一种新型mariner转座子系统、应用和构建枯草芽孢杆菌插入突变株文库的制作方法
1.本发明属于微生物技术领域,尤其是一种新型mariner转座子系统、应用和构建枯草芽孢杆菌插入突变株文库。
背景技术:
2.转座因子或转座子是一类在很多动物中(包括线虫、昆虫和人等)存在的可自由移动的跳跃因子,可以自主复制并跳跃插入到某功能基因中,引起该基因失活,产生突变体。转座子参与很多遗传现象,例如基因重排、质粒与染色体整合等。现在转座子突变技术已经成为微生物分子遗传学研究的重要工具之一,特别是具有随机转座特性的转座子已经成为发现新基因、克隆功能基因、发掘已知基因的新功能以及研究蛋白功能的有效工具。
3.枯草芽孢杆菌是益生菌和微生物发酵的主要工业菌株,构建转座子突变株文库对于枯草芽孢杆菌基因组的功能注释具有重要意义。目前国际上通用的枯草芽孢杆菌的转座子为tn917和tn551,tn55来自金黄色葡萄球菌,tn917来自粪链球菌,这两个转座子均含有红霉素抗性基因,具有同源性,它们的末端反向重复序列与革兰氏阴性的tn
‑
3家族的末端反向重复序列之间有很高的同源性,但是有研究表明,以这两种转座子构建的转座子突变株文库随机性较差,具有转座子插入“热点”,不能得到完全覆盖全基因组的转座子突变株文库;含有mariner转座酶的用于革兰氏阳性菌的转座子系统pbtn,以温度敏感性质粒pbt2作为载体,整合了两端带有重复序列的红霉素耐药基因,在红霉素的耐药基因和来源于mariner真核转座子的转座酶间插入来源于ptx15的启动子,以转座子pbtn构建的突变文库无明显插入热点,具有高度随机性和单一性,转座子pbtn的插入位置确定较难,转座效率不确定。
4.tc1/mariner转座子是自然界中分布最广泛的一类dna转座子超家族,家族成员的全长一般为1300~2400bp,其中包含一个单基因编码的转座酶,侧翼为反向末端重复序列(inverted terminal repeats,itrs)。当转座酶与itrs结合时,可以促进转座的发生。在自然界已经发现14个有活性的tc1/mariner类型转座子,如minos和mos1等。另外通过分子重构也获得高活性的人工转座子,如sleeping beauty,sb。sb和mos1等转座子作为基因转移载体已被广泛应用于转基因、基因捕获和基因治疗等领域的研究中,并取得了很好的应用效果。
5.通过对比,本发明专利申请与上述专利公开文献存在本质的不同。
技术实现要素:
6.本发明的目的在于克服现有技术的不足之处,提供一种新型mariner转座子系统、应用和构建枯草芽孢杆菌插入突变株文库。
7.本发明解决其技术问题所采用的技术方案是:
8.一种新型mariner转座子系统,用于构建枯草芽孢杆菌插入突变株文库,所述系统
包含如下元件:
9.⑴
已测序的mariner转座子,该转座子genbank序列接收号为kj933396;
10.⑵
额外添加在步骤
⑴
中已测序的mariner转座子的itr区域中的壮观霉素抗性基因spc;
11.⑶
能在大肠杆菌细胞内复制的宿主复制起始区;
12.⑷
能在枯草芽孢杆菌细胞内复制的温度敏感性质粒复制起始区;
13.⑸
温度敏感性质粒的载体耐药性基因ermc。
14.而且,所述温度敏感性质粒复制起始区扩增于pe194质粒的温敏复制起始区pe194ori
ts
,能够使质粒在枯草芽孢杆菌细胞内复制,当培养温度超过42℃时,质粒丢失。
15.而且,所述能在大肠杆菌细胞内复制的宿主复制起始区扩增于pbr322质粒的复制起始区cole1 origin,能够使所述新型mariner转座子系统在大肠杆菌细胞内复制。
16.而且,所述已测序的mariner转座子扩增于pkkma质粒,包括转座酶基因tnpa,并在受转座酶识别的两个反向重复序列itrs区域中额外添加了壮观霉素抗性基因spc。
17.而且,所述温度敏感性质粒的载体耐药性基因ermc来源于pmad质粒。
18.而且,所述新型mariner转座子系统的元件的插入具有随机性,将新型mariner转座子系统命名为pzq1902。
