具有从κ基因座表达的有限λ轻链谱系的非人类动物和其用途的制作方法
具有从
κ
基因座表达的有限
λ
轻链谱系的非人类动物和其用途
1.相关申请的交叉引用
2.本技术要求2019年6月5日提出的美国临时申请no.62/857,712的优先权,所述临时申请以引用的方式并入本文中。
3.序列表
4.本技术含有序列表,所述序列表已以ascii格式电子提交并以引用的方式整体并入本文中。ascii拷贝于2020年5月20日建立,名称为2010794
‑
2050_sl.txt,且大小为47,576字节。
背景技术:
5.人类抗体是增长最迅速的治疗剂类别。在当前用于产生人类抗体的技术当中,整体或部分地用编码人类抗体的遗传物质进行工程化的基因工程动物(例如啮齿动物)的发展已经彻底改革了用于治疗各种疾病的人类治疗性单克隆抗体领域。然而,仍然需要发展改良的用于产生人类单克隆抗体的体内系统,所述系统使人类抗体谱系在宿主基因工程动物中最大化。
技术实现要素:
6.本公开提供了基因修饰啮齿动物。在一些实施方案中,如所提供的基因修饰啮齿动物是大鼠或小鼠。在一些实施方案中,所有内源序列是大鼠或小鼠序列。举例来说,在一些实施方案中,基因修饰啮齿动物是大鼠且所有内源序列是大鼠序列。在一些实施方案中,基因修饰啮齿动物是小鼠且所有内源序列是小鼠序列。
7.在一些实施方案中,本公开提供了本文提供的基因修饰啮齿动物的繁殖群,所述繁殖群包含第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物,其中所述第一、第二和第三基因修饰啮齿动物各自是如本文所述的基因修饰啮齿动物。在一些实施方案中,第三基因修饰啮齿动物是第一基因修饰啮齿动物和第二基因修饰啮齿动物的后代。
8.如所提供的基因修饰啮齿动物具有包含有限人类λ轻链可变区谱系的种系基因组。在一些实施方案中,有限人类λ轻链可变区谱系可包含一个或两个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段。在一些实施方案中,有限人类λ轻链可变区谱系可包含两个未重排人类vλ基因区段和四个未重排人类jλ基因区段。在一些实施方案中,有限人类λ轻链可变区谱系可包含两个未重排人类vλ基因区段和五个未重排人类jλ基因区段。在一些实施方案中,有限人类λ轻链可变区谱系可包含单一重排人类免疫球蛋白λ轻链可变区,包括包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区。
9.在一些实施方案中,由如所提供的基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括由有限人类λ轻链可变区谱系表达的人类免疫球蛋白λ轻链可变结构域。在一些实施方案中,由如所提供的基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球
蛋白λ轻链可变结构域。在一些实施方案中,由本文提供的基因修饰啮齿动物的b细胞表达的所有免疫球蛋白轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。在一些实施方案中,由基因修饰啮齿动物的b细胞表达的所有重链均包括人类免疫球蛋白重链可变结构域和啮齿动物免疫球蛋白重链恒定结构域。
10.在一些实施方案中,如所提供的基因修饰啮齿动物包括包含有限人类λ轻链可变区谱系的工程化内源啮齿动物免疫球蛋白轻链基因座。在一些实施方案中,如所提供的基因修饰啮齿动物包括包含有限人类λ轻链可变区谱系的工程化内源啮齿动物免疫球蛋白λ轻链基因座。在一些实施方案中,如所提供的基因修饰啮齿动物包括包含有限人类λ轻链可变区谱系的工程化内源啮齿动物免疫球蛋白κ轻链基因座。在一些实施方案中,基因修饰啮齿动物的种系基因组对于工程化内源免疫球蛋白轻链基因座(例如工程化内源免疫球蛋白λ或κ轻链基因座)是纯合的。在一些实施方案中,基因修饰啮齿动物的种系基因组对于工程化内源免疫球蛋白轻链基因座(例如工程化内源免疫球蛋白λ或κ轻链基因座)是杂合的。
11.在一些实施方案中,基因修饰啮齿动物的种系基因组包括包含两个等位基因的工程化内源免疫球蛋白κ基因座。在一些实施方案中,第一等位基因包含有限人类λ轻链可变区谱系且第二等位基因包含有限人类κ轻链可变区谱系。在一些实施方案中,如本文所述的基因修饰啮齿动物的种系基因组和因此啮齿动物细胞和啮齿动物组织包括包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区的第一工程化内源免疫球蛋白κ轻链基因座等位基因,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。在一些实施方案中,如本文所述的基因修饰啮齿动物和因此啮齿动物细胞或啮齿动物组织包括包含可操作地连接至啮齿动物cκ基因区段的单一重排人类免疫球蛋白κ轻链可变区的第二工程化内源免疫球蛋白κ轻链基因座等位基因,其中单一重排人类免疫球蛋白κ轻链可变区包含人类vκ基因区段和人类jκ基因区段。在一些实施方案中,这种非人类动物或非人类组织可以由第一工程化内源免疫球蛋白κ轻链基因座等位基因表达λ轻链和由第二工程化内源免疫球蛋白κ轻链基因座等位基因表达κ轻链。在一些实施方案中,单一重排人类免疫球蛋白κ轻链可变区包含vκ3
‑
20或vκ1
‑
39且单一重排人类免疫球蛋白λ轻链可变区包含vλ1
‑
51或vλ2
‑
14。在一个实施方案中,单一重排人类免疫球蛋白κ轻链可变区为vκ3
‑
20/jκ1,且单一重排人类免疫球蛋白λ轻链可变区为vλ1
‑
51/jλ2或vλ2
‑
14/jλ2。
12.在一些实施方案中,如所提供的基因修饰啮齿动物包含可操作地连接至轻链恒定区基因区段的有限人类λ轻链可变区谱系。在一些实施方案中,如所提供的基因修饰啮齿动物包含可操作地连接至cκ基因区段的有限人类λ轻链可变区谱系。在一些实施方案中,如所提供的基因修饰啮齿动物包含可操作地连接至cλ基因区段的有限人类λ轻链可变区谱系。
13.在一些实施方案中,如所提供的基因修饰啮齿动物包括包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区的工程化内源免疫球蛋白κ轻链基因座,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。
14.在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ
1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
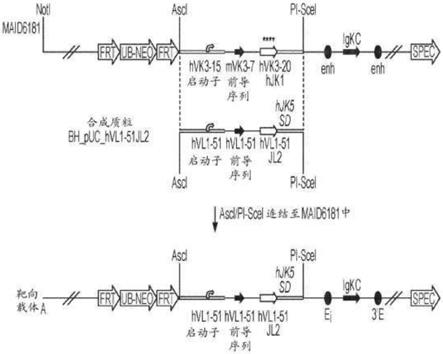
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类vλ基因区段是vλ1
‑
51或vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,人类jλ基因区段是jλ2。
15.在一些实施方案中,如所提供的基因修饰啮齿动物在工程化内源免疫球蛋白κ轻链基因座处缺乏啮齿动物cκ基因。
16.在一些实施方案中,如所提供的基因修饰啮齿动物具有包含工程化内源免疫球蛋白重链基因座的种系基因组。在一些实施方案中,工程化内源免疫球蛋白重链基因座包含一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。在一些实施方案中,一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段可操作地连接至一个或多个啮齿动物免疫球蛋白重链恒定区基因。
17.在一些实施方案中,如所提供的基因修饰啮齿动物具有包含工程化内源免疫球蛋白重链基因座的种系基因组,所述工程化内源免疫球蛋白重链基因座包含可操作地连接至一个或多个啮齿动物免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。在一些实施方案中,如所提供的基因修饰啮齿动物具有对于工程化内源免疫球蛋白重链基因座是纯合的种系基因组。
18.在一些实施方案中,如所提供的基因修饰啮齿动物具有包含一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段代替一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段、一个或多个内源j
h
基因区段或它们的组合的种系基因组。在一些实施方案中,如所提供的基因修饰啮齿动物具有包含分别替代一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段和一个或多个内源j
h
基因区段的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段的种系基因组。
19.在一些实施方案中,一个或多个未重排人类v
h
基因区段包含v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合。在一些实施方案中,一个或多个未重排人类d
h
基因区段包含d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合。在一些实施方案中,一个或多个未重排人类j
h
基因区段包含j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合。
20.在一些实施方案中,(i)一个或多个未重排人类v
h
基因区段包含v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合,(ii)一个或多个未重排人类d
h
基因区段包含d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合,以及(iii)一个或多个未重排人类j
h
基因区段包含j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合。
21.在一些实施方案中,如所提供的基因修饰啮齿动物具有包含一个或多个啮齿动物免疫球蛋白重链恒定区基因的种系基因组。在一些实施方案中,一个或多个啮齿动物免疫球蛋白重链恒定区基因是一个或多个内源啮齿动物免疫球蛋白重链恒定区基因。
22.在一些实施方案中,如所提供的基因修饰啮齿动物具有包含缺乏功能性内源啮齿动物adam6基因的工程化内源免疫球蛋白重链基因座的种系基因组。在一些实施方案中,如所提供的基因修饰啮齿动物具有包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列的种系基因组。在一些实施方案中,如所提供的基因修饰啮齿动物表达一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段。在一些实施方案中,如所提供的基因修饰啮齿动物具有包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列的种系基因组,所述一个或多个核苷酸序列包括在与工程化内源免疫球蛋白重链基因座相同的染色体上。在一些实施方案中,如所提供的基因修饰啮齿动物具有包含工程化内源免疫球蛋白重链基因座的种系基因组,所述工程化内源免疫球蛋白重链基因座包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。在一些实施方案中,如所提供的基因修饰啮齿动物具有包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列代替人类adam6伪基因的种系基因组。在一些实施方案中,如所提供的基因修饰啮齿动物具有包含替代人类adam6伪基因的编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列的种系基因组。
23.在一些实施方案中,如所提供的基因修饰啮齿动物具有包含以下的种系基因组:包含第一和第二人类v
h
基因区段的一个或多个人类v
h
基因区段,和介于第一人类v
h
基因区段与第二人类v
h
基因区段之间的编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。在一些实施方案中,第一人类v
h
基因区段是v
h1‑
2且第二人类v
h
基因区段是v
h6‑
1。
24.在一些实施方案中,编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于人类v
h
基因区段与人类d
h
基因区段之间。
25.在一些实施方案中,啮齿动物cλ基因具有与小鼠cλ1至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。在一些实施方案中,啮齿动物cλ基因具有与小鼠cλ2至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
在一些实施方案中,啮齿动物cλ基因具有与小鼠cλ3基因至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
26.在一些实施方案中,啮齿动物cλ基因是或包含小鼠cλ基因。在一些实施方案中,啮齿动物cλ基因是或包含小鼠cλ1基因。
27.在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ1至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ2至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ3至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ4基因至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
28.在一些实施方案中,啮齿动物cλ基因是或包含大鼠cλ基因。
29.在一些实施方案中,单一重排人类免疫球蛋白λ轻链可变区代替一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。在一些实施方案中,单一重排人类免疫球蛋白λ轻链可变区替代一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。
30.在一些实施方案中,本文提供的基因修饰小鼠包含功能性内源免疫球蛋白λ轻链基因座。在一些实施方案中,本文提供的基因修饰小鼠包含失活内源免疫球蛋白λ轻链基因座。在一些实施方案中,通过缺失或颠倒内源免疫球蛋白λ轻链基因座的全部或一部分使内源免疫球蛋白λ轻链基因座失活。在一些实施方案中,内源vλ基因区段、内源jλ基因区段和内源cλ基因整体或部分缺失。
31.在一些实施方案中,本文提供的基因修饰小鼠不可检测地表达内源免疫球蛋白κ轻链可变结构域。
32.本公开提供了包含如本文所述的基因修饰的啮齿动物胚胎。在一些实施方案中,啮齿动物胚胎具有包含工程化内源免疫球蛋白κ轻链基因座的基因组,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。在一些实施方案中,啮齿动物胚胎的基因组对于工程化内源免疫球蛋白κ轻链基因座是纯合的。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,人类vλ基因区段是vλ1
‑
51或vλ2
‑
14。在一些实施方案中,人类jλ基因区段是jλ2。
33.在一些实施方案中,提供了一种啮齿动物胚胎,所述啮齿动物胚胎的基因组包含工程化内源免疫球蛋白重链基因座,所述工程化内源免疫球蛋白重链基因座包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。在一些实施方案中,啮齿动物胚胎的基因组对于工程化内源免疫球蛋白重链基因座是纯合的。
34.本公开提供了本文所述的基因修饰啮齿动物的b细胞。在一些实施方案中,本文所述的基因修饰啮齿动物的b细胞包含工程化内源κ轻链基因座的单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
35.在一些实施方案中,本公开提供了如本文所述的基因修饰啮齿动物的b细胞,其包含工程化内源κ轻链基因座的单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。在一些实施方案中,如本文所述的基因修饰啮齿动物的b细胞包含重排人类免疫球蛋白重链可变区,所述重排人类免疫球蛋白重链可变区源自于工程化内源重链基因座中一个或多个未重排人类v
h
基因区段的人类v
h
基因区段、一个或多个未重排人类d
h
基因区段的人类d
h
基因区段和一个或多个未重排人类j
h
基因区段的人类j
h
基因区段。
36.本公开提供了由如本文所述的基因修饰啮齿动物的b细胞产生的杂交瘤。
37.本公开提供了来自单个如本文所述的基因修饰啮齿动物的b细胞群体。在一些实施方案中,由b细胞群体表达的所有抗体均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。在一些实施方案中,由b细胞群体表达的抗体包括从至少两个不同的重排人类免疫球蛋白重链可变区或其体细胞超突变型式表达的多个人类免疫球蛋白重链可变结构域。
38.本公开提供了包含如本文所述的基因修饰的干细胞,例如胚胎干(es)细胞。在一些实施方案中,干细胞(例如es细胞)包括包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区的工程化内源免疫球蛋白κ轻链基因座。在一些实施方案中,干细胞(例如es细胞)的基因组对于工程化内源免疫球蛋白κ轻链基因座是纯合的。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,人类vλ基因区段是vλ1
‑
51或vλ2
‑
14。在一些实施方案中,人类jλ基因区段是jλ2。
39.在一些实施方案中,干细胞(例如es细胞)包括包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段的工程化内源免疫球蛋白重链基因座。
40.本公开提供了一种表达抗体的哺乳动物细胞。在一些实施方案中,由哺乳动物细胞表达的抗体包括包含人类免疫球蛋白重链可变结构域的重链和包含人类免疫球蛋白λ轻链可变结构域的轻链,其中人类免疫球蛋白重链可变结构域、人类免疫球蛋白λ轻链可变结构域或两者是从本文所述的基因修饰啮齿动物鉴定、分离或与其同一。
41.在一些实施方案中,通过如下方法制备抗体,所述方法包括以下步骤:(a)将本文所述的基因修饰啮齿动物暴露于所关注抗原;(b)维持基因修饰啮齿动物处于足以使基因修饰啮齿动物对所关注抗原产生免疫反应的条件下;以及(c)从基因修饰啮齿动物回收:(i)结合所关注抗原的抗体,(ii)编码结合所关注抗原的抗体的人类轻链可变结构域、人类重链可变结构域、轻链或重链的核苷酸,或(iii)表达结合所关注抗原的抗体的细胞,其中
(c)的抗体包括人类重链可变结构域和人类λ轻链可变结构域。在一些实施方案中,抗体为双特异性抗体。
42.在一些实施方案中,制造抗体的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于抗原;(b)允许基因修饰啮齿动物对抗原显现免疫反应;以及(c)从基因修饰啮齿动物分离对所述抗原具特异性的抗体、表达对所述抗原具特异性的抗体的b细胞或编码对所述抗原具特异性的抗体的一个或多个核苷酸序列。在一些实施方案中,抗体为双特异性抗体。
43.在一些实施方案中,制造抗体的方法包括以下步骤:(a)在哺乳动物细胞中表达包含两条人类免疫球蛋白λ轻链和两条人类免疫球蛋白重链的所述抗体,其中每条人类免疫球蛋白λ轻链包括人类免疫球蛋白λ轻链可变结构域且每条人类免疫球蛋白重链包括人类免疫球蛋白重链可变结构域,其中从如本文所述的基因修饰啮齿动物鉴定或分离人类免疫球蛋白重链可变结构域中的至少一个、λ轻链可变结构域中的至少一个或它们的组合的氨基酸序列;以及(b)获得抗体。在一些实施方案中,抗体为双特异性抗体。
44.在一些实施方案中,提供了一种制造双特异性抗体的方法,所述方法包括:(a)使第一如本文所述的基因修饰啮齿动物与第一抗原的第一表位接触;(b)使第二如本文所述的基因修饰啮齿动物与第二抗原的第二表位接触;(c)从第一基因修饰啮齿动物分离表达对第一抗原的第一表位具特异性的第一抗体的b细胞并确定第一抗体的第一人类免疫球蛋白重链可变结构域;(d)从第二基因修饰啮齿动物分离表达对第二抗原的第二表位具特异性的第二抗体的b细胞并确定第二抗体的第二人类免疫球蛋白重链可变结构域;(e)将编码第一人类免疫球蛋白重链可变结构域的核苷酸序列可操作地连接至编码第一人类免疫球蛋白恒定结构域的核苷酸序列以产生编码第一人类重链的第一核苷酸序列;(f)将编码第二人类免疫球蛋白重链可变结构域的核苷酸序列可操作地连接至编码第二人类免疫球蛋白恒定结构域的核苷酸序列以产生编码第二人类重链的第二核苷酸序列;(g)在哺乳动物细胞中表达:(i)第一核苷酸序列;(ii)第二核苷酸序列;和(iii)第三核苷酸序列,其包含可操作地连接至人类免疫球蛋白λ轻链恒定区的单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
45.在一些实施方案中,本文提供了一种制造双特异性抗体的方法,所述方法包括(a)在哺乳动物细胞中表达:(i)第一核苷酸序列,其包含可操作地连接至第一人类免疫球蛋白恒定区的第一人类免疫球蛋白重链可变区;(ii)第二核苷酸序列,其包含可操作地连接至第二人类免疫球蛋白恒定区的第二人类免疫球蛋白重链可变区;和(iii)第三核苷酸序列,其包含可操作地连接至人类免疫球蛋白λ轻链恒定区的人类免疫球蛋白λ轻链可变区。在一些实施方案中,第一人类免疫球蛋白重链可变区编码从已经用第一抗原的第一表位免疫接种的第一如本文所述的基因修饰啮齿动物中的第一抗体鉴定、从其分离或与其同一的第一人类重链可变结构域,其中第一抗体特异性结合第一抗原的第一表位。在一些实施方案中,第二人类免疫球蛋白重链可变区编码从已经用第二抗原的第二表位免疫接种的第二如本文所述的基因修饰啮齿动物中的第二抗体鉴定、从其分离或与其同一的第二人类重链可变结构域,其中第二抗体特异性结合第二抗原的第二表位。在一些实施方案中,第三核苷酸的人类免疫球蛋白λ轻链可变区是单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
46.在一些实施方案中,第一基因修饰啮齿动物和第二基因修饰啮齿动物是相同基因修饰啮齿动物。在一些实施方案中,第一基因修饰啮齿动物和第二基因修饰啮齿动物是不同基因修饰啮齿动物。
47.在一些实施方案中,第一抗原和第二抗原是相同抗原,且第一表位和第二表位是不同表位。在一些实施方案中,第一抗原和第二抗原是不同抗原。
48.在一些实施方案中,制造人类免疫球蛋白重链的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;(b)获得特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列;以及(c)将人类免疫球蛋白重链可变结构域序列可操作地连接至人类免疫球蛋白重链恒定结构域序列以形成人类免疫球蛋白重链。在一些实施方案中,提供通过本段落的方法产生的人类免疫球蛋白重链。
49.在一些实施方案中,制备人类免疫球蛋白重链可变结构域的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;和(b)获得特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列。在一些实施方案中,提供通过本段落的方法产生的人类免疫球蛋白重链可变结构域。
50.在一些实施方案中,制造一系列人类免疫球蛋白重链可变结构域的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原,和(b)从基因修饰啮齿动物分离所述系列人类免疫球蛋白重链可变结构域。在一些实施方案中,一系列人类免疫球蛋白重链可变结构域各自结合于从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域,其中人类λ轻链可变结构域与所述系列中的人类免疫球蛋白重链可变结构域中的任一个配对,结合抗原。
51.在一些实施方案中,制备人类免疫球蛋白λ轻链的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;(b)获得特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列;以及(c)将人类免疫球蛋白λ轻链可变结构域序列可操作地连接至人类免疫球蛋白λ轻链恒定结构域序列以形成人类免疫球蛋白λ轻链。在一些实施方案中,提供通过本段落的方法产生的人类免疫球蛋白λ轻链。
52.在一些实施方案中,用于产生人类免疫球蛋白λ轻链可变结构域的方法包括以下步骤:(a)将本文所述的基因修饰啮齿动物暴露于所关注抗原;和(b)获得特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列。在一些实施方案中,提供通过本段落的方法产生的人类免疫球蛋白λ轻链可变结构域。
53.在一些实施方案中,制造编码人类免疫球蛋白重链的核苷酸序列的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;(b)获得编码特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列的人类免疫球蛋白重链可变区;以及(c)将人类免疫球蛋白重链可变区可操作地连接至人类免疫球蛋白重链恒定区序列以形成编码人类免疫球蛋白重链的核苷酸序列。在一些实施方案中,提供通过本段落的方法产生的编码人类免疫球蛋白重链的核苷酸序列。
54.在一些实施方案中,制造包含人类免疫球蛋白重链可变区的核苷酸序列的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;和(b)获得编码特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列的人类免疫球蛋白重链可变区。在一些实施方案中,提供通过本段落的方法产生的包含人
类免疫球蛋白重链可变区的核苷酸序列。
55.在一些实施方案中,制造编码人类免疫球蛋白λ轻链的核苷酸序列的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;(b)获得编码特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列的人类免疫球蛋白λ轻链可变区;和(c)将人类免疫球蛋白λ轻链可变区可操作地连接至人类免疫球蛋白λ轻链恒定区序列以形成编码人类免疫球蛋白λ轻链的核苷酸序列。在一些实施方案中,提供通过本段落的方法产生的编码人类免疫球蛋白λ轻链的核苷酸序列。
56.在一些实施方案中,制造包含人类免疫球蛋白λ轻链可变区的核苷酸序列的方法包括以下步骤:(a)将如本文所述的基因修饰啮齿动物暴露于所关注抗原;和(b)获得编码特异性结合抗原并由基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列的人类免疫球蛋白λ轻链可变区。在一些实施方案中,提供通过本段落的方法产生的包含人类免疫球蛋白λ轻链可变区的核苷酸序列。
57.在一些实施方案中,提供了一种靶向载体。在一些实施方案中,靶向载体包含(i)5'同源臂,所述5'同源臂包含与内源啮齿动物κ轻链基因座中的5'标靶序列相对应的核苷酸序列;(ii)单一重排人类免疫球蛋白λ轻链可变区,所述单一重排人类免疫球蛋白λ轻链可变区包含vλ基因区段和jλ基因区段;(iii)啮齿动物cλ基因区段;和(iv)3'同源臂,所述3'同源臂包含与内源啮齿动物κ轻链基因座中的3'标靶序列相对应的核苷酸序列。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类vλ基因区段是vλ1
‑
51或vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,人类jλ基因区段是jλ2。
58.在一些实施方案中,制造本文所述的基因修饰啮齿动物的方法包括以下步骤:(a)将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中;和(b)使用步骤(a)中产生的啮齿动物es细胞产生啮齿动物。
59.在一些实施方案中,制备本文所述的基因修饰啮齿动物的方法包括以下步骤:(a)将可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段;和(b)使用步骤(a)中产生的啮齿动物es细胞产生啮齿动物。在一些实施方案中,啮齿动物es细胞的基因组包含在工程化内源免疫球蛋白重链基因座处可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多
个未重排人类j
h
基因区段。在一些实施方案中,啮齿动物es细胞的基因组还包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
60.在一些实施方案中,制造基因修饰啮齿动物es细胞的方法包括以下步骤:将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中。在一些实施方案中,制造基因修饰啮齿动物es细胞的方法包括以下步骤:将可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。在一些实施方案中,啮齿动物es细的基因组胞包含在工程化内源免疫球蛋白重链基因座处可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。在一些实施方案中,啮齿动物es细胞的基因组还包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
61.在一些实施方案中,制造如本文所述的基因修饰啮齿动物的方法包括以下步骤:(a)对啮齿动物的种系基因组中的内源免疫球蛋白κ轻链基因座进行工程化以包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段,使得由基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。在一些实施方案中,一种方法还包括以下步骤:(b)对啮齿动物的种系基因组中的内源免疫球蛋白重链基因座进行工程化以包含可操作地连接至一个或多个啮齿动物免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段,使得由基因修饰啮齿动物的b细胞表达的所有重链均包括人类免疫球蛋白重链可变结构域和啮齿动物免疫球蛋白重链恒定结构域。在一些实施方案中,步骤(b)包括对内源免疫球蛋白重链基因座进行工程化以还包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。在一些实施方案中,步骤(a)和/或步骤(b)是在啮齿动物es细胞中进行。
62.在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类vλ基因区段是vλ1
‑
51或vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,人类jλ基因区段是jλ2。
63.在一些实施方案中,内源免疫球蛋白κ轻链基因座缺乏啮齿动物cκ基因。
64.在一些实施方案中,一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段代替一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段、一个或多个内源j
h
基因区段或它们的组合。在一些实施方案中,一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段分别替代一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段和一个或多个内源j
h
基因区段。
65.在一些实施方案中,一个或多个未重排人类v
h
基因区段包含v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合。在一些实施方案中,一个或多个未重排人类d
h
基因区段包含d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合。在一些实施方案中,一个或多个未重排人类j
h
基因区段包含j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合。
66.在一些实施方案中,一个或多个啮齿动物免疫球蛋白重链恒定区基因是一个或多个内源啮齿动物免疫球蛋白重链恒定区基因。
67.在一些实施方案中,内源免疫球蛋白重链基因座缺乏功能性内源啮齿动物adam6基因。在一些实施方案中,基因修饰啮齿动物的种系基因组包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。在一些实施方案中,一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段由基因修饰啮齿动物表达。在一些实施方案中,编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列包括在与工程化内源免疫球蛋白重链基因座相同的染色体上。在一些实施方案中,工程化内源免疫球蛋白重链基因座包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。在一些实施方案中,编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列代替人类adam6伪基因。在一些实施方案中,编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列替代人类adam6伪基因。
68.在一些实施方案中,一个或多个人类v
h
基因区段包含第一和第二人类v
h
基因区段,且编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于第一人类v
h
基因区段与第二人类v
h
基因区段之间。在一些实施方案中,第一人类v
h
基因区段是v
h1‑
2且第二人类v
h
基因区段是v
h6‑
1。
69.在一些实施方案中,编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于人类v
h
基因区段与人类d
h
基因区段之间。
70.在一些实施方案中,啮齿动物cλ基因具有与小鼠cλ1至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
71.在一些实施方案中,啮齿动物cλ基因具有与小鼠cλ2至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
72.在一些实施方案中,啮齿动物cλ基因具有与小鼠cλ3基因至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
73.在一些实施方案中,啮齿动物cλ基因是或包含小鼠cλ基因。
74.在一些实施方案中,啮齿动物cλ基因是或包含小鼠cλ1基因。
75.在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ1至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
76.在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ2至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
77.在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ3至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
78.在一些实施方案中,啮齿动物cλ基因具有与大鼠cλ4基因至少80%、至少85%、至少90%、至少95%、至少98%或至少99%同一的序列。
79.在一些实施方案中,啮齿动物cλ基因是或包含大鼠cλ基因。
80.在一些实施方案中,单一重排人类免疫球蛋白λ轻链可变区代替一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。在一些实施方案中,单一重排人类免疫球蛋白λ轻链可变区替代一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。在一些实施方案中,本文所述的基因修饰啮齿动物的种系基因组还包含失活内源免疫球蛋白λ轻链基因座。在一些实施方案中,通过缺失或颠倒内源免疫球蛋白λ轻链基因座的全部或一部分使内源免疫球蛋白λ轻链基因座失活。在一些实施方案中,内源vλ基因区段、内源jλ基因区段和内源cλ基因整体或部分缺失。
附图说明
81.由以下图构成的本文中所包括的图式仅仅是出于说明的目的而非限制性的。
82.图1a和图1b示出了构建用于产生根据本公开的非人类动物的一实施方案的靶向载体(实施例1和4中描述)的策略的一例示性实施方案(不按比例)的图。除非图中的标记另外表示(例如关于选择盒、loxp位点等),否则实心形状和单线代表小鼠序列,且空心形状和双线代表人类序列。“frt”指示frt重组酶系统位点;“ub
‑
neo”指示具有ub启动子的新霉素(neomycin)抗性基因;“enh”指示增强子;“ei”指示内含子增强子;“3'e”指示3'增强子;“spec”指示壮观霉素(spectinomycin)抗性基因;“sd”指示剪接供体位点;“cds”指示编码序列;“lox”指示lox2372位点;“loxp”指示lox p位点;和“ub
‑
hyg”指示具有ub启动子的潮霉素(hygromycin)抗性基因。
83.图2a和图2b示出了构建用于产生根据本公开的非人类动物的一实施方案的靶向载体(实施例7和10中描述)的策略的一例示性实施方案(不按比例)的图。除非图中的标记另外表示(例如关于选择盒、loxp位点等),否则实心形状和单线代表小鼠序列,且空心形状和双线代表人类序列。“frt”指示frt重组酶系统位点;“ub
‑
neo”指示具有ub启动子的新霉素抗性基因;“enh”指示增强子;“ei”指示内含子增强子;“3'e”指示3'增强子;“spec”指示壮观霉素抗性基因;“sd”指示剪接供体位点;“cds”指示编码序列;“lox”指示lox2372位点;
“
loxp”指示lox p位点;和“ub
‑
hyg”指示具有ub启动子的潮霉素抗性基因。
84.图3示出了靶向载体a(如实施例1中所述)插入啮齿动物胚胎干(es)细胞克隆的工程化igκ轻链基因座(如实施例2中所述)中的图(不按比例),所述es细胞克隆用于产生根据本公开的啮齿动物的一实施方案。图中包括用于证实胚胎干(es)细胞克隆对某些示例性序列呈阳性的各种探针(通过圆形破折号指示)的大致位置。除非图中的标记另外表示(例如关于选择盒、loxp位点等),否则实心形状和单线代表小鼠序列,且空心形状和双线代表人类序列。“frt”指示frt重组酶系统位点;“ub
‑
neo”指示具有ub启动子的新霉素抗性基因;“ei”指示内含子增强子;“3'e”指示3'增强子;和“sd”指示剪接供体位点。“het”指示杂合小鼠,“ho”指示纯合小鼠。
85.图4示出了靶向载体b(如实施例4中所述)插入啮齿动物胚胎干(es)细胞克隆的工程化igκ轻链基因座(如实施例5中所述)中的图(不按比例),所述es细胞克隆用于产生根据本公开的啮齿动物的一实施方案。图中包括用于证实胚胎干(es)细胞克隆对某些示例性序列呈阳性的各种探针(通过圆形破折号指示)的大致位置。除非图中的标记另外表示(例如关于选择盒、loxp位点等),否则实心形状和单线代表小鼠序列,且空心形状和双线代表人类序列。“frt”指示frt重组酶系统位点;“ub
‑
neo”指示具有ub启动子的新霉素抗性基因;“ei”指示内含子增强子;“3'e”指示3'增强子;“sd”指示剪接供体位点;“loxp”指示lox p位点;和“ub
‑
hyg”指示具有ub启动子的潮霉素抗性基因。“het”指示杂合小鼠,“ho”指示纯合小鼠。
86.图5示出了靶向载体c(如实施例7中所述)插入啮齿动物胚胎干(es)细胞克隆的工程化igκ轻链基因座(如实施例8中所述)中的图(不按比例),所述es细胞克隆用于产生根据本公开的啮齿动物的一实施方案。图中包括用于证实胚胎干(es)细胞克隆对某些示例性序列呈阳性的各种探针(通过圆形破折号指示)的大致位置。除非图中的标记另外表示(例如关于选择盒、loxp位点等),否则实心形状和单线代表小鼠序列,且空心形状和双线代表人类序列。“frt”指示frt重组酶系统位点;“ub
‑
neo”指示具有ub启动子的新霉素抗性基因;“ei”指示内含子增强子;“3'e”指示3'增强子;和“sd”指示剪接供体位点。“het”指示杂合小鼠,“ho”指示纯合小鼠。
87.图6示出了靶向载体d(如实施例10中所述)插入啮齿动物胚胎干(es)细胞克隆的工程化igκ轻链基因座(如实施例11中所述)中的图(不按比例),所述es细胞克隆用于产生根据本公开的啮齿动物的一实施方案。图中包括用于证实胚胎干(es)细胞克隆对某些示例性序列呈阳性的各种探针(通过圆形破折号指示)的大致位置。除非图中的标记另外表示(例如关于选择盒、loxp位点等),否则实心形状和单线代表小鼠序列,且空心形状和双线代表人类序列。“frt”指示frt重组酶系统位点;“ub
‑
neo”指示具有ub启动子的新霉素抗性基因;“ei”指示内含子增强子;“3'e”指示3'增强子;“sd”指示剪接供体位点;“loxp”指示lox p位点;和“ub
‑
hyg”指示具有ub启动子的潮霉素抗性基因。“het”指示杂合小鼠,“ho”指示纯合小鼠。
88.图7示出了小鼠cκ的核苷酸序列(seq id no:25)。编码序列由粗体字母表示,且3'非翻译区未加粗。
89.图8示出了大鼠cκ的核苷酸序列(seq id no:27)。编码序列由粗体字母表示,且3'非翻译区未加粗。
90.图9a
‑
9c示出了工程化vλ1
‑
51/jλ2载体(seq id no:31)的核苷酸序列。限制酶位点序列
‑
asci(序列5'末端)和pi
‑
scei(序列3'末端)以大写斜体字母表示;vλ1
‑
51启动子序列由小写字母(非粗体)表示;vλ1 51 5'utr序列由大写、斜体且粗体字母表示;重排的vλ1
‑
51/jλ2序列,其包括:vλ1
‑
51外显子1序列由大写字母(非粗体)表示,其中vλ1
‑
51外显子1起始密码子进一步用斜体表示且加下划线;vλ1
‑
51内含子1序列由小写粗体字母表示;vλ1
‑
51外显子2序列由大写、粗体且加下划线(实下划线)字母表示,且jλ2序列由大写、粗体且加下划线(虚下划线))字母表示;且人类jκ5
‑
cκ内含子序列由小写斜体字母表示。
91.图10示出了包括vλ1
‑
51内含子的重排的vλ1
‑
51/jλ2可变区的核苷酸序列(seq id no:32)。vλ1
‑
51外显子1序列由大写字母表示,其中vλ1
‑
51外显子1起始密码子进一步用斜体表示且加下划线;vλ1
‑
51内含子1序列由小写粗体字母表示;vλ1
‑
51外显子2序列由大写粗体且加下划线(实下划线)字母表示;且jλ2序列由大写、粗体且加下划线(虚下划线)字母表示。
92.图11示出了无vλ1
‑
51内含子的重排的vλ1
‑
51/jλ2可变区的核苷酸序列(seq id no:33)。vλ1
‑
51编码序列由大写字母表示,其中vλ1
‑
51起始密码子进一步用斜体表示且加下划线;且jλ2序列由大写字母和虚下划线表示。
93.图12示出了包括信号序列的重排的vλ1
‑
51/jλ2可变区的核苷酸序列(seq id no:34)。斜体粗体字指示信号肽的序列。
94.图13a
‑
13c示出了工程化vλ2
‑
14/jλ2载体的核苷酸序列(seq id no:36)。限制酶位点序列
‑
asci(序列5'末端)和pi
‑
scei(序列3'末端)以大写斜体字母表示;vλ1
‑
51启动子序列由小写字母(非粗体)表示;vλ1 51 5'utr序列由大写、斜体且粗体字母表示;重排的vλ2
‑
14/jλ2序列,其包括:vλ2
‑
14外显子1序列由大写字母(非粗体)表示,其中vλ2
‑
14外显子1起始密码子进一步用斜体表示且加下划线;vλ2
‑
14内含子1序列由小写粗体字母表示;vλ2
‑
14、外显子2序列由大写、粗体且加下划线(实下划线)字母表示,且jλ2序列由大写、粗体且加下划线(虚下划线))字母表示;且人类jκ5
‑
cκ内含子序列由小写斜体字母表示。
95.图14示出了包括vλ2
‑
14内含子的重排的vλ2
‑
14/jλ2可变区的核苷酸序列(seq id no:37)。vλ2
‑
14外显子1序列由大写字母表示,其中vλ2
‑
14外显子1起始密码子进一步用斜体表示且加下划线;vλ2
‑
14内含子1序列由小写、粗体字母表示;vλ1
‑
51外显子2序列由大写、粗体且加下划线(实下划线)字母表示;且jλ2序列由大写、粗体且加下划线(虚下划线)字母表示。
96.图15示出了无vλ2
‑
14内含子的重排的vλ2
‑
14/jλ2可变区的核苷酸序列(seq id no:38)。vλ2
‑
14序列由大写字母表示,其中vλ2
‑
14起始密码子进一步用斜体表示且加下划线;且jλ2序列由大写字母和虚下划线表示。
97.图16示出了包括信号肽的重排的vλ2
‑
14/jλ2可变结构域的氨基酸序列(seq id no:39)。斜体粗体字指示信号肽的序列。
98.序列表中所选序列的简要描述
99.在如本文所述的非人类动物的一些实施方案中可利用的各种人类vλ和jλ基因区段的代表性核苷酸和氨基酸序列可得自the international immunogenetics information system网站、www.imgt.org或lefranc,m
‑
p.,the immunoglobulin factsbook,academic press,2001年5月23日(本文称为“lefranc 2001”)。
100.以下是在如本文所述的非人类动物的一些实施方案中可利用的各种小鼠、大鼠或人类λ恒定区或结构域的代表性核苷酸和氨基酸序列。
