调控胚乳发育及品质的基因FLO18
调控胚乳发育及品质的基因flo18
技术领域
1.本发明属于生物技术和植物学领域,更具体地,本发明涉及调控胚乳发育及品质的基因flo18。
背景技术:
2.被子植物的胚乳由一个精细胞和两个位于中央细胞的极核双受精后发育形成,是三倍体组织。植物胚乳的发育过程可分为核型胚乳、细胞型胚乳和沼生目型胚乳三种类型。
3.禾本科植物水稻作为世界上最主要的粮食作物之一,拥有巨大的市场和需求。人们所消费的大米是水稻的胚乳部分,水稻胚乳占籽粒的70~80%。同时水稻胚乳在水稻胚的形成和种子萌发过程中起重要作用。探明胚乳遗传发育机制,揭示水稻胚乳发育与水稻产量品质的关系,对于提高水稻产量和品质具有重要的意义。
4.水稻的胚乳发育过程属于核型胚乳类型。即在胚乳发育早期,只进行细胞核分裂不进行细胞质分裂,不形成细胞壁。当发育到一定程度时才进行细胞质分裂和细胞壁形成。
5.水稻的胚乳的发育过程涉及有丝分裂,细胞化,细胞分化,贮藏物质积累,和程序化死亡过程,一般在受精后21~22天内完成整个发育。0~2dap(授粉后天数,days after pollination)时期主要是核分裂但是胞质不分裂,3~5dap主要进行细胞化,6~21dap主要进行胚乳细胞分化和贮藏物质的积累。8dap开始细胞程序性死亡逐渐从胚乳中心蔓延到整个胚乳,到了21dap的时候,只剩下糊粉层细胞具有活性。22~30dap,水稻颖果的鲜重和干重不再增加。淀粉胚乳中随着单个淀粉粒边缘模糊形成了复合淀粉粒。在整个籽粒水平上,淀粉和贮藏的结晶导致了淀粉胚乳在成熟过程结束时形成半透明的淀粉胚乳。
6.水稻胚乳完全成熟后,经过脱水所剩的干物质中,淀粉占80~90%,蛋白质占7~9%,脂质占0.3~0.5%,还含有钾、镁、钙、锌等无机物。
7.淀粉有两种类型——直链淀粉和支链淀粉。直链淀粉以α(1-4)糖苷键相连,每个分子中有200个左右葡萄糖;支链淀粉除α(1-4)糖苷键外还有以α(1-6)糖苷键链接的分支结构;经常以碘液染色区分支链和直链淀粉,直链淀粉遇碘呈蓝色,支链淀粉则为红色。一般品种的水稻胚乳中的淀粉由20~30%的直链淀粉和70~80%的支链淀粉组成。
8.水稻中,淀粉主要是由四类酶协同作用合成:腺苷二磷酸葡萄糖焦磷酸化酶(adp-glucose pyrophosphorylase,agpase),淀粉合成酶(starch synthases,ss),颗粒结合淀粉合成酶(granule bound ss,gbss),淀粉分支酶(starch branching enzymes,sbe),淀粉去分支酶(starch debranching enzymes,dbe)。
9.胚乳中,细胞质基质中的agpase催化glucose-1-phosphate和atp形成adp-glucose,释放ppi;adp-glucose是淀粉合成的原料,这是淀粉合成的第一个关键步骤反应。淀粉合成酶ss催化adp-glucose的葡萄糖基转移到之前存在的以α(1-4)糖苷键连接的葡聚糖引物的非还原端,形成直链淀粉和支链淀粉。淀粉分支酶sbe主要作用是裂解α-(1-4)糖苷键,将释放的还原端转移到c6羟基端生成α-(1-6)糖苷键,形成支链淀粉分子的分支结构。淀粉去分支酶dbe主要用于水解支链淀粉中不正常分支的α-(1-6)糖苷键,用于调整支
链淀粉的分支,维持支链淀粉的结晶度。除此之外,歧化酶(disproportionating enzyme,dpe)和淀粉磷酸化酶(phosphorylase,pho)在淀粉多聚体的起始和延长中起着重要作用。
10.这几类酶协同作用合成水稻中的淀粉,水解不正常的淀粉结构,使淀粉在淀粉体中形成淀粉颗粒,最终形成结构紧密多面体的淀粉晶体结构,这些酶中的任意一个酶发生变化,或者调控这些酶的相关基因发生变化,都会对淀粉的合成与积累产生影响,影响水稻籽粒品质
11.例如:编码agpase亚基的基因发生改变会导致水稻胚乳中的淀粉颗粒不正常,淀粉含量下降。gbssi基因的改变则会影响淀粉的合成,产生waxy胚乳。osssiiia的突变会导致白心粉质胚乳,影响胚乳中淀粉的理化性质。此外淀粉合成酶和淀粉分支酶的不同亚家族的基因发生突变如:osssi,osbei,osbeiib都产生了粉质胚乳性状。
12.除了涉及淀粉合成的酶,涉及其他代谢途径基因发生改变也会间接影响淀粉的合成,导致不正常的胚乳。ospk2,ospkα1都编码一个质体丙酮酸激酶,osppdk编码一个丙酮酸磷酸二激酶,二者都属于糖酵解途径,但是影响淀粉颗粒的形成和籽粒灌浆。du1编码一个mrna前体加工蛋白家族成员,主要在水稻穗部表达,通过特异的影响wxb基因的沉默来调控调控淀粉的合成。wx基因是水稻中控制直链淀粉含量的一个主效qtl位点。pfp1编码一个焦磷酸果糖6-磷酸1-磷酸转移酶的β亚基,在糖酵解途径中焦磷酸果糖6-磷酸1-磷酸转移酶将果糖6-磷酸和焦磷酸转换成果糖1,6二磷酸,pfp1以异四聚体的形式,可逆的调控胚乳的代谢流,调控淀粉合成和籽粒发育。flo15编码一个质体乙二醛酶(osglyi),主要参与甲基乙二醛解毒过程,osglyi的改变会导致淀粉合成相关基因表达改变,产生白心粉质胚乳。flo11编码一个质体热激蛋白(oshsp70cp-2)其突变会导致水稻籽粒白心粉质和外周粉质;该基因在发育中的胚乳上大量表达,oshsp70cp-2与tic复合体作用调控蛋白进淀粉体的运输。flo16编码一个nad依赖的胞质苹果酸脱氢酶,其主要功能是控制水稻胚乳中的氧化还原稳态,而氧化还原稳态对淀粉颗粒的形成和后续的淀粉合成都十分重要。过表达flo16水稻籽粒的千粒重显著的增加。
13.有一些间接影响淀粉合成的基因编码的蛋白则具有特定结构。核定位的基因flo2,具有tpr结合位点结构,通过与具有basic helix-loop-helix(bhlh)结构的转录因子结合,正向调控淀粉合成基因的表达。flo6是一个具有carbohydrate-binding domain(cbm)的蛋白,通过与淀粉异构酶(isa1)相互作用调控淀粉的合成与淀粉颗粒的形成。osnppr1是一个核定位的三角状五肽重复结构蛋白(pentatricopeptie repeat protein,ppr),编码的蛋白与cucacmotif结合,调控特定内含子的剪接,影响胚乳中线粒体的发育,继而影响胚乳的发育。flo10同样编码一个三角状五肽重复结构蛋白(pentatricopeptie repeat protein,ppr),该蛋白由核基因编码定位于线粒体上,主要参与线粒体nad1第一个内含子的反式沉默,由于nad1编码线粒体中nadh电子传递链的复合体i的核心成分,flo10的变化,影响了nad1的正确剪接,影响胚乳的发育,形成粉质胚乳。此外还有osndufa9,osalat1,osbt1等基因通过调控淀粉合成酶间接调控胚乳中淀粉合成。
14.水稻胚乳中的蛋白质主要由醇溶蛋白,球蛋白,谷蛋白,白蛋白组成,其中谷蛋白约占贮藏蛋白的80%左右。主要储存在胚乳中的蛋白体i(protein body i,pbi)和蛋白体ii(protein body ii,pbii)。蛋白合成后谷蛋白和球蛋白以及白蛋白通过液泡分选途径储存在液泡中,最后发育成pbii,pbii没有固定形状;醇溶蛋白通过内质网驻留途径储存在内
质网中,最后发育成pbi,pbi为球形囊泡状。
15.现有的研究中对谷蛋白的运输调控已经有一些进展,谷蛋白前体在粗面内质网合成后通过致密囊泡介导的后高尔基体运输途径或者内质网衍生的前体积累腔转移到pbii。谷蛋白前体的不正常积累会导致水稻胚乳出现粉质胚乳影响稻米品质。例如:got1b编码一个一个高尔基体转运蛋白,作为一个copii的一个组分,调控蛋白从内质网到高尔基体的运输。gpa3编码的蛋白作用类似于一个支架,招募鸟嘌呤核苷酸交换因子osvps9a。vps9a继而激活小g蛋白osrab5a。gpa3,osvps9a,osrab5a在致密囊泡上组合形成一个复合体调控谷蛋白前体的液泡运输。pdil1-1编码一个类二硫异构酶,定位于粗面内质网,调控内质网内的二硫键的形成。glup3编码一个液泡加工蛋白,主要作用是在pbii中将谷蛋白前体加工成酸性和碱性亚基。osnhx5编码一个na+/h+反向转运蛋白,定位在高尔基体,反面网状结构,主要在前液泡腔内维持ph稳态,对由致密囊泡介导的谷蛋白运输具有非常重要的作用。
