一种脂肪酸延长酶基因在酵母合成神经酸中的应用的制作方法
1.本发明涉及生物技术领域,更具体地涉及一种脂肪酸延长酶基因在酵母合成神经酸中的应用。
背景技术:
2.神经酸(cis-15-tetracosenoic acid,24:1δ15)是一种超长链单不饱和脂肪酸(very long chain monounsaturated fatty acids,vlcmfas),主要以鞘糖脂和鞘磷脂的形式存在于动物大脑白质和髓鞘的神经纤维中,是生物膜的重要组成成分。神经酸在医学和保健方面具有重要的作用,具有促进大脑发育、改善记忆、调节血脂、增强免疫等功效,可用于治疗多发性硬化症等神经紊乱病症。此外,研究表明神经酸对神经系统的发育具有促进作用,尤其是在婴幼儿脑神经细胞和视神经细胞生长与发育过程具有重要的作用。
3.目前,木蜡酸(c24:0)、神经酸等超长链脂肪酸的合成在植物中认识较为清楚。植物以油酸为底物,通过超长链脂肪酸链延长循环进行碳链的延长,每次循环能够使原有脂肪酸的碳链延长两个碳原子。这个延长过程受4个酶(kcs、kcr、hcd和ecr)形成的酶复合体的组合催化和调控,目前普遍认为其中的kcs(3-酮酯酰-coa合酶)是决定植物组织中超长链脂肪酸链长和合成速率的关键酶。
4.虽然神经酸用途广泛,对人体健康十分有益,但难以通过人体自身合成,需要从外界摄取。目前通过化学方法合成神经酸,因副产物多、工艺路线长、得率低,价格超过1000美元每公斤,不具有市场竞争力。神经酸市场需求巨大,但植物作为主要来源因生长缓慢,生长环境要求高等特点限制了神经酸的产量,通过微生物生产神经酸具有广阔前景。然而,已知的产神经酸微生物大部分具有致病性或者生产潜力不足,导致不能应用于工业化生产。
5.因此,本领域迫切想要开发一种能够高产神经酸的新方法。
技术实现要素:
6.本发明的目的在于提供一种能够高产神经酸的新方法。
7.在本发明的第一方面,提供了一种用于生产神经酸的工程菌,所述工程菌的基因组中整合有基因表达盒,所述的基因表达盒表达3-酮酯酰-coa合酶(kcs)基因所编码的蛋白。
8.在另一优选例中,所述工程菌包括酵母菌。
9.在另一优选例中,所述工程菌选自下组:解脂耶氏酵母、酿酒酵母、甲醇酵母、毕赤酵母、汉逊酵母、克鲁维酵母、假丝酵母、隐球酵母、东方伊萨酵母、红酵母、或其组合。
10.在另一优选例中,所述3-酮酯酰-coa合酶基因来源于蒜头果(malania oleifera)、碎米芥(cardamine graeca)、元宝枫(acer truncatum)、银扇草(lunaria annua)、海檀木(ximenia caffra)、旱金莲(tropaeolum speciosum)、文冠果(xanthoceras sorbifolia)、油菜(brassica)、荠菜(capsella)。
11.在另一优选例中,所述工程菌为解脂耶氏酵母(yarrowia lipolytica),较佳地,
为解脂耶氏酵母yarrowia lipolytica po1g-g3-maolekcs。
12.在另一优选例中,所述的基因组中整合有1-20个(较佳地,1-10个,更佳地,1-5个)所述的基因表达盒。
13.在另一优选例中,所述基因表达盒整合于选自下组的一种或多种启动子:tef、tefintro、exp、gpd、gpat、yat。
14.在另一优选例中,所述3-酮酯酰-coa合酶(kcs)基因为密码子优化的3-酮酯酰-coa合酶(kcs)基因。
15.在另一优选例中,所述3-酮酯酰-coa合酶(kcs)基因的序列如seq id no.1-11中的任一所示。
16.在另一优选例中,所述的3-酮酯酰-coa合酶(kcs)基因受组成型或诱导型启动子的驱动。
17.在另一优选例中,所述的启动子选自下组:tef、tefintro、exp、gpd、gpat、yat、或其组合。
18.在另一优选例中,所述的工程菌的神经酸的产量≥20g/l,较佳地,20-30g/l。
19.在另一优选例中,所述的工程菌的发酵生物量(细胞干重)≥120g/l,较佳地,130-200g/l。
20.在另一优选例中,所述的工程菌的神经酸/总脂肪酸(神经酸/总脂肪酸)的比例≥20%,较佳地,为20-65%,更佳地,25-65%。
21.本发明第二方面提供了一种生产神经酸的方法,包括步骤:
22.(i)培养本发明第一方面所述的工程菌,从而获得含神经酸的发酵产物;和
23.(ii)从所述发酵产物中分离出神经酸。
24.本发明第三方面提供了一种构建本发明第一方面所述工程菌的方法,包括步骤:
25.(a)构建含有基因表达盒的载体,所述表达盒具有以下元件:筛选标记基因、抗性基因元件、rdna同源重组基因片段,和目的基因,所述的目的基因为3-酮酯酰-coa合酶(kcs)基因;
26.(b)将步骤(a)获得的含有所述基因表达盒的载体转入受体菌株,获得受体基因组整合有所述基因表达盒的菌株。
27.在另一优选例中,还包括步骤(c):pcr和dna测序验证步骤(b)得到的整合有基因表达盒的菌株的基因型;和/或
28.步骤(d):发酵检测得到的整合有基因表达盒菌株的神经酸的产量。
29.在另一优选例中,步骤(a)所述的载体为质粒、黏粒或核酸片段。