101.表5:某些小鼠cλ基因区段的核苷酸序列
[0102][0103][0104]
表6:某些小鼠cλ基因区段的氨基酸序列
[0105][0106]
表7:某些大鼠cλ基因区段的核苷酸序列
[0107]
[0108][0109]
表8:某些大鼠cλ基因区段的氨基酸序列
[0110][0111]
表9:某些人类cλ基因区段的核苷酸序列
[0112]
[0113][0114]
表10:某些人类cλ基因区段的氨基酸序列
[0115]
[0116][0117]
以下是在如本文所述的非人类动物的一些实施方案中可利用的小鼠、大鼠或人类κ恒定区或结构域的代表性核苷酸和氨基酸序列。
[0118]
小鼠cκ的核苷酸序列(seq id no:25)(如图7中再现):
[0119]
gggctgatgctgcaccaactgtatccatcttcccaccatccagtgagcagttaacatctggaggtgcctcagtcgtgtgcttcttgaacaacttctaccccaaagacatcaatgtcaagtggaagattgatggcagtgaacgacaaaatggcgtcctgaacagttggactgatcaggacagcaaagacagcacctacagcatgagcagcaccctcacgttgaccaaggacgagtatgaacgacataacagctatacctgtgaggccactcacaagacatcaacttcacccattgtcaagagcttcaacaggaatgagtgttagagacaaaggtcctgagacgccaccaccagctccccagctccatcctatcttcccttctaaggtcttggaggcttccccacaagcgacctaccactgttgcggtgctccaaacctcctccccacctccttctcctcctcctccctttccttggcttttatcatgctaatatttgcagaaaatattcaataaagtgagtctttgcacttga
[0120]
关于seq id no:25,以下适用:
[0121]
‑
编码序列由粗体字母表示;且
[0122]
‑
3'非翻译区未加粗。
[0123]
小鼠cκ的氨基酸序列(seq id no:26):
[0124]
adaaptvsifppsseqltsggasvvcflnnfypkdinvkwkidgserqngvlnswtdqdskdstysmsstltltkdeyerhnsytceathktstspivksfnrnec
[0125]
大鼠cκ的核苷酸序列(seq id no:27)(如图8中再现):
[0126]
gggctgatgctgcaccaactgtatctatcttcccaccatccacggaacagttagcaactggaggtgcctcagtcgtgtgcctcatgaacaacttctatcccagagacatcagtgtcaagtggaagattgatggcactgaacgacgagatggtgtcctggacagtgttactgatcaggacagcaaagacagcacgtacagcatgagcagcaccctctcgttgaccaaggctgactatgaaagtcataacctctatacctgtgaggttgttcataagacatcatcctcacccgtcgtcaagagcttcaacaggaatgagtgttagacccaaaggtcctgaggtgccacctgctccccagctccttccaatcttccctcctaaggtcttggagacttccccacaagcgacctaccactgttgcggtgctccaaacctcctccccacctcatcctccttcctttccttggctttgatcatgctaatatttggggaatattaaataaagtgaatctttgcacttga
[0127]
关于seq id no:27,以下适用:
[0128]
‑
编码序列由粗体字母表示;且
[0129]
‑
3'非翻译区未加粗。
[0130]
大鼠cκ的氨基酸序列(seq id no:28):
[0131]
adaaptvsifppsteqlatggasvvclmnnfyprdisvkwkidgterrdgvldsvtdqdskdstysmsstlsltkadyeshnlytcevvhktssspvvksfnrnec
[0132]
人类cκ的核苷酸序列(seq id no:29):
[0133]
gaactgtggctgcaccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt
[0134]
人类cκ的氨基酸序列(seq id no:30):
[0135]
tvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[0136]
工程化vλ1
‑
51/jλ2载体的核苷酸序列(seq id no:31)(如图1a中示意性示出和图9a
‑
9c中再现):
[0137]
[0138]
[0139][0140]
关于seq id no:31,以下适用:
[0141]
‑
限制酶位点序列
‑
asci(序列5'末端)和pi
‑
scei(序列3'末端)以大写斜体字母表示;
[0142]
‑
vλ1
‑
51启动子序列由小写字母(非粗体)表示;
[0143]
‑
vλ1
‑
51 5'utr序列由大写、斜体且粗体字母表示;
[0144]
‑
重排的vλ1
‑
51/jλ2序列,其包括:
[0145]
οvλ1
‑
51外显子1序列由大写字母(非粗体)表示,其中vλ1
‑
51外显子1起始密码子进一步用斜体表示且加下划线;
[0146]
οvλ1
‑
51内含子1序列由小写粗体字母表示;
[0147]
οvλ1
‑
51外显子2序列由大写、粗体且加下划线(实下划线)字母表示,且
[0148]
οjλ2序列由大写、粗体且加下划线(虚下划线)字母表示;且
[0149]
‑
人类jκ5
‑
cκ内含子序列由小写斜体字母表示。
[0150]
包括vλ1
‑
51内含子的重排的vλ1
‑
51/jλ2可变区的核苷酸序列(seq id no:32)(如图10中再现):
[0151][0152][0153]
关于seq id no:32,以下适用:
[0154]
‑
vλ1
‑
51外显子1序列由大写字母表示,其中vλ1
‑
51外显子1起始密码子进一步用斜体表示且加下划线;
[0155]
‑
vλ1
‑
51内含子1序列由小写粗体字母表示;
[0156]
‑
vλ1
‑
51外显子2序列由大写粗体且加下划线(实下划线)字母表示;且
[0157]
‑
jλ2序列由大写、粗体且加下划线(虚下划线)字母表示。
[0158]
无vλ1
‑
51内含子的重排的vλ1
‑
51/jλ2可变区的核苷酸序列(seqid no:33)(如图11中再现):
[0159][0160][0161]
关于seq id no:33,以下适用:
[0162]
‑
vλ1
‑
51编码序列由大写字母表示,其中vλ1
‑
51起始密码子进一步用斜体表示且加下划线;且
[0163]
‑
jλ2序列由大写字母和虚下划线表示。
[0164]
包括信号肽的重排的vλ1
‑
51/jλ2可变结构域的氨基酸序列(seq id no:34)(如图12中再现):
[0165][0166]
关于seq id no:34,斜体粗体字指示信号肽的序列。
[0167]
无信号肽的重排的vλ1
‑
51/jλ2可变结构域的氨基酸序列(seq idno:35):
[0168]
qsvltqppsvsaapgqkvtiscsgsssnignnyvswyqqlpgtapklliydnnkrpsgipdrfsgsksgtsatlgitglqtgdeadyycgtwdsslsavvfgggtkltvl
[0169]
工程化vλ2
‑
14/jλ2载体的核苷酸序列(seq id no:36)(如图2a中示意性示出和图13a
‑
13c中再现):
[0170]
[0171]
[0172][0173]
关于seq id no:36,以下适用:
[0174]
‑
限制酶位点序列
‑
asci(序列5'末端)和pi
‑
scei(序列3'末端)以大写斜体字母表示;
[0175]
‑
vλ1
‑
51启动子序列由小写字母(非粗体)表示;
[0176]
‑
vλ1
‑
51 5'utr序列由大写、斜体且粗体字母表示;
[0177]
‑
重排的vλ2
‑
14/jλ2序列,其包括:
[0178]
οvλ2
‑
14外显子1序列由大写字母(非粗体)表示,其中vλ2
‑
14外显子1起始密码子进一步用斜体表示且加下划线;
[0179]
οvλ2
‑
14内含子1序列由小写粗体字母表示;
[0180]
οvλ2
‑
14外显子2序列由大写、粗体且加下划线(实下划线)字母表示,且
[0181]
οjλ2序列由大写、粗体且加下划线(虚下划线))字母表示;且
[0182]
ο人类jκ5
‑
cκ内含子序列由小写斜体字母表示。
[0183]
包括vλ2
‑
14内含子的重排的vλ2
‑
14/jλ2可变区的核苷酸序列(seq id no:37)(如图14中再现):
[0184]
关于seq id no:37,以下适用:
[0185]
‑
vλ2
‑
14外显子1序列由大写字母表示,其中vλ2
‑
14外显子1起始密码子进一步用斜体表示且加下划线;
[0186]
‑
vλ2
‑
14内含子1序列由小写粗体字母表示;
[0187]
‑
vλ1
‑
51外显子2序列由大写、粗体且加下划线(实下划线)字母表示;且
[0188]
‑
jλ2序列由大写、粗体且加下划线(虚下划线)字母表示。
[0189]
无vλ2
‑
14内含子的重排的vλ2
‑
14/jλ2可变区的核苷酸序列(seq id no:38)(如图15中再现):
[0190][0191][0192]
关于seq id no:38,以下适用:
[0193]
‑
vλ2
‑
14序列由大写字母表示,其中vλ2
‑
14起始密码子进一步用斜体表示且加下划线;且
[0194]
‑
jλ2序列由大写字母和虚下划线表示。
[0195]
包括信号肽的重排的vλ2
‑
14/jλ2可变结构域的氨基酸序列(seq id no:39)(如图16中再现):
[0196][0197]
关于seq id no:39,斜体粗体字指示信号肽的序列。
[0198]
无信号肽的重排的vλ2
‑
14/jλ2可变结构域的氨基酸序列(seq id no:40):
[0199]
qsaltqpasvsgspgqsitisctgtssdvggynyvswyqqhpgkapklmiyevsnrpsgvsnrfsgsksgntasltisglqaedeadyycssytssstlvvfgggtkltvl
[0200]
定义
[0201]
本发明的范围由随附权利要求书界定,且不受本文所述的某些实施方案限制。所属领域的技术人员阅读本说明书时,将清楚可等同于这类所描述的实施方案或在权利要求书范围内的各种修改。一般来说,除非另外清楚指示,否则本文中使用的术语是根据其在所属领域中所了解的含义。以下提供了某些术语的明确定义;所属领域的技术人员从上下文将清楚本说明书通篇中特定情况下这些和其它术语的含义。本说明书通篇阐述以下和其它术语的额外定义。本说明书内引用的专利和非专利文献的参考文献或它们的相关部分以引用的方式整体并入本文中。
[0202]
权利要求书中修饰权利要求要素的例如“第一”、“第二”、“第三”等顺序术语的使用本身不暗示一种权利要求要素优于另一要素的任何优先权、优先或顺序,或进行方法动作的暂时顺序,而仅仅用作区分具有某一名称的一种权利要求要素与具有相同名称(如果未使用顺序术语)的另一要素以区分所述权利要求要素。
[0203]
应了解除非相反地清楚指示,否则如本文所用的冠词“一种(a/an)”包括多个指示物。除非相反地指示或另外从上下文显而易见,否则如果超过一个或所有群组成员存在于、用于给定产物或方法中或以其它方式与给定产物或方法有关,那么在群组的一个或多个成员之间包括“或”的权利要求或描述视为符合。在一些实施方案中,群组正好一个成员存在于、用于给定产物或方法中或以其它方式与给定产物或方法有关。在一些实施方案中,超过一个或所有群组成员存在于、用于给定产物或方法中或以其它方式与给定产物或方法有关。应了解,除非另外指示,或除非所属领域的一般技术人员显而易见将出现矛盾或不一致,否则本发明涵盖一个或多个限制、要素、条款、描述性术语等从一个或多个所列出的权利要求引入依附于同一基础权利要求(或相关的任何其它权利要求)的另一权利要求中的所有变体、组合和变换。在要素呈清单呈现(例如马库什群组(markush group)或类似格式)的情况下,应了解还公开了所述要素的每个子组,且可以从所述群组去除任何要素。应了解,一般来说,在实施方案或方面称为“包含”具体要素、特征等的情况下,某些实施方案或方面“由”这种要素、特征等“组成”或“基本上由其组成”。为求简单,那些实施方案不会在每种情况下特定明确地阐述于本文中。还应了解,任何实施方案或方面均可明确地从权利要求书排除掉,不管本说明书中是否叙述所述特定排除。
[0204]
施用:如本文所用,包括向受试者或系统(例如向细胞、器官、组织、生物体或它们的相关组分或组分集合)施用组合物(例如抗原或抗体)。熟练技术人员将了解施用途径可
视例如组合物正施用的受试者或系统、组合物的性质、施用目的等变化。举例来说,在某些实施方案中,向动物受试者(例如向人类或啮齿动物)施用可经支气管(包括通过支气管滴注)、经颊、经肠、皮间(interdermal)、动脉内、皮内、胃内、髓内、肌肉内、鼻内、腹膜内、鞘内、静脉内、心室内、经粘膜、经鼻、经口、经直肠、皮下、舌下、表面、经气管(包括通过气管内滴注)、经皮、经阴道和/或经玻璃体。在一些实施方案中,施用可涉及间歇给药。在一些实施方案中,施用可涉及连续给药(例如灌注)至少所选时段。
[0205]
抗原结合蛋白:如本文所用,是指特异性结合于至少一种所关注抗原的任何蛋白质或多肽。抗原结合蛋白包括(但不限于)抗体、重链、轻链(例如λ或κ轻链)、重链可变结构域、轻链可变结构域(例如λ或κ轻链可变结构域)和单链可变片段(scfv)。在一些实施方案中,抗原结合蛋白可为多特异性的且特异性地结合于两种或更多种表位或抗原。
[0206]
大约:在应用于一个或多个感兴趣值时,包括与所述参考值类似的值。在某些实施方案中,除非另有说明或另外从上下文显而易见,否则术语“大约”或“约”是指在所述参考值
±
10%(大于或小于)内的值的范围(这种数值超过可能值的100%除外)。
[0207]
生物活性:如本文所用,是指在生物系统中、在体外或体内(例如在生物体中)具有活性的任何药剂的特征。举例来说,在存在于生物体中时在所述生物体内具有生物效应的药剂被视为是生物活性的。在特定实施方案中,在蛋白质或多肽为生物活性的情况下,共享蛋白质或多肽的至少一种生物活性的所述蛋白质或多肽的一部分通常称为“生物活性”部分。
[0208]
可比:如本文所用,是指两种或更多种药剂、实体、情形、条件集合等彼此可能不一致,但足够相似,从而允许之间进行比较,以便基于观察到的差异或相似性,可合理地得出结论。所属领域的一般技术人员将理解在上下文中在两种或更多种这种药剂、实体、情形、条件集合等的任何给定情况下何种同一性程度视为可比的。
[0209]
保守:如本文所用,是指当描述保守氨基酸取代时,包括氨基酸残基经侧链r基团具有类似化学特性(例如电荷或疏水性)的另一氨基酸残基取代的情况。一般来说,保守氨基酸取代基本上不会改变蛋白质的所关注功能性,例如受体结合于配体的能力。侧链具有类似化学特性的氨基酸的组的实例包括:脂族侧链,例如甘氨酸(gly、g)、丙氨酸(ala、a)、缬氨酸(val、v)、亮氨酸(leu、l)和异亮氨酸(ile、i);脂族
‑
羟基侧链,例如丝氨酸(ser、s)和苏氨酸(thr、t);含酰胺的侧链,例如天冬酰胺(asn、n)和谷氨酰胺(gln、q);芳族侧链,例如苯丙氨酸(phe、f)、酪氨酸(tyr、y)和色氨酸(trp、w);碱性侧链,例如赖氨酸(lys、k)、精氨酸(arg、r)和组氨酸(his、h);酸性侧链,例如天冬氨酸(asp、d)和谷氨酸(glu、e);和含硫侧链,例如半胱氨酸(cys、c)和甲硫氨酸(met、m)。保守氨基酸取代组包括例如缬氨酸/亮氨酸/异亮氨酸(val/leu/ile、v/l/i)、苯丙氨酸/酪氨酸(phe/tyr、f/y)、赖氨酸/精氨酸(lys/arg、k/r)、丙氨酸/缬氨酸(ala/val、a/v)、谷氨酸/天冬氨酸(glu/asp、e/d)和天冬酰胺/谷氨酰胺(asn/gln、n/q)。在一些实施方案中,保守氨基酸取代可以是蛋白质中的任何天然残基经丙氨酸取代,如例如丙氨酸扫描诱变中所使用。在一些实施方案中,进行在以下中公开的pam250对数似然矩阵中具有正值的保守性取代:gonnet,g.h.等人,1992,science 256:1443
‑
1445,其以引用的方式整体并入本文中。在一些实施方案中,取代是中度保守取代,其中所述取代在pam250对数似然矩阵中具有非负值。
[0210]
对照:如本文所用,是指所属领域中了解的“对照”的含义,作为结果进行比较的标
准。通常,通过分离变数以得出关于这种变数的结论,在实验中使用对照来加强完整性。在一些实施方案中,对照是与测试反应或测定同时进行以提供比较对象的反应或测定。“对照”还包括“对照动物”。“对照动物”可具有如本文所述的修饰,与本文所述不同的修饰或无修饰(即野生型动物)。在一个实验中,施加“测试”参数(例如测试的变数)。在第二实验中,不施加“对照”测试的变数。在一些实施方案中,对照是历史对照(即先前进行的测试或测定,或先前已知的量或结果)。在一些实施方案中,对照是或包含印刷的或以其它方式保存的记录。对照可以是阳性对照或阴性对照。
[0211]
破坏:如本文所用,是指与dna分子(例如与内源同源序列,例如基因或基因座)的同源重组事件的结果。在一些实施方案中,破坏可以实现或表示插入、缺失、取代、替换、错义突变或dna序列的框移或它们的任何组合。插入可包括插入可以是除内源序列以外的来源(例如异源序列)的整个基因或基因片段,例如外显子。在一些实施方案中,破坏可增加基因或基因产物(例如由基因编码的多肽)的表达和/或活性。在一些实施方案中,破坏可减少基因或基因产物的表达和/或活性。在一些实施方案中,破坏可改变基因或编码的基因产物(例如编码的多肽)的序列。在一些实施方案中,破坏可截短基因或编码的基因产物(例如编码的多肽)或将其片段化。在一些实施方案中,破坏可延长基因或编码的基因产物。在一些这种实施方案中,破坏可实现融合多肽的组装。在一些实施方案中,破坏可减少基因或基因产物的水平,而非活性。在一些实施方案中,破坏可影响基因或基因产物的活性,而非水平。在一些实施方案中,破坏可对基因或基因产物的水平无显著作用。在一些实施方案中,破坏可对基因或基因产物的活性无显著作用。在一些实施方案中,破坏可对基因或基因产物的水平或活性无显著作用。
[0212]
测定(determining)、测量、评估、评定、测定(assaying)和分析:在本文中可互换使用,是指任何测量形式,且包括确定要素是否存在。这些术语包括定量和/或定性测定。测定可以是相对或绝对的。“测定存在”可以是确定存在的某物的量,和/或确定它是否存在。
[0213]
内源启动子:如本文所用,是指例如在野生型生物体中与内源基因天然缔合的启动子。
[0214]
工程化:如本文所用,一般是指人为操纵的方面。举例来说,在一些实施方案中,当在自然界中不以某一顺序连接在一起的两种或更多种序列被人为操纵而在工程化多核苷酸中彼此直接连接时,多核苷酸可视为“工程化”。在一些实施方案中,工程化多核苷酸可包含调控序列,发现所述调控序列在自然界中与第一编码序列操作性缔合,但不与第二编码序列操作性缔合,通过人为连接,使其与第二编码序列可操作地缔合。可替代地或另外,在一些实施方案中,各编码在自然界中彼此不连接的多肽元件或结构域的第一和第二核酸序列在单个工程化多核苷酸中可彼此连接。同等地,在一些实施方案中,如果细胞或生物体被操纵以致于其遗传信息改变(例如引入先前不存在的新遗传物质,或先前存在的遗传物质已被改变或去除),那么其视为“工程化”。作为惯例且所属领域的技术人员所了解,工程化多核苷酸或细胞的后代通常仍然称为“工程化”,即使实际操纵是在先前实体上进行。此外,如所属领域的技术人员所了解,可利用能够实现如本文所述的“工程化”的多种方法。举例来说,在一些实施方案中,“工程化”可涉及通过使用被编程成进行分析或比较或以其它方式分析、推荐和/或选择序列、改变等)的计算机系统进行选择或设计(例如核酸序列、多肽序列、细胞、组织和/或生物体)。可替代地或另外,在一些实施方案中,“工程化”可涉及使用
体外化学合成方法和/或重组核酸技术,例如核酸扩增(例如经由聚合酶链反应)杂交、突变、转化、转染等和/或多种控制配对方法中的任一种。如所属领域的技术人员将了解,多种已确立的这种技术(例如重组dna、寡核苷酸合成和组织培养和转化(例如电穿孔、脂质粒转染等))是所属领域中众所周知的并描述于本说明书通篇引用和/或论述的各种通用和更特定的参考文献中。参见例如sambrook等人,molecular cloning:a laboratory manual第2版,cold spring harbor laboratory press,cold spring harbor,n.y.,1989和principles of gene manipulation:an introduction to genetic manipulation,第5版,by old,r.w.和s.b.primrose编辑,blackwell science公司,1994,以引用的方式整体并入本文中。
[0215]
功能:如本文所用,是指展现具体特性(例如形成编码序列的一部分)和/或活性的实体形式或片段(例如基因或基因区段)。举例来说,在免疫球蛋白的上下文中,可变区通过被组装(或重组)成形成功能编码序列的独特的基因区段(即v、d和/或j)来编码。当存在于基因组中时,虽然发生变异,但基因区段组织成簇。“功能”基因区段是在已经分离(即克隆)对应基因组dna的表达的序列(即可变区)中表示并通过序列鉴定的基因区段。一些免疫球蛋白基因区段序列含有开放阅读框且虽然未在表达的谱系中表示,但视为功能的,而其它免疫球蛋白基因区段序列含有引起终止密码子和/或截短库列的突变(例如点突变、插入、缺失等),随后使得这种基因区段序列不能进行与未突变序列相关的特性和/或活性。这种序列在表达的序列中未表示,因此,分类为伪基因。
[0216]
基因:如本文所用,是指染色体中编码产物(例如rna产物和/或多肽产物)的dna序列。在一些实施方案中,基因包括编码序列(即编码特定产物的序列)。在一些实施方案中,基因包括非编码序列。在一些特定实施方案中,基因可包括编码(例如外显子)与非编码(例如内含子)序列。在一些实施方案中,基因可包括一个或多个调控序列(例如启动子、增强子等)和/或内含子序列,所述序列例如可控制或影响基因表达的一个或多个方面(例如细胞类型特异性表达、诱导型表达等)。为求清楚,注意,在用于本公开中时,术语“基因”一般是指编码多肽或其片段的核酸部分;所属领域的一般技术人员将从上下文清楚,所述术语可任选地涵盖调控序列。本定义不意图排除术语“基因”应用于非蛋白质编码的表达单元,而是阐明在大多数情况下,如本文中所用的术语是指编码多肽的核酸。
[0217]
基因修饰非人类动物或基因工程非人类动物:在本文中可互换使用且是指任何非天然存在的非人类动物(例如啮齿动物,例如大鼠或小鼠),其中整体或部分地,所述非人类动物的一个或多个细胞含有异源核酸和/或编码所关注多肽的基因。举例来说,在一些实施方案中,“基因修饰非人类动物”或“基因工程非人类动物”是指含有如本文所述的转基因或转基因构建体的非人类动物。在一些实施方案中,异源核酸和/或基因直接或间接地通过引入前驱细胞中、借助于故意遗传操纵,例如通过显微注射或通过感染重组病毒来引入细胞中。术语遗传操纵不包括经典繁育技术,而是针对引入重组dna分子。所述分子可整合在染色体内。短语“基因修饰非人类动物”或“基因工程非人类动物”是指对于异源核酸和/或基因是杂合或纯合的动物,和/或具有异源核酸和/或基因的单个或多个拷贝的动物。
[0218]
种系构型:如本文所用,是指如在野生型动物(例如小鼠、大鼠或人类)的内源种系基因组中发现的序列排列(例如基因区段)。免疫球蛋白基因区段的种系构型的实例可见于例如lefranc,m
‑
p.,the immunoglobulin factsbook,academic press,2001年5月23日(本
文中称为“lefranc 2001”):
[0219]
人类重链可变区基因区段和人类重链恒定区基因的一种示例性构型可见于lefranc 2001第47页;
[0220]
人类λ轻链可变区基因区段和人类λ轻链恒定区基因的一种示例性构型可见于lefranc 2001第61页;
[0221]
人类κ轻链可变区基因区段和人类κ轻链恒定区基因的一种示例性构型可见于lefranc 2001第53页;
[0222]
小鼠重链可变区基因区段和小鼠重链恒定区基因的一种示例性构型可见于lucas,j.等人,chapter 1:the structure and regulation of the immunoglobulin loci,molecular biology of b cells,第2版,academic press,2015(lucas);
[0223]
小鼠λ轻链可变区基因区段和小鼠λ轻链恒定区基因的一种示例性构型可见于lefranc,m
‑
p等人,chapter 4:immunoglobulin lambda(igl)genes of human and mouse,molecular biology of b cells,第1版,academic press,2004(lefranc 2004);
[0224]
小鼠κ轻链可变区基因区段和小鼠κ轻链恒定区基因的一种示例性构型可见于christele,m
‑
j等人,nomenclature and overview of the mouse(mus musculus and mus sp.)immunoglobulin kappa(igk)genes,exp clin immunogenet 2001,18:255
‑
279(christele);
[0225]
lefranc 2001、lucas、lefranc 2004和christele的每一引述部分以引用的方式并入本文中。
[0226]
种系基因组:如本文所用,是指在动物形成中使用的生殖细胞(例如配子,例如精子或卵)中发现的基因组。种系基因组是动物中细胞的基因组dna的来源。因而,在种系基因组中具有修饰的动物(例如小鼠或大鼠)被视为在其所有细胞的基因组dna中均具有修饰。
[0227]
种系序列:如本文所用,是指如在野生型动物(例如小鼠、大鼠或人类)的内源种系基因组中发现的dna序列,或由如在动物(例如小鼠、大鼠或人类)的内源种系基因组中发现的dna序列编码的rna或氨基酸序列。免疫球蛋白基因区段的代表性种系序列可见于例如lefranc 2001:
[0228]
在如本文所述的一些实施方案中可利用的人类v
h
基因区段的代表性种系核苷酸序列和人类v
h
基因区段的代表性种系氨基酸序列可见于lefranc 2001第107
‑
234页;
[0229]
在如本文所述的一些实施方案中可利用的人类d基因区段的代表性种系核苷酸序列和人类d基因区段的代表性种系氨基酸序列可见于lefranc 2001第98
‑
100页;
[0230]
在如本文所述的一些实施方案中可利用的人类j
h
基因区段的代表性种系核苷酸序列和人类j
h
基因区段的代表性种系氨基酸序列可见于lefranc 2001第104页;
[0231]
在如本文所述的非人类动物的一些实施方案中可利用的人类vλ基因区段的代表性种系核苷酸序列和人类vλ基因区段的代表性种系氨基酸序列可见于lefranc 2001第350
‑
428页;以及
[0232]
在如本文所述的非人类动物的一些实施方案中可利用的人类jλ基因区段的代表性种系核苷酸序列和人类jλ基因区段的代表性种系氨基酸序列可见于lefranc 2001第346页。
[0233]
lefranc 2001的各引述部分以引用的方式并入本文中。
[0234]
异源:如本文所用,是指来自不同来源的剂或实体。举例来说,当提及特定细胞或生物体中存在的多肽、基因或基因产物时使用时,所述术语阐明相关多肽、基因或基因产物:1)被人为工程化;2)人为(例如经由基因工程化)引入细胞或生物体中(或其前驱细胞);和/或3)并非相关细胞或生物体(例如相关细胞类型或生物体类型)天然产生或其中不存在。“异源”还包括特定天然细胞或生物体中通常存在,但已例如通过突变或置于非天然相关和在一些实施方案中非内源调控元件(例如启动子)的控制下而改变或修饰的多肽、基因或基因产物。
[0235]
宿主细胞:如本文所用,是指已引入核酸或蛋白质的细胞。熟练人员在阅读本公开后将了解,这类术语不仅指具体的受试者细胞,还用于指这种细胞的后代。因为在后续世代中由于突变或者环境影响而可能存在某些修饰,所以这种后代事实上可不与母细胞一致,但仍然包括在短语“宿主细胞”范围内。在一些实施方案中,宿主细胞是或包含原核或真核细胞。一般来说,宿主细胞是适合于接受和/或产生异源核酸或蛋白质的任何细胞,不管细胞表示的生命界如何。示例性细胞包括原核生物和真核生物的细胞(单细胞或多细胞)、细菌细胞(例如大肠杆菌、芽孢杆菌属(bacillus spp.)、链霉菌属(streptomyces spp.)等菌株)、分枝杆菌属(mycobacteria)细胞、真菌细胞、酵母细胞(例如酿酒酵母(saccharomyces cerevisiae)、粟酒裂殖酵母(schizosaccharomyces pombe)、巴氏毕赤酵母(pichia pastoris)、甲醇毕赤酵母(pichia methanolica)等)、植物细胞、昆虫细胞(例如sf
‑
9、sf
‑
21、杆状病毒感染的昆虫细胞、粉纹夜蛾(trichoplusia ni)等)、非人类动物细胞、人类细胞或细胞融合物,例如杂交瘤或四价体瘤。在一些实施方案中,细胞是人、猴、猿、仓鼠、大鼠或小鼠细胞。在一些实施方案中,细胞是真核的且选自以下细胞:中国仓鼠卵巢(cho)(例如cho k1、dxb
‑
11cho、veggie
‑
cho)、cos(例如cos
‑
7)、视网膜细胞、vero、cv1、肾(例如hek293、293ebna、msr 293、mdck、hak、bhk)、hela、hepg2、wi38、mrc 5、colo205、hb 8065、hl
‑
60、(例如bhk21)、jurkat、daudi、a431(表皮)、cv
‑
1、u937、3t3、l细胞、c127细胞、sp2/0、ns
‑
0、mmt 060562、sertoli细胞、brl 3a细胞、ht1080细胞、骨髓瘤细胞、肿瘤细胞和源自于前述细胞的细胞系。在一些实施方案中,细胞包含一个或多个病毒基因,例如表达病毒基因的视网膜细胞(例如细胞)。在一些实施方案中,宿主细胞是或包含分离的细胞。在一些实施方案中,宿主细胞是组织的部分。在一些实施方案中,宿主细胞是生物体的部分。
[0236]
同一性:如本文中结合序列比较所用,是指如通过所属领域中已知的可用于测量核苷酸和/或氨基酸序列同一性的许多不同算法确定的同一性。在一些实施方案中,如本文所述的同一性是使用clustalw v.1.83(缓慢)比对采用10.0的开放空位罚分、0.1的延伸空位罚分和使用gonnet相似矩阵(macvector
tm 10.0.2,macvector inc.,2008)确定。
[0237]
代替:如本文所用,是指一种位置取代,其中第一核酸序列位于染色体中第二核酸序列的位置上(例如其中第二核酸序列先前(例如最初)位于染色体中,例如第二核酸序列的内源基因座上)。短语“代替”不要求从例如基因座或染色体去除第二核酸序列。在一些实施方案中,第二核酸序列和第一核酸序列彼此可比,因为例如第一和第二序列彼此同源,含有对应元件(例如蛋白质编码元件、调控元件等)和/或具有类似或相同序列。在一些实施方案中,第一和/或第二核酸序列包括启动子、增强子、剪接供体位点、剪接受体位点、内含子、外显子、非翻译区(utr)中的一个或多个;在一些实施方案中,第一和/或第二核酸序列包括
一个或多个编码序列。在一些实施方案中,第一核酸序列是第二核酸序列的同源物或变体(例如突变体)。在一些实施方案中,第一核酸序列是第二序列的直系同源物或同源物。在一些实施方案中,第一核酸序列是或包含人类核酸序列。在一些实施方案中,包括在第一核酸序列是或包含人类核酸序列下,第二核酸序列是或包含啮齿动物序列(例如小鼠或大鼠序列)。在一些实施方案中,包括在第一核酸序列是或包含人类核酸序列下,第二核酸序列是或包含人类序列。在一些实施方案中,第一核酸序列是第二序列的变体或突变体(即与第二序列相比含有一个或多个序列差异,例如取代的序列)。如此置放的核酸序列可包括作为用于获得如此置放的序列的源核酸序列的一部分的一个或多个调控序列(例如启动子、增强子、5'
‑
或3'
‑
非翻译区等)。举例来说,在多个实施方案中,第一核酸序列是内源序列经异源序列取代,由此从如此置放的核酸序列(包含异源序列)产生基因产物,但不表达内源序列;第一核酸序列具有核酸序列编码具有与由内源序列编码的多肽类似功能的多肽的内源基因组序列(例如内源基因组序列编码非人类可变区多肽整体或部分,且dna片段编码一个或多个人类可变区多肽整体或部分)。在多个实施方案中,人类免疫球蛋白基因区段或其片段代替内源非人类免疫球蛋白基因区段或片段。
[0238]
体外:如本文所用,是指在人造环境中,例如在试管或反应容器中、在细胞培养物中等发生,而非在多细胞生物体内发生的事件。
[0239]
体内:如本文所用,是指在多细胞生物体,例如人类和/或非人类动物内发生的事件。在基于细胞的系统的背景下,该术语可用于指在活细胞内发生的事件(与例如体外系统相对比)。
[0240]
分离:如本文所用,是指已经(1)与在最初产生时所关联的至少一些组分分离(无论在自然界中和/或在实验背景下)和/或(2)人为设计、产生、制备和/或制造的物质和/或实体。分离的物质和/或实体可以与约10%、约20%、约30%、约40%、约50%、约60%、约70%、约80%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%或超过约99%的其最初相关联的其它组分分离。在一些实施方案中,分离的剂与10%至100%、15%
‑
100%、20%
‑
100%、25%
‑
100%、30%
‑
100%、35%
‑
100%、40%
‑
100%、45%
‑
100%、50%
‑
100%、55%
‑
100%、60%
‑
100%、65%
‑
100%、70%
‑
100%、75%
‑
100%、80%
‑
100%、85%
‑
100%、90%
‑
100%、95%
‑
100%、96%
‑
100%、97%
‑
100%、98%
‑
100%或99%
‑
100%的其最初相关联的其它组分分离。在一些实施方案中,分离的剂与10%至100%、10%
‑
99%、10%
‑
98%、10%
‑
97%、10%
‑
96%、10%
‑
95%、10%
‑
90%、10%
‑
85%、10%
‑
80%、10%
‑
75%、10%
‑
70%、10%
‑
65%、10%
‑
60%、10%
‑
55%、10%
‑
50%、10%
‑
45%、10%
‑
40%、10%
‑
35%、10%
‑
30%、10%
‑
25%、10%
‑
20%或10%
‑
15%的其最初相关联的其它组分分离。在一些实施方案中,分离的剂与11%至99%、12%
‑
98%、13%
‑
97%、14%
‑
96%、15%
‑
95%、20%
‑
90%、25%
‑
85%、30%
‑
80%、35%
‑
75%、40%
‑
70%、45%
‑
65%、50%
‑
60%或55%
‑
60%的其最初相关联的其它组分分离。在一些实施方案中,分离的剂约80%、约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%或超过约99%纯。在一些实施方案中,分离的剂80%
‑
99%、85%
‑
99%、90%
‑
99%、95%
‑
99%、96%
‑
99%、97%
‑
99%或98%
‑
99%纯。在一些实施方案中,分离的剂80%
‑
99%、80%
‑
98%、80%
‑
97%、80%
‑
96%、80%
‑
95%、80%
‑
90%或80%
‑
85%纯。在一些实施方案中,分离的剂85%
‑
98%、90%
‑
97%或95%
‑
96%纯。在一些实施方案中,如果物
质基本上不含其它组分,则其是“纯的”。在一些实施方案中,如所属领域的技术人员所了解,物质在与例如一种或多种载体或赋形剂(例如缓冲剂、溶剂、水等)的某些其它组分组合之后仍可视为“分离”或甚至“纯的”;在这类实施方案中,物质的分离或纯度百分比是在不包括这种载体或赋形剂下计算的。仅举一例,在一些实施方案中,在如下情形时,自然界中存在的例如多肽或多核苷酸的生物聚合物视为“分离”:a)其来源或起源不与在自然界中在天然状态下伴随的一些或所有组分相关联;b)其基本上不含来自与在自然界中产生其的物种相同的物种的其它多肽或核酸;或c)由并非在自然界中产生其的物种的细胞或其它表达系统表达或以其它方式与来自所述细胞或其它表达系统的组分相关联。因此,举例来说,在一些实施方案中,化学上合成或在与在自然界中产生其的细胞不同的细胞系统中合成的多肽视为“分离”的多肽。可替代地或另外,在一些实施方案中,在已经与以下的其它组分分离的程度上,已经受一种或多种纯化技术的多肽可视为“分离”的多肽:a)在自然界中相关联的组分;和/或b)最初产生时所关联的组分。
[0241]
基因座:如本文所用,是指基因、dna序列、多肽编码序列的定位,或在生物体的基因组的染色体上的位置。举例来说,“免疫球蛋白基因座”可指免疫球蛋白基因区段(例如v、d、j或c)、免疫球蛋白基因区段dna序列、免疫球蛋白基因区段编码序列的定位,或确定为存在这种序列的生物体的基因组的染色体上的免疫球蛋白基因区段位置。“免疫球蛋白基因座”可包含免疫球蛋白基因区段的调控元件,包括(但不限于)增强子、启动子、5'和/或3'调控序列或区域、或它们的组合。“免疫球蛋白基因座”可包含基因间dna,例如通常存在于或出现在野生型基因座中的基因区段之间的dna。所属领域的一般技术人员将了解,在一些实施方案中,染色体可含有数百或甚至数千基因并证明当在不同物种之间比较时类似遗传基因座的物理共定位。这种遗传基因座可称作共享同线性。
[0242]
天然出现:如本文所用,在提及生物要素(例如核酸序列)时,意指可在细胞或生物体(例如动物)中在规定背景和/或位置中在缺乏工程化(例如基因工程化)下发现该生物要素。换句话说,在规定背景和/或位置中天然出现的序列不在由工程化(例如基因工程化)产生的规定背景和/或位置中。举例来说,天然出现在内源人类免疫球蛋白κ轻链基因座中人类jκ1基因区段相邻处的序列是可在人类中内源人类免疫球蛋白κ轻链基因座中人类jλ1基因区段相邻处找到的序列,缺乏基因工程化。在一些实施方案中,可从在细胞或生物体中天然出现之处获得、得到和/或分离序列。在一些实施方案中,细胞或生物体不是天然出现在细胞或生物体中的序列的直接来源。举例来说,可鉴定细胞或生物体中的对应序列,接着通过所属领域中已知的机制产生或复制。
[0243]
非人类动物:如本文所用,是指非人类的任何脊椎动物生物体。在一些实施方案中,非人类动物是圆口类动物、多骨鱼、软骨鱼(例如鲨鱼或鳐)、两栖动物、爬行动物、哺乳动物和禽类。在一些实施方案中,非人类动物是哺乳动物。在一些实施方案中,非人类哺乳动物是灵长类动物、山羊、绵羊、猪、犬、母牛或啮齿动物。在一些实施方案中,非人类动物是啮齿动物,例如大鼠或小鼠。
[0244]
核酸:如本文所用,是指并入或可并入寡核苷酸链中的任何化合物和/或物质。在一些实施方案中,“核酸”是经由磷酸二酯键并入或可并入寡核苷酸链中的化合物和/或物质。如从上下文显而易见,在一些实施方案中,“核酸”是指个别的核酸残基(例如核苷酸和/或核苷);在一些实施方案中,“核酸”是指包含个别的核酸残基的寡核苷酸链。在一些实施
方案中,“核酸”是或包含rna;在一些实施方案中,“核酸”是或包含dna。在一些实施方案中,“核酸”是、包含一种或多种天然核酸残基或由其组成。在一些实施方案中,“核酸”是、包含一种或多种天然核酸类似物或由其组成。在一些实施方案中,核酸类似物与“核酸”的不同之处在于其不利用磷酸二酯主链。举例来说,在一些实施方案中,“核酸”是、包含一种或多种“肽核酸”或由其组成,“肽核酸”是所属领域中已知并在主链中肽键代替磷酸二酯键。可替代地或另外,在一些实施方案中,“核酸”具有一个或多个硫代磷酸酯键和/或5'
‑
n
‑
氨基磷酸酯键,而非磷酸二酯键。在一些实施方案中,“核酸”是以下、包含以下或由以下组成:一个或多个天然核苷(例如腺苷、胸苷、鸟苷、胞苷、尿苷、脱氧腺苷、脱氧胸苷、脱氧鸟苷和脱氧胞苷)。在一些实施方案中,“核酸”是以下、包含以下或由以下组成:一个或多个核苷类似物(例如2
‑
氨基腺苷、2
‑
硫胸苷、肌苷、吡咯并嘧啶、3
‑
甲基腺苷、5
‑
甲基胞苷、c
‑
5丙炔基
‑
胞苷、c
‑
5丙炔基
‑
尿苷、2
‑
氨基腺苷、c5
‑
溴尿苷、c5
‑
氟尿苷、c5
‑
碘尿苷、c5
‑
丙炔基
‑
尿苷、c5
‑
丙炔基
‑
胞苷、c5
‑
甲基胞苷、2
‑
氨基腺苷、7
‑
脱氮腺苷、7
‑
脱氮鸟苷、8
‑
氧代基腺苷、8
‑
氧代基鸟苷、o(6)
‑
甲基鸟嘌呤、2
‑
硫胞苷、甲基化碱基、插入碱基和它们的组合)。在一些实施方案中,与天然核酸中的糖比较,“核酸”包含一个或多个修饰糖(例如2'
‑
氟核糖、核糖、2'
‑
脱氧核糖、阿拉伯糖和己糖)。在一些实施方案中,“核酸”具有编码例如rna或多肽的功能基因产物的核苷酸序列。在一些实施方案中,“核酸”包括一个或多个内含子。在一些实施方案中,“核酸”包括一个或多个外显子。在一些实施方案中,“核酸”通过以下中的一个或多个来制备:从天然来源分离、通过基于互补模板(体内或体外)聚合而酶促合成、在重组细胞或系统中重现和化学合成。在一些实施方案中,“核酸”至少例如但不限于3个、4个、5个、6个、7个、8个、9个、10个、15个、20个、25个、30个、35个、40个、45个、50个、55个、60个、65个、70个、75个、80个、85个、90个、95个、100个、110个、120个、130个、140个、150个、160个、170个、180个、190个、20个、225个、250个、275个、300个、325个、350个、375个、400个、425个、450个、475个、500个、600个、700个、800个、900个、1000个、1500个、2000个、2500个、3000个、3500个、4000个、4500个、5000个或更多个残基长。在一些实施方案中,“核酸”是单股;在一些实施方案中,“核酸”是双股。在一些实施方案中,“核酸”具有包含至少一个编码多肽或与编码多肽的序列互补的元件的核苷酸序列。在一些实施方案中,“核酸”具有酶活性。
[0245]
可操作地连接:如本文所用,是指组分并置,其中所述组分处于允许其以所欲方式发挥作用的关系中(例如当组分存在于适当组织、细胞类型、细胞活动等中)时。举例来说,如果在b细胞发育中的适当时间,v
h
、d和j
h
基因区段可剪接至重链恒定区,不管这种剪接是否存在于例如免疫系统外的细胞(例如生殖细胞)中,那么一个或多个v
h
基因区段、一个或多个d基因区段和一个或多个j
h
基因区段“可操作地连接”至重链恒定区。“可操作地连接”至编码序列的控制序列以使编码序列在与控制序列相容的条件下表达的方式连结。“可操作地连接”的序列包括与所关注基因相连的表达控制序列和反式作用或隔开一定距离控制所关注基因(或所关注序列)的表达控制序列两者。术语“表达控制序列”包括影响其所连结的编码序列的表达和加工所必需的多核苷酸序列。“表达控制序列”包括:适当转录起始、终止、启动子和增强子序列;高效rna加工信号,例如剪接和多腺苷酸化信号;稳定细胞质mrna的序列;增强翻译效率(即科扎克共有序列(kozak consensus sequence))的序列;增强多肽稳定性的序列;和需要时,增强多肽分泌的序列。这种控制序列的性质视宿主生物体而不同。举例来说,在原核生物中,这种控制序列一般包括启动子、核糖体结合位点和转录终止
序列,而在真核生物中,这种控制序列通常包括启动子和转录终止序列。术语“控制序列”意图包括存在对于表达和加工来说不可或缺的组分,且还可以包括存在有利的额外组分,例如前导序列和融合搭配物序列。
[0246]
多肽:如本文所用,是指氨基酸的任何聚合链。在一些实施方案中,多肽具有在自然界中存在的氨基酸序列。在一些实施方案中,多肽具有在自然界中不存在的氨基酸序列。在一些实施方案中,多肽具有含有在自然界中彼此分开存在的部分的氨基酸序列(即来自两种或更多种不同生物体,例如人类和非人类部分)。在一些实施方案中,多肽具有因通过人为作用设计和/或产生而工程化的氨基酸序列。在一些实施方案中,多肽具有由在自然界中不存在的序列(例如因为通过人为作用设计和/或产生以编码所述多肽而工程化的序列)编码的氨基酸序列。
[0247]
重排:如本文所用,描述dna序列包括两个或更多个接合(直接或间接)在一起的免疫球蛋白基因区段,以便接合在一起的基因区段具有编码免疫球蛋白的可变区的dna序列。重排dna序列的两个或更多个免疫球蛋白基因区段不再与发挥功能的重组信号序列(rss)相关联,因而无法经历进一步重排。所属领域的技术人员将认识到,虽然重排的dna序列的两个或更多个免疫球蛋白基因区段不能进一步重排,但其并不意指同一基因座内的其它免疫球蛋白基因区段无法进行例如第二次重排。所属领域的技术人员将了解,重排的基因区段(例如在重排的免疫球蛋白可变区中)可经由天然vdj重组过程接合在一起。所属领域的技术人员还将了解,重排的基因区段(例如在重排的免疫球蛋白可变区中)可被工程化成例如通过使用标准重组技术接合所述基因区段而接合在一起。重排的免疫球蛋白可变区通常包括两个或更多个接合的免疫球蛋白基因区段。举例来说,重排的免疫球蛋白λ轻链可变区可包括与jλ基因区段接合的vλ基因区段。重排的免疫球蛋白重链可变区可包括接合的v
h
基因区段、d基因区段、j
h
基因区段。所属领域的技术人员还将了解,一般在重排的免疫球蛋白可变区中的免疫球蛋白基因区段之间去除所有或基本上所有基因间序列。所属领域的技术人员将进一步了解,重排的序列可尤其在基因区段中包括内含子。
[0248]
重组:如本文所用,是指由使来自多种来源(例如生物体、组织、细胞、基因组或基因组部分)的遗传物质汇集在一起的基因重组的实验室方法(例如克隆)形成的分子(例如dna、rna或多肽)。在一些实施方案中,通过重组方式设计、工程化、制备、表达、建立或分离重组多肽,所述方式例如使用转染至宿主细胞中的重组表达载体表达的多肽、从重组的组合人类多肽文库分离的多肽(hoogenboom,h.r.,1997,tib tech.15:62