16.成熟的水稻胚乳中,脂质只占胚乳极少部分。但是脂质的变化却会对胚乳额外观和品质造成影响。例如:fse1(floury shrunken endosperm1)编码一个磷脂酶样的蛋白,定位于细胞质基质和胞内膜,当fse1突变时水稻胚乳中淀粉粒不正常发育,总淀粉和支链淀粉含量下降,脂质和总蛋白含量上升。研究表明fse1主要控制水稻胚乳中半乳糖脂的合成,同时也是脂质代谢和淀粉合成之间的联系。osltpl36编码一个脂质转运蛋白,在发育的种皮和糊粉层中特异表达。下调osltpl36的表达量会导致水稻结实率降低,千粒重下降。osltpl36表达抑制以后不仅会导致脂肪含量降低,还会导致水稻胚的发育迟缓。
17.在胚乳的细胞分裂和分化以及物质积累过程中,细胞骨架发挥了重要作用。细胞骨架中的微观及相关的微观结合蛋白结构和功能发生改变都会对水稻的粒型和胚乳品质造成影响。例如水稻mt定位蛋白osiq67-domain14(osiqd14),在水稻种子壳细胞中高度表达。当osiqd14缺乏时,种子短而宽,总产量增加,过度表达却导致种子窄而长,osiqd14通过影响mt动力学来调控介导的mt重排序,从而调控水稻籽粒的局部细胞形态,改变水稻产量性状。
18.在玉米中vks1编码zmkin11,属于kinesin-14亚家族,主要在胚乳发育早期表达。vks1动态定位于细胞核和微管,在细胞的迁移过程中起关键作用在有丝分裂早期的有丝分裂和胞质分裂中的游离核。缺少vks1不会致死,但会导致纺锤体组装、姐妹染色单体分离和隔膜形成等畸形导致细胞增殖减少,从而导致不同的种子大小。
19.尽管本领域对于植物的胚乳发育已有较多的研究和关注,但鉴于水稻等植物为重要粮食作物,本领域亟待找到很多的影响植物胚乳发育的基因,以利于植物的进一步育种改良。
技术实现要素:
20.本发明的目的在于提供一种控制胚乳发育及品质的新型基因flo18。
21.在本发明的第一方面,提供一种flo18蛋白或其调节分子的用途,用于调节禾本科植物的产量性状、胚乳性状或淀粉发育性状。
22.在一个优选例中,所述的flo18蛋白包括其同源物。
23.在另一优选例中,所述调节分子为上调分子,所述flo18蛋白或其上调分子的用途包括选自:(i)提高产量;较佳地包括增加籽粒的粒宽或粒厚,或促进籽粒灌浆;(ii)提高胚
no:2所示氨基酸序列的多肽的活性片段;或(v)在seq id no:2所示氨基酸序列的多肽的n或c末端添加标签序列或酶切位点序列,或在其n末端添加信号肽序列后形成的多肽。
35.在本发明的另一方面,提供flo18蛋白或其编码基因的用途,用作鉴定禾本科植物性状的分子标记物;所述性状包括:产量性状、胚乳性状或淀粉发育性状。
36.在本发明的另一方面,提供一种定向选择或鉴定产量高、胚乳品质高或淀粉发育好的禾本科植物的方法,包括:鉴定测试植物体内flo18蛋白的表达或活性,若该测试植物中flo18蛋白的表达或活性高于或等于该类植物(对照植物)中flo18蛋白的表达或活性的平均值,则其为产量高、胚乳品质高或淀粉发育好(如籽粒饱满、灌浆好、淀粉发育好、淀粉颗粒直径大或能形成角质胚乳)的禾本科植物。
37.在一个优选例中,所述高表达或高活性,是指与同类或同种植物的表达或活性的平均值相比,表达或活性具有统计学意义的提高,如提高10%、20%、40%、60%、80%、90%或更高。
38.在另一优选例中,所述低表达或低活性,是指与同类或同种植物的表达或活性的平均值相比,表达或活性具有统计学意义的降低,如降低10%、20%、40%、60%、80%、90%或更低。
39.在另一优选例中,所述“量高”/“品质高(或好)”是指与同类或同种植物的量相比,有统计学意义的高,如高10%、20%、40%、60%、80%、90%或更高。
40.在另一优选例中,所述量低是指与同类或同种植物的量相比,有统计学意义的低,如低10%、20%、40%、60%、80%、90%或更低。
41.在本发明的另一方面,提供一种筛选促进禾本科植物性状改良的物质(潜在物质)的方法,所述性状改良包括:产量高、胚乳品质高或淀粉发育好;所述方法包括:(1)将候选物质加入到表达flo18蛋白的体系中;(2)检测所述体系,观测其中flo18蛋白的表达或活性,若其表达或活性提高(显著提高,如提高10%、20%、40%、60%、80%、90%或更高),则表明该候选物质为促进禾本科植物性状改良的物质。
42.在一个优选例中,所述方法还包括:设置不添加所述候选物质的对照组,从而明确分辨测试组中所述flo18蛋白表达或活性与对照组的差异。
43.在另一优选例中,所述的候选物质包括(但不限于):针对所述flo18蛋白或其编码基因或其上游或下游蛋白或基因设计的调控分子(如上调剂、小分子化合物基因编辑构建物等。
44.在本发明的另一方面,提供一种禾本科植物细胞、组织或器官,其中含有外源的flo18蛋白的上调分子,所述上调分子包括选自:过表达所述flo18蛋白的表达盒或表达构建物(包括表达载体);或提高所述flo18蛋白翻译效率的表达盒或表达构建物;或与所述flo18蛋白相互作用、从而提高其表达或活性的上调分子。
45.在一个优选例中,所述的植物细胞、组织或器官不具有繁殖能力。
46.本发明的其它方面由于本文的公开内容,对本领域的技术人员而言是显而易见的。
附图说明
47.图1a、突变体材料xa80与9311杂交,构建定位群体的流程示意图。
48.图1b、突变体与野生型相比的突变位点的确定。
49.图1c、neightbor-joining法构建的系统进化树。
50.图2a、突变体与野生型植株的表型考察,左边为野生型,右边为突变体bar=20cm。
51.图2b、获取突变体与野生型植株的籽粒,脱去颖壳后进行外观比较(左图)以及在体式镜下观察(右图)。bar=1cm。
52.图2c、突变体与野生型植株的籽粒的千粒重(左图)和灌浆速率(右图)。
53.图2d、突变体与野生型植株的籽粒的粒宽比较(上图)、粒厚比较(下图)。
54.图2e、突变体(下图)与野生型(上图)植株的籽粒的整粒、横断面以及淀粉粒状态的扫描电镜照片。
55.图2f~g、光学显微镜(f)和透射电镜(g)下胚乳的细胞结构照片。
56.图3a、授粉后6天的胚乳中,突变体与野生型植株的维管束周围细胞中细胞核情况。
57.图3b、zeiss lsm880激光共聚焦显微镜下观察flo18在细胞中的表达位置。
58.图3c、flo18与tub6的共转实验。
具体实施方式
59.本发明人经过广泛的研究,克隆获得了一种新型的与调控植物产量性状或胚乳品质相关的基因flo18,其为产量或胚乳品质的正调控基因,其功能缺失则会导致胚乳产生粉质胚乳表型,产量和品质下降,但是对于植物其他性状没有影响。本发明为植物的性状改良提供了新靶点。
60.术语
61.如本文所用,所述的“植物”包括表达flo18或包含flo18及其所参与的信号通路的植物。根据本领域的知识,表达flo18的植物,其内在存在如本发明所主张的作用机制,可以实现如本发明所主张的技术效果。在一些优选方式中,所述的植物为作物,较佳地为禾谷类作物,所述禾谷类作物为具有籽粒(穗粒)的作物。所述的“禾谷类作物”可以是禾本科植物。较佳地,所述的禾本科植物包括:水稻,大麦、小麦、燕麦、黑麦、玉米、高粱、二穗短柄草等。
62.如本文所用,术语“提高”、“改良”或“增强”是相互可以交换的并且在应用含义上应当意指与本文中定义的对照植物相比较,至少2%、3%、4%、5%、6%、7%、8%、9%或10%、优选的至少15%或20%、更优选25%、30%更高的调节。
