioussubgenomicamplicons)”的技术已被证实能够在哺乳动物或蚊子细胞中实现对单股正链rna病毒的人工拯救,并被应用于日本脑炎病毒、西尼罗病毒、寨卡病毒、黄热病病毒、登革病毒和人科萨奇病毒等病毒的反向遗传学研究中。该技术是一种新型的“无细菌”反向遗传学方法,不需要获得病毒全长cdna质粒以及在体外获得病毒rna转录本,可以直接通过转染具有同源区域的dna片段拯救病毒。具体的的来说,“感染性亚基因组复制子”采用的技术路线是:首先通过pcr方法产生包含整个病毒基因组的重叠的非感染性亚基因组dna片段,这些重叠的亚基因组复制子数量可以为3至10个,相邻之间的片段之间具有100bp左右的重叠区域,同时,第一个片段的5'端和最后一个片段的3'末端分别侧接巨细胞病毒早期启动子(cytomegalovirusimmediateearlypromoter,pcmv)序列、丁型肝炎病毒核酶(hepatitisdeltavirusribozyme,hdvr)序列和猴空泡病毒早期mrna多腺苷酸化信号序列(sv40earlymrnapolyadenylationsignal,sv40pa)序列,这些元件能够帮助亚基因组复制子在被混合转染到易感细胞后开始转录,并利用宿主细胞的同源重组机制自发重组,形成具有感染性的完整病毒转录本,继而导致病毒的复制与增殖,最终获得具有感染性的拯救病毒。
虽然目前已经有关于3型鸭甲肝病毒感染性克隆的构建以及拯救方法的报道,但是研究者采用的仍然是构建病毒全长感染性cdna克隆的策略,同时有研究结果显示含有3型鸭甲肝病毒全基因组克隆的质粒在传代过程中具有较高的突变率,可能是因为病毒部分序列具有潜在毒性以及不稳定性。因此,若能使用“感染性亚基因组复制子”技术建立一种简便、高效的3型鸭甲肝病毒“无细菌”反向遗传系统,这将为研究该病毒的致病机理、开发新型疫苗等提供了有利的工具。
技术实现要素:
本发明的目的是提供一种快速构建3型鸭甲肝病毒反向遗传株的方法,该方法使用“感染性亚基因组复制子”技术,避免现有技术中鸭3型甲肝病毒病毒全长cdna克隆在细菌中不能稳定保存的问题,传统的酶切连接效率较低的问题,以及体外转录导致的转录本异质性的问题;利用该方法可快速、简便构建3型鸭甲肝病毒突变株。
为了实现上述技术目的,本发明句通过以下技术方案实现:
一种快速构建3型鸭甲肝病毒反向遗传株的方法,具体包括以下步骤:
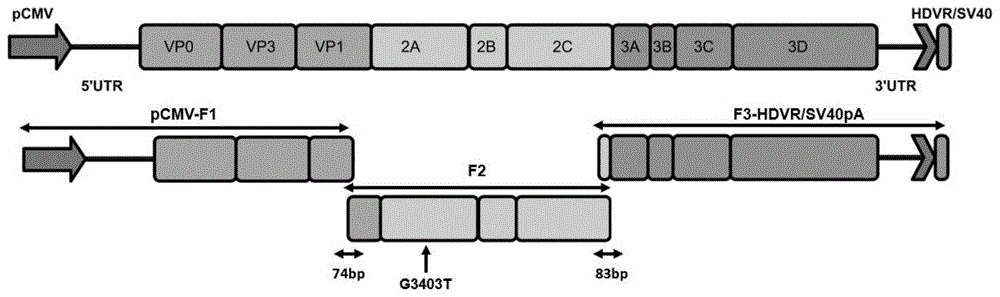
1)将亲本病毒株全基因组分为大小相近的三个片段进行pcr扩增,在病毒基因组第一个片段的5'端添加巨细胞病毒早期启动子(cytomegalovirusimmediateearlypromoter,pcmv),在第二个片段的2a基因中引入同义突变作为遗传标记位点,在第三个片段的3'端添加丁型肝炎病毒核酶(hepatitisdeltavirusribozyme,hdvr)序列和猴空泡病毒早期mrna多腺苷酸化信号(sv40earlymrnapolyadenylationsignal,sv40pa)序列,获得三个dna片段,构成3型鸭甲型肝炎病毒感染性亚基因组复制子;
2)将构建的感染性亚基因组复制子与转染试剂混合后转染鸭胚成纤维细胞,复制子在宿主细胞内转录并利用细胞的同源重组机制自发重组,形成具有感染性的完整病毒转录本,继而导致病毒的复制与增殖,获得含有遗传标记的3型鸭甲型肝炎病毒反向遗传株病毒。
进一步的,所述的第一个片段与第三个片段分别与第二个片段含有74个和83个碱基对的重叠区域。
进一步的,用融合pcr方法在3型鸭甲肝病毒基因组第二个片段的2a基因中引入了一个同义碱基突变,从而反向遗传株病毒的第3403位核苷酸的g突变为t,该位点作为感染性克隆的遗传分子标记,可通过pcr方法结合dna测序来区分反向遗传株病毒与亲本株以及野毒株。
所述的3型鸭甲肝病毒(duckhepatitisavirustype3,dhav-3)保藏于中国典型培养物保藏中心,保藏编号为cctccno:v201305,保藏地址:湖北省武汉市武昌区八一路299号武汉大学校内,保藏日期:2013年3月25日。
