一种草铵膦脱氢酶突变体及在氧化-还原多酶偶联生产L-草铵膦中的应用的制作方法
(一)技术领域
本发明涉及生物化工领域,涉及一种手性纯l-草铵膦的生产方法;是一种利用来源于微生物的氨基酸氧化酶和草铵膦脱氢酶偶联生产光学纯l-草铵膦的方法。
(二)技术背景
草铵膦(glufosinate-ammonium)是世界第二大转基因作物耐受除草剂,由赫斯特公司(几经合并后现归属拜耳公司)开发生产,化学名称为4-[羟基(甲基)膦酰基]-dl-高丙氨酸,又称草铵膦铵盐、basta、buster等,属膦酸类除草剂,是谷氨酰胺合成酶抑制剂,非选择性(灭生性)触杀型除草剂。
目前,世界上的三大除草剂品种是草甘膦、草铵膦和百草枯,相对于百草枯和草甘膦,草铵膦具有优异的除草性能及较小的药害副作用,随着抗草铵膦转基因作物的快速发展,草铵膦在未来一段时间内市场需求巨大,前景非常广阔。
草铵膦有两种光学异构体,分别为l-草铵膦和d-草铵膦。但只有l-型具有生理活性,且在土壤中易分解,对人类和动物的毒性较小,除草谱广,对环境的破坏力小。
草铵膦有d、l两种对映异构体,但只有l-构型具有植物毒性,且在土壤中易分解,对人类和动物的毒性小,对环境的破坏力小。目前,市场上销售的草铵膦一般都是外消旋混合物。如果草铵膦产品能以l-构型的光学纯异构体形式使用,可使草铵膦的使用量降低50%,这对于提高原子经济性、降低成本、减轻环境压力都具有十分重要的意义。目前报道的l-草铵膦的方法主要有化学合成法,包括消旋草铵膦的拆分、手性原料法、手性辅基法和不对称催化法,但存在d-草铵膦不易消旋再利用、合成步骤冗长,反应需要超低温,产物ee值较低,收率低,且手性拆分试剂价格昂贵等问题。相比之下,生物合成法具有立体选择性严格、反应条件温和及产物易分离纯化的优点,因此探索生物法生产l-草铵膦的可行性具有十分重要的产业价值和显著的社会效益。
生物法生产l-草铵膦以起始原料和途径进行分类,主要包括以下3大类:
1)以l-草铵膦的衍生物为底物,通过酶法直接水解获得,主要的优点转化率高,产物ee值较高,但需要昂贵且不易获得的手性原料作为前体(organophosphorusanaloguesandderivativesorthenaturall-aminocarboxylicacidsandpeptides.i.enzymaticsynthesisord-,dl-,andl-phosphinothricinandtheircyclicanalogues[j].bullchemsocjpn,1988,61(10):3699-3704.)。例如生物法制备l-草铵膦最简单的方法就是利用蛋白酶直接水解双丙氨膦。双丙氨膦是一种天然的三肽化合物,在蛋白酶的催化下,双丙氨膦脱去2分子l-丙氨酸,生成l-草铵膦。
2)以外消旋草铵膦的前提为底物,通过酶的选择性拆分获得。主要优点为原料相对易得,催化剂活力高,但其理论收率只能达到50%,会造成原料的浪费。natchev报道了一种采用α-胰凝乳蛋白酶拆分双丙氨膦乙酯制备l-草铵膦的方法,该法首先经过3步反应将外消旋的草铵膦合成双丙氨膦二乙酯,接着用碱性mesinterico肽酶将其c端脂基水解。再经α-胰凝乳蛋白酶催化水解肽键(cheminrormabstract:totalsynthesisandenzyme-substrateinteractionord-,dl-,andl-phosphinotricine,“bialaphos”(sr-1293)anditscyclicanalogues[j].cheminrorm,1989,1(17):125-131.)。在该步反应中,α-胰凝乳蛋白酶能选择性水解l-双丙氨膦乙酯,生成l-草铵膦乙脂。最后利用磷酸二脂酶水解p端酯基得到l-草铵膦。
3)以α-酮酸-2-羰基-4-(羟基甲基膦酰基)丁酸为底物,通过酶的不对称合成获得,主要涉及的酶包括转氨酶与氨基酸脱氢酶。早在研究草铵膦在土壤微生物体内的代谢途径时就已经发现,l-草铵膦在转氨酶的作用下,发生转氨作用被分解成2-羰基-4-(羟基甲基膦酰基)丁酸(简称ppo)。schulza等(stereospeciricproductionortheherbicidephosphinothricin(glurosinate)bytransamination:isolationandcharacterizationoraphosphinothricin-specirictransaminaserromescherichiacoli[j].applied&environmentalmicrobiology,1990,56(1):1-6.)在上世纪90年代就利用从大肠杆菌中克隆的转氨酶,以2-羰基-4-(羟基甲基膦酰基)丁酸为底物,l-谷氨酸作为氨基供体,催化转氨反应生产l-草铵膦。该在转氨酶经固定化并安装至生物反应器,催化制备l-草铵膦,其产物浓度可达76.1g/l,最高产率为50g/(l·h),l-草铵膦的ee值超过99.9%。但利用转氨酶制备l-草铵膦有两大缺陷,其一是原料ppo不能完全转化为l-ppt,转化率最高只有90%;其二是要使可逆反应向生成l-ppt的方向进行,需要4倍当量以上的l-谷氨酸作为氨基供体,过量的谷氨酸给l-草铵膦的分离带来了很大的麻烦。
4)以d,l-草铵膦为原料,其中的d-草铵膦经d-氨基酸氧化酶催化得到l-草铵膦前体2-羰基-4-[羟基(甲基)膦酰基]丁酸,再经氨基酸脱氢酶或转氨酶催化得到l-草铵膦,该方法不仅解决了消旋化的问题,也节约了成本。
d-氨基酸氧化酶是一类特异选择性的将d-氨基酸及其衍生物催化生成α-酮酸的酶,反应由其本身自带的辅酶fad催化完成,因其表现处的优良催化效率和选择性,d-氨基酸氧化酶被广泛用于生物拆分l-氨基酸和α酮酸的生产。例如,d-氨基酸氧化酶转化头孢菌素c为戊二酰-7-氨基头孢烯酸。
草铵膦脱氢酶属于一种氨基酸脱氢酶,氨基酸脱氢酶(ec1.4.1.-,aadh)是一类能将氨基酸可逆的脱氨生成对应酮酸的酶,其反应需要核苷类辅酶的参与(nad(p)+)。根据其底物特异性,可分为氨基酸脱氢酶、亮氨酸脱氢酶、丙氨酸脱氢酶、缬氨酸脱氢酶等。因其表现出的优良催化效率与选择性,氨基酸脱氢酶被广泛的应用于天然与非天然α-氨基酸的合成。例如,li等利用亮氨酸脱氢酶制备l-叔亮氨酸,0.6m的底物能够在5.5h被完全转化,且其产物的ee值高达99%(stereoselectivesynthesisorl-tert-leucinebyanewlyclonedleucinedehydrogenasefromexiguobacteriumsibiicum[j].journalormolecularcatalysisbenzymatic,2014,105(7):11-17.)。hanson等人利用氨基酸脱氢酶制备l-6-羟基正亮氨酸,其最终产率为91~97%,ee值大于99%(enzymaticsynthesisorl-6-hydroxynorleucine.[j].bioorganic&medicinalchemistry,1999,7(10):2247-2252.)。
