水稻粒长基因功能标记及其应用的制作方法
本发明涉及水稻育种,尤其涉及一种水稻粒长基因功能标记及其应用。
背景技术:
垩白度是影响大米外观品质的重要因素,降低垩白度是水稻育种筛选的重要指标之一,可通过增加粒长及长宽比来实现,因此将短粒品种适当变长能够有效提高大米的外观品质。此外,我国不同地区人们对大米形状有不同偏好性,如华南地区偏好长粒型大米,而江浙及东北地区偏好短圆型大米,培育不同粒长水稻品种可满足不同市场消费需求。传统育种主要采用肉眼观察筛选目标性状,具有较强的经验依赖性,且没有统一判断依据,降低了育种的筛选效率,特别是粒长等籽粒性状,由于水稻籽粒较小,肉眼很难分辨差异。而且这些性状的表现具有环境依赖性和背景依赖性,特定遗传位点的性状促进效应在某些种植条件或地点无法显现,必须通过穿梭育种才可满足准确筛选,极大地增加了育种成本。分子标记技术的发展为解决上述问题提供了有效手段,目前水稻中已经克隆了多个粒型相关基因,导致粒型变化关键序列变异已经明确,利用相关变异在水稻苗期即可实现对籽粒性状的准确筛选,排除无效单株,从而提高育种效率。测序技术可以有效鉴定不同粒型基因的关键变异,但该方法成本较高,针对少量样品进行分析尚可,无法开展大量群体的筛选,且测序技术难以有效区分杂合基因型,限制了其在分子育种改良中的应用。分子标记是将特定序列变异转变成基于pcr扩增或酶切鉴定的分子技术,而功能型分子标记则将导致性状变化的关键序列变异进行转化,能够直接对目标性状进行追踪。
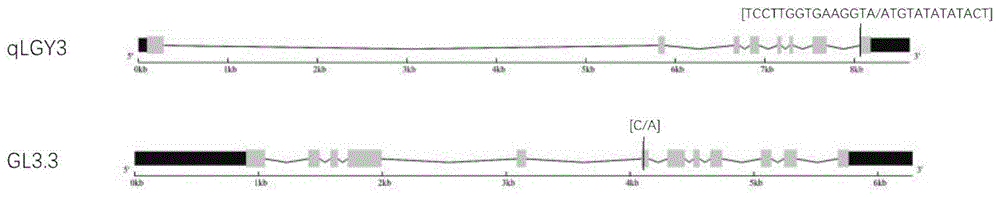
在水稻粒长基因中,gs3和gw5的效应最显著,针对这两个基因的关键变异,目前不同科研及育种机构已发展了多种不同类型的分子标记,极大地促进了育种进程。但很多水稻主栽品种已经包含了上述位点,且gw5在增加粒长的同时降低粒重,限制了其在粒长性状改良中的应用,需要挖掘更多粒长基因进行分子育种应用。近期,水稻中成功克隆了两个新的粒长基因,分别为qlgy3和gl3.3,这两个基因都位于水稻三号染色体,分别对应基因号os03g0215400和os03g0841800。如图1所示,qlgy3基因包含8个外显子,在第七内含子和第八外显子交界处,其野生型的“tccttggtgaaggta”序列被置换成“atgtatatatact”,导致该基因转录本截短,产生长粒表型,并降低垩白度,提高稻米外观品质;gl3.3基因包含12个外显子,存在多种突变类型,其中位于第六外显子672bp处的c到a碱基置换主要在aus稻亚种中检测到,与长粒表型相关,可用于现有栽培品种的粒型改良。上述两个基因目前尚没有有效的功能型分子标记产生,针对qlgy3基因,已有研究根据cdna序列差异设计引物扩增,产生扩增片段大小差异,但需要对鉴定的品种进行rna提取及反转录,费时费力。
技术实现要素:
发明目的:针对现有技术中存在的问题,本发明提供一种水稻粒长基因功能标记及应用,为水稻粒长基因qlgy3和gl3.3提供了有效的功能性分子标记。
技术方案:本发明所述的水稻粒长基因功能标记,为qlgy3功能标记或/和gl3.3功能标记,其中,
所述的qlgy3功能标记为针对第七内含子和第八外显子交界处的tccttggtgaaggta/atgtatatatact突变开发的caps标记;
所述的gl3.3功能标记为针对第六外显子672bp处的c/a碱基突变开发的caps标记。
其中,
所述qlgy3功能标记的引物为:
正向引物:5-’gggtattcttcttttcttcca-3’
反向引物:5-’tgtctgctgcttcattgct-3’;
所述gl3.3功能标记的引物为:
正向引物:5-’tgcactgccaaaatcacatattttgagccg-3’
