III-A型CRISPR-Cas系统抑制剂AcrIIIA2及其应用的制作方法
本发明属于rna编辑技术领域,具体为一种iii-a型crispr-cas系统抑制剂acriiia2及其应用。
背景技术:
在过去的十年中微生物学最激动人心的发现之一是,类似于真核生物,细菌同样具有获得性免疫系统,打破了长久以来“获得性免疫是真核生物所特有的”定论。在外来入侵过程中,细菌已进化出一种新的特有的免疫防御系统,即crispr-cas系统(成簇规律间隔短回文重复序列及相关cas蛋白)。crispr-cas系统通过捕获整合外源核酸片段,并在cas蛋白和crisprrnas(crrna)的共同作用下保护细菌和古生菌免遭外来噬菌体、病毒和质粒的的入侵。crispr-cas系统已被开发用于基因编辑,应用于生物科学、医学诊断和作物育种等领域。但是由于cas蛋白在完成对目标基因的编辑后持续激活,可导致在整个基因水平进行非特异性切割,从而造成未知后果。因此,合理控制crispr-cas的编辑活性,降低脱靶效应,是crispr-cas系统应用于基因编辑、生物治疗等迫切需要解决的科学问题。
技术实现要素:
有鉴于此,本发明的目的之一在于提供一种iii-a型crispr-cas系统抑制剂acriiia2,本发明的目的之二在于提供一种含有acriiia2和/或acriiia2的同源性蛋白的试剂或组合物,本发明的目的之三在于提供所述试剂或组合物在制备抑制iii-a型crispr-cas系统rna编辑活性的药物中的应用。为达到上述目的,本发明提供如下技术方案:
1、iii-a型crispr-cas系统抑制剂acriiia2,所述acriiia2的氨基酸序列为seqidno:8。
作为优选的技术方案之一,所述acriiia2的基因序列为s.argenteus3688stdy6125118细菌中被iii-a型crispr-cas系统靶向识别的序列位点。
作为优选的技术方案之一,所述acriiia2与acriiia2的同源性蛋白的序列相比具有至少70%的序列同一性,并且具有与acriiia2相同的生物学功能。
作为优选的技术方案之一,所述同源性蛋白的氨基酸序列为seqidno:9。
作为优选的技术方案之一,所述同源性蛋白的氨基酸序列为seqidno:10。
作为优选的技术方案之一,所述acriiia2或所述acriiia2的同源性蛋白抑制iii-a型crispr-cas系统切割rna的活性。
2、含有acriiia2和/或acriiia2的同源性蛋白的试剂或组合物。
3、所述试剂或组合物在制备抑制iii-a型crispr-cas系统rna编辑活性的药物中的应用。
本发明的有益效果在于:
本发明从staphylococcusargenteus3688stdy6125118细菌基因序列中筛选出抑制iii-a型crispr-cas基因编辑活性的iii型抗crispr(acr)抑制剂acriiia2,在细菌和哺乳细胞中acriiia2控制iii-a型crispr-cas基因编辑能力的“关闭”。此外,本发明还筛选出acriiia2的同源性蛋白,并具有相同抑制效果,丰富了iii-a型crispr-cas系统抑制剂的种类。acriiia2抑制剂及其同源蛋白可以在时间或空间上控制iii-a型crispr-casrna编辑效率,提高iii-a型crispr-cas编辑技术在生物治疗、生物技术和农业等领域的安全性和实用性。
附图说明
图1为生物信息筛选与txtl鉴定acriiia基因。a为staphylococcusargenteus3688stdy6125118含有自身crrna识别的位点及候选acriiia基因示意图;b为transcription-translation(txtl)反应体系示意图;c、d为gfp荧光检测候选acriiia基因抑制iii-a型crispr-cas切割rna的活性。
图2为acriiia1和acriiia2在细菌中抑制iii-a型crispr-cas切割rna的活性。a为ms2rna噬菌体感染宿主的噬菌斑实验的设计;b为噬菌斑实验验证候选基因orf1-23基因抑制iii-a型crispr-cas抵抗ms2rna噬菌体感染宿主;c为acriiia1和acriiia2基因抑制iii-a型crispr-cas切割rna的活性。
图3为哺乳细胞hek293t中验证acriiias基因抑制iii-a型crispr-cas编辑技术的活性。a为hek293t细胞中建立检测acriiias基因抑制iii-a型crispr-cas切割rna活性的示意图;b为qrt-pcr检测iav病毒rna表达水平评估acriiia1和acriiia2基因抑制iii-a型crispr-cas的活性效率;c为iav病毒滴度评估acriiia1和acriiia2基因抑制iii-a型crispr-cas的活性效率。
图4为acriiia1及同源性蛋白的系统进化树分析。
图5为acriiia2及同源性蛋白的系统进化树分析。
图6为acriiia1和acriiia2的同源性蛋白在细菌中抑制iii-a型crispr-cas切割rna的活性;a为acriiia1同源性蛋白的氨基酸序列比对;b为acriiia2同源性蛋白的氨基酸序列比对;c为噬菌斑实验验证acriiia1和acriiia2同源性基因抑制iii-a型crispr-cas切割rna的活性。