我们克隆了pseudomonasmooreincbi登录号为wp_090325311.1中的草铵膦脱氢酶基因和野生菌rhodotorulatoruloides中的d-氨基酸氧化酶并在大肠杆菌(escherichiacoli)中实现该基因的异源表达,d-氨基酸氧化酶能够催化d-草铵膦生成2-羰基-4-(羟基甲基氧膦基)-丁酸,草铵膦脱氢酶能够催化2-羰基-4-(羟基甲基氧膦基)-丁酸不对称胺化还原为l-草铵膦。但该草铵膦脱氢酶野生酶对2-羰基-4-(羟基甲基氧膦基)-丁酸的活力不够高,从而限制了其工业化应用。通过已报道的草铵膦脱氢酶晶体结构,利用分子模拟手段,确定该酶的空间结构和可能的与活性相关的草铵膦位点,通过定点突变技术提高了草铵膦脱氢酶对2-羰基-4-(羟基甲基氧膦基)-丁酸的催化活力,将具有较强的工业应用价值。
(三)
技术实现要素:
本发明针对现有l-草铵膦合成工艺存在的缺陷,提供了一种草铵膦脱氢酶突变体及其氧化-还原多酶偶联生产l-草铵膦的方法,以d,l-草铵膦或d-草铵膦为底物,在有氧环境及过氧化氢酶存在的情况下,利用离体的d-氨基酸氧化酶或体外表达d-氨基酸氧化酶的细胞催化d-草铵膦得到2-羰基-4-[羟基(甲基)膦酰基]丁酸;2-羰基-4-[羟基(甲基)膦酰基]丁酸再在草铵膦脱氢酶的作用下,催化生成l-草铵膦,催化剂的活性提高了近10倍,底物浓度提高了5倍,该方法原料转化率高、收率高、产物易于分离提纯。
本发明采用的技术方案:
本发明提供一种草铵膦脱氢酶突变体,所述突变体是将seqidno.2所示氨基酸序列第164位单突变位点突变获得的,更优选将第164位精氨酸突变为亮氨酸(a164g)。
本发明还提供一种所述草铵膦脱氢酶突变体在氧化-还原多酶偶联生产l-草铵膦中的应用,所述应用的方法为:以d-氨基酸氧化酶基因工程菌经诱导培养获得的湿菌体或湿菌体超声破碎提取的粗酶液以及草铵膦脱氢酶突变体基因工程菌诱导培养获得的湿菌体或湿菌体超声破碎提取的粗酶液为催化剂,以d-草铵膦为底物,加入过氧化氢酶、无机氨基供体和辅助底物,以ph值为7-8(优选7.5)的缓冲液为反应介质构成反应体系,在35℃-40℃、500-600rpm条件下反应,反应完全,将反应液分离纯化,获得l-草铵膦;所述辅助底物为葡萄糖、甲酸铵或异丙醇;所述无机氨基供体为甲酸铵或硫酸铵;所述反应体系中,催化剂的用量以湿菌体总重量计为20~100g/l(优选75g/l),其中d-氨基酸氧化酶基因工程菌湿菌体与草铵膦脱氢酶突变体基因工程菌湿菌体重量比为1-5:1(优选2:1),所述底物的初始浓度为10~500mm(优选500mm),辅助底物添加量为12-750mm(优选600mm),无机氨基供体添加量为50mm-1.5m(优选800mm);所述草铵膦脱氢酶突变体工程菌是将草铵膦脱氢酶突变体基因分别与葡萄糖脱氢酶基因、醇脱氢酶基因或甲酸脱氢酶基因中的一种共同导入宿主菌构建而成,所述葡萄糖脱氢酶基因核苷酸序列为seqidno.3所示,所述醇脱氢酶基因核苷酸序列为seqidno.4所示,所述甲酸脱氢酶基因核苷酸序列为seqidno.5所示;所述d-氨基酸氧化酶基因核苷酸序列为seqidno.6所示。
本发明反应体系中,催化剂的使用形式为细胞破碎后的粗酶液,也可以为表达重组酶的工程菌静息细胞,还可以采用纯化后的纯酶,或者是经固定化后的酶。
本发明所述草铵膦脱氢酶突变体基因工程菌湿菌体按如下方法制备:将草铵膦脱氢酶突变体基因工程菌(优选为重组大肠杆菌e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-adh、e.colibl21(de3)/petduet-1-pptgdh-fdh)接种至含有50μg/ml氨苄霉素抗性的lb液体培养基,37℃,200rpm下培养12h,再以1%(v/v)接种量接种至新鲜的含有50μg/ml氨苄霉素抗性的lb液体培养基中,于37℃,150rpm下培养至菌体od600达0.6-0.8,加入终浓度为24μg/ml的iptg,18℃下诱导培养16h后,4℃、8000rpm离心20min,弃去上清液,收集沉淀,用ph7.5、20mm磷酸盐缓冲液(pbs)洗涤两次,获得湿菌体;将湿菌体加入ph7.5、100mm的pbs中重悬,在冰水混合物上超声破碎10min,超声破碎条件:功率为400w,破碎1s,暂停5s,获得粗酶液。
本发明所述d-氨基酸氧化酶基因工程菌湿菌体按如下方法制备:将d-氨基酸氧化酶基因工程菌接种至含有50μg/ml卡那霉素的lb液体培养基中,37℃,200rpm下培养12h,再以2%(v/v)接种量接种至新鲜的含有50μg/ml卡那霉素抗性的lb液体培养基中,于37℃,150rpm下培养至菌体od600达0.6-0.8,加入终浓度为24μg/ml的iptg,28℃下诱导培养16h后,4℃、8000rpm离心20min,弃去上清液,收集沉淀,用ph7.5、20mm磷酸盐缓冲液(pbs)洗涤两次,获得湿菌体;将湿菌体加入ph7.5、100mm的pbs中重悬,在冰水混合物上超声破碎10min,超声破碎条件:功率为400w,破碎1s,暂停5s,获得粗酶液。
与现有技术相比,本发明具有以下有益效果:
(1)本发明方法以d-草铵膦为底物,在无机氨基供体、过氧化氢酶与辅酶存在时,通过d-氨基酸氧化酶催化d-草铵膦生成2-羰基-4-(羟基甲基氧膦基)-丁酸,草铵膦脱氢酶催化2-羰基-4-(羟基甲基氧膦基)-丁酸发生还原胺化反应直接制备手性纯的l-草铵膦,草铵膦脱氢酶催化活性提高了近10倍,底物浓度提高了5倍。最终草铵膦脱氢酶突变体偶联葡萄糖脱氢酶完全催化90g/l的2-羰基-4-(羟基甲基氧膦基)-丁酸生产l-草铵膦仅仅需要40分钟(转氨酶通常需要40小时),且ee值大于99%。该方法原料转化率高、收率高、产物易于分离提纯且手性纯度较高。
(2)本发明方法与转氨酶等催化工艺相比,工艺相对简单,原料转化率高,转化率可达100%,且获得的产物易于从反应液中分离提纯。
(四)附图说明
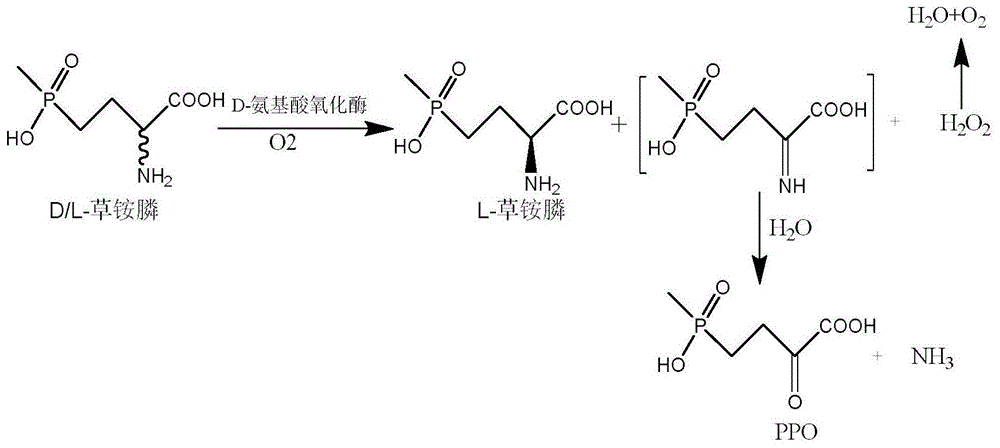
图1为d-氨基酸氧化酶催化d-草铵膦合成2-羰基-4-[羟基(甲基)膦酰基]丁酸以及氨气和过氧化氢的反应式。
图2为利用草铵膦脱氢酶偶联多酶催化2-羰基-4-(羟基甲基氧膦基)-丁酸不对称胺化还原制备l-草铵膦的反应示意图。
图3为利用d-氨基酸氧化酶、草铵膦脱氢酶与辅酶再生酶三酶耦联一锅法催化d-草铵膦制备l-草铵膦的反应示意图。
图4为草铵膦脱氢酶粗酶液sds-page图,m:标准蛋白分子量,泳道1:草铵膦脱氢酶bl21(de3)/petduet-1-pptgdh粗酶液,泳道2:草铵膦脱氢酶bl21(de3)/petduet-1-pptgdh-a164g突变体粗酶液。