反向引物:5-’tcaactaccactaggtccac-3’。
本发明还提供了所述的水稻粒长基因功能标记在辅助选择水稻粒型中的应用。例如选择长粒性状。
本发明还提供了一种水稻选育方法,包括:在育种过程中采用所述的功能标记辅助选择水稻粒型的性状。
具体的,在育种过程中筛选同时携带qlgy3突变型基因和gl3.3突变型基因的水稻材料,可以获得长粒性状,从而达到降低垩白度的目的。
有益效果:
针对水稻基因qlgy3和gl3.3的功能突变位点,本发明开发了有效的caps标记,可实现有效扩增。
利用本发明的分子标记,可以对粒型性状进行辅助选择,加快育种进程和目的性。
本发明发现同时携带qlgy3突变型基因和gl3.3突变型基因的水稻材料的长粒性状突出,可以利用本发明的两个功能标记在育种过程中筛选获得长粒性状。
本发明开发的caps标记,引物特异性好,所选酶都是价格较低的常用内切酶,可极大降低鉴定成本,适用于大量群体基因型分析。
附图说明
图1为qlgy3和gl3.3基因结构及关键变异示意图;
图2为qlgy3基因核心变异caps标记设计图示;
图3为gl3.3基因核心变异caps标记设计图示;
图4为qlgy3功能型标记鉴定品种凝胶电泳图;
图5为gl3.3功能型标记鉴定品种凝胶电泳图。
具体实施方式
下面结合具体实施例,进一步阐明本发明,应理解这些实施例仅用于说明本发明而不用于限制本发明的范围,在阅读了本发明之后,本领域技术人员对本发明的各种等价形式的修改均落于本申请所附权利要求所限定的范围。
为了便于进行引物设计,我们需要获得关键变异的侧翼序列。针对qlgy3基因,我们从http://www.gramene.org/网站下载了对应基因组dna序列信息,将该序列复制到lagergene软件包的editseq工具中并保存,命名为wt;随后用“tccttggtgaaggta”搜索wt序列,找到关键变异位置,将该序列用“atgtatatatact”替换,另存并命名为mut。wt和mut序列大小差异仅有2bp,如果发展成indel标记无法用琼脂糖凝胶电泳区分,无法广泛使用。我们发现“tccttggtgaaggta”中包含一个styi酶切位点,而“atgtatatatact”变异类型该酶切位点消失,且该位点上游和下游400bp序列都不包含该酶切位点,基于此,我们将该序列差异转换成caps标记,设计方法为将变异序列两侧400bp序列复制到premierprimer5.0软件中,正向引物设置在变异前半部分,反向引物设置在变异后半部分,运行后获得得分最高的一对引物作为qlgy3的功能型分子标记,获得的正向引物序列为5-’gggtattcttcttttcttcca-3’,反向引物序列为5-’tgtctgctgcttcattgct-3’。该对引物的预期扩增大小为449bp,styi酶切位点位于扩增产物第373bp处(图2)。
针对gl3.3基因,我们从水稻基因组品种多态性数据库(http://ricevarmap.ncpgr.cn/v2/)找到了第672bp处的c到a突变,对应snp编号为vg0335387952,随后下载其两侧500bp序列,用于进行引物设计。我们发现该c到a突变没有产生或消除任何酶切位点,因此无法转换成caps标记。dcaps标记是与caps标记类似的标记,方法是在引物中引入snp,对应snp与序列原有snp组合产生原有序列中不存在的酶切位点,基于这一设计原理,我们利用dcapsfinder2.0软件搜索snp附近引物序列,产生带突变碱基的引物序列,具体方法为:将包含两种snp的60bp序列分别复制到软件界面中,引物的突变碱基数目设置为1,运行后获得一系列带突变碱基的引物序列,最终我们获得了一条可以产生hpaii酶切位点的正向引物序列5-’tgcactgccaaaatcacatattttgagccg-3’,该引物将原有的t碱基置换为c,与原有snp的c(g)变异结合产生识别序列为ccgg的hpaii酶切位点(图3),hpaii为价格较低的内切酶,便于进行大量基因型鉴定,节约成本。