63.关于“对照植物”,选择合适的对照植物是实验设计的例行部分,可以包括对应的野生型植物或无目的基因的相应转基因植物。对照植物一般是相同的植物物种或甚至是与待评估植物相同或属于同一类的品种。对照植物也可以是因分离而丢失转基因植物的个体。如本文所用的对照植物不仅指完整植物,也指植物部分,包括种子和种子部分。
64.如本发明所用,所述的“籽粒”是指植物的果实或种子,在水稻、玉米、小麦、大麦等作物中也称为穗粒。
65.flo18基因及其所编码的蛋白
66.在本发明中,除非特别说明,所述flo18蛋白包括了其同源物(同源蛋白)。所述flo18为具有seq id no:2所示氨基酸序列的多肽(蛋白)。本发明还包括具有与flo18蛋白相同功能的序列变异形式。
67.所述的变异形式包括(但并不限于):若干个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个,还更佳如1-8个、1-5个)氨基酸的缺失、插入和/或取代,以及在c末端和/或n末端添加或缺失一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。任何与所述的flo18蛋白同源性高(比如与seq id no:2所示的多肽序列的同源性为70%或更高;优选地同源性为80%或更高;更优选地同源性为90%或更高,如同源性95%,98%或99%)的、且具有所述flo18蛋白相同功能的蛋白也包括在本发明内。
68.来源于水稻以外其它物种的与seq id no:2所示序列的多肽序列的同源性较高、或在同样或相近的信号通路中发挥同样或相近作用的多肽也包括在本发明中。
69.本发明中,所述的“flo18蛋白”也包括它们的同源物。应理解,虽然本发明中优选研究了获自特定物种的flo18蛋白,但是获自其它物种、特别是禾本科植物的与所述flo18蛋白高度同源(如具有70%以上,更特别80%,85%、90%、95%、甚至98%以上序列相同性)的其它多肽或基因也在本发明考虑的范围之内。
70.本发明还提供了分离的蛋白,其是flo18蛋白的片段或在两端添加其它蛋白或标签等形成的。
71.本发明还涉及编码本发明的flo18蛋白或其序列变异形式的多核苷酸序列。所述的多核苷酸可以是dna形式或rna形式。dna形式包括cdna、基因组dna或人工合成的dna。dna可以是单链的或是双链的。dna可以是编码链或非编码链。编码成熟多肽的编码区序列可以与seq id no:1所示的编码区序列相同或者是简并的变异体。如本文所用,“简并的变异体”在本发明中是指编码具有seq id no:2序列的多肽,但与seq id no:3所示的基因组序列或seq id no:1所示的编码区序列有差别的核酸序列。本发明还涉及上述多核苷酸的变异体(变体),其编码与本发明有相同的氨基酸序列的多肽或多肽的片段、类似物和衍生物。
72.本发明也涉及包含所述的多核苷酸的载体,以及用所述的载体或多肽编码核酸经基因工程产生的宿主细胞。
73.本发明中,编码本发明的多肽的多核苷酸序列可插入到重组表达载体中。术语“重组表达载体”指本领域熟知的细菌质粒、噬菌体、酵母质粒、植物细胞病毒、哺乳动物细胞病毒或其他载体。总之,只要能在宿主体内复制和稳定,任何质粒和载体都可以用。表达载体的一个重要特征是通常含有复制起点、启动子、标记基因和翻译控制元件。较佳地,所述表达载体还可选择性地加入抗性元件、筛选(选择)元件或报告基因元件,如bar、gus。
74.所述的多核苷酸在高等真核细胞中表达时,如果在载体中插入增强子序列时将会使转录得到增强。增强子是dna的顺式作用因子,通常大约有10到300个碱基对,作用于启动子以增强基因的转录。
75.用重组dna转化宿主细胞可用本领域技术人员熟知的常规技术进行。转化植物可使用农杆菌转化或基因枪转化等方法,例如喷洒法、叶盘法、水稻幼胚转化法等。
76.植物改造
77.本发明人通过大量的系统性研究,克隆获得flo18的全长序列并鉴定了其生物学功能,该基因在水稻胚乳发育过程中发挥了重要作用,具有通过控制胚乳细胞分裂和发育,影响胚乳整个结构的发育,进一步影响贮藏物质的积累从而影响胚乳品质。flo18突变体产量降低但是却对每穗粒数,穗长,穗型没有显著影响。有利于更深入地理解和解析细胞骨架在胚乳发育过程中的作用。
78.在具体的实施例中,本发明人通过基因定位群体构建及表型考察,发现flo18的突变体千粒重显著低于野生型千粒重、粒宽显著小于野生型粒宽、粒厚显著小于野生型粒厚。突变体的灌浆速率前3天与正常个体相比并没有明显,第6天开始干物质积累逐渐减缓;最终成熟后干重明显低于野生型。突变体淀粉发育不完全,淀粉颗粒明显偏小,淀粉粒呈小球状,淀粉粒之间空隙较大。并且,突变体的淀粉体发育明显慢于野生型,而且晶体结构疏松,颗粒小,很少能观察到蛋白体的存在。
79.在具体的实施例中,本发明人通过回补实验发现,flo18回补的xa80突变体呈现粳稻il-9野生型的表型,其粒形饱满光滑,横断面透明晶莹紧致;其千粒重也与野生型植株的千粒重、粒宽和粒厚基本相同。
80.在具体的实施例中,本发明人还发现,在胚乳发育过程中,突变体存在细胞核不均等分裂,从而确定flo18与细胞分裂相关进而调控胚乳品质。
81.基于本发明人的新发现,提供了一种flo18蛋白或其调节分子的用途,用于:调节禾本科植物的产量性状、胚乳性状或淀粉发育性状。
82.同时,本发明也提供了一种调控禾本科植物的产量性状、胚乳性状或淀粉发育性状的方法,包括:在植物中调控flo18蛋白的表达或活性;其中,所述的flo18蛋白包括其同源物。
83.应理解,在得知了所述flo18蛋白在禾本科植物的产量性状、胚乳性状或淀粉发育性状调控中的作用后,可以根据实际所需,采用本领域人员熟知的多种方法来调节所述的flo18蛋白的表达或活性,这些方法均被包含在本发明中。
84.可以利用flo18蛋白的表达或活性的上调剂来上调flo18蛋白的活性。所述的flo18蛋白的表达或活性的上调剂包括了促进剂、激动剂、激活剂。所述的“上调”、“促进”包括了蛋白活性的“上调”、“促进”或蛋白表达的“上调”、“促进”。任何可提高flo18蛋白的活性、提高flo18蛋白基因或蛋白的稳定性、上调flo18蛋白基因的表达、增加flo18蛋白的有效作用时间的物质,这些物质均可用于本发明,作为对于上调flo18蛋白或其编码的蛋白有用的物质。它们可以是化合物、化学小分子、生物分子。所述的生物分子可以是核酸水平(包括dna、rna)的,也可以是蛋白水平的。
85.作为一种优选的实施方式,提供一种上调植物中flo18蛋白的表达的方法,所述的方法包括:将flo18蛋白或其编码的蛋白的表达构建物或载体转入植物中。
86.优选地,提供了一种制备转基因植物的方法,包括:
87.(1)将外源的本发明的多肽的编码核酸转入植物器官或组织,获得转化入所述多肽的编码核酸的植物组织或器官;和
88.(2)将步骤(1)获得的转入了外源的本发明的多肽的编码核酸的植物组织或器官再生成植物植株。
89.作为一种优选的实例,所述的方法包括步骤:
90.(s1)提供携带表达载体的农杆菌,所述的表达载体含有本发明的flo18蛋白的编码核酸;
91.(s2)将植物组织或器官与步骤(s1)中的农杆菌接触,从而使所述多肽的编码核酸转入并且整合到植物细胞的染色体上;
92.(s3)选择出转入所述flo18蛋白的编码核酸的植物组织或器官;以及
93.(s4)将步骤(s3)中的植物组织或器官再生成植物。
94.本发明还包括利用前述任一种方法获得的植物,所述的植物包括:转入了所述多肽的编码核酸的转基因植物。
95.作为一种可选择的方式,敲除所述flo18基因的5’utr上的上游开放阅读框(upstream open reading frame;uorf),提高所述flo18基因翻译效率的表达盒或表达构建物。uorf广泛存在于真核生物mrna中,其为一种翻译调控元件。通常,uorf的翻译优先于morf(主要开放阅读框),导致morf翻译受阻。本发明中,靶向敲除所述flo18基因的5’utr上的uorf,从而可有效提高所述flo18蛋白翻译效率,提高其表达。
