具有对抗半翅目和/或鳞翅目昆虫的活性的昆虫抑制毒素家族的制作方法
本申请是申请日为2012年4月6日、申请号为201280027913.1、发明名称为“具有对抗半翅目和/或鳞翅目昆虫的活性的昆虫抑制毒素家族”的发明专利申请的分案申请。
相关申请案
本申请要求2011年4月7日提交的美国临时申请第61/472,865号的优先权,该美国临时申请以引用的方式完整地并入本文。
序列表的并入
命名为“38-21_58195_a_pct_sequence_listing_st25.txt”的文件含有2012年4月6日创建的序列表。该文件为136kb(在mswindows中计量),以电子提交方式(使用美国专利局efs-web申请系统)同时提交,并且以引用的方式完整地并入本文。
发明领域
本发明整体涉及昆虫抑制蛋白质领域。具体地,本发明涉及表现出对抗农业上相关的作物植物和种子害虫,尤其鳞翅目和/或半翅目昆虫害虫物种的昆虫抑制活性的蛋白质。
发明背景
来源于苏云金芽孢杆菌(bacillusthuringiensis,bt)的昆虫抑制蛋白质是本领域中已知的。通过喷洒含有这些蛋白质的制剂到植物/种子上或通过使这些蛋白质在植物和种子中得到表达来将这些蛋白质用于控制农业上相关的作物植物害虫。
仅有几种bt蛋白质已被开发用于制剂中或用作由农民用于商业用途的转基因性状,用以控制鞘翅目(coleopteran)和鳞翅目(lepidopteran)害虫物种,目前还没有将bt蛋白质用于半翅目害虫物种的商业控制。某些半翅目物种,尤其草盲蝽(lygusbug)是棉花和紫花苜蓿的害虫,并且通常只能使用可持续存在且危害环境的广谱化学品,例如,硫丹(endosulfan)、乙酰甲胺磷(acephate)和草氨酰(oxamyl)来进行控制。然而,对这些数量有限的bt蛋白质的依赖可能会导致出现对这些蛋白质有抗性的新害虫,而且对广谱化学品的依赖可能对环境有害。
因此,持续需要发现和商业开发具有对抗作物植物害虫的活性的新蛋白质。
发明概述
本发明提供了经证实表现出对抗一种或多种作物植物害虫的抑制活性的昆虫抑制多肽(毒素蛋白质)的新群,即,新属。所述蛋白质中的每一种均可单独地或者彼此组合及与其它bt蛋白质和毒剂组合地用于制剂和植物中,从而为目前农业系统中使用的bt蛋白质和杀虫化学品提供替代物。
提供了表现出对抗半翅目和/或鳞翅目害虫物种的昆虫抑制活性的重组多肽,其任选地:
(a)表现出与一种或多种具有如seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22,或seqidno:24中任一个所示的氨基酸序列的蛋白质具有至少约47%至约100%的氨基酸序列同一性,或介于47%与100%之间的任何百分点的氨基酸序列同一性;
(b)表现出与一种或多种具有如seqidno:26、seqidno:136,或seqidno:138中任一个所示的氨基酸序列的蛋白质具有至少约56%至约100%的氨基酸序列同一性,或介于56%与100%之间的任何百分点的氨基酸序列同一性;
(c)在多肽内可操作位置处以连续顺序m0、m1、m2、m3、m4和m5含有六种不同基序肽区段中每一种的至少一个,每种基序肽区段表现出与为相应基序肽区段指定的共有序列具有至少约80%的同一性,其中基序肽区段m0的共有序列如seqidno:31所示,基序肽区段m1的共有序列如seqidno:48所示,基序肽区段m2的共有序列如seqidno:53所示,基序肽区段m3的共有序列如seqidno:62所示,基序肽区段m4的共有序列如seqidno:65所示,以及基序肽区段m5的共有序列如seqidno:139所示;
(d)在多肽内以可操作的连接方式含有以连续顺序的三种不同基序肽区段m1t、m2t和m4t中每一种的至少一个,其中每种基序肽区段表现出与为相应基序肽区段指定的共有序列具有至少约80%的同一性,并且其中基序肽区段m1t的共有序列如seqidno:70所示,基序肽区段m2t的共有序列如seqidno:87所示,以及基序肽区段m4t的共有序列如seqidno:120所示;
(e)含有表现出与如seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22、seqidno:24、seqidno:26、seqidno:136,或seqidno:138中任一个所示的氨基酸序列具有约195至约386个相同氨基酸的氨基酸序列;
(f)含有表现出与seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22,或seqidno:24中任一个所示的氨基酸序列具有至少约56%至约100%的同一性的氨基酸序列;
(g)含有表现出与seqidno:26、seqidno:136,和seqidno:138中任一个所示的氨基酸序列具有至少约56%至约100%的同一性,或介于56%与100%之间的任何百分点的同一性的氨基酸序列;或者
(h)由在严格杂交条件下与一个或多个如seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13、seqidno:15、seqidno:17、seqidno:19、seqidno:20、seqidno:23、seqidno:25、seqidno:135,或seqidno:137中任一个所示的核苷酸序列或其补体杂交的多核苷酸区段编码。
提供了包含前述重组多肽的昆虫抑制组合物,以及使用此类重组多肽控制鳞翅目和/或半翅目物种的方法。
提供了包含编码前述重组多肽的核苷酸序列的重组多核苷酸。还提供了包含此类重组多核苷酸的转基因植物细胞、植物,或植物部件,以及使用此类转基因植物细胞、植物,或植物部件控制鳞翅目和/或半翅目物种害虫的方法。
提供了加工的植物产品,其包含可检出量的重组多核苷酸。此类加工的产品包括但不限于植物生物质、油、粗磨粉(meal)、动物饲料、粉状物(flour)、片状物、糠、皮棉(lint)、外壳(hull)及加工的种子。
还提供了制备转基因植物的方法。此类方法包括将重组多核苷酸导入到植物细胞中,并选择表达昆虫抑制量的由重组多核苷酸编码的重组多肽的转基因植物。
根据以下详细描述、实施例和权利要求书,本发明的其它实施方案、特征和优点将是明显的。
附图简述
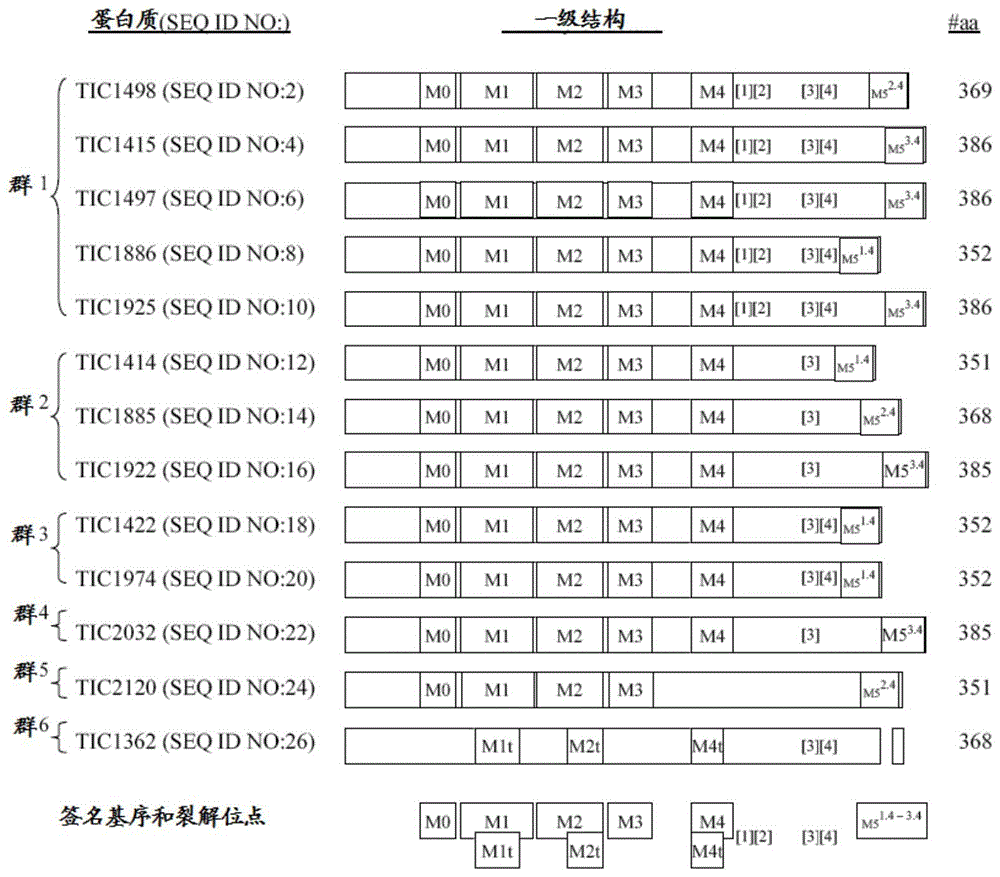
图1示出本申请中例示的蛋白质的一级结构,其示出这个属中的每种蛋白质的典型特征和基序。以名称、seqidno、结构示意图和氨基酸长度(即,氨基酸数目)描述每种蛋白质,并基于各种蛋白质之间的氨基酸同一性将它们组织成群或簇。群内的蛋白质一般表现出至少约90%的氨基酸同一性。一般而言,本文所公开的新型蛋白质属中的大多数蛋白质在多肽内可操作连键中含有六种不同签名基序(m)或基序肽区段中每一种的至少一个,每种基序都是这个蛋白质属所特有的。该基序在本文称为并且连续编号为m0、m1、m2、m3、m4和m5。m0基序邻近每种蛋白质毒素的氨基末端,而m5基序位于最接近每种蛋白质毒素的羧基末端的位置。m5基序也可以以多于一个拷贝的数量存在于每种蛋白质中。每种基序均含有该特定基序所特有的核心氨基酸区段。任何所提到的基序肽区段在来源于苏云金芽孢杆菌或相关芽孢杆菌物种的蛋白质中的存在,及对此种蛋白质的针对一个或多个半翅目和/或鳞翅目物种的毒性活性的观察,足以为将此种蛋白质归类为在本发明范围内作好准备,特别是如果所述蛋白质氨基酸序列在全部长度上表现出与本文实施的任何蛋白质具有至少约47%或更高的同一性,所述的本文实施的任何蛋白质包括如seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22,或seqidno:24所示的蛋白质。在本发明某些蛋白质中的每个一级基序m0-m5内观察到的二级基序在本文称为m1t、m2t和m4t。某些蛋白质还含有蛋白质水解裂解位点kk、eh、tf和fg,每个的相对位置在图1中示出的可适用蛋白质中分别标注为[1]、[2]、[3],[4](这两个字母代表两个支承裂解位点的氨基酸残基),这是在本发明范围内的蛋白质特征。
图2是本发明蛋白质的活性谱的文氏图(venndiagram)。圆圈[a]代表对鳞翅目有活性的蛋白质,而圆圈[b]代表对半翅目有活性的蛋白质。区域[‘c’]是圆圈[a]和[b]的交集,其代表在对抗鳞翅目和半翅目昆虫的活性的测试中呈阳性的蛋白质。区域[‘a’]等于[a]减去[‘c’],代表在对抗鳞翅目昆虫的活性的测试中呈阳性,但是在观察到鳞翅目活性时的剂量下在对抗所指示的半翅目物种的任何活性的测试中不呈阳性的蛋白质。区域[‘b’]等于[b]减去[‘c’],代表在对抗半翅目昆虫的活性的测试中呈阳性,但是在观察到半翅目物种活性时的剂量下在对抗所指示的鳞翅目物种的活性的测试中不呈阳性的蛋白质。
序列简述
seqidno:1为代表来源于苏云金芽孢杆菌(bt)物种的重组多核苷酸的核苷酸序列,其在第1-1107位核苷酸之间具有编码tic1498蛋白质的开放阅读框。
seqidno:2为tic1498蛋白质毒素的氨基酸序列。
seqidno:3为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1158位核苷酸之间具有编码tic1415蛋白质的开放阅读框。
seqidno:4为tic1415蛋白质毒素的氨基酸序列。
seqidno:5为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1158位核苷酸之间具有编码tic1497蛋白质的开放阅读框。
seqidno:6为tic1497蛋白质毒素的氨基酸序列。
seqidno:7为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1056位核苷酸之间具有编码tic1886蛋白质的开放阅读框。
seqidno:8为tic1886蛋白质毒素的氨基酸序列。
seqidno:9为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1158位核苷酸之间具有编码tic1925蛋白质的开放阅读框。
seqidno:10为tic1925蛋白质毒素的氨基酸序列。
seqidno:11为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1053位核苷酸之间具有编码tic1414蛋白质的开放阅读框。
seqidno:12为tic1414蛋白质毒素的氨基酸序列。
seqidno:13为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1104位核苷酸之间具有编码tic1885蛋白质的开放阅读框。
seqidno:14为tic1885蛋白质毒素的氨基酸序列。
seqidno:15为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1155位核苷酸之间具有编码tic1922蛋白质的开放阅读框。
seqidno:16为tic1922蛋白质毒素的氨基酸序列。
seqidno:17为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1056位核苷酸之间具有编码tic1422蛋白质的开放阅读框。
seqidno:18为tic1422蛋白质毒素的氨基酸序列。
seqidno:19为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1056位核苷酸之间具有编码tic1974蛋白质的开放阅读框。
seqidno:20为tic1974蛋白质毒素的氨基酸序列。
seqidno:21为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1155位核苷酸之间具有编码tic2032蛋白质的开放阅读框。
seqidno:22为tic2032蛋白质毒素的氨基酸序列。
seqidno:23为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1104位核苷酸之间具有编码tic2120蛋白质的开放阅读框。
seqidno:24为tic2120蛋白质毒素的氨基酸序列。
seqidno:25为代表来源于bt物种的重组多核苷酸的核苷酸序列,其在第1-1053位核苷酸之间具有编码tic1362蛋白质的开放阅读框。
seqidno:26为tic1362蛋白质毒素的氨基酸序列。
seqidno:27为编码tic1415蛋白质的人工核苷酸序列。
seqidno:28为编码tc1414蛋白质的人工核苷酸序列。
seqidno:29为编码tic1422蛋白质的人工核苷酸序列。