具体实施方式
以下通过特定的具体实例说明本发明的实施方式,本领域技术人员可由本说明书所揭露的内容轻易地了解本发明的其他优点与功效。本发明还可以通过另外不同的具体实施方式加以实施或应用,本说明书中的各项细节也可以基于不同观点与应用,在没有背离本发明的精神下进行各种修饰或改变。
实施例1
acriiia1和acriiia2的鉴定与克隆
(1)生物信息筛选与分析候选acriiias基因
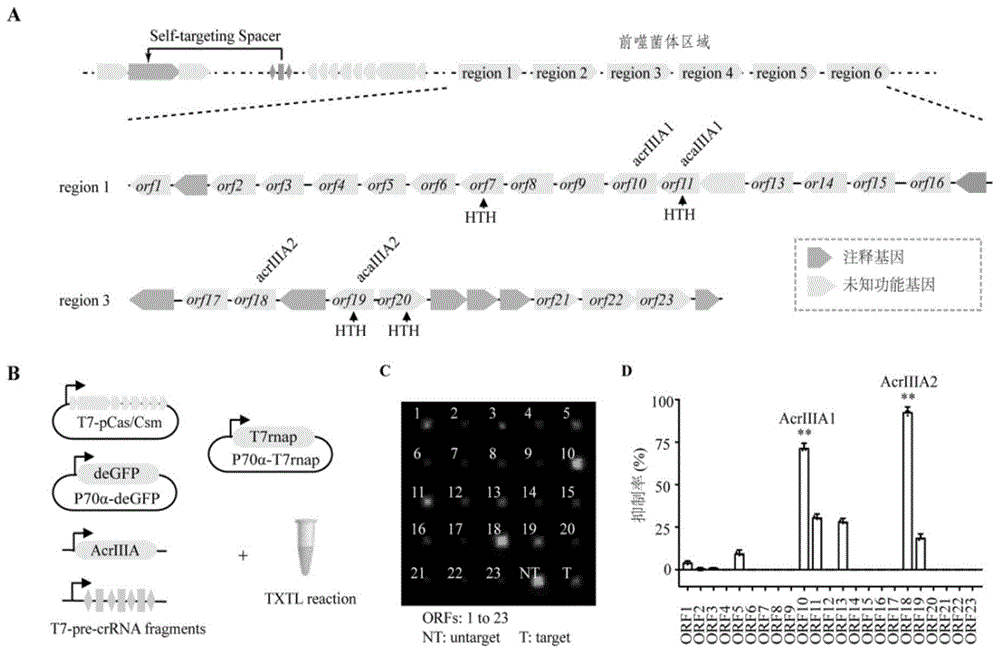
根据在细菌的转录组中存在能够被iii-a型crispr-cas靶向识别的rna序列,暗示在细菌自身基因组中可能包含了阻止iii-a型crispr-cas系统rna编辑活性的抑制剂。依据该原则,在含有iii-a型crispr-cas系统的staphylococcusargenteus3688stdy6125118细菌基因序列(https://www.ncbi.nlm.nih.gov/,登录号:nz_fqly01000002.1),通过self-targetingspacesearchplatform(ssts)分析,获得可以被iii-a型crispr-cas靶向识别的序列位点(图1中a),暗示在s.argenteus3688stdy6125118中可能存在抑制iii-a型crispr-cas切割rna活性的抑制剂。由于acr基因通常与含有保守helix-turn-helix(hth)结构域的acr-associated(aca)基因成簇存在。在此基础上,首先,通过phagesearchtool(phast)在s.argenteus3688stdy6125118菌中鉴定到6个前噬菌体区域(prophageregions),编码204开放阅读框(openreadingframes:orfs);其次,利用pfam(proteinfamiliesdatabase)分析这204orfs的结构域,鉴定获得含有hth结构域的aca基因;最后,以邻近aca基因并且转录方向相同为条件,共筛选获得23个未知功能的基因(orf1-23)作为acriiias的候选基因,用于下一步验证。
(2)候选基因acriiias的克隆及其重组表达载体的构建
由geneuniversal公司合成orf1-23候选基因的全长序列,并通过ncoi和xhoi双酶切并回收基因片段,同时将pet28a载体用ncoi和xhoi双酶切,回收载体骨架,将回收的基因酶切片段和pet28a骨架片段进行连接转化,获得pet28a-orf1至pet28a-orf23重组质粒,经测序确认正确的重组质粒转化大肠杆菌。
gfppre-crrna序列:(seqidno:1)
5’-gatataaacctaattacctcgagaggggacggaaacggacacgctgaacttgtggccgtttacgtcgccgtcgatataaacctaattacctcgagaggggacggaaaccttcagggtcagcttgccgtaggtggcatcgccctcgatataaacctaattacctcgagaggggacggaaacgggtggtcacgagggtgggccagggcacgggcagctgatataaacctaattacctcgagaggggacggaaac-3’
gfppre-crrna序列由geneuniversal公司合成,经kpni和xbai双酶切链接至pbluescriptiisk(+),获得pbluescriptiisk(+)-gfppre-crrna(t7-pre-crrnafragments)重组质粒。
(3)无细胞转录-翻译系统(transcription-translation,txtl)系统筛选acriiias基因
txtl反应按照制造厂商所建议的条件,由degfp报告质粒(p70α-degfp,arborbioscience),iii-a型crispr-cas表达质粒(t7-pcas/csm),gfppre-crrna(t7-pre-crrnafragments),t7rnap表达质粒(ptxtl-p70a-t7rnaphp,arborbioscience),和pet28a-orf1至pet28a-orf23重组质粒组成反应体系(图1中b)。其中,所有质粒用qiagenplasmidminikit(qiagen)提取获得超纯质粒,经ampurexpbeads(beckmancoulter)纯化获得质粒。
每12μl的txtl反应体系含有9μltxtlmastermix,0.1nm的ptxtl-p70a-t7rnaphp,0.125nm的p70α-degfp,1nm的t7-pcas/csm,2nm的pbluescriptiisk(+)-gfppre-crrna,2nm的pet28a-orf1至pet28a-orf23,和5μm的iptg。29℃、反应20小时,用bioteksynergyhtmulti-modemicroplatereader检测gfp荧光值。同时,用ivisxriisystem(perkinelmer)拍摄gfp荧光照片。抑制iii-a型crispr-cas切割活性的算法如下:
gfp20h代表在20h时crrna靶向gfp时的gfp荧光值,gfpmin代表检测过程中最小的gfp荧光值,gfpev,20h代表没acr存在时gfp荧光值,gfpnt,20h代表crrna非靶向gfp时的gfp荧光值,gfpnt,min代表检测过程中最小的gfp荧光值。
txtl反应结果显示,orf10和orf18能够在体外抑制iii-a型crispr-cas介导的rna切割能力,分别命名为acriiia1和acriiia2(图1中c、d)。
实施例2
acriiia1和acriiia2在细菌中抑制iii-a型crispr-cas切割rna的活性
在细菌中验证acriiia1和acriiia2是否抑制iii-a型crispr-cas的活性。首先,构建pcrisprms2重组质粒,获得crrna表达载体用于识别ms2rna噬菌体的reprna。pre-crrna序列由geneuniversal公司合成,如图2中a所示,pre-crrna经t4dna连接酶链接至pbluescriptiisk(+)1载体,获得重组质粒pcrisprms2。其次,将pet28a-acriiia1、pet28a-acriiia2或者pet28a与t7-pcas/csm质粒和pcrisprms2质粒转化至感受态细胞,获得e.colic-3000/t7-pcas/csm/pcrisprms2/pet28a、e.colic-3000/t7-pcas/csm/pcrisprms2/pet28a-acriiia1和e.colic-3000/t7-pcas/csm/pcrisprms2/pet28a-acriiia2菌株用于ms2rna噬菌体入侵的噬菌斑形成实验(图2中a)。将获得重组菌株37℃、220rpm/min培养过夜,接种至含有1mmiptg的lbagar(0.3%)固体培养基上,用10倍梯度稀释的ms2rna噬菌体感染,培养过夜,观察噬菌斑的形成并拍照。
结果如图2中b、c所示,acriiia1和acriiia2抑制iii-a型crispr-cas介导的rna切割能力,导致ms2rna噬菌体感染宿主并生长增殖,导致宿主裂解,最终形成噬菌斑。表明在细菌中acriiia1和acriiia2可以阻止iii-a型crispr-casrna编辑活性。
实施例3
acriiia1和acriiia2在哺乳细胞中抑制iii-a型crispr-casrna的编辑活性
为了验证acriiia1和acriiia2在hek293t细胞中是否抑制iii-a型crispr-casrna编辑活性,在hek293t细胞中检测acriiia1和acriiia2对iii-a型crispr-cas切割流感病毒(influenzaavirus,iav)的效率影响。利用lipofectaminetmcrisprmaxtmtransfectionreagent(thermofisherscienctific)将靶向iav的iii-a型crispr-cas复合物与acriiia1或者acriiia2蛋白转染hek293t细胞后,感染iav病毒(moi=0.01和0.5,multiplicityofinfection),检测iav病毒的rna以及病毒滴度(图3中a)。过程如下:
1)acriiia1、acriiia2和iii-a型crispr-cas蛋白的ni柱亲和层析纯化:
将构建的pet28a-his-acriiia1、pet28a-his-acriiia1、以及t7-pcas/csm/his-csm2质粒转化e.colibl21感受态细胞,获得重组质粒菌,扩大培养后,7000rpm离心10分钟收集,用pbs清洗3次,液氮反复冻融3次后,超声波破碎30min,16000rpm离心30分钟收集上清,上清用0.45μm滤膜抽滤;将抽滤后的上清上柱到用pbs平衡过的ni亲和柱中,分别用30mlpbs溶液和20mm咪唑的pbs快速冲洗柱子,以洗去未结合的蛋白,分别用10ml含100mm、500mm和1m咪唑的pbs缓慢冲洗柱子并收集洗脱液,大约每2ml收集一管,将收集到的洗脱液进行sds-page电泳检测,以丢弃含有杂蛋白的洗脱液,用pbs溶液进行透析36h,每8h换透析液一次,最后进行sds-page电泳检测,结果表明分离纯化获了单一条带的acriiia1和acriiia2蛋白,以及iii-a型crispr-cas复合物蛋白;并用bca蛋白浓度测定试剂盒测定其浓度。
2)iavguiderna序列合成
5’-acggaaacguaaugaaggaucuuauuucuucggagacaau-3’(seqidno:2)
5’-acggaaacggucgguugcucacaaguccugccugccugcc-3’(seqidno:3)
3)转染
参照lipofectaminecrisprmaxtmtransfectionreagent(thermofisherscienctific)转染试剂说明书,用3000ngiii-a型crispr-cas复合物蛋白,1200ngiavgrnas,2000ngacriiia1或者acriiia2蛋白转染hek293t细胞后,感染iav病毒(moi=0.01和0.5,multiplicityofinfection),24h后检测iav病毒的rna以及病毒滴度。