图5为d-氨基酸氧化酶sds-page电泳图,其中泳道1:标准蛋白分子量;泳道2:d-氨基酸氧化酶纯酶;泳道3:d-氨基酸氧化酶粗酶液;泳道4:d-氨基酸氧化酶湿菌体。
图6为e.colibl21(de3)/pet24a-prtdaao与e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-a164g-gdh一锅法制备l-草铵膦的反应进程图。
图7为e.colibl21(de3)/pet24a-prtdaao与e.colibl21(de3)/petduet-1-pptgdh-fdh、e.colibl21(de3)/petduet-1-pptgdh-a164g-fdh一锅法制备l-草铵膦的反应进程图。
图8为e.colibl21(de3)/pet24a-prtdaao与e.colibl21(de3)/petduet-1-pptgdh-adh、e.colibl21(de3)/petduet-1-pptgdh-a164g-adh一锅法制备l-草铵膦的反应进程图。
(五)具体实施方式
以下结合具体实施例,对本发明做进一步描述。应理解,以下实施例仅用于说明本发明而非用于限制本发明的范围。
本发明中的实验方法如无特别说明均为常规方法,基因克隆操作具体可参见j.萨姆布鲁克等编的《分子克隆实验指南》。
上游基因工程操作所用试剂:本发明实施例中使用的一步克隆试剂盒均购自vazyme,南京诺唯赞生物科技有限公司;质粒提取试剂盒、dna回收纯化试剂盒购自axygen杭州有限公司;e.colibl21(de3)、质粒等购自上海生工;dnamarker、fastpfudna聚合酶、低分子量标准蛋白、琼脂糖电泳试剂、引物合成与基因测序工作由杭州擎科梓熙生物技术有限公司完成。以上试剂使用方法参考商品说明书。
下游催化工艺所用试剂:2-羰基-4-(羟基甲基膦酰基)丁酸(简称ppo),d,l-草铵膦、l-草铵膦(l-ppt)标准品购自sigma-aldrich公司;nadph购于邦泰生物工程(深圳)有限公司;其他常用试剂购自国药集团化学试剂有限公司。
实施例1表达载体和工程菌的构建
(1)重组大肠杆菌e.colibl21(de3)/petduet-1-pptgdh
通过文献报道和基因序列同源性,在ncbi数据库中挑选了1株来源于pseudomonasmoorei的草铵膦脱氢酶,ncbi登录号为wp_090325311.1,并进行全基因合成,核苷酸序列为图seqidno.1所示,氨基酸序列为seqidno.2所示。
根据seqidno.1所示核苷酸序列设计引物,并分别在引物中引入了saci和noti限制性酶切位点:
上游引物:5’-gagctcatgattgagagcgtcgagtct-3’;
下游引物:5’-gcggccgcttagacgaccccctgtgcc-3’;
以petduet-1质粒为表达载体,构建大肠杆菌e.colibl21(de3)/petduet-1-pptgdh:
表达质粒的构建:在上述引物的引发下,以pseudomonasmoorei基因组为模板,利用高保真pfudna聚合酶进行扩增,获得草铵膦脱氢酶的基因序列,测序后利用saci和noti限制性内切酶(takara)对扩增片段进行处理,并利用t4dna连接酶(takara)将该片段同用相同的限制性内切酶处理的载体petduet-1进行连接,构建表达载体petduet-1-pptgdh。
感受态细胞的制备:从-80℃冰箱中获得甘油管保藏的e.colibl21(de3)菌株,在无抗lb平板上划线,37℃培养10h,获取单菌落;挑取lb平板的单菌落,接种至含5ml的lb培养基的试管中,37℃、180rpm培养9h;从试管中取200μl菌液,接种到50ml的lb培养基中,37℃、180rpm培养od600至0.4-0.6;将菌液在冰上预冷,取菌液至灭菌的离心管中,冰上放置10min,4℃、5000rpm离心10min;将上清液倒出,注意防止染菌,用预冷的0.1mol/l的cacl2水溶液重悬沉淀细胞,并在冰上放置30min;4℃、5000rpm离心10min,弃上清,用预冷的含15%甘油的0.1mol/l的cacl2水溶液重悬沉淀细胞,取100μl重悬细胞分装至灭菌的1.5ml离心管中,保藏于-80℃冰箱,需要时取出。
重组大肠杆菌的构建:首先将储藏于-80℃的大肠杆菌bl21(de3)(invitrogen)感受态细胞在0℃冰浴10min,然后在超净台内加入5μl的质粒petduet-1-pptgdh,0℃冰浴30min,42℃水浴中热击90s,0℃冰浴2min,加入600μl的lb培养基,在37℃、200rpm摇床培养1h;涂布于含有50μg/ml氨苄霉素抗性的lb平板,37℃下培养8-12h,随机挑取克隆抽提质粒进行测序鉴定,筛选获得含有表达重组质粒的重组大肠杆菌e.colibl21(de3)/petduet-1-pptgdh。
(2)重组大肠杆菌e.colibl21(de3)/pet24a-prtdaao
将d-氨基酸氧化酶基因(ncbi登录号:alm22233.1),根据seqidno.6所示d-氨基酸氧化酶基因核苷酸序列设计引物,并分别在引物中引入了saci和noti限制性酶切位点:
上游引物:5’-gagctcatgcacagccagaagcgtgt-3’;
下游引物:5’-gcggccgcttacagtttgctttcacgcgcc-3’;
以pet-24a(+)质粒为表达载体,构建e.colibl21(de3)/pet-24a-prtdaao:
表达质粒的构建:在上述引物的引发下,以rhodotorulatoruloides基因组为模板,利用高保真pfudna聚合酶进行扩增,获得d-氨基酸氧化酶的基因序列,测序后利用saci和noti限制性内切酶(takara)对扩增片段进行处理,并利用t4dna连接酶(takara)将该片段同用相同的限制性内切酶处理的载体pet-24a(+)进行连接,构建e.colibl21(de3)/pet-24a-prtdaao。
d-氨基酸氧化酶工程菌的获得:首先将储藏于-80℃的大肠杆菌bl21(de3)(invitrogen)感受态细胞在0℃冰浴10min,然后在超净台内加入5μl的pet24a-prtdaao质粒,0℃冰浴30min,42℃水浴中热击90s,0℃冰浴2min,加入600μl的lb培养基,在37℃、200rpm摇床培养1h;涂布于含有50μg/ml卡那霉素抗性的lb平板,37℃下培养8-12h,随机挑取克隆抽提质粒进行测序鉴定,获得含有表达重组质粒的重组大肠杆菌e.colibl21(de3)/pet24a-prtdaao。