30.在另一优选例中,步骤(a)所述的表达盒还含有强启动子元件。
31.在另一优选例中,所述的强启动子元件包括tefintro、gpat、exp、gpd。
32.在另一优选例中,所述的强启动子元件为tefintro。
33.在另一优选例中,所述强启动子元件为1-3个,较佳地,1-2个,更佳地,为2个。
34.在另一优选例中,所述筛选标记基因选自下组:ura、leu、hgr、或其组合。
35.在另一优选例中,所述抗性基因元件包括氨苄抗性基因。
36.在另一优选例中,步骤(a)所述的表达盒具有以下元件:强启动子元件tefintro、目的基因3-酮酯酰-coa合酶(kcs)基因、包含ura、leu或hgr中的一种或多种的酵母筛选标
记基因、和氨苄抗性基因amp。
37.在另一优选例中,步骤(b)所述受体基因组整合的基因表达盒的数量为1-5个,较佳地为1-3个。
38.本发明第四方面提供了一种本发明第一方面所述工程菌株的用途,所述工程菌被用作发酵生产神经酸的菌株。
39.应理解,在本发明范围内中,本发明的上述各技术特征和在下文(如实施例)中具体描述的各技术特征之间都可以互相组合,从而构成新的或优选的技术方案。限于篇幅,在此不再一一累述。
附图说明
40.图1不同3-酮酯酰-coa合酶蛋白序列的系统树
41.图2不同3-酮酯酰-coa合酶蛋白序列比对。方框内为kcs家族的3个功能保守结构域“fgntssss”、“gsgfkcnsavw”和“gmgcsa”。
42.图3重组菌株的诊断pcr验证。a:下游pcr结果。ck为空白对照po1g-g3,数字1-8对应重组菌株po1g-g3-maolekcs398、po1g-g3-maolekcs461或po1g-g3-maolekcs467的不同转化子;b:重组菌株po1g-g3-maolekcs817的不同转化子的下游pcr结果;c:上游pcr结果。ck为空白对照po1g-g3,数字1-8对应重组菌株po1g-g3-maolekcs398、po1g-g3-maolekcs461、po1g-g3-maolekcs467或po1g-g3-maolekcs817的不同转化子。
43.图4重组菌株的摇瓶发酵结果。不同菌株间干重、总脂产量和总脂含量比较。strainck:对照菌株po1g-g3;strain398:po1g-g3-maolekcs398;strain461:po1g-g3-maolekcs461;strain467:po1g-g3-maolekcs467;strain817:po1g-g3-maolekcs817。
44.图5重组菌株的脂肪酸气相分析结果。ck:对照菌株po1g-g3;maolekcs:po1g-g3-maolekcs398。数字1-11代表不同的脂肪酸成分,依次为c16:0,c16:1,c18:0,c18:1,c18:2,c20:0,c20:1,c22:0,c22:1,c24:0和c24:1。
45.图6过表达maolekcs重组菌株的摇瓶发酵结果。a:不同菌株间干重、总脂产量和总脂含量比较。b:不同菌株的脂肪酸成分分析。strainck:对照菌株po1g-g3-maolekcs;strain#1:po1g-g3-3maolekcs398;strain#2:po1g-g3-3maolekcs461;strain#3:po1g-g3-3maolekcs467;strain#4:po1g-g3-3maolekcs817。
46.图7过表达maolekcs重组菌株的脂肪酸成分气相分析图谱。a:对照菌株po1g-g3-maolekcs;b:po1g-g3-3maolekcs398;c:po1g-g3-3maolekcs461;d:po1g-g3-3maolekcs467;e:po1g-g3-3maolekcs817。数字1-11代表不同的脂肪酸成分,依次为c16:0,c16:1,c18:0,c18:1,c18:2,c20:0,c20:1,c22:0,c22:1,c24:0和c24:1。
47.图8重组菌株的发酵罐发酵结果。
48.图9重组菌株发酵罐发酵168h的脂肪酸成分气相分析图谱。数字1-11代表不同的脂肪酸成分,依次为c16:0,c16:1,c18:0,c18:1,c18:2,c20:0,c20:1,c22:0,c22:1,c24:0和c24:1。
具体实施方式
49.本发明人经过广泛而深入地研究,首次意外的构建一种酵母工程菌,所述工程菌
的基因组中整合有基因表达盒,所述的基因表达盒表达3-酮酯酰-coa合酶(kcs)基因所编码的蛋白,本发明的工程菌可显著提高神经酸的产量。在此基础上完成了本发明。
50.术语
51.除非另外定义,否则本文中所用的全部技术与科学术语均具有如本发明所属领域的普通技术人员通常理解的相同含义。
52.如本文所用,在提到具体列举的数值中使用时,术语“约”意指该值可以从列举的值变动不多于1%。例如,如本文所用,表述“约100”包括99和101和之间的全部值(例如,99.1、99.2、99.3、99.4等)。
53.如本文所用,术语“含有”或“包括(包含)”可以是开放式、半封闭式和封闭式的。换言之,所述术语也包括“基本上由
…
构成”、或“由
…
