一种抑制MMP9活性的多肽及其应用的制作方法
本发明属于生物医学领域,涉及一种抑制基质金属蛋白酶9(mmp9)活性的多肽,具体涉及一种对mmp9有特异性结合并具有抑制其活性的多肽及其用途。
背景技术:
目前,癌症是世界范围内的主要死亡原因,因此对具有潜在抗癌特性的新制剂的研究具有重要意义。基质金属蛋白酶(mmps)与细胞凋亡、细胞迁移、血管生成等过程密切相关,在细胞外基质(ecm)的重构和癌症的进展中起着至关重要的作用。同时,mmps在不同类型肿瘤患者的组织和血液中的表达和活性均有所增加,可能作为预测侵袭性的标志,提示远处转移的风险。因此,基质金属蛋白酶抑制剂(mmpis)可能成为抗癌药物的靶点,mmpis的研发为治疗癌症患者创造可更多的可能。
自20世纪90年代以来,世界各大制药公司都在新型抗肿瘤药物mmpis的研发上投入了大量资金。目前,已有十几种mmpis进入临床试验阶段,还有大量化合物正在进行临床前研究。这些化学合成的mmpis可大致分为肽类mmpis、非肽类mmpis、四环素类mmpis和双膦酸盐类mmpis等。广谱mmps抑制剂,如batimastat,在临床前动物模型中显示出非常优越的抗肿瘤作用。然而在临床试验中这些广谱mmps抑制剂常伴随难以忍受的副作用,限制了他们的使用。后期开发了相对特异性的mmps抑制剂,实验证实具有抗肿瘤和抗血管生成作用。然而,这些抑制剂的临床试验结果是令人失望的。明确mmps在肿瘤发展阶段的作用及保证mmpis的特异性是mmpis更好应用的基础。mmp9在肿瘤组织中特异高表达,涉及肿瘤生长、转移的各个阶段,且与预后不良相关,是一个较好的药物设计靶点。由于不同mmps之间的相似性,实现mmp9抑制剂的特异性以避免对其他mmps的脱靶效应是对mmp9抑制剂研发的一个强有力的挑战。到目前为止,还没有有效的特异性mmp9抑制剂应用于临床。我们迫切需要一种安全有效的mmp9抑制剂。
肽类药物具有不良反应低,免疫原性低,易于合成和转化的优点,在本研究中基于对mmp9晶体结构的分析,设计靶向mmp9的肽抑制剂。利用moe软件中的分子对接模块筛选稳定的抑制肽。
技术实现要素:
本发明的目的在于提供一种抑制mmp9活性的新型多肽及其应用,该多肽能与mmp9特异性结合并抑制其活性,在高表达mmp9的套细胞淋巴瘤细胞中可与肿瘤细胞特异性结合,并抑制肿瘤细胞增殖及侵袭转移能力,这在套细胞淋巴瘤的早期诊断和靶向药物的研发等方面具有重要作用。
mmp9在体内通常以酶原形式存在,其前肽部分插入至活性位点裂隙,阻碍催化活性中心的锌离子与底物结合,被激活时前肽脱落,暴露出活性位点,激活mmp9功能。本发明即以前肽的这一特性,选取前肽序列中包含关键氨基酸97-100prcg的氨基酸序列tprcgvpdl作为抑制mmp9激活的模板肽。根据氨基酸理化性质替换模板肽中四个关键位点氨基酸,设计不同序列氨基酸作为候选肽。将候选肽及mmp9结构导入moe软件中,利用分子对接模块将mmp9分别与各候选肽进行对接,根据s评分及相互作用分析筛选靶向抑制肽。该多肽可通过抑制mmp9活性进一步抑制套细胞淋巴瘤细胞增殖及侵袭转移,有望成为治疗套细胞淋巴瘤的新型药物。
为了实现上述目的,本发明采用如下技术方案。
一种抑制mmp9活性的多肽,该多肽为以下任意:(1)多肽的氨基酸序列为:tfkepvpdlc,(2)在(1)所述的多肽分子中经过缺失、插入或置换一个或几个氨基酸且与(1)所述的多肽分子具有相同生物学功能的多肽衍生物。
进一步地,所述多肽对mmp9具有靶向抑制性,与肿瘤细胞特异性结合并抑制肿瘤细胞活性。
进一步地,所述的肿瘤细胞为套细胞淋巴瘤细胞。
一种抑制mmp9活性的多肽在制备肿瘤诊断试剂盒中的应用。
进一步地,该试剂盒中包含所述多肽或多肽偶联物。