实施例2葡萄糖脱氢酶、甲酸脱氢酶、醇脱氢酶分别和草铵膦脱氢酶重组构建,获得重组大肠杆菌e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-adh、e.colibl21(de3)/petduet-1-pptgdh-fdh。
1、采用一步克隆法将葡萄糖脱氢酶(ncbi登录号:km817194.1)、甲酸脱氢酶(ncbi登录号:wp_013726924.1)、醇脱氢酶(ncbi登录号:cad66648.1)分别克隆到质粒petduet-1的第二个多克隆位点上:
带有同源序列的葡糖糖脱氢酶基因、甲酸脱氢酶基因和醇脱氢酶基因的获得:将线性化载体petduet-1开头和末端的各15-20bp序列作为同源序列,以e.colibl21(de3)/pet28b-gdh、e.colibl21(de3)/pet28b-fdh和e.colibl21(de3)/pet28b-adh(根据ncbi登录号,以全基因组为模板,构建菌株)为模板,设计引物1、引物2,引物3,引物4,引物5,引物6并将同源臂分别添加到基因特异性正/反向扩增引物序列的5’端,以此对带有同源臂的葡萄糖脱氢酶基因、甲酸脱氢酶基因和醇脱氢酶基因利用高保真pfudna聚合酶进行扩增,将消化模板后的pcr产物用dna回收纯化试剂盒进行纯化回收,并分别测出核酸浓度,得到带有同源序列的葡萄糖脱氢酶、甲酸脱氢酶和醇脱氢酶基因序列,其中葡糖糖脱氢酶核苷酸序列为seqidno.3所示,甲酸脱氢酶核苷酸序列为seqidno.4所示,醇脱氢酶核苷酸序列为seqidno.5所示。
引物1:
5’-gagatatacatggcagatctcatgggttataattctctgaaa-3’;
引物2:
5’-gtggcagcagcctaggttaattaatcaaccacggccagcctga-3’。
引物3:
5’-gagatatacatggcagatctcatgaccaaagttctggccgtg-3’
引物4:
5’-gtggcagcagcctaggttaattatttttctgcttcgccgcta-3’
引物5:
5’-gagatatacatggcagatctcatgggcatgagcaaccgtctg-3’
引物6:
5’-gtggcagcagcctaggttaatttgcgcggtatagccaccatc-3’
单片段同源重组反应:
最适克隆载体使用量={0.02*克隆载体碱基对数}ng(0.03pmol)
最适插入片段使用量={0.04*插入片段碱基对数}ng(0.06pmol)
反应体系:
注:x表示加入线性化载体的量,y表示插入片段的量。
将配置好的反应体系使用移液器轻轻吸打混匀,短暂离心后将反应液收集到管底。反应体系放在50℃的水浴中,静置5min,然后立即放在冰上冷却。将3种不同体系分别转化至大肠杆菌bl21(de3)中(42℃,90s),涂布于含有50μg/ml氨苄霉素抗性的lb平板,37℃下培养12-16h,随机挑取克隆抽提质粒进行测序鉴定,分别筛选获得含有草铵膦脱氢酶和葡萄糖脱氢酶基因、甲酸脱氢酶基因、醇脱氢酶基因的重组大肠杆菌e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-fdh和e.colibl21(de3)/petduet-1-pptgdh-adh。
实施例3:草铵膦脱氢酶-葡萄糖脱氢酶或醇脱氢酶或甲酸脱氢酶重组菌、d-氨基酸氧化酶的诱导表达
1、含草铵膦脱氢酶基因-葡萄糖脱氢酶基因或醇脱氢酶基因或甲酸脱氢酶基因的湿菌体:分别将实施例1获得的重组大肠杆菌e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-adh、e.colibl21(de3)/petduet-1-pptgdh-fdh、接种至含有50μg/ml氨苄霉素抗性的lb液体培养基,37℃,200rpm下培养12h,再以1%(v/v)接种量接种至新鲜的含有50μg/ml氨苄霉素抗性的lb液体培养基中,于37℃,150rpm下培养至菌体od600达0.6-0.8,加入终浓度为0.1mm的iptg,18℃下诱导培养16h后,4℃、8000rpm离心20min,弃去上清液,收集沉淀,用ph7.5、20mm磷酸盐缓冲液(pbs)洗涤两次,即获得含草铵膦脱氢酶-葡萄糖脱氢酶或醇脱氢酶或甲酸脱氢酶的重组菌株e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-adh、e.colibl21(de3)/petduet-1-pptgdh-fdh的湿菌体;将湿菌体加入ph7.5、100mm的pbs中重悬,在冰水混合物上超声破碎10min,超声破碎条件:功率为400w,破碎1s,暂停5s,获得粗酶液。
2、含d-氨基酸氧化酶的湿菌体:将含有d-氨基酸氧化酶基因的工程菌e.colibl21(de3)/pet24a-prtdaao接种至含有50μg/ml卡那霉素的lb液体培养基中,37℃,200rpm下培养12h,再以2%(v/v)接种量接种至新鲜的含有50μg/ml卡那霉素抗性的lb液体培养基中,于37℃,150rpm下培养至菌体od600达0.6-0.8,加入终浓度为24μg/ml的iptg,28℃下诱导培养16h后,4℃、8000rpm离心20min,弃去上清液,收集沉淀,用ph7.5、20mm磷酸盐缓冲液(pbs)洗涤两次,获得湿菌体;将湿菌体加入ph7.5、100mm的pbs中重悬,在冰水混合物上超声破碎10min,超声破碎条件:功率为400w,破碎1s,暂停5s,获得粗酶液。
粗酶液通过4℃、12000转/分钟离心20min,取上清。使用ni亲和柱(1.6×10cm,bio-rad公司,美国)纯化突变体蛋白,具体操作如下:①用5倍柱体积的结合缓冲液(含0.3mnacl的ph8.0、50mm磷酸钠缓冲液)平衡ni柱,至基线稳定;②样品上样,流速1ml/min,上样量在25-40mg/ml蛋白,使目标蛋白吸附于ni柱上;③用6倍柱体积的缓冲液a(含0.3mnacl、30mm咪唑的ph8.0、50mm磷酸钠缓冲液)冲洗杂蛋白,流速1ml/min,至基线稳定;④用缓冲液b(含0.3mnacl、500mm咪唑的ph8.0、50mm磷酸钠缓冲液)洗脱,流速1ml/min,收集目的蛋白。将目的蛋白置于ph7.5、20mm磷酸盐缓冲液中透析过夜,获得纯化酶,电泳图见图5所示。
lb培养基:胰蛋白胨10g/l、酵母提取物5g/l、氯化钠10g/l,溶剂为水,ph7.4。
lb平板:胰蛋白胨10g/l、酵母提取物5g/l、氯化钠10g/l,18g/l琼脂,溶剂为水,ph7.4。
实施例4:d-氨基酸氧化酶工程菌、草铵膦脱氢酶和葡萄糖脱氢酶gdh、甲酸脱氢酶fdh或醇脱氢酶adh工程菌细胞固定化