seqidno:30为编码tic1362蛋白质的人工核苷酸序列。
seqidno:31为m0基序区段的共有氨基酸序列。
seqidno:32-47为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定(formulating)如seqidno:31所示的共有序列。
seqidno:48为m1基序区段的共有氨基酸序列。
seqidno:49-52为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:48所示的共有序列。
seqidno:53为m2基序区段的共有氨基酸序列。
seqidno:54-61为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:53所示的共有序列。
seqidno:62为m3基序区段的共有氨基酸序列。
seqidno:63-64为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:62所示的共有序列。
seqidno:65为m4基序区段的共有氨基酸序列。
seqidno:66-69为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:65所示的共有序列。
seqidno:70为m1t基序区段的共有氨基酸序列。
seqidno:71-86为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:70所示的共有序列。
seqidno:87为m2t基序区段的共有氨基酸序列。
seqidno:88-119为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:87所示的共有序列。
seqidno:120为m4t基序区段的共有氨基酸序列。
seqidno:121-122为本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:120所示的共有序列。
seqidno:123为代表tic1497的昆虫抑制片段的氨基酸序列,并且对应于seqidno:5的第1至933位核苷酸的氨基酸翻译。
seqidno:124为代表tic1497的昆虫抑制片段的氨基酸序列,并且对应于seqidno:5的第1至885位核苷酸的氨基酸翻译。
seqidno:125为代表tic1497的昆虫抑制片段的氨基酸序列,并且对应于seqidno:5的第1至939位核苷酸的氨基酸翻译。
seqidno:126为代表tic1497的昆虫抑制片段的氨基酸序列,并且对应于seqidno:5的第1至882位核苷酸的氨基酸翻译。
seqidno:127为用于与编码本发明蛋白质的dna的5’端的(+)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:3(tic1415正向引物)的位置1..29。
seqidno:128为用于与编码本发明蛋白质的dna的3’端的(-)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:3(tic1415反向引物)的位置1131..1161。
seqidno:129为用于与编码本发明蛋白质的dna的5’端的(+)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:11(tic1414正向引物)的位置1..40。
seqidno:130为用于与编码本发明蛋白质的dna的3’端的(-)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:11(tic1414反向引物)的位置1015..1056。
seqidno:131为用于与编码本发明蛋白质的dna的5’端的(+)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:17(tic1422正向引物)的位置1..35。
seqidno:132为用于与编码本发明蛋白质的dna的3’端的(-)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:17(tic1422反向引物)的位置1021-1059。
seqidno:133为用于与编码本发明蛋白质的dna的5’端的(+)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:25(tic1362正向引物)的位置1..28。
seqidno:134为用于与编码本发明蛋白质的dna的3’端的(-)链杂交的引物中的寡核苷酸序列,并且对应于seqidno:25(tic1362反向引物)的位置1025-1056。
seqidno:135为代表来源于苏云金芽孢杆菌(bt)物种的重组多核苷酸的核苷酸序列,其在第1-1008位核苷酸之间具有编码tic2335蛋白质的开放阅读框。
seqidno:136为tic2335蛋白质毒素的氨基酸序列。
seqidno:137为代表来源于苏云金芽孢杆菌(bt)物种的多核苷酸的核苷酸序列,其在第1-1014位核苷酸之间具有编码tic2334蛋白质的开放阅读框。
seqidno:138为tic2334蛋白质毒素的氨基酸序列。
seqidno:139为m5基序区段的共有氨基酸序列。
seqidno:140-141为来自本文所公开的各种毒素蛋白质中的每一种的每个单独氨基酸序列,其用于制定如seqidno:139所示的共有序列。
seqidno:142为本发明蛋白质共享的n-末端共有序列。
seqidno:143为本发明蛋白质共享的c-末端共有序列。
具体实施方式
苏云金芽孢杆菌(bt)蛋白质是多种多样的毒素蛋白质的丰富来源;然而,鉴定新bt毒素的工艺存在许多问题。涉及bt菌株的形态学分型(例如,伴孢包涵体的结构分析、细胞外被形态学、可见的颜色,及在不同生长条件下的形态学)的筛选方法不能提供与新型毒素蛋白质的存在的良好相关性。另外,涉及鉴定具有毒性的蛋白质的高矩阵生物测定工艺的筛选方法产生不一致的结果。此类工艺包括但可能不局限于测试在bt生长和发育的各个阶段表达的蛋白质、测试不同的bt蛋白质制品、测试被各种蛋白质水解处理激活的bt蛋白质、测试与其它辅助蛋白质在一起的bt蛋白质,以及测试在各种诱导条件下的bt蛋白质。一些筛选方法依赖于需要劳动和技能密集型程序阐释结构/功能关系的结构和功能设计,并且这些方案经常仅在对完全阐释清楚的毒素进行时才有效。鉴于在寻找新bt毒素蛋白质上的固有问题,已根据生物信息学和基因组序列分析功能上的最新改进对针对编码bt毒素蛋白质的基因的筛选作出修改。
本文的发明人已利用高通量测序及生物信息学分析能力的改进,在bt基因组中筛选出编码新型蛋白质的bt毒素基因,然后克隆该基因并使其在无晶体bt菌株中表达从而产生用于昆虫抑制活性筛选的蛋白质样品。如本文所描述的及使用这种方法,已发现新型蛋白质属及表现出对抗半翅目和/或鳞翅目物种的杀虫活性的示例性蛋白质。本领域技术人员将会理解,本发明的教导使得能够鉴定或工程改造表现出本发明蛋白质的属性和特征的相关基因/蛋白质成员。
本发明多肽/蛋白质通过来源或起源(来自b.t.细菌菌株)、通过对抗半翅目和/或鳞翅目内昆虫害虫的生物毒素活性、通过一级结构(保守的氨基酸序列),以及通过长度(约300至约400个氨基酸)相互联系着。
本发明蛋白质及与本发明蛋白质类似的蛋白质可使用各种本领域中已知的基于计算机的算法进行相互比较来鉴定。本文报告的氨基酸同一性是使用以下缺省参数的clustalw比对的结果:权重矩阵:blosum,空位开放罚分:10.0,空位扩展罚分:0.05,亲水性空位:开,亲水性残基:gpsndqerk,残基特异性空位:开(thompsonetal(1994)nucleicacidsresearch,22:4673-4680)。
如果表现出对抗鳞翅目和/或半翅目昆虫物种的昆虫抑制活性的重组多肽与seqidno:2(tic1498)、seqidno:4(tic1415)、seqidno:6(tic1497)、seqidno:8(tic1886)、seqidno:10(tic1925)、seqidno:12(tic1414)、seqidno:14(tic1885)、seqidno:16(tic1922)、seqidno:18(tic1422)、seqidno:20(tic1974)、seqidno:22(tic2032)、seqidno:24(tic2120)、seqidno:26(tic1362)、seqidno:136(tic2335),和seqidno:138(tic2334)中任一个的比对得出存在至少195、196、197、198、199、200、201、202、203、204、205、206、207、208、209、210、211、212、213、214、215、216、217、218、219、220、221、222、223、224、225、226、227、228、229、230、231、232、233、234、235、236、237、238、239、240、241、242、243、244、245、246、247、248、249、250、251、252、253、254、255、256、257、258、259、260、261、262、263、264、265、266、267、268、269、270、271、272、273、274、275、276、277、278、279、280、281、282、283、284、285、286、287、288、289、290、291、292、293、294、295、296、297、298、299、300、301、302、303、304、305、306、307、308、309、310、311、312、313、314、315、316、317、318、319、320、321、322、323、324、325、326、327、328、329、330、331、332、333、334、335、336、337、338、339、340、341、342、343、344、345、346、347、348、349、350、351、352、353、354、355、356、357、358、359、360、361、362、363、364、365、366、367、368、369、370、371、372、373、374、375、376、377、378、379、380、381、382、383、384、385,或386个相同的氨基酸,则预期所述多肽在本发明的范围内。参见表1。即,在某些实施方案中,本发明重组多肽包含与seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22、seqidno:24、seqidno:26、seqidno:136,和seqidno:138相比较时显示具有195-386个相同氨基酸的氨基酸序列。
表1.本发明毒素蛋白质的配对矩阵显示。
(m)为seqidno和蛋白质名称(tic#)
(n)为seqidno。
所有对之间的氨基酸同一性百分比是相对于(m)计算的,并且由上端的数字表示。
在矩阵中,每个框中的下端数字代表各对之间相同的氨基酸数目。
灰色阴影框代表与表格中的其它蛋白质的关系较远的密切相关的毒素蛋白质。
如果表现出昆虫抑制活性的第一蛋白质与以下任何由seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22、seqidno:24中任一个所示的第二蛋白质的clustalw比对得出第一蛋白质与第二蛋白质之间具有至少约47%的氨基酸序列同一性;或具体地,第一蛋白质与第二蛋白质之间具有至少47%、48%、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.2%、99.5%、99.8%,或100%的氨基酸序列同一性,则同样预期所述第一蛋白质在本发明的发明内;或者任选地,如果表现出昆虫抑制活性的第一蛋白质与以下任何由seqidno:26、seqidno:136,或seqidno:138中任一个所示的第二蛋白质的clustalw比对得出第一蛋白质与第二蛋白质之间具有至少约56%的氨基酸序列同一性;或者,具体地,第一蛋白质与第二蛋白质之间具有至少56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.2%、99.5%、99.8%,或100%氨基酸序列同一性,则所述第一蛋白质在本发明的范围内。