96.本发明中,所述的flo18蛋白或其编码基因的下调剂是指任何可降低flo18蛋白的活性、降低flo18蛋白或其编码基因的稳定性、下调flo18蛋白的表达、减少flo18蛋白有效作用时间、抑制flo18基因的转录和翻译的物质、或降低蛋白的磷酸化/激活水平,这些物质均可用于本发明,作为对于下调flo18蛋白有用的物质。它们可以是化合物、化学小分子、生物分子。所述的生物分子可以是核酸水平(包括dna、rna)的,也可以是蛋白水平的。例如,所述的下调剂是:特异性干扰flo18蛋白或其它信号通路基因表达的干扰rna分子或反义核苷酸;或是特异性编辑flo18基因的基因编辑试剂,等等。
97.作为本发明的一种优选方式,提供一种下调植物中flo18蛋白的方法,包括对flo18蛋白进行靶向性地突变、基因编辑或基因重组,从而实现下调。作为一种更为具体的实施例方式,藉由上述任一的方法,使flo18蛋白转变为其突变体,从而使其不再发挥作用。作为一种更为具体的实施例方式,采用crispr/cas9系统进行基因编辑。合适的sgrna靶位点,会带来更高的基因编辑效率,所以在着手进行基因编辑前,可以设计并找到合适的靶位点。在设计特异性靶位点后,还需要进行体外细胞活性筛选,以获得有效的靶位点用于后续实验。本发明的实施例中提供了优选的基因编辑试剂。
98.作为其它可选的方式,所述下调植物中flo18蛋白的表达的方法可包括:(1)将干扰flo18基因表达的干扰分子转入植物细胞、组织、器官或种子,获得转化入所述干扰分子的植物细胞、组织、器官或种子;(2)将步骤(1)获得的转入了所述干扰分子的植物细胞、组织、器官或种子再生成植物。较佳地,所述方法还包括:(3)选择出转入了所述载体的植物细胞、组织或器官;和(4)将步骤(3)中的植物细胞、组织或器官再生成植物。
99.可采用任何适当的常规手段,包括试剂、温度、压力条件等来实施所述的方法。
100.植物定向筛选及分子标记
101.基于本发明人的新发现,本发明提供了适用于鉴定产量高、胚乳品质高或淀粉发育好的植物的分子标记,即flo18基因。本发明还涉及针对所述flo18基因设计的特异性分子标记,以及鉴定策略。
102.因此,本发明提供了一种定向选择或鉴定农艺性状被调节的植物的方法,包括:鉴定测试植物体内flo18蛋白的表达或活性,若该测试植物中flo18蛋白的表达或活性高于该类植物(对照植物)中flo18蛋白的表达或活性的平均值,则其为产量高,籽粒饱满、灌浆好、淀粉发育好、淀粉颗粒直径大或能形成角质胚乳的禾本科植物;或,若该测试植物中flo18蛋白的表达或活性低于该类植物(对照植物)中flo18蛋白的表达或活性的平均值,则其为产量低,籽粒不饱满、灌浆不理想、淀粉发育不理想好、淀粉颗粒直径小或能形成粉质胚乳的禾本科植物。
103.根据本发明的新发现,本领域技术人员可以采用任何本领域公知的或正在发展的多种技术来进行核酸序列的分析,这些技术均可被包含在本发明中。所述的方法例如包括但不限于:测序法,pcr扩增法,探针法,杂交法,限制性酶切分析法,等位基因多态性分析法(如溶解曲线法)进行核酸序列的鉴定,等等。
104.本发明的鉴定方法,只需进行pcr反应和/或琼脂糖凝胶电泳,并通过判断相应的pcr产物的长度,就可以准确、快速地判断待测样品的表型或产量,成本低廉,适合于大规模鉴定,而且所需的样品量很少。如果需要,本领域技术人员能够设计出鉴定所述分子标记的引物。
105.获取待测样品的dna的方法是本领域技术人员所熟知的技术,例如可采取传统的酚/氯仿/异戊醇法,或者可采用一些商购的dna提取试剂盒,这类试剂盒是本领域技术人员熟知的。聚合酶链反应(pcr)技术是本领域技术人员熟知的技术,其基本原理是体外酶促合成特异dna片段的方法。本发明的方法可采用常规的pcr技术进行。
106.本发明在分子设计育种及利用基因工程技术进行农作物品种改良等方面具有良好的应用前景。
107.在得知了flo18基因的功能以后,可以以其为分子标记物,来进行植物的定向筛选。也可基于该新发现来筛选通过调节这一机制,从而定向调控产量、胚乳品质或淀粉发育的物质或潜在物质。
108.本发明提供了一种筛选促进禾本科植物性状改良的物质(潜在物质)的方法,所述性状改良包括:产量高、胚乳品质高或淀粉发育好;该方法包括:(1)将候选物质加入到表达flo18蛋白的体系中;(2)检测所述体系,观测其中flo18蛋白的表达或活性,若其表达或活性提高,则表明该候选物质为促进禾本科植物性状改良的物质。
109.以蛋白或基因或其上特定的区域作为靶点,来筛选作用于该靶点的物质的方法是本领域人员所熟知的,这些方法均可用于本发明。所述的候选物质可以选自:肽、聚合肽、拟肽、非肽化合物、碳水化合物、脂、抗体或抗体片段、配体、有机小分子、无机小分子和核酸序列等。根据待筛选的物质的种类,本领域人员清楚如何选择适用的筛选方法。
110.检测蛋白与蛋白之间相互作用以及相互作用的强弱可采用多种本领域技术人员熟知的技术,比如gst沉降技术(gst-pull down)、双分子荧光互补实验、酵母双杂交系统或免疫共沉淀技术等。
111.经过大规模的筛选,可以获得一类特异性作用于flo18蛋白或其编码基因,对禾本科植物性状改良有调控作用的物质。
112.下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件如j.萨姆布鲁克等编著,分子克隆实验指南,第三版,科学出版社,2002中所述的条件,或按照制造厂商所建议的条件。
113.材料和方法
114.1、突变体材料xa80的获取和考察
115.对突变体材料xa80,收获种子后进行水稻籽粒脱壳处理。日本satake种子判别仪器和万深种子考察系统测量水稻粒长,粒宽,和粒厚,进行统计分析。
116.2、扫描电镜观察种子表面
117.样品准备:随机选取完全成熟的水稻籽粒在纯水中超声波处理10分钟,42℃烘箱中干燥三天。
118.样品固定:将双面胶黏贴在铜台的合适位置上,用镊子夹取样品,确保观察面向上并粘在铜台的胶上。涂上导电胶,确保观察面干净。
119.镀膜:样品处理好后进行真空喷金镀膜。
120.观察:样品制备好后,使用日本岛津jsm-6360lv型扫描电子显微镜进行观察。
121.3、半薄切片
122.将用于组织学观察的材料放入装有faa固定液的离心管。将离心管置于冰上抽真空,将固定液充分渗入组织中,固定材料内部结构。待所有组织都沉没于固定液后将faa固定液重新换一次,4℃保存。在使用梯度浓度的乙醇脱水后,将材料包埋在环氧树脂中进行半薄切片。
123.4、大肠杆菌转化和农杆菌转化
124.大肠杆菌转化:将保存于-80℃冰箱的的感受态dh5α大肠杆菌感受态细胞置于冰上5min。将适量质粒加入处于冰水混合态的感受态细胞中,轻轻拨动管底,于冰上静置15min。42℃热激40s后迅速将感受态细胞置于冰上。2分钟后加入500微升的无抗lb液体培养基。将感受态细胞置于37℃培养箱孵育1小时,把菌液涂于含有相应抗性的lb固体培养基上。37℃培养过夜。
125.农杆菌转化:将农杆菌感受态细胞eha105从-80℃冰箱中取出,置于冰上解冻。在感受态细胞中加入0.01~1微克的质粒,拨动管底混匀。按照冰上5min,液氮中5min,37℃5min,冰上5min的顺序操作。加入500微升无抗yep液体培养基,28℃孵育2~3小时。5000rpm,1min离心富集菌落,管中留有100微升作用的菌液。将富集后的菌液涂于相应抗性的yep固体培养基上。28℃培养箱培养2天。
126.5、flo18的亚细胞定位
127.(1)flo18-gfp融合蛋白构建
128.为了观察glw7的细胞亚定位,本发明人构建了flo18-gfp融合蛋白。经测序验证正确后,化学方法转化原生质体。
129.水稻原生质体的培养:水稻种子消毒在超净台中晾干后,将种子播于1/2ms培养基上30℃培养7~10天。将水稻幼苗剪去根和叶子,使用茎和叶鞘的绿色组织。用刀片切成0.5mm的小段将切下的小段立刻转移至20~30ml的0.6m d-甘露醇的50ml离心管(锡箔纸包裹好)中黑暗10min。
130.配制酶解液,配方如表1。
131.表1