本发明方法获得的含有遗传标记的拯救病毒在鸭胚上的增殖能力和病毒滴度、对雏鸭的致病力与亲本病毒相似,且遗传稳定性良好。
本发明方法可对3型鸭甲肝病毒的任何碱基进行突变或片段缺失/替换/插入,获得新的突变重组病毒。
本发明的有益效果为:
1、首次证明“感染性亚基因组复制子”技术能够用于禽类病毒的反向遗传拯救中。
2、本发明中获得的拯救病毒具有与亲本病毒相同的特性,在鸭胚中能够稳定增殖传代,且具有与亲本病毒相近的增殖能力和抗原性。
3、发明提供的方法操作简单、快速,可对3型鸭甲肝病毒的任何碱基进行突变或片段缺失/替换/插入而获得新的突变重组病毒,为研究3型鸭甲肝病毒的致病机理等提供了有效的工具,并且对于加快新型疫苗的开发具有重要的意义。
附图说明
图1为3型鸭甲肝病毒反向遗传株“感染性亚基因组复制子”的模式图;
图2为pcmv-f1、f2和f3-hdvr/sv40pa片段琼脂糖凝胶电泳图;m为dl5,000dnamarker,1号至3号泳道依次为pcmv-f1、f2和f3-hdvr/sv40pa片段;
图3为3型鸭甲肝病毒反向遗传株“感染性亚基因组复制子”转染鸭胚成纤维细胞结果图;
图4为拯救病毒与亲本病毒f2基因片段blni酶切结果电泳图;m为dl5,000dnamarker,1号和2泳道分别为拯救病毒与亲本病毒;
图5为拯救病毒的分子遗传标记位点核苷酸测序结果;
图6为3型鸭甲肝病毒突变株“感染性亚基因组复制子”的模式图;
图7为3型鸭甲肝病毒突变株“感染性亚基因组复制子”转染鸭胚成纤维细胞结果图。
具体实施方式
以下结合具体实施例,对本发明进行详细说明,不是对本发明的进一步限定。下述实施例中所使用的实验方法如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中所用的材料和试剂具体如下:
病毒株:
3型鸭甲肝病毒强毒株,本实验室分离得到,已保藏于中国武汉大学的中国典型培养物保藏中心,保藏编号为:cctccno:v201305,分类命名:禽肝病毒属小rna病毒科3型鸭甲肝病毒(duckhepatitisavirustype3,dhav-3)。
试剂及仪器:
takaraminibestuniversalrnaextractionkit,primestarmaxdnapolymerase,dnamarker等购自宝生物工程(大连)有限公司;胶回收试剂盒,质粒抽提试剂盒等购自美国omega公司;脂质体转染试剂盒lipofectamine3000购自invitrogen公司;其他试剂均为国产分析纯。
核酸蛋白检测仪(biorad,smartspec3000)、梯度pcr仪(biometra,tgradient)、电泳仪(biorad,powerpac300)和凝胶成像系统(bioradversadocmodel2000)。
实施例13型鸭甲肝病毒“感染性亚基因组复制子”的构建及病毒拯救
1.1.引物的设计与合成
根据genbank中3型鸭甲肝病毒的全基因组序列,设计了6对引物用于扩增病毒基因组序列、pcmv和sv40pa序列,具体序列信息见表1,引物由上海生工生物工程有限公司合成。
表1构建3型鸭甲肝病毒反向遗传株“感染性亚基因组复制子”所需引物的序列
1.2.病毒rna抽提
参照takaraminibestuniversalrnaextraction试剂盒的使用说明书操作,从鸭胚尿囊液中抽提病毒的全基因组rna,并使用核酸蛋白检测仪(biorad,smartspec3000)测定其的核酸浓度和纯度后,-70℃保存备用。
1.3.基因片段扩增克隆
(1)使用primescriptii1ststrandcdnasynthesis试剂盒将抽提的总rna反转录为cdna模板,然后使用dna高保真pcr酶primestarmaxdnapolymerase,以亲本株病毒总rna的反转录产物为模板,使用引物f1-f和f1-r扩增得到dhav-3-f1片段,使用引物f2-1-f和f2-1-r扩增得到dhav-3-f2-1片段,使用引物f2-2-f和f2-2-r扩增得到dhav-3-f2-2片段,使用引物f3-hdvr-f和f3-hdvr-r扩增得到dhav-3-f3-hdvr片段;以真核表达质粒pegfp-c1为模板,使用引物pcmv-f和pcmv-r扩增得到pcmv片段,使用引物hdvr-sv40pa-f和hdvr-sv40pa-r扩增得到hdvr-sv40pa片段;
