利用β-1,6-葡聚糖酶制备低聚龙胆糖的方法及其应用与流程
本发明涉及利用β-1,6-葡聚糖酶制备低聚龙胆糖的方法及其应用,属于基因工程和发酵工程技术领域。
背景技术:
低聚龙胆糖是葡萄糖以β-1,6糖苷键连接形成的功能性低聚糖,包括龙胆二糖和少量的三、四糖。低聚龙胆糖不被人体肠道消化吸收,但却有利于双歧杆菌和乳酸菌的繁殖和生长,适合糖尿病等人群食用,其高保湿性能有利于保持食品中的水分,可防止淀粉老化;同时低聚龙胆糖耐热性高,适用于需要高温处理的食品;其成分中的龙胆三糖具有独特的、柔和的柔和的提神苦味,具有良好的保健效果。目前,低聚龙胆糖被广泛运用于巧克力、冰淇凌、咖啡、调味品、烘烤食品和饮料中。
低聚龙胆糖酶法生产相关研究主要集中在利用β-葡萄糖苷酶以葡萄糖为底物通过逆水解缩合作用形成产物,但目前报道的β-葡萄糖苷酶在低聚龙胆糖生产过程中主要存在三个问题,一是低聚龙胆糖产率低,二是产物中只含有龙胆二糖,未检测到龙胆三糖及更高聚合度的低聚龙胆糖组分,三是用酶成本高。研究资料显示,龙胆三糖和龙胆四糖具有比龙胆二糖更好的促双歧杆菌益生活性。
β-1,6-葡聚糖酶(ec3.2.1.75)是一种可水解β-1,6葡聚糖的糖苷水解酶,水解终产物为龙胆二糖。然而,部分研究显示,自然界中还存在部分β-1,6-葡聚糖酶对龙胆二糖具有微弱的水解活性,可利用龙胆二糖为底物转苷合成龙胆三糖甚至四糖。如fujimoto等以聚合度为2~6的低聚龙胆糖为底物测试β-葡聚糖酶时发现,随着聚合度的降低,酶对底物的水解活力也会随之下降,假定对六糖的水解活力为100%,那么对于二糖则不到1%,而只有对二糖具备一定水解活力的酶,才能够以龙胆二糖为底物合成更高聚合度的低聚龙胆糖。由此可见,在目前的生产和研究中,对于如何制备聚合度更高的龙胆三糖仍然是一个亟待解决的问题。
技术实现要素:
为解决目前存在的问题,本发明提出一种利用β-1,6-葡聚糖酶制备低聚龙胆糖的方法及其应用,提高低聚龙胆糖的转化率和产物中龙胆三糖的占比,提升产品品质。
本发明的第一个目的是提供一种利用β-1,6-葡聚糖酶单酶法制备低聚龙胆糖的方法。
在本发明的一种实施方式中,所述方法是以葡萄糖为底物。
在本发明的一种实施方式中,所述β-1,6-葡聚糖酶是来源于解纤维素蓝状菌talaromycescellulolyticus的β-1,6-葡聚糖酶tcbgn,其氨基酸序列如seqidno.1所示。
在本发明的一种实施方式中,以70~90%的葡萄糖为底物,β-1,6-葡聚糖酶加酶量为400~1600u/g葡萄糖,在ph3.5~4.5、温度40~50℃下进行酶反应,反应48~96h。
本发明的第二个目的是提供一种利用β-葡萄糖苷酶和β-1,6-葡聚糖酶协同制备低聚龙胆糖的方法。
在本发明的一种实施方式中,所述方法是以葡萄糖为底物。
在本发明的一种实施方式中,所述β-葡萄糖苷酶和β-1,6-葡聚糖酶均来源于解纤维素蓝状菌talaromycescellulolyticus;所述β-葡萄糖苷酶为β-葡萄糖苷酶tcbgl3a;所述β-1,6-葡聚糖酶为β-1,6-葡聚糖酶tcbgn;β-1,6-葡聚糖酶tcbgn的氨基酸序列如seqidno.1;β-葡萄糖苷酶tcbgl3a的氨基酸序列如seqidno.2所示。
在本发明的一种实施方式中,以70~90%葡萄糖为底物,β-葡萄糖苷酶加酶量为200~600u/g葡萄糖,同时β-1,6-葡聚糖酶加酶量为400~800u/g葡萄糖,在ph4.0~5.0、温度50~60℃下反应36~60h。
在本发明的一种实施方式中,所述低聚龙胆糖包括龙胆二糖和龙胆三糖。
本发明第三个目的是提供一种提高龙胆三糖产量的方法,所述方法以葡萄糖为底物。
在本发明的一种实施方式中,加入氨基酸序列如seqidno.1所示的β-1,6-葡聚糖酶。
在本发明的一种实施方式中,加入氨基酸序列如seqidno.1所示的β-1,6-葡聚糖酶,和氨基酸序列如seqidno.2所示的β-葡萄糖苷酶。
本发明的第四个目的是提供表达所述β-1,6-葡聚糖酶的基因工程菌,所述基因工程菌以毕赤酵母为宿主,表达氨基酸序列如seqidno.1所示的β-1,6-葡聚糖酶。
本发明的第五个目的是提供一种构建所述基因工程菌的方法,所述方法是将核苷酸序列为seqidno.3的编码β-1,6-葡聚糖酶的基因连接到表达载体ppic9k上,得到重组质粒;将重组质粒转化至毕赤酵母km71中,得到表达β-1,6-葡聚糖酶的基因工程菌。