结果如图3中b、c所示,在hek293t细胞中acriiia1和acriiia2阻止iii-a型crispr-cas介导的rna编辑活性,acriiia1和acriiia2可有效控制iii-a型crispr-cas基因编辑的“关闭”。这将有助于改进iii-a型crispr-cas介导的rna编辑技术在临床治疗和应用研究的安全性。
实施例4
acriiia1和acriiia2的同源性蛋白分析
利用acriiia1的蛋白序列对所有ncbi中的蛋白序列进行blastp比对,选择e-value小于10-3以及同源性比例大于70%的蛋白为acriiia1的同源性蛋白。鉴定获得acriiia1的同源蛋白包括wp_001552317.1,yp_009196757.1,yp_002332371.1,ewj86503.1,eho90800.1,kmr53579.1,euq10906.1,scu38681.1,ewa35716.1,ewr63129.1,ewk80326.1,axj28344.1,shd87588.1,kfa43737.1,awq90359.1,evd55746.1,eur30676.1,sgs29864.1,ewh71077.1,coe55786.1,arm68199.1,wp_095376943.1,kfb80258.1,evg06959.1,eje28889.1,evv21927.1,czq83597.1和wp_037544580.1;其中arm68199.1(acriiia1ime1367)、yp_009196757.1(acriiia123mra)和yp_002332371.1(acriiia1ipla35)是来自噬菌体病毒的同源性蛋白。同时,用mega7进行系统进化树分析,制作acriiia1及其同源性蛋白系统进化树(图4)。
利用acriiia2的蛋白序列对所有ncbi中的蛋白序列进行blastp比对,选择e-value小于10-3以及同源性比例大于70%的蛋白。鉴定获得acriiia2的同源蛋白包括:
wp_072465245.1,wp_072539211.1,wp_031868661.1,wp_064131496.1,wp_117232106.1,wp_001077670.1,wp_001573838.1,wp_106104614.1,wp_053005550.1,wp_001077638.1,wp_070059026.1,wp_015978251.1,wp_103259238.1,wp_103252633.1,wp_103147425.1,wp_072458005.1,wp_031921279.1,wp_129934257.1,wp_101766656.1,wp_145340959.1,wp_070848524.1,wp_053031406.1,wp_105967223.1,wp_107399202.1,wp_119504741.1,wp_050331550.1,ptf96982.1,wp_075778679.1,wp_002468630.1,wp_021298890.1,wp_145449609.1,wp_002501272.1,wp_141489545.1,wp_002469096.1,wp_049391373.1,wp_115343401.1,wp_049387433.1,wp_002469451.1,wp_064587828.1,wp_002439153.1,ptg35301.1,wp_015365401.1,wp_031765295.1,wp_017804551.1,wp_024273300.1,wp_029625613.1,wp_072599561.1,wp_048527588.1,yp_006382263.1,wp_001837400.1,wp_000896616.1,yp_003857099.1,wp_110179714.1,wp_111762068.1,wp_050961184.1,wp_042856227.1,wp_032099440.1,wp_094969788.1,wp_093514686.1,wp_072527761.1,wp_070859732.1,wp_049401137.1,wp_046467714.1,wp_115287758.1,sum72483.1,wp_072492559.1,rcv80954.1,wp_114288318.1,wp_032604936.1,wp_002495917.1,wp_002502889.1,wp_124263453.1,wp_069996864.1,wp_135789161.1,egs40332.1,wp_060556001.1,wp_031764150.1,wp_070481548.1,wp_046597470.1,agz24991.1,wp_099816467.1,wp_002475547.1,ehm65174.1,eje02307.1,wp_129531134.1,wp_087437151.1,wp_070664144.1;