一种抑制mmp9活性的多肽在制备用于治疗套细胞淋巴瘤药物中的应用,该药物包含所述的多肽与药物活性成分,或包含所述的多肽与递药载体。
进一步地,该药物为任何药物治疗学上可接受的剂型。
进一步地,该药物优选的剂型为注射制剂。
进一步地,该药物为任何药物治疗学上可接受的剂量。
与现有技术相比,本发明的有益效果如下。
(1)本发明使用moe软件分子对接技术具有操作简便、高通量的淘选、高效率,可以模拟多肽与靶蛋白结合、筛选最优结合构象、成本较低等优点。寻找具有特定生物活性的新化合物需要付出巨大的人力和成本。这种付出源于大量的化合物需要被合成,并随后进行生物评估。因此,制药企业对使药物的合理设计成为可能的理论方法表现出极大的兴趣。近年来,随着专业软件的发展和计算机能力的提高,生物信息学经历了巨大的演变。moe软件分子对接模块在配体和大分子靶标之间寻找有利的结
合构型,已经用于新药的设计和虚拟筛选。
(2)本发明通过研究pro-mmp9激活模式及前肽功能,运用moe分子对接技术所筛选出的多肽能够与激活型mmp9蛋白特异性结合并抑制其活性,可抑制肿瘤细胞活性,而且作用效果明显,为临床诊断、治疗及新药研发提供了可靠的科学依据。
附图说明
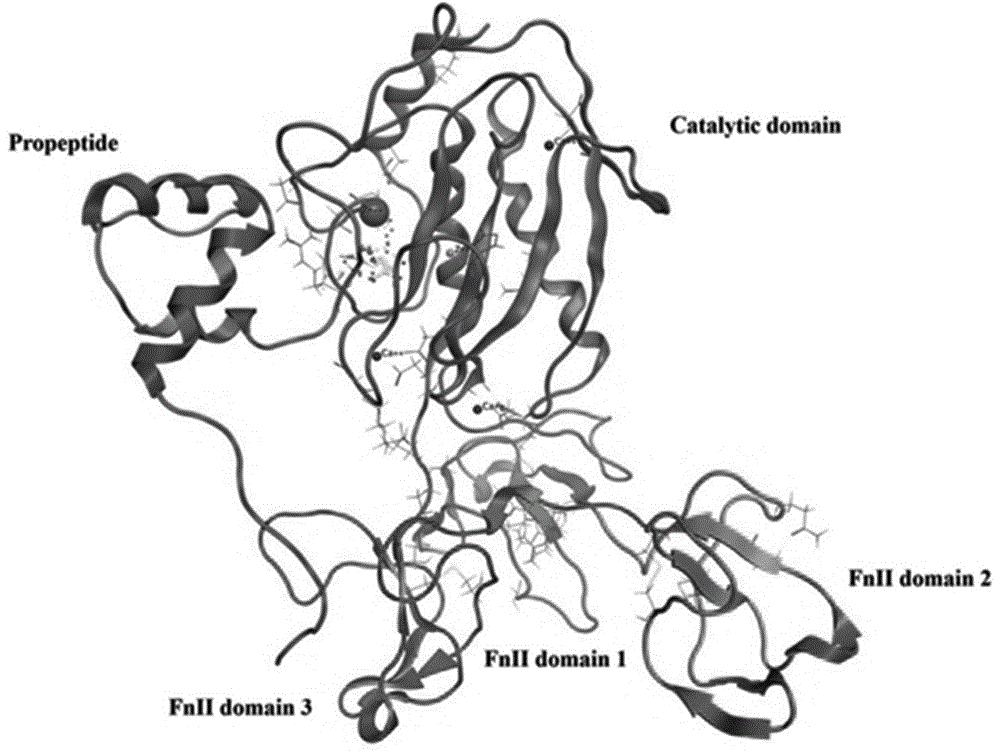
图1是pro-mmp93d晶体结构带状图,其中催化域显示为粉色,三个fnii域分别显示为红色、黄色及蓝色,活性中心的钙显示为红色球体而锌显示为橙色球体,疏水区s1,s2,s1',s2'氨基酸残基显示为红白色小球体。
图2是设计候选肽的氨基酸替换方案,其中a为非极性芳香族氨基酸(f/y/w)替换第一位氨基酸p;b为非极性芳香族氨基酸(f/y/w)替换第一位氨基酸g。
图3是受体与配体氨基酸残基相互作用,m0多肽。
图4是受体与配体氨基酸残基相互作用,m1多肽。
图5是受体与配体氨基酸残基相互作用,m2多肽。
图6是受体与配体氨基酸残基相互作用,m3多肽。
图7是多肽与mmp9活性位点相互作用的3d结构,其中a-d分别对应m0-m3,设计的抑制肽显示为绿色骨架肽段。
图8是m0多肽结构式。
图9是m1多肽结构式。
图10是m2多肽结构式。
图11是m3多肽结构式。
图12是m0多肽性质预测,m0序列:tprcgvpdlc,chemicalformula:c43h73n13o14s2,消光系数:0m-1cm-1,gravy:0.07,平均分子量:1060.24g/mol,理论等电点:ph6.13。gravy=疏水性的算术平均值,负数代表亲水,越小越亲水。
图13是m1多肽性质预测,m1序列:tfkcgvpdlc,chemicalformula:c47h75n11o14s2,消光系数:0m-1cm-1,gravy:0.57,平均分子量:1082.29g/mol,理论等电点:ph6.13。gravy=疏水性的算术平均值,负数代表亲水,越小越亲水。
图14是m2多肽性质预测,m2序列:tfkegvpdlc,chemicalformula:c49h77n11o16s,消光系数:0m-1cm-1,gravy:-0.03,平均分子量:1108.26g/mol,理论等电点:ph4.07。gravy=疏水性的算术平均值,负数代表亲水,越小越亲水。
图15是m3多肽性质预测,tfkepvpdlc,chemicalformula:c52h81n11o16s,消光系数:0m-1cm-1,gravy:-0.15,平均分子量:1148.32g/mol,理论等电点:ph4.07。gravy=疏水性的算术平均值,负数代表亲水,越小越亲水。
图16是流式细胞仪术检测m0肽与套细胞淋巴瘤结合特异性,将细胞分为jeko-1组、5μm肽、50μm肽、100μm肽、150μm肽及mmp9抗体组,随着肽浓度的增加,结合率逐渐增高,但都低于mmp9抗体组。浓度单位:μm;c:jeko-1组;ab:mmp9抗体组。