本发明的第六个目的是提供一种生产β-1,6-葡聚糖酶的方法,所述方法包括:(1)分批发酵阶段:将种子液以8%-12%接种量接种于发酵罐中,控制温度28-30℃、初始转速180-220rpm、初始通气量7l/min、溶氧28-32%、ph4.5-5.5;(2)补料发酵阶段:待溶氧上升至80-100%,以恒速流加甘油的方式进行补料培养,控制温度28-30℃、溶氧28-32%、ph4.5-5.5。
在本发明的一种实施方式中,当菌体od600在150~200,添加体积比为1~1.5%的甲醇、在25~30℃下诱导产酶,同时控制溶氧28-32%、ph4.5-5.5,诱导96-144h。
本发明还保护所述利用β-1,6-葡聚糖酶单酶法制备低聚龙胆糖的方法,或利用β-葡萄糖苷酶和β-1,6-葡聚糖酶协同制备低聚龙胆糖的方法,或提高龙胆三糖产量的方法,或表达所述β-1,6-葡聚糖酶的基因工程菌,或生产β-1,6-葡聚糖酶的方法在制备低聚龙胆糖中的应用。
本发明的有益效果:
目前已报道以葡萄糖为底物酶法制备低聚龙胆糖最高水平为,绿色木霉trichodermaviride来源的β-葡萄糖苷酶在80%葡萄糖的底物浓度下,加酶量为900u/g葡萄糖,制备低聚龙胆糖转化率为16.25%。
本发明所提供的β-1,6-葡聚糖酶tcbgn能以葡萄糖为底物单酶制备低聚龙胆糖,在加酶量400~800u/g葡萄糖时,低聚龙胆糖转化率可达19.21%~19.84%;在最佳反应条件下,以80%葡萄糖为底物制备低聚龙胆糖转化率可达到19.96%,高于目前已知报道以葡萄糖为底物单酶制备低聚龙胆糖的最高转化率,并且,三糖的比例可达1.35%~1.93%。
本发明还提出了一种利用β-1,6-葡聚糖酶和β-葡萄糖苷酶双酶复配制备低聚龙胆糖的方法,以上述β-1,6-葡聚糖酶和β-葡萄糖苷酶构建协同反应体系,在最佳反应条件下,以80%葡萄糖为底物,低聚龙胆糖转化率最高可达到23.83%,且产品组分中龙胆三糖含量明显增加,三糖比例可达2.97%~4.00%,为目前报道中以葡萄糖为底物制备得到的最高产率,极大提升了产品品质。
附图说明
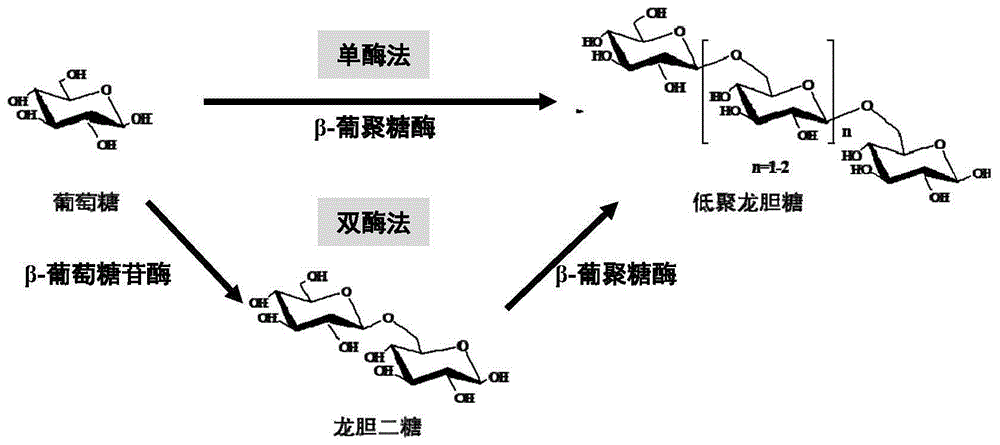
图1是单酶法或双酶法制备低聚龙胆糖的方法示意图。
图2是重组β-葡萄糖苷酶tcbgl3a在3.6l罐发酵上清酶活曲线和sds-page电泳图。
图3是重组β-1,6葡聚糖酶tcbgn在3.6l罐发酵上清酶活曲线和sds-page电泳图;
图4是温度和ph对重组β-葡萄糖苷酶tcbgl3a酶活性的影响。
图5是温度和ph对重组β-1,6葡聚糖酶tcbgn酶活性的影响。
具体实施方式
培养基:
(1)md固体培养基:ynb13.4g/l,生物素4×10-4g/l,葡萄糖20g/l,琼脂20g/l。
(2)ypd培养基:蛋白胨20g/l,酵母提取物10g/l,葡萄糖20g/l,固体培养基添加20g/l琼脂
(3)bmgy培养基:ynb13.4g/l,甘油10g/l,生物素4×10-4g/l,0.1mol/l磷酸钾缓冲溶液(ph6.0),蛋白胨20g/l,酵母粉10g/l
(4)bmmy培养基:ynb13.4g/l,甲醇1%,生物素4×10-4g/l,0.1mol/l磷酸钾缓冲溶液(ph6.0),蛋白胨20g/l,酵母粉10g/l
(5)发酵种子培养基:酵母粉5.0g/l,胰蛋白胨10.0g/l,葡萄糖10.0g/l,甘油30g/l。
(6)bsm培养基:85%磷酸26.7ml/l,caso40.93g/l,k2so418.2g/l,mgso4·7h2o14.9g/l,koh4.13g/l,甘油30.0g/l,微量元素盐溶液4.32ml/l。
(7)生长阶段补料培养基:80%甘油,4.92ml/l微量元素液。
(8)诱导阶段补料培养基:100%甲醇,12.5ml/l微量元素液。高密度发酵用100%氨水和50%磷酸来调节ph。
β-葡萄糖苷酶酶活力分析:
(1)酶活单位定义
每毫升酶液每分钟水解pnpg产生1μmol的对硝基苯酚的酶活力为一个酶活单位。
(2)酶活力测定步骤
反应体系为1ml,ph5.0的醋酸缓冲液960μl,加入适度稀释的粗酶液20μl,再加入20μl100mmol/l的pnpg,在60℃恒温水浴中反应10min,10min后立即加入200μl的1mol/l的na2co3溶液终止反应,冰浴5min,于405nm处测光吸收值。以加热失活的酶液按照同样的方法处理作空白。
β-1,6-葡聚糖酶活力分析:
(1)酶活单位定义
每毫升酶液每分钟水解石耳多糖产生1μmol的葡萄糖的酶活力为一个酶活单位。
(2)酶活力测定步骤