(2)通过融合pcr技术,如图1所示,将pcmv和dhav-3-f1片段融合为pcmv-f1,将dhav-3-f2-1和dhav-3-f2-2片段融合为f2片段,将dhav-3-f3-hdvr和hdvr-sv40pa片段融合为f3-hdvrz/sv40pa片段,琼脂糖凝胶电泳结果如图2所示,扩增片段经1%琼脂糖凝胶电泳分离后采用凝胶回收试剂盒(omega)切胶回收目的片段,并将dna片段送至上海生工生物工程有限公司进行测序,测序结果表明成功完成反向遗传株“感染性亚基因组复制子”的构建。
1.4.转染反向遗传株“感染性亚基因组复制子”拯救病毒
使用9日龄鸭胚制备原代鸭胚成纤维细胞,当3.5cm培养皿中的细胞生长至90%融合时,将等量(1.5μg)的pcmv-f1、f2和f3-hdvrz-sv40pa基因片段混合后,使用lipofectamine3000(invitrogen)按说明书转染细胞。细胞置于37℃5%co2培养箱中进行培养观察,16小时后更换培养基,在转染72小时后,转染组细胞出现细胞碎裂现象,而对照组细胞生长状况良好。在转染鸭胚成纤维细胞120小时后,细胞生长状况如图3所示,拍照记录后将细胞反复冻融3次,然后将细胞培养液上清通过尿囊腔途径接种5枚9日龄鸭胚,0.2ml/枚,石蜡封口后放入孵化箱内继续孵育,每隔8小时照胚1次,观察鸭胚在接种后的死亡情况,弃去24小时内死亡鸭胚。结果显示鸭胚在接种后48小时至120小时之间死亡,鸭胚死亡胚体出血严重,收集尿囊液作为第一代反向遗传株进行保存。
实施例2反向遗传株病毒的鉴定和特性
2.1.反向遗传株病毒的传代
为观察拯救出来的拯救病毒的是否能够在鸭胚中增殖传代,将第1代的拯救病毒用灭菌生理盐水做1:100稀释,接种5枚9日龄鸭胚。结果显示鸭胚死亡时间集中在接种后48小时至120小时之间,胚体病变明显,收集死亡鸭胚尿囊液,并连续传代5次,鸭胚均病变明显,与亲本病毒相似。
2.2.反向遗传株病毒中遗传标记的鉴定
为了排除反向遗传株病毒可能来自于转染或传代过程中亲本病毒或野毒株污染的可能,在复制子构建过程中,我们采用融合pcr方法在f2基因片段的2a基因中引入一个碱基突变(g突变为t),突变后产生1个病毒本身不具有的遗传标记blni限制性内切酶位点,同时该突变不改变2a蛋白相应氨基酸的组成。拯救病毒在鸭胚上传代5次。从尿囊液中提取总rna,用f2-f和f2-r引物扩增含有突变位点的dna片段,并按照限制性内切酶blni推荐的酶切体系,将f2基因片段进行酶切后琼脂糖凝胶电泳结果如图4所示,结果表明,dna转染拯救病毒的f2片段用blni酶切可产生0.9kb和1.7kb的两个片段,而亲本病毒的2.6kb扩增片段无法用blni酶切。扩增片段经1%琼脂糖凝胶电泳分离后采用凝胶回收试剂盒(omega)切胶回收目的片段,并将dna片段送至上海生工生物工程有限公司进行测序,测序结果显示扩增的产物含有引入的沉默突变(g3403t),如图5所示,表明我们获得了正确的拯救病毒,而非野生型毒株污染。
2.3.病毒增殖及含量测定
用无菌生理盐水溶液倍比稀释3型鸭甲肝病毒反向遗传株与亲本株,选择10-3、10-4、10-5、10-6、10-7、10-8这6个稀释度,分别以每胚0.2ml的量经尿囊腔途径接种5枚9日龄的鸭胚,另设灭菌生理盐水对照5枚,接种后置于37℃恒温培养箱内孵育,24小时内死亡鸭胚不计,观察并记录7天内接种鸭胚的死亡和存活情况,按reed-muech法计算eld50,结果显示拯救病毒与亲本株病毒具有相似的增殖能力,每0.2ml尿囊液中病毒含量分别为10-4.33eld50和10-4.50eld50,拯救病毒在鸭胚中增殖能力与亲本病毒相似。
2.4.血清中和试验检测病毒的抗原性