随后设计与正向引物匹配的反向引物,方法为将正向引物及其下游300bp序列拷贝到premierprimer5.0软件,正向引物范围仍为原来的30bp区间,反向引物设置在30bp区间范围之外,运行后得到得分最高的反向引物序列5-’tcaactaccactaggtccac-3’。该对引物预期条带扩增大小为300bp,hpaii酶切位点位于正向引物第30bp处(图3)。至此,我们成功设计出两个基因的功能型分子标记。
为了验证两个标记的pcr扩增及酶切效果,我们挑选了八个品种进行了分子标记鉴定,分别为yyp1,日本晴(nip),珍汕97(zs97),稻花香(dhx),kasalath(kasa),wy3,宝大粒(bdl)和katy。苗期对8个不同水稻品种叶片取样,放入带钢珠的2ml离心管中,采用tps小量抽提法提取dna,具体步骤为:1、将叶片用球磨仪震荡破碎,加入500ultps缓冲液,65℃放置45分钟;2、12000转速离心10分钟,吸300ul上清液到新的离心管中,加入等体积异丙醇,室温放置45分钟;3、12000转速离心10分钟,获得dna沉淀,倒掉上清液,加入500ul75%乙醇;4、7500转速离心5分钟,倒掉上清液并吸干残液,室温放置30分钟后加入100ul双蒸水获得dna溶液。
随后以提取的dna为模板分别对两对引物进行pcr扩增,反应体系为20ul包含dna模板2ul、pcrbuffer(北京鼎国生物)2ul、正向引物0.5ul、反向引物0.5ul、taq聚合酶(北京鼎国生物)0.3ul及14.7ul双蒸水。pcr反应条件为:94℃变性3分钟,随后进行35个循环的“94℃变性20秒-55℃退火30秒-72℃延伸20秒”,最后72℃延伸5分钟完成反应。pcr扩增产物条带差异几乎无差异,需进一步对其酶切才可识别,酶切反应体系20ul,包括pcr产物10ul、内切酶(styi用于qlgy3基因、hpaii用于gl3.3基因,内切酶购自thermoscientific公司)0.5ul、对应buffer2ul、双蒸水7.5ul,37摄氏度酶切1小时。
pcr及酶切完成后吸取10ul进行3%浓度的琼脂糖凝胶电泳,电泳20分钟后进行拍照,比较不同品种pcr带型差异,如图4所示,qlgy3分子标记在八个品种中都能扩增出条带,且酶切后表现条带大小的多态性,其中dhx,wy3和katy三个品种pcr扩增条带酶切后大小约为449bp,说明无法被酶切,为突变类型;而剩余5个品种条带大小约为373bp,说明包含styi酶切位点,为野生型类型。如图5所示,gl3.3分子标记在八个品种中都能扩增出条带,条带大小约为270bp,可被hpaii酶切,说明八个品种都为c突变类型,所挑选品种没有a突变类型;为了进一步证明该标记的效果,我们增加了一个已知包含a突变类型的品种jhona349进行同步比较,发现该品种pcr产物无法被酶切,条带大小为300bp,与预期一致。上述结果表明两个功能型分子标记都能明确区分特定变异类型,用于准确筛选品种中的长粒型变异。