观察到,本发明多肽/蛋白质通过已知只存在于这种特定昆虫抑制蛋白质家族成员中的六种签名氨基酸序列基序区段的存在而相互联系。所述签名基序区段中的每一种的相对位置在图1中示为“m0”、“m1”、“m2”、“m3”、“m4”,和“m5”。seqidno:31代表m0基序共有序列,其中x1为n或t,x2为d或a,x3为i或t,并且x4为r或s。每个m0基序区段均由seqidno:32-47所示的相应氨基酸序列表示。m0基序区段对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第48至70位氨基酸序列,及seqidno:12、seqidno:14、seqidno:16、seqidno:22,和seqidno:24的第47至69位氨基酸序列。核心氨基酸序列qx2fqtx3px4l嵌入在m0基序区段内。这种核心序列(或m0基序区段),或表现出与这种核心序列(或m0基序区段)具有至少约80%的氨基酸序列同一性的肽区段在来源于bt的特定毒素蛋白质中的单独存在或与本文所描述及可操作地定位于任何此种毒素蛋白质的一级序列内的其它基序区段的组合存在,决定了所述毒素蛋白质是本文所描述的蛋白质属的成员,特别是当该蛋白质还被证明表现出昆虫抑制性能时。
seqidno:48代表m1基序共有序列,其中x1为v或i,并且x2为r或k。每个m1基序均由seqidno:49-52所示的对应氨基酸序列表示。m1基序对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第76至118位氨基酸序列,及seqidno:12、seqidno:14、seqidno:16、seqidno:22,和seqidno:24的第75至117位氨基酸序列,或这种m1基序的核心氨基酸序列qtx1sfnex2tt的任何较短区段。这种核心序列(或m1基序),或表现出与这种核心序列(或m1基序)具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。在本文所例示的蛋白质属内的某些蛋白质包括m1基序以及二级核心基序区段mt1,所述二级核心基序区段mt1由如seqidno:70所示的共有氨基酸序列表示,其中x1为v或f,x2为s或t,x3为h或t,并且x4为v或t。每个m1t二级核心基序区段均由seqidno:71-86所示的氨基酸序列表示。m1t对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第94至112位氨基酸,seqidno:12、seqidno:14、seqidno:16、seqidno:22,和seqidno:24的第93至111位氨基酸,及seqidno:26的第83至101位氨基酸。这种二级核心基序m1t,或表现出与这种二级核心基序具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。
seqidno:53代表m2基序共有序列,其中x1为s或a,x2为v或t,并且x3为t或s。每个m2基序均由seqidno:54-61所示的对应氨基酸序列表示。m2基序对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第134至170位氨基酸,及seqidno:12、seqidno:14、seqidno:16、seqidno:22,和seqidno:24的第133至169位氨基酸。这种m2基序,或表现出与这种m2基序具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。在本文所例示的蛋白质属内的某些蛋白质包括作为m2基序的一部分的二级基序m2t,所述二级基序m2t如seqidno:87所示,其中x1为e或a,x2为g或s,x3为v或t,x4为t或s,x5为l或i。每个m2t基序均由seqidno:88–119所示的氨基酸序列表示。m2t对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第153至168位氨基酸,seqidno:12、seqidno:14、seqidno:16、seqidno:22,和seqidno:24的第152至167位氨基酸,及seqidno:26的第139至154位氨基酸。这种基序m2t,或表现出与这种基序m2t具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。
seqidno:62代表m3基序共有序列,其中x1为d或n。每个m3基序均由seqidno:63-64所示的氨基酸序列表示。m3对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第172至200位氨基酸,及seqidno:12、seqidno:14、seqidno:16、seqidno:22,和seqidno:24的第171至199位氨基酸,或包含这种m3基序的核心氨基酸序列agsvx1vpid的任何较短区段。这种m3基序或其核心,或表现出与这种m3基序或其核心序列具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。
seqidno:65代表m4基序共有序列,其中x1为p或t,并且x2为d或n。每个m4基序均由seqidno:66-69所示的氨基酸序列表示。m4对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第267至294位氨基酸,及seqidno:12、seqidno:14、seqidno:16,和seqidno:22的第266至293位氨基酸,或包含这种m4基序的核心氨基酸序列slatx1x2qils的任何较短区段。这种m4基序或其核心,或表现出与这种m4基序或其核心序列具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。在本文所例示的蛋白质属内的某些蛋白质包括作为m4基序的一部分的二级基序m4t,所述二级基序m4t如seqidno:120所示,其中x1为a或t。每个m4t基序均由氨基酸序列seqidno:121和seqidno:122表示。m4t基序对应于seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:18,和seqidno:20的第267至281位氨基酸,seqidno:12、seqidno:14、seqidno:16,和seqidno:22的第266至280位氨基酸,及seqidno:26的第254至268位氨基酸,或包含核心氨基酸序列pgftgetr的任何较短区段。这种m4t基序,或表现出与这种m4t基序具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。
seqidno:139代表m5基序共有序列区段,其中x1为c、y或r,x2为h或r,x3为n、d或h,x4为y或h,x5为r或g,并且x6为d或n。seqidno:142-143是两个示例性m5基序。m5对应于seqidno:12、14、16、22和24的第327至343位氨基酸,seqidno:2、4、6、8、10、18和20的第328至344位氨基酸,seqidno:14、16、22,和24的第344至360位氨基酸,seqidno:2、4、6和10的第345至361位氨基酸,seqidno:12、16和22的第361至377位氨基酸,seqidno:4、6和10的第362至378位氨基酸,或包含这种m5基序的核心氨基酸序列(c/y)ehnyde的任何较短区段。这种m5基序的核心氨基酸序列(c/y)ehnyde对应于seqidno:2、4、6、8、10、12、14、16、18、20、22和24中最后8个n-末端氨基酸残基中的七个。这种m5基序或其核心,或表现出与这种m5基序具有至少80%的氨基酸序列同一性的肽区段在来源于bt的特定蛋白质中的单独存在或与本文所描述的其它基序的组合存在,决定了所述蛋白质是本文所描述的蛋白质属的成员,特别是当所述蛋白质还被证明表现出昆虫抑制性能时。本发明多肽/蛋白质通过这种m5基序而相互联系着,这种m5基序可出现一次,以seqidno:8、12、18和20为例;可重复出现二次,以seqidno:2、14和24为例;并且可重复出现三次,以seqidno:4、6、10、12、16和22为例。有趣地,tic1414与tic1922可以以相对于tic1414的100%同一性比对(表1),这可能是因为tic1414与tic1922之间的差异是两个重复的m5基序(配对比对中允许的空位)。
本发明提供了重组多肽,其包含表现出与选自由seqidno:31(基序m0)、seqidno:48(基序m1)、seqidno:53(基序m2)、seqidno:62(基序m3)、seqidno:65(基序m4)、seqidno:141(基序m5)、seqidno:70(基序m1t)、seqidno:87(基序m2t)、seqidno:120(基序m4t)及其任何组合组成的组的氨基酸序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%,或100%的氨基酸序列同一性的肽区段。此种多肽表现出对抗鳞翅目和/或半翅目物种的昆虫抑制活性。如本文所使用的术语“昆虫抑制活性”是指当被提供在害虫的食物中并被靶标(预期)害虫摄入时在抑制害虫,优选一种或多种作物植物的害虫上有效的蛋白质或其片段的活性。作物植物的害虫包括线虫和节肢动物,包括昆虫。本发明蛋白质在抑制特定靶标害虫,特别是昆虫害虫的生长、发育、生活力或繁殖力上有效,所述昆虫害虫包括但不限于鳞翅目和半翅目的昆虫。
在某些实施方案中,昆虫抑制多肽/蛋白质含有表现出与本发明蛋白质的任一种或多种签名基序(seqidno:31-122、142和143)具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性的氨基酸区段。
在某些实施方案中,重组多肽包含如seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22、seqidno:24、seqidno:26、seqidno:136,或seqidno:138所示的氨基酸序列,或其昆虫抑制片段。示例性昆虫抑制片段包括但不限于包含如seqidno:123、seqidno:124、seqidno:125,和seqidno:126所示的氨基酸序列的那些。
另外的签名基序包括蛋白水解裂解位点“kk”、“eh”、“tf”和“fg”,每个的相对位置分别在图1中示为[1]、[2]、[3]和[4]。
另外的签名基序包括如seqidno:142所示的n-末端共有序列,其中x1为a或e,x2为n或d,x3为q或e,并且x4为s或l,所述的n-末端共有序列是作为本文例示的蛋白质属的成员的蛋白质所共享的,但具有如seqidno:26所示的氨基酸序列的蛋白质除外。正向寡核苷酸引物,例如,seqidno:127、seqidno:129、seqidno:131和seqidno:133可被设计成与为本发明蛋白质的n-末端共有序列编码的dna序列的正链杂交。如seqidno:143所示的其中x1为h或e并且x2为n或y的c-末端共有序列是本发明蛋白质共享的签名基序。反向寡核苷酸引物,例如,seqidno:128、seqidno:130、seqidno:132和seqidno:134,可被设计成为本发明蛋白质的c-末端共有序列编码的dna序列的负链杂交。寡核苷酸引物可被设计成与本发明蛋白质的任一种或多种签名基序(seqidno:31-122、140和141)的正链或负链杂交。
当组合时,正向和反向引物可用于扩增编码本发明蛋白质(或其片段)的核苷酸序列。
使用文氏图(图2)连同证明毒性活性的生物测定证据,用共同功能和针对半翅目和/或鳞翅目昆虫物种的杀虫活性阐释了本发明蛋白质的关系。表2用昆虫物种将图2文氏图中示出的蛋白质与昆虫抑制活性联系起来。从表2汇编的结果更详细地描述在实施例中。
在某些实施方案中,害虫具体地为昆虫害虫。昆虫害虫包括选自以下目的昆虫:鞘翅目、双翅目、膜翅目(hymenoptera)、鳞翅目、食毛目(mallophaga)、同翅目(homoptera)、半翅目、蜚蠊目(blattodea)、直翅目(orthoptera)、缨翅目(thysanoptera)、革翅目(dermaptera)、等翅目(isoptera)、虱目(anoplura)、蚤目(siphonaptera),及毛翅目(trichoptera)。