19.而且,所述已测序的mariner转座子的核苷酸序列为seq id no.1。
20.而且,所述新型mariner转座子系统的核苷酸序列为seq id no.2。
21.如上所述的新型mariner转座子系统在枯草芽孢杆菌插入突变株文库的构建方面中的应用。
22.利用如上所述的新型mariner转座子系统构建获得的枯草芽孢杆菌插入突变株文库,其特征在于:构建步骤如下:
23.将新型mariner转座子系统在大肠杆菌dh5α中构建成功后,转化到枯草芽孢杆菌中,培养含有新型mariner转座子系统的枯草芽孢杆菌,获得转化子30℃扩增培养后,得到枯草芽孢杆菌插入突变株文库。
24.本发明取得的优点和积极效果为:
25.1、本新型mariner转座子系统插入位点随机性强,序列清晰,插入位置可快速鉴定,且受受体菌内源性转座子的影响较小,与现有的转座子相比,实用性更强,具有插入位点随机性强,能够利用反向pcr方法鉴定转座子在基因组的插入位置的优点。通过高温培养(42℃),可以快速消除细胞内质粒携带的转座子系统。
26.2、本新型mariner转座子系统使用到的tc1/mariner转座子质粒pkkma,该质粒携带一个tc1/mariner类型转座子,该转座子经测序,genbank序列接收号为kj933396,该转座子经测序,序列清晰,使用便利,在铜绿假单胞菌、鲍曼不动杆菌、弗氏柠檬酸菌、粘质沙雷氏菌等革兰氏阴性菌中转座效率高,随机性好,能够利用反向pcr方法鉴定转座子在基因组的插入位置。为探究mariner转座酶在革兰氏阳性菌中能否使用,使用效率是否高效,本发明基于革兰氏阴性菌转座子pkkma改造提供了一种用于构建枯草芽孢杆菌插入突变株文库。。
27.3、本新型mariner转座子系统能够用于构建枯草芽孢杆菌插入突变株文库,可以系统高效的得到覆盖全基因组的革兰氏阳性菌枯草芽孢杆菌转座子文库。
附图说明
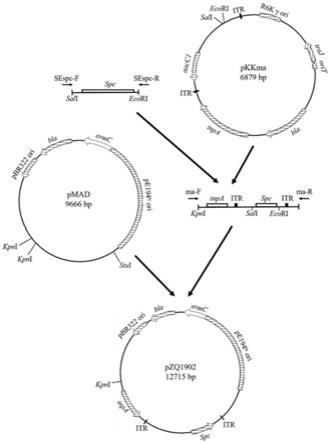
28.图1为本发明中新型mariner转座子系统的一种构建过程图。
具体实施方式
29.下面结合实施例,对本发明进一步说明,下属实施例是叙述性的,不是限定性的,不能以下述实施例来限定本发明的保护范围。
30.本发明中所使用的原料,如无特殊说明,均为常规市售产品,本发明中所使用的方法,如无特殊说明,均为本领域常规方法,本发明所用各物质质量均为常规使用质量。
31.一种新型mariner转座子系统,所述系统包含如下元件:
32.⑴
已测序的mariner转座子,该转座子genbank序列接收号为kj933396,在革兰氏阴性菌中转座效率较高;
33.⑵
额外添加在步骤
⑴
中已测序的mariner转座子的itr区域中的壮观霉素抗性基因spc;
34.⑶
能在大肠杆菌细胞内复制的宿主复制起始区;
35.⑷
能在枯草芽孢杆菌细胞内复制的温度敏感性质粒复制起始区;
36.⑸
温度敏感性质粒的载体耐药性基因ermc。
37.较优地,所述温度敏感性质粒复制起始区扩增于pe194质粒的温敏复制起始区pe194 ori
ts
,能够使质粒在枯草芽孢杆菌细胞内复制,当培养温度超过42℃时,质粒丢失。
38.较优地,所述能在大肠杆菌细胞内复制的宿主复制起始区扩增于pbr322质粒的复制起始区cole1 origin,能够使所述新型mariner转座子系统在大肠杆菌细胞内复制。
39.较优地,所述已测序的mariner转座子扩增于pkkma质粒,包括转座酶基因tnpa,并在受转座酶识别的两个反向重复序列itrs区域中额外添加了壮观霉素抗性基因spc。
40.较优地,所述温度敏感性质粒的载体耐药性基因ermc来源于pmad质粒,有助于获得携带mariner转座系统的枯草芽孢杆菌细胞。
41.较优地,所述新型mariner转座子系统的元件的插入具有随机性,将新型mariner转座子系统命名为pzq1902。
42.较优地,所述已测序的mariner转座子的核苷酸序列为seq idno.1。