‑
70;azzazy,h.和w.e.highsmith,2002,clin.biochem.35:425
‑
45;gavilondo,j.v.和j.w.larrick,2002,biotechniques 29:128
‑
45;hoogenboom h.和p.chames,2000,immunol.today 21:371
‑
8,以引用的方式整体并入本文中)、从已被基因工程化成包括人类免疫球蛋白基因的动物(例如小鼠)分离的抗体(参见例如taylor,l.d.等人,1992,nucl.acids res.20:6287
‑
95;kellermann,s
‑
a.和l.l.green,2002,curr.opin.biotechnol.13:593
‑
7;little,m.等人,2000,immunol.today 21:364
‑
70;osborn,m.j.等人,2013,j.immunol.190:1481
‑
90;lee,e
‑
c.等人,2014,nat.biotech.32(4):356
‑
63;macdonald,l.e.等人,2014,proc.natl.acad.sci.u.s.a.111(14):5147
‑
52;murphy,a.j.等人,2014,proc.natl.acad.sci.u.s.a.111(14):5153
‑
8,各以引用的方式整体并入本文中)或通过涉及将所选序列元件剪接至彼此的任何其它方式制备、表达、建立或分离的多肽。在一些实施
方案中,在自然界中发现一个或多个这类所需序列元件。在一些实施方案中,一个或多个这类所需序列元件是电脑模拟设计的。在一些实施方案中,一个或多个这类所选序列元件由例如来自天然或合成(例如人造)来源的已知序列元件的诱变(例如体内或体外)产生。举例来说,在一些实施方案中,重组多肽由在所关注源生物体(例如人类、小鼠等)的基因组中发现的序列构成。在一些实施方案中,重组多肽具有由诱变(例如体外或体内,例如在非人类动物中)产生的氨基酸序列,使得所述重组多肽的氨基酸序列是虽然来源于多肽序列并与多肽序列有关,但可能不天然存在于体内非人类动物的基因组内的序列。
[0249]
参考:如本文所用,是指所关注的剂、动物、群、个体、群体、样品、序列或值进行比较的标准或对照剂、动物、群、个体、群体、样品、序列或值。在一些实施方案中,基本上与所关注的剂、动物、群、个体、群体、样品、序列或值的测试或测定同时测试和/或确定参考剂、动物、群、个体、群体、样品、序列或值。在一些实施方案中,参考剂、动物、群、个体、群体、样品、序列或值是历史参考,任选地收录在有形媒介中。在一些实施方案中,参考可指对照。“参考”还包括“参考动物”。“参考动物”可具有如本文所述的修饰、与本文所述不同的修饰或无修饰(即野生型动物)。通常,如所属领域的技术人员将了解,参考剂、动物、群、个体、群体、样品、序列或值是在与用于测定或表征所关注的剂、动物(例如哺乳动物)、群、个体、群体、样品、序列或值的条件相当的条件下测定或表征的。
[0250]
替代:如本文所用,是指在宿主基因座中(例如基因组中)发现的“被替代”的核酸序列(例如基因)从所述基因座去除且不同的“替代”核酸位于其位置的过程。在一些实施方案中,被替代的核酸序列和替代核酸序列彼此可比,因为例如它们彼此同源,含有对应元件(例如蛋白质编码元件、调控元件等)和/或具有类似或相同序列。在一些实施方案中,被替代的核酸序列包括启动子、增强子、剪接供体位点、剪接受体位点、内含子、外显子、非翻译区(utr)中的一个或多个;在一些实施方案中,替代核酸序列包括一个或多个编码序列。在一些实施方案中,替代核酸序列是被替代的核酸序列的同源物或变体(例如突变体)。在一些实施方案中,替代核酸序列是被替代的序列的直系同源物或同源物。在一些实施方案中,替代核酸序列是或包含人类核酸序列。在一些实施方案中,包括在替代核酸序列是或包含人类核酸序列下,被替代的核酸序列是或包含啮齿动物序列(例如小鼠或大鼠序列)。在一些实施方案中,包括在替代核酸序列是或包含人类核酸序列下,被替代的核酸序列是或包含人类序列。在一些实施方案中,替代核酸序列为被替代的序列的变体或突变体(即与被替代的序列相比,含有一个或多个序列差异,例如取代的序列)。如此置放的核酸序列可包括作为用于获得如此置放的序列的源核酸序列的一部分的一个或多个调控序列(例如启动子、增强子、5'
‑
或3'
‑
非翻译区等)。举例来说,在多个实施方案中,替代是内源序列经异源序列取代,从如此置放的核酸序列产生基因产物(包含异源序列),但不表达内源序列;替代是内源基因组序列经编码与由内源序列编码的多肽具有类似功能的多肽的核酸序列(例如内源基因组序列编码非人类可变区多肽整体或部分,且dna片段编码一种或多种人类可变区多肽整体或部分)替代。在多个实施方案中,内源非人类免疫球蛋白基因区段或其片段经人类免疫球蛋白基因区段或其片段替代。
[0251]
基本上:如本文所用,是指展现全部或靠近全部程度的所关注的特征或特性的定性条件。生物领域中的一般技术人员将了解,生物和化学现象很少(如果有过)完整和/或进行完整或实现或避免绝对结果。因此,术语“基本上”在本文中用于记录在许多生物和化学
现象中固有的潜在完整缺乏。
[0252]
基本类似性:如本文所用,是指氨基酸或核酸序列之间的比较。如所属领域的一般技术人员将了解,如果两个序列在对应位置中含有类似残基(例如氨基酸或核苷酸),那么其一般被视为“基本上类似”。如所属领域中所了解,虽然类似残基可以是相同残基(又见以下基本同一性),但类似残基还可以是具有适当可比的结构和/或功能特征的不相同残基。举例来说,如所属领域的一般技术人员所众所周知,某些氨基酸通常分类为“疏水性”或“亲水性”氨基酸,和/或具有“极性”或“非极性”侧链。一种氨基酸经另一相同类型取代常可视为“保守”取代。典型的氨基酸分类概述于下表中。
[0253]
丙氨酸alaa非极性中性1.8精氨酸argr极性正
‑
4.5天冬酰胺asnn极性中性
‑
3.5天冬氨酸aspd极性负
‑
3.5半胱氨酸cysc非极性中性2.5谷氨酸glue极性负
‑
3.5谷氨酰胺glnq极性中性
‑
3.5甘氨酸glyg非极性中性
‑
0.4组氨酸hish极性正
‑
3.2异亮氨酸ilei非极性中性4.5亮氨酸leul非极性中性3.8赖氨酸lysk极性正
‑
3.9甲硫氨酸metm非极性中性1.9苯丙氨酸phef非极性中性2.8脯氨酸prop非极性中性
‑
1.6丝氨酸sers极性中性
‑
0.8苏氨酸thrt极性中性
‑
0.7色氨酸trpw非极性中性
‑
0.9酪氨酸tyry极性中性
‑
1.3缬氨酸valv非极性中性4.2
[0254][0255][0256]
如所属领域中所众所周知,可使用多种算法中的任一种比较氨基酸或核酸序列,所述算法包括可用于商业计算机程序中的算法,例如用于核苷酸序列的blastn、和用于氨基酸序列的blastp、空位blast和psi
‑
blast。示例性这类程序描述于altschul,s.f.等人,
1990,j.mol.biol.,215(3):403
‑
10;altschul,s.f.等人,1996,meth.enzymol.266:460
‑
80;altschul,s.f.等人,1997,nucleic acids res.,25:3389
‑
402;baxevanis,a.d.和b.f.f.ouellette(编辑)bioinformatics:a practical guide to the analysis of genes and proteins,wiley,1998;和misener等人(编辑)bioinformatics methods and protocols,methods in molecular biology,第132卷,humana press,1998,以引用的方式整体并入本文中。除鉴定类似序列外,以上提及的程序通常指示相似度。在一些实施方案中,如果在残基相关延伸段上两个序列的对应残基的至少、例如但不限于80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更多相似(例如同一或包括保守取代),那么所述两个序列被视为基本上类似。在一些实施方案中,相关延伸段是完整序列(例如基因序列、基因区段、编码结构域的序列、多肽或结构域)。在一些实施方案中,相关延伸段是至少9个、10个、11个、12个、13个、14个、15个、16个、17个或更多个残基。在一些实施方案中,相关延伸段是至少10个、15个、20个、25个、30个、35个、40个、45个、50个或更多个残基。在一些实施方案中,相关延伸段包括沿着完整序列的相连残基。在一些实施方案中,相关延伸段包括沿着完整序列的不相连残基,例如通过多肽或其一部分的折叠构形而汇集在一起的非相连残基。
[0257]
基本同一性:如本文所用,是指氨基酸或核酸序列之间的比较。如所属领域的一般技术人员将了解,如果两个序列在对应位置中含有相同残基(例如氨基酸或核苷酸),则其一般被视为“基本上同一”。如所属领域中所众所周知,可使用多种算法中的任一种比较氨基酸或核酸序列,所述算法包括可用于商业计算机程序中的算法,例如用于核苷酸序列的blastn、和用于氨基酸序列的blastp、空位blast和psi
‑
blast。示例性这类程序描述于altschul,s.f.等人,1990,j.mol.biol.,215(3):403
‑
10;altschul,s.f.等人,1996,meth.enzymol.266:460
‑
80;altschul,s.f.等人,1997,nucleic acids res.,25:3389
‑
402;baxevanis,a.d.和b.f.f.ouellette(编辑)bioinformatics:a practical guide to the analysis of genes and proteins,wiley,1998;和misener等人(编辑)bioinformatics methods and protocols,methods in molecular biology,第132卷,humana press,1998,各以引用的方式整体并入本文中。除鉴定相同序列外,以上提及的程序通常指示同一性程度。在一些实施方案中,如果在残基相关延伸段上两个序列的对应残基的至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更多同一,则所述两个序列被视为基本上同一。在一些实施方案中,残基相关延伸段是完整序列。在一些实施方案中,残基相关延伸段是例如但不限于至少10个、15个、20个、25个、30个、35个、40个、45个、50个或更多个残基。
[0258]
靶向构建体或靶向载体:如本文所用,是指包含靶向区域的多核苷酸分子。靶向区域包含与标靶细胞、组织或动物中的序列同一或基本上同一且提供靶向构建体经由同源重组整合至所述细胞、组织或动物的基因组内部的位置中的序列。本文还包括和描述使用位点特异性重组酶识别位点(例如loxp或frt位点)靶向的靶向区域。在一些实施方案中,如本文所述的靶向构建体还包含特别关注的核酸序列或基因、选择标记、控制和/或调控序列和允许经由外源添加帮助或促进这种序列重组的蛋白质而介导的重组的其它核酸序列。在一些实施方案中,如本文所述的靶向构建体还包含所关注基因整体或部分,其中所关注基因是编码具有与由内源序列编码的蛋白质类似的功能的多肽整体或部分的异源基因。在一些
实施方案中,如本文所述的靶向构建体还包含所关注人源化基因整体或部分,其中所关注人源化基因编码具有与由内源序列编码的多肽类似的功能的多肽整体或部分。在一些实施方案中,靶向构建体(或靶向载体)可包含通过人为操纵的核酸序列。举例来说,在一些实施方案中,靶向构建体(或靶向载体)可以被构建成含有工程化或重组多核苷酸,所述工程化或重组多核苷酸含有两个或更多个在自然界中不以所述顺序连接在一起但通过人为操纵而在所述工程化或重组多核苷酸中直接彼此连接的序列。
[0259]
转基因或转基因构建体:如本文所用,是指已例如通过本文所述的方法人为引入细胞中的核酸序列(编码例如所关注多肽整体或部分)。转基因对于引入其的基因工程动物或细胞来说可以是部分或完全异源,即外来的。转基因可包括一个或多个转录调控序列和表达所选核酸序列所必需的任何其它核酸,例如内含子或启动子。
[0260]
未重排:如本文所用,描述dna序列包括两个或更多个尚未经历重组事件或以其它方式接合,因此在之间包括基因间序列的免疫球蛋白基因区段。所属领域的技术人员将了解,未重排v基因区段和j基因区段可与完整重组信号序列(rss)缔合。未重排d基因区段两侧可以是两个完整重组信号序列(rss)。所属领域的技术人员将进一步了解,未重排基因区段可尤其包括内含子。
[0261]
载体:如本文所用,是指能够输送所相关联的另一核酸的核酸分子。在一些实施方案中,载体能够在例如真核和/或原核细胞的宿主细胞中在染色体外复制和/或表达其所连接的核酸。能够指导可操作地连接的基因的表达的载体在本文中称为“表达载体”。
[0262]
野生型:如本文所用,是指具有如在自然界中在“正常”(与突变、患病、改变、工程化等相对比)状态或背景下发现的结构和/或活性的实体。所属领域的一般技术人员将了解,野生型基因和多肽常以多种不同形式(例如等位基因)存在。
具体实施方式
[0263]
本公开洞察到非人类动物中的内源抗体形成机制,包括免疫球蛋白链配对和亲和力成熟,可用于产生抗原特异性的高亲和力抗体,包括外源人类免疫球蛋白序列。这种动物可产生正常和稳固的免疫反应,所述免疫反应可用于制造例如人类抗体治疗剂。本公开认识到,基因修饰非人类动物提供了一个产生包括人类可变结构域,包括人类重链、κ轻链和λ轻链结构域的抗体的有效且高效的平台。本公开进一步认识到,基因修饰非人类动物能够成功地利用人类重链、κ轻链和λ轻链可变区基因区段来产生亲和力成熟的人类重链、κ轻链和λ轻链可变结构域。
[0264]
本公开认识到在非人类动物中包括人类λ轻链可变结构域的抗体的产生先前面临挑战,因为某些非人类动物中λ轻链的表达低。举例来说,小鼠利用κ轻链显著超过λ轻链(即,约95:5的κ:λ比率)。本公开进一步认识到通过限制非人类动物的轻链可变区谱系来产生包括通用轻链(例如能够结合于多个重链的轻链)的抗体可能是挑战性的,因为它消除了一些对于高亲和力抗体产生来说最强大的多样性产生机制,例如组合多样性、连接多样性和第二次重排。它还显著地使另外由“混合和匹配”重链和轻链配对所引起的多样性产生作用最小。鉴于这些认识,在所属领域中仍然需要用于在非人类动物中从有限人类轻链可变区产生人类λ可变区的平台和方法。
[0265]
本公开洞察到在κ轻链基因座处包括有限人类λ轻链可变区谱系的非人类动物(例
如啮齿动物,例如大鼠或小鼠)能够有效产生高亲和力的抗原特异性抗体。这个结果是意外的,因为非人类动物中的有限人类λ轻链可变区谱系会迫使非人类动物利用λ轻链可变区序列。λ轻链可变区序列的使用与某些非人类动物的自然偏好是相反的。如上所提及,非人类动物中使用有限人类λ轻链可变区谱系还消除了产生高亲和力的抗原特异性抗体的许多天然机制。
[0266]
本公开提供了表达人类免疫球蛋白λ轻链可变结构域的基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠),其中非人类动物具有有限人类λ轻链可变区谱系(包含一个或两个人类vλ基因区段)。在一些实施方案中,本公开提供了表达人类免疫球蛋白λ轻链可变结构域和人类免疫球蛋白重链可变结构域的基因修饰非人类动物,其中非人类动物具有有限人类λ轻链可变谱系。还提供了一种用于产生人类λ轻链可变结构域的生物系统,所述人类λ轻链可变结构域由有限人类λ轻链可变区谱系表达,与亲和力成熟的人类重链可变结构域的多种谱系缔合。提供了用于制造包含人类免疫球蛋白可变结构域的抗原结合蛋白(例如抗体、重链、轻链(例如λ轻链)、重链可变结构域、轻链可变结构域(例如λ轻链可变结构域)、单链可变片段(scfv))的方法。在一些实施方案中,一种方法包括用所关注抗原对本文所述的非人类动物进行免疫接种。在一些实施方案中,一种方法包括在抗原结合蛋白(例如特异性结合所关注抗原的结合蛋白)中采用本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)的免疫球蛋白可变区基因序列。方法包括用于制造适用于制造多特异性抗原结合蛋白的人类免疫球蛋白重链和/或λ轻链可变结构域的方法。
[0267]
提供了基因工程非人类动物(例如啮齿动物,例如大鼠或小鼠),所述基因工程非人类动物由人类λ轻链可变区基因区段的有限谱系表达人类λ轻链可变结构域的有限谱系。在一些实施方案中,本文所述的非人类动物被基因工程化成包括单一重排人类λ轻链可变区序列(vλ/jλ序列)。在一些实施方案中,本文所述的非人类动物被基因工程化成包括仅仅一个或两个人类未重排λ轻链可变区基因区段。在一些实施方案中,本文所述的非人类动物被基因工程化成包括仅仅一个或两个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段。在某些实施方案中,本文所述的非人类动物包括四个或五个未重排人类jλ基因区段。由本文所述的非人类动物表达的重排人类λ轻链可变结构域能够与由这种非人类动物表达的多个亲和力成熟的人类重链配对,其中所述多个重链可变区能够特异性结合不同表位。在一些实施方案中,如本文所述的非人类动物表达并选择源自于未重排人类重链可变区基因区段的谱系的合适的亲和力成熟的人类免疫球蛋白重链可变结构域,其中所述亲和力成熟的人类重链可变结构域与源自于非人类动物的有限人类免疫球蛋白λ轻链可变区基因谱系的人类λ轻链可变结构域缔合并一起表达。本公开洞察到在多特异性抗体、尤其双特异性抗体的产生中可利用人类λ轻链可变结构域和人类重链可变结构域(与编码的人类λ轻链可变区和人类重链可变区一起)。
[0268]
非人类动物中的抗体谱系
[0269]
免疫球蛋白(又称为抗体)是大的(约150kd)y形糖蛋白,由宿主免疫系统的b细胞产生,用于压制病原体(例如病毒、细菌等)。每种免疫球蛋白(ig)由两条一致的重链和两条一致的轻链构成,各具有两个结构组分
‑
可变结构域和恒定结构域。在由不同b细胞产生的抗体中重链和轻链可变区是不同的,但对由单个b细胞或b细胞克隆产生的所有抗体来说是一样的。每种抗体的重链和轻链可变区一起构成抗原结合区(或抗原结合位点)。免疫球蛋
白可呈不同种类存在,所述种类基于其所含有的重链恒定区(或结构域)而被称为同种型或类别。在相同同种型的所有抗体中重链恒定区均一致,在不同同种型的抗体中则不同。下表概述了小鼠和人类中的九种抗体同种型。
[0270][0271]
已经在其它物种中鉴定出额外的同种型。由于不同同种型中不同的结构特性,同种型赋予抗体专门生物特性,并在动物体内不同位置(细胞、组织等)发现。最初,b细胞产生具有一致抗原结合区的igm和igd。活化后,通过称为类别转换的方法b细胞转换成不同同种型,所述方法涉及改变由b细胞产生的抗体的恒定区,而可变区保持相同,从而保存原始抗体的抗原特异性(b细胞)。
[0272]
两个独立基因座(igκ和igλ)含有在重排后编码抗体轻链并展现等位基因和同种型排斥的基因区段。κ
+
与λ
+
b细胞的表达比率在物种当中有所变化。举例来说,人类显示约60:40的比率(κ:λ)。在小鼠和大鼠中,观察到95:5的比率(κ:λ)。有趣地,在猫中观察到的κ:λ比率(5:95)与小鼠和大鼠中相反。已经进行了几项研究以解释在这些观察到的比率背后的可能原因,并且已经提出基因座的复杂性(即基因区段、尤其是v基因区段的数目)和基因区段重排效率是基本原因。人类免疫球蛋白λ轻链基因座延伸超过1,000kb并含有大约70个vλ基因区段(29至33个功能性)和七个jλ
‑
cλ基因区段对(四至五个功能性),组织成三簇(参见例如美国专利no.9,006,511的图1,其以引用的方式整体并入本文中)。在表达的抗体谱系中大部分观察到的vλ区域由最近端的簇(称为a簇)内所含的基因区段编码。小鼠免疫球蛋白λ轻链基因座明显不同于人类基因座,且取决于品系,仅仅含有几个vλ和jλ基因区段,组织成两个不同的基因簇(参见例如美国专利no.9,006,511的图2,其以引用的方式整体并入本文中)。
[0273]
用于治疗各种人类疾病的治疗性抗体的发展基本上以建立工程化非人类动物品系、尤其是工程化啮齿动物品系为主题,所述品系在其与人类免疫球蛋白基因相对应的基因组中具有变化量的遗传物质(例如br
ü
ggemann,m.等人,2015,arch.immunol.ther.exp.63:101
‑
8中所评述,其以引用的方式整体并入本文中)。建立这种基因工程啮齿动物的初始工作集中在整合可独立支撑基因区段重组和产生完全人类的重链和/或轻链的人类免疫球蛋白基因座部分,同时使内源免疫球蛋白基因座失活(参见例如br
ü
ggemann,m.等人,1989,proc.nat.acad.sci.u.s.a.86:67
‑
09
‑
13;br
ü
ggemann,m.等人,1991,eur.j.immunol.21:1323
‑
6;taylor,l.d.等人,1992,nucl.acids res.20:6287
‑
6295;davies,n.p.等人,1993,biotechnol.11:911
‑
4;green,l.l.等人,1994,
nat.genet.7:13
‑
21;lonberg,n.等人,1994,nature 368:856
‑
9;taylor,l.d.等人,1994,int.immunol.6:579
‑
91;wagner,s.d.等人,1994,eur.j.immunol.24:2672
‑
81;fishwild,d.m.等人,1996,nat.biotechnol.14:845
‑
51;wagner,s.d.等人,1996,genomics 35:405
‑
14;mendez,m.j.等人,1997,nat.genet.15:146
‑
56;green,l.l.等人,1998,j.exp.med.188:483
‑
95;xian,j.等人,1998,transgenics 2:333
‑
43;little,m.等人,2000,immunol.today 21:364
‑
70;kellermann,s.a.和l.l.green,2002,cur.opin.biotechnol.13:593
‑
7,各以引用的方式整体并入本文中)。具体地说,一些工作包括整合人类免疫球蛋白λ轻链序列(参见例如美国专利申请公布no.2002/0088016a1、2003/0217373a1和2011/0236378a1;美国专利no.6,998,514和7,435,871;nicholson,i.c.等人,1999,j.immunol.163:6898
‑
906;popov,a.v等人,1999,j.exp.med.189(10):1611
‑
19,各以引用的方式整体并入本文中)。这类工作集中在随机整合含有人类vλ、jλ和cλ序列的酵母人造染色体,从而建立表达完全人类免疫球蛋白λ轻链(即人类vλ和cλ结构域)的小鼠品系。更多近来的工作已采用类似的策略,使用同样含有人类vλ、jλ和cλ序列的构建体(osborn,m.j.等人,2013,j.immunol.190:1481
‑
90;lee,e
‑
c.等人,2014,nat.biotech.32(4):356
‑
63,各以引用的方式整体并入本文中)。
[0274]
其它工作包括人类vλ和jλ基因区段特异性插入内源啮齿动物免疫球蛋白轻链基因座(κ和λ)中以使得所述人类vλ和jλ基因区段可操作地连接至内源免疫球蛋白轻链恒定区基因(参见例如美国专利no.9,006,511、9,012,717、9,029,628、9,035,128、9,066,502、9,150,662和9,163,092;均以引用的方式整体并入本文中)。在这种动物的一些实施方案中,来自a簇和b簇的所有人类vλ基因区段和一或四个人类jλ基因区段插入内源免疫球蛋白κ和免疫球蛋白λ轻链基因座中。几种不同的人类vλ和jλ基因区段显示在两种工程化啮齿动物免疫球蛋白轻链基因座处适当重排以形成在啮齿动物抗体谱系中表达的功能性轻链,在内源cκ和cλ区域情况下所述轻链包括人类vλ结构域(参见例如美国专利no.9,006,511的表7和图11
‑
13,其以引用的方式整体并入本文中)。具体地说,具有含人类vλ和jλ基因区段的工程化免疫球蛋白κ轻链基因座的小鼠显示脾室中约1:1的人类λ与内源λ比率(如通过igcκ与igcλ比率所测量)(参见例如美国专利no.9,006,511的表4,其以引用的方式整体并入本文中)。实际上,两种工程化小鼠品系(即工程化免疫球蛋白κ或工程化免疫球蛋白λ轻链基因座)均显示人类vλ结构域可以从啮齿动物中的内源免疫球蛋白轻链基因座表达,内源免疫球蛋白轻链基因座通常对轻链表达展现出极大偏好(参见上文)。本公开认识到可产生交替的工程化免疫球蛋白轻链基因座结构以最大化人类λ轻链可变结构域从有限人类λ轻链可变区谱系的表达。这种交替的工程化免疫球蛋白轻链基因座结构提供了由其设计产生独特抗体谱系的能力。
[0275]
本公开提供了一种非人类动物,所述非人类动物的种系基因组含有工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区。单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。在一些实施方案中,由基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至非人类或人类免疫球蛋白λ或免疫球蛋白κ
轻链恒定区基因的单一重排人类免疫球蛋白λ轻链可变区。在一些实施方案中,这种轻链的表达可通过将所述单一重排人类免疫球蛋白λ轻链可变区插入内源免疫球蛋白κ轻链基因座(或等位基因)中来实现。在一些实施方案中,所提供的非人类动物被工程化,使得内源免疫球蛋白λ轻链可变区的表达失活(例如通过基因缺失)。在一些实施方案中,所提供的非人类动物被工程化,使得内源免疫球蛋白κ轻链可变区的表达失活(例如通过插入、替代或取代)。
[0276]
通用轻链
[0277]
多个问题已经阻碍了先前制造有用的多特异性抗原结合蛋白、例如双特异性抗体的工作,所述问题常共享一共同范例:体外选择或操纵序列以合理地进行工程化,或通过试误法(trial
‑
and
‑
error)对适用于异源二聚体双特异性人类免疫球蛋白配对的格式进行工程化。不幸地,如果不是全部的话,至少大多数体外工程化途径提供了适合于(如果有的话)个别分子的基本上专门方位。
[0278]
已经研发出了采用复杂的生物体选择能够产生人类治疗剂的适当配对的体内方法(参见例如美国专利no.10,143,186,其以引用的方式整体并入)。然而,先前尚未产生能够产生稳固免疫反应,包括产生充足滴度水平的具有人类免疫球蛋白λ轻链可变结构域的高亲和力通用λ轻链的非人类动物。一般地,天然非人类序列常常并非人类治疗性序列的优良来源。至少出于此原因,产生与通用人类轻链配对的非人类免疫球蛋白重链可变结构域具有有限的实用性。更多的体外工程化工作将花费在试误法中,以试图将非人类重链可变序列人源化,同时希望保持表位特异性和亲和力以及与共同人类轻链偶合的能力,结果不确定。在这种方法结束时,最终产物可能维持一些特异性和亲和力,并与通用轻链缔合,但在人类中的免疫原性最终将可能仍然有着极度的风险。
[0279]
因此,适合制造人类治疗剂的非人类动物包括适当大的人类重链可变区基因区段谱系代替内源非人类重链可变区基因区段。人类重链可变区基因区段应能够重排和剪接至内源非人类重链恒定区以形成反向嵌合重链(即包含人类可变结构域和非人类恒定结构域的重链)。重链基因座应能够进行类别转换和体细胞超突变以使得适当大的重链可变结构域谱系可用于非人类动物,以选择可与由有限人类λ轻链可变区谱系编码的人类λ轻链可变结构域缔合的重链可变结构域谱系。
[0280]
为多个重链选择通用轻链的非人类动物具有实用性。在多个实施方案中,在只能表达通用轻链的非人类动物中表达的抗体将具有能够与相同或基本上相同的轻链缔合并一起表达的重链。这特别可用于制造双特异性抗体。举例来说,这种非人类动物可用第一免疫原进行免疫接种,以产生表达特异性地结合第一表位的抗体的b细胞。非人类动物(或包括相同的对重链和λ轻链基因座的基因修饰的非人类动物)可用第二免疫原进行免疫接种,以产生表达特异性地结合第二表位的抗体的b细胞。可以从b细胞克隆重链可变区并与相同重链恒定区和相同轻链一起表达,且在细胞中表达以制造双特异性抗体,其中已经通过非人类动物选择双特异性抗体的轻链组分以与轻链组分缔合和一起表达。已经研发出表达通用κ轻链的非人类动物(参见例如美国专利no.10,143,186,其以引用的方式整体并入)。然而,仍然需要研发能够表达通用λ轻链,包括人类λ轻链可变结构域的非人类动物。
[0281]
本公开提供了工程化的非人类动物(例如啮齿动物,例如大鼠或小鼠),用于产生将适合与相当不同的家族的重链,包括可变区背离种系序列,例如亲和力成熟或进行体细
胞超突变的可变区的重链配对的免疫球蛋白λ轻链。在多个实施方案中,如本文所述的非人类动物被工程化成表达人类λ轻链可变结构域并将人类λ轻链可变结构域与包含体细胞突变的人类重链可变结构域配对,因此能够提供了一种获得适用作人类治疗剂的高亲和力结合蛋白(例如抗体)的途径。
[0282]
通过在生物体内进行长期和复杂的抗体选择过程,如本文所述的基因工程非人类动物(例如啮齿动物,例如大鼠或小鼠)在一系列不同的人类重链可变结构域与有限数目的轻链选项配对上进行生物学上适当的选择。为了实现这一点,非人类动物被工程化成呈现有限数目的人类λ轻链可变结构域选项,与多种人类重链可变结构域选项一起。在用免疫原激发后,本文所述的非人类动物可最大化其谱系中解决方案的数目以针对免疫原形成抗体,基本上或仅仅受其谱系中数目或轻链选项限制。在多个实施方案中,这包括允许非人类动物实现轻链可变结构域合适和相容的体细胞突变,尽管如此,这些突变仍与相对大量的人类重链可变结构域,尤其包括体细胞超突变的人类重链可变结构域相容。
[0283]
为了实现有限人类λ轻链选项谱系,如本文所述的非人类动物可被工程化成使其不能或基本上不能制造或重排天然非人类λ和/或κ轻链可变结构域。在一些实施方案中,这可以例如通过非人类动物的λ和/或κ轻链可变区基因区段缺失来实现。在一些实施方案中,然后内源非人类基因座可以用可操作地连接至内源非人类轻链恒定结构域的适合精选外源人类λ轻链可变区序列修饰。在一些实施方案中,外源人类可变区基因区段未重排(例如两个vλ基因区段和一个或多个jλ基因区段)并且可重排和剪接至内源非人类轻链恒定区基因以形成重排的反向嵌合轻链基因(人类可变、非人类恒定)。在一些实施方案中,外源人类可变基因区段进行重排(例如一个vλ基因区段和一个jλ基因区段)并且可以剪接至内源非人类轻链恒定区基因以形成包含人类可变区和非人类恒定区的反向嵌合轻链基因。在一些实施方案中,外源人类可变基因区段进行重排(例如一个vλ基因区段和一个jλ基因区段)并且可剪接至外源人类轻链恒定区基因以形成包含人类可变区和人类恒定区的人源化轻链基因。在多个实施方案中,轻链可变区能够进行体细胞超突变。在多个实施方案中,适当增强子保留在非人类动物中。已经报告增强子最大化轻链可变区获得体细胞突变的能力。举例来说,在用人类λ可变区基因区段替代内源非人类动物κ可变区基因区段来修饰非人类动物的κ基因座中,非人类κ内含子增强子和非人类κ3'增强子在功能上得以维持,或未被破坏。虽然本公开涵盖去除或破坏增强子的实施方案,但预计这种实施方案将减少或消除体细胞超突变。在这种实施方案中,轻链可变区的体细胞超突变相对于例如包含一个或多个内源非人类增强子的轻链可变区减少。
[0284]
提供了一种基因工程非人类动物,所述非人类动物表达与多种反向嵌合(人类可变、非人类恒定)重链缔合的有限反向嵌合(人类可变、非人类恒定)轻链谱系。在多个实施方案中,内源非人类κ轻链可变区基因区段缺失并且经单个(或两个)人类λ轻链可变区基因区段替代,可操作地连接至内源非人类κ恒定区基因。在一些实施方案中,维持非人类κ内含子增强子和非人类κ3'增强子。不限于任一理论,增强子可尤其最大化人类λ轻链可变区基因区段的体细胞超突变。在多个实施方案中,非人类动物还包含非功能性λ轻链基因座或其缺失,或使基因座不能制造λ轻链的缺失。
[0285]
提供了一种基因工程非人类动物,在多个实施方案中,所述非人类动物包含缺乏内源非人类轻链可变基因区段且包含可操作地连接至非人类cλ基因区段的人类可变基因
区段,在一些实施方案中,重排人类免疫球蛋白λ轻链可变区的轻链可变区基因座,其中基因座能够进行体细胞超突变,并且其中基因座表达包含连接至非人类cλ基因区段的人类免疫球蛋白λ轻链可变区的轻链。因此,在多个实施方案中,基因座包含与正常或野生型水平的体细胞超突变相关的非人类κ3'增强子。
[0286]
在多个实施方案中,基因工程非人类动物在用所关注抗原进行免疫接种时产生了展现与一个或与两个重排的轻链一起表达和发挥功能的人类免疫球蛋白重链可变区的多种重排的b细胞。在一些实施方案中,人类λ轻链可变区包含体细胞超突变。在一些实施方案中,人类λ轻链可变区各包含1至5个体细胞超突变。在多个实施方案中,由本文所述的非人类动物表达的轻链能够与包括在非人类动物中表达的人类免疫球蛋白重链可变区的任何重链缔合并一起表达。
[0287]
所提供的非人类动物、细胞和组织
[0288]
提供了表达(例如其b细胞表达)抗体的非人类动物,所述抗体含有在非人类动物的种系基因组中的内源非人类λ轻链基因座处非人类免疫球蛋白κ轻链可变区序列的位置中包括源自于以下的人类λ轻链可变结构域的轻链:(i)一个或两个未重排vλ基因区段和一个或多个未重排jλ基因区段,或(ii)单一重排人类λ轻链可变区。应了解,在本文所述的各种实施方案中,基因修饰非人类动物是啮齿动物,例如大鼠或小鼠,并且本文所述的非人类元件(增强子、恒定区等)是啮齿动物,例如大鼠或小鼠元件。本文所述的非人类动物的合适实例包括但不限于啮齿动物,例如大鼠或小鼠,尤其小鼠。
[0289]
本公开提供了改良的体内系统,所述系统用于鉴定和研发新抗原结合蛋白、抗体、抗体组分(例如抗原结合部分和/或包括其的组合物或格式),和/或可用于例如治疗多种影响人类的疾病的抗体类治疗剂。此外,本公开涵盖以下认识:具有工程化免疫球蛋白基因座,例如包括有限λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的非人类动物(例如啮齿动物,例如大鼠或小鼠)是适用的。在一些实施方案中,本文所述的非人类动物提供了改良的体内系统,所述系统用于研发施用于人类的抗体和/或抗体类治疗剂。在一些实施方案中,本文所述的非人类动物提供了改良的体内系统,所述系统用于研发含有人类λ轻链可变结构域的抗体和/或抗体类治疗剂,所述抗体和/或抗体类治疗剂的特征在于与从含有人类vλ区序列的现有的体内系统获得的抗体和/或抗体类治疗剂相比,具有改善和/或不同的性能(例如抗原特异性抗体谱系的表达和/或表示)。
[0290]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括有限人类λ轻链可变区谱系的内源免疫球蛋白κ轻链基因座。在一些实施方案中,有限人类λ轻链可变区谱系的序列可操作地连接至非人类轻链恒定区。在一些实施方案中,非人类轻链恒定区是啮齿动物(例如小鼠或大鼠)轻链恒定区。在一些实施方案中,非人类轻链恒定区是κ或λ轻链恒定区。在一些实施方案中,有限人类λ轻链可变区谱系的序列可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ。在一些实施方案中,有限人类λ轻链可变区谱系的序列可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ(例如cλ1)。在一些实施方案中,非人类λ轻链恒定区(例如小鼠cλ,例如小鼠cλ1)代替内源非人类cκ。
[0291]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例
如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括两个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,一个或多个未重排人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个或多个未重排人类jλ基因区段包括jλ1、jλ2、jλ3和jλ7。在一些实施方案中,一个或多个未重排人类jλ基因区段包括jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,两个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段可操作地连接至非人类轻链恒定区。在一些实施方案中,非人类轻链恒定区是啮齿动物(例如小鼠或大鼠)轻链恒定区。在一些实施方案中,非人类轻链恒定区是κ或λ轻链恒定区。在一些实施方案中,两个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ。在一些实施方案中,两个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ(例如cλ1)。在一些实施方案中,非人类λ轻链恒定区(例如啮齿动物,例如大鼠或小鼠,例如小鼠cλ,例如小鼠cλ1)代替内源非人类(例如啮齿动物,例如大鼠或小鼠)cκ。
[0292]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ的两个未重排人类vλ基因区段和四个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。
[0293]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ的两个未重排人类vλ基因区段和五个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。
[0294]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ(例如cλ1)
的两个未重排人类vλ基因区段和四个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。
[0295]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ(例如cλ1)的两个未重排人类vλ基因区段和五个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,两个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。
[0296]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括一个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,一个或多个未重排人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个或多个未重排人类jλ基因区段包括jλ1、jλ2、jλ3和jλ7。在一些实施方案中,一个或多个未重排人类jλ基因区段包括jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段可操作地连接至非人类轻链恒定区。在一些实施方案中,非人类轻链恒定区是啮齿动物(例如小鼠或大鼠)轻链恒定区。在一些实施方案中,非人类轻链恒定区是κ或λ轻链恒定区。在一些实施方案中,一个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ。在一些实施方案中,一个未重排人类vλ基因区段和一个或多个未重排人类jλ基因区段可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ(例如cλ1)。在一些实施方案中,非人类λ轻链恒定区(例如啮齿动物,例如大鼠或小鼠,例如小鼠cλ,例如小鼠cλ1)代替内源非人类(例如啮齿动物,例如大鼠或小鼠)cκ。
[0297]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ的一个未重排人类vλ基因区段和四个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ
1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ1
‑
51,且四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ2
‑
14,且四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。
[0298]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ的一个未重排人类vλ基因区段和五个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ1
‑
51,且五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ2
‑
14,且五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。
[0299]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ的一个未重排人类vλ基因区段和四个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ1
‑
51,且四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ2
‑
14,且四个未重排人类jλ基因区段是jλ1、jλ2、jλ3和jλ7。
[0300]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ的一个未重排人类vλ基因区段和五个未重排人类jλ基因区段的内源免疫球蛋白κ轻链基因座。在一些实施方案中,一个未重排人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ1
‑
51,且五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,一个未重排人类vλ基因区段是vλ2
‑
14,且五个未重排人类jλ基因区段是jλ1、jλ2、jλ3、jλ6和jλ7。
[0301]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括包含人类vλ基因区段和人类jλ基因区段的单一重排人类λ轻链可变区(v/j)的内源免疫球蛋白κ轻链基因座。在一些实施方案中,单一重排人类λ轻链可变区的人类vλ基因区段选自由以下组成的组:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,单一重排人类λ轻链可变区的人类vλ基因区
段选自由以下组成的组:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,单一重排人类λ轻链可变区的人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,单一重排人类λ轻链可变区的人类vλ基因区段是vλ1
‑
51。在一些实施方案中,单一重排人类λ轻链可变区的人类vλ基因区段是vλ2
‑
14。在一些实施方案中,单一重排人类λ轻链可变区的人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,单一重排人类λ轻链可变区的人类jλ基因区段是jλ1。在一些实施方案中,单一重排人类λ轻链可变区的人类jλ基因区段是jλ2。在一些实施方案中,单一重排人类λ轻链可变区的人类jλ基因区段是jλ3。在一些实施方案中,单一重排人类λ轻链可变区可操作地连接至非人类轻链恒定区。在一些实施方案中,非人类轻链恒定区是啮齿动物(例如小鼠或大鼠)轻链恒定区。在一些实施方案中,非人类轻链恒定区是κ或λ轻链恒定区。在一些实施方案中,单一重排人类λ轻链可变区可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cκ。在一些实施方案中,单一重排人类λ轻链可变区可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ(例如cλ1)。在一些实施方案中,非人类λ轻链恒定区(例如小鼠cλ,例如小鼠cλ1)代替内源非人类(例如啮齿动物,例如大鼠或小鼠)cκ。
[0302]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至小鼠cκ的单一重排人类λ轻链可变区的内源免疫球蛋白κ轻链基因座,其中单一重排人类λ轻链可变区包括人类vλ基因区段和人类jλ基因区段。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,人类vλ基因区段是vλ1
‑
51且人类jλ基因区段是jλ2。在一些实施方案中,人类vλ基因区段是vλ2
‑
14且人类jλ基因区段是jλ2。
[0303]
本公开尤其提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其具有被工程化成包括可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)cλ的单一重排人类λ轻链可变区的内源免疫球蛋白κ轻链基因座,其中单一重排人类λ轻链可变区包括人类vλ基因区段和人类jλ基因区段。在一些实施方案中,人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,人类vλ基因区段是vλ1
‑
51且人类jλ基因区段是jλ2。在一些实施方案中,人类vλ基因区段是vλ2
‑
14且人类jλ基因区段是jλ2。
[0304]
本公开尤其提供了一种小鼠、小鼠细胞或小鼠组织,其具有被工程化成包括可操作地连接至小鼠cλ的单一重排人类λ轻链可变区的内源免疫球蛋白κ轻链基因座,其中单一重排人类λ轻链可变区包括人类vλ基因区段和人类jλ基因区段。在一些实施方案中,人类vλ基因区段包含vλ1
‑
51。在一些实施方案中,人类vλ基因区段包含vλ2
‑
14。在一些实施方案中,人类jλ基因区段包含jλ2。在一些实施方案中,人类vλ基因区段是vλ1
‑
51且人类jλ基因区段是jλ2。在一些实施方案中,人类vλ基因区段是vλ2
‑
14且人类jλ基因区段是jλ2。