采用固定病毒稀释血清法测定兔抗3型鸭甲肝病毒血清对反向遗传株与亲本株的中和效价,首先将发明人所在实验室前期制备的兔抗dhav-3标准血清(效价≥1:128)用灭菌生理盐水作10倍倍比稀释从2-1到2-9这9个稀释度,同时将经毒价滴定的病毒稀释成每个单位剂量(0.2ml)含200eld50,然后两者等量混合并加入5%的青、链霉素双抗37℃水浴l小时,然后以每胚0.2ml的计量接种9日龄健康鸭胚尿囊腔内,每个稀释度接种5枚鸭胚。另设阴性血清对照(健康兔血清与病毒混合)和空白对照组(灭菌生理盐水与病毒混合),弃去24小时内死亡的鸭胚,观察并记录7天内的鸭胚死亡和存活的情况,然后计算病毒的中和效价。
结果拯救病毒与亲本株病毒的空白对照组和阴性血清对照组鸭胚均在48小时至96小时间全部死亡;当血清稀释度在2-1~2-6之间时,拯救病毒和亲本病毒中和组的鸭胚全部健活,当稀释度在2-7时鸭胚开始失去保护,此时开始,随稀释度越高,鸭胚保护率越低,直到稀释度为2-9时鸭胚彻底失去保护,表明拯救病毒和亲本株病毒具有相同的抗原性。
2.5.对易感雏鸭的致病力试验
使用亲本株和拯救毒株进行致病力试验,采集实施例2.3中死亡鸭胚的肝脏组织匀浆处理,按体积比1:100的比例加入灭菌磷酸盐缓冲溶液,研磨,反复冻融3次,12000g离心10min,上清液经0.22μm滤器过滤除菌后经尿囊腔途径接种9日龄鸭胚测定其eld50,然后将病毒液稀释至103.0eld50/0.4ml。另外将1日龄健康雏鸭30只随机分为3组,试验组的雏鸭经肌肉注射途径接种0.4ml亲本株或拯救毒株,对照组的雏鸭则接种等体积灭菌生理盐水,不同组雏鸭在不同动物房内隔离饲养,自由饮水采食,接种后每日观察,记录雏鸭的发病、死亡情况,及时剖检死亡雏鸭,观察7天后剖检存活雏鸭,记录雏鸭肝脏、肾脏等器官的病变情况。
7天内对照组雏鸭未出现临床症状,雏鸭进食和饮水状况正常;拯救病毒和亲本株病毒对1日龄雏鸭具有相似的致病力,接种后雏鸭发病率均为80%,死亡率均为60%,同时,存活雏鸭剖检肝脏也均可见部分针尖大小出血点。
实施例3应用3型鸭甲肝病毒“感染性亚基因组复制子”方法拯救isa-a117c突变株病毒
3.1.引物的设计与合成
根据genbank中3型鸭甲肝病毒的全基因组序列,设计了7对引物用于扩增病毒全基因组序列、pcmv和sv40pa序列,具体序列信息见表2,引物由上海生工生物工程有限公司合成。
表2构建3型鸭甲肝病毒突变株isa-a117c“感染性亚基因组复制子”所需引物的序列
3.2.病毒抽提
参照takaraminibestuniversalrnaextraction试剂盒的使用说明书操作,从鸭胚尿囊液中抽提3型鸭甲肝病毒分离株的病毒全基因组rna,并使用核酸蛋白检测仪(biorad,smartspec3000)测定其的核酸浓度和纯度后,-70℃保存备用。
3.3.基因片段扩增克隆
(1)使用primescriptii1ststrandcdnasynthesis试剂盒将抽提的总rna反转录为cdna模板,然后使用dna高保真pcr酶primestarmaxdnapolymerase,使用引物f1-f和a117c-r、a117c-f和f1-r以亲本株病毒总rna的反转录产物为模板扩增得到dhav-3-f1-a117c-f和dhav-3-f1-a117c-r片段,使用引物f2-1-f和f2-1-r、f2-2-f和f2-2-r以亲本株病毒总rna的反转录产物为模板分别扩增得到dhav-3-f2-1片段和dhav-3-f2-2片段,使用引物f3-hdvr-f和f3-hdvr-r以亲本株病毒总rna的反转录产物为模板扩增得到dhav-3-f3-hdvr片段;以真核表达质粒pegfp-c1为模板,使用引物pcmv-f和pcmv-r扩增得到pcmv片段,使用引物hdvr-sv40pa-f和hdvr-sv40pa-r扩增得到hdvr-sv40pa片段。(2)通过融合pcr技术,如图6所示,先将dhav-3-f1-a117c-f和dhav-3-f1-a117c-r片段融合为dhav-3-f1-a117c,然后将pcmv和dhav-3-f1-a117c片段融合为pcmv-f1;将dhav-3-f2-1和dhav-3-f2-2片段融合为f2片段;将dhav-3-f3-hdvr和hdvr-sv40pa片段融合为f3-hdvrz/sv40pa片段,最终获得的三个dna片段构成突变株isa-a117c“感染性亚基因组复制子”。扩增片段经1%琼脂糖凝胶电泳分离后,分别采用凝胶回收试剂盒(omega)切胶回收目的片段。将dna片段送至上海生工生物工程有限公司进行测序。