其中yp_006382263.1(acriiia2tem123)和yp_003857099.1(acriiia2sap26)是来自噬菌体病毒的同源性蛋白。同时,用mega7进行系统进化树分析,制作acriiia2及其同源性蛋白系统进化树(图5)。
为了检测表1中来源于噬菌体的acriiia1和acriiia2同源性蛋白是否抑制iii-a型crispr-cas的活性,按照实施例2的方法进行ms2rna噬菌体入侵的噬菌斑形成实验。结果如图6中a、b、c所示,acriiia1的同源性蛋白(acriiia1ime1367、acriiia123mra和acriiia1ipla35)和acriiia2的同源性蛋白(acriiia2tem123和acriiia2sap26)抑制iii-a型crispr-cas介导的rna切割能力,导致ms2rna噬菌体感染宿主并生长增殖,导致宿主裂解,最终形成噬菌斑。表明在噬菌体病毒来源的中acriiia1和acriiia2同源性蛋白也可以阻止iii-a型crispr-casrna编辑活性。
表1acriiia1和acriiia2及同源性蛋白的氨基酸序列表
最后说明的是,以上实施例仅用以。说明本发明的技术方案而非限制,尽管参照较佳实施例对本发明进行了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本技术方案的宗旨和范围,其均应涵盖在本发明的权利要求范围当中。
序列表
<110>中国人民解放军陆军特色医学中心
<120>iii-a型crispr-cas系统抑制剂acriiia2及其应用
<160>10
<170>siposequencelisting1.0
<210>1
<211>252
<212>dna
<213>人工序列(artificialsequence)
<400>1
gatataaacctaattacctcgagaggggacggaaacggacacgctgaacttgtggccgtt60
tacgtcgccgtcgatataaacctaattacctcgagaggggacggaaaccttcagggtcag120
cttgccgtaggtggcatcgccctcgatataaacctaattacctcgagaggggacggaaac180
gggtggtcacgagggtgggccagggcacgggcagctgatataaacctaattacctcgaga240
ggggacggaaac252
<210>2
<211>40
<212>rna
<213>人工序列(artificialsequence)
<400>2
acggaaacguaaugaaggaucuuauuucuucggagacaau40
<210>3
<211>40
<212>rna
<213>人工序列(artificialsequence)
<400>3
acggaaacggucgguugcucacaaguccugccugccugcc40
<210>4
<211>58
<212>prt
<213>人工序列(artificialsequence)
<400>4
metasngluglulysleulysmetileleuleuleuleugluaspile
151015
proargaspglutrpasnargleuvalasngluvalasnasnglntyr
202530
sertyrglnalaasplysvalglyleualaseraspasncysglngln
354045
ilealaasnasntyrlyshistyrglyphe
5055
<210>5
<211>58
<212>prt
<213>人工序列(artificialsequence)
<400>5
metasngluglulysleulysmetileleuleuleuleugluaspile
151015
proargaspglutrpasnargleuvalasngluvalasnasnglntyr
202530
sertyrglnalaasplysvalglyleualaseraspasncysglngln
354045
ilealaasnasntyrlyshistyrglyphe
5055
<210>6
<211>58
<212>prt
<213>人工序列(artificialsequence)
<400>6
metasngluglulysleulysmetileleuleuleuleugluaspval
151015
proargaspglutrpasnargleuvalasngluvalasnasnglntyr
202530
sertyrglnalaasplysvalglyleualaseraspasncysglngln
354045
ilealaasnasntyrlyshistyrglyphe
5055
<210>7
<211>58
<212>prt
<213>人工序列(artificialsequence)
<400>7
metarggluglulysleulysmetleuleuleuleuleugluaspval
151015