构成”。
54.如本文所用,术语“以上”和“以下”包括本数,例如“20%以上”指≥20%,“8:1以上”指≥8:1。
55.出发菌株
56.如本文所用,术语“本发明出发菌株”或“本发明出发微生物”可以互换使用,都是指解脂耶氏酵母y.lipolytica po1g(mata,leu2-270,ura3-302::ura3,xpr2-3),购自yeastern biotech co.,ltd。
57.应理解,出发菌株不仅包括本发明的解脂耶氏酵母y.lipolytica po1g,还包括其衍生菌株。
58.神经酸
59.神经酸(cis-15-tetracosenoic acid,24:1δ15)是一种超长链单不饱和脂肪酸(very long chain monounsaturated fatty acids,vlcmfas),主要以鞘糖脂和鞘磷脂的形式存在于动物大脑白质和髓鞘的神经纤维中,是生物膜的重要组成成分。神经酸在医学和保健方面具有重要的作用,具有促进大脑发育、改善记忆、调节血脂、增强免疫等功效,可用于治疗多发性硬化症等神经紊乱病症。此外,研究表明神经酸对神经系统的发育具有促进作用,尤其是在婴幼儿脑神经细胞和视神经细胞生长与发育过程具有重要的作用。
60.3-酮酯酰-coa合酶(kcs)基因
61.在本发明中,本文所述“3-酮酯酰-coa合酶(kcs)基因”编码的蛋白产物为3-酮酯酰-coa合酶,参与催化神经酸的合成。
62.3-酮酯酰-coa合酶(kcs)基因编码的蛋白产物为3-酮酯酰-coa合酶,催化最终合成神经酸。3-酮酯酰-coa合酶(kcs)包含单一开阅框,编码512-653个氨基酸的蛋白质。
63.本发明经过大量的创造性实验发现在工程菌中整合表达3-酮酯酰-coa合酶(kcs)基因的表达盒可显著提高神经酸的产量、降低不饱和脂肪酸的含量。因此本方案提供的改造后的工程菌具有广阔的实际应用价值及前景。
64.本领域技术人员能使用通用的方法获得3-酮酯酰-coa合酶(kcs)基因的序列,例如从ncbi上获取。
65.本发明的3-酮酯酰-coa合酶(kcs)基因还包括不同来源的3-酮酯酰-coa合酶(kcs)基因、同一来源的不同的同源序列,以及密码子优化的3-酮酯酰-coa合酶(kcs)基因。
66.引物
67.如本文所用,术语“引物”指的是在与模板配对,在dna聚合酶的作用下能以其为起
点进行合成与模板互补的dna链的寡居核苷酸的总称。引物可以是天然的rna、dna,也可以是任何形式的天然核苷酸。引物甚至可以是非天然的核苷酸如lna或zna等。
68.引物“大致上”(或“基本上”)与模板上一条链上的一个特殊的序列互补。引物必须与模板上的一条链充分互补才能开始延伸,但引物的序列不必与模板的序列完全互补。比如,在一个3’端与模板互补的引物的5’端加上一段与模板不互补的序列,这样的引物仍大致上与模板互补。只要有足够长的引物能与模板充分的结合,非完全互补的引物也可以与模板形成引物-模板复合物,从而进行扩增。
69.工程菌构建
70.本发明提供了一种构建基因工程菌神经酸高产菌株的方法,包括(但不局限于)以下步骤:
71.1.构建含有基因表达盒的载体
72.经密码子优化后合成基因,设计3-酮酯酰-coa合酶(kcs)基因的pcr引物,克隆获得目的基因,通过gibson assembly无缝连接方法构建质粒。2.将获得的含有基因表达盒的载体转入受体菌株,获得受体基因组整合有基因表达盒的菌株。在一个优选例中,将构建的具有表达盒的载体通过属间接合转移,转入出发菌株中,在一定抗性筛选条件下挑选阳性接合子。
73.在一个优选例中,还包括步骤3:pcr验证步骤2得到的整合有基因表达盒的菌株的基因型;和/或步骤4:发酵检测得到的整合有基因表达盒菌株的神经酸的产量。在一个优选例中,将具有一定抗性的阳性接合子在液体培养基培养,提取菌体总dna,进行基因型的pcr验证。可以针对目的基因设计多对引物,以验证目标基因组中整合入相应基因。回收pcr扩增得到的正确大小碱基片段,采用dna测序的方法最终确证基因工程菌株正确。
74.本发明获得的菌株解脂耶氏酵母菌株po1g-g3
--
maolekcs,发酵生产神经酸的能力得到显著提高。
75.在一优选实施方式中,本发明采用技术方案为:
76.(1)通过生物信息学分析,对蒜头果3-酮酯酰-coa合酶进行筛选和分析。
77.(2)以解脂耶氏酵母为出发菌株,导入蒜头果3-酮酯酰-coa合酶基因(见seq id no.1-11),使解脂耶氏酵母具有或者增强合成神经酸的能力。
78.进一步说,以解脂耶氏酵母po1g-g3为出发菌株,采用同源重组方式导入maolekcs基因,获得菌株po1g-g3-maolekcs。
79.再进一步说,先构建同源重组质粒pylex-rdna-maolekcs-ura,再将所得质粒酶切后转化解脂耶氏酵母菌株po1g-g3-δura,通过尿嘧啶营养缺陷型平板筛选转化子,获得导入maolekcs基因的重组菌株po1g-g3-maolekcs。
80.再进一步说,所述同源重组质粒pylex-rdna-maolekcs-ura以质粒pylex-ura作为骨架模板,maolekcs表达盒的上下游分别为解脂耶氏酵母26s rdna的两个片段(见seq id no.12和seq id no.13)。