图17是流式细胞仪术检测m1肽与套细胞淋巴瘤结合特异性,将细胞分为jeko-1组、5μm肽、50μm肽、100μm肽、150μm肽及mmp9抗体组,随着肽浓度的增加,结合率逐渐增高,但都低于mmp9抗体组。浓度单位:μm;c:jeko-1组;ab:mmp9抗体组。
图18是流式细胞仪术检测m2肽与套细胞淋巴瘤结合特异性,将细胞分为jeko-1组、5μm肽、50μm肽、100μm肽、150μm肽及mmp9抗体组,随着肽浓度的增加,结合率逐渐增高,但都低于mmp9抗体组。浓度单位:μm;c:jeko-1组;ab:mmp9抗体组。
图19是流式细胞仪术检测m3肽与套细胞淋巴瘤结合特异性,将细胞分为jeko-1组、5μm肽、50μm肽、100μm肽、150μm肽及mmp9抗体组,随着肽浓度的增加,结合率逐渐增高,但都低于mmp9抗体组,同时加入无荧光mmp9抗体及150μmm3时,m3结合率降低。浓度单位:μm;c:jeko-1组;ab:mmp9抗体组。
图20是明胶酶谱法检测模板肽m0抑制mmp9活性,m0可明显抑制mmp9活性,并不影响mmp9含量。
图21是明胶酶谱法检测肽m3抑制mmp9活性,m3可明显抑制mmp9活性,并不影响mmp9含量。
图22是elisa法检测加入不同浓度m3后jeko-1细胞培养基中mmp9含量。
图23是100μmm3加入jeko-1细胞中培养48h后,免疫荧光检测荧光肽m3(fitc)表达,细胞膜用dil复染,放大倍数×200。
图24是cck-8检测jeko-1细胞加入50μm、100μmm3及mmp9抗体后细胞增殖变化,*p<0.05,**p<0.01,与jeko-1细胞相比。
图25是流失细胞术分别检测jeko-1细胞组细胞周期。
图26是流失细胞术分别检测50μmm3。组细胞周期。
图27是流失细胞术分别检测10μm3组细胞周期。
图28是流失细胞术分别检测mmp9抗体组细胞周期。
图29是jeko-1细胞、50μmm3、/10μm3及mmp9抗体组细胞周期定量分析数据,*p<0.05,**p<0.01,与jeko-1细胞相比;#p<0.05,##p<0.01,与mmp9抗体组相比。
图30是transwell实验检测jeko-1细胞加入50μm、100μmm3及mmp9抗体后细胞侵袭能力变化,**p<0.01,与jeko-1细胞相比;#p<0.05,与mmp9抗体组相比。
图31是transwell实验检测jeko-1细胞加入50μm、100μmm3及mmp9抗体后细胞迁移能力变化,*p<0.05,**p<0.01,与jeko-1细胞相比;#p<0.05,##p<0.01,与mmp9抗体组相比。数据以均值±标椎差表示,每个处理组结果均重复三次。
具体实施方式
下面结合实施例和附图对本发明作进一步的阐述,以下实施例仅为本发明的优选实施例,并不限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
实施例。
1.实验材料。
1.1细胞。
jeko-1细胞购自中国科学院上海细胞库。
1.2实验试剂。
mmpzymographyassaykit:普利莱基因技术有限公司。
mmp9抗体:英国abcam公司。
人类mmp9elisa试剂盒:英国abcam公司。
碘化丙啶(pi):美国sigma公司。
rnasea:美国sigma公司。
多聚甲醛:北京索莱宝科技有限公司。
dil:美国invitrogen公司。
rpmi-1640培养基:美国gibco公司。
胰蛋白酶:美国gibco公司。
胎牛血清(fbs):美国hyclone公司。