某些品种可能在引物结合位点存在变异,从而影响扩增效果,我们进一步明确了两对引物对大量品种的鉴定效果,并对长粒突变类型的分布频率和粒长效应进行了分析。我们收集了分布于世界各地的280份品种,进行田间种植,苗期取叶片提取dna,提取方法同上。随后分别用qlgy3和gl3.3功能型分子标记进行pcr扩增及酶切鉴定,最终获得不同品种的变异类型。qlgy3标记鉴定结果显示(表1),280份品种中有70份品种包含“atgtatatatact”长粒突变类型,205份品种为野生型,5份品种为杂合型,证明该标记可用于杂交稻及f2分离群体基因型鉴定;gl3.3标记鉴定结果显示(表2),280份品种中有32份品种包含长粒的a突变类型,245份品种为野生型,3份品种为杂合型,证明该标记可用于杂交稻及f2分离群体基因型鉴定;上述结果表明两个功能型标记在鉴定大量品种中也表现出良好的扩增效果,没有出现无法扩增的现象,具有广泛的适用性。上述品种可进一步被分成六个亚种,分别为香稻(aromatic)、秋稻(aus)、籼稻(ind)、温带粳稻(tej)、热带粳稻(trj)和混合类群(admix,基因组构成包含不同亚种血缘),亚种内比较发现qlgy3的长粒型突变主要集中于热带粳稻中,而在其它几个亚种中分布很少或者没有;gl3.3的长粒突变型主要集中于秋稻中,而在籼稻和粳稻亚种中很少。上述结果说明两种长粒突变类型尚未广泛用于现代籼稻和温带粳稻栽培种的育种实践中,具有较高的分子育种价值。
表1.qlgy3基因两种突变类型在280份品种中的分布频率
表2.gl3.3基因两种突变类型在280份品种中的分布频率
进一步,我们对两个功能标记的粒长区分效果进行了分析比较,考虑到不同亚种的群体结构影响,我们只在亚种内进行基因型-表型关联分析。粒型及粒重性状表现易受环境影响,因此我们在上海和海南两个完全不同的种植环境进行了性状测量,在植株完全成熟后对不同品种进行单株收种并晒干,随后采用万深sc-g自动种子考种分析及千粒重仪进行分析,自动获得粒长、粒宽、长宽比和千粒重数据,随后比较亚种内两种突变型的性状差异。具体方法为根据每个标记的两种基因型对表型数据进行分组,计算每组平均值并进行student’st-test差异检测,若p<0.05则认为该标记与对应粒型性状显著相关。qlgy3基因两种突变类型在admix亚种中表现出较为平衡的分布,野生型和突变型数目分别为35和13个,统计分析发现该亚种内两种突变类型没有产生明显的粒型变化(表3),这说明qlgy3的粒长促进效应具有遗传背景依赖性,对该位点的选择无法借助表型观察筛选,必须利用分子标记辅助选择技术进行;而gl3.3基因的两种突变类型在aus亚种中表现出平衡的分布,野生型和突变型数目分别为23和20个,统计分析显示,在两种种植条件下a突变类型都能显著促进长粒和长宽比性状产生,而对粒宽和粒重无显著影响,粒长区分效果明显(表4)。
表3.admix亚种中qlgy3基因两种突变类型与粒型表型的相关性
表4.aus亚种中gl3.3基因两种突变类型与粒型表型的相关性
基因聚合是分子设计育种的一种重要策略,本发明设计的两个功能型分子标记可以促进这一目标的实现。基于针对280个品种的单标记分析结果,我们进一步比较了两个基因的组合变异模式在群体中的分布频率。结果显示有8份品种为杂合组合类型,180份品种包含tccttggtgaaggta/c类型,即两个基因都为野生型;69份品种包含atgtatatatact/c类型,即qlgy3基因为突变型,而gl3.3基因为野生型;29份品种为tccttggtgaaggta/a类型,即qlgy3基因为野生型,而gl3.3基因为突变型;仅有两份品种为atgtatatatact/a类型,即同时包含两个基因的长粒突变型,对应品种名称分别为日本品种hiderisirazu和美国品种saber,hiderisirazu在上海和海南的粒长分别为9.43mm和9.25mm,saber在上海和海南的粒长分别为9.77mm和8.12mm,都表现出明显的长粒表型。该结果说明在今后育种中,可以上述两个品种作为供体品种,通过杂交、回交并利用本发明创造的两个标记同时筛选两种粒长变异,快速增加常规籼稻和粳稻栽培种的粒长性状,最终改良高垩白度品种大米的外观品质。
- 还没有人留言评论。精彩留言会获得点赞!