81.(3)以解脂耶氏酵母为出发菌株,过表达蒜头果3-酮酯酰-coa合酶基因(见seq id no.1-11),使解脂耶氏酵母具有或增强合成神经酸的能力。
82.进一步说,以解脂耶氏酵母po1g-g3-maolekcs为出发菌株,采用同源重组方式导入maolekcs基因,获得菌株po1g-g3-3maolekcs。
83.再进一步说,先构建同源重组质粒pylex-fad2-maolekcs-ura,再将所得质粒酶切后转化解脂耶氏酵母菌株po1g-g3-maolekcs-δura,通过尿嘧啶营养缺陷型平板筛选转化子,获得导入maolekcs基因的重组菌株po1g-g3-3maolekcs。
84.再进一步说,所述同源重组质粒pylex-fad2-maolekcs-ura以质粒pylex-ura作为骨架模板,maolekcs表达盒的上下游分别为解脂耶氏酵母fad2基因的两个片段(见seq id no.14和seq id no.15)。
85.(4)将上述重组菌株进行发酵罐培养,神经酸占总脂肪酸比例可达36.9%,神经酸产量达到25.7g/l。
86.发酵生产神经酸
87.本发明的基因改造菌株可以用来发酵生产神经酸,在一个优选例中,培养基可以包括(但不限于),以培养基的总体积计,重量体积比:
88.ypd液体培养基:葡萄糖20g/l、蛋白胨20g/l、酵母提取物10g/l,若需要固体培养基则添加1.5%的琼脂;用于常规培养。
89.尿嘧啶营养缺陷型培养基(ynb-δura):ynb(yeast nitrogen base without amino acids and ammonium)1.7g/l,葡萄糖20.0g/l,csm-ura(mp biomedicals)0.67g/l,硫酸铵5g/l,若需要固体培养基则添加1.5%的琼脂;用于重组菌株的筛选和活化培养。
90.摇瓶发酵培养基:葡萄糖150g/l,酵母提取物6g/l,硫酸铵12g/l。
91.大肠杆菌所用培养温度为37℃,液体培养时摇床转速为200rpm,所用培养基如下:
92.lb培养基:胰蛋白胨10g/l,酵母提取物5g/l,氯化钠10g/l,若需要固体培养基则添加1.5%的琼脂;用于常规培养。
93.amp抗性培养基:lb培养基中加入高浓度amp溶液,使amp终浓度为100μg/ml,若需要固体培养基则添加1.5%的琼脂;用于重组细菌菌株的筛选和扩大培养。
94.本发明菌株的发酵产物可用于制备神经酸。
95.药物组合物和施用方法
96.本发明菌株发酵产物中的神经酸可以用作制备药物。本发明化合物可施用于哺乳动物(如人),可以口服、直肠、肠胃外(静脉内、肌肉内或皮下)、局部等方式给药。所述化合物可以单独给药,或者与其他药学上可接受的化合物联合给药。需要指出,本发明的化合物可以混合给药。
97.用于口服给药的固体剂型包括胶囊剂、片剂、丸剂、散剂和颗粒剂。在这些固体剂型中,活性化合物与至少一种常规惰性赋形剂(或载体)混合,如柠檬酸钠或磷酸二钙,或与下述成分混合:(a)填料或增容剂,例如,淀粉、乳糖、蔗糖、葡萄糖、甘露醇和硅酸;(b)粘合剂,例如,羟甲基纤维素、藻酸盐、明胶、聚乙烯基吡咯烷酮、蔗糖和阿拉伯胶;(c)保湿剂,例如,甘油;(d)崩解剂,例如,琼脂、碳酸钙、马铃薯淀粉或木薯淀粉、藻酸、某些复合硅酸盐、和碳酸钠;(e)缓溶剂,例如石蜡;(f)吸收加速剂,例如,季胺化合物;(g)润湿剂,例如鲸蜡醇和单硬脂酸甘油酯;(h)吸附剂,例如,高岭土;和(i)润滑剂,例如,滑石、硬脂酸钙、硬脂酸镁、固体聚乙二醇、十二烷基硫酸钠,或其混合物。胶囊剂、片剂和丸剂中,剂型也可包含缓冲剂。
98.固体剂型如片剂、糖丸、胶囊剂、丸剂和颗粒剂可采用包衣和壳材制备,如肠衣和其它本领域公知的材料。它们可包含不透明剂,并且,这种组合物中活性化合物或化合物的
g3-δura,具体为:菌株po1g-g3是通过过表达解脂耶氏酵母内源基因dgat1、acc1和scd基因而获得(qiao and stephanopoulos.metabolic engineering,2015,29:56-65),再通过5-foa法反向筛选得到尿嘧啶营养缺陷型菌株po1g-g3-δura。
115.解脂耶氏酵母所用培养温度为28℃,液体培养时摇床转速为220rpm,所用培养基如下:
116.ypd液体培养基:葡萄糖20g/l、蛋白胨20g/l、酵母提取物10g/l,若需要固体培养基则添加1.5%的琼脂;用于常规培养。
117.尿嘧啶营养缺陷型培养基(ynb-δura):ynb(yeast nitrogen base without amino acids and ammonium)1.7g/l,葡萄糖20.0g/l,csm-ura(mp biomedicals)0.67g/l,硫酸铵5g/l,若需要固体培养基则添加1.5%的琼脂;用于重组菌株的筛选和活化培养。
118.摇瓶发酵培养基:葡萄糖150g/l,酵母提取物6g/l,硫酸铵12g/l。
119.大肠杆菌所用培养温度为37℃,液体培养时摇床转速为200rpm,所用培养基如下:
120.lb培养基:胰蛋白胨10g/l,酵母提取物5g/l,氯化钠10g/l,若需要固体培养基则添加1.5%的琼脂;用于常规培养。
121.amp抗性培养基:lb培养基中加入高浓度amp溶液,使amp终浓度为100μg/ml,若需要固体培养基则添加1.5%的琼脂;用于重组细菌菌株的筛选和扩大培养。
122.1.2蒜头果3-酮酯酰-coa合酶基因的筛选与表达
123.1.2.1蒜头果3-酮酯酰-coa合酶基因的生物信息分析
124.从文献(xu,et al.gigascience,2019,8:1
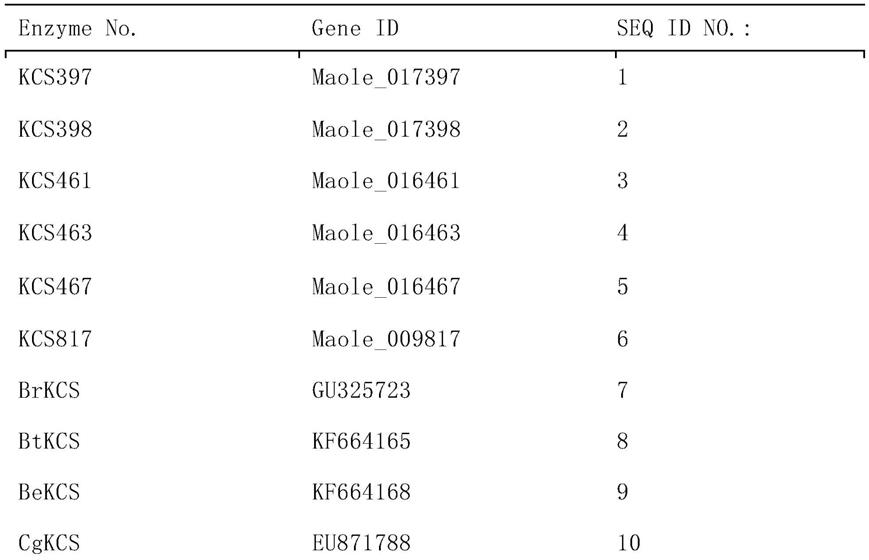
–
14)基因组数据中获取了12个潜在的蒜头果3-酮酯酰-coa合酶基因,分别为maole_002411、maole_003085、maole_004215、maole_005654、maole_008020、maole_009817、maole_016461、maole_016463、maole_016466、maole_016467、maole_017397和maole_017398。然后选取了其中6个基因,分别命名为kcs397(maole_017397)、kcs398(maole_017398)、kcs461(maole_016461)、kcs463(maole_016463)、kcs467(maole_016467)和kcs817(maole_009817),并且获取了相应的氨基酸序列,依次为maole_017397.t1、maole_017398.t1、maole_016461.t1、maole_016463.t1、maole_016467.t1和maole_009817.t1,具体见表1。从ncbi数据库中搜寻到已经被确认为3-酮酯酰-coa合酶的mokcs(genbank:qda34238.1,来源于m.oleifera)、cgkcs(genbank:acj61778.1,来源于c.graeca)、lakcs(genbank:eu871787.1,来源于lunaria annua)、brkcs(genbank:gu325723,来源于brassica rapa)、btkcs(genbank:kf664165,来源于brassica tournefortii)、bekcs(genbank:kf664168,来源于brassica elongata),以及其基因和氨基酸序列。用mega5.1中自带的clusterw对kcs397、kcs398、kcs461、kcs463、kcs467、kcs817、mokcs、cgkcs、lakcs、brkcs、btkcs和bekcs的氨基酸序列进行蛋白序列多重比对后,采用neighbor-joining算法(自检举1000次),绘制出系统树,见图1。选取4个kcs(kcs398、kcs461、kcs467和kcs817)基因,利用ncbi在线结构域分析软件(https://www.ncbi.nlm.nih.gov/structure/cdd/wrpsb.cgi)以及dnaman软件对cgkcs、mokcs和选出的4个kcs蛋白序列进行比对,见图2。
125.1.2.2载体构建
126.kcs398、kcs461、kcs467和kcs817基因由无锡青兰生物科技有限公司进行密码子优化后合成(优化后的基因序列依次为seq id no.2、3、5、6),得到带有maolekcs基因和氨
苄抗性基因的穿梭质粒,命名为pmv-maolekcs#-amp(注:#代表不同的maolekcs基因编号,即指398,461,467或817,后面亦然)。
127.质粒载体的构建采用gibson assembly方法,所有装配片段采用pcr扩增方法获得。质粒骨架源自pylex质粒(yeastern biotech co.,ltd)。所有pcr扩增采用kapa hifi高保真dna聚合酶,扩增体系皆为50μl(2
×
kapa mix,25ul;10μm引物各1.5ul;模板1μl;加水补至50μl),即得到各dna序列。扩增条件为:95℃预变性3分钟;98℃变性20秒、60-72℃退火15秒、72℃延伸,延伸时间按照30秒每kb计算,循环数为29-35;72℃延伸10分钟。
128.表1.蒜头果及从数据库筛选的3-酮酯酰-coa合酶
[0129][0130][0131]
质粒pylex-ura是采用gibson assembly方法,用ura3基因替代pylex质粒上的leu2基因而获得;然后再以pylex-ura为骨架构建同源重组质粒pylex-rdna-cgkcs-ura(cgkcs为1.2中的碎米芥属kcs基因),其中cgkcs表达盒上下游分别为解脂耶氏酵母26s rdna的部分序列(见seq id no.12和seq id no.13),以获得定点整合位点在26s rdna区域的同源重组质粒。以质粒pylex-rdna-cgkcs-ura作为骨架,采用gibson assembly方法,用maolekcs基因替代cgkcs基因,构建同源重组质粒pylex-rdna-maolekcs-ura。
[0132]
具体构建过程如下:
[0133]