[0305]
本公开尤其提供了一种基因修饰小鼠、小鼠细胞或小鼠组织,其种系基因组包含工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至小鼠cλ1基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ1
‑
51基因区段和人类jλ2基因区段,其中由所述基因修饰小鼠的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0306]
本公开尤其提供了一种基因修饰小鼠、小鼠细胞或小鼠组织,其种系基因组包含工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至小鼠cλ1基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ2
‑
14基因区段和人类jλ2基因区段,其中由所述基因修饰小鼠的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0307]
在一些实施方案中,所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)的特征在于从所述非人类动物的种系基因组中的内源免疫球蛋白κ轻链基因座表达抗体,所述抗体含有(1)人类vλ结构域和(2)非人类cλ结构域。在一些实施方案中,所提供的非人类动物的特征在于与一个或多个参考工程化非人类动物相比,从包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座使用人类vλ区的使用率提高(例如但不限于约2倍)。
[0308]
在一些实施方案中,提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其种系基因组包括包含以下的内源免疫球蛋白κ轻链基因座:(a)单一重排人类免疫球蛋白λ轻链可变区,和(b)cλ基因,其中(a)可操作地连接至(b),并且其中非人类动物在内源免疫球蛋白κ轻链基因座处缺乏非人类动物cκ基因。
[0309]
在一些实施方案中,提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,其基因组包括包含单一重排人类免疫球蛋白λ轻链可变区和cλ基因插入的内源免疫球蛋白κ轻链基因座,所述单一重排人类免疫球蛋白λ轻链可变区可操作地连接至所述cλ基因,并且所述cλ基因插入在内源免疫球蛋白κ轻链基因座处非人类cκ基因的位置中。在非人类动物、非人类细胞或非人类组织的许多实施方案中,插入在内源免疫球蛋白κ轻链基因座处非人类cκ基因的位置中的cλ基因是非人类或人类cλ基因。在一些实施方案中,非人类cλ基因是或包含选自由以下组成的组的哺乳动物cλ基因:灵长类动物、山羊、绵羊、猪、犬、母牛或啮齿动物(例如大鼠或小鼠)cλ基因。
[0310]
在一些实施方案中,非人类cλ基因是或包含啮齿动物cλ基因。
[0311]
在一些实施方案中,啮齿动物cλ基因是或包含小鼠cλ基因。在一些实施方案中,小鼠cλ基因包含与选自由小鼠cλ1、小鼠cλ2和小鼠cλ3组成的群的小鼠cλ基因至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,小鼠cλ基因包含与选自由小鼠cλ1、小鼠cλ2和小鼠cλ3组成的群的小鼠cλ基因基本上同一或同一的序列。在一些实施方案中,小鼠cλ1基因是或包含seq id no:1。在某些实施方案中,小鼠cλ2基因是或包含seq id no:2。在某些实施方案中,小鼠cλ3基因是或包含seq id no:3。在某些实施方案中,小鼠cλ基因包含与小鼠cλ1基因同一的序列。
[0312]
在一些实施方案中,小鼠cλ基因包含与选自由小鼠cλ1、小鼠cλ2和小鼠cλ3组成的群的小鼠cλ基因80%至100%、85%至100%、90%至100%、95%至100%或98%至100%同一的序列。在一些实施方案中,小鼠cλ基因包含与选自由小鼠cλ1、小鼠cλ2和小鼠cλ3组成的群的小鼠cλ基因80%至98%、80%至95%、80%至90%或80%至85%同一的序列。在一些实施方案中,小鼠cλ基因包含与选自由小鼠cλ1、小鼠cλ2和小鼠cλ3组成的群的小鼠cλ基因85%至98%、90%至95%或88%至93%同一的序列。
[0313]
在一些实施方案中,啮齿动物cλ基因是或包含大鼠cλ基因。在一些实施方案中,大鼠cλ基因包含与选自由大鼠cλ1、大鼠cλ2、大鼠cλ3和大鼠cλ4基因组成的群的大鼠cλ基因至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,大鼠cλ基因包含与选自由大鼠cλ1、大鼠cλ2、大鼠cλ3和大鼠cλ4基因组成的群的大鼠cλ基因基本上同一或同一的序列。在某些实施方案中,大鼠cλ基因是或包含seq id no:7。在某些实施方案中,大鼠cλ2基因是或包含seq id no:8。在某些实施方案中,大鼠cλ3基因是或包含seq id no:9。在某些实施方案中,大鼠cλ4基因是或包含seq id no:10。
[0314]
在一些实施方案中,大鼠cλ基因包含与选自由大鼠cλ1、大鼠cλ2、大鼠cλ3和大鼠cλ4基因组成的群的大鼠cλ基因80%至100%、85%至100%、90%至100%、95%至100%或98%至100%同一的序列。在一些实施方案中,大鼠cλ基因包含与选自由大鼠cλ1、大鼠cλ2、大鼠cλ3和大鼠cλ4基因组成的群的大鼠cλ基因80%至98%、80%至95%、80%至90%或80%至85%同一的序列。在一些实施方案中,大鼠cλ基因包含与选自由大鼠cλ1、大鼠cλ2、大鼠cλ3和大鼠cλ4基因组成的群的大鼠cλ基因85%至98%、90%至95%或88%至93%同一的序列。
[0315]
在一些实施方案中,人类cλ基因包含与选自由人类cλ1、人类cλ2、人类cλ3、人类cλ6和人类cλ7基因组成的群的人类cλ基因至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,人类cλ基因包含与选自由人类cλ1、人类cλ2、人类cλ3、人类cλ6和人类cλ7基因组成的群的人类cλ基因基本上同一或同一的序列。在一些实施方案中,人类cλ基因包含与选自由人类cλ1、人类cλ2、人类cλ3、人类cλ6和人类cλ7基因组成的群的人类cλ基因同一的序列。在某些实施方案中,人类cλ1基因是或包含seq id no:15。在某些实施方案中,人类cλ2基因是或包含seq id no:16。在某些实施方案中,人类cλ3基因是或包含seq id no:17。在某些实施方案中,人类cλ6基因是或包含seq id no:18。在某些实施方案中,人类cλ7基因是或包含seq id no:18。在某些实施方案中,人类cλ基因是或包含人类cλ2基因。
[0316]
在一些实施方案中,人类cλ基因包含与选自由人类cλ1、人类cλ2、人类cλ3、人类cλ6和人类cλ7基因组成的群的人类cλ基因80%至100%、85%至100%、90%至100%、95%至100%或98%至100%同一的序列。在一些实施方案中,人类cλ基因包含与选自由人类cλ1、人类cλ2、人类cλ3、人类cλ6和人类cλ7基因组成的群的人类cλ基因80%至98%、80%至95%、80%至90%或80%至85%同一的序列。在一些实施方案中,人类cλ基因包含与选自由人类cλ1、人类cλ2、人类cλ3、人类cλ6和人类cλ7基因组成的群的人类cλ基因85%至98%、90%至95%或88%至93%同一的序列。
[0317]
在所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织的一些实施方案
中,所述非人类动物、非人类细胞或非人类组织的种系基因组还包含被修饰成包含一个或多个人类v
h
基因区段、一个或多个人类d
h
基因区段和一个或多个人类j
h
基因区段的内源免疫球蛋白重链基因座,所述人类v
h
、d
h
和j
h
基因区段在内源免疫球蛋白重链基因座处可操作地连接至非人类免疫球蛋白重链恒定区(参见例如macdonald,l.e.等人,“precise and in situ genetic humanization of 6 mb of mouse immunoglobulin genes,”proc.natl.acad.sci.u.s.a,111(14):5147
‑
5152(2014年4月8日);美国专利no.6,596,541、8,642,835、8,697,940和8,791,323,各以引用的方式整体并入本文中)。
[0318]
在一些实施方案中,一个或多个人类v
h
基因区段、一个或多个人类d
h
基因区段和一个或多个人类j
h
基因区段的插入整体或部分地代替或替代非人类v
h
、d
h
和j
h
基因区段(例如在位置上非人类v
h
、d
h
和j
h
基因区段的编码序列经人类v
h
、d
h
和j
h
基因区段的编码序列替代或取代)。在一些实施方案中,非人类免疫球蛋白重链恒定区是或包含内源非人类免疫球蛋白重链恒定区。在许多实施方案中,非人类免疫球蛋白重链恒定区(例如内源)包括一个或多个非人类免疫球蛋白重链恒定区基因或基因区段(例如igm、igd、igg、ige、iga等)。在某些实施方案中,插入包括天然出现在人类v
h
、d
h
和j
h
基因区段之间的人类非编码dna以及其组合。在某些实施方案中,如本文所述的免疫球蛋白重链基因座包含人类v
h
基因区段v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合,人类d
h
基因区段d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合,以及人类j
h
基因区段j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合的插入。在某些实施方案中,插入包括天然出现在内源重链基因座中与人类v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2或v
h6‑
1相邻处的人类非编码dna,天然出现在与人类d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26或d
h7‑
27相邻处的人类非编码dna,以及天然出现在内源重链基因座中与人类j
h
1、j
h
2、j
h
3、j
h
4、j
h
5或j
h
6相邻处的人类非编码dna。
[0319]
在一些实施方案中,本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织在其基因组(例如其种系基因组)中包括adam6基因,所述adam6基因编码adam6多肽、其功能直系同源物、功能同源物或功能片段(参见例如美国专利no.8,642,835和8,697,940,各以引用的方式整体并入本文中)。在一些实施方案中,本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织在其基因组(例如其种系基因组)中包括啮齿动物(例如小鼠或大鼠)adam6基因,所述adam6基因编码啮齿动物(例如小鼠或大鼠)adam6多肽、其功能直系同源物、功能同源物或功能片段(参见例如美国专利no.8,642,835和8,697,940,各以引用的
方式整体并入本文中)。在一些实施方案中,adam6多肽、其功能直系同源物、功能同源物或功能片段从adam6基因表达。在一些实施方案中,如本文所述的基因修饰非人类动物中的adam6基因并非源自于所述特定非人类动物(例如包括大鼠adam6基因或从另一品系小鼠获得的小鼠adam6基因的小鼠)。在一些实施方案中,本文所述的非人类动物包括异位adam6基因。如本文所用,“异位”adam6基因是指处于与adam6基因出现在野生型非人类动物中不同的背景下的adam6基因。举例来说,adam6基因可位于不同染色体上,位于不同基因座上,或位于不同序列相邻处。一种示例性异位adam6基因是位于人类免疫球蛋白序列(例如人类重链可变区基因区段)内的小鼠adam6基因。在一些实施方案中,本文所述的非人类动物包括插入或整合的adam6基因。
[0320]
在一些实施方案中,本文所述的非人类动物、非人类细胞或非人类组织在其基因组(例如其种系基因组)中包括编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列的插入。
[0321]
在一些实施方案中,本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织在其基因组(例如其种系基因组)中包括编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。在一些实施方案中,本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织在其基因组(例如其种系基因组)中包括小鼠adam6a基因和/或小鼠adam6b基因。在一些实施方案中,本文所述的非人类动物、非人类细胞或非人类组织包括编码小鼠adam6a、其功能直系同源物、功能同源物或功能片段,和/或小鼠adam6b、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
[0322]
在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列插入和/或位于与内源免疫球蛋白重链基因座相同的染色体上。在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列插入和/或位于使编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列与人类免疫球蛋白重链可变区基因区段相连的位置中。在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列插入和/或位于使编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列与人类免疫球蛋白重链可变区基因区段相邻的位置中。在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列插入和/或位于使编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列位于人类免疫球蛋白重链可变区基因区段之间的位置中。在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列插入和/或位于第一与第二人类v
h
基因区段之间。在一些实施方案中,第一人类v
h
基因区段是人类v
h1‑
2且第二人类v
h
基因区段是人类v
h6‑
1。在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列插入和/或位于人类adam6伪基因的位置处。在一些实施方案中,编码一种或多种非人类adam6多肽、其功能直系同源物、功能
同源物或功能片段的一个或多个核苷酸序列插入人类v
h
基因区段与人类d
h
基因区段之间。
[0323]
在一些实施方案中,本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织包括使adam6活性恢复或增强的adam6基因。在一些实施方案中,adam6基因使adam6活性恢复至包括功能性内源adam6基因的可比非人类动物的水平。在一些实施方案中,adam6基因使adam6活性增强至不包括功能性adam6基因的可比非人类动物的adam6活性的至少2倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍或至少10倍的水平。
[0324]
在一些实施方案中,本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织包括当在雄性非人类动物中表达时使生育力恢复或增强的adam6基因。在一些实施方案中,adam6基因使雄性非人类动物中的生育力恢复至包括功能性内源adam6基因的可比非人类动物的水平。在一些实施方案中,adam6基因使雄性非人类动物的生育力恢复,以使得通过所述雄性非人类动物交配所产生的幼仔数目是由不包括功能性adam6基因的可比雄性非人类动物的可比交配产生的幼仔数目的至少70%、至少80%、至少90%、至少95%。在一些实施方案中,adam6基因增强雄性非人类动物的生育力,以使得通过所述雄性非人类动物交配所产生的幼仔数目是由不包括功能性adam6基因的可比雄性非人类动物的可比交配产生的幼仔数目的至少2倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍或至少10倍。
[0325]
在一些实施方案中,如本文所述的非人类免疫球蛋白重链基因座缺乏至少一个内源非人类adam6基因。在一些实施方案中,至少一个内源非人类adam6基因的缺乏降低缺乏内源非人类adam6基因的雄性啮齿动物(例如小鼠或大鼠)的adam6活性和/或生育力。在一些实施方案中,如本文所述的非人类免疫球蛋白重链基因座包括至少一个内源非人类adam6基因的破坏。在一些实施方案中,至少一个内源非人类adam6基因的破坏降低缺乏内源非人类adam6基因的雄性啮齿动物(例如小鼠或大鼠)的adam6活性和/或生育力。
[0326]
在如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织的一些实施方案中,非人类动物、非人类细胞或非人类组织对于如本文所述的包含人类重链可变区基因区段的工程化内源免疫球蛋白重链基因座是纯合或杂合的。
[0327]
在如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织的一些实施方案中,非人类动物、非人类细胞或非人类组织对于如本文所述的包含人类轻链可变基因区段(人类可变λ轻链基因区段)的工程化内源免疫球蛋白κ轻链基因座是纯合或杂合的。
[0328]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织包含第一工程化内源免疫球蛋白κ轻链基因座等位基因,所述第一工程化内源免疫球蛋白κ轻链基因座等位基因包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。在一些实施方案中,非人类动物、非人类细胞或非人类组织包含第二工程化内源免疫球蛋白κ轻链基因座等位基因,所述第二工程化内源免疫球蛋白κ轻链基因座等位
基因包含可操作地连接至啮齿动物cκ基因区段的单一重排人类免疫球蛋白κ轻链可变区,其中单一重排人类免疫球蛋白κ轻链可变区包含人类vκ基因区段和人类jκ基因区段。在一些实施方案中,这种非人类动物或非人类组织可以从第一工程化内源免疫球蛋白κ轻链基因座等位基因表达λ轻链并从第二工程化内源免疫球蛋白κ轻链基因座等位基因表达κ轻链。在一些实施方案中,单一重排人类免疫球蛋白κ轻链可变区包含vκ3
‑
20或vκ1
‑
39且单一重排人类免疫球蛋白λ轻链可变区包含vλ1
‑
51或vλ2
‑
14。在一个实施方案中,单一重排人类免疫球蛋白κ轻链可变区是vκ3
‑
20/jκ1或vκ1
‑
39/jκ5,且单一重排人类免疫球蛋白λ轻链可变区是vλ1
‑
51/jλ2或vλ2
‑
14/jλ2。
[0329]
在所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织的一些实施方案中,内源免疫球蛋白λ轻链基因座整体或部分缺失。在所提供的非人类动物、非人类细胞或非人类组织的一些实施方案中,内源免疫球蛋白λ轻链基因座是功能沉默的或在其它方面无功能的(例如通过基因靶向)。在所提供的非人类动物、非人类细胞或非人类组织的某些实施方案中,非人类动物、非人类细胞或非人类组织对于功能沉默或在其它方面无功能的如本文所述的内源免疫球蛋白λ轻链基因座是纯合的。
[0330]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织不可检测地表达内源免疫球蛋白λ轻链。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织不可检测地表达内源免疫球蛋白κ轻链。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织不可检测地表达内源免疫球蛋白λ轻链或内源免疫球蛋白κ轻链。
[0331]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织不可检测地表达内源免疫球蛋白重链。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织不可检测地表达内源免疫球蛋白λ轻链、内源免疫球蛋白κ轻链和内源免疫球蛋白重链。
[0332]
在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织具有还包含可操作地连接至转录控制元件的编码外源末端脱氧核苷酸转移酶(tdt)的核酸序列的基因组。(参见例如wo 2017/210586和美国公布no.2017/0347633,各以引用的方式整体并入本文中)。
[0333]
在一些实施方案中,转录控制元件包括rag1转录控制元件、rag2转录控制元件、免疫球蛋白重链转录控制元件、免疫球蛋白κ轻链转录控制元件、免疫球蛋白λ轻链转录控制元件或它们的任何组合。
[0334]
在一些实施方案中,编码外源tdt的核酸序列位于免疫球蛋白κ轻链基因座、免疫球蛋白λ轻链基因座、免疫球蛋白重链基因座、rag1基因座或rag2基因座处。
[0335]
在一些实施方案中,tdt是人类tdt。在一些实施方案中,tdt是tdt的短亚型(tdts)。
[0336]
在一些实施方案中,将单一重排人类免疫球蛋白λ轻链可变区以维持靠近插入点的非人类免疫球蛋白κ轻链增强子区域(或增强子序列)(例如非人类免疫球蛋白κ内含子增
强子和/或非人类免疫球蛋白κ3'增强子)的完整性的方式引入内源免疫球蛋白κ轻链基因座中。因此,这种非人类动物具有可操作地连接至人类和非人类免疫球蛋白λ轻链序列(例如人类vλ和jλ基因区段,和非人类cλ或cκ)或可操作地连接至人类免疫球蛋白λ轻链序列(例如人类vλ和jλ基因区段,和人类cλ或cκ)的野生型免疫球蛋白κ轻链增强子区域(或增强子序列)。
[0337]
在一些实施方案中,如本文所述的经一个或多个人类免疫球蛋白λ轻链序列改变、置换、破坏、缺失、替代或工程化的非人类免疫球蛋白κ轻链基因座是鼠类免疫球蛋白κ轻链基因座。在一些实施方案中,如本文所述的一个或多个人类免疫球蛋白λ轻链序列插入所述非人类免疫球蛋白κ轻链基因座的两个拷贝中的非人类免疫球蛋白κ轻链基因座的一个拷贝(即等位基因)中,产生关于人类免疫球蛋白κ轻链序列是杂合的非人类动物。在一些实施方案中,提供了一种对于如本文所述的包括一个或多个人类免疫球蛋白λ轻链序列的免疫球蛋白κ轻链基因座是纯合的非人类动物。
[0338]
在一些实施方案中,内源非人类免疫球蛋白λ轻链基因座的一个或多个内源非人类免疫球蛋白λ轻链序列(或其部分)未缺失。在一些实施方案中,内源非人类免疫球蛋白λ轻链基因座的一个或多个内源非人类免疫球蛋白λ轻链序列(或其部分)缺失。在一些实施方案中,内源非人类免疫球蛋白λ轻链基因座的一个或多个内源非人类免疫球蛋白λ轻链序列(例如v、j和/或c或它们的任何组合)被改变、置换、破坏、缺失或替代以使得所述非人类免疫球蛋白λ轻链基因座是功能沉默的。在一些实施方案中,内源非人类免疫球蛋白λ轻链基因座的一个或多个内源非人类免疫球蛋白λ轻链序列(例如v、j和/或c或它们的任何组合)经靶向载体改变、置换、破坏、缺失或替代以使得所述非人类免疫球蛋白λ轻链基因座是功能失活的(即不能产生在如本文所述的非人类动物的抗体谱系中表达和/或可检测的抗体的功能性轻链)。关于内源非人类免疫球蛋白λ轻链基因座失活的指导提供于例如美国专利no.9,006,511(参见例如图2)中,所述专利以引用的方式整体并入本文中。
[0339]
工程化免疫球蛋白κ轻链基因座或转基因(例如包括如本文所述的有限人类λ轻链可变区谱系)或其表达产物可以使用多种方法检测,包括例如pcr、dna印迹法、限制性片段长度多态性(rflp)、等位基因得失测定法、蛋白质印迹法、facs分析等。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织关于如本文所述的工程化免疫球蛋白κ轻链基因座是杂合的。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织关于如本文所述的工程化免疫球蛋白κ轻链基因座为半合子的。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织含有如本文所述的工程化免疫球蛋白κ轻链基因座或转基因的一个或多个拷贝。在一些实施方案中,如本文所述的非人类动物、非人类细胞或非人类组织含有如图式中所描绘的工程化内源免疫球蛋白κ轻链基因座。
[0340]
本公开认识到,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织在抗体选择和产生机制(例如重组和体细胞超突变)中将利用其基因组中所包括的人类重链和λ轻链可变区基因区段。因而,在多个实施方案中,由本文所述的非人类动物、非人类细胞或非人类组织产生的人类免疫球蛋白人类重链和λ轻链可变结构域分别由其基因组中所包括的人类重链和λ轻链可变区基因区段或其体细胞超突变变体编码。
[0341]
在一些实施方案中,提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠),所
述非人类动物的基因组包含工程化免疫球蛋白κ轻链基因座,其中所述非人类动物包括包含进行体细胞超突变的人类重链可变区序列和/或人类λ轻链可变区序列的b细胞。在一些实施方案中,本公开的非人类动物(例如啮齿动物,例如大鼠或小鼠)的b细胞中存在的人类重链可变区序列和/或人类λ轻链具有1个、2个、3个、4个、5个或更多个体细胞超突变。所属领域的技术人员知道用于鉴定成熟抗体序列中的源基因区段的方法。举例来说,多种工具可用于帮助进行这项分析,例如dnaplot、imgt/v
‑
quest、joinsolver、soda和ab
‑
origin。
[0342]
本公开尤其提供了来自本文所述的非人类动物(例如啮齿动物,例如大鼠、小鼠)的细胞和组织。在一些实施方案中,提供了来自如本文所述的非人类动物的脾细胞(和/或其它淋巴组织)。在一些实施方案中,提供了来自如本文所述的非人类动物的b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的祖b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的前b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的未成熟b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的成熟幼稚b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的激活b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的记忆b细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的b谱系淋巴细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的血浆或血浆细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的干细胞。在一些实施方案中,干细胞是胚胎干细胞。在一些实施方案中,提供了来自如本文所述的非人类动物的生殖细胞。在一些实施方案中,生殖细胞是卵母细胞。在一些实施方案中,生殖细胞是精细胞。在一些实施方案中,来自如本文所述的非人类动物的精细胞表达一种或多种adam6多肽、其功能直系同源物、功能同源物或功能片段。在一些实施方案中,来自如本文所述的非人类动物的任何细胞或组织可以分离。在一些实施方案中,提供了来自如本文所述的非人类动物的分离的细胞和/或分离的组织。在一些实施方案中,提供了一种杂交瘤,其中所述杂交瘤用如本文所述的非人类动物的b细胞制造。在一些实施方案中,杂交瘤用已经用所关注抗原进行免疫接种的非人类动物的b细胞制造。在一些实施方案中,杂交瘤用表达结合(例如特异性结合)于所关注抗原上的表位的抗体的非人类动物的b细胞制造。
[0343]
如本文所述的任何非人类动物(例如啮齿动物,例如大鼠或小鼠)可以用一种或多种所关注抗原进行免疫接种,条件和时间足以使非人类动物显现对所述一种或多种所关注抗原的免疫反应。所属领域的技术人员知道用于对非人类动物进行免疫接种的方法。用于对非人类动物进行免疫接种的一种示例性非限制性方法可见于美国专利no.7,582,298,所述专利以引用的方式整体并入本文中。
[0344]
本公开尤其提供了进行免疫接种的如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠),以及从其分离的细胞和组织。在一些实施方案中,本文所述的非人类动物对用包括一个或多个表位的抗原进行的免疫接种起反应,产生b细胞群体。在一些实施方案中,非人类动物产生表达结合(例如特异性结合)于所关注抗原的一个或多个表位的抗体的b细胞群体。在一些实施方案中,由对抗原起反应而产生的b细胞群体表达的抗体包括具有由人类重链可变区序列编码的人类重链可变结构域的重链和/或具有由如本文所述的人类λ轻链可变区序列编码的人类λ轻链可变结构域的λ轻链。在一些实施方案中,由对抗原起反应而产生的b细胞群体表达的抗体包括(i)具有由人类重链可变区序列编码的人类重链可
变结构域的重链,和(ii)具有由如本文所述的人类λ轻链可变区序列编码的人类λ轻链可变结构域的λ轻链。
[0345]
在一些实施方案中,非人类动物(例如啮齿动物,例如大鼠或小鼠)产生表达结合于所关注抗原的一个或多个表位的抗体的b细胞群体,其中由对抗原起反应而产生的b细胞群体表达的抗体包括:(i)具有由人类重链可变区序列编码的人类重链可变结构域的重链,和(ii)具有由如本文所述的人类λ轻链可变区序列编码的人类λ轻链可变结构域的λ轻链。在一些实施方案中,如本文所述的人类重链可变区序列和/或人类λ轻链可变区序列进行体细胞超突变。在一些实施方案中,对抗原起反应而产生的b细胞群体中的b细胞的至少约5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%包括进行体细胞超突变的人类重链可变区序列和/或人类λ轻链可变区序列。
[0346]
特定示例性实施方案
‑
免疫球蛋白κ轻链基因座
[0347]
在一些实施方案中,所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)包含工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含单一重排人类免疫球蛋白λ轻链可变区且插入在非人类或人类cλ基因区段上游并可操作地连接至非人类或人类cλ基因区段,所述非人类或人类cλ基因区段插入在非人类cκ基因的位置中。如本文所述,这种工程化内源免疫球蛋白κ轻链基因座还包括非人类免疫球蛋白κ轻链增强子区域(或增强子序列)。在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)包含包括在人类免疫球蛋白λ轻链基因座的a簇中出现的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区。在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)包含包括在人类免疫球蛋白λ轻链基因座的b簇中出现的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区。在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)包含包括在人类免疫球蛋白λ轻链基因座的c簇中出现的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区。在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)包含包括选自由以下组成的组的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,工程化κ轻链基因座(或等位基因)包含包括选自由以下组成的组的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
39、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,工程化κ轻链基因座(或等位基因)包含包括选自由以下组成的组的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1。在一些实施方案中,工程化κ轻链基因座(或等位基因)包含包括选自由以下组成的组的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。在一些实施方案中,工程化κ轻链基因座(或等位基因)包
含包括选自由以下组成的组的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:vλ1
‑
51、vλ1
‑
40和vλ2
‑
14。在一些实施方案中,工程化κ轻链基因座(或等位基因)包含包括选自vλ1
‑
51或vλ2
‑
14的人类vλ基因区段的单一重排人类免疫球蛋白λ轻链可变区。在一些实施方案中,工程化免疫球蛋白κ轻链基因座(或等位基因)包含包括选自由以下组成的组的人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:jλ1、jλ2、jλ3、jλ6和jλ7。在一些实施方案中,工程化免疫球蛋白κ轻链基因座(或等位基因)包含包括选自由以下组成的组的人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区:jλ1、jλ2、jλ3和jλ7。在一些实施方案中,工程化免疫球蛋白κ轻链基因座(或等位基因)包含包括人类jλ2基因区段的单一重排人类免疫球蛋白λ轻链可变区。
[0348]
本公开认识到,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)在抗体选择和产生机制(例如重组和体细胞超突变)中将利用其基因组中所包括的人类λ轻链可变区基因区段。因而,在多个实施方案中,由本文所述的非人类动物产生的人类免疫球蛋白λ轻链可变结构域由其基因组中所包括的人类λ轻链可变区基因区段或其体细胞超突变变体编码。
[0349]
在一些实施方案中,提供了一种非人类动物(例如啮齿动物,例如大鼠或小鼠),所述非人类动物的基因组包含工程化内源免疫球蛋白κ轻链基因座,其中非人类动物包括包含进行体细胞超突变的人类重链可变区序列和/或人类λ轻链可变区序列的b细胞。在一些实施方案中,本公开的非人类动物(例如啮齿动物,例如大鼠或小鼠)的b细胞中存在的人类重链可变区序列和/或人类λ轻链可变区序列具有1个、2个、3个、4个、5个或更多个体细胞超突变。所属领域的技术人员知道用于鉴定成熟抗体序列中的源基因区段的方法。举例来说,多种工具可用于帮助进行这项分析,例如dnaplot、imgt/v
‑
quest、joinsolver、soda和ab
‑
origin。
[0350]
在许多实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)含有在野生型免疫球蛋白κ轻链基因座(或等位基因)中出现的非人类免疫球蛋白κ轻链增强子区域(或增强子序列)。在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)含有在不同物种(例如不同啮齿动物物种)的野生型免疫球蛋白κ轻链基因座(或等位基因)中出现的非人类免疫球蛋白κ轻链增强子区域(或增强子序列)。
[0351]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)在其种系基因组中包含可操作地连接至一个或多个非人类免疫球蛋白κ轻链增强子(即增强子序列或增强子区域)的有限人类λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)。在某些实施方案中,所述有限人类λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)可操作地连接至鼠类免疫球蛋白κ轻链内含子增强子区域(igκei或eiκ)。在某些实施方案中,所述有限人类λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)可操作地连接至鼠类免疫球蛋白κ轻链3'增强子区域(igκ3'e或3'eκ)。在某些实施方案中,所述有限人类λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)可操作地连接至鼠类eiκ且可操作地连接至鼠类3'eκ。
[0352]
在一些实施方案中,工程化内源免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因是啮齿动物cλ基因,例如小鼠cλ基因或大鼠cλ基因。在某些实施方案中,工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因是或包含来自包括129品系、balb/c品
系、c57bl/6品系、混合129xc57bl/6品系或它们的组合的遗传背景的小鼠cλ基因。
[0353]
在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因包含与seq id no:1(小鼠cλ1)、seq id no:2(小鼠cλ2)或seq id no:3(小鼠cλ3)至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因包含与seq id no:1(小鼠cλ1)、seq id no:2(小鼠cλ2)或seq id no:3(小鼠cλ3)基本上同一或同一的序列。在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因是或包含小鼠cλ1基因的序列。
[0354]
在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)处的序列编码的非人类cλ结构域包含与seq id no:4(小鼠cλ1)、seq id no:5(小鼠cλ2)或seq id no:6(小鼠cλ3)至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)处的序列编码的非人类cλ结构域包含与seq id no:4(小鼠cλ1)、seq id no:5(小鼠cλ2)或seq id no:6(小鼠cλ3)基本上同一或同一的序列。在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)处的序列编码的非人类cλ基因是或包含小鼠cλ1域多肽。
[0355]
在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因包含与seq id no:7(大鼠cλ1)、seq id no:8(大鼠cλ2)、seq id no:9(大鼠cλ3)或seq id no:10(大鼠cλ4)至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在某些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因包含与seq id no:7(大鼠cλ1)、seq id no:8(大鼠cλ2)、seq id no:9(大鼠cλ3)或seq id no:10(大鼠cλ4)基本上同一或同一的序列。在某些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的非人类cλ基因是或包含大鼠cλ1基因的序列。
[0356]
在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)处的序列编码的非人类cλ结构域包含与seq id no:11(大鼠cλ1)、seq id no:12(大鼠cλ2)、seq id no:13(大鼠cλ3)或seq id no:14(大鼠cλ4)至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)处的序列编码的非人类cλ结构域包含与seq id no:11(大鼠cλ1)、seq id no:12(大鼠cλ2)、seq id no:13(大鼠cλ3)或seq id no:14(大鼠cλ4)基本上同一或同一的序列。在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)处的序列编码的非人类cλ结构域是或包含大鼠cλ1域多肽。
[0357]
在一些实施方案中,工程化免疫球蛋白κ轻链基因座(或等位基因)的人类cλ基因包括人类cλ基因,例如人类cλ1基因、人类cλ2基因、人类cλ3基因、人类cλ6基因或人类cλ7基因。在某些实施方案中,工程化免疫球蛋白κ轻链基因座(或等位基因)的人类cλ基因是或包含人类cλ2基因。
[0358]
在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的人类cλ基因包含与seq id no:15(人类cλ1)、seq id no:16(人类cλ2)、seq id no:17(人类cλ3)、seq id no:18(人类cλ6)或seq id no:19(人类cλ7)至少80%、至少85%、至少
90%、至少95%或至少98%同一的序列。在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的人类cλ基因包含与seq id no:15(人类cλ1)、seq id no:16(人类cλ2)、seq id no:17(人类cλ3)、seq id no:18(人类cλ6)或seq id no:19(人类cλ7)基本上同一或同一的序列。在一些实施方案中,如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的人类cλ基因是或包含人类cλ2基因的序列。
[0359]
在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的序列编码的人类cλ结构域包含与seq id no:20(人类cλ1)、seq id no:21(人类cλ2)、seq id no:22(人类cλ3)、seq id no:23(人类cλ6)或seq id no:24(人类cλ7)至少80%、至少85%、至少90%、至少95%或至少98%同一的序列。在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的序列编码的人类cλ结构域包含与seq id no:20(人类cλ1)、seq id no:21(人类cλ2)、seq id no:22(人类cλ3)、seq id no:23(人类cλ6)或seq id no:24(人类cλ7)基本上同一或同一的序列。在一些实施方案中,由位于如本文所述的工程化免疫球蛋白κ轻链基因座(或等位基因)的序列编码的人类cλ结构域是或包含人类cλ2域多肽。
[0360]
特定示例性实施方案
‑
免疫球蛋白重链基因座
[0361]
在一些实施方案中,所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)包含如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座,并且还包含特征在于存在以种系构型排列且可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)免疫球蛋白重链恒定区基因、增强子和调控区域的多个人类v
h
、d
h
和j
h
基因区段的工程化免疫球蛋白重链基因座(或等位基因)。在一些实施方案中,如本文所述的工程化免疫球蛋白重链基因座(或等位基因)包含可操作地连接至非人类免疫球蛋白重链恒定区的一个或多个人类v
h
基因区段、一个或多个人类d
h
基因区段和一个或多个人类j
h
基因区段。在某些实施方案中,工程化免疫球蛋白重链基因座(或等位基因)包含至少人类v
h
基因区段v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合。在某些实施方案中,工程化免疫球蛋白重链基因座(或等位基因)包含至少人类d
h
基因区段d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合。在某些实施方案中,工程化免疫球蛋白重链基因座(或等位基因)包含至少人类j
h
基因区段j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合。
[0362]
本公开认识到,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)在抗体选择和产生机制(例如重组和体细胞超突变)中将利用其基因组中所包含的人类重链可变区基因区段。因而,在多个实施方案中,由本文所述的非人类动物产生的人类免疫球蛋白重链可变结构域由其基因组中所包括的人类重链可变区基因区段或其体细胞超突变变体编码。
[0363]
在一些实施方案中,非人类动物(例如啮齿动物,例如大鼠或小鼠)包括包含进行体细胞超突变的人类重链可变区序列和/或人类λ轻链可变区序列的b细胞。在一些实施方
案中,本公开的非人类动物(例如啮齿动物,例如大鼠或小鼠)的b细胞中存在的人类重链可变区序列和/或人类λ轻链可变区序列具有1个、2个、3个、4个、5个或更多个体细胞超突变。所属领域的技术人员知道用于鉴定成熟抗体序列中的源基因区段的方法。举例来说,多种工具可用于帮助进行这项分析,例如dnaplot、imgt/v
‑