昆虫可包括鳞翅目幼虫,例如但不限于夜蛾科的粘虫、地老虎、尺蠖,及棉铃虫类(例如,草地贪夜蛾(fallarmyworm,spodopterafrugiperda)、甜菜夜蛾(beetarmyworm,spodopteraexigua)、蓓带夜蛾(berthaarmyworm,mamestraconfigurata)、小地老虎(blackcutworm,agrotisipsilon)、粉纹夜蛾(cabbagelooper,trichoplusiani)、大豆夜蛾(soybeanlooper,pseudoplusiaincludens)、藜豆夜蛾(velvetbeancaterpillar,anticarsiagemmatalis)、苜蓿绿夜蛾(greencloverworm,hypenascabra)、烟芽夜蛾(tobaccobudworm,heliothisvirescens))、颗粒地老虎(granulatecutworm,agrotissubterranea)、一星粘虫(armyworm,pseudaletiaunipuncta)、西部地老虎(westerncutworm,agrotisorthogonia);螟蛾科的螟虫、鞘蛾、结网毛虫、锥虫(coneworm)、菜青虫和雕叶虫(例如,欧洲玉米螟(europeancornborer,ostrinianubilalis)、脐橙螟(navelorangeworm,amyeloistransitella)、玉米根结网毛虫(cornrootwebworm,crambuscaliginosellus)、草地结网毛虫(sodwebworm,herpetogrammalicarsisalis)、日葵螟(sunflowermoth,homoeosomaelectellum)、小玉米茎蛀虫(lessercornstalkborer,elasmopalpuslignosellus);卷蛾科的卷叶虫(leafroller)、蚜虫(budworm)、种子蠕虫(seedworm),和果实蠕虫(例如,苹果蠹蛾(codlingmoth,cydiapomonella)、葡萄果实蛾(grapeberrymoth,endopizaviteana)、东方果蛾(orientalfruitmoth,grapholitamolesta)、向日葵芽蛾(sunflowerbudmoth,suleimahelianthana);及许多其它经济上重要的鳞翅目物种(例如,小菜蛾(diamondbackmoth,plutellaxylostella)、红铃虫(pinkbollworm,pectinophoragossypiella)、舞毒蛾(gypsymoth,lymantriadispar)。鳞翅目的其它昆虫害虫包括,例如,棉叶虫(alabamaargillacea,cottonleafworm)、果树卷叶蛾(archipsargyrospila,fruittreeleafroller)、欧洲卷叶蛾(a.rosana,europeanleafroller)和其它黄卷蛾属(archips)物种、水稻二化螟(chilosuppressalis,asiaticriceborer或ricestemborer)、水稻卷叶蛾(cnaphalocrocismedinalis,riceleafroller)、玉米根结网毛虫(crambuscaliginosellus,cornrootwebworm)、蓝草结网毛虫(c.teterrellus,bluegrasswebworm)、西南玉米螟(diatraeagrandiosella,southwesterncornborer)、甘蔗螟(d.saccharalis,surgarcaneborer)、埃及金钢钻(eariasinsulana,spinybollworm)、翠纹金钢钻(e.vittella,spottedbollworm)、美洲棉铃虫(helicoverpaarmigera,americanbollworm)、玉米穗虫(h.zea,cornearworm或cottonbollworm)、烟芽夜蛾(heliothisvirescens,tobaccobudworm)、草地结网毛虫(herpetogrammalicarsisalis,sodwebworm)、欧洲葡萄蛾(lobesiabotrana,europeangrapevinemoth)、红铃虫(pectinophoragossypiella,pinkbollworm)、柑橘潜叶蛾(phyllocnistiscitrella,citrusleafminer)、大菜粉蝶(pierisbrassicae,largewhitebutterfly)、小菜粉蝶(p.rapae,importedcabbageworm或smallwhitebutterfly)、小菜蛾(plutellaxylostella,diamondbackmoth)、甜菜夜蛾(spodopteraexigua,beetarmyworm)、斜纹夜蛾(s.litura,tobaccocutworm,clustercaterpillar)、草地贪夜蛾(s.frugiperda,fallarmyworm)),及番茄斑潜蝇(tutaabsoluta,tomatoleafminer)。
昆虫可包括半翅目和同翅目的成虫和若虫,例如但不限于盲蝽科(miridae)的盲蝽,蝉科(cicadidae)的蝉,叶蝉科(cicadellidae)的叶蝉(例如,小绿叶蝉属某些种(empoascaspp.)),蜡蝉总科(fulgoroidea)和飞虱科(delphacidae)的飞虱(planthopper),角蝉科(membracidae)的角蝉,木虱科(psyllidae)的木虱,粉虱科(aleyrodidae)的粉虱,蚜科(aphididae)的蚜虫,根瘤蚜科(phylloxeridae)的根瘤蚜,粉蚧科(pseudococcidae)的粉介壳虫,蚧科(coccidae)、盾蚧科(diaspididae)和硕蚧科(margarodidae)的介壳虫,纲蝽科(tingidae)的纲蝽,蝽科(pentatomidae)的椿象,长蝽科(lygaeidae)的麦长蝽(cinchbug)(例如,土长蝽属某些种(blissusspp.))和其它长蝽,沫蝉科(cercopidae)的沫蝉,缘蝽科(coreidae)的南瓜缘蝽(squashbug),以及红蝽科(pyrrhocoridae)的红蝽和棉红蝽。半翅目的其它害虫包括:绿蝽象(acrosternumhilare,greenstinkbug)、南瓜缘蝽(anasatristis)、麦长蝽(blissusleucopterusleucopterus)、棉网蝽(corythucagossypii,cottonlacebug)、cyrtopeltismodesta(tomatobug)、棉红蝽(dysdercussuturellus))、褐臭蝽(euschistusservus,brownstinkbug)、一斑臭蝽(euschistusvariolarius,one-spottedstinkbug)、长蝽复合物(graptostethus某些种,complexofseedbugs)、松籽喙缘蝽(leptoglossuscorculus,leaf-footedpineseedbug)、牧草盲蝽(lyguslineolaris,tarnishedplantbug)、豆荚草盲蝽(lygushesperus,westerntarnishplantbug)、南方绿蝽(nezaraviridula,southerngreenstinkbug)、稻臭蝽(oebaluspugnax,ricestinkbug)、大马利筋长蝽(oncopeltusfasciatus,largemilkweedbug),及棉跳盲蝽(pseudatomoscelisseriatus,cottonfleahopper)。
在某些实施方案中,本发明重组多肽表现出对抗选自由以下组成的组的鳞翅目物种的昆虫抑制活性:玉米穗虫、欧洲玉米螟、甘蔗螟、西南玉米螟、藜豆夜蛾、草地贪夜蛾、甜菜夜蛾、小地老虎、粉纹夜蛾、大豆夜蛾、烟芽夜蛾、小菜蛾、红铃虫、美洲棉铃虫、小玉米茎蛀虫和柑橘潜叶蛾,和/或对抗选自由以下组成的组的半翅目物种的昆虫抑制活性:豆荚草盲蝽、牧草盲蝽、绿蝽象、褐臭蝽、南方绿蝽、桃蚜(m.persicae)、大豆蚜(a.glycines)和棉蚜(a.gossypii)。
本发明蛋白质代表与本领域中已知的任何其它bt蛋白质表现出的氨基酸同一性不大于56%的cry蛋白质的新类别和类。表现出与本发明任何蛋白质具有最接近的同一性的蛋白质为cry15aa1(gi:142726,accession:aaa22333)(brown和whiteley,journalofbacteriology,jan.1992,p.549-557,vol.174,no.2)。使用clustalw对cry15aa1与本发明中例示的每种蛋白质进行比对,结果在表3中给出。
表3.蛋白质与cry15aa1的比对。
*在clustalw比对中
cry15aa1不含本发明蛋白质所共享的任何签名基序(seqidno:31-122、140和141)。cry15aa1没有显示出如图1中所示的本发明蛋白质所共享的蛋白水解裂解位点[2]、[3]和[4]。cry15aa1显示出约7.3pi的计算等电点,相比之下,本发明蛋白质各自显示出约5至6pi的计算等电点。cry15aa1在中性ph下仅显示出3个正电荷,而本发明蛋白质在中性ph下显示出3至10个负电荷。
本发明蛋白质可用于制备与这种蛋白质属特异性结合的抗体,并且可用于筛选及寻找该属的其它成员。
可使用编码这些蛋白质的核苷酸序列作为探针和引物使用热扩增或等温扩增和/或杂交方法筛选鉴定该属的其它成员,例如,如seqidno:127-134所示的寡核苷酸,及与编码本发明签名基序的序列杂交的寡核苷酸。核苷酸序列同源物,即,由与本文所公开的每个或任何序列在严格杂交条件下杂交的核苷酸序列编码的杀虫蛋白质特别意欲包括在本发明的范围内。本发明还提供了检测与第二核苷酸序列杂交的第一核苷酸序列的方法,其中第一核苷酸序列编码杀虫蛋白质或其杀虫片段,并且与第二核苷酸序列在严格杂交条件下杂交。在此种情况下,第二核苷酸序列可为在严格杂交条件下的本文所公开的任何序列。核苷酸编码序列彼此在适当杂交条件下杂交,并且由这些核苷酸序列编码的蛋白质与针对任一种其它蛋白质产生的抗血清交叉反应。如本文所定义的严格杂交条件至少包括在42℃下杂交,接着在室温下用2xssc、0.1%sds洗涤两次,每次五分钟,再接着在65℃下用0.5xssc、0.1%sds洗涤两次,每次三十分钟。当然,本领域技术人员将会认识到,由于遗传密码的冗余,许多其它序列也能够编码此类相关蛋白质,那些序列如果在芽孢杆菌菌株中或在植物细胞中起到表达杀虫蛋白质的作用,则意欲包括在本发明内,当然认识到许多此类冗余编码序列在这些条件下不能与编码tic1498、tic1415、tic1497、tic1886、tic1925、tic1414、tic1885、tic1922、tic1422、tic1974、tic2032、tic2120、tic1362、tic2335,和tic2334的天然bt序列杂交。
在某些实施方案中,表现出对抗鳞翅目和/或半翅目昆虫物种的昆虫抑制活性的重组多肽在本发明的范围内,所述多肽由在严格杂交条件下与一个或多个如seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13、seqidno:15、seqidno:17、seqidno:19、seqidno:21、seqidno:23、seqidno:25、seqidno:135或seqidno:137中任一个所示的核苷酸序列或其补体杂交的多核苷酸区段编码。
本发明方面提供了发现相关蛋白质的方法,此类方法包括对bt基因组测序、汇编序列数据、鉴定和克隆编码此类昆虫抑制蛋白质的bt基因,及表达和测试新bt蛋白质以测定昆虫抑制活性。本发明另一方面采用分子方法工程改造和克隆商业上有用的蛋白质,包括来自昆虫抑制蛋白质类的蛋白质和改进变体的嵌合体,例如,可由来自介于签名基序之间的间隙内的各种蛋白质中的每一种的区段组装嵌合体以获得改进的实施方案。可将本发明蛋白质彼此之间或与其它bt昆虫抑制蛋白进行比对,并且可从每种此种蛋白质中鉴定出可用于在比对的蛋白质之间进行替换从而导致嵌合蛋白质的构建的区段。可对此类嵌合蛋白质进行害虫生物测定分析,并进行表征以确定与嵌合体中每个此种区段来源于其的母体蛋白质相比较是否存在生物活性增加或靶标害虫谱扩大的情况。可通过与其它蛋白质交换结构域或区段,对所述多肽的昆虫抑制活性进行进一步工程改造以获得针对特定害虫或更广谱的害虫的改进活性。