用50mm的ph4.0柠檬酸-磷酸氢二钠缓冲液配制40g/l的石耳多糖溶液,取180μl的缓冲液并加入200μl的石耳多糖溶液混匀后置于一定温度的水浴锅中预热5min,再加入20μl一定稀释倍数的酶液,准确计时30min后加入600μldns终止反应,冰浴7min后加入2ml去离子水并于540nm处测吸光值,热处理使酶液失活并以同样的反应体系作为空白。
实施例1:β-1,6葡聚糖酶及β-葡萄糖苷酶毕赤酵母基因工程菌的构建
(1)构建β-1,6葡聚糖酶毕赤酵母基因工程菌
化学合成β-1,6-葡聚糖酶tcbgn编码基因(核苷酸序列如seqidno.3所示)与毕赤酵母表达载体ppic9k相连,得到重组质粒ppic9k-tcbgn并转入escherichiacolijm109中,酶切验证后,将重组质粒ppic9k-tcbgn电转整合到毕赤酵母km71中,然后将转化液涂布到md平板上,在md平板长出单克隆,然后挑取96个转化子转到新的md板上并用10ml规格的小管筛选酶活力较高的转化子,即为得到的β-1,6葡聚糖酶毕赤酵母基因工程菌。
pcr扩增β-1,6-葡聚糖酶tcbgn基因所用引物(下划线处为酶切位点):
f:5’-gggaattcatccacaaacgagttacaactccg-3’,(seqidno.5);
r:5’-gggcggccgcttagacagcaggcaacacccatgt-3’,(seqidno.6)。
(2)构建β-葡萄糖苷酶毕赤酵母基因工程菌
合成β-葡萄糖苷酶的编码基因(核苷酸序列如seqidno.4所示),构建所用的酶切位点、具体步骤等参照上述β-1,6葡聚糖酶毕赤酵母基因工程菌的构建,构建得到β-葡萄糖苷酶毕赤酵母基因工程菌。
实施例2:3.6l罐发酵生产β-葡萄糖苷酶和β-1,6-葡聚糖酶
(1)将实施例1制备得到的β-1,6葡聚糖酶毕赤酵母基因工程菌和β-葡萄糖苷酶毕赤酵母基因工程菌分别接种至ypd培养基中,在30℃、200rpm培养24h,培养至种子液的od600为1.3-1.5。
(2)分批发酵阶段:将种子液以8%-12%接种量接种于发酵罐中,控制温度28-30℃、初始转速180-220rpm、初始通气量7l/min、溶氧28-32%、ph4.5-5.5;
(3)补料发酵阶段:待溶氧上升至80-100%,以恒速流加甘油的方式进行补料培养,控制温度28-30℃、溶氧28-32%、ph4.5-5.5;
(4)诱导培养阶段:当菌体细胞浓度od600在100-200范围内,用甲醇流加仪流加甲醇诱导产酶,甲醇浓度控制在0.5-1.5%(v/v),控制温度20-30℃、溶氧28-32%、ph4.5-5.5,诱导96-144h。将发酵液离心取上清液,得到粗酶液。
在高密度发酵的诱导阶段,分别对初始诱导菌体od600、甲醇浓度、诱导温度做优化。
①初始诱导菌体od600的优化:设置初始诱导菌体od600为100、150、200,发酵144h测定β-1,6-葡聚糖酶酶活。
表1不同初始od600下蛋白酶酶活
②甲醇浓度的优化:在初始诱导菌体od600为150的条件下,分别设置甲醇浓度为0.5%(v/v)、1.0%、1.5%,发酵144h测定β-1,6-葡聚糖酶酶活。
表2不同甲醇浓度下β-1,6-葡聚糖酶酶活
③诱导温度的优化:在初始诱导菌体od600为150、甲醇浓度为1.0%(v/v),设置诱导温度为20、25、30℃,发酵144h测定蛋白酶活。
表3不同诱导温度下β-1,6-葡聚糖酶酶活
根据同样的方法,对β-葡萄糖苷酶的发酵条件进行优化,结果表明:在初始诱导菌体od600为150,甲醇浓度1.0%,诱导温度为25℃的条件下,β-葡萄糖苷酶的产酶效果也较其他的条件更好。
在初始诱导菌体od600为150,甲醇浓度1.0%,诱导温度为25℃最佳发酵条件下,测得β-葡萄糖苷酶在发酵108h时达到最高酶活为350u/ml,β-1,6-葡聚糖酶在发酵144h时最高酶活1795u/ml,重组β-葡萄糖苷酶的sds-page电泳图见图2,β-1,6-葡聚糖酶蛋白sds-page电泳图见图3。
实施例3:β-葡萄糖苷酶的酶学性质
将得到的β-葡萄糖苷酶酶液用上述所述的酶活测定方法进行酶学定性,以pnpg为底物,在不同的温度下测酶活,结果表明β-葡萄糖苷酶的最适温度为60℃;然后在最适温度条件下设置不同ph梯度测β-葡萄糖苷酶的酶活,得出该酶的最适ph为4.5(图4)。
实施例4:β-1,6-葡聚糖酶的酶学性质
将得到的β-葡萄糖苷酶酶液进行酶学定性,以石耳多糖为底物,在不同的温度下测酶活,结果表明β-1,6-葡聚糖酶的最适温度为45℃。然后在最适温度条件下设置不同ph梯度测β-1,6-葡聚糖酶的酶活,得出该酶的最适ph为4.0(图5)。
实施例5:β-1,6-葡聚糖酶在低聚龙胆糖制备中的应用