根据实施例3方法获得的湿菌体100g加入2l生理盐水中,充分溶解,获得细胞悬液。将细胞菌悬液中加入终浓度为20g/l的海藻酸钠,接着加入终浓度为20g/l的cacl2,并在4℃中固化12h。利用蒸馏水将小球清洗三次,加入终浓度为5g/l的戊二醛,20min后用蒸馏水清洗三次,加入终浓度为10g/l的聚乙烯亚胺,搅拌混匀20min,再次利用蒸馏水清洗三次,得到固定化细胞。
实施例5:高效液相检测方法
高效液相色谱(hplc)检测底物浓度,分析方法为:色谱柱型号:qs-c18,5μm,4.6×250mm。流动相:将50mm磷酸二氢胺溶于800ml超纯水中,加入10ml四丁基氢氧化铵(10%)用水稀释并定容至1000ml,用磷酸调ph到3.8,与乙腈以体积比88:12混合。检测波长为232nm,流速:1.0ml/min。柱温:40℃,2-羰基-4-(羟基甲基氧膦基)-丁酸出峰时间为:9.7分钟。
产物的手性分析及浓度分析通过柱前衍生化高效液相色谱进行,具体的分析方法为:
(1)色谱条件:色谱柱型号:qs-c18,5μm,4.6×250mm。流动相:50mm乙酸铵溶液:甲醇=10:1。荧光检测波长:λex=340nm,λem=455nm。流速:1ml/min。柱温:30℃,l-草铵膦出峰时间为8.5min,d-草铵膦出峰时间为10.2min。
(2)衍生化试剂:分别称取0.1g邻苯二甲醛与0.12gn-乙酰-l-半胱氨酸,用10ml乙醇助溶,再加入40ml0.1mol/l硼酸缓冲液(ph9.8),振荡使其充分溶解,4℃冰箱保存备用(不超过4天)。
(3)衍生化反应与测定:取100μl样品加入400μl衍生化试剂,在振荡器上500rpm,30℃震荡5min,再加入400μl超纯水混匀,进样10μl进行hplc分析。
实施例6:草铵膦脱氢酶基因突变文库的建立
1、定点饱和突变
以实施例2构建的e.colibl21(de3)/petduet-1-pptgdh-gdh、e.colibl21(de3)/petduet-1-pptgdh-adh、e.colibl21(de3)/petduet-1-pptgdh-fdh为出发菌株。
草铵膦脱氢酶突变体文库的制备通过1轮定点饱和突变来实现,引物设计如表2,转化,涂平板,通过筛选获得草铵膦脱氢酶优势突变体。
突变pcr体系(100μl)为:2*phantamax缓冲液25μl,dntps1μl,突变上下引物各1μl,模板(出发菌株)1μl,pfudna聚合酶0.5μl,补ddh2o至50μl。pcr条件为:95℃预变性3min,经30个循环:95℃15s,60℃15s,72℃7min20s,最后72℃终延伸10min。pcr结果分别进行dna琼脂糖凝胶电泳阳性验证,将pcr产物进行dpni酶消化模板,37℃,1小时,200转/分钟,65℃,1分钟灭活,将pcr产物热击转化,并将大肠杆菌e.colibl21(de3)活化,置于37℃、200转/分钟,培养1小时,涂布于含50μg/ml氨苄霉素抗性的lb平板上,37℃倒置培养过夜,对获得的突变体进行优势突变体的筛选,将获得优势菌株送杭州擎科生物技术有限公司进行测序确认,并保存。
表2草铵膦脱氢酶定点饱和突变引物设计
实施例7:草铵膦脱氢酶基因突变文库高通量筛选
一、草铵膦脱氢酶高通量筛选方法的建立
配制50ml工作液:邻苯二甲醛0.013g,n乙酰-l-半胱氨酸0.032g,用ph=9.8硼酸缓冲液溶解定容到50ml,作为高通量工作液。用ph=9.8硼酸缓冲液配置1mm外消旋草铵膦溶液50μl与50μl工作液震荡反应30s,再加入100μlddh2o。
二、高通量筛选
将实施例6获得的突变体文库转入大肠杆菌e.colibl21(de3)的感受态细胞中,转化条件:将pcr产物加入到感受态细胞中,冰浴30min,热击温度42℃,热击90秒,在含有50μg/ml氨苄霉素的lb抗性平板上挑取单克隆,用灭菌的牙签挑取单菌落到灭好菌的96深孔板中,每孔有1ml含有50μg/ml氨苄抗性的lb培养基,在37℃、200rpm摇床上培养8h后,每孔吸取500μl的菌液,转移到另一每孔有500μl的含有终浓度为50μg/ml氨苄和终浓度为24μg/mliptg诱导剂的lb培养基的96深孔板中,然后放在18℃、200rpm的摇床上培养16h后,离心,收取菌体于96孔板底。
1、初筛:
配置反应液:终浓度10mm底物ppo(2-羰基-4-(羟基甲基膦酰基)丁酸),50mm的硫酸铵和12mm的葡萄糖,以ph7.5的磷酸缓冲液为反应介质构成反应液。96深孔板每孔加入500μl的反应液,同时用移液枪反复吹打,将96孔板收集的菌体重悬,然后将96深孔板放入35℃、200rpm的摇床上反应4小时后,离心取上清,测荧光值,λex=340nm,λem=455nm,筛选荧光值高于原始菌株的菌株。
2、复筛:
将初筛获得的菌株的超声破碎(超声破碎条件:功率为400w,破碎1s,暂停5s)获得的粗酶液作为催化剂,以2-羰基-4-(羟基甲基氧膦基)-丁酸为底物,以葡萄糖为辅助底物,不添加外源性nadph或nadp+,运用菌体内源型nadph,建立起辅酶循环系统。反应体系选择为10ml,催化剂用量以破碎前湿菌体总重量计50g/l,底物终浓度300mm,葡萄糖终浓度450mm,硫酸铵终浓度1m,30℃、600转/分钟反应10min取样,取反应液100μl,加5μl盐酸终止反应,以超纯水补足至1ml,即反应液稀释10倍,稀释后的样品先经过衍生化处理,取200μl稀释后的反应液加400μl衍生化试剂30℃衍生化5min,再加400μl超纯水补足至1ml,12000转/分钟离心1分钟,取上清,过0.22μm微滤膜,作为液相样品,hplc检测2-羰基-4-(羟基甲基氧膦基)-丁酸、l-草铵膦、d-草铵膦及ee值。以产物l-草铵膦和ee值为指标,筛选优势突变体,实验结果示于表3。
表3pptgdh及其突变体的催化性能和立体选择性
通过表3可以看出e.colibl21(de3)/petduet-1-pptgdh-a164g-gdh、e.colibl21(de3)/petduet-1-pptgdh-a164g-adh、e.colibl21(de3)/petduet-1-pptgdh-a164g-fdh突变株的l-草铵膦生成量明显高于原始菌株,催化效率最好的是e.colibl21(de3)/petduet-1-pptgdh-a164g-gdh。
实施例8:d-氨基酸氧化酶工程菌、草铵膦脱氢酶突变体-葡萄糖脱氢酶重组工程菌e.colibl21(de3)/petduet-1-pptgdh-a164g-gdh以d-草铵膦为底物,采用一锅法制备l-草铵膦。