3.4.突变株isa-a117c“感染性亚基因组复制子”的转染拯救
使用9日龄鸭胚制备制备原代鸭胚成纤维细胞,当3.5cm培养皿中的细胞生长至90%融合时,将等量(1.5μg)的pcmv-f1、f2和f3-hdvrz-sv40pa基因片段与lipofectamine3000(invitrogen)混合后转染长满90%的鸭胚成纤维细胞,对照组仅使用lipofectamine3000(invitrogen)进行转染。细胞置于37℃5%co2培养箱中进行培养观察,16小时后更换培养基,在转染72小时后,转染组细胞出现细胞碎裂现象,而对照组细胞生长状况良好。在转染鸭胚成纤维细胞120小时后,细胞生长状况如图7所示,拍照记录后将细胞反复冻融3次,细胞培养液通过尿囊腔途径接种5枚9日龄鸭胚,0.2ml/枚,石蜡封口后放入孵化箱内继续孵育,每隔8小时照胚1次,观察鸭胚在接种后的死亡情况,弃去24小时内死亡鸭胚。结果显示鸭胚在接种后24小时至48小时之间死亡,鸭胚死亡胚体出血严重,收集尿囊液作为第一代反向遗传病毒株进行保存。
3.5.反向遗传突变株中遗传标记和突变位点的鉴定
为了排除反向遗传突变株病毒可能来自于转染或传代过程中亲本病毒或野毒株污染的可能,利用反向遗传学方法,突变株基因组的3403位碱基由突变g突变为t,该突变不改变2a蛋白相应氨基酸的组成,该位点作为分子遗传标记位点,可通过pcr方法结合dna测序来区分突变株与亲本株以及野毒株。拯救病毒在鸭胚上通过有限稀释法传代纯化5次。从尿囊液中提取总rna,反转录后使用f2-f和f2-r引物pcr扩增含有遗传标记位点的dna片段,使用f1-f和f1-r引物pcr扩增含有目标突变位点的dna片段,扩增片段经1%琼脂糖凝胶电泳分离,然后使用凝胶回收试剂盒(omega)切胶回收,并将dna片段送至上海生工生物工程有限公司进行测序,测序结果显示扩增的产物含有引入的遗传标记位点(g3403t)和目标突变位点(a117c),表明我们获得了正确的拯救病毒,而非亲本株或者野生型毒株污染。
尽管已经示出和描述了本发明的实施例,对于本领域的普通技术人员而言,可以理解在不脱离本发明的原理和精神的情况下可以对这些实例进行多种变化、修改、替换和变型,本发明的范围由所附权利要求及其等同物限定。
序列表
<110>四川农业大学
<120>一种快速构建3型鸭甲肝病毒反向遗传株的方法
<160>17
<170>siposequencelisting1.0
<210>1
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>1
tagttattaatagtaatcaattacggggtca31
<210>2
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>2
acaccacagccgctttcaaacggttcactaaaccagctct40
<210>3
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>3
agagctggtttagtgaaccgtttgaaagcggctgtggtgt40
<210>4
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>4
caacctgccaaaagtcaaacca22
<210>5
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>5
attctgttacacctttacgccccaca26
<210>6
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>6
caacctaggtaagtgagcacgat23
<210>7
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>7