43.较优地,所述新型mariner转座子系统的核苷酸序列为seq id no.2。
44.如上所述的新型mariner转座子系统在枯草芽孢杆菌插入突变株文库的构建方面中的应用。
45.利用如上所述的新型mariner转座子系统构建获得的枯草芽孢杆菌插入突变株文库,构建步骤如下:
46.将新型mariner转座子系统在大肠杆菌dh5α中构建成功后,转化到枯草芽孢杆菌中,培养含有新型mariner转座子系统的枯草芽孢杆菌,获得转化子30℃扩增培养后,得到枯草芽孢杆菌插入突变株文库。
47.本发明中新型mariner转座子系统的构建过程可以如图1所示。
48.本发明可以基于转座子pkkma构建了一种新型mariner转座子系统,用于构建枯草芽孢杆菌插入突变株文库。该系统由四部分组成:
⑴
pe194质粒的温敏复制起始区pe194
ts ori,可以在枯草芽孢杆菌细胞内复制,当培养温度超过42℃时,质粒的复制受到抑制;
⑵
pbr322质粒的复制起始区cole1 origin,可以在大肠杆菌体内复制;
⑶
mariner转座子,包括转座酶基因tnpa和壮观霉素抗性基因spc,以及受转座酶识别的两个反向重复序列itrs;
⑷
红霉素抗性基因ermc。该mariner转座子序列已知,与现有枯草芽孢杆菌转座子系统相比,具有插入位点随机性强,能够利用反向pcr方法鉴定转座子在基因组的插入位置。通过高温培养(42℃),可以快速消除细胞内质粒携带的转座子系统。
49.下面对本发明的新型mariner转座子系统构建过程详细描述。
50.一、新型mariner转座子系统(pzq1902)的构建
51.第一步、设计引物扩增质粒pic333的壮观霉素抗性基因
52.以质粒pic333为模板,利用上游引物:5
‘‑
cgcgtcgaccacctagatccttttgactc
‑3’
和下游引物5
‘‑
cggaattcaaagtaagcacctgttattgc
‑3’
扩增壮观霉素抗性基因spc的序列,扩增产物大小为1033bp,利用切胶的方式纯化回收目的片段,与pkkma质粒载体进行连接,在引物两端设计限制性内切酶位点sali和ecori,在回收目的片段后用限制性内切酶sali和ecori双酶切扩增纯化产物,再利用切胶的方式纯化回收目的片段。
53.第二步、线性化mariner转座子载体质粒pkkma,并利用酶切重组的方式将壮观霉素抗性基因连接到转座子载体质粒pkkma上
54.用限制性内切酶sali和ecori双酶切mariner转座子载体质粒pkkma,再利用切胶的方式纯化回收目的片段。用t4dna连接酶将第一步纯化壮观霉素抗性基因片段和线性化载体pkkma片段在16℃进行过夜连接。
55.第三步、设计引物扩增重组pkkma质粒的mariner转座子跳跃基因序列,包括转座酶基因tnpa、壮观霉素抗性基因spc和转座子两端的反向重复序列itr。
56.以第二步得到的重组pkkma质粒为模板,使用上游引物:5
‘‑
ggggtaccgcattggtaactgtcagaccaag
‑3’
和下游引物5
‘‑
acccgacccgaaacaccat
‑3’
引物扩增mariner转座子跳跃基因序列(包括转座酶基因tnpa、壮观霉素抗性基因spc和转座子两端的反向重复序列itr),扩增产物大小为5342bp,利用切胶的方式纯化回收目的片段。为与质粒载体pmad进行连接,在引物一端设计限制性内切酶位点kpni,将回收纯化片段用限制性内切酶kpni进行酶切,再利用切胶的方式纯化回收目的片段。
57.第四步、线性化pmad质粒,并利用酶切重组的方式将重组的mariner转座子跳跃基因片段连接到pmad质粒载体上
58.使用限制性内切酶kpni和stui双酶切pmad质粒,其中stui酶切后产生的末端为平末端,酶切后回收大小为7376bp的目的条带。纯化回收的酶切载体片段和重组的mariner转座子跳跃基因片段用t4 dna连接酶在16℃进行过夜连接。