quest、joinsolver、soda和ab
‑
origin。
[0364]
在一些实施方案中,非人类免疫球蛋白重链恒定区包括一种或多种非人类免疫球蛋白重链恒定区基因,例如免疫球蛋白m(igm)、免疫球蛋白d(igd)、免疫球蛋白g(igg)、免疫球蛋白e(ige)和免疫球蛋白a(iga)。在某些实施方案中,非人类免疫球蛋白重链恒定区包括啮齿动物igm、啮齿动物igd、啮齿动物igg3、啮齿动物igg1、啮齿动物igg2b、啮齿动物igg2a、啮齿动物ige和啮齿动物iga恒定区基因。在一些实施方案中,所述人类v
h
、d
h
和j
h
基因区段可操作地连接至一个或多个非人类免疫球蛋白重链增强子(即增强子序列或增强子区域)。在一些实施方案中,所述人类v
h
、d
h
和j
h
基因区段可操作地连接至一个或多个非人类免疫球蛋白重链调控区域(或调控序列)。在一些实施方案中,所述人类v
h
、d
h
和j
h
基因区段可操作地连接至一个或多个非人类免疫球蛋白重链增强子(或增强子序列)和一个或多个非人类免疫球蛋白重链调控区域(或调控序列)。
[0365]
在一些实施方案中,如本文所述的工程化免疫球蛋白重链基因座不含内源adam6基因。在一些实施方案中,如本文所述的工程化免疫球蛋白重链基因座在与在相同物种的野生型非人类动物的种系基因组中所发现相同的种系基因组位置中不含内源adam6基因(或编码adam6的序列)。在一些实施方案中,如本文所述的工程化免疫球蛋白重链基因座不含人类adam6伪基因。在一些实施方案中,如本文所述的工程化免疫球蛋白重链基因座包含编码一种或多种非人类(例如啮齿动物)adam6多肽、其功能直系同源物、功能同源物或功能片段的至少一个核苷酸序列的插入。在一些实施方案中,所述插入可在如本文所述的工程化免疫球蛋白重链基因座外部(例如但不限于在最5'v
h
基因区段上游),在工程化免疫球蛋白重链基因座内或在非人类动物(例如但不限于随机引入的非人类adam6编码序列)、细胞或组织的种系基因组中的其它地方。
[0366]
在多个实施方案中,所提供的如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)不可检测地表达抗体分子中的内源非人类v
h
区域整体或部分。在多个实施方案中,所提供的如本文所述的非人类动物不含(或缺乏,或含有缺失)编码抗体分子中的内源非人类v
h
区域(例如v
h
、d
h
和/或j
h
)整体或部分的一个或多个核苷酸序列。在多个实施方案中,所提供的如本文所述的非人类动物具有包括内源非人类v
h
、d
h
和j
h
基因区段整体或部分缺失的种系基因组。在多个实施方案中,所提供的非人类动物有生殖力。
[0367]
关于建立具有这种工程化免疫球蛋白重链基因座(或等位基因)的靶向载体、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和动物的指导可见于以下中:macdonald(2014);美国专利no.6,596,541、8,642,835、8,697,940和8,791,323,各以引用的方式整体并入本文中。所属领域的技术人员知道所属领域已知的用于实现非人类(例如哺乳动物)基因组的这种基因工程化和/或操纵或者以其它方式制备、提供或制造这种序列以供引入非人类动物的种系基因组中的多种技术。
[0368]
特定示例性实施方案
‑
免疫球蛋白基因座的组合
[0369]
在一些实施方案中,如本文所提供的非人类动物(例如啮齿动物,例如大鼠或小
鼠)在其种系基因组中包含如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座,并且还包含一个或多个包括人类免疫球蛋白基因区段或其它人类或人源化基因(例如编码tdt的人类基因)的额外免疫球蛋白基因座(例如经由杂交繁育或多个基因靶向策略)。这种非人类动物可以如上所述或使用所属领域中已知的方法制备,以取决于非人类动物的预期用途,实现期望的工程化基因型。在其它免疫球蛋白基因座处的额外人类免疫球蛋白基因区段或其它人类或人源化基因(例如编码tdt的人类基因)可以通过进一步改变具有如上所述的基因修饰的细胞(例如胚胎干细胞)的基因组或通过所属领域中已知的繁育技术利用其它期望的基因修饰品系来引入。
[0370]
在一些实施方案中,如本文所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)在其种系基因组中包含如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座,并且在其种系基因组中还包含包括可操作地连接至一个或多个非人类动物免疫球蛋白重链恒定区基因的一个或多个人类v
h
基因区段、一个或多个人类d基因区段和一个或多个人类j
h
基因区段的工程化内源免疫球蛋白重链基因座。在一些实施方案中,非人类动物对于如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座是杂合或纯合的。在一些实施方案中,非人类动物对于如本文所述的工程化内源免疫球蛋白重链基因座是杂合或纯合的。在一些实施方案中,非人类动物对于如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座是纯合的并且对于如本文所述的工程化内源免疫球蛋白重链基因座是纯合的。
[0371]
在一些实施方案中,如本文所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)在其种系基因组中包含如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座、包含可操作地连接至一个或多个非人类动物免疫球蛋白重链恒定区基因的一个或多个人类v
h
基因区段、一个或多个人类d基因区段和一个或多个人类j
h
基因区段的工程化内源免疫球蛋白重链基因座,并且还包含功能失活(例如整体或部分缺失,或以其它方式引起无功能)的内源免疫球蛋白λ轻链基因座。在一些实施方案中,非人类动物对于如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座是杂合或纯合的。在一些实施方案中,非人类动物对于如本文所述的工程化内源免疫球蛋白重链基因座是杂合或纯合的。在一些实施方案中,非人类动物对于功能失活(例如整体或部分缺失,或以其它方式引起无功能)的内源免疫球蛋白λ轻链基因座是杂合或纯合的。在一些实施方案中,非人类动物对于如本文所述的包括有限人类免疫球蛋白λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化内源免疫球蛋白κ轻链基因座是纯合的,对于如本文所述的工程化内源免疫球蛋白重链基因座是纯合的,并且对于功能失活(例如整体或部分缺失,或以其它方式引起无功能)的内源免疫球蛋白λ轻链基因座是纯合的。
[0372]
在一些实施方案中,基因修饰非人类动物啮齿动物(例如小鼠或大鼠)的种系基因组包括包含两个等位基因的工程化内源免疫球蛋白κ基因座。在一些实施方案中,第一等位
基因包含有限人类λ轻链可变区谱系并且第二等位基因包含有限人类κ轻链可变区谱系。在一些实施方案中,如本文所述的基因修饰啮齿动物(例如小鼠或大鼠)、啮齿动物(例如小鼠或大鼠)细胞或啮齿动物(例如小鼠或大鼠)组织的种系基因组包含第一工程化内源免疫球蛋白κ轻链基因座等位基因,所述等位基因包含可操作地连接至啮齿动物(例如小鼠或大鼠)cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。在一些实施方案中,如本文所述的基因修饰啮齿动物(例如小鼠或大鼠)和因此啮齿动物(例如小鼠或大鼠)细胞或啮齿动物(例如小鼠或大鼠)组织包含第二工程化内源免疫球蛋白κ轻链基因座等位基因,所述等位基因包含可操作地连接至啮齿动物(例如小鼠或大鼠)cκ基因区段的单一重排人类免疫球蛋白κ轻链可变区,其中单一重排人类免疫球蛋白κ轻链可变区包含人类vκ基因区段和人类jκ基因区段。在一些实施方案中,这种啮齿动物(例如小鼠或大鼠)组织可以从第一工程化内源免疫球蛋白κ轻链基因座等位基因表达λ轻链并且从第二工程化内源免疫球蛋白κ轻链基因座等位基因表达κ轻链。在一些实施方案中,单一重排人类免疫球蛋白κ轻链可变区包含vκ3
‑
20或vκ1
‑
39且单一重排人类免疫球蛋白λ轻链可变区包含vλ1
‑
51或vλ2
‑
14。在一个实施方案中,单一重排人类免疫球蛋白κ轻链可变区是vκ3
‑
20/jκ1,且单一重排人类免疫球蛋白λ轻链可变区是vλ1
‑
51/jλ2或vλ2
‑
14/jλ2。
[0373]
在一些实施方案中,所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)具有包含以下的种系基因组:(a)纯合或杂合的免疫球蛋白重链基因座,所述重链基因座包含可操作地连接至一个或多个内源非人类免疫球蛋白重链恒定区的人类v
h
、d
h
和j
h
基因区段,以使得所述非人类动物表达包含与非人类c
h
结构域序列融合的人类v
h
结构域序列的免疫球蛋白重链;(b)免疫球蛋白κ轻链基因座,所述轻链基因座包含可操作地连接至非人类(例如啮齿动物)免疫球蛋白cλ基因区段的单一重排人类免疫球蛋白λ轻链,以使得所述非人类动物表达包含与非人类cλ结构域序列融合的人类vλ结构域序列的免疫球蛋白轻链。
[0374]
在一些实施方案中,所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)具有包含以下的种系基因组:(a)纯合或杂合的免疫球蛋白重链基因座,所述重链基因座包含可操作地连接至一个或多个内源非人类免疫球蛋白重链恒定区的人类v
h
、d
h
和j
h
基因区段,以使得所述非人类动物表达包含与非人类c
h
结构域序列融合的人类v
h
结构域序列的免疫球蛋白重链;(b)免疫球蛋白κ轻链基因座,所述轻链基因座包含可操作地连接至非人类(例如啮齿动物)免疫球蛋白cλ基因区段的单一重排人类免疫球蛋白λ轻链,以使得所述非人类动物表达包含与非人类cλ结构域序列融合的人类vλ结构域序列的免疫球蛋白轻链;和(c)纯合或杂合的功能失活或整体或部分缺失的内源免疫球蛋白λ轻链基因座。
[0375]
举例来说,如本文所述,如本文所述的包含工程化内源免疫球蛋白κ轻链基因座的非人类动物还可包含(例如经由杂交繁育或多个基因靶向策略)如美国专利no.8,642,835、8,697,940、9,006,511、9,035,128、9,066,502、9,150,662和9,163,092中所述的一个或多个修饰;各专利全文引用的方式并入。
[0376]
核酸构建体
[0377]
通常,含有人类免疫球蛋白λ轻链序列(例如单一重排人类λ轻链可变区,或一个或两个未重排vλ基因区段与至少一个未重排jλ基因区段)或其部分的多核苷酸分子与载体、优选地dna载体连接(例如插入其中),以在宿主细胞中复制所述多核苷酸分子。
[0378]
人类免疫球蛋白λ轻链序列可以直接从已知的序列或来源(例如文库)克隆,或者从基于可从genbank或其它公共可用的数据库(例如imgt)获得的公开序列,电脑模拟设计的种系序列合成。可替代地,细菌人工染色体(bac)文库可提供所关注免疫球蛋白dna序列(例如人类vλ和jλ序列和它们的组合)。bac文库可含有100
‑
150kb的插入尺寸,且能够具有大至300kb的插入物(shizuya等人,1992,proc.natl.acad.sci.,usa 89:8794
‑
8797;swiatek等人,1993,genes and development 7:2071
‑
2084;kim等人,1996,genomics 34 213
‑
218;以引用的方式整体并入本文中)。举例来说,已经描述了具有164
‑
196kb的平均插入物尺寸的人类bac文库(osoegawa,k.等人,2001,genome res.11(3):483
‑
96;osoegawa,k.等人,1998,genomics 52:1
‑
8,论文编号ge985423,各以引用的方式整体并入本文中)。已经构建人类和非人类动物基因组bac文库且可购得(例如thermofisher)。基因组bac文库还可以用作免疫球蛋白dna序列以及转录控制区的来源。
[0379]
可替代地,可以从酵母人工染色体(yac)分离、克隆和/或转移免疫球蛋白dna序列。举例来说,已经确定人类免疫球蛋白λ轻链基因座的核苷酸序列(参见例如dunham,i.等人,1999,nature 402:489
‑
95,其以引用的方式整体并入本文中)。此外,yac先前已经用以组装人类免疫球蛋白λ轻链基因座转基因(参见例如popov,a.v.等人,1996,gene 177:195
‑
201;popov,a.v.等人,1999,j.exp.med.189(10):1611
‑
19,各以引用的方式整体并入本文中)。可对整个免疫球蛋白λ轻链基因座(人类或非人类)进行克隆并含于几个yac内。不管所包括的序列如何,如果采用多个yac且含有重叠相似区,那么它们都可以在酵母宿主菌株内重组,产生表示整个基因座或基因座的期望部分(例如靶向载体所靶向的区域)的单个构建体。yac臂可另外通过翻断,用哺乳动物选择盒来修饰,以帮助通过所属领域中已知和/或本文所述的方法将构建体引入胚胎干细胞或胚胎中。
[0380]
用于构建如本文所述的工程化免疫球蛋白κ轻链基因座的人类免疫球蛋白λ轻链基因区段的dna和氨基酸序列可以从公开数据库(例如genbank、imgt等)和/或公开的抗体序列获得。
[0381]
在某些实施方案中,核酸构建体含有可操作地连接至人类或非人类(例如啮齿动物,例如大鼠或小鼠)免疫球蛋白λ或免疫球蛋白κ轻链恒定区(分别cλ或cκ)基因的人类免疫球蛋白λ轻链基因区段(例如单一重排人类λ轻链可变区,或一个或两个未重排vλ基因区段与至少一个未重排jλ基因区段)。在某些实施方案中,核酸构建体含有可操作地连接至一个或多个非人类(例如啮齿动物,例如大鼠或小鼠)免疫球蛋白κ或免疫球蛋白λ轻链增强子区域(或增强子序列)的人类免疫球蛋白λ轻链基因区段(例如单一重排人类λ轻链可变区,或一个或两个未重排vλ基因区段与至少一个未重排jλ基因区段)。在一些实施方案中,核酸构建体含有可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)或人类cλ区域基因和非人类免疫球蛋白κ轻链增强子区域(或增强子序列)的人类免疫球蛋白λ轻链基因区段(例如单一重排人类λ轻链可变区,或一个或两个未重排vλ基因区段与至少一个未重排jλ基因区段)。
[0382]
在一些实施方案中,含有未重排人类vλ和jλ序列的核酸构建体还包含来源于人类和/或鼠类的基因间dna。在一些实施方案中,基因间dna是或包含非编码鼠类免疫球蛋白κ轻链序列、非编码人类免疫球蛋白κ轻链序列、非编码鼠类免疫球蛋白λ轻链序列、非编码人类免疫球蛋白λ轻链序列或它们的组合。
[0383]
可使用所属领域中已知的方法制备核酸构建体。举例来说,核酸构建体可以制备为较大的质粒的一部分。这种制备允许以如所属领域中已知的高效方式克隆和选择正确的构建。如本文所述的含有人类免疫球蛋白λ轻链序列整体或部分的核酸构建体可位于质粒上的限制位点之间,以便可以将其与剩余的质粒序列分离以并入期望的非人类动物(例如啮齿动物,例如大鼠或小鼠)中。
[0384]
所属领域中已知各种用于制备核酸构建体(例如质粒)和宿主生物体转化的方法。关于适用于原核和真核细胞的其它表达系统以及通用重组程序,参见principles of gene manipulation:an introduction to genetic manipulation,第5版,由old,r.w.和s.b.primrose编辑,blackwell science公司,1994和molecular cloning:a laboratory manual,第2版,sambrook,j.等人编辑,cold spring harbor laboratory press:1989,各以引用的方式整体并入本文中。
[0385]
靶向载体
[0386]
靶向载体可用以将核酸构建体引入标靶基因组基因座中。靶向载体可包含核酸构建体和在所述核酸构建体侧面的同源臂;所属领域的技术人员将清楚一般适用于靶向载体的设计、结构和/或使用的多种选项和特征。举例来说,靶向载体可以呈线性形式或呈圆形形式,且其可以是单股或双股。靶向载体可以是脱氧核糖核酸(dna)或核糖核酸(rna)。为了便于提及,同源臂在本文中称为5'和3'(即上游和下游)同源臂。所述术语是指在靶向载体内同源臂相对核酸构建体的位置。5'和3'同源臂与靶向基因座内的区域对应或另一靶向载体内的区域对应,在本文中分别称为“5'标靶序列”和“3'标靶序列”。在一些实施方案中,同源臂也可以充当5'或3'标靶序列。
[0387]
在一些实施方案中,本文所述的方法采用两种、三种或更多种能够彼此重组的靶向载体。在多个实施方案中,靶向载体是大的靶向载体(ltvec),如本文中其它地方描述。在一些实施方案中,第一、第二和第三靶向载体各自包含5'和3'同源臂。第一靶向载体的3'同源臂包含与第二靶向载体的5'同源臂重叠的序列(即重叠序列),这允许第一和第二ltvec之间同源重组。
[0388]
在双重靶向法的情况下,第一靶向载体的5'同源臂和第二靶向载体的3'同源臂可类似于标靶基因组基因座内的对应区段(即标靶序列),这能够促进第一和第二靶向载体与对应基因组区段的同源重组并修饰标靶基因组基因座。
[0389]
在三重靶向法的情况下,第二靶向载体的3'同源臂可包含与第三靶向载体的5'同源臂重叠的序列(即重叠序列),这能够允许第二与第三ltvec之间同源重组。第一靶向载体的5'同源臂和第三靶向载体的3'同源臂类似于标靶基因组基因座内的对应区段(即标靶序列),这能够促进第一和第三靶向载体与对应基因组区段的同源重组且修饰标靶基因组基因座。
[0390]
当两个区域享有彼此足够水平的序列同一性,以便其能够充当同源重组反应的底物时,同源臂与标靶序列或两个同源臂“相对应”或彼此“对应”。给定标靶序列与基于靶向载体的对应同源臂(即重叠序列)之间或两个同源臂之间的序列同一性可以是允许同源重组发生的任何程度的序列同一性。仅举一例,靶向载体(或其片段)的同源臂与另一靶向载体的标靶序列或标靶基因组基因座(或其片段)的标靶序列共享的序列同一性的量可以是例如但不限于至少50%、55%、60%、65%、70%、75%、80%、81%、82%、83%、84%、85%、
86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%序列同一性,以使得序列进行同源重组。
[0391]
此外,同源臂与对应标靶序列之间的类似性(例如同一性)对应区域可具有足够促进标靶基因组基因座处同源重组的任何长度。举例来说,给定同源臂和/或对应标靶序列所包含的类似性对应区域可例如为但不限于约5
‑
10kb、5
‑
15kb、5
‑
20kb、5
‑
25kb、5
‑
30kb、5
‑
35kb、5
‑
40kb、5
‑
45kb、5
‑
50kb、5
‑
55kb、5
‑
60kb、5
‑
65kb、5
‑
70kb、5
‑
75kb、5
‑
80kb、5
‑
85kb、5
‑
90kb、5
‑
95kb、5
‑
100kb、100
‑
200kb或200
‑
300kb长(例如本文中其它地方描述),以便同源臂具有足够类似性,从而与细胞的标靶基因组基因座内或另一靶向载体内的对应标靶序列进行同源重组。在一些实施方案中,给定同源臂和/或对应标靶序列所包含的类似性对应区域例如为但不限于约10
‑
100kb、15
‑
100kb、20
‑
100kb、25
‑
100kb、30
‑
100kb、35
‑
100kb、40
‑
100kb、45
‑
100kb、50
‑
100kb、55
‑
100kb、60
‑
100kb、65
‑
100kb、70
‑
100kb、75
‑
100kb、80
‑
100kb、85
‑
100kb、90
‑
100kb或95
‑
100kb长(例如本文中其它地方描述),以便同源臂具有足够类似性,从而与细胞的标靶基因组基因座内或另一靶向载体内的对应标靶序列进行同源重组。
[0392]
第一靶向载体的3'同源臂与第二靶向载体的5'同源臂或第二靶向载体的3'同源臂与第三靶向载体的5'同源臂的重叠序列可具有足够促进所述靶向载体之间同源重组的任何长度。举例来说,同源臂的给定重叠序列包含的对应重叠区可为约1
‑
5kb、5
‑
10kb、5
‑
15kb、5
‑
20kb、5
‑
25kb、5
‑
30kb、5
‑
35kb、5
‑
40kb、5
‑
45kb、5
‑
50kb、5
‑
55kb、5
‑
60kb、5
‑
65kb、5
‑
70kb、5
‑
75kb、5
‑
80kb、5
‑
85kb、5
‑
90kb、5
‑
95kb、5
‑
100kb、100
‑
200kb或200
‑
300kb长,以便同源臂的重叠序列具有足够类似性,从而与另一靶向载体内的对应重叠序列进行同源重组。在一些实施方案中,同源臂的给定重叠序列包含的重叠区为约1
‑
100kb、5
‑
100kb、10
‑
100kb、15
‑
100kb、20
‑
100kb、25
‑
100kb、30
‑
100kb、35
‑
100kb、40
‑
100kb、45
‑
100kb、50
‑
100kb、55
‑
100kb、60
‑
100kb、65
‑
100kb、70
‑
100kb、75
‑
100kb、80
‑
100kb、85
‑
100kb、90
‑
100kb或95
‑
100kb长,以便同源臂的重叠序列具有足够类似性,从而与另一靶向载体内的对应重叠序列进行同源重组。在一些实施方案中,重叠序列为1
‑
5kb(包括端点)。在一些实施方案中,重叠序列为约1kb至约70kb(包括端点)。在一些实施方案中,重叠序列为约10kb至约70kb(包括端点)。在一些实施方案中,重叠序列为约10kb至约50kb(包括端点)。在一些实施方案中,重叠序列为至少10kb。在一些实施方案中,重叠序列为至少20kb。举例来说,重叠序列可为约1kb至约5kb(包括端点)、约5kb至约10kb(包括端点)、约10kb至约15kb(包括端点)、约15kb至约20kb(包括端点)、约20kb至约25kb(包括端点)、约25kb至约30kb(包括端点)、约30kb至约35kb(包括端点)、约35kb至约40kb(包括端点)、约40kb至约45kb(包括端点)、约45kb至约50kb(包括端点)、约50kb至约60kb(包括端点)、约60kb至约70kb(包括端点)、约70kb至约80kb(包括端点)、约80kb至约90kb(包括端点)、约90kb至约100kb(包括端点)、约100kb至约120kb(包括端点)、约120kb至约140kb(包括端点)、约140kb至约160kb(包括端点)、约160kb至约180kb(包括端点)、约180kb至约200kb(包括端点)、约200kb至约220kb(包括端点)、约220kb至约240kb(包括端点)、约240kb至约260kb(包括端点)、约260kb至约280kb(包括端点)、或约280kb至约300kb(包括端点)。仅举一例,重叠序列可为约20kb至约60kb(包括端点)。可替代地,重叠序列可为至少1kb、至少5kb、至少10kb、至少15kb、至少20kb、至少25kb、至少30kb、至少35kb、至少40kb、至少45kb、至少50kb、至少60kb、至少
70kb、至少80kb、至少90kb、至少100kb、至少120kb、至少140kb、至少160kb、至少180kb、至少200kb、至少220kb、至少240kb、至少260kb、至少280kb或至少300kb。在一些实施方案中,重叠序列可为至多400kb、至多350kb、至多300kb、至多280kb、至多260kb、至多240kb、至多220kb、至多200kb、至多180kb、至多160kb、至多140kb、至多120kb、至多100kb、至多90kb、至多80kb、至多70kb、至多60kb或至多50kb。
[0393]
在一些实施方案中,同源臂可与细胞天然的基因座(例如靶向基因座)对应,或可替代地,其可与整合至细胞基因组中的dna的异源或外源区段的区域,包括例如转基因、表达盒或dna的异源或外源区域对应。在一些实施方案中,同源臂可在一些实施方案中与细胞中靶向载体上的区域对应。在一些实施方案中,靶向载体的同源臂可与酵母人工染色体(yac)、细菌人工染色体(bac)、人类人工染色体的区域或适当宿主细胞中含有的任何其它工程化区域对应。更进一步,靶向载体的同源臂可对应于或源自于bac文库、黏粒文库或p1噬菌体文库。在某些实施方案中,靶向载体的同源臂与对原核生物、酵母、禽类(例如鸡)、非人类哺乳动物、啮齿动物、人类、大鼠、小鼠、仓鼠兔、猪、牛、鹿、绵羊、山羊、猫、犬、雪貂、灵长类动物(例如狨、恒河猴)、家养哺乳动物、农业哺乳动物或所关注其它生物体来说是天然、异源或外源的基因座对应。在一些实施方案中,同源臂与显示使用常规方法的可靶向性有限,或在缺乏核酸酶剂(例如cas蛋白质)诱发的切口或双股断裂下显示在标靶位点相对低水平的成功整合和/或显著水平的脱靶整合的细胞基因座对应。在一些实施方案中,同源臂被设计成包括工程化dna。
[0394]
在一些实施方案中,靶向载体的5'和3'同源臂与靶向基因组对应。可替代地,同源臂与相关基因组对应。举例来说,靶向基因组是第一品系的小鼠基因组,且靶向臂与第二品系的小鼠基因组对应,其中第一品系与第二品系不同。在某些实施方案中,同源臂与相同动物的基因组对应,或来自于相同品系的基因组,例如靶向基因组是第一品系的小鼠基因组,且靶向臂与来自相同小鼠或来自相同品系的小鼠基因组对应。
[0395]
靶向载体的同源臂可具有足够促进与对应标靶序列的同源重组事件的任何长度,包括例如1
‑
5kb(包括端点)、5
‑
10kb(包括端点)、5
‑
15kb(包括端点)、5
‑
20kb(包括端点)、5
‑
25kb(包括端点)、5
‑
30kb(包括端点)、5
‑
35kb(包括端点)、5
‑
40kb(包括端点)、5
‑
45kb(包括端点)、5
‑
50kb(包括端点)、5
‑
55kb(包括端点)、5
‑
60kb(包括端点)、5
‑
65kb(包括端点)、5
‑
70kb(包括端点)、5
‑
75kb(包括端点)、5
‑
80kb(包括端点)、5
‑
85kb(包括端点)、5
‑
90kb(包括端点)、5
‑
95kb(包括端点)、5
‑
100kb(包括端点)、100
‑
200kb(包括端点)或200
‑
300kb(包括端点)长。在一些实施方案中,靶向载体的同源臂具有足够促进与对应标靶序列的同源重组事件的长度,为1
‑
100kb(包括端点)、5
‑
100kb(包括端点)、10
‑
100kb(包括端点)、15
‑
100kb(包括端点)、20
‑
100kb(包括端点)、25
‑
100kb(包括端点)、30
‑
100kb(包括端点)、35
‑
100kb(包括端点)、40
‑
100kb(包括端点)、45
‑
100kb(包括端点)、50
‑
100kb(包括端点)、55
‑
100kb(包括端点)、60
‑
100kb(包括端点)、65
‑
100kb(包括端点)、70
‑
100kb(包括端点)、75
‑
100kb(包括端点)、80
‑
100kb(包括端点)、85
‑
100kb(包括端点)、90
‑
100kb(包括端点)或95
‑
100kb(包括端点)长。如本文所述,大的靶向载体可采用更大长度的靶向臂。
[0396]
核酸酶剂(例如crispr/cas系统)可以与靶向载体组合使用来促进标靶基因座的修饰(例如免疫球蛋白κ轻链基因座的修饰,或先前修饰或工程化免疫球蛋白κ轻链基因座的修饰)。这种核酸酶剂可促进靶向载体与标靶基因座之间的同源重组。当核酸酶剂与靶向
载体组合使用时,靶向载体可包含与位置足够接近核酸酶裂解位点以便促进标靶序列与同源臂之间的同源重组事件在核酸酶裂解位点处切口或双股断裂后发生的5'和3'标靶序列相对应的5'和3'同源臂。术语“核酸酶裂解位点”包括由核酸酶剂建立切口或双股断裂的dna序列(例如cas9裂解位点)。如果距离达到促进5'和3'标靶序列与同源臂之间的同源重组事件在识别位点处切口或双股断裂后发生,那么与靶向载体的5'和3'同源臂对应的靶向基因座内的标靶序列“位置足够接近”核酸酶裂解位点。因此,在某些实施方案中,与靶向载体的5'和/或3'同源臂相对应的标靶序列在给定识别位点的至少一个核苷酸内,或在给定识别位点的至少10个核苷酸至约14kb内。在一些实施方案中,核酸酶裂解位点紧邻标靶序列中的至少一个或两者。
[0397]
与靶向载体的同源臂对应的标靶序列与核酸酶裂解位点的空间关系可以变化。举例来说,标靶序列可位于核酸酶裂解位点的5',标靶序列可位于识别位点的3',或标靶序列可在核酸酶裂解位点侧面。
[0398]
与仅仅使用靶向载体相比,靶向载体(包括例如大靶向载体)与核酸酶剂的组合使用可增加靶向效率。举例来说,当靶向载体联合核酸酶剂使用时,与仅仅使用靶向载体相比,靶向载体的靶向效率可增加至少两倍、至少三倍、至少四倍、至少五倍、至少六倍、至少七倍、至少八倍、至少九倍、至少十倍或在由这些整数形成的范围内,例如2
‑
10倍。
[0399]
一些靶向载体是“大的靶向载体”或“ltvec”,其包括同源臂与比意图在细胞中进行同源重组的其它途径通常所用的核酸序列大的核酸序列对应且源自于这些核酸序列的靶向载体。ltvec可为例如至少10kb长,或5'同源臂与3'同源臂总计可达例如至少10kb。ltvec还包括核酸构建体比意图在细胞中进行同源重组的其它途径通常所用的核酸构建体大的靶向载体。举例来说,ltvec令大基因座的修饰成为可能,这些大基因座因尺寸限制而无法被传统的基于质粒的靶向载体容纳。举例来说,靶向基因座可为(即5'和3'同源臂可对应于)使用常规方法无法靶向或在缺乏由核酸酶剂(例如cas蛋白)诱发的切口或双股断裂下靶向不正确或效率显著低的细胞基因座。
[0400]
在一些实施方案中,本文所述的方法可采用两种或三种ltvec,这些ltvec能够以三路或四路重组事件彼此重组和与标靶基因组基因座重组。这种方法令使用单个ltvec无法实现的大基因座的修饰成为可能。
[0401]
ltvec的实例包括源自于细菌人工染色体(bac)、人类人工染色体或酵母人工染色体(yac)的载体。ltvec可以呈线性形式或呈圆形形式。ltvec和它们的制造方法的实例描述于例如macdonald(2014)的美国专利no.6,586,251、6,596,541和7,105,348;和国际专利申请公布no.wo 2002/036789,各以引用的方式整体并入本文中。
[0402]
制造所提供的非人类动物的方法
[0403]
提供了用于制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的组合物和方法,所述非人类动物的种系基因组包含工程化免疫球蛋白κ轻链基因座,所述工程化免疫球蛋白κ轻链基因座包括一个或多个人类免疫球蛋白λ轻链序列(例如人类vλ和jλ基因区段)代替非人类免疫球蛋白κ轻链序列,包括包含人类vλ和jλ区段的特定多态形式(例如特定v和/或j等位基因或变体)的人类免疫球蛋白λ轻链编码序列,所述组合物和方法包括用于制造表达如下抗体的非人类动物的组合物和方法,所述抗体包含含有人类可变区和非人类或人类恒定区的免疫球蛋白λ轻链,所述免疫球蛋白λ轻链从含有可操作地连接至非人类或人类
免疫球蛋白λ轻链恒定区基因的单一重排人类免疫球蛋白λ轻链可变区的免疫球蛋白κ轻链基因座组装,所述非人类或人类免疫球蛋白λ轻链恒定区基因位于在野生型非人类免疫球蛋白κ轻链基因座中正常出现的非人类免疫球蛋白κ轻链恒定区基因的位置中。在一些实施方案中,还提供了用于制造在内源免疫球蛋白κ增强子和/或内源免疫球蛋白κ调控序列控制下表达这种抗体的非人类动物的组合物和方法。在一些实施方案中,还提供了用于制造在异源免疫球蛋白κ增强子和/或异源免疫球蛋白κ调控序列控制下表达这种抗体的非人类动物的组合物和方法。
[0404]
本文所述的方法包括在非人类或人类免疫球蛋白λ轻链恒定区基因(例如啮齿动物,例如鼠类,例如大鼠或小鼠)或人类cλ区域基因)上游插入编码人类免疫球蛋白λ轻链可变结构域的单一重排人类免疫球蛋白λ轻链可变区,所述非人类或人类免疫球蛋白λ轻链恒定区基因位于在野生型非人类免疫球蛋白κ轻链基因座中正常出现的非人类免疫球蛋白κ轻链恒定区基因的位置中,以便表达抗体,所述抗体的特征在于存在含有人类λ轻链可变结构域和非人类cλ结构域(例如啮齿动物(例如鼠类,例如大鼠或小鼠)cλ结构域)的轻链或存在含有人类λ轻链可变和人类cλ结构域的轻链,且在b细胞表面上和非人类动物血流中均表达。
[0405]
在一些实施方案中,方法包括将含有单一重排人类免疫球蛋白λ轻链可变区的遗传物质插入非人类动物(例如啮齿动物,例如大鼠或小鼠)的免疫球蛋白κ轻链基因座(例如野生型、修饰或工程化免疫球蛋白κ轻链基因座)中。在一些实施方案中,方法包括将含有单一重排人类免疫球蛋白λ轻链可变区的遗传物质插入非人类动物(例如啮齿动物,例如大鼠或小鼠)的修饰或工程化品系的免疫球蛋白κ轻链基因座中。
[0406]
在一些实施方案中,方法包括单个es细胞克隆中多次插入。在一些实施方案中,方法包括连续es细胞克隆中进行依次插入。在一些实施方案中,方法包括工程化es细胞克隆中进行单次插入。
[0407]
在一些实施方案中,方法包括在非人类(例如啮齿动物,例如大鼠或小鼠)cλ1基因(或人类cλ2基因)上游插入dna以使得所述dna插入可操作地连接至所述非人类(例如啮齿动物,例如大鼠或小鼠)cλ1基因(或人类cλ2基因),所述dna插入包含选自由以下组成的组的一个或两个人类vλ基因区段:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
39、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1,和选自由以下组成的组的一个或多个人类jλ基因区段:jλ1、jλ2、jλ3、jλ6和jλ7,并且所述非人类(例如啮齿动物,例如大鼠或小鼠)cλ1基因(或人类cλ2基因)位于内源免疫球蛋白κ轻链基因座的非人类(例如啮齿动物,例如大鼠或小鼠)cκ基因的位置中。在一些实施方案中,方法包括在非人类(例如啮齿动物,例如大鼠或小鼠)cλ1基因(或人类cλ2基因)上游插入dna以使得所述dna插入可操作地连接至所述非人类(例如啮齿动物,例如大鼠或小鼠)cλ1基因(或人类cλ2基因),所述dna插入包含单一重排人类λ轻链可变区,所述单一重排人类λ轻链可变区包括选自由以下组成的组的人类vλ基因区段:vλ4
‑
69、vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
39、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3和vλ3
‑
1,和选
自由以下组成的组的人类jλ基因区段:jλ1、jλ2、jλ3、jλ6和jλ7,且所述非人类(例如啮齿动物,例如大鼠或小鼠)cλ1基因(或人类cλ2基因)位于内源免疫球蛋白κ轻链基因座的非人类(例如啮齿动物,例如大鼠或小鼠)cκ基因的位置中。
[0408]
适当时,编码人类免疫球蛋白λ轻链可变结构域的人类免疫球蛋白λ轻链序列(即含有人类vλ和jλ基因区段的序列)可单独进行修饰以包括针对在非人类动物中表达进行优化的密码子(例如参见美国专利no.5,670,356和5,874,304,各以引用的方式并入本文中)。密码子优化的序列是工程化序列,且优选地编码由未密码子优化的亲本多核苷酸编码的同一多肽(或具有与全长多肽基本上相同活性的全长多肽的生物活性片段)。在一些实施方案中,编码人类免疫球蛋白λ轻链可变结构域的人类免疫球蛋白λ轻链序列可单独包括改变序列以针对具体细胞类型(例如啮齿动物细胞,例如大鼠或小鼠细胞)优化密码子。举例来说,有待插入如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)的基因组中的每个核苷酸序列的密码子可针对在非人类动物的细胞中表达进行优化。这种序列可称为密码子优化序列。
[0409]
编码人类免疫球蛋白λ轻链可变结构域的核苷酸序列的插入采用如本文所述的非人类动物的种系基因组的最小修饰并引起包含具有人类vλ结构域的轻链的抗体的表达,所述人类免疫球蛋白λ轻链可变结构域从内源工程化免疫球蛋白κ轻链基因座表达。产生工程化非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法,包括敲除和敲入,是所属领域中已知的(参见例如gene targeting:a practical approach,joyner编辑,oxford university press公司,2000;以引用的方式整体并入本文中)。举例来说,基因工程啮齿动物的产生可任选地涉及一个或多个内源啮齿动物基因(或基因区段)的遗传基因座的破坏和一个或多个异源基因(或基因区段或核苷酸序列)引入啮齿动物基因组中,在一些实施方案中,引入与内源啮齿动物基因(或基因区段)相同的位置上。在一些实施方案中,编码人类vλ结构域的核苷酸序列引入在啮齿动物种系基因组中随机插入的工程化轻链转基因的非人类(例如啮齿动物,例如大鼠或小鼠)或人类免疫球蛋白λ轻链恒定区基因上游。在一些实施方案中,编码人类vλ结构域的核苷酸序列引入在啮齿动物种系基因组中内源免疫球蛋白κ轻链基因座的非人类(例如啮齿动物,例如大鼠或小鼠)或人类免疫球蛋白λ轻链恒定区基因上游;在某些实施方案中,内源免疫球蛋白κ轻链基因座被改变、修饰或工程化成含有可操作地连接至小鼠cλ1基因或可操作地连接至人类cλ2基因的人类免疫球蛋白λ基因区段(例如人类v和j)。
[0410]
用于构建如本文所述的工程化免疫球蛋白κ轻链基因座的示例性方法的示意图(不按比例)提供于图1
‑
6中。具体地说,图1
‑
6阐述了用于构建工程化免疫球蛋白κ轻链基因座的示例性策略,其特征在于插入含有单一重排人类免疫球蛋白λ轻链可变区的核苷酸序列,以及相应靶向载体。靶向载体可线性化且电穿孔至啮齿动物胚胎干细胞(es)中以建立种系基因组包含工程化免疫球蛋白κ轻链基因座的啮齿动物。如以下实施例部分中所述,靶向载体的电穿孔中所采用的啮齿动物es细胞含有如先前在美国专利no.10,143,186中所述的工程化免疫球蛋白κ轻链基因座;以引用的方式整体并入本文中。使用所属领域已知的筛选方法确认阳性啮齿动物es细胞克隆。任何剩余的选择盒可以按照要求经由重组介导的缺失而缺失。
[0411]
可替代地,人类cλ基因可代替小鼠cλ基因用于靶向载体中。除了编码人类cλ基因
(例如cλ2)的序列可工程化至靶向载体中之外,靶向载体可以按照与上述(或以下实施例中)类似的方式构建。使用这种方法允许研发人类抗体治疗剂,因为编码轻链可变和恒定区的dna可以一起分离,从而消除了连接至人类轻链恒定区以制备完整人类抗体的任何后续克隆步骤。
[0412]
用于构建如本文所述的工程化免疫球蛋白κ轻链基因座的靶向载体可并入非人类细胞(例如啮齿动物(例如大鼠或小鼠)胚胎干细胞)的种系基因组中。在一些实施方案中,如本文所述的靶向载体并入非人类(例如啮齿动物,例如大鼠或小鼠)细胞的种系基因组中的野生型免疫球蛋白κ轻链基因座中,所述种系基因组还含有与一个或多个免疫球蛋白重链恒定区基因可操作地连接的人类v
h
、d
h
和j
h
基因组dna(例如含有多个人类v
h
、d
h
和j
h
基因区段)(例如参见macdonald(2014),美国专利no.6,596,541、8,642,835、8,697,940和8,791,323,各以引用的方式整体并入本文中)。在一些实施方案中,如本文所述的靶向载体并入非人类细胞的种系基因组中修饰或工程化免疫球蛋白κ轻链基因座中,所述种系基因组还含有与一个或多个免疫球蛋白重链恒定区基因可操作地连接的人类v
h
、d
h
和j
h
基因组dna(例如含有多个人类v
h
、d
h
和j
h
基因区段)(例如参见macdonald(2014),美国专利no.6,596,541、8,642,835、8,697,940、8,791,323、9,006,511、9,012,717、9,029,628、9,035,128、9,066,502、9,150,662和9,163,092,各以引用的方式整体并入本文中)。
[0413]
靶向载体通过电穿孔引入非人类(例如啮齿动物,例如小鼠或大鼠)胚胎干细胞中,以使得靶向载体中所含的序列使非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类动物(例如啮齿动物,例如大鼠或小鼠)能够表达如下抗体,所述抗体具有包括人类免疫球蛋白λ轻链可变结构域和非人类或人类轻链恒定(cλ或cκ)结构域的轻链,并且所述轻链从工程化内源免疫球蛋白κ轻链基因座表达。如本文所述,产生基因工程非人类动物,其中已经在非人类动物的种系基因组中建立工程化免疫球蛋白κ轻链基因座(例如含有可操作地连接至在内源啮齿动物cκ基因位置处的啮齿动物或人类cλ基因的人类免疫球蛋白λ轻链序列(即重排人类λ轻链可变区)的内源免疫球蛋白κ轻链基因座)。抗体在非人类动物b细胞表面上和所述非人类动物血清中表达,所述抗体的特征在于轻链具有人类vλ结构域和非人类或人类cλ结构域。当靶向载体未靶向如本文所述的非人类动物的种系基因组中的内源免疫球蛋白κ轻链基因座时,工程化免疫球蛋白κ轻链转基因优选地插入除内源非人类动物免疫球蛋白κ轻链基因座外的位置处(例如随机插入的转基因)。
[0414]
如上所述在非人类动物中建立工程化免疫球蛋白κ轻链基因座提供了一种工程化非人类动物品系,所述品系产生如下抗体,所述抗体包括从这种工程化免疫球蛋白κ轻链基因座表达的具有人类vλ结构域和非人类(例如啮齿动物,例如大鼠或小鼠)或人类cλ结构域的免疫球蛋白λ轻链。利用包括可操作地连接至免疫球蛋白重链恒定区基因的多个人类v
h
、d
h
和j
h
基因区段的工程化免疫球蛋白重链基因座的存在,建立了产生用于研发人类抗体类治疗剂的抗体和抗体组分的工程化非人类动物品系。因此,实现了单个工程化非人类动物品系,它能够提供采用人类vλ结构域的替代体内系统来研发新的抗体类药物以治疗人类疾病。
[0415]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入非人类(例如啮齿动物,例如大鼠或小鼠)胚胎干(es)细胞的基因组中的工程化内
源免疫球蛋白κ轻链基因座中。在一些实施方案中,制造基因修饰非人类动物的方法包括将非人类(例如啮齿动物,例如大鼠或小鼠)cλ基因区段引入非人类es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中。在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括使用如上所述的非人类(例如啮齿动物,例如大鼠或小鼠)es细胞产生非人类动物。
[0416]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入非人类es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中。在一些实施方案中,制造基因修饰非人类动物的方法包括将非人类cλ基因区段引入非人类es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中。
[0417]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括以下步骤:(a)将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入非人类es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中;以及(b)将非人类cλ基因区段引入非人类es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中。
[0418]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括以下步骤:(a)将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入非人类胚胎干(es)细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中;(b)将非人类cλ基因区段引入非人类es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中;以及(c)使用步骤(a)和(b)中产生的非人类es细胞产生非人类动物。
[0419]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括将可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段引入非人类es细胞的基因组中的工程化内源免疫球蛋白重链基因座中。
[0420]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法还包括以下步骤:(d)将可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段引入非人类es细胞的基因组中的工程化内源免疫球蛋白重链基因座中。
[0421]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括对非人类动物的种系基因组中的内源免疫球蛋白κ轻链基因座进行工程化以包含可操作地连接至非人类cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段,使得由基因修饰非人类动物的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0422]
在一些实施方案中,制造基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法包括对非人类动物(例如啮齿动物,例如大鼠或小鼠)的种系基因组中的内源免疫球蛋白重链基因座进行工程化以包含可操作地连接至一个或多个非人类(例如啮齿动物,例如大鼠或小鼠)免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或
多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段,使得由基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)的b细胞表达的所有重链均包括人类免疫球蛋白重链可变结构域和非人类(例如啮齿动物,例如大鼠或小鼠)免疫球蛋白重链恒定结构域。