根据实施例3的方法,通过发酵获得d-草铵膦氧化酶湿菌体1.5g(50gdcw/l)、草铵膦脱氢酶突变体-葡萄糖脱氢酶重组工程菌e.colibl21(de3)/petduet-1-pptgdh-a164g-gdh湿菌体0.75g(25gdcw/l),过氧化氢酶(3000units/mg)0.3g,加入终浓度200mm的d-草铵膦,终浓度300mm的葡萄糖,终浓度600mm的硫酸铵构成反应体系30ml,在35℃、磁力搅拌转速为500rpm下进行反应8h,流加氨水使反应液ph维持在7.5。同样条件下,以e.colibl21(de3)/petduet-1-pptgdh-gdh为对照。通过液相方法检测反应过程中产物l-草铵膦的生成和ee值的变化,反应进程曲线如图6所示。该图显示,产物浓度随时间的推移而逐渐升高,8h内反应完成,底物转化率大于99%,产物ee值始终保持在99.5%以上。
实施例9:d-氨基酸氧化酶、草铵膦脱氢酶突变体-甲酸脱氢酶重组工程菌e.colibl21(de3)/petduet-1-pptgdh-a164g-fdh以d-草铵膦为底物,采用一锅法制备l-草铵膦。
根据实施例3的方法,通过发酵获得d-草铵膦氧化酶湿菌体1.5g(50gdcw/l)、草铵膦脱氢酶突变体-甲酸脱氢酶重组工程菌湿菌体e.colibl21(de3)/petduet-1-pptgdh-a164g-gdh0.75g(25gdcw/l),过氧化氢酶0.3g,加入终浓度200mm的d-草铵膦,终浓度600mm的甲酸铵构成反应体系30ml,在35℃、磁力搅拌转速为500rpm下进行反应10h,流加氨水使反应液ph维持在7.5。通过液相方法检测反应过程中产物l-草铵膦的生成和ee值的变化,反应进程曲线如图7所示。同样条件下,以e.colibl21(de3)/petduet-1-pptgdh-fdh为对照。该图显示,产物浓度随时间的推移而逐渐升高,10h内反应完成,底物转化率大于99%,产物ee值始终保持在99.5%以上。
实施例10:d-氨基酸氧化酶、草铵膦脱氢酶突变体-醇脱氢酶重组工程菌e.colibl21(de3)/petduet-1-pptgdh-a164g-adh以d-草铵膦为底物,采用一锅法制备l-草铵膦。
根据实施例3的方法,通过发酵获得d-草铵膦氧化酶湿菌体1.5g(50gdcw/l)、草铵膦脱氢酶突变体-醇脱氢酶重组工程菌湿菌体e.colibl21(de3)/petduet-1-pptgdh-a164g-adh0.75g(25gdcw/l),过氧化氢酶0.3g,加入终浓度500mm的d-草铵膦,终浓度750mm的异丙醇,终浓度1.5m的硫酸铵构成反应体系30ml,在35℃、磁力搅拌转速为500rpm下进行反应11h,流加氨水使反应液ph维持在7.5。通过液相方法检测反应过程中产物l-草铵膦的生成和ee值的变化,反应进程曲线如图8所示。同样条件下,以e.colibl21(de3)/petduet-1-pptgdh-adh为对照。该图显示,产物浓度随时间的推移而逐渐升高,11h内反应完成,底物转化率大于99%,产物ee值始终保持在99.5%以上。
序列表
<110>浙江工业大学
<120>一种草铵膦脱氢酶突变体及在氧化-还原多酶偶联生产l-草铵膦中的应用
<160>6
<170>siposequencelisting1.0
<210>1
<211>1338
<212>dna
<213>未知(unknown)
<400>1
atgattgagagcgtcgagtctttcttggcccgccttaaaaagcgcgaccctgaccagccg60
gagtttcatcaggcagttgaggaagtcttacgctcattatggccgttcctggaagctaac120
ccccgttatttgactagcggcattcttgaacgtatttgcgagccggaacgtgccatcgtt180
ttccgtgtgagctgggtagacgaccaaggaaaggtgcaagtgaaccgtggcttccgcatc240
cagatgaactcagctatcggcccatataaaggcgggttgcgttttcatccaagcgttaat300
ttgggtgtcttaaaattcttagcgttcgagcaaacatttaaaaacagcttaacatcgtta360
cccatgggtggaggaaagggtggtagtgacttcgacccaaaggggaagagcgatgcggaa420
gtcatgcgtttctgccaggcattcatgtcagagctttaccgtcacatcggggcggacgtc480
gatgtgccagcgggagatattggcgtgggtgcgcgcgagattggatttttattcggtcag540
tataagcgtctgtctaaccagttcacctcggtacttacgggtaagggaccgtcatatggc600
ggcagtttgattcgcccagaagctaccggatttggttgtgtttattttgccgaagaaatg660
cttaagcgccgtggagaaaccgtggaaggcaagcgtgttgccattagtggctctgggaac720
gtagcgcagtatgcggcccgcaaggtgatggatcttggcggaaaagtcatttctttatca780
gacagcgagggcacattatactgcgaatccggtttgactgaagctcaatggcaagcagtg840
ttggaactgaagaatgtacaacgtggccgtatttcagaattagccggacgctttggtctt900
gaatttttagcgggccaacgcccctggggtttatcttgcgatatcgcccttccttgcgcg960
acgcagaacgagcttgacgccgaagctgcgcgtgctttacttcgtaatggatgcacgtgc1020
gtcgctgaaggggcgaacatgccgacaacccttgaggcggttgatctgtttatcgaagcg1080
ggtattctgttcgctccaggtaaagcctcgaatgctggcggggttgcagtgtcgggttta1140
gagatgtcgcaaaacgcaatgcgtttattgtggacagggggcgaggttgactcaaaattg1200
catgctatcatgcagagcatccatcatgcttgcgtacattacggtgaagagaacggtcag1260
gtaaactacgtaaagggggcgaatattgctggattcgtgaaggttgctgatgcaatgctg1320
gcacagggggtcgtctaa1338
<210>2
<211>445
<212>prt
<213>未知(unknown)
<400>2
metilegluservalgluserpheleualaargleulyslysargasp
151015
proaspglnprogluphehisglnalavalglugluvalleuargser
202530