技术人员理解氨基酸替换的概念,并且会认识到这需要非常规的实验,因为存在可接受替换而不会对蛋白质结构或功能产生明显影响的氨基酸位置;然而,在意外情况下,甚至保守性替换也可能会被判定为显著改变蛋白质的结构或功能,而且将接受此类变化的氨基酸区段中的准确位置往往是未知的。因此,可通过丙氨酸扫描诱变鉴定沿着蛋白质序列长度的影响功能的位置处的氨基酸替换,此类位置经常可用作氨基酸插入和/或缺失,或n-或c-末端缺失的点。因此,本发明蛋白质包括由本发明氨基酸序列表示的蛋白质的功能等效片段(n-或c-末端缺失)。tic1497和tic1922的n-末端蛋白质片段(seqidno:123-126,16)已展现出昆虫抑制活性(分别见表2及实施例6、10和11)。考虑了该属任何成员的相应n-末端蛋白质片段。
与本发明蛋白质在功能上等效(具有基本上等效的昆虫抑制活性)的蛋白质包括在本发明蛋白质序列中具有保守性氨基酸替换的蛋白质。在此类氨基酸序列中,起始序列中的一个或多个氨基酸被另外的一个(多个)与天然氨基酸有相似电荷和极性的氨基酸替换,即,如本文例示的保守性氨基酸替换,所述替换从电荷和极性角度来看导致保守性变化,但是其可能会导致蛋白质的生物活性发生变化,优选与此类位置上具有原始氨基酸的起始蛋白质相比较增加蛋白质的活性,或者导致变体蛋白质的生物活性谱发生变化而不发生昆虫抑制活性的任何损失。可接受替换的氨基酸或末端缺失来获得生物等效物的蛋白质的实例包括但不限于如seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:10、seqidno:12、seqidno:14、seqidno:16、seqidno:18、seqidno:20、seqidno:22、seqidno:24、seqidno:26和seqidno:123-126中任一个所示的蛋白质序列。
考虑了在植物中或通过包括在用于表达/产生本发明重组多肽/蛋白质的条件下培养重组bt细胞的工艺富集本发明蛋白质。此种工艺可包括通过干燥、冷冻干燥、均质化、提取、过滤、离心、沉降或浓缩表达/产生所述重组多肽的重组苏云金芽孢杆菌细胞的培养物制备。此种工艺可产生bt细胞提取物、细胞悬浮液、细胞匀浆液、细胞裂解液、细胞上清液、细胞滤液,或细胞沉淀物。通过获得如此制得的重组多肽/蛋白质,包括重组多肽/蛋白质的组合物可包括细菌细胞、细菌孢子和伴孢包涵体,并且可调配用于各种用途,包括农业昆虫抑制喷雾产品或在食物生物测定中用作昆虫抑制制剂。
包含前述重组多肽/蛋白质的昆虫抑制组合物/制剂意欲在本发明的范围内。在某些实施方案中,此种组合物可进一步包含至少一种表现出对抗相同鳞翅目和/或半翅目昆虫物种的昆虫抑制活性但是不同于所述重组多肽的杀虫剂。此种杀虫剂选自由以下组成的组:昆虫抑制蛋白质、昆虫抑制dsrna分子,和辅助蛋白质。此类杀虫剂的实例包括但不限于tic807蛋白质、tic853蛋白质、axmi-171蛋白质,和cry51aa1蛋白质。还考虑了将其它组合物与本发明蛋白质及与上文所提供的蛋白质组合进行组合。例如,设计用于控制也受本发明蛋白质控制的害虫的局部施用的杀虫化学品可与本发明蛋白质一起用于种子处理及用于可直接施用(土壤浇灌)至土壤的喷涂式/滴注式/或涂抹式制剂中,可施用至表达本发明蛋白质的生长中的植物,或者可调配用于施用至含有一个或多个编码本发明的一种或多种蛋白质的转基因的种子。此类用于种子处理的制剂可与本领域中已知的各种粘着剂和增粘剂一起施用。此类制剂可含有与本发明蛋白质在作用模式上协同的杀虫剂,也即该制剂杀虫剂通过不同的作用模式起到控制与受本发明蛋白质控制的害虫相同或相似的害虫的作用,或者此类杀虫剂起到控制较广泛宿主范围内的害虫的作用,所述较广泛宿主范围内的害虫例如鳞翅目或半翅目物种或其它不能被本发明蛋白质有效控制的植物害虫物种如鞘翅目物种。
前述组合物/制剂可进一步包含农业上可接受的载体,例如毒饵(bait)、粉末、粉尘、团粒、细粒、喷雾剂、乳状液、胶体悬浮液、水溶液、芽孢杆菌孢子/结晶制品、种子处理剂、被转化成表达一种或多种所述蛋白质的重组植物细胞/植物组织/种子/植物,或被转化成表达一种或多种所述蛋白质的细菌。根据重组多肽固有的昆虫抑制或杀虫抑制水平及待应用至植物或食物测定中的制剂水平,所述组合物/制剂可包括各种按重量计的量的重组多肽,例如从0.0001重量%至0.001重量%至0.01重量%至1重量%至99重量%的重组多肽。
本发明蛋白质可被组合成以下制剂:用于局部施用至植物表面、土壤的制剂、用于种子处理的制剂、含有对半翅目和鳞翅目物种的靶标害虫有毒性的其它试剂的制剂。此类试剂包括但不限于tic807蛋白质、tic853蛋白质、axmi-171蛋白质,和cry51aa1蛋白质,它们都在控制与受本发明昆虫抑制蛋白质控制的半翅目害虫相同的半翅目害虫上有效。
控制鳞翅目和/或半翅目物种害虫的方法也意欲在本发明的范围内。此种方法包括使害虫与昆虫抑制量的重组多肽/蛋白质接触的步骤。在某些实施方案中,鳞翅目和半翅目物种害虫在作物田地中。
本发明实施方案包括编码所述属的昆虫抑制蛋白质成员的重组多核苷酸。就“重组”多核苷酸而言,多核苷酸分子意欲由人工方式或人工干预通过分子生物学工程技术制备,所述分子生物学工程技术可包括在将此类分子导入到宿主细胞中后扩增或复制此类分子,以及随后从代表性宿主细胞中提取和/或纯化出多核苷酸。本发明多核苷酸实施方案包括核糖核酸(rna)和脱氧核糖核酸(dna)。本发明蛋白质可从这样的dna构建体表达,在所述dna构建体中,编码所述蛋白质的开放阅读框可操作地连接至诸如启动子和在所述构建体预期用于其中的特定系统中起表达作用的任何其它调控元件的元件。例如,植物可表达的启动子可以可操作地连接至用于在植物中表达蛋白质的蛋白质编码序列,而bt可表达的启动子可以可操作地连接至用于在bt中表达蛋白质的蛋白质编码序列。可以可操作地连接至蛋白质编码序列的其它有用元件包括但不限于增强子、内含子、蛋白质固定标签(his-标签)、翻译后修饰酶的靶标位点、dsrna编码区段、sirna、mirna、核糖体结合位点、前导元件和mirna靶标位点。
由此提供的示例性重组多核苷酸分子包括但不限于seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:9、seqidno:11、seqidno:13、seqidno:15、seqidno:17、seqidno:19、seqidno:21、seqidno:23、seqidno:25、seqidno:27、seqidno:28、seqidno:29、seqidno:30和seqidno:135。
本发明的方面提供了包括一种或多种前述多核苷酸的重组dna构建体,其可另外用可转录的区域或不可转录的区域或二者工程改造。可操作地组装此类区域以通过体内或体外系统促进dna表达成rna,从而产生本发明的新型rna转录本实施方案。本发明以rna转录本为特征,所述rna转录本包括但不限于编码蛋白质的rna和另外的rna区域,所述另外的rna区域为可翻译的或不可翻译的rna区域或二者。此类另外的rna区域包括经工程改造成翻译至末端或肽内区域的可翻译区域,及经工程改造成促进转录、翻译或二者的不可翻译的区域。
在某些实施方案中,前述重组dna构建体位于用在大肠杆菌或bt表达系统中的表达盒中。表达盒通常被设计成在表达盒的5’端在编码本发明蛋白质的所需多核苷酸区段上游具有启动子。启动子可由多个不同的启动子元件组成,这些不同的启动子元件可操作地连接从而为编码本发明蛋白质的序列的转录的启动作好准备。由启动子–蛋白质-编码dna组成的dna序列可在其3’端可操作地连接至在大肠杆菌和/或bt细胞中起作用的转录终止信号序列从而产生重组dna构建体。
在某些实施方案中,前述重组dna构建体位于用于在植物中表达的表达盒中。表达盒被设计成在表达盒的5’端在编码本发明蛋白质的所需多核苷酸区段上游具有启动子。5’未转录的dna可包含可由多个不同启动子和增强子元件组成的启动子,所述的多个不同启动子和增强子元件可操作地连接从而为包括编码本发明多肽的序列的下游序列的转录的启动作好准备。可将一个或多个转录但未翻译的dna序列可操作地连接在包括前导和/或内含子序列的表达盒中的启动子的3’端。任选地将内含子序列提供在前导序列的3’端或者在一些情况下提供在编码所需蛋白质的开放阅读框内。可将编码任选易位多肽(例如信号肽或叶绿体转运肽)的多核苷酸区段插入到本发明蛋白质的编码序列的5’端,以便将本发明蛋白质定位至特定亚细胞位置。任选地将编码本发明蛋白质的核苷酸序列,连同在植物细胞中起作用的任何必要的可操作地连接的多聚腺苷酸化(多聚a)和/或转录终止序列一起可操作地定位在前述表达盒内。连续地布置前述元件,并且前述元件可根据所需的表达效果以各种组合形式使用。
本发明以在植物中起作用的启动子为特征,所述在植物中起作用的启动子包括但不限于组成型启动子、非组成型启动子、空间特异性启动子、时间特异性启动子、组织特异性启动子、发育特异性启动子、诱导型启动子和病毒启动子。在植物中起作用的启动子的实例包括玉米蔗糖合成酶1启动子、玉米乙醇脱氢酶1启动子、玉米集光复合体启动子、玉米热激蛋白质启动子、豌豆小亚基rubp羧化酶启动子、ti质粒甘露氨酸合成酶启动子、ti质粒胭脂氨酸合成酶启动子、矮牵牛查尔酮异构酶启动子、菜豆富甘氨酸蛋白质1启动子、马铃薯糖蛋白启动子、凝集素启动子、camv35s启动子、s-e9小亚基rubp羧化酶启动子、香石竹蚀环病毒启动子,及大丽花花叶病毒启动子。
包含蛋白质编码序列的重组dna构建体也可进一步包含编码一种或多种昆虫抑制剂的dna区域,该dna区域可被构造成与编码本发明蛋白质、不同于前述蛋白质的蛋白质、昆虫抑制dsrna分子或辅助蛋白质的dna序列同时表达或共表达。辅助蛋白质包括辅因子、酶、结合伴侣,或其它昆虫抑制剂,其通过辅助昆虫抑制剂的表达、影响它在植物中的稳定性、优化用于寡聚化的自由能、增强它的毒性及增大它的活性谱而协同地起到辅助提高它的有效性的作用。
可通过载体将包含蛋白质编码序列的重组多核苷酸或重组dna构建体递送至宿主细胞。将重组dna构建体转移进和转移出宿主细胞的方法是本领域中已知的,所述宿主细胞包括大肠杆菌、苏云金芽孢杆菌和农杆菌属物种。此类载体被设计成促进载体dna的摄取以及进一步在体外或体内系统中提供dna到rna到蛋白质的瞬时或稳定表达。载体的实例包括但不限于质粒、杆状病毒、人工染色体、病毒体、粘粒、噬菌粒、噬菌体或病毒载体。此类载体可用来在宿主细胞中获得蛋白质编码序列的稳定或瞬时表达,并且如果这样的话,随后获得多肽的表达。包含蛋白质编码序列并且被导入到宿主细胞中的外源重组多核苷酸或重组dna构建体在本文也还称为“转基因”。
质粒可被设计成在大肠杆菌或苏云金芽孢杆菌或二者中复制。此类质粒含有允许此类质粒在任一物种中的复制和保持及允许转基因,例如,前述重组dna构建体在任一物种中的表达的遗传元件。
植物转化载体可被设计成允许农杆菌介导的包含前述重组dna构建体的t-dna,即转移的dna的转移。此类植物转化载体含有允许此类质粒载体在大肠杆菌和/或农杆菌中的复制和保持并且是t-dna转移到植物基因组中所必需的遗传元件。
还考虑了包含编码本发明毒素蛋白质的重组dna构建体的转基因宿主细胞。转基因宿主细胞可进一步被定义为原核宿主细胞,即,细菌细胞,例如,苏云金芽孢杆菌、枯草芽孢杆菌(bacillussubtilis)、巨大芽胞杆菌(bacillusmegaterium)、蜡样芽胞杆菌(bacilluscereus)、侧孢芽孢杆菌(bacilluslaterosperous)、埃希氏菌(escherichia)、沙门氏菌(salmonella)、农杆菌、假单胞菌,或根瘤菌细胞,或真核宿主细胞,例如,植物细胞,并且这些类型的细胞中的每一种还在本文称为微生物细胞、微生物,或微生物体。
如本文所使用的“宿主细胞”意指例如通过电穿孔或通过农杆菌介导的转化或通过使用包裹有重组dna的纳米粒子轰击,或通过转导或通过质粒转移或通过其它方式用外源重组dna转化或转染的细胞。本发明的宿主细胞可为转化的细菌,例如,大肠杆菌宿主细胞或bt宿主细胞或农杆菌宿主细胞或植物宿主细胞。
因此,宿主细胞可以是作为微生物体或作为后代植物细胞存在的原始转化的植物细胞,所述后代植物细胞再生为分化组织,例如,再生为具有稳定整合的非天然重组dna的转基因植物,或来源于后代转基因植物的种子或花粉。如本文所使用的“转基因植物”包括基因组已经因重组dna的稳定整合而发生改变的植物、植物部件、植物细胞或种子。转基因植物包括从原始转化的植物细胞再生的植物及来自转化的植物的后世或杂交的后代转基因植物。因此,植物部件的实例为叶、树枝、树皮、叶片、花粉粒、梗、细胞、茎、花、萼片、果实、根、或种子。