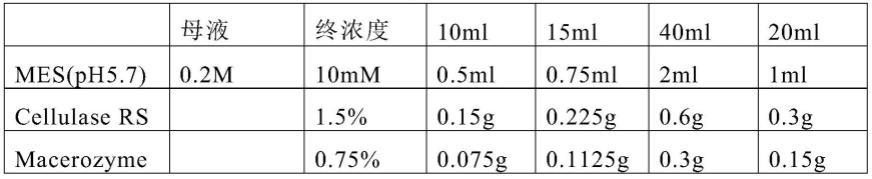
132.[0133][0134]
酶解液配制过程如下(以20ml为例,具体用量以样品量为参照):取1ml 0.2m mes(ph5.7)溶液加热到70℃,3~5分钟;分别加入0.3g cellulase rs,0.15g macerozyme r-10,15ml 0.8m甘露醇,溶液55℃,10min;冷却至室温(25℃),分别加入200ul 1m cacl2和200ul 100mg/ml bsa,加水补齐至20ml;用40um的注射器和0.45um的滤头将酶液过滤到另一个50ml离心管中。
[0135]
准备一个用锡箔纸包住的小锥形瓶,除去步骤4中甘露醇,将小段浸在酶液中。可将锥形瓶真空抽滤25min,以达到使酶解液充分浸润水稻材料。放在室温为25℃的摇床上,转速为60,放置4小时。向消化后的锥形瓶中加入20ml的w5(与酶解液同体积),手中剧烈震荡10s。w5的配制如表2。
[0136]
表2
[0137] 终浓度母液浓度100ml200mlnacl154mm(0.9%) 0.9g1.8gcacl2125mm(1.84%)1m12.5ml25ml无水葡萄糖5mm(0.1%) 0.090.18gkcl5mm1m500ul1mlmes(ph5.7)2mm0.2m1ml2ml
[0138]
用40um尼龙网过滤原生质体入50ml离心管中,用w52溶液将锥形瓶冲洗干净,平衡离心,100xg,(acc 8;dec 5),离心两分钟,吸掉上清。加入10ml w5,温和混匀,100xg,(acc 8;dec 5),离心两分钟,吸掉上清。用1ml的mmg重悬,将离心管中的原生质体转移至2ml ep管,血球计数板计数。mmg配方如表3。
[0139]
表3
[0140] 母液浓度终浓度2ml5ml8ml10ml甘露醇0.8m0.5m1250ul3.125ml5ml6.25mlmgcl21m15mm30ul75ul120ul150ulmes(ph5.7)0.2m4mm40ul100ul160ul200ul
[0141]
100xg,(acc 8;dec 5),离心两分钟,吸掉上清,根据之前镜检的情况加入相应体积的mmg重悬原生质体。
[0142]
(2)peg介导的转染
[0143]
peg溶液的配方如表4。
[0144]
表4
[0145] 母液浓度终浓度5mlpeg4000 40%(w/v)2g甘露醇0.8m0.2m1.25ml
[0146]
peg溶液配好后,应放于65℃烘箱使其融化,使用时冷却至室温即可。
[0147]
转染步骤:
①
准备2ml离心管,将之前准备好的10ugdna加到管底(一般100μl的原生质体加10ugdna,对于bifc或共表达系统dna含量可达10ug和15ug),加入100ul的原生质体,暂时不要混匀。
②
加入110ul新配的peg溶液,加peg溶液时,将pe管稍微平一点放,然后顺着管壁,缓慢加入,peg加完一个样,就立即温和混匀。
③
室温黑暗放置10-20min。
④
缓慢加入440ul w5溶液,终止反应,100xg,升8降5(acc 8;dec 5),离心两分钟;弃上清。
⑤
重复步骤
④
。
⑥
500ul w5溶液重悬。
⑦
平放在纸盒中,放入26℃培养箱,培养6-16小时。
[0148]
6、序列信息
[0149]
flo18基因序列,第1~126位碱基为5’utr,第4716~5171位为3’utr,黄色为蛋白编码区。
[0150]
osflo18(loc_os07g32390)基因序列(seq id no:3):
[0151]
gatccaaccaccaccaccttcttctcctcctcttccgctcgacgccatcaccacgcacttgagagagagagagagagagagagagagagaaaccctaagcctcgcccgctatccagccaagcaccgatggcgacggacgcgaacccggaggcggcggcgccgccgccgcagttactggtggacgagggctacgagttctgcgcgcccaagttcttcgacttcgtctgcgacgagacggaggaggaaatccgcgccgccgagcgctggttcgaggcctccgccagccacgccccttcccgtacgctccccaatccccaccccgccgcctcgatcatccccccctctcccgtgttttgctttttttgcgctgattagggctcttcttcttttacctgcttgcagcgttcgctccgaggatcaaggagtcgagggcggaggtcaagatcgagagcctctgcgacttcaccgacgcggagccgattccgaaggtacgaagcagaaagctgcgggaataaccccccaattttgccctaatttctgtaaatctccccccttcttattgtccgttggcctctgcgttcgtcatgataggaggtagcagtggaggaggcagcaggaagcgccgccaatccctcgcagaattctgatgggtaagagttggagaactaaagaaattgtttatgatgattcgttgtgatctgatgaagttaatcgatttctcgtgtgtttttcatcttcacttttggatgttcaggaatgtgcaacagaataaggacggctccatcaaacttgtacgtgatttcttgatgttttcatgttttctatgttctcagtttcagtaagtactagtgttgggtttctagcagctcacatcctgaattcttcaggtccatgaagcaaatcccagtgaaaattgtgtcactgacggcgatcataagcaccaagaaaggtgcatttcaagacctcaatttgttggttcattccgtttcttgctggttttggcatgtgaatgactaattggttgttttgggcagtgatgcaatgttggagtcgccaccagcagaggaggatgagaaggaatcgccaaaatccttcgagtttgtcccttccaatgcaaaatgtaagttgctagttatatatgagcaatggcagtggggaagaactctgatccctggattttgatatcaatgttgcatggctttagcagcagatgttgcttctagcacaccgaagattcagaggcctccacccgtcaaagctgtcactacggtgcctacctgtcccaagctgacagtgaagacagaagccttcactccgaaggtgcaggcaacgaactcctccagaggtcttgcacccttgactggctcaagggcacatccatctgctttgaagcagtcgatgagcgtcaagaggagtgtgatcaagtaagagtcatgctcactcataaaaaaatatattaatctggagattattttcttactaatgtctgaattgttggtttttcagatgccctcgtgagttgctggctgggaaggccgctactgctgcgaatgaaatcgcacaagaaaatcaagctgtcaagagacagaagcttgatgatgggaggacaagacaggtaatactgggatgagattaaagatggcacttcaaatacatgtgtttgcgttgcgcgagatttctgagaaacattgtgtttgccgtcgtggttagatactgaacgtgaagacaaggaccctgcctcacaaggggagaggaggtggtctagcaggaagcaccgagatgagcctgtcggccatgaggaagcaccgtgacgattcacactctctcaaggtagctcccaagttgctcttctgactcttaaatgatagacttaattgtaatgtttcttgtgagaatgcaagtataatgtgattttgttattgtgagcttggcacttagcaacttttgtttctgccatgtgcaggaagtgactcactatatatcggcagcagagatggtaaagaagttcgagtctgggactagagagttggccatccctcacaacagatcgctttctcatgttagcactcatgtgccctttattattaattttaaattgtgtgcatttctccaattttattaatatacatgtcttctcagtcttgttgt