质粒pylex-rdna-cgkcs-ura从专利申请人实验室中直接获得。首先用p#-f和p#-r引物以pylex-rdna-cgkcs-ura质粒为模板,扩增得到带amp和ura筛选标记的骨架dna片段;然后用pmv#-f和pmv#-r引物,以带有maolekcs基因的质粒pmv-maolekcs#-amp为模板,分别扩增maolekcs基因片段;将骨架和maolekcs基因片段gibson装配后转化大肠杆菌感受态trans-t1(北京全式金生物技术有限公司),再用引物rdna-sf和rdna-sr进行菌落pcr和测序验证后,获得同源重组质粒pylex-rdna-maolekcs#-ura(所述引物见表2)。
[0134]
表2.构建质粒和菌落pcr所用引物
[0135][0136][0137]
1.2.3酵母转化
[0138]
利用not i限制性内切酶对重组质粒pylex-rdna-maolekcs-ura进行酶切,通过琼脂糖凝胶电泳进行确认,转化用片段胶回收后利用liac转化法转化解脂耶氏酵母po1g-g3-δura,转化体系如表3所示,方法步骤描述如下:
[0139]
表3.解脂耶氏酵母转化体系
[0140][0141]
(1)按照表3的转化体系添加各组分,混匀;
[0142]
(2)30℃水浴1h后涡旋震荡,39℃热激10min;
[0143]
(3)直接取转化体系的混合液涂布在ynb-δura培养基的筛选平板上,于28℃培养
2-4天,获得导入maolekcs基因的重组菌株,命名为po1g-g3-maolekcs#。
[0144]
1.2.4重组菌株的诊断pcr验证
[0145]
对上述获得的各重组菌株的不同转化子进行诊断pcr验证。
[0146]
从筛选平板上挑菌重组菌株单菌落于ypd平板上,生长约24小时后,挑取菌体,应用真菌基因组快提试剂盒(生工生物工程股份有限公司)提取重组菌株的基因组dna,以基因组dna为模板,导入基因maolekcs的上游部分由引物ylr-duf和ylr-dur进行pcr验证,下游部分由引物ylr-ddf和ylr-ddr进行验证(所述引物见表1)。
[0147]
pcr扩增采用easytaq dna聚合酶,扩增体系皆为20μl(2
×
easytaq mix,10μl;10μm引物各0.6μl;模板1μl;加水补至20μl),扩增片段通过琼脂糖凝胶电泳进行确定。扩增条件为:94℃预变性3分钟;94℃变性30秒、55℃退火30秒、72℃延伸,延伸时间按照60秒每kb计算,循环数为36;72℃延伸10分钟。
[0148]
1.2.5重组菌株的摇瓶发酵培养
[0149]
将上下游同时验证成功(见图3)的重组菌株接种于20ml ypd培养基(250ml锥形瓶中),生长约24小时后转接到ypd培养基平板上,生长约24小时后接种到5ml ynb-δura培养基(50ml锥形瓶)中,生长36小时后吸取100μl转接到新鲜的5ml ynb-δura培养基中培养36小时,再转接到30ml摇瓶发酵培养基,使初始od统一为0.08,每组三个平行,培养168小时。菌株干重由称重法获得。
[0150]
1.2.6酸热法提取总油脂
[0151]
(1)50ml离心管收集解脂耶氏酵母发酵培养液,6000rpm高速离心5min收集酵母菌体;
[0152]
(2)1g湿菌体大约加入10ml 4m的盐酸,振荡均匀,放于28℃摇床震荡约1-2h;
[0153]
(3)沸水浴6-8min,立即放于-20℃冷却30min;
[0154]
(4)加入氯仿:甲醇=1:1(v/v)20ml充分混合,
[0155]
(5)分离下层氯仿并称量体积,加入等体积的0.15%的氯化钠,4000rpm离心10min;
[0156]
(6)取下层转移到新的已称重的玻璃管中,氮吹仪吹干溶剂相,再次称重计算油脂产量,总脂含量为总脂产量占干重的比例。
[0157]
1.2.7脂肪酸甲脂化
[0158]
(1)在玻璃管中添加2.6ml的甲醇:硫酸=98:2(v/v)溶液,混匀后放于85℃反应3h,之后在冰箱内冷却;
[0159]
(2)添加1ml饱和nacl溶液和1ml正己烷,振荡后5000rpm高速离心5min,吸取上清到ep管中,并用有机相滤膜进行过滤后进行气相色谱实验。
[0160]
1.2.8气相色谱分析脂肪酸成分
[0161]
脂肪酸成分的分析采用气相色谱法(agilent 7890b-gc)。色谱检测条件:色谱柱为hp-inowax(30m
×
0.32mm
×
0.5μm);进样温度:250℃;检测器温度:250℃;进样体积:1μl;起始柱温为140℃,保持1min,以10℃/min升温至180℃,保持2min,5℃/min升温至210℃,保持4min,然后5℃/min升温至250℃,保持4min。以面积归一法得到各脂肪酸组分的相对含量。
[0162]
1.3解脂耶氏酵母中过表达maolekcs
[0163]
1.3.1过表达基因重组质粒构建
[0164]