表达本发明的一种(多种)蛋白质的转基因植物表现出害虫耐受性。此类植物及其细胞包括紫花苜蓿、香蕉、大麦、豆科植物(bean)、浆果、芸苔属植物(brassica)、西兰花、卷心菜、仙人掌、胡萝卜、木薯、蓖麻、菜花、芹菜、鹰嘴豆、大白菜、柑橘、三叶草、椰子、咖啡、玉米、棉花、黄瓜、葫芦、花旗松(douglasfir)、茄子、桉树、亚麻科植物(flax)、水果、大蒜、葡萄、啤酒花、木棉、韭葱、豆科植物(legume)、莴苣(lettuce)、火炬松(loblollypine)、瓜类植物(melon)、小米(millet)、坚果、燕麦、橄榄、洋葱、观赏性植物、棕榈、牧草、豌豆、花生、胡椒、木豆、松树、杨树、马铃薯、南瓜、辐射松(radiatapine)、萝卜、油菜籽、水稻、砧木、黑麦、红花、灌木、高粱、南方松(southernpine)、大豆、菠菜、南瓜属植物(squash)、草莓、多汁植物、甜菜、甘蔗、向日葵、甜玉米、甜胶皮枫香树(sweetgum)、红薯、柳枝稷、茶、烟草、番茄、树、黑小麦、草坪草、蔬菜、西瓜、及小麦植物和细胞。
可构建可用于在植物细胞中表达这些蛋白质的核苷酸序列,并且可将此类植物细胞再生为可产生含有此类核苷酸序列的种子的转基因植物,该转基因植物可商业化,并且可与表达不同bt昆虫抑制蛋白质或其它对作物害虫有毒性的试剂的其它转基因植物一起育种。
植物细胞可通过本领域技能范围内的多种机制转化,所述多种机制包括但不限于细菌转化系统,例如农杆菌或根际细菌(rhizobacterium)、电穿孔、冲击(ballistic)介导的系统等。微粒轰击方法在美国专利5,015,580(大豆)、5,550,318(玉米)、5,538,880(玉米)、5,914,451(大豆)、6,160,208(玉米)、6,399,861(玉米)、6,153,812(小麦)和6,365,807(水稻)中有举例说明,而农杆菌属介导的转化在美国专利5,159,135(棉花)、5,824,877(大豆)、5,463,174(卡诺拉(canola))、5,591,616(玉米)、5,846,797(棉花)、6,384,301(大豆)、7,026,528(小麦)和6,329,571(水稻),及美国专利申请公布2004/0087030a1(棉花)和美国专利申请公布2001/0042257a1(甜菜)及arencibia等人(1998)transgenicres.7:213-222(甘蔗)中有描述,其它更新的方法描述于美国专利申请公布2009/0138985a1(大豆)、2008/0280361a1(大豆)、2009/0142837a1(玉米)、2008/0282432(棉花)、2008/0256667(棉花)、2003/0110531(小麦),及美国专利5,750,871(卡诺拉(canola))、7,026,528(小麦)和6,365,807(水稻)中。植物材料的转化可在组织培养物中在营养介质,例如将允许细胞在体外生长的营养物混合物中进行。受体细胞靶标包括但不限于分生组织细胞、下胚轴、愈伤组织、未成熟胚及配子细胞如小孢子、花粉、精子和卵子细胞。愈伤组织可能起源于包括但不限于以下的组织源:未成熟胚、下胚轴、幼苗茎尖分生组织、小孢子等。含有转基因细胞核的细胞生长成转基因植物。
除了用重组dna直接转化植物材料之外,还可通过使具有含有含重组dna的转基因细胞核的细胞的第一植物与缺乏转基因细胞核的第二植物杂交来制备转基因植物细胞核。例如,可将重组dna导入到适合于在细胞中转化成转基因细胞核的来自第一植物品系的细胞核中,该转基因细胞核生长成转基因植物,可使该转基因植物与第二植物品系杂交从而将重组dna种质渗入到第二植物品系中。可使具有提供增强的性状,例如,增加的产率的重组dna的转基因植物与具有赋予另一种性状,例如除草剂抗性或害虫抗性的其它重组dna的转基因植物品系杂交,以产生具有赋予这两种性状的重组dna的后代植物。通常,在此种组合性状的育种中,供给额外性状的转基因植物为雄性品系,而携带基础性状的转基因植物为雌性品系。这种杂交的后代将发生分离,使得一些植物将携带双方亲本性状的dna,而一些只携带一方亲本性状的dna;可通过与亲本重组dna关联的标记来鉴定此类植物,例如,通过针对重组dna的分析鉴定标记,或者在可选择标记连接至重组dna的情况下,通过应用选择剂,例如用于与除草剂耐受性标记一起使用的除草剂,或通过针对昆虫抑制性状进行选择来鉴定标记。可将携带双方亲本性状的dna的后代植物与雌性亲本品系多次回交,例如通常6到8代,以产生除了另一个转基因亲本品系的重组dna之外还具有与一个原始转基因亲本品系基本相同的基因型的后代植物。
在植物转化实践中,通常在任一个转化实验中,仅将外源dna导入到小百分比的靶标植物细胞中。可通过多种熟知的dna检测方法或通过多种熟知的测试昆虫抑制活性的生物活性测定(进一步描述于实施例部分中)直接测试本发明的细胞来证实外源dna的稳定整合。标记基因可用来提供用于鉴定通过接受重组dna分子并将其整合到其基因组中而被稳定转化的细胞的有效系统。优选的标记基因提供赋予选择剂如抗生素或除草剂抗性的选择标记。本发明植物对其可具有抗性的任何除草剂都可用作供选择标记用的试剂。使潜在转化的细胞暴露于选择剂中。在存活细胞群体中的细胞将是这样的细胞,其中一般整合了赋予抗性的基因并且该基因以允许细胞存活的足够水平表达。可进一步测试细胞以证实外源dna的稳定整合。常用的选择标记基因包括赋予抗生素抗性或除草剂抗性的基因,所述抗生素例如卡那霉素和巴龙霉素(nptii)、潮霉素b(aphiv)、壮观霉素(aada)和庆大霉素(aac3和aacc4),所述除草剂例如草铵膦(bar或pat)、麦草畏(dmo)和草甘膦(aroa或epsps)。此种可选择标记的实例在美国专利5,550,318、5,633,435、5,780,708和6,118,047中有举例说明。还可采用提供目视筛选转化体的能力的标记,例如,表达有色蛋白或荧光蛋白,例如荧光素酶或绿色荧光蛋白(gfp)的基因。
可在再生培养基中培养暴露于选择剂后存活的植物细胞或在筛选测定中评分为正的植物细胞,并让它们成熟为植物。可将从转化的植物细胞再生的发育中的植物转移至植物生长混合物(plantgrowthmix)中,并且例如在大约85%的相对湿度、600ppmco2和25-250微爱因斯坦m2s-1的光下在可控环境箱中使之受冷而变得耐寒,然后后将其转移至温室或生长箱中以进行成熟。这些生长条件随着植物物种变化,并且是本领域技术人员已知的。在鉴定转化体后再生植物约6周至10个月,这取决于原始组织和植物物种。可使用本领域技术人员已知的常规植物育种方法对植物传粉并产生种子,例如对于转基因玉米通常使用自花传粉。可针对重组dna的表达测试再生的转化的植物或其后代种子或植物,并针对昆虫抑制活性的存在对它们进行选择。
栽种编码并表达本发明一种或多种蛋白质的转基因植物以(i)产生具有与对照植物相比增强的性状的转基因植物,及(ii)产生本发明的转基因种子和单倍体花粉。通过针对增强的性状选择转化的植物或后代种子来鉴定此类具有增强的性状的植物。为有效起见,设计选择方法以评价包含重组dna的多个转基因植物(事件),例如来自2至20或更多个转基因事件的多个植物。从本文所提供的转基因种子生长的转基因植物展现出促成增加的昆虫抑制耐受性或增加的收获产量的改进农艺学性状,或提供增加的植物价值(包括例如改进的种子或圆荚质量)的其它性状。特别感兴趣的是具有增强的对抗一种或多种鳞翅目和/或半翅目昆虫的昆虫抑制抗性的棉花、紫花苜蓿、玉米、大豆,或甘蔗植物。特别感兴趣的是具有增强的对抗半翅目昆虫的昆虫抑制抗性的棉花植物。
本发明提供了从包含编码本发明昆虫抑制多肽的重组多核苷酸分子的种子产生植物和收获作物的方法。特别感兴趣的是具有增强的对抗一种(多种)抗鳞翅目和/或半翅目昆虫的昆虫抑制抗性的棉花、紫花苜蓿、玉米、大豆,或甘蔗植物。该方法包括以下步骤:使表达本发明重组多肽的抗昆虫植物与另一种植物杂交,获取至少一种来源于这种杂交的后代植物,及选择表达本发明重组多肽的后代,其中所述后代对昆虫有抗性。这包括种植种子、从由种子生长的植物产生作物及收获作物的步骤,其中至少50%的作物包含含有重组多核苷酸分子的种子。
在本发明的一个方面,本文提供了包含表达任一种或多种所述蛋白质编码序列的重组多核苷酸(即转基因)的转基因植物细胞、转基因植物和转基因植物部件。预期“细菌细胞”或“细菌”可包括但不限于农杆菌、芽孢杆菌、埃希氏菌、沙门氏菌、假单胞菌或根瘤菌细胞。预期“植物细胞”或“植物”包括紫花苜蓿、香蕉、大麦、豆、西兰花、卷心菜、芸苔属植物、胡萝卜、木薯、蓖麻、菜花、芹菜、鹰嘴豆、大白菜、柑橘、椰子、咖啡、玉米、三叶草、棉花、葫芦、黄瓜、花旗松、茄子、桉树、亚麻科植物、大蒜、葡萄、啤酒花、韭葱、莴苣、火炬松、小米、瓜类植物、坚果、燕麦、橄榄、洋葱、观赏性植物、棕榈、牧草、豌豆、花生、胡椒、木豆、松树、马铃薯、杨树、南瓜、辐射松、萝卜、油菜籽、水稻、砧木、黑麦、红花、灌木、高粱、南方松、大豆、菠菜、南瓜属植物、草莓、甜菜、甘蔗、向日葵、甜玉米、甜胶皮枫香树、红薯、柳枝稷、茶、烟草、番茄、黑小麦、草坪草、西瓜,及小麦植物细胞或植物。在某些实施方案中,提供了从转基因植物细胞再生的转基因植物和转基因植物部件。在某些实施方案中,转基因植物可从转基因种子获得。在某些实施方案中,转基因植物部件可通过将其从植物上切断、掰断、碾断或以其它方式分离出来来获得。在某些实施方案中,植物部件可为种子、圆荚、叶,花、茎、根,或其任何部分。在某些实施方案中,本文所提供的转基因植物部件为转基因植物部件的不可再生部分。如上下文中使用的转基因植物部件的“不可再生”部分是不能被诱导形成整株植物或不能被诱导形成能够有性繁殖和/或无性繁殖的整株植物的部分。在某些实施方案中,植物部件的不可再生部分为转基因种子、圆荚、叶,花、茎、根的部分。
本文还提供了制备包含昆虫抑制量的本发明的一种(多种)蛋白质的转基因植物的方法。此类植物可通过如下方式制得:将编码本文所提供的任何蛋白质的重组多核苷酸导入到植物细胞中,并选择从所述植物细胞获得的表达昆虫抑制量的蛋白质的植物。可通过再生、种子、花粉,或分生组织转化技术从植物细胞获得植物。
本文还提供了表达昆虫抑制量的本发明的一种或多种蛋白质从而控制鳞翅目和/或半翅目物种害虫的转基因植物的用途。任何前述转基因植物都可用在本文所提供的保护植物免遭昆虫侵扰的方法中。
本文还提供了任何前述转基因宿主细胞在生产本发明蛋白质中的用途。
本发明另外的方面包括检测本发明的dna、rna或蛋白质的方法和/或试剂盒、鉴定本文所描述的蛋白质属的成员的方法、鉴定与属家族成员有关的新型蛋白质的方法、测试对昆虫生长和/或侵扰的控制的方法,及向植物和其它受体宿主提供此种控制的方法。这些蛋白质可用来产生与这个蛋白质类/属特异性结合的抗体,并且这些抗体可用来筛选和寻找该属的其它成员。考虑了与本发明重组多肽的靶标特异性结合的单独抗体或抗体混合物;并且还考虑了使用这种单独抗体或抗体混合物检测或定量共享本发明蛋白质的表位的蛋白质的方法。此种检测或定量方法可包括以下步骤:使样品与抗体接触及使用本领域中熟知的检测方式检测抗体与样品中多肽靶标的结合。当考虑在此种方法中使用一个或多个表位及它们的组合时,识别不同表位的抗体或抗体混合物的结合可鉴定表现出与本发明重组多肽具有同源性的多肽。
提供了用于在怀疑含有多肽靶标的样品中检测多肽靶标的存在的试剂盒。此类试剂盒将包括一种(多种)用于表位检测的试剂及一种(多种)用于证明该检测是在统计差异内运作的对照试剂。考虑了可包括在此类试剂盒中的试剂储存器、检测方式和试剂使用的说明书,及另外的部件和工具。
编码本发明蛋白质,即,所描述的属的蛋白质的多核苷酸区段,特别是来源于野生型bt菌株的区段,可用作用于使用热扩增和/或杂交方法筛选和鉴定该属内其它成员的探针和引物。核苷酸探针或引物的长度、序列、浓度、主链和构成(formulation)可以不同,这取决于所使用的样品检测方法。本发明以可用于检测和分离编码该属的昆虫抑制蛋白质成员的同源基因的引物和探针为特征。考虑了dna检测试剂盒,其使技术人员更容易执行本发明同源基因的检测和/或分离。本发明提供了此类试剂盒和方法的用途,及用前述检测方式检测和分离的新型基因和由此类基因编码的昆虫抑制多肽。
本发明进一步提供了测试本发明多肽的昆虫抑制活性的方法,在本文称为“生物测定”。本文描述了测量生长抑制、致死性或二者的组合的昆虫定性生物测定。在以下实施例中测试的昆虫目包括鞘翅目、双翅目、鳞翅目和半翅目。食物配方和制备、测试和对照样品的制备、昆虫制备,及用于进行测定的程序通常取决于经受任何特定评价的昆虫和/或害虫的类型和大小。以下实施例举例说明并详细描述了此类方法。
在某些实施方案中,植物产品可包括从转基因植物或转基因植物部件获得的日用品或其它商业产品,其中可通过检测编码或包含本发明蛋白质区分部分的核苷酸区段或表达的rna或蛋白质经由商务追踪该日用品或其它产品。此类日用品或其它商业产品包括但不限于植物部件、生物质、油、粗磨粉、糖、动物饲料、粉状物、片状物、糠、皮棉、加工的种子,及种子。