transwell小室:美国corning公司。
matrigel胶:美国corning公司。
cck-8试剂盒:碧云天公司。
1.3实验工作液。
明胶酶谱实验试剂配制。
(1)洗脱液(1×):量取tritonx-10025ml,加去离子水定容至1000ml即可。
(2)孵育液(1×):称取tris碱6.06g,cacl2·2h2o0.74g(或cacl20.555g),brij-350.2g,nacl11.7g。加水至800ml。用浓hcl调ph至7.6,定容至1000ml。
(3)2×上样缓冲液(不含β巯基乙醇)。
0.5mtris-hcl,ph6.8:2.5ml。
甘油:2.0ml。
10%(w/v)sds:4.0ml。
0.1%溴酚蓝:0.5ml。
dh2o:1ml。
total:10ml。
(4)5×上样缓冲液(不含β巯基乙醇):量取下列试剂,置于15ml塑料离心管中。
1mtris-hcl(ph6.8):2.5ml。
sds:1g。
溴酚蓝:0.05g。
甘油:5ml。
加dh2o定容至10ml,vortax振荡混匀,小份分装(1ml/管),-20℃保存。
(5)0.5%(w/v)考马斯亮蓝r-250400ml:称取2g考马斯亮蓝r-250,置于1l烧杯中,量取100ml异丙醇加入上述烧杯中,搅拌溶解,加入40ml冰醋酸,搅拌均匀,加入260ml去离子水,搅拌溶解,用滤纸过滤除去颗粒物质后,室温保存。
(6)考马斯亮蓝脱色液(400ml):甲醇:乙酸:水=200:40:160=5:1:4。
(7)明胶储液(10mg/ml):称取明胶0.1g,加去离子水10ml溶解。配好后储存于4℃。若凝固成胶冻状可用55℃水浴解冻。
(8)10%sds-page凝胶之分离胶(含1.0mg/ml明胶)。
dh2o:3ml。
30%丙烯酰胺溶液:3.3ml。
1.5mtris-hcl(ph8.8):2.5ml。
10%sds:0.1ml。
10%过硫酸铵:0.1ml。
temed:0.004ml。
10mg/ml明胶:1ml。
total:10ml。
浓缩胶的配置。
h2o:4.1ml。
30%丙烯酰胺溶液:1ml。
1.0mol/ltris-hcl:0.75ml。
10%sds:0.06ml。
10%过硫酸铵:0.06ml。
temed:0.006ml。
2.实验方法。
2.1moe(分子操作环境)软件中进行受体优化。
应用分子操作环境(moe)软件将肽与激活型mmp9进行分子对接。从蛋白数据库(pdb)中获取mmp9的3d晶体结构(pdbid:1l6j)。通过软件添加h原子对结构进行优化。同时为了优化结构,将h2o分子从结构中去除,随后进行三维质子化,使其处于离子化水平。此外,分子对接前需能使用特定的参数将受体进行能量最小化。
2.2选择模板肽及设计候选肽。
mmp9通常存在于酶原形式(prommp9),prommp9结构包括前肽、催化域及三纤连蛋白ii型(fnii)f域如图1所示。前肽97-100位氨基酸残基插入催化域活性位点的裂隙中,阻断了催化中心zn2+与底物的结合。在体内或者体外,通过有机汞化合物或其他蛋白酶刺激,prommp9可通过自我蛋白酶降解裂解前肽部分成为激活型mmp9。前肽的裂解开放了活性位点,促进催化域与底物或抑制剂的相互作用。通过对前肽的研究,从前肽中筛选出氨基酸序列tprcgvpdl作为模板肽段,其中包含97-100位关键氨基酸残基(prcg)。根据氨基酸理化性质优化关键氨基酸残基。在关键氨基酸中添加非极性芳香氨基酸(f/y/w),有两种选择:取代第一位置氨基酸p或者第四位置g。关键氨基酸的第二位残基与带正电荷的极性氨基酸残基k有相似的相互作用,而第三位置氨基酸残基属于极性氨基酸,可以选择极性氨基酸进行替代(r/n/d/q/e/h/c/s/t)(图2)。设计三十余种候选肽用于后续与激活型mmp9分子对接。利用chemsketch软件将设计的肽段转换成各自的3d结构。