gggtatttggttccatacaaagaccgatgtcctttttgtgatactgatatctgcatatcaaatattgcacagttattacatctgaaccgttgacgacgaaatcattattggtactaagaattttgacatattgcaatttgtgcacaatgcaggaggatgcagcaaccgcattacaaagaagaacaaagctgatgctaacaaggccaaaggaacctgagttccagacctcccacagggtccgtgctgtcagagtgaaaagctctgctgagctagaggaggagatgctagccaaaattccgaagttcagagcacggccgtttaacaaaaaagtgagaaatttgtttgctttggttttgattatactaactactcaatccaggatctgaagactaactatttgtttgcttctcctttcagattgcggaggccccttcatttcctcctcttccaaggaaagctccacagcttcctgaatttaatgtacgtcttgtttggctgattcagaaatttatgagctccacagctttgtttggctgattcagttattcctttccaacacctacaggagtttcatcttaaaacgatggagagagctacacgacatgcagacacctgttccgaagcttcttccgtggggactatcagggtaagctcccttcacttaacctggttgctaatatcagtatgtaaattcttcttcttattagtctcctaaaaattaaaattcttggcattactgaaagtttaatcaaatcttgcagagtcagagcagtaagccactgaccttgacagcgccaaaaccaccccaacttgagacagcattacgagctagaccaccaaggtttccctttgccacttgcatcattcagaaaacaagccattttatgttattgattattttcgattactctaacatcatctttctgcagggtgaaaagttctcaggaattggaactagaagaattagaaaaggctcccaagttcaaggcaaagccactgaacaaaaaggtctccttatcttcttgaagcttgtacatttgatacattgattgcattgtgcttcaaaacatccagtctgaacatcatttcttgcagattctcgagagcaagggtgatattggtgtgtttccacatctaaaggctcaaccaacagcccccaaggaattccatttctctaccgatgatcgcttgggtcctcctgcagttgtggatctatttgataaggtgagggttctaagtcatgctagtaagctcattattttttttccagaaacctttcttctaagtccttttctattttgtagctctctctttgctcagaatcttcatatcatagcaagaaagatgtgcccagattgacaataccaaaccccttcaatttacatacagatgtaagcagtatgaaagttatttgttttcccaacacattgcatttacctatctgttgcaggcaaataactttgttatccatgttacaggaaagagggcatgagaaagagaggcagctagctgcacaactgctgcaaaagcaattacaggaagagaaagccaggattccgaaggctaatccttatccctacacaactgactatccagtggtacgtgcaaagtaacataccgaatattacatgtcctggttgctgaccatctcatattctgctagccttaacattttctctctcttcagataccaccgaagcctgaaccaaaaccatgcacgaggccagaaggctttcagttagagagtttggtgaggcatgagatggagcaacagaggataatggaagaaagagagaggatggagagggaagaggcccagaggagagtagtgaaggcacaccccataatgaaagagtaagccttcacattacggtttctcttgtcccaacagttcctggtttcccttataagatgcatattaatactgttaattttgttgcagggatcccattcctcttccagagaaagagaggaagcctctcactgaagttcaaccacttaagttacatgttgatgaaagggcggtccaaagatcagaatttgacaatatggtcagcaaagcaagcttgttctattatctggtttggaggagcctgccaaattggctgaatttactgagcaattcgttgtcgtgcaggtgaaggaaaaggaaataacttacaagagattgcgagaggaaaatgaattcgcacaaaaggttattattttcttatgccttcttctcaaaatccctacattgttggatgcattttcatctgttctggcaactaacttgctgtgatcttttcatgtgaagattgaggaagagaaagcgttgaagcagctcaggaggaccttggtgccacaagcacggcctctcccgaagtttgacagaccatttcgtccccagaggtaagagcagaaacggccaagatttttgcctacaatgtcatcgacctttccgtttcatctcacaaacgtgaactcgttgtttcgcacacagatcaaccaagcaggtgacgaggccgaagtccccacagcttcaggtcgaccaaagaggggcaagaaggcacgccttcatcagatgatccaactctccggctcatcttgttgtcgtctatcggttacccactggctgctgctgttttctcttctgttcttgacgtcaagatcatccccaattctcgcagcgattctttctgaccaatacataaataggccgatccaatcctgttatatagtcagtcaacacagctgacggcgtgtgtatgctcctttagggttctgctgatgcttgactgcattattgtaaattcagtttgtgtgctgtgtgtgtgtgctgttcgatgaacgacaataggatgacaccctgtcagtgtcctcctcgtctgtttcgacgaatgcaagttgtattcattatttgtttatcatctctgaactatcctggagttctgtcgatgttgaataatctaattcatatctcggcacttggaaaactctcgtgagaaacatatgtaccaagaa
aaactgattcatgtcaacaa
[0152]
osflo18 cds序列(seq id no:1):
[0153]