[0423]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,dna片段引入非人类胚胎干细胞中,所述非人类胚胎干细胞的种系基因组包含一个或多个工程化免疫球蛋白基因座(例如免疫球蛋白重链、免疫球蛋白κ轻链、免疫球蛋白λ轻链和它们的组合)。在某些实施方案中,工程化免疫球蛋白基因座是内源工程化免疫球蛋白基因座。
[0424]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,dna片段包含包括免疫球蛋白λ轻链序列和/或免疫球蛋白κ轻链序列的工程化序列。在制造非人类动物的方法的一些实施方案中,dna片段包含包括免疫球蛋白λ轻链序列和/或免疫球蛋白κ轻链序列和非免疫球蛋白序列(例如重组信号序列、抗性基因和它们的组合)的工程化序列。
[0425]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,dna片段引入非人类胚胎干细胞中,所述非人类胚胎干细胞的基因组包括包含一个或多个人类v
h
基因区段、一个或多个人类d
h
基因区段和一个或多个人类j
h
基因区段的插入的内源免疫球蛋白重链基因座,所述人类v
h
、d
h
和j
h
基因区段可操作地连接至非人类免疫球蛋白重链恒定区。在一些实施方案中,es细胞还包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的核苷酸序列。
[0426]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,dna片段引入非人类胚胎干细胞中,所述非人类胚胎干细胞的种系基因组包括包含单一重排人类λ轻链可变区的插入的内源免疫球蛋白κ轻链基因座,所述人类vλ和jλ基因区段可操作地连接至非人类免疫球蛋白κ或λ轻链恒定区基因。
[0427]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,在非人类胚胎干细胞中对非人类动物的种系基因组进行修饰以便其包括包含有限人类λ轻链可变区谱系(例如单一重排人类免疫球蛋白λ轻链可变区)的工程化免疫球蛋白κ轻链基因座,所述非人类胚胎干细胞的基因组包括包含一个或多个人类v
h
基因区段、一个或多个人类d
h
基因区段和一个或多个人类j
h
基因区段的插入的内源免疫球蛋白重链基因座,所述人类v
h
、d
h
和j
h
基因区段可操作地连接至非人类免疫球蛋白重链恒定区。
[0428]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,在非人类胚胎干细胞中对非人类动物的种系基因组进行修饰以便其包括包含有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座,所述非人类胚胎干细胞的种系基因组包括包含一个或多个人类vλ和一个或多个人类jλ基因区段的插入的内源免疫球蛋白κ轻链基因座,所述人类vλ和jλ基因区段可操作地连接至非人类免疫球蛋白κ轻链恒定区基因。在制造非人类动物的方法的一些实施方案中,在非人类胚胎干细胞中对非人类动物的种系基因组进行修饰以便其包含包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座,所述非人类胚胎干细胞的种系基因组包括包含一个或多个人类vλ和一个或多个人类jλ基因区段的插入的内源免疫球蛋白κ轻链基因座且人类免疫球蛋白κ轻链序列位于、置于或定位在所述一个或多个人类vλ基因区段与所述一个或多个人类jλ基因区段之间,所述人类vλ
和jλ基因区段可操作地连接至非人类免疫球蛋白κ轻链恒定区基因。
[0429]
在制造非人类动物(例如啮齿动物,例如大鼠或小鼠)的方法的一些实施方案中,一个或多个人类v
h
基因区段、一个或多个人类d
h
基因区段和一个或多个人类j
h
基因区段的插入包括内源人类免疫球蛋白基因座中天然出现在人类v
h
基因区段相邻处的人类非编码dna、天然出现在人类d
h
基因区段相邻处的人类非编码dna和天然出现在人类j
h
基因区段相邻处的人类非编码dna。
[0430]
在一些实施方案中,提供了从如本文所述的方法制造、产生(generate)、产生(produce)、获得或可获得的非人类动物(例如啮齿动物,例如大鼠或小鼠)。
[0431]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)的基因组还包含如以下中所述的一个或多个人类免疫球蛋白重链可变区:macdonald(2014),美国专利no.6,596,541、8,642,835、8,697,940和8,791,323,各以引用的方式整体并入本文中。可替代地,如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座可以工程化至不同修饰品系的胚胎干细胞中,例如品系(参见例如macdonald(2014),美国专利no.6,596,541和/或8,642,835;以引用的方式整体并入本文中)。如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的纯合性随后可以通过繁育实现。可替代地,在随机插入的工程化免疫球蛋白κ轻链转基因(上述)的状况下,可尤其基于人类vλ结构域从转基因的表达来选择非人类动物品系。在一些实施方案中,小鼠可以是1(vi
‑
1)小鼠,其包括十八个人类v
h
基因区段、所有人类d
h
基因区段和所有人类j
h
基因区段。vi
‑
1小鼠还可以包括十六个人类vκ基因区段和所有人类jκ基因区段。在一些实施方案中,小鼠可以是2(vi
‑
2)小鼠,其包括三十九个人类v
h
基因区段、所有人类d
h
基因区段和所有人类j
h
基因区段。vi
‑
2小鼠还可以包括人类三十个vκ基因区段和所有人类jκ基因区段。在一些实施方案中,小鼠可以是3(vi
‑
3)小鼠,其包括八十个人类v
h
基因区段、所有人类d
h
基因区段和所有人类j
h
基因区段。vi
‑
3小鼠还可以包括人类四十个vκ基因区段和所有人类jκ基因区段。
[0432]
可替代地,和/或另外,在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)的种系基因组还包含缺失、失活、功能沉默或在其它方面无功能的内源免疫球蛋白λ轻链基因座。使用本文所述的方法和/或所属领域已知的方法可实现使基因或遗传基因座缺失或无功能的基因修饰。
[0433]
基因工程创始非人类动物(例如啮齿动物,例如大鼠或小鼠)可基于其种系基因组中存在如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座和/或所述非人类动物的组织或细胞中表达具有人类λ轻链可变结构域和非人类或人类cλ或cκ结构域的抗体来鉴定。接着基因工程创始非人类动物可用于繁育携带如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的额外非人类动物,从而建立一群各自携带包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的一个或多个拷贝的非人类动物。此外,携带如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的基因工程非人类动物可按照要求进一步繁育成携带其它转基因(例如人类免疫球蛋白基因)或工程化免疫球蛋白基因座的其它基因工程非人类动物。
[0434]
还可以产生基因工程非人类动物(例如啮齿动物,例如大鼠或小鼠),其含有允许转基因或整合序列的调控性、指导性、诱导性和/或细胞类型特异性表达的选定系统。举例来说,如本文所述的非人类动物可以被工程化成含有编码抗体人类λ轻链可变结构域的条件性表达的一个或多个序列(例如rajewski,k.等人,1996,j.clin.invest.98(3):600
‑
3中评述,以引用的方式整体并入本文中)。示例性系统包括噬菌体p1的cre/loxp重组酶系统(参见例如lakso,m.等人,1992,proc.natl.acad.sci.u.s.a.89:6232
‑
6,以引用的方式整体并入本文中)和酿酒酵母(s.cerevisiae)的flp/frt重组酶系统(o’gorman,s.等人,1991,science 251:1351
‑
5,以引用的方式整体并入本文中)。可通过构建“双重”基因工程化动物,例如通过使两种基因工程化动物交配来提供这种动物,一种动物含有包含选定修饰的转基因(例如如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座)且另一种动物含有编码重组酶(例如cre重组酶)的转基因。
[0435]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)可如上所述或使用所属领域中已知的方法制备,以包含额外人类、人源化或以其它方式进行工程化的基因,常取决于非人类动物的预期用途。这种人类、人源化或以其它方式进行工程化的基因的遗传物质可通过进一步改变具有如上所述的基因修饰的细胞(例如胚胎干细胞)的基因组或通过所属领域中已知的繁育技术按照要求利用其它基因修饰或工程化品系来引入。在一些实施方案中,如本文所述的非人类动物被制备成还包含人类重链基因区段(参见例如murphy,a.j.等人,(2014)proc.natl.acad.sci.u.s.a.111(14):5153
‑
5158;macdonald(2014),美国专利no.6,596,541、8,642,835、8,697,940和8,791,323;美国专利no.8,791,323;和美国专利申请公布no.2013/0096287 a1;各以引用的方式整体并入本文中)。
[0436]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)可通过将本文所述的靶向载体引入来自修饰或工程化品系的细胞中来制备。举例来说,如本文所述的靶向载体可引入小鼠中。小鼠表达具有完全人类可变结构域和小鼠恒定结构域的抗体。在另一实例中,如本文所述的靶向载体可引入如美国专利no.9,006,511、9,012,717、9,029,628、9,035,128、9,066,502、9,150,662和9,163,092(以引用的方式整体并入本文中)中的任一个中所述的工程化小鼠中。在一些实施方案中,如本文所述的非人类动物被制备成还包含人类免疫球蛋白基因(可变和/或恒定区基因)。在一些实施方案中,如本文所述的非人类动物包含如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座和来自异源物种(例如人类)的遗传物质,其中遗传物质编码一个或多个人类重链可变区整体或部分。
[0437]
举例来说,如本文所述,包含如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的非人类动物(例如啮齿动物,例如大鼠或小鼠)还可包含(例如经由杂交繁育或多个基因靶向策略)一个或多个修饰,如以下中所述:murphy,a.j.等人,(2014)proc.natl.acad.sci.u.s.a.111(14):5153
‑
8;macdonald(2014);美国专利no.6,596,541、8,642,835、8,697,940和8,791,323;均以引用的方式整体并入本文中。在一些实施方案中,使包含如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的非人类动物与包含人源化免疫球蛋白重链可变区基因座的非人类动物杂交(参见例如macdonald(2014),美国专利no.6,596,541、8,642,835、8,697,940和/或8,791,323;以引用的方式整体并入本文中)。在一些实施方案中,使包含如本文所述的包括有限人类λ轻
链可变区谱系的工程化免疫球蛋白κ轻链基因座的非人类动物与包含人源化免疫球蛋白重链可变区基因座(参见例如macdonald(2014),美国专利no.6,596,541、8,642,835、8,697,940和/或8,791,323;以引用的方式并入本文中)和失活内源免疫球蛋白λ轻链基因座(参见例如美国专利no.9,006,511、9,012,717、9,029,628、9,035,128、9,066,502、9,150,662和9,163,092,以引用的方式整体并入本文中)的非人类动物杂交。
[0438]
虽然本文中广泛论述了描述在小鼠(即,具有特征为存在与小鼠或人类cλ或cκ基因可操作地连接的有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座以便表达含有人类λ轻链可变结构域和小鼠或人类cλ或cκ结构域的抗体的小鼠)中构建工程化免疫球蛋白κ轻链基因座的实施方案,还提供了包含工程化免疫球蛋白κ轻链基因座的其它非人类动物。这种非人类动物包括可进行基因修饰以表达如本文所述的抗体的那些动物中的任一者,包括例如哺乳动物,例如小鼠、大鼠、兔、猪、牛(例如母牛、公牛、水牛)、鹿、绵羊、山羊、鸡、猫、犬、雪貂、灵长类动物(例如绒、恒河猴)等。举例来说,对于不易获得合适的可基因修饰的es细胞的那些非人类动物,采用其它方法来制造包含基因修饰的非人类动物。这类方法包括例如修饰非es细胞基因组(例如纤维母细胞或诱导性多能干细胞)和采用体细胞核转移(scnt)将基因修饰基因组转移至适合细胞,例如去核卵母细胞,和在非人类动物中在合适条件下孕育修饰细胞(例如修饰卵母细胞)以形成胚胎。
[0439]
用于修饰非人类动物的种系基因组(例如猪、母牛、啮齿动物、鸡等基因组)的方法包括例如采用锌指核酸酶(zfn)、转录激活因子样效应物核酸酶(talen)或cas蛋白(即crispr/cas系统)来包括如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座。关于修饰非人类动物的种系基因组的方法指导可见于例如美国专利no.9,738,897和美国专利申请公布no.us 2016/0145646(2016年5月26日公开)和us 2016/0177339(2016年6月23日公开)中;各以引用的方式整体并入本文中。
[0440]
在一些实施方案中,如本文所述的非人类动物是哺乳动物。在一些实施方案中,如本文所述的非人类动物是小型哺乳动物,例如跳鼠科(dipodoidea)或鼠总科(muroidea)。在一些实施方案中,如本文所述的基因修饰动物是啮齿动物。在一些实施方案中,如本文所述的啮齿动物选自小鼠、大鼠和仓鼠。在一些实施方案中,如本文所述的啮齿动物选自鼠总科。在一些实施方案中,如本文所述的基因修饰动物来自选自以下的科:丽仓鼠科(calomyscidae)(例如小鼠样仓鼠)、仓鼠科(cricetidae)(例如仓鼠、新世界大鼠和小鼠、野鼠)、鼠科(muridae)(真正小鼠和大鼠、沙鼠、刺毛鼠、冠毛鼠)、马岛鼠科(nesomyidae)(攀鼠、岩鼠、白尾鼠、马达加斯加大鼠(malagasy rat)和小鼠)、刺山鼠科(platacanthomyidae)(例如多刺睡鼠)和鼹形鼠科(spalacidae)(例如鼹鼠(mole rate)、竹鼠和鼢鼠)。在某些实施方案中,如本文所述的基因修饰啮齿动物选自真正小鼠或大鼠(鼠科)、沙鼠、刺毛鼠和冠毛鼠。在某些实施方案中,如本文所述的基因修饰小鼠来自鼠科成员。在一些实施方案中,如本文所述的非人类动物是啮齿动物。在某些实施方案中,如本文所述的啮齿动物选自小鼠和大鼠。在一些实施方案中,如本文所述的非人类动物是小鼠。
[0441]
在一些实施方案中,如本文所述的非人类动物是啮齿动物,其为选自c57bl/a、c57bl/an、c57bl/grfa、c57bl/kalwn、c57bl/6、c57bl/6j、c57bl/6byj、c57bl/6nj、c57bl/10、c57bl/10scsn、c57bl/10cr和c57bl/ola的c57bl品系的小鼠。在某些实施方案中,如本文所述的小鼠是选自由以下组成的组的129
‑
品系:品系129p1、129p2、129p3、129x1、129s1
(例如129s1/sv、129s1/svim)、129s2、129s4、129s5、129s9/svevh、129/svjae、129s6(129/svevtac)、129s7、129s8、129t1、129t2(参见例如festing等人,1999,mammalian genome 10:836;auerbach,w.等人,2000,biotechniques 29(5):1024
‑
1028,1030,1032,各以引用的方式整体并入本文中)。在某些实施方案中,如本文所述的基因修饰小鼠是前述129品系与前述c57bl/6品系的混合物。在某些实施方案中,如本文所述的小鼠是前述129品系的混合物,或前述bl/6品系的混合物。在某些实施方案中,如本文所述的混合物的129品系是129s6(129/svevtac)品系。在一些实施方案中,如本文所述的小鼠是balb品系,例如balb/c品系。在一些实施方案中,如本文所述的小鼠是balb品系与另一前述品系的混合物。
[0442]
在一些实施方案中,如本文所述的非人类动物是大鼠。在某些实施方案中,如本文所述的大鼠选自威斯塔鼠(wistar rat)、lea品系、斯普雷格多利品系(sprague dawley strain)、费雪品系(fischer strain)、f344、f6和黑刺豚鼠(dark agouti)。在某些实施方案中,如本文所述的大鼠品系是两种或更多种选自由以下组成的组的品系的混合物:威斯塔鼠、lea、斯普雷格多利、费雪、f344、f6和黑刺豚鼠。
[0443]
大鼠多能和/或全能细胞可来自任何大鼠品系,包括例如aci大鼠品系(源自于八月(august)和哥本哈根(copenhagen)品系的近交系)、黑刺豚(da)大鼠品系、威斯塔大鼠品系、lea大鼠品系、斯普雷格多利(sd)大鼠品系或费雪大鼠品系,例如费雪f344或费雪f6。大鼠多能和/或全能细胞还可以从源自于上述两种或更多种品系的混合物的品系获得。举例来说,大鼠多能和/或全能细胞可来自da品系或aci品系。aci大鼠品系的特征在于具有黑色环纹,具有白色腹部和足以及rt1av1单倍型。这种品系可从多种来源获得,包括harlan laboratories。来自aci大鼠的大鼠es细胞系的一实例是aci.g1大鼠es细胞。da大鼠品系的特征在于具有环纹皮毛和rt1av1单倍型。这种大鼠可从多种来源获得,包括charles river和harlan laboratories。来自da大鼠的大鼠es细胞系的实例是da.2b大鼠es细胞系和da.2c大鼠es细胞系。在一些实施方案中,大鼠多能和/或全能细胞来自近交大鼠品系(参见例如美国专利申请公布no.2014
‑
0235933 a1,2014年8月21日公开,以引用的方式整体并入本文中)。关于使用如本文所述的方法和/或构建体在大鼠基因组中(例如在大鼠es细胞中)制造修饰的指导可见于例如美国专利申请公布no.2014
‑
0310828和2017
‑
0204430;均以引用的方式整体并入本文中。
[0444]
使用所提供的非人类动物、细胞或组织的方法
[0445]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织可用作研发抗体的平台。具体地说,如本文所述的非人类动物、非人类细胞和非人类组织代表了特别有利于产生和鉴定与从有限人类λ轻链谱系表达的人类λ轻链可变结构域配对的人类重链可变结构域和包括这种人类重链可变结构域的抗体的平台。这种人类重链可变结构域可用于例如产生双特定抗原结合蛋白。
[0446]
因此,本公开提供了本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织可用于制造抗体的方法中。根据本公开制造的抗体可包括例如人类抗体、嵌合抗体、反向嵌合抗体、这些抗体中的任一抗体的片段或它们的组合。
[0447]
在一些实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、
非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织可用于制造人类抗体(例如完全人类抗体),所述人类抗体包含源自于由如本文所述的非人类动物的细胞的遗传物质编码的核酸序列的可变结构域。在一些实施方案中,用所关注抗原对如本文所述的非人类动物(例如基因修饰啮齿动物,例如基因修饰大鼠或小鼠)进行免疫接种,条件和时间足以使非人类动物显现对所关注抗原的免疫反应。从进行免疫接种的非人类动物(或一个或多个细胞,例如一个或多个b细胞)分离和/或鉴定抗体和/或抗体序列(即,编码抗体一部分的序列,例如可变区序列)且使用测量例如亲和力、特异性、表位定位、阻断配体
‑
受体相互作用的能力、抑制受体活化等多种测定法表征。在多个实施方案中,由如本文所述的非人类动物、非人类细胞和非人类组织产生的抗体包含源自于从非人类动物、非人类细胞或非人类组织分离的一个或多个人类可变区核苷酸序列的一个或多个人类可变区。在一些实施方案中,可在如本文所述的非人类动物、非人类细胞和非人类组织中激起抗药物抗体(例如抗独特型抗体)。在多个实施方案中,由如本文所述的非人类动物、非人类细胞和非人类组织产生的抗体是包括人类轻链可变结构域和非人类(例如啮齿动物,例如大鼠或小鼠)轻链恒定结构域和/或人类重链可变结构域和非人类(例如啮齿动物,例如大鼠或小鼠)重链恒定结构域的反向嵌合抗体。
[0448]
在多个实施方案中,由非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织产生的抗体包括具有人类可变结构域和非人类恒定结构域的重链和轻链。在一些实施方案中,由如本文所述的非人类动物、非人类细胞和非人类组织产生的抗体是包括人类轻链可变结构域和非人类(例如啮齿动物)轻链恒定结构域的反向嵌合抗体。在一些实施方案中,由如本文所述的非人类动物、非人类细胞和非人类组织产生的抗体是包括人类重链可变结构域和非人类(例如啮齿动物,例如大鼠或小鼠)重链恒定结构域的反向嵌合抗体。
[0449]
在一些实施方案中,所提供的方法包括用所关注抗原对如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)进行免疫接种。在一些实施方案中,所提供的方法包括从所述非人类动物鉴定淋巴细胞(例如通过克隆选择的淋巴细胞),其中淋巴细胞表达结合(例如特异性结合)所关注抗原的抗体。在一些实施方案中,淋巴细胞是b细胞。在一些实施方案中,从淋巴细胞(例如b细胞)获得和/或鉴定(例如基因分型,例如测序)人类重链可变区序列和/或人类λ轻链可变区序列。在一些实施方案中,从淋巴细胞(例如b细胞)获得和/或鉴定(例如测序)人类重链可变结构域和/或人类λ轻链可变结构域的氨基酸序列。在一些实施方案中,来自非人类动物的b细胞的人类重链可变区序列和/或人类λ轻链可变区序列在宿主细胞中表达。在一些实施方案中,来自非人类动物的b细胞的人类重链可变区序列和/或人类λ轻链可变区序列的变体在宿主细胞中表达。在一些实施方案中,变体包括一个或多个突变。在一些实施方案中,一个或多个突变可改善包括变体在内的抗体的药物动力学和/或药效学性质。在一些实施方案中,一个或多个突变可改善包括变体在内的抗体的特异性、亲和力和/或免疫原性。
[0450]
在一些实施方案中,制造人类抗体的方法包括从本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织鉴定编码人类免疫球蛋白重链可变结构域和/或人类免疫球蛋白轻链可变结构域的核苷酸序列;以及(i)将编码人类免疫球蛋白重链可变结构域的核
苷酸序列接合或连结至编码人类免疫球蛋白重链恒定结构域的核苷酸序列,从而形成编码完全人类免疫球蛋白重链的人类免疫球蛋白重链序列,和/或(ii)将编码人类免疫球蛋白λ轻链可变结构域的核苷酸序列接合或连结至编码人类免疫球蛋白λ轻链恒定结构域的核苷酸序列,从而形成编码完全人类免疫球蛋白λ轻链的人类免疫球蛋白λ轻链序列。在某些实施方案中,人类免疫球蛋白重链序列和人类免疫球蛋白λ轻链序列在细胞(例如宿主细胞、哺乳动物细胞)中表达以便表达完全人类免疫球蛋白重链和完全人类免疫球蛋白λ轻链并形成人类抗体。在一些实施方案中,从细胞或包括细胞的培养基分离人类抗体。
[0451]
在一些实施方案中,可以采用如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)用于制造双特异性抗体的方法,所述方法包括:(a)使第一如本文所述的基因修饰非人类动物与第一抗原的第一表位接触,所述非人类动物包含可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)恒定区的单一重排人类igλ轻链可变区;(b)使第二如本文所述的基因修饰非人类动物与第二抗原的第二表位接触,所述非人类动物包含可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)恒定区的单一重排人类igλ轻链可变区(例如与第一非人类动物中所存在相同的单一重排人类igλ轻链可变区);(c)从第一基因修饰非人类动物分离表达对第一抗原的第一表位具特异性的第一抗体的b细胞并确定第一抗体的第一人类免疫球蛋白重链可变结构域;(d)从第二基因修饰非人类动物分离表达对第二抗原的第二表位具特异性的第二抗体的b细胞并确定第二抗体的第二人类免疫球蛋白重链可变结构域;(e)将编码第一人类免疫球蛋白重链可变结构域的核苷酸序列可操作地连接至编码第一人类免疫球蛋白恒定结构域的核苷酸序列以产生编码第一人类重链的第一核苷酸序列;(f)将编码第二人类免疫球蛋白重链可变结构域的核苷酸序列可操作地连接至编码第二人类免疫球蛋白恒定结构域的核苷酸序列以产生编码第二人类重链的第二核苷酸序列;(g)在哺乳动物细胞中表达:(i)第一核苷酸序列;(ii)第二核苷酸序列;和(iii)第三核苷酸序列,所述第三核苷酸序列包含可操作地连接至人类免疫球蛋白λ轻链恒定区的单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
[0452]
在一些实施方案中,可以采用如本文所述的非人类(例如啮齿动物,例如大鼠或小鼠)细胞用于制造双特异性抗体的方法,所述方法包括:(a)在非人类细胞中表达:(i)第一核苷酸序列,所述第一核苷酸序列包含可操作地连接至第一人类免疫球蛋白恒定区的第一人类免疫球蛋白重链可变区;(ii)第二核苷酸序列,所述第二核苷酸序列包含可操作地连接至第二人类免疫球蛋白恒定区的第二人类免疫球蛋白重链可变区;以及(iii)第三核苷酸序列,所述第三核苷酸序列包含可操作地连接至人类免疫球蛋白λ轻链恒定区的人类免疫球蛋白λ轻链可变区;其中第一人类免疫球蛋白重链可变区编码在已经用第一抗原的第一表位进行免疫接种的第一基因修饰非人类动物中鉴定的第一人类重链可变结构域,且第二人类免疫球蛋白重链可变区编码在已经用第二抗原的第二表位进行免疫接种的第二基因修饰非人类动物中鉴定的第二人类重链可变结构域,其中第一和第二基因修饰非人类动物各自是如本文所述的基因修饰非人类动物,所述非人类动物包含可操作地连接至非人类(例如啮齿动物,例如大鼠或小鼠)恒定区的单一重排人类igλ轻链可变区;并且其中第三核苷酸的人类免疫球蛋白λ轻链可变区是单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。所鉴定的氨基酸或核酸用于制造含有本文所鉴定的可变结构域的抗体或抗原结合蛋白。
[0453]
可以采用如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织来鉴定编码由本文所述的非人类动物、非人类细胞或非人类组织产生的人类可变结构域的核苷酸或核酸序列,所述人类可变结构域例如作为针对表位或抗原的抗体的一部分。
[0454]
可以采用如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织来鉴定由本文所述的非人类动物、非人类细胞或非人类组织产生的人类可变结构域的氨基酸序列,所述人类可变结构域例如作为针对表位或抗原的抗体的一部分。
[0455]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织提供了改良的体内系统和生物材料来源(例如细胞、核苷酸、多肽、蛋白质复合物),用于产生可用于多种测定法的人类抗体。在多个实施方案中,如本文所述的非人类动物、非人类细胞和非人类组织用于研发治疗剂,所述治疗剂靶向所关注多肽(例如跨膜或分泌多肽)和/或调节一种或多种与所述所关注多肽相关的活性和/或调节所述所关注多肽与其它结合搭配物(例如配体或受体多肽)的相互作用。举例来说,在多个实施方案中,如本文所述的非人类动物、非人类细胞和非人类组织用于研发治疗剂,所述治疗剂靶向一种或多种受体多肽,调节受体多肽活性和/或调节受体多肽与其它结合搭配物的相互作用。在多个实施方案中,如本文所述的非人类动物、非人类细胞和非人类组织用于鉴定、筛选和/或研发结合一种或多种所关注多肽的候选治疗剂(例如抗体、scfv等)。在多个实施方案中,如本文所述的非人类动物、非人类细胞和非人类组织用于筛选和研发阻断一种或多种所关注多肽的活性或阻断一种或多种所关注受体多肽的活性的候选治疗剂(例如抗体、scfv等)。在多个实施方案中,如本文所述的非人类动物、非人类细胞和非人类组织用于确定一种或多种所关注多肽的拮抗剂和/或激动剂的结合特征。在一些实施方案中,如本文所述的非人类动物、非人类细胞和非人类组织用于确定一种或多种候选治疗性抗体的结合一种或多种所关注多肽的表位。
[0456]
在多个实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织用于确定一种或多种候选人类抗体的药物动力学特征。在多个实施方案中,一个或多个如本文所述的非人类动物、非人类细胞和非人类组织和一个或多个对照或参考非人类动物、非人类细胞和非人类组织各自暴露于各种剂量的一种或多种候选人类抗体(例如0.1mg/kg、0.2mg/kg、0.3mg/kg、0.4mg/kg、0.5mg/kg、1mg/kg、2mg/kg、3mg/kg、4mg/kg、5mg/mg、7.5mg/kg、10mg/kg、15mg/kg、20mg/kg、25mg/kg、30mg/kg、40mg/kg或50mg/kg或更多)。候选治疗性抗体可经由包括肠胃外和非肠外的任何期望施用途径给与如本文所述的非人类动物。肠胃外途径包括例如静脉内、动脉内、门静脉内、肌肉内、皮下、腹膜内、脊柱内、鞘内、脑室内、颅内、胸膜内或其它注射途径。非肠外途径包括例如经口、经鼻、经皮、经肺、经直肠、经颊、经阴道、经眼。施用还可以通过连续输注、局部施用、从植入物(凝胶、膜等)持续释放和/或静脉内注射。在多个时间点(例如0小时、6小时、1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天或长达30天或更多天)从非人类动物(人源化和对照)分离血液。可以使用从如本文所述的非人类动物、非人类细胞和非人类组织获得的样品执行多种测定法以确定所施用的候选治疗性抗体的药物动力学特征,包括但不限于总igg、抗治疗性抗体反应、
凝集等。
[0457]
在多个实施方案中,如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞和非人类(例如啮齿动物,例如大鼠或小鼠)组织用于测量:阻断或调节所关注多肽的活性的治疗作用;以及细胞改变或在受体多肽的情况下非人类动物、非人类细胞或非人类组织中细胞表面上受体多肽的密度对基因表达的影响。在多个实施方案中,如本文所述的非人类动物(或从其分离的细胞)、非人类细胞或非人类组织暴露于结合所关注多肽的候选治疗剂,并在后续时间段之后,分析对与所关注多肽相关的特定细胞过程,例如配体
‑
受体相互作用或信号转导的影响。
[0458]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)表达人类抗体可变区,因此,可产生细胞、细胞系和细胞培养物,用作结合和功能测定法中使用的抗体可变区的来源,所述测定法例如测定拮抗剂或激动剂的结合或功能,尤其是拮抗剂或激动剂对所关注人类抗原具有特异性或对在配体
‑
受体相互作用(结合)中起作用的表位具有特异性的情况下。在多个实施方案中,可以使用从如本文所述的非人类动物分离的细胞确定被候选治疗性抗体或片段(例如fab片段,由vh、vl、ch1和cl结构域组成的单价片段;(ii)f(ab')2片段,包含两个由铰链区二硫桥连接的fab片段的二价片段;(iii)fd片段,由vh和ch1结构域组成;(iv)fv片段,由抗体单臂的vh和vl结构域组成;(v)dab片段(ward等人,(1989)nature 341:544
‑
546),其包含单个可变结构域;和(vi)分离的互补决定区(cdr))结合的表位。
[0459]
可以从所提供的非人类动物(例如啮齿动物,例如大鼠或小鼠)分离细胞并根据具体情况使用,或可维持培养状态,历经许多代。在多个实施方案中,使来自所提供的非人类动物的细胞永生化(例如经由使用病毒)并且无限期地维持培养状态(例如连续培养)。
[0460]
在一些实施方案中,非人类(例如啮齿动物,例如大鼠或小鼠)细胞是非人类淋巴细胞。在一些实施方案中,非人类细胞选自b细胞、树突状细胞、巨噬细胞、单核细胞和t细胞。在一些实施方案中,非人类细胞是未成熟b细胞、成熟幼稚b细胞、激活b细胞、记忆b细胞和/或血浆细胞。
[0461]
在一些实施方案中,非人类(例如啮齿动物,例如大鼠或小鼠)细胞是非人类胚胎干(es)细胞。在一些实施方案中,非人类es细胞是啮齿动物es细胞。在某些实施方案中,啮齿动物es细胞是小鼠es细胞并且来自129品系、c57bl品系、balb/c或其混合物。在某些实施方案中,啮齿动物胚胎干细胞是小鼠胚胎干细胞并且是129和c57bl品系的混合物。在某些实施方案中,啮齿动物胚胎干细胞是小鼠胚胎干细胞并且是129、c57bl和balb/c品系的混合物。
[0462]
在一些实施方案中,提供如本文所述的非人类(例如啮齿动物,例如大鼠或小鼠)es细胞用于制造非人类动物的用途。在某些实施方案中,非人类es细胞是小鼠es细胞并且用于制造包含如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的小鼠。在某些实施方案中,非人类es细胞是大鼠es细胞并且用于制造包含如本文所述的包括有限人类λ轻链可变区谱系的工程化免疫球蛋白κ轻链基因座的大鼠。
[0463]
在一些实施方案中,非人类(例如啮齿动物,例如大鼠或小鼠)组织选自脂肪、膀胱、脑、乳房、骨髓、眼睛、心脏、肠、肾、肝、肺、淋巴结、肌肉、胰腺、血浆、血清、皮肤、脾、胃、胸腺、睾丸、卵子和它们的组合。
[0464]
在一些实施方案中,提供了一种永生化细胞,所述永生化细胞是从如本文所述的分离的非人类细胞或组织制造、产生(generated)、产生(produced)或获得。
[0465]
在一些实施方案中,提供了非人类(例如啮齿动物,例如大鼠或小鼠)胚胎,所述非人类胚胎是从如本文所述的非人类es细胞制造、产生(generated)、产生(produced)或获得。在某些实施方案中,非人类胚胎是啮齿动物胚胎;在一些实施方案中,小鼠胚胎;在一些实施方案中,大鼠胚胎。
[0466]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)提供了一种体内系统,所述体内系统用于产生结合所关注多肽的人类抗体可变区的变体(例如人类vλ结构域变体)。这种变体包括具有所需功能性、特异性、与所关注多肽的两种或更多种变体所共有的常见表位的交叉反应性低的人类抗体可变区。在一些实施方案中,如本文所述的非人类动物用于产生人类抗体可变区小组,所述人类抗体可变区小组含有针对期望或改良功能性进行筛选的一系列变异可变区。
[0467]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)提供了一种体内系统,所述体内系统用于产生人类抗体可变区文库(例如人类vλ结构域文库)。这种文库提供了重链和/或轻链可变区序列的一种来源,所述重链和/或轻链可变区序列可基于所需效应功能被移植至不同fc区上,用作使用所属领域中已知的技术(例如定点诱变、易错pcr等)对可变区序列进行亲和力成熟的来源,和/或用作用于产生例如以下的抗体类治疗性分子的抗体组分的来源:嵌合抗原受体(即使用抗体组分进行工程化的分子,例如scfv)、多特异性结合剂(例如双特异性结合剂)和融合蛋白(例如单结构域抗体、scfv等)。
[0468]
在一些实施方案中,提供了一种在非人类(例如啮齿动物,例如大鼠或小鼠)动物中产生抗体的方法,所述方法包括以下步骤:(a)用所关注抗原对如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)进行免疫接种;(b)将非人类动物(例如啮齿动物,例如大鼠或小鼠)维持在足以使非人类动物对所关注抗原产生免疫反应(例如表达抗体)的条件下;以及(c)从非人类动物(例如啮齿动物,例如大鼠或小鼠)或非人类动物(例如啮齿动物,例如大鼠或小鼠)的细胞(例如b细胞)回收结合所关注抗原的抗体。
[0469]
在非人类动物中产生抗体的方法的一些实施方案中,非人类细胞是b细胞。在非人类动物中产生抗体的方法的一些实施方案中,非人类细胞是杂交瘤。
[0470]
在一些实施方案中,提供了通过一种方法制备的抗体,所述方法包括以下步骤:(a)提供如本文所述的非人类动物;(b)用所关注抗原对非人类动物进行免疫接种;(c)将非人类动物维持在足以使非人类动物对所关注抗原产生免疫反应的条件下;以及(d)从非人类动物或非人类细胞回收结合所关注抗原的抗体,其中(d)的抗体包括人类v
h
和vλ结构域。
[0471]
在通过一种方法制备的抗体的一些实施方案中,由重排人类重链可变区编码的人类重链可变结构域包含人类v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2 v
h6‑
1或其体细胞超突变变体。
[0472]
在通过一种方法制备的抗体的一些实施方案中,从如本文所述的包含有限人类免疫球蛋白λ轻链可变区谱系的非人类动物获得的人类λ轻链可变结构域由包含人类vλ4
‑
69、
vλ8
‑
61、vλ4
‑
60、vλ6
‑
57、vλ10
‑
54、vλ5
‑
52、vλ1
‑
51、vλ9
‑
49、vλ1
‑
47、vλ7
‑
46、vλ5
‑
45、vλ1
‑
44、vλ7
‑
43、vλ1
‑
40、vλ5
‑
39、vλ5
‑
37、vλ1
‑
36、vλ3
‑
27、vλ3
‑
25、vλ2
‑
23、vλ3
‑
22、vλ3
‑
21、vλ3
‑
19、vλ2
‑
18、vλ3
‑
16、vλ2
‑
14、vλ3
‑
12、vλ2
‑
11、vλ3
‑
10、vλ3
‑
9、vλ2
‑
8、vλ4
‑
3vλ3
‑
1或其体细胞超突变变体的重排人类λ轻链可变区编码。
[0473]
在一些实施方案中,提供了如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,用于制造和/或研发用于治疗或诊断的药物(例如抗体或其片段)。
[0474]
在一些实施方案中,提供了如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织,用于制造用以治疗、预防或改善疾病、病症或疾患的药剂。
[0475]
在一些实施方案中,提供了如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织用于制造和/或研发用于医学中,例如用作药剂的药物或疫苗的用途。
[0476]
在一些实施方案中,提供了如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)、非人类(例如啮齿动物,例如大鼠或小鼠)细胞或非人类(例如啮齿动物,例如大鼠或小鼠)组织用于制造和/或研发抗体或其片段的用途。
[0477]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)提供了一种用于分析和测试药物或疫苗的体内系统。在多个实施方案中,候选药物或疫苗可递送至一个或多个如本文所述的非人类动物,接着监测所述非人类动物以确定对药物或疫苗的一个或多个免疫反应、药物或疫苗的安全特征或对疾病或疾患和/或疾病或疾患的一种或多种症状的作用。用于确定安全特征的示例性方法包括测量毒性、最佳剂量浓度、抗体(即抗药)反应、药物或疫苗的功效和可能的风险因素。在可以在这种非人类动物中改良和/或研发这类药物或疫苗。
[0478]
疫苗功效可以用多种方式确定。简单地说,使用所属领域已知的方法对如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)进行疫苗接种且接着用疫苗激发,或向已感染的非人类动物施用疫苗。可以通过对非人类动物(或从其分离的细胞)进行监测和/或进行一个或多个测定来测量非人类动物对疫苗的反应,以确定疫苗的功效。接着使用所属领域中已知和/或本文所述的一个或多个量度,将非人类动物对疫苗的反应与对照动物进行比较。
[0479]
疫苗功效可以进一步通过病毒中和测定法来确定。简单地说,对如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)进行免疫接种并在免疫接种后的多天收集血清。将血清的连续稀释液与病毒一起预先孵育,在此期间血清中对病毒具有特异性的抗体将结合于病毒。接着将病毒/血清混合物添加至允许细胞中以通过空斑测定法或微量中和测定法来确定感染性。如果血清中的抗体中和病毒,那么与对照组相比空斑较少或相对荧光素酶单位较低。
[0480]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)产生人类抗体可变区,因此提供了一种用于产生供诊断应用(例如免疫学、血清学、微生物学、细胞病理学等)中使用的人类抗体的体内系统。在多个实施方案中,如本文所述的非人类动物可用于产生结合相关抗原位点的人类抗体可变区,以鉴定细胞变化,例如指示病理学变化的特定细胞
表面标记的表达。这种抗体可与多种化学实体(例如放射性示踪剂)缀合并且根据需要用于多种体内和/或体外测定法中。
[0481]
如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)提供了一种改良的体内系统,用于研发和选择用于肿瘤学和/或感染性疾病的人类抗体。在多个实施方案中,如本文所述的非人类动物和对照非人类动物(例如具有与本文所述不同的基因修饰或无基因修饰,即野生型)可植入肿瘤(或肿瘤细胞)或感染病毒(例如流感、hiv、hcv、hpv等)。在植入感染后,可以向非人类动物施用候选治疗剂。在施用候选治疗剂之前可留有足够的时间在非人类动物内的一个或多个位置建立肿瘤或病毒。可替代地和/或另外,可在这种非人类动物中监测免疫反应,以便表征和选择可发展成治疗剂的潜在人类抗体。
[0482]
结合超过一个表位的抗原结合蛋白
[0483]
本文所述的组合物和方法可用于制造结合超过一个表位的结合蛋白,例如双特异性抗体。本发明的优点包括能够选择各自将与单个轻链缔合的适当高结合水平(例如亲和力成熟)的重链免疫球蛋白链。
[0484]
双特异性结合蛋白的合成和表达是有问题的,这在一定程度上归因于与鉴定能够与两条不同重链缔合并一起表达的合适轻链有关的问题,并且在一定程度上归因于分离的问题。本文所述的方法和组合物允许基因修饰非人类动物(例如啮齿动物,例如大鼠或小鼠)通过另外的天然方法来选择包括人类λ轻链可变结构域的合适轻链,所述轻链能够与超过一条重链,包括进行体细胞超突变(例如亲和力成熟)的重链缔合并一起表达。可以从表达具有反向嵌合重链(即,人类可变结构域和非人类(例如啮齿动物,例如大鼠或小鼠)恒定结构域)的亲和力成熟抗体的如本文所述的免疫接种小鼠的合适b细胞鉴定人类vλ和v
h
序列,并在表达载体中与合适人类恒定区基因序列(例如人类igg1)在框内克隆。可制备两种这样的构建体,其中每种构建体编码结合不同表位的人类重链可变结构域。包括具有种系序列的基因区段的重排人类λ轻链可变区(例如人类vλ1
‑
51/jλ2或人类vλ2
‑
14/jλ2)或其体细胞超突变型式(例如如从b细胞鉴定)可与合适人类恒定区基因(例如人类λ恒定基因)在框内融合。这三种完全人类重链和轻链构建体可处于合适的细胞中来表达。细胞将表达两个主要种类:同源二聚体重链与相同轻链,和异源二聚体重链与相同轻链。为了便于分离这些主要种类,对重链之一进行修饰以省略结合蛋白a的决定子,使同源二聚体结合蛋白与异源二聚体结合蛋白的亲和力有差别。解决这个问题的组合物和方法描述于美国专利no.8,586,713、9,309,326和9,982,013中,各专利以引用的方式并入本文中。
[0485]
在一些实施方案中,提供了一种如本文所述的抗原结合蛋白,其中人类vλ和v
h
序列源自于已经用包含所关注表位的抗原或所关注抗原进行免疫接种的本文所述的小鼠。
[0486]
在一些实施方案中,提供了一种抗原结合蛋白,所述抗原结合蛋白包含第一和第二多肽,第一多肽从n端至c端包含选择性结合第一表位的第一抗原结合结构域,接着是包含选自igg1、igg2和igg4的人类igg的第一c
h
3结构域的恒定结构域;并且第二多肽从n端至c端包含选择性结合第二表位的第二抗原结合结构域,接着是包含选自igg1、igg2和igg4的人类igg的第二c
h
3结构域的恒定结构域,其中第二c
h
3结构域包含减少或消除第二c
h
3结构域与蛋白a的结合的修饰。
[0487]
在一些实施方案中,提供了一种抗原结合蛋白,所述抗原结合蛋白包含第一重链和第二重链。在一些实施方案中,第一重链从n端至c端包含选择性结合第一表位的第一人
类重链可变结构域(与人类λ轻链可变结构域),接着是包含选自igg1、igg2和igg4的人类igg的第一c
h
3结构域的恒定结构域。在一些实施方案中,第二重链从n端至c端包含选择性结合第二表位(其可在与第一表位相同或不同的抗原上)的第二人类重链可变结构域(与相同人类λ轻链可变结构域),接着是包含选自igg1、igg2和igg4的人类igg的第二c
h
3结构域的恒定结构域,其中第二c
h
3结构域包含减少或消除第二c
h
3结构域与蛋白a的结合的修饰。
[0488]
在一些实施方案中,c
h
3结构域包含减少或消除c
h
3结构域与蛋白a的结合的修饰。在一些实施方案中,减少或消除c
h
3结构域与蛋白a的结合的修饰包含h95r修饰(通过imgt外显子编号;通过eu编号,为h435r)。在一些实施方案中,减少或消除c
h
3结构域与蛋白a的结合的修饰包含y96f修饰(imgt;通过eu,为y436f)。在一些实施方案中,减少或消除c
h