2.3moe中进行肽优化。
利用moe软件对所有候选肽段进行了优化。首先与受体优化过程类似,需要为肽段添加h原子。设置gradient:0.05、forcefield:mmff94x、chiralconstrzaint和currentgeometry等参数实现对肽的能量最小化。此外,利用lowmodemd方法对这些肽进行构象搜索,并将搜索结果保存在mdb数据库中,以便进一步对接分析。
2.4moe中分子对接模块进行肽与受体对接。
利用moe软件的算法将这些肽段与激活型mmp9进行分子对接。参数设置为re-scoringfunction:londondg、placement:trianglematcher、retain:5、refinement:forcefield及re-scoring2:londondg。moe的分子对接程序通过化学键的旋转及柔性分子结构为配体提供了正确的构象。s评分是选择最优肽段以及最优构象的依据。最优构象通过氨基酸相互作用进一步研究氢键及π-π共轭情况。
2.5流式细胞技术检测肽肽与肿瘤细胞结合特异性。
取jeko-1细胞1×106个,pbs洗涤2次后分别加入荧光标记肽m0(50μm、100μm、150μm)、m1(50μm、100μm、150μm)、m2(50μm、100μm、150μm)、m3(50μm、100μm、150μm)及mmp9抗体,37℃孵育1h。pbs低速离心1000rpm8min洗涤3次后,上机检测。
2.6明胶酶谱实验。
取对数期的jeko-1细胞在的无血清培养基中培养24h。次日收集上清液,将上清液移入离心管中2000rpm离心10min,-70℃储存备用。根据细胞计数调整各组细胞培养上清液中的蛋白浓度。与5×上样缓冲液混合,13ul样本+4ul上样缓冲液。配制分离胶和浓缩胶,16ul/孔上样,4℃进行sds-page电泳100v约1.5h。电泳结束后,将凝胶置于洗脱液中振荡洗脱2次,然后用漂洗液漂洗2次,后将凝胶置于孵育液中37℃孵育42h。孵育结束后经染色液染色3h,及脱色液a、b、c(甲醇浓度分别为30%、20%、10%,乙酸浓度分别为10%、10%、5%)分别脱色0.5、1、2h后,显示出mmp-9(92kd)为位于蓝色背景上的透亮带。
2.7mmp9elisa检测。
细胞上清样品至少要求进行100倍稀释后检测,稀释可采用10µl样品+90µl稀释剂混合得到100倍稀释样品。从已平衡至室温的密封袋中取出微孔板,每孔加入100µl检测液。分别将不同浓度标准品,实验样本加入相应孔中,每孔100µl。用封板胶纸封住反应孔,放置震荡仪(直径为3mm)上,500rpm/min,室温孵育2h。将板内液体吸去后洗板。每孔加洗涤液400µl,然后将板内洗涤液吸去。重复操作4次。最后一次洗板结束,将板内所有液体吸干。在每个微孔内加入200µl酶标检测抗体。用封板胶纸封住反应孔,放置震荡仪(直径为3mm)上,500rpm/min,室温孵育1h,洗板后在每个微孔内加入200µl显色底物,室温孵育30min。在每个微孔内加入50µl终止液1,孔内溶液颜色会从蓝色变为黄色。加入终止液1后30min内,使用酶标仪测量450nm的吸光度值,设定540nm或570nm作为校正波长。计算结果:每一个标准品及样本的复孔校正吸收光值(od450-od540/od570)取平均值,再减去标准曲线零点的od值。利用酶标仪携带的软件,绘制一个4参数(4-pl)线性标准曲线,曲线横坐标为标准曲线点的人mmp-9浓度值,纵坐标为标准曲线点的od平均值。通过样本的od值,可从标准曲线上得到样本中人mmp-9的浓度。