atggcgacggacgcgaacccggaggcggcggcgccgccgccgcagttactggtggacgagggctacgagttctgcgcgcccaagttcttcgacttcgtctgcgacgagacggaggaggaaatccgcgccgccgagcgctggttcgaggcctccgccagccacgccccttccccgttcgctccgaggatcaaggagtcgagggcggaggtcaagatcgagagcctctgcgacttcaccgacgcggagccgattccgaaggaggtagcagtggaggaggcagcaggaagcgccgccaatccctcgcagaattctgatgggaatgtgcaacagaataaggacggctccatcaaacttgtccatgaagcaaatcccagtgaaaattgtgtcactgacggcgatcataagcaccaagaaagtgatgcaatgttggagtcgccaccagcagaggaggatgagaaggaatcgccaaaatccttcgagtttgtcccttccaatgcaaaatcagcagatgttgcttctagcacaccgaagattcagaggcctccacccgtcaaagctgtcactacggtgcctacctgtcccaagctgacagtgaagacagaagccttcactccgaaggtgcaggcaacgaactcctccagaggtcttgcacccttgactggctcaagggcacatccatctgctttgaagcagtcgatgagcgtcaagaggagtgtgatcaaatgccctcgtgagttgctggctgggaaggccgctactgctgcgaatgaaatcgcacaagaaaatcaagctgtcaagagacagaagcttgatgatgggaggacaagacagatactgaacgtgaagacaaggaccctgcctcacaaggggagaggaggtggtctagcaggaagcaccgagatgagcctgtcggccatgaggaagcaccgtgacgattcacactctctcaaggaagtgactcactatatatcggcagcagagatggtaaagaagttcgagtctgggactagagagttggccatccctcacaacagatcgctttctcatgaggatgcagcaaccgcattacaaagaagaacaaagctgatgctaacaaggccaaaggaacctgagttccagacctcccacagggtccgtgctgtcagagtgaaaagctctgctgagctagaggaggagatgctagccaaaattccgaagttcagagcacggccgtttaacaaaaaaattgcggaggccccttcatttcctcctcttccaaggaaagctccacagcttcctgaatttaatgagtttcatcttaaaacgatggagagagctacacgacatgcagacacctgttccgaagcttcttccgtggggactatcaggagtcagagcagtaagccactgaccttgacagcgccaaaaccaccccaacttgagacagcattacgagctagaccaccaagggtgaaaagttctcaggaattggaactagaagaattagaaaaggctcccaagttcaaggcaaagccactgaacaaaaagattctcgagagcaagggtgatattggtgtgtttccacatctaaaggctcaaccaacagcccccaaggaattccatttctctaccgatgatcgcttgggtcctcctgcagttgtggatctatttgataagctctctctttgctcagaatcttcatatcatagcaagaaagatgtgcccagattgacaataccaaaccccttcaatttacatacagatgaaagagggcatgagaaagagaggcagctagctgcacaactgctgcaaaagcaattacaggaagagaaagccaggattccgaaggctaatccttatccctacacaactgactatccagtgataccaccgaagcctgaaccaaaaccatgcacgaggccagaaggctttcagttagagagtttggtgaggcatgagatggagcaacagaggataatggaagaaagagagaggatggagagggaagaggcccagaggagagtagtgaaggcacaccccataatgaaagaggatcccattcctcttccagagaaagagaggaagcctctcactgaagttcaaccacttaagttacatgttgatgaaagggcggtccaaagatcagaatttgacaatatggtgaaggaaaaggaaataacttacaagagattgcgagaggaaaatgaattcgcacaaaagattgaggaagagaaagcgttgaagcagctcaggaggaccttggtgccacaagcacggcctctcccgaagtttgacagaccatttcgtccccagagatcaaccaagcaggtgacgaggccgaagtccccacagcttcaggtcgaccaaagaggggcaagaaggcacgccttcatcagatga
[0154]
osflo18蛋白序列(seq id no:2)
[0155]
matdanpeaaapppqllvdegyefcapkffdfvcdeteeeiraaerwfeasashapspfaprikesraevkieslcdftdaepipkevaveeaagsaanpsqnsdgnvqqnkdgsiklvheanpsencvtdgdhkhqesdamlesppaeedekespksfefvpsnaksadvasstpkiqrpppvkavttvptcpkltvkteaftpkvqatnssrglapltgsrahpsalkqsmsvkrsvikcprellagkaataaneiaqenqavkrqklddgrtrqilnvktrtlphkgrggglagstemslsamrkhrddshslkevthyisaaemvkkfesgtrelaiphnrslshedaatalqrrtklmltrpkepefqtshr
vravrvkssaeleeemlakipkfrarpfnkkiaeapsfpplprkapqlpefnefhlktmeratrhadtcseassvgtirsqsskpltltapkppqletalrarpprvkssqeleleelekapkfkakplnkkileskgdigvfphlkaqptapkefhfstddrlgppavvdlfdklslcsessyhskkdvprltipnpfnlhtderghekerqlaaqllqkqlqeekaripkanpypyttdypvippkpepkpctrpegfqleslvrhemeqqrimeerermereeaqrrvvkahpimkedpiplpekerkpltevqplklhvderavqrsefdnmvkekeitykrlreenefaqkieeekalkqlrrtlvpqarplpkfdrpfrpqrstkqvtrpkspqlqvdqrgarrhafir
[0156]
用于扩增osflo18全长基因的扩增引物:
[0157]
primerf:gatccaaccaccaccaccttcttctcctcct(seq id no:4);
[0158]
primerr:ttgttgacatgaatcagtttttcttggtac(seq id no:5)。
[0159]
实施例1、候选基因分析
[0160]
本发明人利用γ射线诱变的粳稻il-9(tohoku il9),获得了稳定遗传的粉质胚乳突变体株系,本发明人将其命名为xa80。
[0161]