59.第五步、将连接产物化转到大肠杆菌dh5α中,筛选鉴定阳性转化子,从转化子中提取质粒进行酶切验证,如果质粒pzq1902构建成功,则能被限制性内切酶ecori和kpni双酶切出两条大小分别为8646bp和4069bp的条带。
60.二、构建枯草芽孢杆菌168的转座子插入突变株文库
61.1、枯草芽孢杆菌168感受态的制备和转化
62.(1)挑取枯草芽孢杆菌168单菌落接种于5ml液体lb培养基中,37℃振荡培养12h。
63.(2)取300μl菌液转接至10mlspi培养基中,37℃,220rpm培养至对数末期(od
600
约为1.6)。
64.(3)快速取1mlspi菌液转接至9mlspii培养基中,37℃,100rpm培养1.5h。
65.(4)加入100μl0.1megta溶液混匀,继续培养10min后,分装为500μl每管。
66.2、将新构建的温度敏感性转化子质粒pzq1902转入到枯草芽孢杆菌168中
67.(1)向500μl枯草芽孢杆菌168感受态细胞中加入适量质粒,轻轻混匀后,30℃,100rpm培养40min。
68.(2)将培养液转至250rpm,30℃继续培养2h。
69.(3)4000rpm离心10min收集菌体,弃部分上清,留100μl重悬菌体,涂布相应的抗性平板,30℃过夜培养。
70.(4)长出的转化子用质粒提取试剂盒提取质粒,通过酶切验证质粒是否成功转入枯草芽孢杆菌中。
71.3、转座子突变株插入文库的建立
72.(1)挑取168/pzq1902单菌落接种到5ml液体lb培养基中,培养基含有红霉素10μg/ml、壮观霉素100μg/ml,30℃,220rpm培养12h。
73.(2)菌液按照3%的转接量转接到100ml液体lb(无抗生素)培养基中,30℃振荡培养3h至od
600
约为0.8。
74.(3)培养液12000rpm离心10min,弃上清,用2ml液体lb将沉淀重悬,梯度稀释后涂布含有壮观霉素100μg/ml的lb平板,50℃过夜培养。
75.(4)将平板上长出来的菌落经过划线纯化后分别点到含有红霉素10μg/ml的lb平板上和含有壮观霉素100μg/ml的lb平板上,37℃过夜培养。
76.(5)挑选在含有红霉素的平板上不长,含有壮观霉素的平板上生长的菌落,可以初步确定为枯草芽孢杆菌168转座子插入突变株。
77.4、转座子突变株插入位点的鉴定
78.(1)通过苯酚
‑
氯仿抽提法提取枯草芽孢杆菌168突变株的基因组,用限制性内切酶taqi酶切突变株的基因组,65℃酶切5h。200μl酶切体系:10xbuffer 20μl、基因组dna 50μl、taqi 5μl、去离子水125μl。
79.(2)用苯酚
‑
氯仿抽提法纯化回收酶切产物,将酶切产物用t4 dna连接酶16℃连接16h,使酶切片段发生自身环化。20μl连接体系:10xbuffer 2μl、酶切产物5μl、t4 dna连接酶1μl、去离子水12μl。
80.(3)以连接产物为模板,根据mariner转座子序列的末端设计一对引物taq3
‑
f:tttgcgccgacatcataacg和taq3
‑
r:ccataccacagcttccgatgg通过反向pcr扩增连接产物,将pcr产物送去测序,鉴定突变株基因组中mariner转座子的插入位点。pcr产物应该包括mariner转座子的部分末端序列、168基因组的部分序列和taqi酶切位点。
81.seq1
82.已测序的mariner转座子的核苷酸序列
83.[0084][0085]
seq2
[0086]
新型mariner转座子系统的核苷酸序列
[0087]
[0088]
[0089]
[0090]
[0091]
[0092]
[0093][0094]
尽管为说明目的公开了本发明的实施例,但是本领域的技术人员可以理解:在不脱离本发明及所附权利要求的精神和范围内,各种替换、变化和修改都是可能的, 因此,本发明的范围不局限于实施例所公开的内容。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1