2.8细胞免疫荧光实验。
将jeko-1细胞于4℃1000rpm离心5min,吸去培养基后pbs重悬。pbs重悬3次后离心吸去pbs,以1ml4%多聚甲醛重悬细胞,固定30min。离心。吸去固定液,pbs重悬细胞3次。加入1ml100μmm34℃放置1h。pbs清洗3次。加入dil2μl30min后pbs清洗2次。离心细胞并以少量pbs重悬,将细胞悬液滴于载玻片上,加上盖玻片,激光共聚焦显微镜(nikon,japan)下观察荧光强度。
2.9cck-8细胞活性检测。
取对数生长期的jeko-1细胞接种于6孔板,每孔接种1×105个细胞,置于37℃、5%co2培养箱中过夜,通过entransertm-r将0.67μgsi-h-mmp9转染细胞24h后,将细胞以4×103个细胞/孔的密度接种到96孔板上,每组细胞设3个复孔,分别于24h、48h、72h收集细胞。每孔分别加入10μlcck-8(dojindomoleculartechnologiesinc.,japan)处理1h。使用多模式读取酶标仪(ld942,北京,中国)测量450nm处吸光度。以时间为横轴,吸光度为纵轴绘制曲线。
2.10transwell实验。
在24孔板中放置含有8μm微孔的transwell小室,上室中铺有1:43.5稀释的matrigel胶。细胞转染24h后,取细胞悬液200μl加入上室,细胞数均为1×104/孔,下室加入含30%fbs的培养液800μl。将24孔板置于37℃、5%co2、饱和湿度条件下培养24h。收集下室细胞悬液,取10μl加入到总体积100μl的pbs中(滴入一滴台盼蓝染料)。从稀释后的细胞悬液中取出10μl加入计数板中,显微镜下计数。
2.11流式细胞术检测细胞周期与凋亡。
常规方法计数并收集细胞(约2×106),pbs洗涤一次(1000rpm,5min)。
应用预冷75%乙醇固定,4℃孵育过夜。弃去75%乙醇,pbs洗涤一次(1000rpm,5min)。重悬细胞于800ul1×pbs+1%bsa溶液中,加入100ulpi染液(3.8×10-2sodiumcitrate,ph7.0),加入100ulrna酶(rnasea,10mg/ml),37℃避光孵育30min,上机检测。各放大器接收信号的hcv值<2%,采用cellmodifit软件进行分析。细胞增殖指数pi=(s+g2/m)/(s+g2/m+g0/g1),pi指数代表细胞增殖水平。亚g0细胞比例代表细胞凋亡百分比。
3.实验结果。
3.1设计模板肽段与候选肽段。
mmp9通常存在于酶原形式(prommp9),pdb数据库中prommp9结构包括前肽、催化域及三纤连蛋白ii型(fnii)f域,如图1所示。根据prommp9激活过程,从前肽中筛选出氨基酸序列tprcgvpdl作为模板肽段(m0),其中包含97-100位关键氨基酸残基(prcg)。根据氨基酸理化性质优化关键氨基酸残基,替换方案如图2所示,设计三十余种候选肽用于后续对接筛选。
3.2候选肽m3的对接结果优于模板肽m0。
moe分子对接程序中设定每个肽段保存十种构象,选择s值最小的构象为最优构象。根据s评分,肽段m3的最优构象被认为结合较稳定其次是m1和m2,见表1。该三种肽段的最优构象进一步进行相互作用分析发现氢键和π-π共轭相互作用。结果表明,m1、m2及m3均能与活性中心的zn2+结合,影响酶活性。m3除了具有最优s值外,还与激活型mmp9的活化中心的三个氨基酸残基(pro193、his405和leu409)产生相互作用。因此,m3可以作为抑制激活型mmp9的候选药物。此外,m1的s值评分排在第二位,它与活性中心氨基酸his411有潜在的相互作用。其他肽段(m0和m2)与活性位点没有潜在的相互作用,但与催化域的活性残基有疏水性作用。氨基酸残基与活性位点的相互作用见表1。受体与配体之间的相互作用如图3-6所示。配体与受体蛋白的结合方式如图7所示。固相合成法合成m0、m1、m2及m3四种多肽,结构式及多肽性质预测如图8-11、图12-15所示。