同时还提供了包含可检测量的编码本发明任一种蛋白质、其昆虫抑制片段或其任何区分部分的重组核苷酸的加工的植物产品。在某些实施方案中,加工的产品选自由以下组成的组:植物生物质、油、粗磨粉、动物饲料、粉状物、片状物、糠、皮棉、外壳,及加工的种子。在某些实施方案中,加工的产品是不可再生的。在某些实施方案中,它的区分部分可包含任何编码seqidno:2、4、6、8、10、12、14、16、18、20、22、24、26、136或138的氨基酸序列的至少20、30、50或100个氨基酸的多核苷酸。
本文还提供了控制昆虫的方法。此类方法可包括栽种包含昆虫抑制量的本发明蛋白质的植物。在某些实施方案中,此类方法可进一步包括以下任何一者或多者:(i)将包含或编码本发明蛋白质的任何组合物施用至植物或产生植物的种子;和/或(ii)用编码本发明蛋白质的多核苷酸转化植物或产生植物的植物细胞。在某些实施方案中,该植物为包含表达昆虫抑制量的本发明蛋白质的转基因的瞬时或稳定转化的转基因植物。在某些实施方案中,该植物为包含已施用本发明蛋白质的组合物的非转基因植物。
根据以下详细描述、实施例和权利要求书,本发明的其它特征和优点将是明显的。
实施例
鉴于前述内容,本领域技术人员应理解,可在所公开的具体方面作出改变并且仍获得相似或类似的结果,而不偏离本发明的精神和范围。因而,本文所公开的具体细节不应被解释为限制性的。
实施例1:昆虫抑制蛋白质的发现
鉴定相互比较时具有独特的属性,例如,推测的毒性、蛋白质组学多样性和形态变异的各种bt菌株,从每种此种菌株中获取dna并使其为dna测序作好制备。为每种此种菌株获得dna序列信息,加工原始序列读段(reads),由经加工的读段组装重叠群(contig),鉴定开发阅读框,并分析推导的氨基酸序列。
实施例2:克隆和表达昆虫抑制蛋白质
本实施例举例说明了编码昆虫抑制蛋白质的多核苷酸区段的克隆,及在重组宿主细胞中的插入和表达。
通过扩增从实施例1自其鉴定每个开放阅读框的相应基因组样品中获取核苷酸区段。将扩增的核苷酸区段插入到重组质粒中,并将其转化成无晶体bt宿主细胞或转化成大肠杆菌表达菌株,观察到所得一种(多种)重组菌株表达重组蛋白质。
观察到本文例示的重组蛋白质表现出针对如下文实施例3–13中所描述的多种害虫物种的昆虫抑制性能。证实了如seqidno:1、seqidno:3、seqidno:5、seqidno:7、seqidno:11、seqidno:15、seqidno:17、seqidno:19、seqidno:25、seqidno:135和seqidno:137所示的核苷酸序列编码具有如seqidno:2、seqidno:4、seqidno:6、seqidno:8、seqidno:12、seqidno:16、seqidno:18、seqidno:20、seqidno:26、seqidno:136和seqidno:138所示的氨基酸序列的蛋白质(分别为tic1498、tic1415、tic1497、tic1886、tic1414、tic1922、tic1422、tic1974、tic1362、tic2335和tic2334)。
还构建了含有具有如seqidno:9、seqidno:13、seqidno:21和seqidno:23所示的序列的多核苷酸区段的重组质粒和菌株,证实了该重组质粒和菌株编码具有如seqidno:10、seqidno:14、seqidno:22和seqidno:24所示的氨基酸序列的蛋白质(分别为tic1925、tic1885、tic2032和tic2120)。
实施例3:tic1886的鳞翅目活性
本实施例举例说明了来自表达具有seqidno:8的推导氨基酸序列的重组蛋白质tic1886的重组菌株的样品表现出玉米穗虫(helicoverpazea,hz)活性。
在14%(w/v)琼脂基质(serva#11393)中制备具有16.5%(w/v)“multiplespecies”食物(southlandproducts,201stuartislandroad,lakevillage,arkansas71653)的食物。熔化琼脂基质并将其与食物和净化水掺合至一定体积(1.4%(w/v)琼脂)。将食物悬浮液分配到各个生物测定隔室中。
如实施例2中所描述的那样制备重组蛋白质tic1886的测试样品,并将其覆盖在24个分隔的食物表面上方,每个隔室近似约2890ug/ml。将缓冲液对照样品覆盖在96个分隔的食物表面上方。总共120个分隔的食物表面包含为玉米穗虫生物测定作准备的本实施例的测试装置(testset)。
将单个新生幼虫转移到本实施例的测试装置的每个单独隔室的食物表面中(总共120个新生幼虫),并且用通风罩将每个隔室密封。将测试装置在27℃和60%rh下在无光条件下放置在可控环境中5-7天,并对致命性(mortality)和发育不良进行评分。视觉评价与未经处理的昆虫相比较的发育不良,并将其评分为严重发育不良(>67%发育不良)、中度发育不良(33-67%发育不良)或发育不良(<33%)。
对于缓冲液对照样品未观察到发育不良或致死性。在2890ug/mltic1886情况下,观察到对24只hz幼虫的致死性。结论是,蛋白质tic1886展现出玉米穗虫(helicoverpazea,cornearworm)活性(参见图2和表2)。
还使用本发明蛋白质(来自实施例2,tic1498、tic1415、tic1497、tic1414、tic1422和tic1362)在来自欧洲玉米螟、甘蔗螟(diatraeasaccharalis)、西南玉米螟和藜豆夜蛾物种的幼虫组合上应用本实施例所描述的鳞翅目生物测定程序,结果描述在实施例4-7中。
实施例4:tic1498、tic1497和tic1422的鳞翅目活性
使用实施例3中所描述的方法和生物测定技术,针对新生的欧洲玉米螟(on)、甘蔗螟(ds)、西南玉米螟(dg)和藜豆夜蛾(ag)昆虫物种测试重组蛋白质tic1498(seqidno:2)、tic1497(seqidno:6)和tic1422(seqidno:18)。
在2500ug/ml剂量下对于24只幼虫,tic1498表现出对欧洲玉米螟的致死性,并且存活者显著发育不良。在3700ug/ml剂量下对于24只幼虫,tic1497表现出对欧洲玉米螟的致死性,并且存活者显著发育不良。在1300ug/ml剂量下对于24只幼虫,tic1422表现出对欧洲玉米螟的致死性,并且存活者显著发育不良。结论是,tic1498、tic1497和tic1422展现出对抗欧洲玉米螟(ostrinianubilalis,europeancornborer)的活性(参见图2和表2)。
在3000ug/ml剂量下对于24只幼虫,tic1498表现出对甘蔗螟的致死性,并且存活者显著发育不良。在2000ug/ml剂量下对于24只幼虫,tic1497表现出对甘蔗螟的致死性,并且存活者显著发育不良。在300ug/ml剂量下对于24只幼虫,tic1422表现出对甘蔗螟的100%致死性。结论是,重组蛋白质tic1498、tic1497和tic1422展现出对抗甘蔗螟(diatraeasaccharalis,surgarcaneborer)的活性(参见图2和表2)。
在3000ug/ml剂量下对于24只幼虫,tic1498表现出对西南玉米螟的致死性,并且存活者中度发育不良。在2000ug/ml剂量下对于24只幼虫,tic1497表现出对西南玉米螟的致死性,并且存活者严重发育不良。在300ug/ml剂量下对于24只幼虫,tic1422表现出对西南玉米螟的致死性。结论是,tic1498、tic1497和tic1422展现出对抗西南玉米螟(diatraeagrandiosella,southwesterncornborer)的活性(参见图2和表2)。
在2500-3000ug/ml剂量下对于48只幼虫,tic1498表现出对藜豆夜蛾的致死性,并且存活者严重发育不良。在2000-3700ug/ml剂量下对于48只幼虫,tic1497表现出对藜豆夜蛾的致死性,并且存活者中度至严重发育不良。在300-1300ug/ml剂量下对于48只幼虫,tic1422表现出对藜豆夜蛾的致死性,并且存活者严重发育不良。结论是,tic1498、tic1497和tic1422展现出对抗藜豆夜蛾(anticarsiagemmatalis,velvetbeancaterpillar)的活性(参见图2和表2)。
实施例5:tic1415的鳞翅目活性
针对新生的欧洲玉米螟(on)和藜豆夜蛾(ag)昆虫物种测试tic1415(seqidno:4)。在1500ug/ml剂量下对于24只幼虫,tic1415表现出对欧洲玉米螟的致死性,并且存活者中度发育不良。结论是,tic1415展现出对抗欧洲玉米螟(ostrinianubilalis,europeancornborer)的活性(参见图2和表2)。
在1500ug/ml剂量下对于24只幼虫,tic1415表现出对藜豆夜蛾的致死性,并且存活者中度发育不良。结论是,tic1415展现出对抗藜豆夜蛾(anticarsiagemmatalis,velvetbeancaterpillar)的活性(参见图2和表2)。
实施例6:tic1414的鳞翅目活性
针对新生的藜豆夜蛾(ag)昆虫物种测试tic1414(seqidno:12)。在870ug/ml剂量下对于24只幼虫,tic1414表现出对藜豆夜蛾的致死性,并且存活者发育不良。结论是,tic1414展现出对抗藜豆夜蛾(anticarsiagemmatalis,velvetbeancaterpillar)的活性(参见图2和表2)。
实施例7:tic1362的鳞翅目活性
针对新生的甘蔗螟(ds)和西南玉米螟(dg)昆虫物种测试tic1362(seqidno:26)。在400ug/ml剂量下对于24只幼虫,tic1362表现出对甘蔗螟的致死性,并且存活者严重发育不良。tic1362展现出甘蔗螟(藜豆夜蛾)(diatraeasaccharalis,velvetbeancaterpillar)活性(参见图2和表2)。
在400ug/ml剂量下对于24只幼虫,tic1362表现出对西南玉米螟的100%致死性。tic1362展现出西南玉米螟(diatraeagrandiosella,southwesterncornborer)活性(参见图2和表2)。
实施例8:tic1498的半翅目活性
本实施例举例说明了当提供在半翅目昆虫食物中时tic1498(seqidno:2)的昆虫抑制活性,所述半翅目昆虫包括但不限于半翅目盲蝽科的成员,包括草盲蝽属,例如,豆荚草盲蝽和牧草盲蝽。本实施例更具体地举例说明了来自表达重组蛋白质tic1498的重组菌株的样品表现出豆荚草盲蝽(lh)和牧草盲蝽(ll)活性。
制备具有7.81%(w/v)的“lygusdiet”食物(bio-serv#f9644b,one8thstreet,suiteone,frenchtown,nj08825)和两只完整新鲜卵的液体内容物的食物。将该食物冷却并在4℃下储存在湿度受控条件下直至准备使用。这种食物制品在制备后的2天内使用。
制备包封在拉伸石蜡膜和聚酯薄膜片之间的含有tic1498蛋白质的测试样品(~40ul)(小袋)。
在24℃下温育豆荚草盲蝽和牧草盲蝽卵直至它们在0至约12小时之间达到预孵(pre-hatch)阶段。将预孵卵浸泡在无菌水中并用无菌水冲洗,然后将其紧靠着(confinedproximity)所制备的小袋在24℃和60%rh下在无光条件下放置在可控环境中4-7天,并对致死百分率和任何存活者的发育不良进行评分。视觉评价与未经处理的昆虫相比较的发育不良,并将其评分为严重发育不良(>67%发育不良)、中度发育不良(33-67%发育不良)或发育不良(<33%)。
在10ug/mltic1498情况下,对于24只新生若虫,观察到对牧草盲蝽的致死性,并且存活者发育不良。在50ug/mltic1498情况下,对于24只新生若虫,观察到对牧草盲蝽的致死性,并且存活者发育不良。对于24只新生若虫,在100ug/ml剂量下tic1498表现出对牧草盲蝽的致死性,并且存活者中度发育不良。
在10ug/mltic1498情况下,对于24只新生若虫,观察到对豆荚草盲蝽的致死性,并且存活者发育不良。在50ug/mltic1498情况下,对于24只新生若虫,观察到对豆荚草盲蝽的致死性,并且存活者发育不良。对于24只新生若虫,在100ug/ml剂量下tic1498表现出对豆荚草盲蝽的致死性,并且存活者中度发育不良。在2300ug/mltic1498情况下,对于24只新生若虫,观察到对豆荚草盲蝽的100%致死性。