以80%葡萄糖为底物,在ph4.0、45℃反应条件下反应72h,设置不同β-1,6-葡聚糖酶加酶量对酶反应的影响(表4),低聚龙胆糖累积含量在一定范围内随加酶量升高而增加。当加酶量为1600u/g时,低聚龙胆糖的产量能达到159.7g/l,转化率为19.96%,高于目前已知报道以葡萄糖为底物单酶制备低聚龙胆糖的最高转化率。
表4重组β-1,6葡聚糖酶加酶量对低聚龙胆糖转化率的影响
实施例6:β-葡萄糖苷酶和β-1,6-葡聚糖酶在协同制备低聚龙胆糖中的应用
以80%葡萄糖为底物,在ph4.5、60℃反应条件下反应48h,确定β-葡萄糖苷酶的加酶量为400u/g,设置不同β-1,6-葡聚糖酶加酶量对酶反应的影响(表5),低聚龙胆糖累积含量在一定范围内随加酶量升高而增加,但当β-1,6-葡聚糖酶的加酶量达到400u/g葡萄糖时,增加加酶量反而降低低聚龙胆糖的产量。所以选择β-1,6-葡聚糖酶的加酶量为400u/g葡萄糖时最合适,此时低聚龙胆糖的产量能达到190.6g/l,转化率为23.83%,其中龙胆二糖为19.83%,龙胆三糖为4.00%。双酶法制备低聚龙胆糖转化率显著高于单酶法,且产品组分中龙胆三糖含量明显提高,产品品质提升。
表5加酶量对双酶复配低聚龙胆糖转化率的影响
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
sequencelisting
<110>江南大学
<120>利用β-1,6-葡聚糖酶制备低聚龙胆糖的方法及其应用
<160>6
<170>patentinversion3.3
<210>1
<211>470
<212>prt
<213>talaromycescellulolyticus
<400>1
ilehislysargvalthrthrproalaalatyralaserasnserasp
151015
glysertyrserleuthrserilethralaproileglnglyalaala
202530
serproglysergluserthrtrpgluleuserileaspaspserleu
354045
serglytyrlysglnthrilethrglypheglyalaalavalthrasp
505560
alathrvalthrserpheasnthrleusersersergluleusergln
65707580
leuleuasnvalleumetthrseralaglyalaasppheserleumet
859095
arghisthrileglyserseraspleuserglyaspproalatyrthr
100105110
tyraspaspasnglyglyalavalaspthrsermetserglypheasn
115120125
leuglyaspargglythralametalaglumetleualalysmetlys
130135140
serleuglnseraspleulysvalleuglysersertrpserpropro
145150155160
glytrpmetlysleuasnseralaileaspglythrthrthrasnasn
165170175
asnleuasnaspglytyrleuglythrglyvalglyseralaglytyr
180185190
sersergluphealaglntyrphevallystyrileglnalatyrglu
195200205
alaleuglyalaasnileaspalailethrileglnasngluproleu
210215220
asnserglnalaglytyrprothrmettyrmetpheasptyrglugln
225230235240
glyaspleuileglnsertyrileglyproalaleualaasnalagly
245250255
leuaspthralavaltrpalatyrasphisasnthrasnvalproser
260265270
pheproglnasnvalleuaspthralaserglntyrvalaspthrval
275280285
alatrphiscystyraspaspserleuasptrpservalleuthrasp
290295300
phelysasnserasnproglyvalthrglntyrmetthrglucystrp
305310315320
thrproalaserglyalatrptyrglnalaserasnphethrilegly