leutrppropheleuglualaasnproargtyrleuthrserglyile
354045
leugluargilecysgluprogluargalailevalpheargvalser
505560
trpvalaspaspglnglylysvalglnvalasnargglypheargile
65707580
glnmetasnseralaileglyprotyrlysglyglyleuargphehis
859095
proservalasnleuglyvalleulyspheleualaphegluglnthr
100105110
phelysasnserleuthrserleuprometglyglyglylysglygly
115120125
serasppheaspprolysglylysseraspalagluvalmetargphe
130135140
cysglnalaphemetsergluleutyrarghisileglyalaaspval
145150155160
aspvalproalaglyaspileglyvalglyalaarggluileglyphe
165170175
leupheglyglntyrlysargleuserasnglnphethrservalleu
180185190
thrglylysglyprosertyrglyglyserleuileargprogluala
195200205
thrglypheglycysvaltyrphealagluglumetleulysargarg
210215220
glygluthrvalgluglylysargvalalaileserglyserglyasn
225230235240
valalaglntyralaalaarglysvalmetaspleuglyglylysval
245250255
ileserleuseraspsergluglythrleutyrcysgluserglyleu
260265270
thrglualaglntrpglnalavalleugluleulysasnvalglnarg
275280285
glyargilesergluleualaglyargpheglyleuglupheleuala
290295300
glyglnargprotrpglyleusercysaspilealaleuprocysala
305310315320
thrglnasngluleuaspalaglualaalaargalaleuleuargasn
325330335
glycysthrcysvalalagluglyalaasnmetprothrthrleuglu
340345350
alavalaspleupheileglualaglyileleuphealaproglylys
355360365
alaserasnalaglyglyvalalavalserglyleuglumetsergln
370375380
asnalametargleuleutrpthrglyglygluvalaspserlysleu
385390395400
hisalailemetglnserilehishisalacysvalhistyrglyglu
405410415
gluasnglyglnvalasntyrvallysglyalaasnilealaglyphe
420425430
vallysvalalaaspalametleualaglnglyvalval
435440445
<210>3
<211>789
<212>dna
<213>未知(unknown)
<400>3
atgggttataattctctgaaaggcaaagtcgcgattgttactggtggtagcatgggcatt60
ggcgaagcgatcatccgtcgctatgcagaagaaggcatgcgcgttgttatcaactatcgt120
agccatccggaggaagccaaaaagatcgccgaagatattaaacaggcaggtggtgaagcc180
ctgaccgtccagggtgacgtttctaaagaggaagacatgatcaacctggtgaaacagact240
gttgatcacttcggtcagctggacgtctttgtgaacaacgctggcgttgagatgccttct300
ccgtcccacgaaatgtccctggaagactggcagaaagtgatcgatgttaatctgacgggt360
gcgttcctgggcgctcgtgaagctctgaaatacttcgttgaacataacgtgaaaggcaac420
attatcaatatgtctagcgtccacgaaatcatcccgtggcctactttcgtacattacgct480
gcttctaagggtggcgttaaactgatgacccagactctggctatggaatatgcaccgaaa540
ggtatccgcattaacgctatcggtccaggcgcgatcaacactccaattaatgcagaaaaa600
ttcgaggatccgaaacagcgtgcagacgtggaaagcatgatcccgatgggcaacatcggc660
aagccagaggagatttccgctgtcgcggcatggctggcttctgacgaagcgtcttacgtt720
accggcatcaccctgttcgcagatggtggcatgaccctgtacccgagctttcaggctggc780
cgtggttga789
<210>4
<211>1197
<212>dna
<213>未知(unknown)
<400>4
atgaccaaagttctggccgtgctgtatccggatccggtggatggttttccgccgaaatat60
gttcgtgatgatattccgaaaatcacccattatccggatggcagtaccgttccgaccccg120
gaaggcattgattttaaaccgggtgaactgctgggtagcgttagtggcggtctgggcctg180
aaaaaatatctggaaagtaaaggtgtggaatttgttgttaccagtgataaagaaggcccg240
gatagtgtgtttgaaaaagaactgccgaccgccgatgtggttattagtcagccgttttgg300
ccggcctatctgaccgcagatctgattgataaagcaaaaaagctgaaactggcaattacc360
gccggtattggcagcgatcatgtggatctgaatgccgccaatgaacataatattaccgtt420