3结构域与蛋白a的结合的修饰包含h95r修饰(通过imgt外显子编号;通过eu编号,为h435r)和y96f修饰(imgt;通过eu,为y436f)。
[0489]
在一些实施方案中,c
h
3结构域是包含h95r修饰(通过imgt外显子编号;通过eu编号,为h435r)、y96f修饰(imgt;通过eu,为y436f)或两者的igg1结构域,并且还包含d16e修饰、l18m修饰、n44s修饰、k52n修饰、v57m修饰、v82i修饰或它们的组合(imgt;通过eu,为d356e、l358m、n384s、k392n、v397m、v422i或它们的组合)。
[0490]
在一些实施方案中,c
h
3结构域是包含h95r修饰(通过imgt外显子编号;通过eu编号,为h435r)、y96f修饰(imgt;通过eu,为y436f)或两者的igg2结构域,并且还包含n44s修饰、k52n修饰、v82i修饰或它们的组合(imgt;通过eu,为n384s、k392n、v422i或它们的组合)。
[0491]
在一些实施方案中,c
h
3结构域是包含h95r修饰(通过imgt外显子编号;通过eu编号,为h435r)、y96f修饰(imgt;通过eu,为y436f)或两者的igg4结构域,并且还包含q15r修饰、n44s修饰、k52n修饰、v57m修饰、r69k修饰、e79q修饰和v82i修饰(imgt;通过eu,为q355r、n384s、k392n、v397m、r409k、e419q、v422i或它们的组合)。
[0492]
一种制造结合超过一个表位的抗原结合蛋白的方法是用包含所关注第一表位的第一抗原对如本文所述的具有有限人类免疫球蛋白λ轻链可变区谱系的第一非人类动物(例如啮齿动物,例如大鼠或小鼠)进行免疫接种,其中所述非人类动物(例如啮齿动物,例如大鼠或小鼠)包括可操作地连接至非人类免疫球蛋白重链恒定区的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d基因区段和一个或多个未重排人类j
h
基因区段。当进行免疫接种时,这种非人类动物将制造反向嵌合抗体,所述反向嵌合抗体包含从有限人类免疫球蛋白λ轻链可变区谱系表达的人类λ轻链可变结构域(例如单个人类重排λ轻链可变区)。可鉴定编码结合所关注表位的人类免疫球蛋白重链可变结构域的b细胞。在一些实施方案中,可提取编码人类免疫球蛋白重链可变结构域的核苷酸序列(例如通过pcr)并与合适的人类免疫球蛋白重链恒定结构域在框内克隆至表达构建体中。在一些实施方案中,可以重复这个过程以鉴定结合第二抗原的第二表位的第二人类免疫球蛋白重链可变结构域,并且可提取第二人类免疫球蛋白重链可变结构域且与合适的第二免疫球蛋白恒定区在框内克隆至表达载体中。在一些实施方案中,可提取编码人类免疫球蛋白λ轻链可变结构域的核苷酸序列(例如通过pcr)并与合适的人类免疫球蛋白λ轻链恒定区在框内克隆至表达构建体中。在一些实施方案中,第一和第二免疫球蛋白人类重链恒定结构域可以是相同或不同的同种型。在一些实施方案中,免疫球蛋白人类重链恒定结构域之一(但另一个不这
样)可以如本文或美国专利no.8,586,713、9,309,326和9,982,013中所述来修饰,各专利以引用的方式并入本文中。在一些实施方案中,抗原结合蛋白可以在合适的细胞中表达并基于它与同源二聚体抗原结合蛋白相比有差异的对蛋白a的亲和力来分离,例如如美国专利no.8,586,713、9,309,326和9,982,013中所述,各专利以引用的方式并入本文中。在一些实施方案中,第一抗原与第二抗原可以相同且第一表位与第二表位可以不同。在一些实施方案中,第一抗原与第二抗原不同。在一些实施方案中,双特异性抗原结合蛋白结合抗原的亲和力比单特异性抗原结合蛋白结合相同抗原的亲和力高。在一些实施方案中,双特异性抗原结合蛋白以不同亲和力结合第一表位和第二表位。
[0493]
在一些实施方案中,提供了一种制备双特异性抗原结合蛋白的方法,所述方法包括从如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)鉴定第一亲和力成熟的(例如包含一个或多个体细胞超突变)人类重链可变结构域,所述非人类动物包括在内源κ轻链基因座处的单个人类重排λ轻链可变区,和可操作地连接至非人类免疫球蛋白重链恒定区的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。在一些实施方案中,制备双特异性抗原结合蛋白的方法包括从如本文所述的非人类动物(例如啮齿动物,例如大鼠或小鼠)鉴定第二亲和力成熟的(例如包含一个或多个体细胞超突变)重链可变结构域,所述非人类动物包括在内源κ轻链基因座处的单个人类重排λ轻链可变区(例如与用于产生第一可变结构域的非人类动物中相同的单个人类重排λ轻链可变区),和可操作地连接至非人类免疫球蛋白重链恒定区的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。在一些实施方案中,编码第一亲和力成熟的人类重链可变结构域的核苷酸序列与缺乏蛋白a决定子修饰的人类重链恒定区在框内克隆,以形成编码完全人类重链的第一核苷酸序列,如美国专利no.8,586,713、9,309,326和9,982,013中所述,各专利以引用的方式并入本文中。在一些实施方案中,编码第二亲和力成熟的人类重链可变结构域的核苷酸序列与包含蛋白a决定子的人类重链恒定区在框内克隆,以形成第二完全人类重链,如美国专利no.8,586,713、9,309,326和9,982,013中所述,各专利以引用的方式并入本文中。在一些实施方案中,将如本段中所述的编码第一完全人类重链的核苷酸和如本段中所述的编码第二完全人类重链的核苷酸引入宿主细胞(例如哺乳动物细胞)中。在一些实施方案中,在第一表达载体上包括如本段中所述的编码第一完全人类重链的核苷酸。在一些实施方案中,在第二表达载体上包括如本段中所述的编码第二完全人类重链的核苷酸。在一些情况下,第一表达载体与第二表达载体相同或不同。在一些实施方案中,制造双特异性抗体的方法包括在宿主细胞中表达如本段中所述的编码第一完全人类重链的核苷酸、如本段中所述的编码第二完全人类重链的核苷酸和编码包含人类λ轻链可变结构域的人类λ轻链的核苷酸,以便形成双特异性抗体,其中人类λ轻链可变结构域具有由如本文所述的具有有限人类λ轻链可变区谱系的非人类动物产生的序列(例如单个人类重排λ轻链可变区)。在其它实施方案中,人类λ轻链可变结构域具有源自于与非人类动物中所存在相同的人类v/j重排(例如与非人类动物中所存在相同的单个人类重排λ轻链可变区)的序列。在一些实施方案中,制造双特异性抗体的方法包括例如从宿主细胞或其培养基获得双特异性抗体。
[0494]
在一些实施方案中,宿主细胞可以是原核生物和真核生物的细胞(单细胞或多细胞)、细菌细胞(例如大肠杆菌、芽孢杆菌属、链霉菌属等菌株)、分枝杆菌属细胞、真菌细胞、
酵母细胞(例如酿酒酵母、粟酒裂殖酵母、巴氏毕赤酵母、甲醇毕赤酵母等)、植物细胞、昆虫细胞(例如sf
‑
9、sf
‑
21、杆状病毒感染的昆虫细胞、粉纹夜蛾等)、非人类动物细胞、人类细胞或细胞融合物,例如杂交瘤或四价体瘤。在一些实施方案中,细胞是人、猴、猿、仓鼠、大鼠或小鼠细胞。在一些实施方案中,细胞是真核的并且选自以下细胞:cho(例如cho k1、dxb
‑
11cho、veggie
‑
cho)、cos(例如cos
‑
7)、视网膜细胞、vero、cv1、肾(例如hek293、293ebna、msr 293、mdck、hak、bhk)、hela、hepg2、wi38、mrc 5、colo205、hb 8065、hl
‑
60、(例如bhk21)、jurkat、daudi、a431(表皮)、cv
‑
1、u937、3t3、l细胞、c127细胞、sp2/0、ns
‑
0、mmt 060562、sertoli细胞、brl 3a细胞、ht1080细胞、骨髓瘤细胞、肿瘤细胞和源自于前述细胞的细胞系。在一些实施方案中,细胞包含一个或多个病毒基因,例如表达病毒基因的视网膜细胞(例如细胞)。
[0495]
药物组合物
[0496]
在一些实施方案中,可以向受试者(例如人类受试者)施用由本文公开的非人类动物产生或源自于由本文公开的非人类动物(例如啮齿动物,例如大鼠或小鼠)产生的抗体、核酸或其治疗相关部分的抗原结合蛋白、编码抗原结合蛋白的核酸或其治疗相关部分。在一些实施方案中,药物组合物包括由本文公开的非人类动物产生的抗体。在一些实施方案中,药物组合物可包括缓冲剂、稀释剂、赋形剂或它们的任何组合。在一些实施方案中,必要时,组合物还可以含有一种或多种额外的治疗活性物质。
[0497]
虽然对本文提供的药物组合物的描述主要是针对适合于伦理上可施用于人类的药物组合物,但熟练技术人员将了解,这种组合物一般适合于施用于各种动物。众所周知,为了使组合物适合于施用于多种动物,可对适合于施用于人类的药物组合物进行修改,并且一般熟练兽医学药理学家可利用常规实验(如果有的话)设计和/或进行这种修改。
[0498]
举例来说,本文提供的药物组合物可以呈无菌可注射形式(例如适合于皮下注射或静脉内输注的形式)。举例来说,在一些实施方案中,药物组合物呈适合于注射的液体剂型提供。在一些实施方案中,药物组合物任选地在真空下呈粉剂(例如冻干和/或杀菌)提供,在注射之前可以用水性稀释剂(例如水、缓冲液、盐溶液等)复原。在一些实施方案中,药物组合物在水、氯化钠溶液、乙酸钠溶液、苯甲醇溶液、磷酸盐缓冲盐水等中稀释和/或复原。在一些实施方案中,粉剂应轻轻地与水性稀释剂混合(例如不震荡)。
[0499]
本文所述的药物组合物的制剂可以通过药理学领域中已知或此后开发的任何方法制备。一般来说,这种制备方法包括如下步骤:使活性成分与稀释剂或另一赋形剂和/或一种或多种其它辅助剂缔合,接着必要和/或需要时,使产物成形和/或包装成所需的单剂量或多剂量单元。
[0500]
在一些实施方案中,包括由本文公开的非人类动物(例如啮齿动物,例如大鼠或小鼠)产生的抗体的药物组合物可包括在用于存储或施用的容器中,例如小瓶、注射器(例如iv注射器)或袋(例如iv袋)。根据本公开的药物组合物可以呈单个单位剂量和/或呈多个单位剂量制备、包装和/或整批出售。如本文所用,“单位剂量”是包含预先确定量的活性成分的药物组合物的离散量。活性成分的量一般等于将施用于受试者的活性成分的剂量,和/或这种剂量的合宜分数,例如这种剂量的一半或三分之一。
[0501]
根据本公开的药物组合物中活性成分、药学上可接受的赋形剂和/或任何额外成分的相对量将视所治疗的受试者的身份、体型和/或状态而变化,并且进一步视施用组合物
的途径而变化。举例来说,组合物可包含0.1%与100%(w/w)之间的活性成分。
[0502]
药物组合物可另外包含药学上可接受的赋形剂,如本文所用,药学上可接受的赋形剂包括适合于期望的具体剂型的任何和所有溶剂、分散介质、稀释剂或其它液体媒介物、分散或悬浮助剂、表面活性剂、等张剂、增稠或乳化剂、防腐剂、固体粘合剂、润滑剂等。remingto n's the science and practice of pharmacy,第21版,a.r.gennaro(lippincott,williams&wilkins,baltimore,md,2006)公开了多种用于配制药物组合物的赋形剂和已知的用于制备药物组合物的技术。除非例如因产生任何不良的生物效应或另外以有害的方式与药物组合物的任何其它组分相互作用,使得任何常规赋形剂介质与物质或其衍生物不相容,否则本公开的范围内涵盖赋形剂介质的使用。
[0503]
在一些实施方案中,药学上可接受的赋形剂是至少95%、至少96%、至少97%、至少98%、至少99%或100%纯。在一些实施方案中,赋形剂被批准用于人类和兽医学用途。在一些实施方案中,赋形剂被美国食品与药物管理局(the united states food and drug administration)批准。在一些实施方案中,赋形剂是医药级的。在一些实施方案中,赋形剂满足美国药典(the united states pharmacopoeia,usp)、欧洲药典(the european pharmacopoeia,ep)、英国药典(the british pharmacopoeia)和/或国际药典(the international pharmacopoeia)的标准。
[0504]
在制造药物组合物中使用的药学上可接受的赋形剂包括(但不限于)惰性稀释剂、分散和/或成粒剂、表面活性剂和/或乳化剂、崩解剂、粘合剂、防腐剂、缓冲剂、润滑剂和/或油类。这种赋形剂可任选地包括在药物制剂中。根据配制师的判断,组合物中可存在例如可可脂和栓剂蜡、着色剂、包衣剂、甜味剂、调味剂和/或芳香剂这样的赋形剂。
[0505]
在一些实施方案中,所提供的药物组合物包含一种或多种药学上可接受的赋形剂(例如防腐剂、惰性稀释剂、分散剂、表面活性剂和/或乳化剂、缓冲剂等)。在一些实施方案中,药物组合物包含一种或多种防腐剂。在一些实施方案中,药物组合物不含防腐剂。
[0506]
在一些实施方案中,药物组合物呈可冷藏和/或冷冻的形式提供。
[0507]
在一些实施方案中,药物组合物呈不可冷藏和/或冷冻的形式提供。在一些实施方案中,复原的溶液和/或液体剂型可在复原后存储达某一段时间(例如2小时、12小时、24小时、2天、5天、7天、10天、2周、一个月、两个月或更长时间)。在一些实施方案中,抗体组合物存储超过规定时间会引起抗体降解。
[0508]
液体剂型和/或复原的溶液在施用之前可能包含颗粒物和/或褪色。在一些实施方案中,如果褪色或混浊和/或如果在过滤后仍然有颗粒物,那么不应使用溶液。
[0509]
药剂的配制和/或制造中的一般考虑可见于例如remington:the science and practice of pharmacy第21版,lippincott williams&wilkins,2005。
[0510]
试剂盒
[0511]
本公开进一步提供了包装或试剂盒,所述包装或试剂盒包含一个或多个的容器,所述容器至少装有如本文所述的非人类细胞、蛋白质(单一或复杂(例如抗体或其片段))、dna片段、靶向载体或它们的任何组合。试剂盒可用于任何适用方法(例如研究法)中。任选地,通告可以与这类容器相联,所述通告呈管制药物或生物制品的制造、使用或销售的政府机构指定的形式,它反映了:(a)被制造、使用或销售的机构批准用于施用于人类,(b)使用说明,和/或(c)管理材料和/或生物制品(例如,如本文所述的非人类动物或非人类细胞)在
两个或更多个实体和它们的组合之间的转移的合同。
[0512]
在一些实施方案中,提供了包含如本文所述的非人类细胞、非人类组织、永生化细胞、非人类es细胞或非人类胚胎的试剂盒。在一些实施方案中,提供了包含来自如本文所述的非人类动物、非人类细胞、非人类组织、永生化细胞、非人类es细胞或非人类胚胎的氨基酸(例如抗体或其片段)的试剂盒。在一些实施方案中,提供了包含来自如本文所述的非人类动物、非人类细胞、非人类组织、永生化细胞、非人类es细胞或非人类胚胎的核酸(例如编码抗体或其片段的核酸)的试剂盒。在一些实施方案中,提供了包含从如本文所述的非人类动物、非人类细胞、非人类组织、永生化细胞、非人类es细胞或非人类胚胎鉴定的序列(氨基酸和/或核酸序列)的试剂盒。
[0513]
在一些实施方案中,提供了如本文所述的试剂盒,所述试剂盒用于制造和/或研发用以治疗或诊断的药物(例如抗体或其片段)。
[0514]
在一些实施方案中,提供了如本文所述的试剂盒,所述试剂盒用于制造和/或研发用以治疗、预防或改善疾病、病症或疾患的药物(例如抗体或其片段)。
[0515]
在以下对示例性实施方案的描述过程中某些实施方案的其它特征将变得显而易见,所述示例性实施方案出于说明而给出且不意图限制。
[0516]
实施例
[0517]
提供了以下实施例以便对熟练技术人员描述如何制造和使用本文所述的方法和组合物;且不意图限制本公开的范围。除非另外指示,否则温度以摄氏度指示且压力处于大气压下或靠近大气压。
[0518]
一般来说,使用技术制造携带重排人类λ轻链可变区的靶向载体(参见例如美国专利no.6,586,251和valenzuela等人(2003)high
‑
throughput engineering of the mouse genome coupled with high
‑
resolution expression analysis,nature biotech.21(6):652
‑
659,各以引用的方式并入本文中)。小鼠基因组的细菌人工染色体(bac)克隆被修饰成产生含有单一重排人类λ轻链可变区的基因组构建体。将所述构建体插入到事先被修饰成缺失内源vκ和jκ基因区段的内源κ轻链基因座中。
[0519]
实施例1
‑
包括重排人类vλ1
‑
51/jλ2和小鼠cκ的靶向载体的产生
[0520]
使用所属领域中公认的标准分子生物学技术制造重排人类vλ1
‑
51/jλ2。重排人类vλ1
‑
51/jλ2中所包括的人类vλ1
‑
51基因区段和人类jλ2基因区段分别具有人类vλ1
‑
51种系序列和人类jλ2种系序列。
[0521]
通过从头dna合成(blue heron biotech)制造dna片段。所述片段含有asci位点、2kb人类vλ1
‑
51启动子区域、人类vλ1
‑
51 5'非翻译区(utr)、人类vλ1
‑
51基因区段的外显子1的编码序列(其包括编码前导肽的序列)、人类vλ1
‑
51基因区段的内含子1、包括人类vλ1
‑
51基因区段的第二外显子和jλ2基因区段的重排人类vλ1
‑
51/jλ2可变序列、人类jκ5
‑
cκ内含子的开始412bp(包括jκ5剪接供体位点(sd))和pi
‑
scei位点(seq id no:31;又参见图1a和图9a
‑
c)。
[0522]
将上一段中描述的片段连结至先前描述的靶向载体中以产生图1a中的靶向载体a。先前描述的靶向载体含有:源自于bac ct7
‑
302g12的约23kb 5'小鼠同源臂、frt
‑
ub
‑
neo
‑
frt盒、asci位点、重排人类vκ3
‑
20/jκ1可变区、pi
‑
scei位点和源自于bac ct7
‑
254m04的约75kb 3'小鼠同源臂(参见例如美国专利no.9,334,334的图14b,和美国专利no.10,
143,186的图2,各以引用的方式并入本文中)。所述小鼠同源臂与先前macdonald等人(2014)proc.natl.acad.sci.usa,111:5147
‑
52描述的同源臂相同,所述文献以引用的方式并入本文中。5'臂在内源κ基因座上游含有约23kb基因组序列,且3'臂在小鼠κ基因座下游含有约2.4kb小鼠jκ
‑
cκ内含子、小鼠cκ外显子和约72kb基因组序列。参见图1a。
[0523]
靶向载体a从5'至3'含有源自于bac ct7
‑
302g12的约23kb 5'小鼠同源臂、frt
‑
ub
‑
neo
‑
frt盒、asci位点、2kb人类vλ1
‑
51启动子区域、人类vλ1
‑
51 5'非翻译区(utr)、人类vλ1
‑
51基因区段的外显子1的编码序列(其包括编码前导肽的序列)、人类vλ1
‑
51基因区段的内含子1、包括人类vλ1
‑
51基因区段的第二外显子和jλ2基因区段的重排人类vλ1
‑
51/jλ2可变序列、人类jκ5
‑
cκ内含子的开始412bp(包括jκ5剪接供体位点(sd))、pi
‑
scei位点和源自于bac ct7
‑
254m04的约75kb 3'小鼠同源臂,所述3'小鼠同源臂在小鼠κ基因座下游包括约2.4kb小鼠jκ
‑
cκ内含子、小鼠cκ外显子和约72kb基因组序列。
[0524]
实施例2
‑
包括重排人类vλ1
‑
51/jλ2和小鼠cκ的es细胞的产生
[0525]
如实施例1中所述的靶向载体a被电穿孔至小鼠es细胞中,在所述小鼠es细胞中内源vκ和jκ基因区段已经被重排人类vκ3
‑
20/jκ1替代,并且其中免疫球蛋白重链基因座包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段。小鼠es细胞的免疫球蛋白重链基因座还包括功能性小鼠adam6a和adam6b基因(参见美国专利no.10,130,081,以引用的方式并入本文中)。对已经进行同源重组的克隆的选择产生修饰es细胞,用于产生从重排人类vλ1
‑
51/jλ2和小鼠cκ表达抗体的嵌合小鼠。
[0526]
通过taqman
tm
筛选,使用对插入内源κ基因座中的重排人类vλ1
‑
51/jλ2具特异性的引物和探针对阳性es细胞克隆进行确认(参见上述valenzuela等人,以引用的方式并入本文中)。图3中通过带圆圈破折号示出了可用于检测的多种探针的大致位置。下表a中尤其列出了测定法中所用的引物和探针。
[0527]
表a
‑
等位基因修饰测定法上所用的引物和探针
[0528]
[0529][0530]
将载有包括重排人类vλ1
‑
51/jλ2的工程化内源κ轻链基因座的es细胞(参见图3)用表达flp的构建体转染以去除由靶向构建体引入的frt化新霉素盒。任选地,通过繁育成表达flp重组酶的小鼠来去除新霉素盒(例如us 6,774,279,其以全文引用的方式并入)。任选地,新霉素盒被保留在小鼠中。
[0531]
实施例3
‑
表达源自于重排人类vλ1
‑
51/jλ2和小鼠cκ的轻链的小鼠的产生
[0532]
以上实施例2中所述的靶向es细胞用作供体es细胞并通过方法引入8细胞期的小鼠胚胎中(参见例如美国专利no.7,294,754和poueymirou等人(2007)f0 generation mice that are essentially fully derived from the donor gene
‑
targeted es cells allowing immediate phenotypic analyses,nature biotech.25(1):91
‑
99,各以引用的方式并入本文中)。通过使用等位基因修饰测定法(上述valenzuela等人,以引用的方式并入本文中)进行基因分型来鉴定载有包括连接至小鼠cκ的重排人类vλ1
‑
51/jλ2的工程化内源κ轻链基因座的所述测定法检
测重排人类vλ1
‑
51/jλ2的存在。载有在一个基因座等位基因处包括连接至小鼠cκ的重排人类vλ1
‑
51/jλ2的工程化内源κ轻链基因座的小鼠可载有在第二基因座等位基因处包含连接至小鼠小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座,并且含有包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座。
[0533]
小鼠被繁育成对于包括重排人类vλ1
‑
51/jλ2和小鼠cκ的工程化内源κ轻链基因座是纯合的,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的。在一种方法中,这可以通过以下来实现:首先将包含在一个轻链基因座等位基因处包括连接至小鼠cκ的重排人类vλ1
‑
51/jλ2的工程化内源κ轻链基因座、在第二轻链基因座等位基因处包含连接至小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座和包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因(在两个重链等位基因处)的工程化免疫球蛋白重链基因座的小鼠繁育成包含内源重链和轻链κ基因座敲除(ko)并且任选地还包含内源λ基因座ko的小鼠,如下文进一步所述。可替代地,可将小鼠繁育成包含内源轻链κ基因座ko、任选地内源λ基因座ko,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的小鼠。将所得到的小鼠繁育成在免疫球蛋白基因座,例如修饰和/或内源免疫球蛋白基因座,例如人源化λ轻链和重链基因座处是纯合的。
[0534]
对幼仔进行基因分型并选择对于包括重排人类vλ1
‑
51/jλ2和小鼠cκ的工程化内源κ轻链基因座是杂合或纯合的幼仔,并且评估和表征人类λ轻链可变结构域从重排人类vλ1
‑
51/jλ2或其体细胞超突变变体的表达。使用facs分析来检测包含本文所述的工程化轻链的小鼠中人类vλ1
‑
51/jλ2的b细胞表面表达。
[0535]
实施例4
‑
包括重排人类vλ1
‑
51/jλ2和小鼠cλ1的靶向载体的产生
[0536]
为了用小鼠cλ1替代靶向载体a中的小鼠cκ,进行额外的克隆步骤(参见图1b)。在体外用两种cas9:grna复合物切割靶向载体a:第一次切割在内含子κ增强子(e
i
)上游,且第二次切割在cκ下游。接着进行吉普森组装(gibson assembly)以用4.9kb合成片段替代缺失区域,所述片段含有45bp与cas9切割的靶向载体的3'末端的重叠、e
i
、小鼠cλ1、loxp
‑
ub
‑
hyg
‑
loxp盒和80bp与cas9切割的靶向载体的5'末端的重叠。合成片段是从pct/us2018/063841的图1a的质粒pe的限制片段获得的,所述文献以引用的方式并入本文中。图1b中示出了所得靶向载体,靶向载体b。
[0537]
除了小鼠cκ用小鼠cλ1替代并且loxp
‑
ub
‑
hyg
‑
loxp盒插入cκ多聚a信号下游之外,靶向载体b与靶向载体a相同。
[0538]
实施例5
‑
包括重排人类vλ1
‑
51/jλ2和小鼠cλ1的es细胞的产生
[0539]
如实施例4中所述的靶向载体b被电穿孔至小鼠es细胞中,在所述小鼠es细胞中内源vκ和jκ基因区段已经被重排人类vκ3
‑
20/jκ1替代,并且其中免疫球蛋白重链基因座包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段。小鼠es细胞的免疫球蛋白重链基因座还包括功能性小鼠adam6a和adam6b基因(参见美国专利no.10,130,081,以引用的方式并入本文中)。对已经进行同源重组的克隆的选择产生修饰es细胞,用于产生
从重排人类vλ1
‑
51/jλ2和小鼠cλ1表达抗体的嵌合小鼠。
[0540]
通过taqman
tm
筛选,使用对插入内源κ基因座中的重排人类vλ1
‑
51/jλ2和小鼠cλ1具特异性的引物和探针对阳性es细胞克隆进行确认(参见上述valenzuela等人,以引用的方式并入本文中)。图4中通过带圆圈破折号示出了用于检测的多种探针的大致位置。上表a中尤其列出了测定法中所用的引物和探针。
[0541]
将载有包括重排人类vλ1
‑
51/jλ2和小鼠cλ1的工程化内源κ轻链基因座的es细胞(参见图4)用表达flp和/或cre的构建体转染以去除frt化新霉素盒或flox化潮霉素盒。任选地,可以通过繁育成表达flp重组酶的小鼠来去除新霉素盒(例如us 6,774,279,以全文引用的方式并入),和/或可以通过繁育成表达cre重组酶的小鼠来去除潮霉素盒。任选地,新霉素盒和/或潮霉素盒被保留在小鼠中。
[0542]
实施例6
‑
表达源自于重排人类vλ1
‑
51/jλ2和小鼠cλ1的轻链的小鼠的产生
[0543]
以上实施例5中所述的靶向es细胞用作供体es细胞并通过方法引入8细胞期的小鼠胚胎中(参见例如美国专利no.7,294,754和poueymirou等人(2007)f0 generation mice that are essentially fully derived from the donor gene
‑
targeted es cells allowing immediate phenotypic analyses,nature biotech.25(1):91
‑
99,各以引用的方式并入本文中)。通过使用等位基因修饰测定法(参见上述valenzuela等人,以引用的方式并入本文中)进行基因分型来鉴定载有包括重排人类vλ1
‑
51/jλ2和小鼠cλ1的工程化内源κ轻链基因座的所述测定法检测重排人类vλ1
‑
51/jλ2或小鼠cλ1的存在。载有在一个基因座等位基因处包括连接至小鼠cλ1的重排人类vλ1
‑
51/jλ2的工程化内源κ轻链基因座的小鼠可载有在第二基因座等位基因处包含连接至小鼠小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座,并且含有包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座。
[0544]
小鼠被繁育成对于包括重排人类vλ1
‑
51/jλ2和小鼠cλ1的修饰κ轻链基因座是纯合的,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的。在一种方法中,这可以通过以下来实现:首先将包含在一个轻链基因座等位基因处包括连接至小鼠cλ1的重排人类vλ1
‑
51/jλ2的工程化内源κ轻链基因座、在第二轻链基因座等位基因处包含连接至小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座和包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因(在两个重链等位基因处)的工程化免疫球蛋白重链基因座的小鼠繁育成包含内源重链和轻链κ基因座敲除(ko)并且任选地还包含内源λ基因座ko的小鼠,如下文进一步所述。可替代地,可将小鼠繁育成包含内源轻链κ基因座ko、任选地内源λ基因座ko,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的小鼠。将所得到的小鼠繁育成在免疫球蛋白基因座,例如修饰和/或内源免疫球蛋白基因座,例如人源化λ轻链和重链基因座处是纯合的。
[0545]
对幼仔进行基因分型并选择对于包括重排人类vλ1
‑
51/jλ2和小鼠cλ1的工程化内源κ轻链基因座是杂合或纯合的幼仔,并且评估和表征人类λ轻链可变结构域从重排人类vλ
1
‑
51/jλ2或其体细胞超突变变体的表达。使用facs分析来检测包含本文所述的工程化轻链的小鼠中人类vλ1
‑
51/jλ2的b细胞表面表达。
[0546]
实施例7
‑
包括重排人类vλ2
‑
14/jλ2和小鼠cκ的靶向载体的产生
[0547]
使用所属领域中公认的标准分子生物学技术制造重排人类vλ2
‑
14/jλ2。重排人类vλ2
‑
14/jλ2中所包括的人类vλ2
‑
14基因区段和人类jλ2基因区段分别具有人类vλ2
‑
14种系序列和人类jλ2种系序列。
[0548]
通过从头dna合成(blue heron biotech)制造dna片段。所述片段含有asci位点、2kb人类vλ1
‑
51启动子区域、人类vλ1
‑
51 5'非翻译区(utr)、人类vλ2
‑
14基因区段的外显子1的编码序列(其包括编码前导肽的序列)、人类vλ2
‑
14基因区段的内含子1、包括人类vλ2
‑
14基因区段的第二外显子和jλ2基因区段的重排人类vλ2
‑
14/jλ2可变序列、人类jκ5
‑
cκ内含子的开始412bp(包括jκ5剪接供体位点(sd))和pi
‑
scei位点(seq id no:36;又参见图2a和图13a
‑
c)。
[0549]
将上一段中描述的片段连结至先前描述的靶向载体中以产生图2a中的靶向载体c。先前描述的靶向载体含有:源自于bac ct7
‑
302g12的约23kb 5'小鼠同源臂、frt
‑
ub
‑
neo
‑
frt盒、asci位点、重排人类vκ3
‑
20/jκ1可变区、pi
‑
scei位点和源自于bac ct7
‑
254m04的约75kb 3'小鼠同源臂(参见例如美国专利no.9,334,334的图14b,和美国专利no.10,143,186的图2,各以引用的方式并入本文中)。所述小鼠同源臂与先前macdonald等人(2014)proc.natl.acad.sci.usa,111:5147
‑
52描述的同源臂相同,所述文献以引用的方式并入本文中。5'臂在内源κ基因座上游含有约23kb基因组序列,且3'臂在小鼠κ基因座下游含有约2.4kb小鼠jκ
‑
cκ内含子、小鼠cκ外显子和约72kb基因组序列。参见图2a。
[0550]
靶向载体c从5'至3'含有源自于bac ct7
‑
302g12的约23kb 5'小鼠同源臂、frt
‑
ub
‑
neo
‑
frt盒、asci位点、2kb人类vλ1
‑
51启动子区域、人类vλ1
‑
51 5'utr、vλ2
‑
14基因区段的外显子1的编码序列(其包括编码前导肽的序列)、人类vλ2
‑
14基因区段的内含子1、包括人类vλ2
‑
14基因区段的第二外显子和jλ2基因区段的重排人类vλ2
‑
14/jλ2可变序列、人类jκ5
‑
cκ内含子的开始412bp(包括jκ5剪接位点)、pi
‑
scei位点和源自于bac ct7
‑
254m04的约75kb 3'小鼠同源臂,所述3'小鼠同源臂在小鼠κ基因座下游包括约2.4kb小鼠jκ
‑
cκ内含子、小鼠cκ外显子和约72kb基因组序列。
[0551]
实施例8
‑
包括重排人类vλ2
‑
14/jλ2和小鼠cκ的es细胞的产生
[0552]
靶向载体c被电穿孔至小鼠es细胞中,在所述小鼠es细胞中内源vκ和jκ基因区段已经被重排人类vκ3
‑
20/jκ1替代,并且其中免疫球蛋白重链基因座包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段。小鼠es细胞的免疫球蛋白重链基因座还包括功能性小鼠adam6a和adam6b基因(参见美国专利no.10,130,081,以引用的方式并入本文中)。对已经进行同源重组的克隆的选择产生修饰es细胞,用于产生从重排人类vλ2
‑
14/jλ2和小鼠cκ表达抗体的嵌合小鼠。
[0553]
通过taqman
tm
筛选,使用对插入内源κ基因座中的重排人类vλ2
‑
14/jλ2具特异性的引物和探针对阳性es细胞克隆进行确认(参见上述valenzuela等人,以引用的方式并入本文中)。图5中通过带圆圈破折号示出了用于检测的多个探针的大致位置。上表a中尤其列出了测定法中所用的引物和探针。
[0554]
将载有包括重排人类vλ2
‑
14/jλ2的工程化内源κ轻链基因座的es细胞(参见图5)
用表达flp的构建体转染以去除由靶向构建体引入的frt化新霉素盒。任选地,通过繁育成表达flp重组酶的小鼠来去除新霉素盒(例如us 6,774,279,以全文引用的方式并入)。任选地,新霉素盒被保留在小鼠中。
[0555]
实施例9
‑
表达源自于重排人类vλ2
‑
14/jλ2和小鼠cκ的轻链的小鼠的产生
[0556]
以上实施例8中所述的靶向es细胞用作供体es细胞并通过方法引入8细胞期的小鼠胚胎中(参见例如美国专利no.7,294,754和poueymirou等人(2007)f0 generation mice that are essentially fully derived from the donor gene
‑
targeted es cells allowing immediate phenotypic analyses,nature biotech.25(1):91
‑
99,各以引用的方式并入本文中)。通过使用等位基因修饰测定法(上述valenzuela等人,以引用的方式并入本文中)进行基因分型来鉴定载有包括重排人类vλ2
‑
14/jλ2和小鼠cκ的工程化内源κ轻链基因座的所述测定法检测重排人类vλ2
‑
14/jλ2的存在。载有在一个基因座等位基因处包括连接至小鼠cκ的重排人类vλ2
‑
14/jλ2的工程化内源κ轻链基因座的小鼠可载有在第二基因座等位基因处包含连接至小鼠小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座,并且含有包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座。
[0557]
小鼠被繁育成对于包括重排人类vλ2
‑
14/jλ2和小鼠cκ的工程化内源κ轻链基因座是纯合的,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的。在一种方法中,这可以通过以下来实现:首先将包含在一个轻链基因座等位基因处包括重排人类vλ2
‑
14/jλ2和小鼠cκ的工程化内源κ轻链基因座、在第二轻链基因座等位基因处包含连接至小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座和包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因(在两个重链等位基因处)的工程化免疫球蛋白重链基因座的小鼠繁育成包含内源重链和轻链κ基因座敲除(ko)并且任选地还包含内源λ基因座ko的小鼠,如下文进一步所述。可替代地,可将小鼠繁育成包含内源轻链κ基因座ko、任选地内源λ基因座ko,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的小鼠。将所得到的小鼠繁育成在免疫球蛋白基因座,例如修饰和/或内源免疫球蛋白基因座,例如人源化λ轻链和重链基因座处是纯合的。
[0558]
对幼仔进行基因分型并选择对于包括重排人类vλ2
‑
14/jλ2和小鼠cκ的工程化内源κ轻链基因座是杂合或纯合的幼仔,并且评估和表征人类λ轻链可变结构域从重排人类vλ2
‑
14/jλ2或其体细胞超突变变体的表达。使用facs分析来检测包含本文所述的工程化轻链的小鼠中人类vλ2
‑
14/jλ2的b细胞表面表达。
[0559]
实施例10
‑
包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的靶向载体的产生
[0560]
为了用小鼠cλ1替代靶向载体c中的小鼠cκ,进行额外的克隆步骤(参见图2b)。在体外用两种cas9:grna复合物切割靶向载体c:第一次切割在内含子κ增强子(e
i
)上游,且第二次切割在cκ下游。接着进行吉普森组装以用4.9kb合成片段替代缺失区域,所述片段含有45bp与cas9切割的靶向载体的3'末端的重叠、e
i
、小鼠cλ1、loxp
‑
ub
‑
hyg
‑
loxp盒和80bp与cas9切割的靶向载体的5'末端的重叠。合成片段是从pct/us2018/063841的图1a的质粒pe
的限制片段获得的,所述文献以引用的方式并入本文中。图2b中示出所得靶向载体,靶向载体d。
[0561]
除了小鼠cκ用小鼠cλ1替代并且loxp
‑
ub
‑
hyg
‑
loxp盒插入cκ多聚a信号下游之外,靶向载体d与靶向载体c相同。
[0562]
实施例11
‑
包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的es细胞的产生
[0563]
靶向载体d被电穿孔至小鼠es细胞中,在所述小鼠es细胞中内源vκ和jκ基因区段已经被重排人类vκ3
‑
20/jκ1替代,并且其中免疫球蛋白重链基因座包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段。小鼠es细胞的免疫球蛋白重链基因座还包括功能性小鼠adam6a和adam6b基因(参见美国专利no.10,130,081,以引用的方式并入本文中)。对已经进行同源重组的克隆的选择产生修饰es细胞,用于产生从重排人类vλ2
‑
14/jλ2和小鼠cλ1表达抗体的嵌合小鼠。
[0564]
通过taqman
tm
筛选,使用对插入内源κ基因座中的重排人类vλ2
‑
14/jλ2和小鼠cλ1具有特异性的引物和探针对阳性es细胞克隆进行确认(参见上述valenzuela等人,以引用的方式并入本文中)。图6中通过带圆圈破折号示出了用于检测的多种探针的大致位置。上表a中尤其列出了测定法中所用的引物和探针。
[0565]
将载有包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的工程化内源κ轻链基因座的es细胞(参见图6)用表达flp和/或cre的构建体转染以去除frt化新霉素盒或flox化潮霉素盒。任选地,通过繁育成表达flp重组酶的小鼠来去除新霉素盒(例如us 6,774,279,以全文引用的方式并入),和/或通过繁育成表达cre重组酶的小鼠来去除潮霉素盒。任选地,新霉素盒和/或潮霉素盒被保留在小鼠中。
[0566]
实施例12
‑
表达源自于重排人类vλ2
‑
14/jλ2和小鼠cλ1的轻链的小鼠的产生
[0567]
以上实施例11中所述的靶向es细胞用作供体es细胞并通过方法引入8细胞期的小鼠胚胎中(参见例如美国专利no.7,294,754和poueymirou等人(2007)f0 generation mice that are essentially fully derived from the donor gene
‑
targeted es cells allowing immediate phenotypic analyses,nature biotech.25(1):91
‑
99,各以引用的方式并入本文中)。通过使用等位基因修饰测定法(参见上述valenzuela等人,其以引用的方式并入本文中)进行基因分型来鉴定载有包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的工程化内源κ轻链基因座的所述测定法检测重排人类vλ2
‑
14/jλ2或小鼠cλ1的存在。载有在一个基因座等位基因处包括连接至小鼠cλ1的重排人类vλ2
‑
14/jλ2的工程化内源κ轻链基因座的小鼠可载有在第二基因座等位基因处包含连接至小鼠小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座,并且含有包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座。
[0568]
小鼠被繁育成对于包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的修饰κ轻链基因座是纯合的,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的。在一种方法中,这可以通过以下来实现:首先将包含在一个轻链基因座等位基因处包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的工程化内源κ轻链基因座、在第二轻链基因座等位基因处包含连接至小鼠cκ的重排人类vκ3
‑
20/jκ1的工程化内源κ轻链基因座和包含可操作地连接至小鼠重链恒
定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因(在两个重链等位基因处)的工程化免疫球蛋白重链基因座的小鼠繁育成包含内源重链和轻链κ基因座敲除(ko)并且任选地还包含内源λ基因座ko的小鼠,如下文进一步所述。可替代地,可将小鼠繁育成包含内源轻链κ基因座ko、任选地内源λ基因座ko,并且对于包含可操作地连接至小鼠重链恒定区基因的未重排人类v
h
、d和j
h
基因区段和功能性小鼠adam6a和adam6b基因的工程化免疫球蛋白重链基因座是纯合的小鼠。将所得到的小鼠繁育成在免疫球蛋白基因座,例如修饰和/或内源免疫球蛋白基因座,例如人源化λ轻链和重链基因座处是纯合的。
[0569]
对幼仔进行基因分型并选择对于包括重排人类vλ2
‑
14/jλ2和小鼠cλ1的工程化内源κ轻链基因座是杂合或纯合的幼仔,并且评估和表征人类λ轻链可变结构域从重排人类vλ2
‑
14/jλ2或其体细胞超突变变体的表达。使用facs分析来检测包含本文所述的工程化轻链的小鼠中人类vλ12
‑
14/jλ2的b细胞表面表达。
[0570]
实施例13
‑
小鼠的繁育
[0571]
本实施例描述了几种基因修饰小鼠品系,它们可被繁育成本文所述的任一种基因修饰小鼠,以建立具有多个基因修饰免疫球蛋白基因座的多个基因修饰小鼠品系。
[0572]
设想了许多不同的繁育方案,这些方案产生在本文所述的工程化轻链和重链基因座处是纯合的小鼠,以便小鼠表达具有重排人类igλ可变结构域的轻链和具有重排人类重链可变结构域的重链。在一个这样的方案中,以上实施例3、6、9和12中所述的基因修饰小鼠对于一个轻链基因座等位基因上(在内源κ基因座处)的重排人类vλ1
‑
51/jλ2或重排人类vλ2
‑
14/jλ2轻链和第二轻链基因座等位基因处(在内源κ基因座处)的重排人类vκ3
‑
20/jκ1是杂合的,并且对于上述包含未重排人类v
h
、d和j
h
基因区段的工程化免疫球蛋白重链基因座是纯合的,所述小鼠被繁育成对于内源小鼠igκ、igλ和ig重链基因座的缺失是纯合的小鼠。所得到的表达重排λ轻链可变结构域的后代彼此繁育,获得对于重排人类vλ1
‑
51/jλ2或重排人类vλ2
‑
14/jλ2轻链是纯合并且对于包含未重排人类v
h
、d和j
h
基因区段(并且如上所述还包含功能性小鼠adam6a和adam6b基因)的工程化免疫球蛋白重链基因座是纯合的小鼠。通过所属领域中公认的标准技术且可替代地通过商业育种公司进行繁育。
[0573]
内源igκ敲除(igκko).举例来说,可将上述包含重排人类λ轻链可变区和人类重链可变区的小鼠繁育成所有或一部分内源igκ基因座缺失,使得内源igκ轻链基因座不能表达内源igκ轻链的小鼠。这种示例性小鼠描述于macdonald等人(2014)proc.natl.acad.sci.usa,111:5147
‑
52中,所述文献以引用的方式并入本文中。
[0574]
内源igλ敲除(igλko).为了优化有限人类λ轻链可变区谱系的使用,将上述包含重排人类λ轻链可变区和人类重链可变区的小鼠繁育成所有或一部分内源igλ轻链基因座缺失,使得内源igλ轻链基因座不能表达内源λ轻链的小鼠(参见例如美国专利no.9,066,502的实施例1,以引用的方式并入本文中)。通过这种方式获得的后代将表达具有从有限人类λ轻链可变区谱系表达的可变结构域的轻链,作为其唯一的λ轻链(例如,如实施例3、6、9和12中所述的重排人类vλ/jλ)。针对有限人类λ轻链可变区谱系的存在和内源小鼠λ轻链的缺乏,筛选载有有限人类λ轻链可变区谱系和所有或一部分内源λ轻链基因座缺失的小鼠品系。
[0575]
内源ig重链敲除(igh ko).举例来说,可将上述包含重排人类λ轻链可变区和未重排人类重链可变区的小鼠繁育成所有或一部分内源ig重链基因座缺乏,使得内源ig重链基
因座不能表达内源ig重链的小鼠。这种示例性小鼠描述于美国专利no.10,457,960的实施例7中,其以引用的方式并入本文中。
[0576]
因此,如上所述,可将如上述包含重排人类λ轻链可变区和人类重链可变区的小鼠繁育成所有或一部分内源igλ、igκ和/或ig重链基因座缺失的小鼠。后来的后代被繁育成在免疫球蛋白基因座,例如修饰和/或内源免疫球蛋白基因座,例如人源化λ轻链和重链基因座处是纯合的。
[0577]
实施例14
–
重链基因使用和λ轻链的体细胞超突变频率
[0578]
可经由所属领域已知的数种方法确定本文所述的包含重排人类vλ1
‑
51/jλ2或重排人类vλ2
‑