325330335
proleuglnasntrpalaserglyvalmetalatrpthrleuglythr
340345350
aspserserasnglyprohisleuserserglyglycysaspthrcys
355360365
glnglyleuvalthrileasnseraspglythrtyrthrleugluarg
370375380
alatyrtyrmetmetalaglntyrserlyspheileprothrglyala
385390395400
ileileleuaspglyserglysertyrthrtyrserglyvalglygly
405410415
ileglnservalalaserleuasnproaspglythrargthrvalval
420425430
ileglnasnthrpheserasnaspvaltyrvalthrvalserthrser
435440445
serglyglnglutrpserglyasnileprothrgluservalvalthr
450455460
trpvalleuproalaval
465470
<210>2
<211>795
<212>prt
<213>talaromycescellulolyticus
<400>2
glnseralasertrpseralaalatyrserlysalathralaalaleu
151015
serlysleuserglnasnasplysileglymetvalthrglyvalgly
202530
trpglylysglyprocysvalglyasnthralaalaproserglyile
354045
serpheproserleucysileglnaspserproleuglyvalargtyr
505560
alaasnprovalthralapheproalaglythrasnalaglymetthr
65707580
trpaspargthrleumetasnglnargglyalaalaleuglyalaglu
859095
serlysglyleuglyvalhisvalglnleuglyprovalalaglypro
100105110
leuglylysilealaglnglyglyargglytrpgluglypheglythr
115120125
aspprotyrleuserglyvalalametilegluthrileserglymet
130135140
glnserserglythrglnalacysalalyshistyrileglyasnglu
145150155160
glngluleuasnargglusermetserserasnileaspaspargthr
165170175
leuhisgluleutyrleutrpprophealaaspalavalargalaasn
180185190
valalaservalmetcyssertyrasnglnileasnglythrpheser
195200205
cysgluasngluglusermetthrglyileleulysthrgluleugly
210215220
pheproglytyrilemetserasptrpaspalaglnhisthrthrval
225230235240
thrseralaasnserglyleuaspmetthrmetproglyserasptyr
245250255
seraspthrproserservalleutrpglyglnasnleualaasnala
260265270
ileserserglyglnvalalaglnserargleuaspaspmetvalthr
275280285
argileleualaalatrptyrleuvalglyglnaspglnglyphepro
290295300
alavalalapheasnsertrpthrglyglyglnalaservalasnval
305310315320
thrserasnhisasnglnvalalaargalavalalaargaspserile
325330335
valleuleulysasnthrasnserthrleuproleuasnlysproser
340345350
serilealaileileglythraspalaglnthrasnproserglypro
355360365
asnalacysthraspargglycysaspthrglythrleualametgly