本发明人利用突变体材料xa80,通过与9311杂交,构建定位群体(图1a)。通过图位克隆将基因定位在水稻7号染色体19.25m至19.26m范围内,通过rap-db数据库检索相应位置的日本晴序列,设计引物扩增覆盖式片段,利用一代测序仪测序将突变体和野生型这一区域内的序列测通,本发明人发现,在突变体序列上有一个t到a的单碱基突变(图1b)。该突变正好位于loc_os07g32390的第18个外显子上。本发明人将loc_os07g32390基因定为候选基因,称为flo18基因。
[0162]
flo18基因编码一个微管结合蛋白flo18。全长5171个碱基。在突变体植株(xa80)中,第18个外显子上碱基t到a的突变使其氨基酸编码赖氨酸(aag)变成了终止密码子(tag),导致195个氨基酸的缺失,基因功能缺失。在ricexpro(https://ricexpro.dna.affrc.go.jp/)数据库中对flo18的基因时空表达谱进行比对,发现flo18基因在水稻生殖生长阶段有特异表达,与植株表型相吻合。
[0163]
针对flo18基因,本发明人进行了在水稻中的同源性分析,利用flo18蛋白序列在msu网站数据库上获取拥有与flo18相同结构功能域一共9个基因的蛋白序列,属于tpx2蛋白家族,该家族有的基因拥有多个转录本翻译成多个蛋白。随后用mega做聚类分析,采用neightbor-joining法构建系统进化树。bootstrap值设置为2000(如图1c),从结果中可以看出tpx2蛋白在水稻中分为两支,tpx2家族具有一定的功能保守性。
[0164]
实施例2、基因定位群体构建及表型考察
[0165]
本实施例中,本发明人对xa80进行表型考察,将之与野生型植株进行比较。
[0166]
通过表型考察,本发明人发现,突变体与野生型植株相比,株高、株型、抽穗期无明显差异。代表性的植株的表型照片如图2a,左边为野生型,右边为突变体。
[0167]
本发明人获取突变体与野生型植株的籽粒,脱去颖壳后进行比较。结果显示,大小籽粒差异明显,野生型籽粒透明均一(图2b的左上图),而突变体则是胚乳不透明(图2b的左下图),籽粒偏小。在体式镜下观察的突变体和野生型籽粒,野生型粒形饱满光滑,横断面透明晶莹紧致(图2b的右上图),为角质胚乳;突变体粒形皱缩、形状偏小,横断面粗糙疏松呈粉白色,为粉质胚乳(图2b的右下图)。
[0168]
通过万深种子扫描系统测量,野生型千粒重为19.6835
±
0.211克,突变体千粒重为15.9428
±
1.551克,突变体千粒重小于野生型19%,p-value=0.004147,突变体千粒重
显著低于野生型千粒重(图2c,左图)。
[0169]
在水稻开花后,从第一天开始每隔3天,测定野生型与突变体水稻籽粒带颖壳的干重,绘制水稻灌浆速率(图2c,右图)。从结果可以看出,突变体的灌浆速率前3天与正常个体相比并没有明显,第6天开始干物质积累逐渐减缓(即:灌浆速率降低)。最终成熟后干重明显低于野生型。
[0170]
将脱壳后的籽粒进行统计和粒型的比较(图2d)。脱壳后的野生型粒长为4.8614
±
0.044cm,脱壳后突变体粒长为4.8771
±
0.040cm,p-value=0.7625,二者没有区别。脱壳后野生型粒宽为2.4604
±
0.027cm,突变体粒宽为2.3099
±
0.040cm,p-value=3.716e-11,突变体粒宽显著小于野生型粒宽。脱壳后的野生型粒厚为1.8923
±
0.016cm,脱壳后突变体粒厚为1.5811
±
0.038cm,p-value《2.2e-16,突变体粒厚显著小于野生型粒厚。
[0171]
在扫描电镜下对种子进行观察,从整粒种子来看,突变体种子干瘪,除纵轴方向的棱以外,在横轴方向也出现棱,从横断面看,野生型灌浆饱满,断裂面整齐。突变体灌浆不完全,断裂面粗糙,种皮和胚乳间存在空隙。从淀粉粒状态看,野生型淀粉粒发育完好,呈规则的结晶状,淀粉粒之间紧密排列。突变体淀粉发育不完全,淀粉颗粒明显偏小,淀粉粒呈小球状,淀粉粒之间空隙较大(图2e)。
[0172]
通过对开花后2天(2daf),4daf,6daf,8daf,10daf,12daf进行取样,在光学显微镜和透射电镜下观察胚乳的细胞结构。通过半薄切片本发明人并没有观察到突变体与野生型相比明显的细胞结构差异,但是发现其染色的深浅存在一定的差别。在透射电镜下观察发现,突变体的淀粉体发育明显慢于野生型,而且晶体结构疏松,颗粒小;并且很少能观察到蛋白体的存在(图2f,g)。
[0173]
实施例3、回补植株的建立
[0174]
本发明人以il9野生型植物的基因组为模板,以下述基因互补序列扩增引物进行扩增,获得含有flo18基因、其启动子以及上下游部分区域(不含编码区)的序列(8619bp),将之克隆到pcambia1300载体骨架上,构建获得互补载体flo18-complementary(flo18-cp)。之后,本发明人通过诱导突变体xa80成熟胚分化成愈伤组织,采用农杆菌转化的方法,将目标载体进行转化,通过共培养、筛选、预分化、分化、生根一系列步骤,获得转基因植株。利用潮霉素引物和突变位点特异引物对转基因植株进行检测,筛选到转基因阳性植株。在t0和t1代对表型进行确认。
[0175]
基因互补序列扩增引物:
[0176]
primerf:atgtgttcttaattgtggggatcagaattt(seq id no:6);
[0177]
primerr:ttagaaatctctttgttgctagctgttctc(seq id no:7)。
[0178]
对获得的t1回补植株的表型进行观测。结果显示,flo18回补的xa80突变体呈现粳稻il-9野生型的表型,其粒形饱满光滑,横断面透明晶莹紧致;其千粒重也与野生型植株的千粒重、粒宽和粒厚基本相同。
[0179]
实施例4、flo18与细胞分裂相关进而调控胚乳品质
[0180]
根据前述分析,本发明人考虑flo18的功能缺失导致突变体植株的胚乳在细胞分裂时,细胞或者染色体不均等分裂。
[0181]
为了观察是否存在这一现象,本发明人选取xa80与il9不同受精天数的胚乳。将材料固定后通过超薄切片在透射电镜下观察。本发明人发现在水稻6dap(授粉后6天)的胚乳
中,维管束周围的细胞中可以观察到细胞核。图中左边为正常个体,右边为突变体(如图3a),圆圈内为标识的细胞核。正常个体中的胚乳细胞核为深灰色,大小均一;而突变体个体的胚乳细胞核大小不一,有的和正常个体大小相差不大,有的则明显小得多。说明在胚乳发育过程中,突变体的确存在细胞核不均等分裂。
[0182]
因此,flo18与细胞分裂相关进而调控胚乳品质。
[0183]
实施例5、flo18的亚细胞定位
[0184]
扩增flo18全长cds接在pa7-gfp载体的上游,构建pa7-gfp融合蛋白,将该融合蛋白转化水稻原生质体瞬时表达,同时共同转化mcherry-tub6载体。在zeiss lsm880激光共聚焦显微镜下观察。结果显示,flo18主要在细胞质中大量表达,细胞膜和中央大液泡上没有表达(图3b)。
[0185]
mcherry-tub6主要用于标记微管蛋白,通过共转实验发现flo18与tub6存在共定位现象,说明flo18与微管蛋白存在互作(图3c)。
[0186]
总结
[0187]
本发明人克隆到基因flo18,该基因在水稻胚乳发育过程中发挥了重要作用,具有通过控制胚乳细胞分裂和发育,影响胚乳整个结构的发育,进一步影响贮藏物质的积累从而影响胚乳品质。flo18突变体产量降低但是却对每穗粒数,穗长,穗型没有显著影响。传统的品质育种中,随着品质的提升,产量会不可避免的下降。针对该基因的研究有利于本领域人员更深入地理解和解析细胞骨架在胚乳发育过程中的作用,为水稻等植物的高产优质育种提供了新的途径。
[0188]
在本发明提及的所有文献都在本技术中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本技术所附权利要求书所限定的范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1