14/jλ2轻链可变区并且用人类重链可变区替代内源重链可变区的小鼠的重链基因使用情况。在一种这样的方法中,使用下一代测序技术。
[0579]
针对抗体的重链可变区区段使用情况,分析从上文所述的包含通用重排人类vλ1
‑
51/jλ2或重排人类vλ2
‑
14/jλ2轻链小鼠并且用人类重链可变区替代内源重链可变区的小鼠收获的脾细胞或骨髓。通过用含有2.5%胎牛血清(fbs)的1x磷酸盐缓冲盐水(pbs,gibco)冲洗股骨,从股骨收集骨髓,并且在必要时,针对不同的b细胞亚型,对骨髓细胞进行facs分选。从小鼠的脾制备单细胞悬浮液。将来自脾的红细胞和骨髓制剂用ack裂解缓冲液(gibco)溶解。通过使用抗cd19(小鼠,b细胞标志物)磁珠和柱(miltenyi biotech)进行磁性细胞分选,从总脾细胞进行脾b细胞的阳性富集。使用rneasy plus rna分离试剂盒(qiagen),根据制造商的说明书,从骨髓和纯化的脾b细胞分离总rna。
[0580]
使用smarter
tm race cdna扩增试剂盒(clontech)和igm或igg特异性引物进行逆转录以产生含有igm或igg恒定区序列的人类重链cdna。在逆转录期间,作为模板转换(ts)引物的反向互补序列的dna序列连接于新合成的cdna的3
′
末端。纯化的cdna通过两轮半巢式pcr扩增,以产生编码由获得mrna的细胞表达的总igm或igg可变结构域互补物的多个cdna,接着进行第三轮pcr,以连接测序引物和标志。使用pippin prep(sage science)对人类可变结构域cdna进行尺寸选择,并且通过qpcr使用kapa文库定量试剂盒(kapa biosystems)进行定量,然后将样品装载至miseq测序仪(illumina)上进行测序。
[0581]
对于生物信息学分析,将原始illumina序列多路分离并基于品质、长度和与对应igm或igg恒定区基因引物的匹配进行过滤。将重叠双端读段合并使用定制的内部管线分析。管线使用igblast(ncbi,v2.2.25+)的本地安装,以将重排重链序列与人类种系重链v、d和j基因区段数据库进行比对,并表示产生性接合和非产生性接合以及中止密码子的存在。使用如国际免疫遗传信息系统(international immunogenetics information system,imgt)中定义的边界提取cdr3序列和预期非模板核苷酸。
[0582]
以上方法允许确定多种人类v、d和j区段的使用情况,以及本文所述的通用重排人类轻链小鼠中特定人类vdj重排的频率。
[0583]
可以使用类似的下一代测序技术确定包含通用重排人类vλ1
‑
51/jλ2或vλ2
‑
14/jλ2轻链可变区的小鼠的λ轻链中体细胞超突变的存在和频率。举例来说,对于包含连接至小鼠cλ(例如cλ1)恒定区的通用重排人类vλ1
‑
51/jλ2或vλ2
‑
14/jλ2轻链可变区的小鼠,使用免疫球蛋白cλ(例如cλ1)特定引物进行上述逆转录pcr以产生含有免疫球蛋白λ恒定区(例如cλ1)基因序列的cdna,并进行生物信息学分析以将重排轻链序列与人类种系vλ和jλ基因区段数据库进行比对。对于连接至小鼠cκ恒定区的通用重排人类vλ1
‑
51/jλ2或vλ2
‑
14/jλ2
轻链可变区,改为使用小鼠cκ特定引物。对λ轻链中体细胞超突变的存在和频率进行定量。
[0584]
实施例15
‑
从小鼠产生人类抗体
[0585]
在将包含有限人类λ轻链可变区谱系(例如,如实施例3、6、9和12中所述的重排人类vλ/jλ)的小鼠繁育成含有其它内源ig基因座的修饰和缺失(如例如实施例13中所述)的各种期望品系后,可用所关注抗原对所选小鼠进行免疫接种。
[0586]
一般来说,用所关注抗原对包含有限人类λ轻链可变区谱系的小鼠(在其种系基因组中包含人类重链可变区基因区段并且用工程化vλ1
‑
51/jλ2人类轻链区域或工程化vλ2
‑
14/jλ2人类轻链区域替代内源小鼠κ轻链基因座(上述),去除或不去除内源λ基因座的小鼠)进行免疫接种。收集血液并通过标准的抗原特异性elisa免疫测定法来监测抗体免疫反应。
[0587]
当实现期望免疫反应时,在一种方法中,收获脾细胞(和/或其它淋巴组织)并与小鼠骨髓瘤细胞融合以保存其活力且形成永生杂交瘤细胞系。对产生的杂交瘤细胞系进行筛选(例如通过elisa测定法)并进行选择以鉴定产生抗原特异性抗体的杂交瘤细胞系。可根据需要,针对相对结合亲和力和同种型进一步表征杂交瘤。使用这项技术和上述免疫原,获得数种抗原特异性嵌合抗体(即,具有人类可变结构域和小鼠恒定结构域的抗体)。
[0588]
可替代地,在另一方法中,可直接从抗原特异性b细胞分离编码由本文中描述和/或例示的工程化小鼠的b细胞产生的抗原特异性嵌合抗体和/或其λ轻链和/或重链可变结构域的dna。举例来说,可分离和表征具有人类可变区和小鼠恒定区的高亲和力嵌合抗体,并界定所关注特定抗体(和/或产生其的b细胞)。仅举几例,这类抗体和/或其可变和/或恒定区的评定特征可以是或包括亲和力、选择性、表位的同一性等的一种或多种。在不与骨髓瘤细胞融合下,直接从抗原阳性b细胞(来自免疫接种小鼠)分离抗原特异性抗体,如例如美国专利no.7,582,298中所述,所述专利特定地以引用的方式整体并入本文中。
[0589]
可分离或以其它方式制备编码重链和λ轻链可变区的dna,并且可分别连接于人类重链(例如期望同种型)和人类λ轻链恒定区以制备完全人类抗体。分离的重链和λ轻链可变区可进行体细胞突变。这为抗原特异性谱系增添了额外的多样性。可在细胞、通常例如cho细胞的哺乳动物细胞中产生完全人类抗体(和/或其重链或λ轻链)。接着可针对相对结合亲和力和/或所关注抗原的中和活性,表征完全人类抗体。
[0590]
实施例16
‑
双特异性抗体的结合亲和力
[0591]
使用所属领域已知的标准重组dna技术从上述对所关注抗原具特异性的所选单特异性抗体的克隆人类重链可变区构建完全人类双特异性抗体。简单地说,在合适的细胞系中,将如下两条人类重链与如下人类轻链一起表达,所述两条人类重链包含来自从包含工程化人类vλ1
‑
51/jλ2轻链或工程化人类vλ2
‑
14/jλ2轻链区域的小鼠获得的两种所选单特异性抗体的人类重链可变区,所述人类轻链包含与小鼠中所存在相同的λ可变区或其体细胞超突变型式;并且获得包含期望特征(例如对两种抗原期望的亲和力)的双特异性抗体。
[0592]
在biacore
tm 2000仪器(ge healthcare)上使用实时表面等离子共振生物传感器测定法确定对所关注抗原具特异性的双特异性或亲本单特异性抗体与抗原整体或一部分的结合。
[0593]
某些实施方案
[0594]
实施方案1.一种基因修饰啮齿动物,所述啮齿动物的种系基因组包含:
[0595]
工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段,
[0596]
其中由所述基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0597]
实施方案2.实施方案1的基因修饰啮齿动物,其中所述基因修饰啮齿动物的种系基因组对于所述工程化内源免疫球蛋白κ轻链基因座是纯合的。
[0598]
实施方案3.实施方案1的基因修饰啮齿动物,其中所述基因修饰啮齿动物的种系基因组对于所述工程化内源免疫球蛋白κ轻链基因座是杂合的。
[0599]
实施方案4.实施方案1
‑
3中任一项的基因修饰啮齿动物,所述啮齿动物的种系基因组还包含:
[0600]
工程化内源免疫球蛋白重链基因座,所述工程化内源免疫球蛋白重链基因座包含可操作地连接至一个或多个啮齿动物免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段,
[0601]
其中由所述基因修饰啮齿动物的b细胞表达的所有重链均包括人类免疫球蛋白重链可变结构域和啮齿动物免疫球蛋白重链恒定结构域。
[0602]
实施方案5.实施方案4的基因修饰啮齿动物,其中所述基因修饰啮齿动物的种系基因组对于所述工程化内源免疫球蛋白重链基因座是纯合的。
[0603]
实施方案6.实施方案1
‑
5中任一项的基因修饰啮齿动物,其中所述基因修饰啮齿动物在所述工程化内源免疫球蛋白κ轻链基因座处缺乏啮齿动物cκ基因。
[0604]
实施方案7.实施方案1
‑
6中任一项的基因修饰啮齿动物,其中所述人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。
[0605]
实施方案8.实施方案1
‑
7中任一项的基因修饰啮齿动物,其中所述人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。
[0606]
实施方案9.实施方案4
‑
8中任一项的基因修饰啮齿动物,其中所述一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段代替一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段、一个或多个内源j
h
基因区段或它们的组合。
[0607]
实施方案10.实施方案4
‑
9中任一项的基因修饰啮齿动物,其中所述一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段分别替代一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段和一个或多个内源j
h
基因区。
[0608]
实施方案11.实施方案9或10的基因修饰啮齿动物,其中所述一个或多个啮齿动物免疫球蛋白重链恒定区基因是一个或多个内源啮齿动物免疫球蛋白重链恒定区基因。
[0609]
实施方案12.实施方案4
‑
11中任一项的基因修饰啮齿动物,其中:
[0610]
(i)所述一个或多个未重排人类v
h
基因区段包含v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、
v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合,
[0611]
(ii)所述一个或多个未重排人类d
h
基因区段包含d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合,以及
[0612]
(iii)所述一个或多个未重排人类j
h
基因区段包含j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合。
[0613]
实施方案13.实施方案4
‑
12中任一项的基因修饰啮齿动物,其中所述工程化内源免疫球蛋白重链基因座缺乏功能性内源啮齿动物adam6基因。
[0614]
实施方案14.实施方案4
‑
13中任一项的基因修饰啮齿动物,其中所述基因修饰啮齿动物的种系基因组包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
[0615]
实施方案15.实施方案14的基因修饰啮齿动物,其中所述一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段由所述基因修饰啮齿动物表达。
[0616]
实施方案16.实施方案14或15的基因修饰啮齿动物,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列包括在与所述工程化内源免疫球蛋白重链基因座相同的染色体上。
[0617]
实施方案17.实施方案14
‑
16中任一项的基因修饰啮齿动物,其中所述工程化内源免疫球蛋白重链基因座包含所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
[0618]
实施方案18.实施方案14
‑
17中任一项的基因修饰啮齿动物,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列代替人类adam6伪基因。
[0619]
实施方案19.实施方案14
‑
18中任一项的基因修饰啮齿动物,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列替代人类adam6伪基因。
[0620]
实施方案20.实施方案14
‑
19中任一项的基因修饰啮齿动物,其中所述一个或多个人类v
h
基因区段包含第一人类v
h
基因区段和第二人类v
h
基因区段,且所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于所述第一人类v
h
基因区段与所述第二人类v
h
基因区段之间。
[0621]
实施方案21.实施方案20的基因修饰啮齿动物,其中所述第一人类v
h
基因区段是v
h1‑
2且所述第二人类v
h
基因区段是v
h6‑
1。
[0622]
实施方案22.实施方案14
‑
17中任一项的基因修饰啮齿动物,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于人类v
h
基因区段与人类d
h
基因区段之间。
[0623]
实施方案23.实施方案1
‑
22中任一项的基因修饰啮齿动物,其中所述啮齿动物cλ基因具有与以下至少80%同一的序列:(i)小鼠cλ1,(ii)小鼠cλ2,或(iii)小鼠cλ3基因。
[0624]
实施方案24.实施方案1
‑
23中任一项的基因修饰啮齿动物,其中所述啮齿动物cλ基因包含小鼠cλ基因。
[0625]
实施方案25.实施方案1
‑
23中任一项的基因修饰啮齿动物,其中所述啮齿动物cλ基因包含小鼠cλ1基因。
[0626]
实施方案26.实施方案1
‑
22的基因修饰啮齿动物,其中所述啮齿动物cλ基因具有与以下至少80%同一的序列:(i)大鼠cλ1,(ii)大鼠cλ2,(iii)大鼠cλ3,或(iv)大鼠cλ4基因。
[0627]
实施方案27.实施方案1
‑
22中任一项的基因修饰啮齿动物,其中所述啮齿动物cλ基因包含大鼠cλ基因。
[0628]
实施方案28.实施方案1
‑
27中任一项的基因修饰啮齿动物,其中所述单一重排人类免疫球蛋白λ轻链可变区代替一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。
[0629]
实施方案29.实施方案1
‑
28中任一项的基因修饰啮齿动物,其中所述单一重排人类免疫球蛋白λ轻链可变区替代一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。
[0630]
实施方案30.实施方案1
‑
29中任一项的基因修饰啮齿动物,所述基因修饰啮齿动物还包含失活内源免疫球蛋白λ轻链基因座。
[0631]
实施方案31.实施方案1
‑
30中任一项的基因修饰啮齿动物,其中所述内源vλ基因区段、所述内源jλ基因区段和所述内源cλ基因整体或部分缺失。
[0632]
实施方案32.实施方案1
‑
31中任一项的基因修饰啮齿动物,其中所述啮齿动物不可检测地表达内源免疫球蛋白κ轻链可变结构域。
[0633]
实施方案33.实施方案1
‑
32中任一项的基因修饰啮齿动物,其中由所述基因修饰啮齿动物的b细胞表达的所有免疫球蛋白轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0634]
实施方案34.实施方案1
‑
33中任一项的基因修饰啮齿动物,其中所述啮齿动物是大鼠或小鼠。
[0635]
实施方案35.一种基因修饰啮齿动物的繁殖群,所述繁殖群包含第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物,其中所述第三基因修饰啮齿动物是所述第一基因修饰啮齿动物和所述第二基因修饰啮齿动物的后代,并且其中所述第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物在它们的种系基因组中各自包含:
[0636]
工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段,
[0637]
其中由所述第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括从单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0638]
实施方案36.实施方案35的繁殖群,其中所述第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物的种系基因组对于所述工程化内源免疫球蛋白重链基因座是纯合的。
[0639]
实施方案37.实施方案35或36的繁殖群,其中所述第一基因修饰啮齿动物、第二基
因修饰啮齿动物和第三基因修饰啮齿动物每一者的所述工程化内源免疫球蛋白κ轻链基因座中的所述人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。
[0640]
实施方案38.实施方案35至37中任一项的繁殖群,其中所述第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物每一者的所述工程化内源免疫球蛋白κ轻链基因座中的所述人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。
[0641]
实施方案39.实施方案35至38中任一项的繁殖群,其中所述第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物在它们的种系基因组中各自包含:
[0642]
工程化内源免疫球蛋白重链基因座,所述工程化内源免疫球蛋白重链基因座包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段,
[0643]
其中由所述基因修饰啮齿动物的b细胞表达的所有重链均包括人类免疫球蛋白重链可变结构域和啮齿动物免疫球蛋白重链恒定结构域。
[0644]
实施方案40.实施方案39的繁殖群,其中所述第一基因修饰啮齿动物、第二基因修饰啮齿动物和第三基因修饰啮齿动物的种系基因组对于所述工程化内源免疫球蛋白重链基因座是纯合的。
[0645]
实施方案41.一种啮齿动物胚胎,所述啮齿动物胚胎的基因组包含工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。
[0646]
实施方案42.实施方案41的啮齿动物胚胎,其中所述啮齿动物胚胎的基因组对于工程化内源免疫球蛋白κ轻链基因座是纯合的。
[0647]
实施方案43.实施方案41或42的啮齿动物胚胎,其中所述人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。
[0648]
实施方案44.实施方案41
‑
43中任一项的啮齿动物胚胎,其中所述人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。
[0649]
实施方案45.实施方案41
‑
44中任一项的啮齿动物胚胎,所述啮齿动物胚胎还包含工程化内源免疫球蛋白重链基因座,所述工程化内源免疫球蛋白重链基因座包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。
[0650]
实施方案46.实施方案45的啮齿动物胚胎,其中所述啮齿动物胚胎的基因组对于所述工程化内源免疫球蛋白重链基因座是纯合的。
[0651]
实施方案47.一种实施方案1
‑
34中任一项的基因修饰啮齿动物的b细胞,所述b细胞包含:
[0652]
所述工程化内源κ轻链基因座的所述单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
[0653]
实施方案48.一种实施方案4
‑
34中任一项的基因修饰啮齿动物的b细胞,所述b细胞包含:
[0654]
所述工程化内源κ轻链基因座的所述单一重排人类免疫球蛋白λ轻链可变区或其
体细胞超突变型式;和
[0655]
重排人类免疫球蛋白重链可变区,所述重排人类免疫球蛋白重链可变区源自于所述工程化内源重链基因座中的所述一个或多个未重排人类v
h
基因区段的人类v
h
基因区段、所述一个或多个未重排人类d
h
基因区段的人类d
h
基因区段和所述一个或多个未重排人类j
h
基因区段的人类j
h
基因区段。
[0656]
实施方案49.一种杂交瘤,所述杂交瘤由实施方案47或48的b细胞产生。
[0657]
实施方案50.一种单个基因修饰啮齿动物的b细胞群体,所述基因修饰啮齿动物在其种系基因组中包含:
[0658]
(a)工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中所述单一重排人类免疫球蛋白λ轻链可变区包含可操作地连接至人类jλ基因区段的人类vλ基因区段,和
[0659]
(b)工程化内源免疫球蛋白重链基因座,其包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段,
[0660]
其中所有由所述b细胞群体表达的抗体均包括:
[0661]
(i)从所述单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域,和
[0662]
(ii)从至少两个不同的重排人类免疫球蛋白重链可变区或其体细胞超突变型式表达的多个人类免疫球蛋白重链可变结构域。
[0663]
实施方案51.一种干细胞,所述干细胞包含工程化内源免疫球蛋白κ轻链基因座,所述工程化内源免疫球蛋白κ轻链基因座包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。
[0664]
实施方案52.实施方案51的干细胞,其中所述干细胞的基因组对于所述工程化内源免疫球蛋白κ轻链基因座是纯合的。
[0665]
实施方案53.实施方案51或52的干细胞,其中所述人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。
[0666]
实施方案54.实施方案51
‑
53中任一项的干细胞,其中所述人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。
[0667]
实施方案55.实施方案51
‑
54中任一项的干细胞,所述干细胞还包含工程化内源免疫球蛋白重链基因座,所述工程化内源免疫球蛋白重链基因座包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。
[0668]
实施方案56.一种表达抗体的哺乳动物细胞,其中所述抗体包括包含人类免疫球蛋白重链可变结构域的重链和包含人类免疫球蛋白λ轻链可变结构域的轻链,其中所述人类免疫球蛋白重链可变结构域、所述人类免疫球蛋白λ轻链可变结构域或两者是从实施方案4
‑
34中任一项的基因修饰啮齿动物鉴定的。
[0669]
实施方案57.一种抗体,所述抗体通过包括以下步骤的方法制备:
[0670]
(a)将实施方案1
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;
[0671]
(b)维持所述基因修饰啮齿动物处于足以使所述基因修饰啮齿动物对所述所关注抗原产生免疫反应的条件下;以及
[0672]
(c)从所述基因修饰啮齿动物回收:
[0673]
(i)结合所述所关注抗原的抗体,
[0674]
(ii)编码结合所述所关注抗原的抗体的人类轻链或重链可变结构域、轻链或重链的核苷酸,或
[0675]
(iii)表达结合所述所关注抗原的抗体的细胞,
[0676]
其中(c)的抗体包括人类重链可变结构域和人类λ轻链可变结构域。
[0677]
实施方案58.一种制造抗体的方法,所述方法包括:
[0678]
(a)将实施方案1
‑
34中任一项的基因修饰啮齿动物暴露于抗原;
[0679]
(b)允许所述基因修饰啮齿动物对所述抗原显现免疫反应;以及
[0680]
(c)从所述基因修饰啮齿动物分离对所述抗原具特异性的抗体、表达对所述抗原具特异性的抗体的b细胞或编码对所述抗原具特异性的抗体的一个或多个核苷酸序列。
[0681]
实施方案59.一种制造抗体的方法,所述方法包括以下步骤:
[0682]
(a)在哺乳动物细胞中表达包含两条人类免疫球蛋白λ轻链和两条人类免疫球蛋白重链的所述抗体,其中每条人类免疫球蛋白λ轻链包括人类免疫球蛋白λ轻链可变结构域且每条人类免疫球蛋白重链包括人类免疫球蛋白重链可变结构域,其中在实施方案1
‑
34中任一项的基因修饰啮齿动物中鉴定所述人类免疫球蛋白重链可变结构域中的至少一个、所述λ轻链可变结构域中的至少一个或它们的组合的氨基酸序列;以及
[0683]
(b)获得所述抗体。
[0684]
实施方案60.实施方案58或59的方法,其中所述抗体是双特异性抗体。
[0685]
实施方案61.一种制造双特异性抗体的方法,所述方法包括:
[0686]
(a)使根据实施方案1
‑
35中任一项的第一基因修饰啮齿动物与第一抗原的第一表位接触,
[0687]
(b)使根据实施方案1
‑
35中任一项的第二基因修饰啮齿动物与第二抗原的第二表位接触,
[0688]
(c)从所述第一基因修饰啮齿动物分离表达对所述第一抗原的所述第一表位具特异性的第一抗体的b细胞并确定所述第一抗体的第一人类免疫球蛋白重链可变结构域;
[0689]
(d)从所述第二基因修饰啮齿动物分离表达对所述第二抗原的所述第二表位具特异性的第二抗体的b细胞并确定所述第二抗体的第二人类免疫球蛋白重链可变结构域;
[0690]
(e)将编码所述第一人类免疫球蛋白重链可变结构域的核苷酸序列可操作地连接至编码第一人类免疫球蛋白恒定结构域的核苷酸序列以产生编码第一人类重链的第一核苷酸序列;
[0691]
(f)将编码所述第二人类免疫球蛋白重链可变结构域的核苷酸序列可操作地连接至编码第二人类免疫球蛋白恒定结构域的核苷酸序列以产生编码第二人类重链的第二核苷酸序列;
[0692]
(g)在哺乳动物细胞中表达:
[0693]
(i)所述第一核苷酸序列;
[0694]
(ii)所述第二核苷酸序列;和
[0695]
(iii)第三核苷酸序列,所述第三核苷酸序列包含可操作地连接至人类免疫球蛋白λ轻链恒定区的所述单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
[0696]
实施方案62.一种制造双特异性抗体的方法,所述方法包括:
[0697]
(a)在哺乳动物细胞中表达:
[0698]
(i)第一核苷酸序列,所述第一核苷酸序列包含可操作地连接至第一人类免疫球蛋白恒定区的第一人类免疫球蛋白重链可变区;
[0699]
(ii)第二核苷酸序列,所述第二核苷酸序列包含可操作地连接至第二人类免疫球蛋白恒定区的第二人类免疫球蛋白重链可变区;和
[0700]
(iii)第三核苷酸序列,所述第三核苷酸序列包含可操作地连接至人类免疫球蛋白λ轻链恒定区的人类免疫球蛋白λ轻链可变区;
[0701]
其中所述第一人类免疫球蛋白重链可变区编码从已经用第一抗原的第一表位进行免疫接种的第一基因修饰啮齿动物中的第一抗体鉴定的第一人类重链可变结构域,其中所述第一抗体特异性结合所述第一抗原的所述第一表位;且
[0702]
其中所述第二人类免疫球蛋白重链可变区编码从已经用第二抗原的第二表位进行免疫接种的第二基因修饰啮齿动物中的第二抗体鉴定的第二人类重链可变结构域,其中所述第二抗体特异性结合所述第二抗原的所述第二表位;
[0703]
其中所述第一基因修饰啮齿动物和第二基因修饰啮齿动物各自是根据实施方案4
‑
34中任一项的基因修饰啮齿动物;且
[0704]
其中所述第三核苷酸的所述人类免疫球蛋白λ轻链可变区是所述单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式。
[0705]
实施方案63.实施方案59或62的方法,其中所述第一基因修饰啮齿动物和第二基因修饰啮齿动物是相同的基因修饰啮齿动物。
[0706]
实施方案64.实施方案59或62的方法,其中所述第一基因修饰啮齿动物和第二基因修饰啮齿动物是不同的基因修饰啮齿动物。
[0707]
实施方案65.实施方案59
‑
64中任一项的方法,其中所述第一抗原和第二抗原是相同抗原,且所述第一表位和第二表位是不同表位。
[0708]
实施方案66.实施方案59
‑
64中任一项的方法,其中所述第一抗原和第二抗原是不同抗原。
[0709]
实施方案67.一种制造人类免疫球蛋白重链的方法,所述方法包括以下步骤:
[0710]
(a)将实施方案4
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;
[0711]
(b)获得特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列;以及
[0712]
(c)将所述人类免疫球蛋白重链可变结构域序列可操作地连接至人类免疫球蛋白重链恒定结构域序列以形成人类免疫球蛋白重链。
[0713]
实施方案68.一种人类免疫球蛋白重链,所述人类免疫球蛋白重链由实施方案67的方法产生。
[0714]
实施方案69.一种制造人类免疫球蛋白重链可变结构域的方法,所述方法包括以下步骤:
[0715]
(a)将实施方案4
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;和
[0716]
(b)获得特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列。
[0717]
实施方案70.一种人类免疫球蛋白重链可变结构域,所述人类免疫球蛋白重链可变结构域由实施方案69的方法产生。
[0718]
实施方案71.一种制造一系列人类免疫球蛋白重链可变结构域的方法,所述方法包括
[0719]
(a)将实施方案4
‑
35中任一项的基因修饰啮齿动物暴露于所关注抗原,以及
[0720]
(b)从所述基因修饰啮齿动物分离所述系列人类免疫球蛋白重链可变结构域,
[0721]
其中所述系列人类免疫球蛋白重链可变结构域各自结合于从所述单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域,且
[0722]
其中与所述系列中的所述人类免疫球蛋白重链可变结构域中的任一者配对的所述人类λ轻链可变结构域结合所述抗原。
[0723]
实施方案72.一种制造人类免疫球蛋白λ轻链的方法,所述方法包括以下步骤:
[0724]
(a)将实施方案1
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;
[0725]
(b)获得特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列;以及
[0726]
(c)将所述人类免疫球蛋白λ轻链可变结构域序列可操作地连接至人类免疫球蛋白λ轻链恒定结构域序列以形成人类免疫球蛋白λ轻链。
[0727]
实施方案73.一种人类免疫球蛋白λ轻链,所述人类免疫球蛋白λ轻链由实施方案72的方法产生。
[0728]
实施方案74.一种用于产生人类免疫球蛋白λ轻链可变结构域的方法,所述方法包括以下步骤:
[0729]
(a)将实施方案1
‑
35中任一项的基因修饰啮齿动物暴露于所关注抗原;以及
[0730]
(b)获得特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列。
[0731]
实施方案75.一种人类免疫球蛋白λ轻链可变结构域,所述人类免疫球蛋白λ轻链可变结构域由实施方案74的方法产生。
[0732]
实施方案76.一种制造编码人类免疫球蛋白重链的核苷酸序列的方法,所述方法包括以下步骤:
[0733]
(a)将实施方案4
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;
[0734]
(b)获得编码特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列的人类免疫球蛋白重链可变区;以及
[0735]
(c)将所述人类免疫球蛋白重链可变区可操作地连接至人类免疫球蛋白重链恒定区序列以形成编码人类免疫球蛋白重链的核苷酸序列。
[0736]
实施方案77.一种编码人类免疫球蛋白重链的核苷酸序列,所述核苷酸序列由实施方案76的方法产生。
[0737]
实施方案78.一种制造包含人类免疫球蛋白重链可变区的核苷酸序列的方法,所
述方法包括以下步骤:
[0738]
(a)将实施方案4
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;以及
[0739]
(b)获得编码特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白重链可变结构域序列的人类免疫球蛋白重链可变区。
[0740]
实施方案79.一种包含人类免疫球蛋白重链可变区的核苷酸序列,所述核苷酸序列由实施方案78的方法产生。
[0741]
实施方案80.一种制造编码人类免疫球蛋白λ轻链的核苷酸序列的方法,所述方法包括以下步骤:
[0742]
(a)将实施方案1
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;
[0743]
(b)获得编码特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列的人类免疫球蛋白λ轻链可变区;以及
[0744]
(c)将所述人类免疫球蛋白λ轻链可变区可操作地连接至人类免疫球蛋白λ轻链恒定区序列以形成编码人类免疫球蛋白λ轻链的核苷酸序列。
[0745]
实施方案81.一种编码人类免疫球蛋白λ轻链的核苷酸序列,所述核苷酸序列由实施方案80的方法产生。
[0746]
实施方案82.一种制造包含人类免疫球蛋白λ轻链可变区的核苷酸序列的方法,所述方法包括以下步骤:
[0747]
(a)将实施方案1
‑
34中任一项的基因修饰啮齿动物暴露于所关注抗原;以及
[0748]
(b)获得编码特异性结合所述抗原并由所述基因修饰啮齿动物产生的抗体的人类免疫球蛋白λ轻链可变结构域序列的人类免疫球蛋白λ轻链可变区。
[0749]
实施方案83.一种包含人类免疫球蛋白λ轻链可变区的核苷酸序列,所述核苷酸序列由实施方案82方法产生。
[0750]
实施方案84.一种靶向载体,所述靶向载体包含:
[0751]
(i)5'同源臂,所述5'同源臂包含与内源啮齿动物κ轻链基因座中的5'标靶序列相对应的核苷酸序列;
[0752]
(ii)单一重排人类免疫球蛋白λ轻链可变区,所述单一重排人类免疫球蛋白λ轻链可变区包含vλ基因区段和jλ基因区段;
[0753]
(iii)啮齿动物cλ基因区段;以及
[0754]
(iv)3'同源臂,所述3'同源臂包含与内源啮齿动物κ轻链基因座中的3'标靶序列相对应的核苷酸序列。
[0755]
实施方案85.实施方案84的靶向载体,其中所述人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。
[0756]
实施方案86.实施方案84或85的靶向载体,其中所述人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。
[0757]
实施方案87.一种制造基因修饰啮齿动物的方法,所述方法包括以下步骤:
[0758]
(a)将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中;以及
[0759]
(b)使用步骤(a)中产生的所述啮齿动物es细胞产生啮齿动物。
[0760]
实施方案88.一种制造基因修饰啮齿动物的方法,所述方法包括以下步骤:
[0761]
(a)将可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段;以及
[0762]
(b)使用步骤(a)中产生的所述啮齿动物es细胞产生啮齿动物。
[0763]
实施方案89.实施方案87或88的方法,其中所述啮齿动物es细胞的基因组在工程化内源免疫球蛋白重链基因座处包含可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。
[0764]
实施方案90.一种制造基因修饰啮齿动物es细胞的方法,所述方法包括以下步骤:
[0765]
将包含人类vλ基因区段和人类jλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中。
[0766]
实施方案91.一种制造基因修饰啮齿动物es细胞的方法,所述方法包括以下步骤:
[0767]
将可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区引入啮齿动物es细胞的基因组中的工程化内源免疫球蛋白κ轻链基因座中,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段。
[0768]
实施方案92.实施方案90或91的方法,其中所述啮齿动物es细胞的基因组在工程化内源免疫球蛋白重链基因座处包含:
[0769]
可操作地连接至一个或多个内源免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段。
[0770]
实施方案93.一种制造基因修饰啮齿动物的方法,所述方法包括以下步骤:
[0771]
(a)对啮齿动物的种系基因组中的内源免疫球蛋白κ轻链基因座进行工程化以包含可操作地连接至啮齿动物cλ基因区段的单一重排人类免疫球蛋白λ轻链可变区,其中所述单一重排人类免疫球蛋白λ轻链可变区包含人类vλ基因区段和人类jλ基因区段,
[0772]
使得由所述基因修饰啮齿动物的b细胞表达的所有免疫球蛋白λ轻链均包括从所述单一重排人类免疫球蛋白λ轻链可变区或其体细胞超突变型式表达的人类免疫球蛋白λ轻链可变结构域。
[0773]
实施方案94.实施方案93的方法,其中所述方法还包括以下步骤:
[0774]
(b)对啮齿动物的种系基因组中的内源免疫球蛋白重链基因座进行工程化以包含可操作地连接至一个或多个啮齿动物免疫球蛋白重链恒定区基因的一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段,
[0775]
使得由所述基因修饰啮齿动物的b细胞表达的所有重链均包括人类免疫球蛋白重链可变结构域和啮齿动物免疫球蛋白重链恒定结构域。
[0776]
实施方案95.实施方案93或94的方法,其中步骤(a)和/或步骤(b)是在啮齿动物es细胞中进行的。
[0777]
实施方案96.实施方案93
‑
95中任一项的方法,其中所述内源免疫球蛋白κ轻链基因座缺乏啮齿动物cκ基因。
[0778]
实施方案97.实施方案93
‑
96中任一项的方法,其中所述人类vλ基因区段选自由以下组成的组:vλ1
‑
51、vλ5
‑
45、vλ1
‑
44、vλ1
‑
40、vλ3
‑
21和vλ2
‑
14。
[0779]
实施方案98.实施方案93
‑
97中任一项的方法,其中所述人类jλ基因区段选自由以下组成的组:jλ1、jλ2、jλ3、jλ6和jλ7。
[0780]
实施方案99.实施方案94
‑
98中任一项的方法,其中所述一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段代替一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段、一个或多个内源j
h
基因区段或它们的组合。
[0781]
实施方案100.实施方案94
‑
99中任一项的方法,其中所述一个或多个未重排人类v
h
基因区段、一个或多个未重排人类d
h
基因区段和一个或多个未重排人类j
h
基因区段分别替代一个或多个内源v
h
基因区段、一个或多个内源d
h
基因区段和一个或多个内源j
h
基因区段。
[0782]
实施方案101.实施方案94
‑
100中任一项的方法,其中所述一个或多个啮齿动物免疫球蛋白重链恒定区基因是一个或多个内源啮齿动物免疫球蛋白重链恒定区基因。
[0783]
实施方案102.实施方案94
‑
101中任一项的方法,其中:
[0784]
(i)所述一个或多个未重排人类v
h
基因区段包含v
h3‑
74、v
h3‑
73、v
h3‑
72、v
h2‑
70、v
h1‑
69、v
h3‑
66、v
h3‑
64、v
h4‑
61、v
h4‑
59、v
h1‑
58、v
h3‑
53、v
h5‑
51、v
h3‑
49、v
h3‑
48、v
h1‑
46、v
h1‑
45、v
h3‑
43、v
h4‑
39、v
h4‑
34、v
h3‑
33、v
h4‑
31、v
h3‑
30、v
h4‑
28、v
h2‑
26、v
h1‑
24、v
h3‑
23、v
h3‑
21、v
h3‑
20、v
h1‑
18、v
h3‑
15、v
h3‑
13、v
h3‑
11、v
h3‑
9、v
h1‑
8、v
h3‑
7、v
h2‑
5、v
h7‑4‑
1、v
h4‑
4、v
h1‑
3、v
h1‑
2、v
h6‑
1或它们的任何组合,
[0785]
(ii)所述一个或多个未重排人类d
h
基因区段包含d
h1‑
1、d
h2‑
2、d
h3‑
3、d
h4‑
4、d
h5‑
5、d
h6‑
6、d
h1‑
7、d
h2‑
8、d
h3‑
9、d
h3‑
10、d
h5‑
12、d
h6‑
13、d
h2‑
15、d
h3‑
16、d
h4‑
17、d
h6‑
19、d
h1‑
20、d
h2‑
21、d
h3‑
22、d
h6‑
25、d
h1‑
26、d
h7‑
27或它们的任何组合,以及
[0786]
(iii)所述一个或多个未重排人类j
h
基因区段包含j
h
1、j
h
2、j
h
3、j
h
4、j
h
5、j
h
6或它们的任何组合。
[0787]
实施方案103.实施方案94
‑
102中任一项的方法,其中所述内源免疫球蛋白重链基因座缺乏功能性内源啮齿动物adam6基因。
[0788]
实施方案104.实施方案94
‑
103中任一项的方法,其中所述基因修饰啮齿动物的种系基因组包含编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
[0789]
实施方案105.实施方案104的方法,其中所述一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段由所述基因修饰啮齿动物表达。
[0790]
实施方案106.实施方案104或105的方法,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列包括在与所述工程化内源免疫球蛋白重链基因座相同的染色体上。
[0791]
实施方案107.实施方案104
‑
106中任一项的方法,其中所述工程化内源免疫球蛋白重链基因座包含所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列。
[0792]
实施方案108.实施方案104
‑
107中任一项的方法,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列代替人类adam6伪基因。
[0793]
实施方案109.实施方案104
‑
108中任一项的方法,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列替代人类adam6伪基因。
[0794]
实施方案110.实施方案104
‑
109中任一项的方法,其中所述一个或多个人类v
h
基因区段包含第一人类v
h
基因区段和第二人类v
h
基因区段,且所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于所述第一人类v
h
基因区段与所述第二人类v
h
基因区段之间。
[0795]
实施方案111.实施方案110的方法,其中所述第一人类v
h
基因区段是v
h1‑
2且所述第二人类v
h
基因区段是v
h6‑
1。
[0796]
实施方案112.实施方案104
‑
111中任一项的方法,其中所述编码一种或多种啮齿动物adam6多肽、其功能直系同源物、功能同源物或功能片段的一个或多个核苷酸序列介于人类v
h
基因区段与人类d
h
基因区段之间。
[0797]
实施方案113.实施方案93
‑
112中任一项的方法,其中所述啮齿动物cλ基因具有与以下至少80%同一的序列:(i)小鼠cλ1,(ii)小鼠cλ2,或(iii)小鼠cλ3基因。
[0798]
实施方案114.实施方案93
‑
113中任一项的方法,其中所述啮齿动物cλ基因包含小鼠cλ基因。
[0799]
实施方案115.实施方案93
‑
114中任一项的方法,其中所述啮齿动物cλ基因包含小鼠cλ1基因。
[0800]
实施方案116.实施方案93
‑
112中任一项的方法,其中所述啮齿动物cλ基因具有与以下至少80%同一的序列:(i)大鼠cλ1,(ii)大鼠cλ2,(iii)大鼠cλ3,或(iv)大鼠cλ4基因。
[0801]
实施方案117.实施方案93
‑
112中任一项的方法,其中所述啮齿动物cλ基因包含大鼠cλ基因。
[0802]
实施方案118.实施方案93
‑
117中任一项的方法,其中所述单一重排人类免疫球蛋白λ轻链可变区代替一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。
[0803]
实施方案119.实施方案93
‑
118中任一项的方法,其中所述单一重排人类免疫球蛋白λ轻链可变区替代一个或多个啮齿动物vκ基因区段、一个或多个啮齿动物jκ基因区段或它们的任何组合。
[0804]
实施方案120.实施方案93
‑
119中任一项的方法,其中所述啮齿动物的种系基因组还包含失活内源免疫球蛋白λ轻链基因座。
[0805]
实施方案121.实施方案93
‑
120中任一项的方法,其中所述内源vλ基因区段、所述内源jλ基因区段和所述内源cλ基因整体或部分缺失。
[0806]
实施方案122.实施方案93
‑
121中任一项的方法,其中所述啮齿动物是大鼠或小鼠。
[0807]
等效物
[0808]
所属领域的技术人员应了解,所属领域的技术人员容易想到对本公开的多种改变、修改和改良。这种改变、修改和改良意图为本公开的一部分,并且意图在本发明的精神和范围内。因此,以上描述和图式仅作为示例,且本公开中描述的任何发明由随后权利要求书进一步详细描述。
[0809]
所属领域的技术人员将了解典型标准差或误差可归因于在测定法或如本文所述的其它方法中获得的值。本文中提及的描述本发明背景和提供有关本发明实施的额外细节的公布、网站和其它参考资料以引用的方式整体并入本文中。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1