370375380
trpglyserglythrcysglnpheprotyrleuthraspproleuthr
385390395400
alailelysthrargalaalaseraspglythrthrilethrthrser
405410415
ileseraspasnglyseralaglyalaservalalaglnseralaglu
420425430
tyralailevalpheileasnseraspserglygluglytyrilethr
435440445
valgluglyvalalaglyaspargasnasnleuaspprotrphisser
450455460
glyasnalaleuvalglnservalalaalavalasnlyslysthrile
465470475480
valvalilehisservalglyprovalileleugluthrileleuala
485490495
glnproasnvalvalalavalvaltrpalaglyileproglyglnglu
500505510
serglyseralaleuthraspileleutyrglyserthralaproser
515520525
glylysleuthrtyrthrilealalysglnalaserasptyrglythr
530535540
alavalvalserglyseraspasntyrprogluglyleupheileasp
545550555560
tyrarghispheasplysserasnilegluproargtyrgluphegly
565570575
tyrglyleusertyrthrthrpheglytyrthrasnleualaileasp
580585590
ilethrvalserthrglyprothrthrglyglnilevalproglygly
595600605
proseraspleuphegluservalglythrvalthrvalglnvalala
610615620
asnthrglyservalalaglysergluvalalaglnleutyrilegly
625630635640
leuproserseralaproserserproprolysglnleuargglyphe
645650655
asplysleuserleualaalaglyalaserglythralathrpheasp
660665670
leuthrargargaspleusertyrtrpaspvalserlysglnlystrp
675680685
valvalproserglyalaphethrvaltyrvalglyalaserserarg
690695700
aspileargleuglnglythrphethrproglyglyserserthrthr
705710715720
serthrilethrserserlysthrserthrthrileserthrserval
725730735
thrthrserserserthrthralalysthrthrthrthrserserthr
740745750
thrserseralaglyprothrglnthrprotyrglyglncysglygly
755760765
glnglytrpthrglyprothrvalcysserserglytrpthrcyslys
770775780
valthrasnglntrptyrserglncysleugln
785790795
<210>3
<211>1413
<212>dna
<213>人工序列
<400>3
attcataaaagggttacaacaccagcagcttatgctagtaactcagatggcagttactcc60
cttacgtcaataacagcccctatccagggagccgcttcacccggttccgagtcaacttgg120
gaactttccatagacgactcattgagtggctacaaacagaccatcaccggcttcggtgcc180
gctgttactgatgcaaccgtcacgtcctttaatactctgtcaagttctgagctgtctcag240
ttgttaaatgttttaatgacatccgccggcgccgacttttccctgatgagacatacgatc300
ggttcaagtgatttgtctggcgatcctgcctatacttatgatgacaatggaggcgcagtt360