3.3mmp9靶向抑制肽m3可与套细胞淋巴瘤细胞亲和,并抑制mmp9活性。
3.3.1流式细胞术鉴定四种荧光合成肽与jeko-1的结合特异性。
为了鉴定m0、m1、m2及m3肽与套细胞淋巴瘤细胞系jeko-1结合的特异性及寻找各荧光肽与jeko-1的优势结合,本部分实验通过流式细胞技术检测荧光修饰肽与jeko-1细胞的结合情况。通过设定不同的孵育时间,选取最适的孵育时间1h。
图16-19结果显示四种荧光修饰多肽m0、m1、m2及m3均与jeko-1细胞有特异性结合,随着肽浓度的增加,结合率增高,但都低于mmp9抗体结合率。当加入mmp9无荧光抗体后再加入150μmm3时,m3结合率明显降低,如图19所示。
3.3.2靶向抑制肽m3可特异性抑制mmp9活性。
如图20-21所示,随着肽m0和m3浓度的增加(5μm、50μm、100μm及150μm),mmp9活性逐渐降低,且m3对mmp9活性的抑制作用强于m0。然而,这两种多肽的抑制作用都弱于mmp9抑制剂。而加入肽m1及m2后细胞mmp9活性并未出现明显改变。每组jeko-1细胞添加不同浓度m3后mmp9表达量并未出现明显改变如图22所示,说明多肽m3只抑制mmp9的活性而没有降低其表达水平。将100μmm3与jeko-1细胞共培养48h后,应用细胞免疫荧光检测m3与细胞亲和性,证实m3确实可与jeko-1细胞亲和,如图23所示。
mmp2也属于mmps家族,与mmp9结构相似。实验结果提示分泌到无血清培养基中的mmp2水平没有受影响,如图20-21所示。结果表明,m3对mmp9具有特异性。
3.4mmp9靶向抑制肽m3可抑制套细胞淋巴瘤细胞增殖、凋亡及侵袭迁移。
为了进一步验证肽m3在mcl细胞系中的功能,将jeko-1细胞分为mmp9抗体组、50μmm3组及100μmm3组进行细胞增殖功能测定。随着jeko-1细胞中m3浓度的增加,细胞增殖受到明显抑制,呈剂量依赖性关系,抑制程度在48h时最为明显,如图24所示。
应用流式细胞术检测细胞凋亡和细胞周期,评估m3对mcl细胞功能影响。细胞凋亡检测,jeko-1细胞分别加入50、100μmm3或mmp9抗体培养48h。流式细胞仪分析显示,50及100μmm3jeko-1细胞组凋亡细胞为12.42%及20.31%,而mmp9抗体组为25.87%。在细胞周期分析中,随着m3浓度从0增加到50或100μm,jeko-1细胞增殖指数从0.51下降到0.48或0.46如图25-29所示。
侵袭实验中,jeko-1细胞分别加入mmp9抗体、50及100μmm3培养1h后转移到提前放置于24孔板的包被明胶的小室中继续培养24小时。m3组(50及100μm)显著地抑制jeko-1细胞侵袭能力(54.5%及63.6%),但抑制效果不如mmp9抗体明显。迁移实验中,50及100μmm3组jeko-1细胞迁移能力下降27.3%及36.4%。同样,这种抑制作用也不如mmp9抗体显著如图30-31所示。
sequencelisting
<110>中国医科大学
<120>一种抑制mmp9活性的多肽及其应用
<130>1
<160>1
<170>patentinversion3.5
<210>1
<211>10
<212>prt
<213>artificialsequence
<400>1
thrphelysgluprovalproaspleucys
1510
- 还没有人留言评论。精彩留言会获得点赞!