质粒载体的构建采用gibson assembly方法,所有装配片段采用pcr扩增方法获得,具体方法与1.2.2中相同。以pylex-ura为骨架构建的同源重组质粒pylex-fad2-cgkcs-ura从专利申请人实验室中直接获得,其中cgkcs表达盒的上下游分别为解脂耶氏酵母fad2基因的部分序列(见seq id no.14和seq id no.15),以获得定点整合位点在fad2基因座的同源重组质粒。以质粒pylex-fad2-cgkcs-ura为骨架,采用gibson assembly方法,用连续的2个maolekcs表达盒替代cgkcs表达盒,构建同源重组质粒pylex-fad2-maolekcs-ura。
[0165]
1.3.2酵母的转化、重组菌株的诊断pcr验证
[0166]
通过5-foa法反向筛选到po1g-g3-maolekcs的尿嘧啶营养缺陷型菌株po1g-g3-maolekcs-δura。利用not i限制性内切酶对重组质粒pylex-fad2-maolekcs-ura进行酶切,通过琼脂糖凝胶电泳进行确认,转化用片段胶回收后利用liac转化法转化解脂耶氏酵母po1g-g3-maolekcs-δura,获得导入maolekcs基因的重组菌株,命名为po1g-g3-3maolekcs。挑取不同的转化子进行诊断pcr验证,方法同1.2.4,上游部分由引物fad-df(5
’-
ctacggcacgataaagatgg-3’)和ylr-dur进行pcr验证,下游部分由引物ylr-ddf和fad-dr(5
’-
ttacaatgggaccgtgcttg-3’)进行验证,上下游同时验证同成功的不同转化子进行摇瓶发酵培养,培养方法以及培养后的各种指标分析方法与1.2中相同。
[0167]
2.结果与分析
[0168]
2.1蒜头果3-酮酯酰-coa合酶基因的筛选与表达
[0169]
2.1.1蒜头果3-酮酯酰-coa合酶基因的生物信息分析
[0170]
从蒜头果基因组中检索到12个3-酮酯酰-coa合酶,但是这些基因的功能和应用没有报道,其中6个3-酮酯酰-coa合酶被文献作者注释为同时具有脂肪酸延长过程中的kcs、kcr、hcd和ecr作用。将6个kcs的氨基酸序列与ncbi数据库中的mokcs(genbank:qda34238.1)和cgkcs(genbank:acj61778.1)进行蛋白序列多重比对后,采用neighbor-joining算法绘制出系统树(见图1),发现maolekcs463与mokcs氨基酸序列完全一致,且maolekcs397与maolekcs398相似度很高。由图2可以看出,kcs398、kcs461、kcs467和kcs817与已报道的cgkcs和mokcs均含有kcs家族的3个功能保守结构域“fgntssss”“gsgfkcnsavw”和“gmgcsa”(红框部分),表明这些蛋白属于kcs家族蛋白,选取kcs398、kcs461、kcs467和kcs817做后续研究。将这4个kcs的基因由无锡青兰生物科技有限公司进行密码子优化后合成,得到带有maolekcs基因和氨苄抗性基因的穿梭质粒pmv-maolekcs#-amp(#代表398、463、467、817数字编号)。
[0171]
2.1.2重组酵母菌株的诊断pcr验证
[0172]
从筛选平板上挑取重组菌株的不同转化子于ypd平板上,其中菌株po1g-g3-maolekcs398挑取7个,菌株po1g-g3-maolekcs461、po1g-g3-maolekcs467和po1g-g3-maolekcs817各挑取8个,同时将对照菌株po1g-g3接种于ypd平板,生长约24小时后提取重组菌株的不同转化子和对照菌株的基因组dna。以基因组dna为模板,对导入基因maolekcs的上下游部分进行pcr扩增,验证基因的整合表达。将po1g-g3的基因组dna作为对照,pcr产物的琼脂糖凝胶电泳图如图3所示。首先对照组po1g-g3中没有上游或下游pcr产物(见图3a和图3c的最左边泳道ck);然后下游部分pcr结果呈阳性(目的条带为1632bp)的转化子有:
[0173]
po1g-g3-maolekcs398的1、3、4、5和6号,
[0174]
po1g-g3-maolekcs461的1、2、3和4号,
[0175]
po1g-g3-maolekcs467的2、3、4、6和7号,
[0176]
po1g-g3-maolekcs817的2、4、6和8号(图3b);
[0177]
接着再对下游部分呈阳性的重组菌株的转化子进行上游部分的整合验证,上游部分呈阳性(目的条带为1555bp)的有:
[0178]
po1g-g3-maolekcs398的1、3、4、5和6号,
[0179]
po1g-g3-maolekcs461的1、2和4号,
[0180]
po1g-g3-maolekcs467的2、3、4、6和7号,
[0181]
po1g-g3-maolekcs817的2、4、6和8号;
[0182]
结合上下游部分pcr产物的亮度,最后选取po1g-g3-maolekcs
[0183]
398的1号,po1g-g3-maolekcs461的2号,po1g-g3-maolekcs467的3号以及po1g-g3-maolekcs817的8号做后续的摇瓶发酵培养。
[0184]
2.1.3重组菌株的摇瓶发酵结果
[0185]