gacacttccatgtcaggtttcaacttgggagaccgtggcacggcaatggccgaaatgttg420
gcaaaaatgaagagtctgcagtctgacctgaaagtcttgggctcatcttggtccccccca480
ggatggatgaaacttaactctgctatcgatggaaccacaacaaacaacaacttaaatgac540
ggttatttaggtacgggagtcggttctgccggttactcctctgagtttgcccagtatttc600
gtcaaatatatacaagcctatgaagccttaggcgcaaatatagacgcaattacaattcaa660
aatgaacccctaaactcccaggccggctaccctacaatgtacatgtttgactatgagcaa720
ggcgaccttatacagtcttatatcggtcccgcattggcaaatgcaggtttggacacggca780
gtgtgggcctacgaccataacaccaacgtgccatcattcccccaaaatgtgttggatacg840
gcatcccaatacgttgacacggttgcctggcattgttatgatgacagtcttgattggtca900
gtattgactgacttcaagaactctaaccccggagtaacacagtacatgacagaatgttgg960
accccagcatctggtgcatggtatcaggcctccaattttaccataggcccactacaaaat1020
tgggcaagtggcgtgatggcctggactctgggtactgactcatctaatggtccccatctg1080
tccagtggtggctgtgatacgtgtcaaggtctggtgacgatcaactcagatggtacatat1140
acattggagagagcttactatatgatggcacagtactcaaaatttattcctacgggagcc1200
atcattctggacggctcaggttcctacacctatagtggagtcggtggtattcagtcagta1260
gcctccttgaaccctgatggtaccaggacggttgtcatacaaaatacatttagtaacgac1320
gtttacgttacagtcagtacttcctccggtcaggagtggtcaggcaatatacctaccgag1380
tccgtagtcacctgggttttgccagcagtgtaa1413
<210>4
<211>2388
<212>dna
<213>人工序列
<400>4
caatccgctagttggtctgcagcatattctaaggctacagcagccctttccaagctgtcc60
caaaacgataaaataggaatggttacgggagtgggatggggcaaaggcccctgtgtgggt120
aacactgccgctcccagtggcatctctttcccctccttatgcatacaggattcaccctta180
ggcgtaagatacgccaatcccgtaactgccttccccgcaggaaccaacgcaggcatgacg240
tgggacaggactctgatgaaccagaggggcgcagctcttggtgccgaaagtaagggccta300
ggtgtgcatgtgcaactaggacccgtcgccggcccacttggcaaaattgcacaaggaggt360
aggggttgggaaggctttggaacggacccatacctgtctggtgtagcaatgattgaaaca420
atatctggtatgcaatcatcaggaacccaagcctgcgccaaacattatataggtaatgag480
caagagctaaaccgtgaatctatgtctagtaatatagatgacagaacgctacacgaattg540
tatctatggcccttcgctgatgccgttagggctaacgtcgcctccgttatgtgctcttac600
aaccaaatcaatggaacattctcatgtgaaaacgaagaaagtatgacaggtattctaaag660
acggaattaggattccccggatatatcatgtcagattgggacgcacaacacacaactgtt720