gcagaagtgacctatagcaatagtgttagtgttgcagaagcagaagtgatgcagctgctg480
gccctggtgcgtaattttattccggcacatgatattgtgaaagccggtggctggaatatt540
gcagatgcagttagccgtgcctatgatctggaaggtatgaccgttggtgtgattggtgca600
ggccgcattggtcgtgccgttctggaacgtctgaaaccgtttggcgttaaactggtgtat660
aatcagcgccatcagctgccggatgaagttgaaaatgaactgggcctgacctattttccg720
gatgttcatgaaatggtgaaagttgtggatgccgttgttctggcagcaccgctgcatgca780
cagacctatcatctgtttaatgatgaagttctggccaccatgaaacgtggcgcctatatt840
gtgaataatagccgcggcgaagaagttgatcgcgatgcaattgttcgcgcactgaatagc900
ggtcagattggcggttatagtggcgatgtttggtatccgcagccggcaccgaaagatcat960
ccgtggcgtaccatgccgaatgaagcaatgaccccgcatatgagtggcaccaccctgagt1020
gcccaggcacgctatgccgcaggtgcacgtgaaattctggaagattttctggaagataaa1080
ccgattcgtccggaatatctgattgcccagggtggtagtctggccggtaccggtgccaaa1140
agttataccgtgaaaaaaggcgaagaaaccccgggtagcggcgaagcagaaaaataa1197
<210>5
<211>765
<212>dna
<213>未知(unknown)
<400>5
atgggcatgagcaaccgtctggacggcaaggtggcgatcattaccggtggcaccctgggt60
attggtctggcgattgcgaccaagttcgtggaggaaggtgcgaaagttatgatcaccggc120
cgtcacagcgacgtgggcgagaaggcggcgaaaagcgttggcaccccggaccagattcaa180
ttctttcagcacgatagcagcgacgaggatggttggaccaagctgttcgatgcgaccgaa240
aaagcgtttggcccggttagcaccctggttaacaacgcgggtattgcggtgaacaagagc300
gttgaggaaaccaccaccgcggagtggcgtaaactgctggcggtgaacctggatggtgtt360
ttctttggcacccgtctgggtatccaacgtatgaagaacaaaggtctgggcgcgagcatc420
attaacatgagcagcattgaaggtttcgttggtgacccgagcctgggtgcgtacaacgcg480
agcaagggtgcggttcgtatcatgagcaaaagcgcggcgctggattgcgcgctgaaggac540
tacgatgtgcgtgttaacaccgtgcacccgggctatattaaaaccccgctggttgacgat600
ctgccgggtgcggaggaagcgatgagccagcgtaccaagaccccgatgggtcacatcggc660
gaaccgaacgacatcgcgtacatttgcgtttatctggcgagcaacgagagcaaattcgcg720
accggtagcgaatttgtggttgatggtggctataccgcgcaataa765
<210>6
<211>1104
<212>dna
<213>未知(unknown)
<400>6
atgcacagccagaagcgtgtggttgtgctgggtagcggcgttatcggtctgagcagcgcg60
ctgattctggcgcgtaaaggctacagcgttcacatcgtggcgcgtgacctgccggaggat120
gtgagcagccagacctttgcgagcccgtgggcgggtgcgaactggaccccgtttatgagc180
ctgaccgatggtccgcgtcaagcgaagtgggaggaactgaccttcaagaaatgggttgag240
ctggtgccgaccggtcaggttatgtggctgaagggcacccgtcgttttgcgcaaaacgaa300
gacggtctgctgggccactggtacaaagatatcaccccgaactatcgtccgctgccgagc360
agcgagtgcccgccgaacagcattggtgttacctatgacaccctgagcgtgcacgcgccg420
aagtactgccagtatctggcgcgtggtctgcagaaactgggcgcgaccttcgaacgtcgt480
accgttaccagcgtggagcaggcgtttgaaggtgtggatctggttgtgaacgcgaccggt540
ctgggtgcgaagagcatcgcgggtattgacgatcaggcggcggaaccgatccgtggccaa600
accgttctggtgaagagcgcgtgcaaacgttgcaccatggacagcagcgatccgagcagc660
ccggcgtacatcattccgcgtccgggtggcgaggttatttgcggtggcacctatggtgtg720
ggcgactgggatctgagcgttaacccggaaaccgtgcaacgtatcctgaaacactgcctg780
cgtctggacccgagcattagcagcgatggtaccatcgagggcattgaagttctgcgtcat840
aacgttggcctgcgtccggcgcgtcgtggtggcccgcgtgttgaagcggaacgtctggtg900
ctgccgctggaccgtagcaagagcccgctgagcctgggtaaaggcaccacccgtgcggcg960
aaggagaaagaagttaccctggtgcacgcgtacggtttcagcagcgcgggctatcagcaa1020
agctggggtgcggcggaggatgttgcgctgctggttgaggaagcgttccaacgttaccac1080
ggcgcggcgcgtgaaagcaaactg1104
- 还没有人留言评论。精彩留言会获得点赞!