tic1498展现出牧草盲蝽(lyguslineolaris,tarnishedplantbug)和豆荚草盲蝽(lygushesperus,westerntarnishplantbug)活性(参见图2和表2)。还使用tic1415、tic1497、tic1886、tic1414、tic1922、tic1974和tic1362执行本实施例中所描述的半翅目生物测定程序。
实施例9:tic1922和tic1974的半翅目活性
针对牧草盲蝽(ll)测试tic1922(seqidno:16)和tic1974(seqidno:20)。在3组24个小袋中测试了tic1922,并且在3000ug/ml剂量下,tic1922表现出对牧草盲蝽的致死性,并且存活者表现出发育不良。在使用3000ug/ml剂量的24只新生若虫评价中,tic1974没有对牧草盲蝽表现出致死性,但存活者发育不良。tic1922和tic1974展现出牧草盲蝽(lyguslineolaris,tarnishedplantbug)活性(参见图2和表2)。
实施例10:tic1497的半翅目活性
针对豆荚草盲蝽(lh)测试tic1497(seqidno:6)。在以2000ug/ml剂量评价48只新生若虫的实验中,tic1497表现出对豆荚草盲蝽的致死性,并且存活者中度发育不良。
如下制备tic1497片段制品:用嗜热菌蛋白酶、糜蛋白酶、胰蛋白酶或glu-c处理tic1497,从而产生具有以下分子量的tic1497片段:32411da(seqidno:64)、32557da(seqidno:62)、34225da(seqidno:61)和34485da(seqidno:63)。在离子交换柱上分离(tic1497.32411)来自嗜热菌蛋白酶处理的制品的蛋白质洗脱物,并将其用于针对半翅目物种的生物测定中。
在使用24只新生若虫和100ug/ml剂量的实验中,tic1497.32411表现出对牧草盲蝽的致死性,并且存活者中度发育不良。tic1497.32411在1000ug/ml剂量下表现出100%的致死性。
在使用24只新生若虫和2300ug/ml剂量的实验中,tic1497.32411表现出对牧草盲蝽的致死性,并且存活者中度发育不良。
tic1497展现出豆荚草盲蝽(lygushesperus,westerntarnishplantbug)活性(参见图2和表2)。片段tic1497.32411既展现出牧草盲蝽(lyguslineolaris,tarnishedplantbug)活性,又展现出豆荚草盲蝽(lygushesperus,westerntarnishplantbug)活性(参见图2和表2)。
实施例11:tic1886、tic1415、tic1414和tic1362的半翅目活性
针对牧草盲蝽和豆荚草盲蝽测试tic1886(seqidno:8)、tic1415(seqidno:4)、tic1414(seqidno:12)和tic1362(seqidno:24)。tic1886在相当于124ug/ml的剂量下表现出对牧草盲蝽和豆荚草盲蝽的致死性。tic1415在相当于150ug/ml的剂量下表现出对牧草盲蝽和豆荚草盲蝽的致死性,并且存活者发育不良。tic1414在相当于95ug/ml的剂量下表现出对牧草盲蝽的致死性。tic1362在相当于370ug/ml的剂量下表现出对牧草盲蝽的致死性。
tic1886、tic1415和tic1362既展现出牧草盲蝽活性,又展现出豆荚草盲蝽活性。tic1414展现出牧草盲蝽活性(参见图2和表2)。
实施例12:其它蛋白质成员的昆虫抑制活性
制备本发明属的其它蛋白质成员,例如但不限于tic1925(seqidno:10)、tic1885(seqidno:14)、tic2032(seqidno:22)和tic2120(seqidno:24)用于针对植物害虫的生物测定中,所述植物害虫包括线虫动物门(phylumnematoda)的害虫、鳞翅目的害虫,和半翅目的害虫。
实施例13:蛋白质在植物中的表达
本实施例示出本发明蛋白质在植物中的表达。用于使本发明蛋白质在植物中表达的多核苷酸区段可根据美国专利第7,741,118号中所示出的方法制备。例如,在植物中,具有如seqidno:4、seqidno:12、seqidno:18和seqidno:26所示的氨基酸序列的毒素蛋白质可由具有分别如seqidno:27、seqidno:28、seqidno:29和seqidno:30所示的序列的多核苷酸区段产生。设计用于在植物中使用并用于编码本发明蛋白质、包括如seqidno:27、seqidno:28、seqidno:29和seqidno:30所示的序列的多核苷酸区段可操作地连接至用于在植物中表达的必要表达元件,并且转化成植物细胞,优选棉花、紫花苜蓿、玉米和大豆细胞的基因组。
为了获得抵御害虫侵扰,特别是昆虫害虫侵扰的植物和植物细胞,预期单独地使用编码以下所列举蛋白质、其昆虫抑制片段,及表现出与这些所列举蛋白质中的一种或多种具有本文上文具体指定的同一度的蛋白质中的每一者的多核苷酸区段(或多核苷酸分子),将该多核苷酸区段彼此组合使用,或将该多核苷酸区段与设计用于与本发明蛋白质以协同或相同方式起作用的其它毒素蛋白质或毒剂例如dsrna介导的基因抑制分子组合使用。在本发明范围内的具体的列举的蛋白质包括tic1498(seqidno:2)、tic1415(seqidno:4)、tic1497(seqidno:6)、tic1886(seqidno:8)、tic1925(seqidno:10)、tic1414(seqidno:12)、tic1885(seqidno:14)、tic1922(seqidno:16)、tic1422(seqidno:18)、tic1974(seqidno:20)、tic2032(seqidno:22)、tic2120(seqidno:24)、tic1362(seqidno:26)、tic2335(seqidno:136)、tic2334(seqidno:138)及其昆虫抑制片段,例如但不限于tic1497.34225(seqidno:61)、tic1497.32557(seqidno:62)、tic1497.34485(seqidno:63)和tic1497.32411(seqidno:64)。
例如,可将本发明tic1415属的蛋白质与其它杀虫剂组合,所述其它杀虫剂包括靶向的害虫与tic1415蛋白质靶向的害虫重叠的杀虫剂。另外,其它杀虫剂可包括靶向的害虫与tic1415蛋白质靶向的害虫不重叠的杀虫剂。在任一情况下,预期单独地使用tic1415蛋白质或将其与其它杀虫剂组合使用。在下面描述的实施例中,使tic1415与tic807毒素蛋白质在棉花植物中共表达,并进行植物内(inplanta)生物测定。除了tic807毒素蛋白质之外,可与tic1415蛋白质组合使用的其它杀虫剂包括(1)以半翅目为中心的试剂,例如,针对褐飞虱(nilaparvatalugens)v-atpase-e,21e01的半翅目直系同源物的dsrna(li,jie等人,2011,pestmanagsci);针对五种不同基因的半翅目直系同源物–肌动蛋白直系同源物、adp/atp转位酶、α-管蛋白、核糖体蛋白质l9(rpl9)和v-atp酶a亚基的dsrna(upadhyay,s.k.,等人,2011,j.biosci.36(1),p.153-161);axmi-171(us20100298207a1);被发现无论就死亡率还是生长速率而言都对豌豆蚜虫和豌豆长管蚜(acyrthosiphonpisum)表现出低度至中度毒性的bt内毒素,例如cry3a、cry4aa、cry11aa和cyt1aa(porcar,m.等人,appliedandenvironmentalmicrobiology,july2009,p.4897–4900,vol.75,no.14);(2)其它鞘翅目杀虫剂,一般为,例如,dig11和dig5、cry7、ecry3.1ab、mcry3a、cry8、cry34/cry35,及cry3毒素,以及(3)其它鳞翅目杀虫剂,例如dig2、cry1毒素、cry1a.105、cry2毒素,特别是cry2a毒素、cry1f毒素、vip3毒素,及cry9毒素。表达其它杀虫剂的转基因作物事件也可与表达tic1415蛋白质的作物事件组合使用,其实例包括mon88017,mon89034,mon863、mon15985、mon531、mon757、cot102、tc1507、das59122-7、3006-210-23、281-24-236、t304-40、ghb119、cot67b、mir162;玉米事件5307等。此类与表达tic1415属蛋白质中的一种或多种蛋白质的事件的组合提供了更持久的害虫防护,提供了用于靶标害虫控制的抗性管理策略,并减少了农民投入,从而节省在时间和货币价值上的花费。
从本实施例的转化植物细胞产生重组植物,并评价该重组植物或其后代对害虫侵扰的抗性,例如对半翅目和/或鳞翅目的耐受性。选择为例如半翅目和鳞翅目提供害虫抗性的转基因植物和种子,并且将此类植物和种子继续用于进一步发育。
实施例14:tic1415的植物内生物测定
在本研究中,在植物中,具有如seqidno:4所示的氨基酸序列的毒素蛋白质tic1415由具有如seqidno:27所示的序列的多核苷酸区段产生。将具有编码tic1415的seqidno:27序列的dna,连同用于在棉花细胞中转化和表达的必要启动子和调控元件一起克隆到农杆菌介导的植物转化载体中。产生转基因棉花植物(重组棉花植物)并测试其功效。将再生的(r0)转基因植物转移到土壤中,并从拷贝数低并表达tic1415蛋白质的转化事件中选择组织样品。将来自三个事件的r0植物的冷冻组织样品称重,并将其以1:50和1:100(重量:缓冲液)的比率与25mmph10.5的碳酸钠/碳酸氢钠缓冲液组合以从该组织中提取可溶性蛋白质。通过蛋白质印迹证实样品中存在tic1415蛋白质。使用实施例8中所描述的生物活性测定,将样品提取物喂给牧草盲蝽。还制备来自缺乏tic1415蛋白质的dp393棉花组织的提取物作为阴性对照。来自所有三个事件的样品提取物都表现出对牧草盲蝽的致死性,并且存活者发育不良。致死性和发育不良得分与用来自dp393阴性对照的样品提取物喂养的昆虫的生物活性得分相比较是显著的。
栽种r0植物并使其自花传粉从而获得对于所导入的转基因dna纯合的种子。选择来自三个不同单一拷贝事件的纯合植物,每个事件种植五颗种子,并在整株植物笼养测定中进行评价。让植物生长至开花期,并将每个整株棉花植物包封在由多孔塑料制成的网笼中。将两对雄性和雌性豆荚草盲蝽成虫放入每个笼子中,并允许它们繁殖。让所得昆虫后代侵扰笼养的棉花植物3周。在21天期结束时,对处于各个发育阶段的草盲蝽属昆虫计数,并以每株植物为基础计算平均值。来自所有三个事件的植物具有与dp393阴性对照相比较显著较少的昆虫。参见表4。
表4.tic1415的植物内生物测定.
第2代草盲蝽平均数目是在笼养整株植物测定中从每个事件的五株棉花植物中恢复的第2代草盲蝽的平均数目。具有相同字母的事件在第2代草盲蝽数目上不具有统计学差异(p<0.05,斯氏t)。
实施例15:tic1415和tic807半翅目毒性蛋白质的蛋白质生物测定
制备含有tic1415与tic807半翅目毒性蛋白质的各种混合物的蛋白质样品,并在生物测定中对它们进行测试。还制备单独的tic1415蛋白质和单独的tic807蛋白质作为阳性对照。缓冲液用作阴性对照。使用生物活性测定将样品喂给牧草盲蝽。所有三个含有毒素蛋白质的制品都表现出对牧草盲蝽的致死性,并且存活者发育不良。致死性和发育不良得分与用缓冲液喂养的昆虫的生物活性得分相比较是显著的(参见表5)。该数据揭示不存在拮抗作用。对混合物执行另外的生物测定测试以证明协同和/或相加效应。
表5.蛋白质混合物:tic1415与tic807毒素蛋白质的组合的生物测定数据
*在95%的置信区间内。
实施例16:tic1415和tic807的植物内生物测定
设计用于共表达相应蛋白质tic1415(seqidno:4)和tic807蛋白质的转基因棉花事件。在用牧草盲蝽侵扰的笼养整株植物测定中评价此类植物。十个事件各取五株植物笼养,用2对雄性和雌性牧草盲蝽侵扰每株植物。将测定物在生长箱中在棉花植物发育的正常环境条件下温育21天。以类似方式栽种dp393阴性对照植物。在3周结束时,对处于各个发育阶段的草盲蝽进行计数。计算每株植物上的处于每个发育阶段的豆荚草盲蝽昆虫的平均数目,结果在表6中给出。在这里,使用不同的植物可表达的启动子驱动编码tic1415的转录本在相应构建体12和13中的表达。
表6.蛋白质混合物:tic1415与tic807蛋白质毒素的组合的植物内数据
第2代草盲蝽平均数目是在笼养整株植物测定中从每个事件的五株棉花植物中恢复的第2代草盲蝽的平均数目。具有相同字母的事件在第2代草盲蝽数目上不具有统计学差异(p<0.05,斯氏t)。
- 还没有人留言评论。精彩留言会获得点赞!