如图4所示,与对照菌株po1g-g3相比,菌株po1g-g3-maolekcs817的干重、总脂产量和总脂含量都有大幅下降;菌株po1g-g3-maolekcs398和po1g-g3-maolekcs467的干重、总脂产量和总脂含量略有提高;菌株po1g-g3-maolekcs461的干重、总脂产量和总脂含量无显著变化。
[0186]
脂肪酸组成分析表明,与对照菌株po1g-g3相比,所有菌株明显生成了神经酸和木蜡酸,并且产量相近,占总脂肪酸比例分别达到8.9%和6.2%(图5)。同时菌株的c18:1和c18:0比例与对照相比明显降低,还有少量c20和c22脂肪酸被检测到(见图5),这说明maolekcs能够将c18:1和c18:0的碳链逐步延长,生成神经酸和木蜡酸。因此,本发明通过脂肪酸延长酶的筛选和酵母体内表达分析,发现了蒜头果来源的脂肪酸延长酶maolekcs能够催化神经酸的合成,对应基因可用于构建高产神经酸酵母。
[0187]
2.2解脂耶氏酵母中过表达maolekcs
[0188]
如图6a所示,与对照菌株的29.2g/l相比,重组菌株不同转化子的干重都有不同程度的降低,依次为26.7g/l、27.4g/l、25.8g/l和28.7g/l。总脂产量方面,对照菌株为14.16g/l,1、2和3号转化子下降明显,其中3号最明显,下降了32%;4号转化子的总脂产量为14.18g/l,没有显著变化。总脂含量方面与总脂产量情况相似,1、2和3号转化子下降明显,其中3号最明显,4号转化子的油脂含量没有显著变化。
[0189]
重组菌株的各转化子的不同脂肪酸比例如图6b和图7所示。对照菌株的神经酸占总脂肪酸酸比例为8.96%(图7a),1、2和4号转化子的神经酸占比都有大幅提高,分别为13.27%、13.26%和12.60%(图7b,c,e);但是3号转化子的比例略有降低,为8.61%(图7d)。同时,4个转化子的木蜡酸占总脂肪酸的比例都大幅提高,其中3号转化子最突出,与对照相比提高了将近2倍。相对应的是,对照菌株的c18:0和c18:1的比例都有不同程度降低,说明过表达maolekcs进一步催化了c18:0和c18:1的碳链延长,其中4号转化子的神经酸合成效果最为显著。4号转化子的总脂产量与对照相比没有显著变化,神经酸占比提高40%,增强了酵母合成神经酸的能力。
[0190]
实施例2:重组菌株的发酵罐发酵
[0191]
1.材料与方法
[0192]
1.1发酵培养基
[0193]
种子培养基(ynb培养基):ynb 3g/l,葡萄糖60g/l,硫酸铵35g/l,ynb与硫酸铵灭菌后过滤除菌加入。
[0194]
发酵培养基(fm培养基):葡萄糖50g/l,硫酸铵35g/l,酵母粉38g/l,磷酸二氢钾8g/l,十二水合磷酸氢二钠6g/l,七水硫酸镁5g/l,消泡剂1ml/l。
[0195]
补料控制:根据发酵罐中的残糖浓度进行控制,使用灭菌后的600g/l的葡萄糖补料。
[0196]
1.2 50l发酵罐的发酵工艺
[0197]
保藏菌种从-80℃取出后,放于冰上解冻。吸取100μl接种到250ml的ynb培养基中,220rpm、28℃下摇床培养24h后,以接种后od约0.01为标准转接到5l种子发酵罐中,种子罐的培养基为fm发酵培养基,装液量为2.5l,种子罐的培养条件为温度28℃,使用制冷机循环制冷;ph使用5mol/l的氢氧化钠控制为5.5,设置do为20%,培养24h后移种至50l发酵罐(镇江汇能达公司,hnd-bj-5l-50l型)中开始发酵。
[0198]
根据实验方案配制fm发酵培养基,充分混匀后加入50l发酵罐中。发酵罐的装液量为25l。接种量为10%,温度降低至28℃后,从接种口接入150ml种子液。
[0199]
发酵条件:发酵温度为28℃,使用制冷机循环制冷;ph使用5mol/l的氢氧化钠控制为5.5;初始通气量为1vvm,前24h内逐渐提高至最大通气量2vvm。溶氧控制方式采用和转速级联控制,设置do为20%,48h之后调节为10%,转速范围为200rpm-600rpm。补料葡萄糖浓度为600g/l,根据罐中残糖浓度变化来补加,待其浓度降低至30g/l以下补料。
[0200]
发酵过程中的指标分析方法参考实施例1。粗神经酸产量(g/l)=总脂产量(g/l)*神经酸占总脂肪酸的比例(%)。
[0201]
2.结果与分析
[0202]
在摇瓶发酵基础上,使用50l发酵罐进行了重组菌株po1g-g3-3maole398的发酵培养,结果见图8。通过对重组菌的油脂、干重和脂肪酸的组成进行检测,得到以下结论:经过168h的发酵,干重(dcw)达到了149.4g/l,总脂产量(lipid titer)为69.6g/l,总脂含量(lipid content)达到46.6%,神经酸占总脂肪酸的比例为36.9%,脂肪酸成分气相分析图谱结果见图9,神经酸产量达到了25.7g/l。
[0203]
在本发明提及的所有文献都在本技术中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本技术所附权利要求书所限定的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1