acttccgctaatagtggcctagatatgacgatgccaggcagtgattactctgacactccc780
tcttccgttctttggggccagaacttggccaatgccatatcatctggacaagtcgcccag840
agtaggttagacgacatggtcacacgtatcttagccgcatggtatctagtcggacaagat900
caaggatttcccgcagtcgcttttaatagttggacaggaggacaagcatctgttaatgtc960
accagtaatcataaccaagtagccagggctgttgccagagactccattgtgttattgaaa1020
aacaccaattcaacgctgccattaaacaaaccctcttctatagcaattatcggcacggat1080
gcccagaccaatccctctggacccaatgcatgcacggataggggttgtgatactggaacc1140
ttagctatgggttggggctccggtacgtgtcagttcccatatttgacagacccattgaca1200
gctatcaagacaagagcagcatccgacggaactacaatcacaactagtatttccgataat1260
ggctccgcaggcgctagtgtggcccagtccgctgagtacgcaattgtatttatcaattcc1320
gattctggagaaggttacattacagtcgaaggagtggcaggtgacaggaataatcttgat1380
ccctggcactcaggaaacgcccttgtccaaagtgtggccgctgtgaacaaaaaaacaata1440
gtggtaattcatagtgtgggccccgtaatcttagagactatattggcccagcccaatgtg1500
gtagcagtcgtttgggcaggtatacctggacaggagagtgggtccgccttaactgacatt1560
ctttacggaagtactgctcctagtggtaagctaacttatactattgcaaaacaggcttct1620
gattacggtacagcagtcgtgtcaggttcagataattaccccgaaggtttattcattgat1680
taccgtcacttcgacaaaagtaatattgagcccagatacgagttcggttacggcttgtca1740
tatacgacatttggatacactaacctagcaatcgatataacggtaagtaccggccccact1800
accggccaaatagtaccaggaggacccagtgatctttttgagtcagtgggcaccgtaacc1860
gtccaggtagctaatacagggtccgtagcaggctccgaagtagctcagttatatatcgga1920
ctgccctccagtgcaccatcatccccaccaaagcaactacgtggcttcgataagctgtcc1980
cttgctgccggtgctagtggaacagctacttttgatttgacccgtcgtgatctatcatat2040
tgggatgtttcaaaacagaagtgggttgtcccttctggtgcttttaccgtttacgtgggt2100
gcatcttctagggacattagactgcagggtacctttacacccggtggatctagtacaaca2160
tccacgatcacgtcaagtaaaacgtccactaccatctccacatctgttactacctcatca2220
tcaacaacggcaaagaccacaaccacgtcatccaccacatcttcagcaggaccaacccag2280
actccatacggacagtgcggcggtcagggttggacgggtcccacggtatgttcatcaggt2340
tggacatgtaaagtaactaatcaatggtactcccagtgtctgcagtaa2388
<210>5
<211>32
<212>dna
<213>人工序列
<400>5
gggaattcatccacaaacgagttacaactccg32
<210>6
<211>34
<212>dna
<213>人工序列
<400>6
gggcggccgcttagacagcaggcaacacccatgt34
- 还没有人留言评论。精彩留言会获得点赞!