proargaspasptrpasnargleuvalasngluvalasnasnglntyr
202530
sertyrglnalaasplysvalglyleualaseraspasncyslysgln
354045
ilealaasnasntyrlysargtyrglyphe
5055
<210>8
<211>163
<212>prt
<213>人工序列(artificialsequence)
<400>8
metasnglnvalserasnasplysleuthrvallysglusertrpthr
151015
alaglylysileargglylysleuasnlysglyglnlysglnvalphe
202530
aspargmetalaileserglulysargaspileileglulyspheasn
354045
asnasnvalprophegluvalglugluilegluargasnglngluthr
505560
lystyrlysileileglulysthrleuasnlysarggluleuasnthr
65707580
metseraspserglylysaspmetleuleulysasnlysvalglygln
859095
pheileaspserpheserthrargpheserserserpheserasnpro
100105110
asnasnalaglyglnmetphethrtyrglumetileasnglnasnphe
115120125
valleuileglumetleuaspgluhisleulysasngluasnlysile
130135140
ilegluglnasnasngluileileaspleuleulysglnilealaasn
145150155160
lysglyval
<210>9
<211>172
<212>prt
<213>人工序列(artificialsequence)
<400>9
metlysglulysleulysmetalaglyasplysleuthrphelysglu
151015
ileleuthrgluthrlysmetpheserlysleusertyrarglysile
202530
aspmettyrlyslysmetthrthraspglulysarglysileleuasn
354045
aspphelysgluglylysgluleuaspileglnleutyrlysserglu
505560
asnphelysasnthrasngluglutyrgluserlysseralalysser
65707580
leuasnglyglnglyilelysglualathraspvalthrthrtyrala
859095
tyrglnlysglnasnileasnprothrleuleulysvaltyrasngly
100105110
leuglythrphethrthrasnvalasplysglnalalysphevalphe
115120125
tyraspthrglnleulysglnasnphevalserilealaglnargasp
130135140
gluleuilelysglnasnasnargileilegluglnasnasngluval
145150155160
ileaspleuleulysglnilealaasnlysglyval
165170
<210>10
<211>180
<212>prt
<213>人工序列(artificialsequence)
<400>10
mettrpargpheasnileasnglnmetlysglulysleulysmetala
151015
glyasplysleuthrphelysgluileleuthrgluthrlysmetphe
202530
serlysleuserasnarglysileaspmettyrlyslysmetthrthr
354045
aspglulysarglysileleuasnaspphelysgluglylysgluleu
505560
aspileglnleutyrlyssergluasnphelysasnthrasngluglu
65707580
tyrgluserlysseralalysserleuasnglyglnglyilelysglu
859095
alathraspvalthrthrtyralatyrglnlysglnasnileasnpro
100105110
thrleuleulysvaltyrasnglyleuglythrphethrthrasnval
115120125
asplysglnalalysphevalphetyraspthrglnleulysglnasn
130135140
phevalserilealaglnargaspgluleuilelysglnasnasnarg
145150155160
ileilegluglnasnasngluvalileaspleuleulysglnileala
165170175
asnlysglyval
180
- 还没有人留言评论。精彩留言会获得点赞!