gtgctcacttacctaggttggtt23
<210>8
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>8
tggcaacttcctgtctaacctg22
<210>9
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>9
ccttgaacactggaacccaa20
<210>10
<211>69
<212>dna
<213>人工序列(artificialsequence)
<400>10
aagtagcccaggtcggaccgcgaggaggtggagatgccatgccgaccctttttttttttt60
ttagggtgg69
<210>11
<211>59
<212>dna
<213>人工序列(artificialsequence)
<400>11
cggtccgacctgggctacttcggtaggctaagggagaagaacttgtttattgcagctta59
<210>12
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>12
taagatacattgatgagtttgga23
<210>13
<211>583
<212>dna
<213>巨细胞病毒早期启动子(pcmv)
<400>13
tagttattaatagtaatcaattacggggtcattagttcatagcccatatatggagttccg60
cgttacataacttacggtaaatggcccgcctggctgaccgcccaacgacccccgcccatt120
gacgtcaataatgacgtatgttcccatagtaacgccaatagggactttccattgacgtca180
atgggtggagtatttacggtaaactgcccacttggcagtacatcaagtgtatcatatgcc240
aagtacgccccctattgacgtcaatgacggtaaatggcccgcctggcattatgcccagta300
catgaccttatgggactttcctacttggcagtacatctacgtattagtcatcgctattac360
catggtgatgcggttttggcagtacatcaatgggcgtggatagcggtttgactcacgggg420
atttccaagtctccaccccattgacgtcaatgggagtttgttttggcaccaaaatcaacg480
ggactttccaaaatgtcgtaacaactccgccccattgacgcaaatgggcggtaggcgtgt540
acggtgggaggtctatataagcagagctggtttagtgaaccgt583
<210>14
<211>67
<212>dna
<213>丁型肝炎病毒核酶序列(hdvr)
<400>14
gggtcggcatggcatctccacctcctcgcggtccgacctgggctacttcggtaggctaag60
ggagaag67
<210>15
<211>122
<212>dna
<213>猴空泡病毒早期mrna多腺苷酸化信号(sv40pa)
<400>15
aacttgtttattgcagcttataatggttacaaataaagcaatagcatcacaaatttcaca60
aataaagcatttttttcactgcattctagttgtggtttgtccaaactcatcaatgtatct120
ta122
<210>16
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>16
gcctagtcctagcgctataggactccc27
<210>17
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>17
gggagtcctatagcgctaggactaggc27
- 还没有人留言评论。精彩留言会获得点赞!